During the last half of 2008 I decided that I would make a last concerted effort to try and further clarify the uncertainty of classification of these plants. This involves the usual introspection, retrospection and reflection. Where are we now and what do we understand? I have had considerable correspondence and interchange of ideas with many enthusiasts. There is a huge disparity between what I write and what other authors do and there are definitely massive misunderstandings. The one reason is the obvious one that we each create our own realities and can only interpret the world around us in terms of our own experience and individual capacities. The second reason is that there are flaws in the entire information system in which we operate.
Much of my correspondence has involved trying to bring someone through the very simple barrier of what a species is and what a Latin binomial is supposed to convey. Several correspondents have remarked that they follow my revision Haworthia Revisited and cannot reconcile themselves with what I have subsequently written. Particularly revealing is the reaction to a very brief item I wrote entitled “A reality check” (reprinted in this publication). In that piece I submitted six pictures of very different plants and stated that they were all the same one species. Respondents expressed surprise that I could say that these were the same (thing), basing their opinions only on the fact that the plants looked so different. They lose sight of the fact that this is the reality of “species”. The members of a species are not all the same and especially not outwardly; and to confuse the sameness of members of a genetic system with the sameness of superficial resemblance, lies at the heart of classification and identification problems.
Gordon Rowley kindly sent me a copy of an article by Sandra Knapp FLS entitled … ”Naming Nature: The future of the Linnaean System”. Erudite and academic as the article is, it exposes the fact that there is a problem inherent in the system without actually defining what the problem is. It seems to me that the alarm bells of a sinking ship are being taken for sounds inherent to the structure and performance of the ship.The problem is that here is not the universal definition of what a species is and Knapp informs us that attempting to arrive at one is ”ultimately not practical in the short or long term”. My opinion is that a statement like this simply adds fuel to the fire. We have to define the word species and in doing do recognize that in their very nature species are not equivalent.
It is botanists themselves who have led us into this morass of names where the whole process of classification and nomenclature is an intellectual and juristic minefield. As collectors, growers and enthusiasts we have come to believe that a Latin name pins a plant into place simply by virtue of superficial resemblance. This is where we fail. Knapp cites Darwin as being unconcerned that the exact definition of a species was difficult to pin down. My opinion is that this statement is at the core of the matter. It is not the definition that is difficult to pin down. What is difficult to pin down is where any plant (organism) fits in any of the boxes we want to fabricate. There is absolutely no need to vacillate about a species definition because we do not know enough about the things we want to organize accordingly.
My plea to Haworthiophiles is now to try and think clearly and recognize that the formal system of latin binomials is primarily the domain of scientists. They have not set us a very good example and we need to adapt our outlook and the way in which we approach the use of Latin names.
In these now twilight notes of mine I will use an informal system of naming and describe some really interesting experiences and finds that follow the very path that Knapp suggests will lead us forward. Gordon Rowley wryly observed that “Bayer changes his mind”. This is because I have been getting on with the job and using names, just as Knapp proposes; as hypotheses, subject to continual testing and change as I gain better insights into processes and pattern in the field where the plants are. There is a misplaced confidence in technology and laboratory based methods that simply are no substitute for the information that is needed from direct field observation and experience.
There is one very disturbing aspect. This is that there are commercial and egocentric interests in opposition to mine that may be using my information for their own interests and to the detriment of sensitive habitats and populations. It is thus very distressing to know that the joy and delight these plants have given me on so many occasions is threatened or marred by the need I have felt to write about them. I have been taken to task for revealing localities. The thing is that there can be no understanding of these plants unless the spatial and distributional aspects are known and understood. There is no way in which my writing can stand as a valid hypothesis that can be tested and refined without this critical element in the constitution of a sensible view of what species are. I firmly believe that there is no merit in secrecy. It is not my role to play policeman and neither do I think that strict conservation laws and implementation have any merit where they deny the expression of natural curiosity and wonder about this incredible creation around us. What is important is for us as individuals to realize that we live in an extraordinary creation that is a conscious one and that we are individually tied to it and responsive to it. We are responsible for it and to it: we need to get back to caring for it rather than scraping what we can out of it.
I have also not written all this because of what someone really unkindly suggested was my need to write. I am writing now because I feel an obligation to do so, having started in an era where the available classification was simply woeful and ending in a situation that is a history repeating itself. It is not about Haworthia, it is about what is true and what is meaningful. It is about how we understand nature and about what nature should mean to us. We have an incredible creation and it is time we woke up to the reality of its beauty and its purpose. God did not write a single book that so many different elements of our society claim as one that only they possess and understand. If there is any book it is everywhere around us as our birthright. ♦
Clichés may often fall into the category of often repeated untruths that come to be believed. One I have used too often is … ”The problem is …”, without ever seemingly being to explain what it actually was. I studied Oxalis and it seemed that where there was an awful amount of detail to explain difference, all this detail simply obscured the fact of similarity. So what I did is “reverse engineer” the process and apply the principle to Haworthia where I have for so long tried fruitlessly to explain that we were all explaining and accepting that there was difference based on detail.
I made some progress in finding facts to support this contention in the approximate 40 populations of Haworthia in the Zuurberg that seem to suggest that plants looking as different as H. cooperi and H. cymbiformis may be the same species. However, this was not very convincing.
Recently I had reason to explore more populations in the arena of a larger problem in H. mirabilis where I may be considering as many as 400 populations or many more. Illustrated by the following sample pictures: … It is now my contention that different as all these single plants appear to be, they are in fact members of one species. The inference is drawn from observations of approximately 150 populations occurring in a geographically coherent pattern in the restricted area between Worcester and Riversdale and southward to the coast. The inference is strengthened by the observations of similar continual intergradation of variation in similar sets of populations throughout the distribution range of the genus.
I do think that this is evidence to put to rest the disputes about the classification of this group of plants. But I do think that there are implications for plant taxonomy in general perhaps starting simply in the Alooideae where there is such confoundment in the understanding of simply the genera. ♦
1. Haworthia mirabilis ‘paradoxa’. Vermaaklikheid2. H. mirabilis ‘paradoxa’. Vermaaklikheid (per Ismael Ebrahim)3. H. mirabilis ‘badia’. Napier4. H. mirabils ‘pilosa’. Lower Beede River.5. H. mirabilis ‘atrofusca’. W.Heidelberg.6. H. mirabilis ‘meiringii’. W Bonnievale
Steven Hammer, in his inimitable style, put a very fresh face on Haworthia in Cact. Succ. J. (Los Angeles) 80:140 (2008). He drew attention to the wonder of the plants in cultivation for the collector, contrasted to a reality of unglamorous scruffiness in the field as per the lens and pen of Bayer. It has fallen to my lot as a very unwilling taxonomist to reduce the fascination these plants have for me, and for so many others, to the mundane vortex of labels, their proliferation and continual amendment. The fact is whether on a label or on the tongue, a name is a part of any language we use to talk to each other; but we are not learning anything from a well-documented history and in Haworthia seem to remain lost in a maze.
The unhappy truth for Haworthia is that by the time von Poellnitz in Germany, G.G. Smith in South Africa, F. Resende in Portugal, A.J.A. Uitewaal in Holland, W. Triebner in Namibia, J.W. Dodson and J.R. Brown in USA had either left or abandoned the scene, there were any number of names that meant very little more than the Latin they were written in. J.R. Brown presented a talk, A brief review of the Genus Haworthia, to the Los Angeles Cactus and Succulent Society that was published in the Cactus and Succulent Journal of America 29:125-135 (1957). He noted the number of species and varietal names at 160 and 370 (!) respectively, arranged in 20 sections.
While J.R. Brown was winding down (his last note on Haworthia was published in 1970), I was busy trying to make sense of a two large files that seemed to form the body of a manuscript by G.G. Smith for which Mrs. M. Courtenay-Latimer had drafted a title… “A monograph of the genus Haworthia.” This manuscript comprised a collection of all current species descriptions arranged in the purported twenty sections of Berger and accompanied by many illustrations from the original publications, as well as by many of Smith’s own photographs and those of H.G. Fourcade. We know that Smith retired in a huff, but was there really good reason for his exit?
It is quite evident from the legacy of notes that the manuscript could not have been anywhere near complete. I drew attention to this in an article…”Reflections on G.G. Smith’s Haworthia collection” that was published in Excelsa 8:46-52 (1978). While stuck in the paradigm of 20 sections, he envisaged many more species and varieties and I cannot imagine how he could ever have reconciled this with the evidence of his compiled manuscript. I suspect he realized this too, hence the (self-directed?) huff.
It took me several years to absorb the specimens in the Compton Herbarium while at the same time taking Smith’s meticulous (in the sense of the literature from which it was extracted) collation of material to pieces. Putting it all back together as best I could, I eventually produced Haworthia Handbook (1976) published by National Botanic Gardens of South Africa. It was not a great work and neither was it very well-received at the time (though it has had a curiously perky afterlife once it went out of print). Charles Glass wrote to me that he was a bit disappointed (by its elliptic compressions?). Gordon Rowley was kind enough to point out many nomenclatural errors and a page of errata had to be inserted.
However, I was actually committed to helping Col. Scott write a formal revision as I had no aspirations to do so myself. He had been refused access to Smith’s collection, as the Curator of the Compton Herbarium had concurred with Mrs. Courtenay-Latimer (and perhaps Smith himself) that Col Scott would not do credit to a formal revision. It became quite problematic for me when I realized that this estimate of Scott’s botanical capability was uncomfortably close to true. This led me to re-write my handbook as The New Haworthia Handbook (1981), also published by National Botanic Gardens, hoping that Col. Scott would follow the ideas expressed in my optimistic foreword.
This did not happen and Scott’s…The Genus Haworthia – A Taxonomic Revision, appeared in 1983 published by Aloe Books, Johannesburg, as a closely and clumsily kept secret from me.
My honest opinion is that this book was a total disaster, taxonomically though not photographically, and it was most disheartening and confounding to consider that the flattering introduction was by Dr. L. E. Codd, then head of the national Division of Botany and of the Pretoria Herbarium. Not only that, Dr. Beth Gibbs Russell later telephoned me to ask whose version of Haworthia the herbarium and the institute should follow! I do not know what my response was other than thinking how unfair it was to place the onus on me to judge. Whatever I might have said, that did not stop the institute from publishing an extraordinary mish-mash of names for Haworthia in a national species list taken from Scott, myself and other sources. It was simultaneously no comfort to welcome the grand old man of Haworthia, J.W. Dodson, to the Karoo Garden. Bewildered by what he saw in the collection there, he could do no more than express his conviction that Col. Scott had produced a very workable revision!
As far as I was concerned, Haworthia was all emptied back into Smith’s “monograph,” and readers may imagine why I found this so disturbing. Around 1993-4 news of a new succulent lexicon to be compiled by Urs Eggli to replace the famous Jacobsen set was circulated. At the same time, by virtue of my contact with the Compton Herbarium, I was asked to write a synopsis of Haworthia for a list of plant species for the Cape Flora. These were the defining factors that led me to writing a formal revision. I cannot bring myself to detail the communications with other aspirants to taxonomic fame then and since, as these border on the bizarre and do nothing more than add to the disrepute of taxonomy in the minds of plant collectors who care about and are compelled to use Latin names.
When I eventually wrote my revision I had no illusion that it would be the last word on the subject. Indeed it seems in retrospect that it may well have been but the first! Not only has there been an explosion of new names and opinions, I have also been driven to examine and re-examine those areas of my understanding built not only on available field knowledge and exploration, but also on Smith’s vast legacy. It is necessary to note that the contributions of people like Jay Dodson and J.R. Brown played no small part either. The little book written by Brown, viz., Succulents for the Amateur, played such a significant part in cultivating my interest in plants that I have been very sensitive to the resonance of the devotion and interest Brown showed for Haworthia. This respect for others like F.R. Long and H.G. Fourcade, whom I never met, extended to people like Frank Stayner, G.J. Payne, Gordon King and so many others I met solely as a consequence of these plants. That the human relations aspect of their classification and identification took such a downturn in the wake of Smith’s criticism of, primarily, Resende in 1947 is a sad record in history. Relations never recovered and my involvement in and distress at the later half-repetitions of history is at the heart of this article.
Since 1995 my accession records have climbed from 6400 to 7700 and these numbers represent almost entirely new population records for Haworthia and I trust that I have not used my own numbers where a very few contributing collectors have helped me. I have written innumerable articles for Haworthia journals while Umdaus published Haworthia Update Vol 1 in 2002. This seems to have been a commercial failure but my belief is that editors and publishers have never understood the issues. I could not help because despite my excessive attempts to explain the problems causing strife and confusion in the literature, I really never exposed the heart of the matter. Fortunately for me Harry Mays of Alsterworthia kindly published Haworthia Updates Vols 2, 3 and 4 between 2003 and 2007.
In these volumes I tried to address the realities of variation and the confoundment that exists in my own and the communal mind, about what species are or might be as related to a perceived botanical and scientific process for naming plants. But if 40 delegates could be brought together as happened recently in an effort to create a united front just for the single genus Aloe, it does seem fairly senseless to be functioning so alone in Haworthia where a greater need exists. In my “unsparing” view of things I find it extraordinary that the symposium did not seem to address the wider problem of genera in the Alooideae. I am quite sure that if this were done first, it would facilitate the common understanding of lower ranks. In Haworthia there has been absolutely no common will to reach any kind of consensus and no reason has been found for that. My conviction has grown that Latin names, and the way the nomenclatural system is used, do nothing more than strain good will in that they implies a reality in a staged relationships of plants that is simply not true. If I were a more dedicated taxonomist I would have simply worked towards a functional generic arrangement: Haworthia for the glamour queens, Jayarbia (honoring J.R. Brown) for the Hexangulares, and Catevala for the Robustipedunculares.
I recently submitted a short list of names that I think should serve Haworthia to the Haworthia Society in England. As Gordon Rowley pointed out in a letter to me… “The problem is that Bruce Bayer changes his mind.” Well, I do want to change my mind about that list again. I would advise incorporating H. heidelbergensis, H. magnifica and H. maraisii and their variants all under H. mirabilis. Furthermore I would suggest abandoning the formal lower ranks of subspecies, variety or forma for appended third epithets in inverted commas. The list of species is thus reduced to a realistic 54.
But there is still a wealth of undiscovered things out there and the many existing names mean very little to me in understanding what I have seen and continue to find. So I give a brief overview of the product of very recent fieldwork where in less than 10 days my wife and I found 24 previously unrecorded Haworthia populations. These included two populations of H. floribunda, one of H. retusa ‘nigra’ (implementing a name change and a new way of expressing difference), five of H. rossouwii, one of H. minima, one of H. marginata, two of H. mirabilis ‘paradoxa’ and 12 of H. mirabilis. While each of these has a story attached, I am only going to touch on the latter.
I have written in detail about H. mirabilis in Haworthia Update 3:87 in a chapter that deals primarily with the populations in the western area of the distribution. I also wrote in detail of associated plants in Update Vol. 2:51-79 dealing with populations east of Heidelberg in the eastern area. So these new populations are relevant to the central zone and I will add several others that were found earlier but subsequent to the Update 3 publication, plus another by Marc Mougin (brought to my attention by Diederik Van den Abbeele). They all deal with an area extending eastwards from Stormsvlei and the Bromberg Mountain to the Karringmelks River southwest of Heidelberg and southwards to Koppies and Stuurmanskraal, where there are populations already covered in Updates.
Five of the populations can be unequivocally assigned to H. mirabilis ‘atrofusca’ and yet with a fair degree of variation. These are all just west of Heidelberg. The other seven that do nothing more than confound understanding of anything that can be construed as H. maraisii, H. magnifica, H. heidelbergensis or H. mirabilis, occur southwest of Heidelberg and extend westward to Diamant (east of the Bromberg). The only significant thing about the two groups is that where ‘atrofusca’ is evident in the populations, H. floribunda may be in the vicinity. Where H. floribunda is not present the seedlings resemble that species as an odd indication that perpetuated juvenile characters underlie the species floribunda. The variation within the populations is quite dramatic and it is indeed difficult to verbally report and sensibly convey this to a reader. The plants vary in size, texture, markings, leaf armature, color and translucence. So I submit pictures with the manuscript and offer the following comment as best I can:-
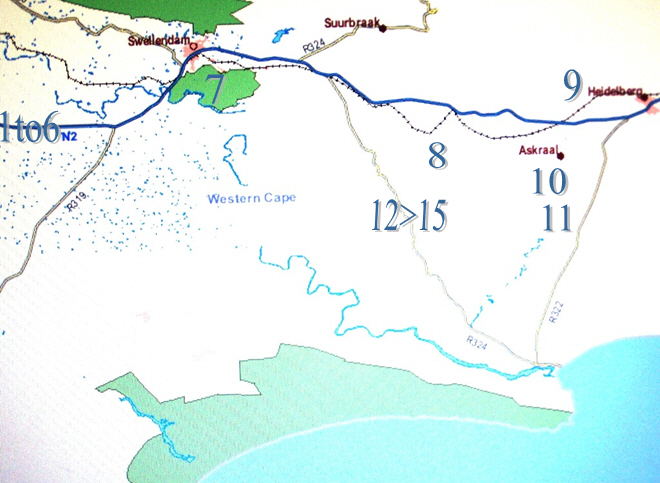
Fig . 1 Map of greater Swellendam area showing population points
A map is provided as Fig.1. Greater Swellendam area to show positions of Haworthia populations noted.
1. MBB7254 Bromberg. (Fig. 2). This really belongs with a large set of populations extending westwards and north along the Riviersonderend Mountains and into the Robertson Karoo. I start with it because it serves as a virtual ‘outgroup’ — and this is because the southern slopes of the Bromberg are different to the northern slopes. They are quartzite (silicon oxides) sandstones and the northern slopes felspathic (other silica salts). This difference in mother material manifests itself in the Haworthia populations and can be used to explain why H. maraisii v.Poelln. in the shales and siltstones at Stormsvlei are so different to H. mirabilis ‘depauperata’ VPoelln. in the quartzitic sandstones less than 1km north. The plants are relatively small and have the reddish color often associated with H. mirabilis south of the Riviersonderend Mountains.
Fig. 2 7254 H. mirabilis. Bromberg.
2. MBB7612 Diamant W (Figs 3 & 4). These are very big plants up to 110mm in diameter with a similarity to ‘maraisii’ by virtue of their blackish-green color and more retused (bent back) leaves. The classic difference of ‘maraisii’ and mirabilis as being separated by translucence absent in the leaves or present, is again flouted in this population as in so many others; and similarly leaf surface texture is variable. The significant thing is that the mountain they are on is an extension of the Bromberg (Riviersonderend Mountain tail) but it is the northern slope and the sandstones are felspathic.
Fig. 3 7612.3 H. mirabilis. Diamant W.Fig. 4 7612.4 H. mirabilis. Diamant W.
3. MBB7626 Diamant Mid (Figs 5 & 6). This population is less than a kilometer east of the receding on a more eastern and thus cooler aspect. The plants are a little smaller, the leaves generally more erect and are a little more reminiscent of mirabilis to the southwest in that spines are better developed on the margins and surfaces than in the case of classic ‘maraisii’ (I am trying to show that ultimately there is no infallible distinction).
Fig. 5 7626.1 H. mirabilis. Diamant Mid.Fig. 6 7626.2 H. mirabilis. Diamant Mid.
4. MBB7718 Diamant E (Figs 7, 8, 9 & 10). This is a huge population on a lower elevation than the preceding and the skeletal habitat is significantly different. It is on the interface of the Felspathic sandstone and shales and siltstones of an older formation while a strong vein of quartzite transects it. In addition there is evidence of an eroded away, and later, ferricrete deposit. The plants are also extremely variable, reflecting the range of divergent surfaces although generally much smaller than preceding. Leaf surfaces are mostly roughish.
Fig. 7 7718 H. mirabilis. Diamant E.Fig. 8 7718 H. mirabilis. Diamant E.Fig. 9 7718 H. mirabilis. Diamant E.Fig. 10 7718 H. mirabilis. Diamant E.
5. MBB7633 Luiperdsberg (Figs 11 & 12). A small population quite high up on a slope of felspathic sandstone. These are small plants and because of quite a grassy habitat the leaves are a little elongated generating the range of forms that I have generally assigned to a little understood or known range of plants and populations in H. ‘heidelbergensis’ Smith.
Fig. 11 7633 H. mirabilis. Luiperdskop.Fig. 12 7633 H. mirabilis. Luiperdskop.
6. MBB7719 Dagbreek (Figs 13, 14, 15 & 16). An established population in felspathic sandstone but marginal to shale and siltstone. Very grassy but with occasional rocky ridges so that the plants are concealed in grass tufts or wedged in crevices. The plants are moderately sized to 60mm diameter and differ in that many have a rather smooth surface with corresponding finer and more closely spaced marginal spines. Some plants develop a shiny opacity about the leaf tips that suggest surfaces perhaps comparable with the smoothness of the H. retusa complex.
Fig. 13 7719 H. mirabilis. Dagbreek.Fig. 14 7719 H. mirabilis. Dagbreek.Fig. 15 7719 H. mirabilis. Dagbreek.Fig. 16 7719 H. mirabilis. Dagbreek.
7. MBB7704 Bontebok Park (Figs.17, 18, 19, & 20). There is an earlier collection (6644) from the ferricrete/shale interface alongside the N2 highway near the northeastern corner of the park, which I considered belonged in a series of five populations that seemed to be neither H. ‘maraisii’ nor generally accepted H. mirabilis. It is on ferricretes but there is some problem how to relate this apparent river terrace gravel with the solid caps of the inselbergs to the east and west. There is nothing common in the general suite of species that could be described as Fynbos on the former and grassy Renosterveld on the latter. The plants are moderate in size; surfaces can be quite rough with a tendency to spination, and the marginal spines quite coarse. Leaves erect or spreading.
Fig. 17 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 18 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 19 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 20 7704 H. mirabilis ‘maraisii’. Bontebok Nat.Pk.
8. MBB7705, 7706, 7713, 7714, 7715 Uitvlugt (Figs 21, 22, 23, 24, 25, & 26). These are five quite independent populations on the one farm that is unusual in that it seems to be on predominantly an earlier variant of Cenozoic ferricretes. The plants are generally related to H. ‘atrofusca’ with rounded leaftips and roughish surfaces. The actual habitats are all different in respect of surface detail and associated vegetation. The plants vary in size between the different populations from small (30mm diam) to large (70mm diam). In the one site where the soil is whitish bentonite, the plants are small and also have a curious similarity of color to that of the clay and the associated plant Gibbaeum haaglenii (austricola).
Fig. 21 7705 H. mirabilis ‘atrofusca’. Uitvlugt NW.Fig. 22 7706 H. mirabilis ‘atrofusca’. Uitvlugt.Fig. 23 7713 H. mirabilis ‘atrofusca’. Uitvlugt W.Fig. 24 7713 H. mirabilis cf ‘magnifica’. Uitvlugt W.Fig. 25 7714 H. mirabilis ‘atrofusca’. Uitvlugt E.Fig. 26 7715 H. mirabilis ‘atrofusca’. Uitvlugt N.
9. VA-RW07-27 (Figs 27, 28, 29 & 30). This population located by M Mougin, was reported to me by D. Van den Abbeele. It is much closer to Heidelberg and obviously in a ferricrete related habitat. The plants are all very ‘atrofusca‘-like with some evidence of the pointed leaf-tips that would have been ‘magnifica’. The surfaces have the slightly riffled texture that one associates with ‘atrofusca‘ rather than the tendency to roughish surfaces or almost spinescent tubercles associated with, but not peculiar only to ‘magnifica’.
Fig. 27 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 28 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 29 VA-RW07-27 H. mirabilis cf ‘magnifica”. Basryhoogte. M.Mougin.Fig. 30 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. D. Van den Abbeele.
10. MBB7728, 7729 Lilliendal (Figs 31, 32, 33, 34 & 35). The significance of these two populations must be emphasized. The place is the Karringmelks River southwest of Heidelberg. This river drains into the Slangriver that flows then southwest into the Breede River. It is quite a different drainage basin to the Klip and Duiwenhoks rivers that drain the area north and west of Heidelberg and enters the sea south of Heidelberg. Matjestoon and the origin of H. ‘heidelbergensis var. toonensis’ is south of Lilliendal and on the same drainage system. The plants would at the time have made it difficult to imagine H. ‘heidelbergensis’ as a discrete system, and these plants force a direct comparison with H. ‘maraisii’, if that system could be generalized. The plants are small, dark-green, rough and with a degree of spination on the surface tubercles. Surfaces may be opaque or translucent. There is a striking resemblance of the seedlings to H. floribunda. The populations are about 1km apart. Both are on Bokkeveld but at the second population it is curious to observe that the plants are not on the ferricrete only meters away.
Fig. 31 7728 H. mirabilis. Lilliendal.Fig. 32 7728 H. mirabilis. Lilliendal.Fig. 33 7729 H. mirabilis. Lilliendal.Fig. 34 7729 H. mirabilis. Lilllendal.Fig. 35 7729 H. mirabilis. Lilliendal.
11. JDV87-3, MBB7165 Matjestoon(Figs 36, 37 & 38). This is one of the original populations known to G. G. Smith and it is an uncomfortable fit anywhere on account of the length and spiny-ness of the leaves. That the plants were placed in ‘heidelbergensis’ is a reflection of the doubtful and composite nature that the species gained by default. There is a semblance of similarity to H. rossouwii but what may be more notable is that at Dassieklip to the southeast there is a very aberrant population that is assigned to H. floribunda simply for want of a more likely category. I say this is notable because in my field observations of Haworthia there is a very strong and predictive geographic continuity between populations. This is evidenced in this general discussion.
Fig. 36 JDV87-3 ‘H. heidelbergensis var toonensis’. Matjestoon.Fig. 37 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.Fig. 38 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.
12. MBB7736 Koppies (Figs 39 & 40). These are small dark-green plants with strongly deflexed leaves and a little unlike an earlier record from 2km west (MBB6879 ex E. Esterhuizen) where the plants have more pronounced surface tubercles becoming quite robust and dark in cultivation. The habitat is among white quartz occasionally associated with ferricrete.
Fig. 39 7736 H. mirabilis. Koppies.Fig. 40 7736 H. mirabilis. Koppies.
13. MBB7723 Stuurmanskraal (Figs 41 & 42). These rank among what might be regarded as the general body of H. ‘maraisii’. Small, dark roughish plants with strong evidence of floribunda in the way the leaf-tips tend to be flat, twisted and rounded. The habitat is Bokkeveld shale.
Fig. 41 7723 H. mirabilis. Stuurmanskraal.Fig. 42 7723 H. mirabilis. Stuurmanskraal.
14. MBB7734 Stuurmanskraal N (Figs 43 & 44). This is a previously known population similar in its responses to the Koppies plants (MBB6879) that become quite robust and dark in cultivation. The habitat is white bentonitic clay that develops under the ferricrete sills from decomposure of underlying shale.
Fig. 43 7734 H. mirabilis. Stuurmanskraal.Fig. 44 7734 H. mirabilis. Stuurmanskraal.
15. MBB7731 Oudekraalkop (Figs 45, 46 & 47). Finding this population at all suggests that there may be many more evading the eye. The plants are small and dark and completely hidden among dense but short vegetation on a rocky scree-like area. As in other populations one is compelled to look at every plant and consider its departure from a hoped for norm for the plants there. Three illustrations hardly do justice to the variability and this true for all these populations discussed here.
Fig. 45 7731 H. mirabilis. Oudekraalkop.Fig. 46 7731 H. mirabilis. Oudekraalkop.Fig. 47 7731 H. mirabilis. Oudekraalkop.
Conclusions My despair over the years that I have been involved with Haworthia, is that the classification model and nomenclatural system we use, does not seem to allow me the freedom to express what I have seen and experienced. My argument has been that we are dealing with a biological system in which things that seem different are the same; and conversely things that seem the same are different. It is not a problem to apply a formal classification system except that a ranked or tiered system suggests quantum differences too, and this is simply not true. The greater problem is that most plant enthusiasts, and those who feel strongly about classification and names, confuse the reality of the sameness that suggests ‘species’ with the sameness that plants of different species may exhibit.
It is with a source of considerable curiosity that I now take note that a taxonomist is one who classifies by appearances and a systematist one who classifies by evolutionary pathways. In truth I never considered that classification was anything but an attempt to accomplish both aims so that meaningful names could be established. The issue is clouded in complexity when it is considered that neither taxonomist nor systematist may be sure when a species is a species. Probably hence the observation that species (binomials) carry a reality they do not possess. My proposal, supported by the observations in this article, is that we adopt a far more informal and flexible approach to the use of all the names we now have at our disposal. This suggestion is also supported by an experienced taxonomist/systematist/botanist who wrote: “The informal system that I prefer allows for endless forms that are identifiable by their locality and do not clog up the literature. Essentially it is a reversion to the polynomials of the pre-Linnaean era.”
My contention is that the only rational classification of the above 20 populations is within a greatly expanded concept of H. mirabilis that includes H. maraisii, H. magnifica, and H. heidelbergensis. This corroborates the observations reported in the chapters in Haworthia Updates and specifically Chapter 6, Update 2,1:50-79 “How to understand H. mutica var. nigra”, and Chapter 1, Update 3,1:1-87 “H. mirabilis and the interface with H. maraisii”.
Acknowledgements The population MBB7704 was shown to me by Ms. Tineke Kraaij of RSA National Parks Board. The concurring botanist is Dr John Manning, who has authored many botanical works and who is best known for his familiarity with the highly complex Cape bulb flora. Dr. Syd Rhamdani introduced me to the new direction that molecular biology is driving plant systematics. Gerhard Marx has been a great help in getting me to resign to the unrealities of popular taxonomy. Steven Hammer, considerably more successful than I have at adapting to the fact that our perceptions about many things are very different to that of others, helped me with the manuscript. Rhett Hiseman of Cape Nature, Jaap Viljoen, Henniie and Elma van Deventer of Koppies, the Odendaals of Dagbreek, Diamant and Dankbaar, Alfred and Andrea of Lilliendal, Dirk Papendorp of Uitvlugt, D. van As of Stuurmanskraal, Anna and Arno Steenkamp of Oudekraalskop (Anna’s Farm) are sincerely thanked for information and/or access to plants. ♦
It has long been my stated contention that H. turgida is in fact a rock face ecotype as opposed to the solitary flat growing H. retusa. Thus one should expect the multiplicity of forms that are found between, and consequently superfluous to say within each, these two primary types. There are problems outside of this and I will deal with those in the Chapter Haworthia enigma. Here I am simply going to present pictures representing plants in nine populations of the species. Most of these populations are of the “typical” solitary form and they all demonstrate variation to greater to lesser degree. Perhaps some special mention should be made of the element H. mutica var. nigra. I have written at length about this and in doing so strayed widely into H. magnifica and its var. atrofusca (both falling now under H. mirabilis). This is because it is quite certain that there is an element of interaction in the field between the prime elements H. retusa and H. mirabilis that this summation is intended to expose. The first known H. mutica var nigra from Kransriviermond is possibly the product of such interaction, whereas all the subsequent collections from northwards and westwards are now perceived by me to be variants of H. retusa and H. retusa ’turgida’ (to use a more informal and flexible way of communicating).
1.MBB7776 Pienaarsrivier. This is a population just west of Kweekkraal and the agglomeration of populations that I dealt with in a series of articles covering the diffuse nature of ‘nigra’. The population was first brought to my attention by Etwin Aslander but finally shown to me by Kobus Venter. It is extraordinary that it is not better known in the literature and again a vivid demonstration of how thoroughly one has to explore to uncover the mysteries of what constitutes species. It has always been difficult to adequately illustrate plants and probably the very weak appreciation we have for variation is precisely due to the fact that often species ere even described without any accompanying illustration. If there was one it was taken to be the exact image of many such that could all go by the same one name and this a Latin binomial. This is the paradigm that needs to change.
Pienaarsrivier – truffeling.
At Pienaarsrivier, the habitat can be described as an erosion product of the ferricreteinselbergs that are so characteristic of the lower areas of the Southern Cape. However, it is an extremely complex situation as there are high and low level gravel terraces as products of direct erosion off mountain sides, river gravels as a product of countryside drainage and then the iron rich ferricretes themselves as the products of deposition along groundwater movement. Here at Pienaarsrivier there is dense Renosterbos and grass among fairly large sized stones. The plants are abundant and they flower in October. I have elsewhere shown pictures of H. retusa from both eastwards at Kweekkraal (two populations) and from Heidelberg itself (many). The Pienaarsriver plants tend to have a very purplish hue and often the leaf tips are very rounded (mutica!) ♦
2. MBB7754 H. retusa ‘turgida’. Brakkekuil. What is most significant about this population is its whereabouts that highlights the overwhelming importance of distribution and geography. The drainage systems (or parts of them) of the Southern Cape drain southwards from the mountains to the sea e.g. Gouritz, Goukou, Duiwenhoks and Lower Breede. These are important especially when it comes to the habitats in the way of exposed rock and steep faces that favour plants requiring skeletal soils. Brakkekuil is on the Slang River that drains southwestwards from near Heidelberg to flow into the Breede River near Malgas. H. retusa ‘turgida’ has not been reported for this entire river system before, while it is present on the Breede River and even westwards at Bredasdorp. So the Brakkekuil population is significant and also significantly different. The plants are neither strictly solitary nor greatly clump-forming and it is not really surprising as this mirrors what happens with H. cooperi in the Eastern Cape in situations that are neither fully cliff face nor plain. The Brakkekuil plants are on the surface of a rocky shale knoll with plants enduring direct exposure to northwestern sun as well as obtaining refuge in the more vegetated and protected slightly southern aspect. It is quite difficult to make reference of individual plants to Latin names, in that variation is already ensconced in the existing system viz. ‘longibracteata’. I gladly concede that all the old names, as Rowley has suggested, can be paraded out again and made use of. In fact I have also said that this is how the contribution of Breuer and Hayashi can be fruitfully used. For my reality this population is H. retusa ‘turgida’ Slangrivier. It is quite the most variable population of the ‘turgida’ side of H. retusa that I have ever seen and there are plants that resemble the more sandstone associated variants (‘caespitosa’) at, say, Tradouw Pass as well as individuals that compare with some of the other populations I will cover from the ferricrete inselbergs. Another very significant observation is the similarity of some plants to those that can be found in H. mirabilis ‘paradoxa’ that is not very far away to the southeast at Vermaaklikheid. There is no doubt that if a full and real understanding of natural systems is to be found it will lie in the realization that even my suggestion favouring a “super species concept” may be conservative. It is actually curious how my treatment of that has been met by readers who have been kind and considerate enough to communicate with me on the issue. The ‘super species” proposal actually comes from Prof. Canio Vosa. It is and was not, any attempt to confound anyone or obfuscate the issue. Prof. Vosa is directly addressing the issue that we have a classification that is a sorry marriage of scientist and layman user groups – both ignorant of the full extent of the field situation.
3. MBB7556 H. retusa ‘longibracteata‘. Dassieklip. There are ‘turgida’ populations all along the Duiwenhoks River. Some of these could be ‘turgida’ and some could be the variant ‘longibracteata‘. These at Dassieklip are close to the latter and are reminiscent of some of the large slender leaved plants one finds close to Heidelberg itself
However, the two plants illustrated are a few hundred meters west of the river and survivors of road materials quarrying. There are many more plants along the river itself and also at Brakrivier between Dassieklip,and Vermaaklikheid. But south of Dassieklip the river drainage passes from shale to calcrete and something really interesting may occur on the interface of these two geological formations northwest of Vermaaklikheid. Curiously no H. retusa is recorded at all between Brakrivier and Stilbaai to the east and this may be due to the calcrete substrate and the absence of suitable habitat.
4. MBB7758 H. retusa ‘retusa’. Skietbaan. Riversdale. This is a very well known population and one of many in the general area extending slightly south and then west rather than east, although there is a population in the rocks of the Goukou river itself to the east. The plants are on a shale substrate and huge with again the expected variation that has been so obscured by the limited illustrative material of the historical past. The habitat is highly disturbed and it is in fact surprising that it has survived here so successfully. A road and a fence cross the site that follows the scale of the relatively confined areas the plants are usually found in. There are a heap of epithets that could be mobilized to personalize these plants and even ‘solitaria’, ‘geraldii’ and ‘fouchei’ could probably find application despite being out of true context.
5. MBB7240 H. retusa. Skeiding. This population seems to be very relictual and I did not see many plants. I include it just to indicate that these ferricrete associated forms are here and they are trespassing into the series of populations that I discussed in the ‘mutica var. nigra’ story. That I now could refer to these as H. retusa ‘nigra’ should be seen as evidence of the flexibility and pragmatism that is really required to allow acceptance of a nature that we can understand.
6. JDV93/57 H. retusa. Plattekop. There are several populations in the ferricrete inselbergs. They are inclined to shiny yellow colours and rather longer, more erect leaves than one would expect in H. retusa. In fact one can see a drift from the latter to ‘longibracteata’ in the broader context that I originally used the name to really suggest populations that were neither ‘retusa’ nor ‘turgida’. This particular habitat is unusual in that the rock is larger than usually accepted by the plants and there is quite an infusion of the white clay of decomposed shale. The plants seemingly, and consequently, take on a rather powdery and milky appearance.
7. MBB7772 H. retusa. Platkop Annex. Where Skeiding is west of Heidelberg, Plattekop and Platkop Annex are east of Riversdale. These and the next two populations form an important extension of the distribution of the species eastwards and to perhaps emphasize a distinction between H. retusa as it grows on the inselbergs as opposed to H. retusa ‘turgida’ that is riverine.
8. MBB7765a H. retusa. Plaatjieskop. These plants are very similar to the two preceding. A similar population was shown to me by Japie Dekenah at Rooikoof to the north. What is significant here is the presence of hybrids with H. floribunda (fig. 74) and that in turn by the fact that H. retusa flowers in spring and H. floribunda in summer. Such hybrids are known at Cooper siding between H. pygmaea ‘argenteomaculosa’ and H. floribunda and also at Blackdown, N. Heidelberg between H. retusa ‘turgida’ and H. floribunda. Because I consider that there is a deep-seated ‘problem’ of interaction like this between major role players, I will discuss this again elsewhere.
7766H.retusaXfloribunda. Plaatjieskop JDV 0682Habitat H. retisa. Plaatjieskop JDV 0711
9. MBB7771 H. retusa. W Soetmelksrivier. There are seemingly always surprises in Africa (“always something new”) and this is one. Not only does H. retusa grow in direct association with H. mirabilis cf. ‘splendens’ but the habitat encroaches on the highly ferruginous rocks that form the hardest layer of the ferricrete inselbergs. It is really only two populations in the Kweekkkraal area where haworthias can be said to be truly in this kind of rock. ♦
I trust that it is clear by now that I consider H. magnifica, H, maraisii, and H. heidelbergensis to be essentially the same one species and emphasize that it is really a self-evident truth that species are complex systems and not simply a randomly occurring set of similar looking things. In a recent manuscript submitted to Haworthiad, I wrote about new finds elaborating this point of view. The essence of this chapter is to discuss exploration focused on clarifying the position in the eastern area between Riversdale and the very problematic H. pygmaea “squadron” that I have also discussed at length. Prior to the trip I had an ongoing communication with Gerhard Marx and we agreed that H. ‘splendens’ was in fact better fitted in H. mirabilis, the major obstacle for me being the fact that there was no field record nearer than Riversdale itself to substantiate such a view. I do, however, want to here also record two further populations east of the Breede River and other populations of H. mirabilis west of the Breede and south of anything previously noted.
1. MBB7442 H. mirabilis. Dankbaar. This population is of small plants but they are not substantially outside of anything noted in any of four compass directions although they do have their own look about them. I found it quite odd that Cameron McMaster had sent me pictures of a population of H. mirabilis from Fonteinskloof to the southwest of Stormsvlei that reflected the same co-similarities with H. mirabilis and H. marasii that I had recorded at Rondeheuwel south of Stormsvlei. The Dankbaar plants are small versions of this and of course tie up with both older and newer (MBB7704) records for the Bontebok National Park.
2. MBB7744 H. mirabilis. Bontebok Park. Having written that, we did in fact locate still another population and of course it looked different as the area where it occurs had been recently burned and being on a northwest aspect the plants were very exposed and even more cryptic than usual.
3. MBB7749 and MBB7751 H.mirabilis. Kadies Landing.: These two populations are in my opinion tremendously significant. While H. mirabilis has been reported from this the east bank of the Breede River, it was a very small form such as I recorded for the lower west bank (Ziekenhuis, Aalwee). The plants we located are down the river from Malgas but on the east bank. They are not any smaller than the general size of H. mrabilis anywhere in the central area of the range and highly similar to the plants especially in the Bontebok park quite some distance upstream from a very winding river course. There is thus now a very substantial connection and continuity from west to east and into ‘atrofusca’ that occupies the inland upper areas from west of Heidelberg to east of Riversdale.
4.MBB7753 H. mirabilis ‘toonensis’. Brakkekuil. It is important to note how I have named this as I have no intention of going through the formalities of nomenclature to change anything. I have been terrorized to my limits by “law” and I think that ethic and common sense can now prevail. ‘Toonensis’ is only known from Matjestoon southwest of Heidelberg and upstream along the Slang River. As I explained in the chapter on H. retusa, the Slang River has its own peculiarities and besides which it is quite unexplored. So it is no surprise to find this rather smaller version of H. mirabilis in a second population, differing from the general body by more and smaller more slender leaves. Naturally there is considerable variation within the population too. The differences relate to the problem of the Dassieklip and Vermaaklikheid areas where H. retusa and H. mirabilis ‘paradoxa’ and the problem should not be deferred to me as a bad taxonomist but as the reality that there is a major geological contributor to the problem of difference viz. shale vs calcrete. It is quite obvious in the series of pictures that one can see resemblances to ‘turgida’ and ‘paradoxa’.
5. MBB7761 H. mirabilis ‘splendens’? Plattekop Farm; MBB7762 do. W Platkop; MBB7763 do. E Plattekop Farm, MBB7765 do. NNW Platkop: MBB7769 do. Toringskop; MBB7770 do. W Soetmelksrivier. I am grouping these together and using the epither ‘splendens’ in an apparently loose and provocative fashion. This is to deliberately hew away at this mental sludge that isolates the really magnificent and incredible plants from their equally interesting but less glamorous close relatives. These populations are all on ferricrete inselbergs in virtually any of the three or four main geomorphological derivatives. The official geosciences description of the Riversdale area makes specific mention of the difference between these and the Soutpan (true ‘splendens’ habitat) by saying that the latter is a pan-area and the relatively thin ferricrete layer apparently developed in a marshy area and close to the surface (opposed to the inselbergs as deeper deposits in underground water systems in sandstone?). These populations are what we predicted and they are the direct link between the most easterly known only true H. mirabilis ’magnifica’ at the Frehse Reserve just south of Riversdale. The element ‘magnifica’ is a phantasm (available as an optional cultivar name as there are many plants in that parent population that do not fit the description?) based on an initial description and limited illustration and has been maintained by myself while trying to resolve the issues in my own mind. It has been kept alive for me by populations at KomseRante and Kruisrivier that I am about to further explore after writing this particular piece, and that will be a separate chapter.
I cannot say that I have resolved the entire complex puzzle but these populations validate the decision reached with Gerhard that it “feels” right to really include ‘splendens’ with H. mirabilis rather than with H. pygmaea. This is where the actual relation of H. retusa and ‘turgida’ takes on another dimension similar to the H. mutica situation to the west where it is confounded by H. mirabilis.
The most significant population is the last noted one viz MBB7770. Not only are the plants larger than in other populations they also manifest the general pattern in Haworthia of “flow”. They resemble true ‘splendens’ that much more than the other populations because they are also geographically closer. Another oddity is their occurrence in the pure ferricrete at the top of the inselberg that I discussed under H. retusa. The most extraordinary thing is that this population shares habitat with H. retusa and while I cannot say there are hybrids, it is evident from some of the plants that hybridization may well have occurred despite the big differences in flowering times. ♦
Again this piece is written against the background of a detailed discussion in Haworthia UpdateVol. 2. Again I am not able to say the situation is fully comprehensible and neither do I want to encourage the daffy view that nature is just too much for us all. It is really curious that this species is woven into the fabric of H. mirabilis and also into that of H. chloracantha, H. parksiana and H. variegata. New finds have not clarified the picture so much as added another dimension to an extraordinary display. Not that H. floribunda is a spectacular species. In the field it can be extraordinarily cryptic and obscure while in cultivation it is an unlikely favourite. I do not want to repeat what I have already written while I hope that this will not contradict that either. H. floribunda seems to occupy a clear niche along the base of the mountains between Albertinia in the east and Swellendam in the west. It occurs as discrete from any other species although hybrids with both H. retusa and H. mirabilis do occur. South and west of Heidelberg it seems to lose its identity within H. mirabilis and then emerges briefly in a limited area near the Potberg in the southwest in a ‘mirabilis’ context as well as in H. variegata context. At Klipfontein farm at the western end of the Potberg it seems to be recognizable in relatively the same form as the very original description. But let us look at new information.
1. MBB7722 H. floribunda ‘major’. Appelbos. This is just west of Swellendam and we came across this locality while trying to confirm a very old record of the form ‘dentata’ once brought to me from Buffeljagts very nearby. The plants are quite large and have a clear green colour quite unlike the normal dark green or purplish hue of ‘typical’ floribunda. It should not be lost to the reader that this same colour difference characterizes the difference between H. retusa and H. mirabilis. Or that both colour variation is apparent in H. mirabilis ‘paradoxa’. Kobus Venter found a similar population northwest of Swellendam and further so than my rather poorly noted collection of proliferous dark plants with predominantly pointed leaves. In fact this population is hidden in its citation in my Revision under H. variegata. This will perturb those readers who are imprisoned in the paradigm of rigid similarities that define species. The Appelbos plants generally have the straight acuminate leaves that H. variegata has, but before arguing the issue and without actually properly looking at the whole H. variegata milieu, here are two connected records …
2. MBB7738 H. floribunda ‘major’. Swellendam. These plants were in fact small when first collected and in cultivation grew so large that I coined the name ‘major’ for them.They do still exist in a very small and disturbed area close to gum trees but curiously in moss free of leaf litter. I did also find them a little further away in a more grassy area where they are/were more typically small and dark coloured. I should note that I also recorded this ’dentata’-like version within the Bontebok Park close to where H. mirabilis occurs and I am still committed to again finding that population in the light of this new material.
3.MBB7774 H. floribunda ‘major’. Swellendam. We were concerned about the disappearance of an interesting form of H. minima from the area and in extending our search for plants came across H. floribunda in gumtree leaf litter about 400m east of the previous H. floribunda population. At first glance I did a double-take and then another because I actually thought I was looking at a slender form of H. marginata as occurs at Drew. The plants we saw were really large forms of H. floribunda with pointed leaves exceeding 100mm in length. They were quite proliferous and not all nearly as big as the first surprise ones. We also located smaller plants in numbers on the opposite side of the road also under gumtrees.
4. MBB7708 H. floribunda ‘dentata’. Goedverwagting. Apart from the Buffeljagts and Bontebok Park records there is an inexplicable hiatus in the appearance of the species west of Heidelberg. This is probably simply due to inadequate exploration that these new records now expose. It is quite evident that in the general area north of an imaginary line connecting Swellendam and Albertinia, H. floribunda does occur independently and in near company of H. mirabilis and the latter invariably in the form of ‘atrofusca’. This Goedverwagting record fills this gap as an association with ’atrofusca’ that pushes southwards.
5. JDV93/58 H. floribunda ‘dentata’. RoseInnis Drive; JDV93/56 do. Plattekop; MBB7760 do. Witkleikop; MBB7764 do. NW Platkop; MBB7767 do. Plaatjieskop. Being so cryptic may be a reason for the non-record of this species. It is associated with the ferricrete inselbergs and has the really curious association with H. mirabilis already remarked upon. In these four records, three of which are new, the plants occur independently of other Haworthia species and there is no obvious habitat preference that suggests why this is so. I also think that there is significance in which the characteristic rounded leaf tip is replicated especially in the juvenile leaves of H. mirabilis. The last cited record is the dramatic one. The plants are in very close association with H. retusa in two small geomorphologically different and adjacent sites, and we observed what seem to be reciprocal hybrids.
JDV93/58 H. floribunda ‘dentata’. RoseInnis Drive
JDV93/56 H. floribunda ‘dentata’. Plattekop.
MBB7760 H. floribunda ‘dentata’. Witkleikop.
MBB7764 H. floribunda ‘dentata’. NW Platkop.
MBB7767 H. floribunda ‘dentata’. Plaatjieskop.
Hybrid MBB7768 H. floribunda x retusa, Platjieskop
There is not much to write about this less glamorous of plants. It is one of the Robustipedunculares. It is very widely distributed and even occurs north of the Langeberg Mountains in the Little Karoo – as does its sister species H. pumila. The pictures cover a huge range of variants from DeMond in the southwestern area to Riversdale. I found them all fascinating. Mostly so perhaps the very few large globose plants we saw at Kleinberg near Malgas (Diepkloof). But the plants at Koppies are really interesting because they were overlooked for so long. Koppies is the second known home of H. serrata (now H. rossouwii) and we came to know it also as a refuge for a small population of H. marginata. It was while we were investigating this that Hennie van Deventer casually pointed to plants of H. attenuata var caespitosa in his garden and said they were also on the farm. Because of our surprise and of course doubt, he took us to show the plants that he said he had not looked at nor thought of for 20 years – and there they were. A very small population of large dark coloured plants.
Also interesting were the very small solitary plants on the farm Sandfontein in the Slang River Valley. At Klipheuwel marginal to the coastal calcretes we also came across the species. Initially we saw only two plants while searching for ‘retusa-like’ plants. Failing to find anything else in a very severly grazed field we thought we would return to commiserate with the few H. minima we saw and enjoy the ambience we always feel in the presence of Haworthia. One can barely explain how cryptic these plants are and how easily they avoid detection. We found another 40 plants in the same area abut 10m diameter where we had found but two a short while earlier.
I found it very curious that in our very extensive exploration of the area east of Riversdale, that we saw neither H. minima nor H. marginata. Gasteria was also conspicuous by its absence and I remark on this because I do not think that taxonomy can remotely afford to ignore fundamental patterns in the general environment. ♦
MBB7737 H. minima, Koppies.
MBB7747 H. minima, Sandfontein.
MBB7757 H. minima, Klipheuwel.
MBBsn H. minima, Bontebok National Park.
MBBsn H. minima, Stoffelsrivier.
Others
H. minima, VermaaklikheidMBB7100 H. minima. Nuutbegin, Swellendam.MBB7519 H. minima. S Kleinberg.MBBsn H. minima. Diepkloof.MBBsn Haworthia minima. De Mond
What always comes to mind as I travel through the countryside is the realization of just how much there is to explore. It takes only an hour of driving from Cape Town to get to the start of Haworthia habitats from any of the three main routes inland. The roads do not always take in the best routes in respect of suitable or inviting habitats to explore, and besides there is the question of landownership and permission for access. In recent months my wife and I decided to really make an issue of new exploration and investing time and energy in contacting landowners and looking at places that we have ignored before because of access difficulties. The result has been a massive set of new finds in respect of populations not previously known to us. Having other interests such as in Drosanthemum and chameleons has also led us into places we might not have otherwise ventured.
This particular note arises from another chance find. Heidelberg is very rich as far as Haworthia is concerned and I have written quite a lot about the area or referred to populations there. It is a very important area for both H. retusa and H. mirabilis, as well as for H. floribunda and one assumes that the area has been fairly well explored. The Duiwenhoks River offers much suitable Haworthia habitat and I can count at least 30 populations along the drainage system that can be referred to those three species systems.
While checking on two populations of H. retusa ‘nigra’ (the informal way in which I now cite my own combination H. mutica var nigra derived from a G.G. Smith record south of Heidelberg), we took the opportunity to venture into a field from which we had on the previous occasion been excluded by the presence of ostriches. This was on the farm Diepkloof that I have cited as Morning Star because the two units are now farmed by the same owner and there is also confusion with farms on the east bank of the river also referred to as Diepkloof. The populations we looked at were of H. mirabilis and H. retusa ‘nigra’. The former appears to incorporate H. floribunda in all its populations down the Duiwenhoks River to as far as the change from Bokkeveld shale to Calcareous limestone near Vermaaklikheid. H. retusa var turgida occurs in various guises along the cliff habitats along the river while in non-riverine flatter areas H. retusa manifests.
We were specifically looking for H. mirabilis or H. retusa when my attention was drawn to an old flower stalk. There are a few bulb species that have a very similar dry inflorescence as Haworthia so I was a bit cautious when I bent closer to examine its source. With my search pattern set on the expected, I at first had difficulty focusing clearly on what was there before me – a small form of H. rossouwii (see MBB7803 Figs 1-9). We started looking wider and found a few plants under bushes until our eyes began to get the message as to what we should be looking for. To our amazement plants began to spring up all over the place among the dense pebbles.
Why Haworthia rossouwii? Firstly I have no sympathy with contending classifications that set out to do nothing more than weakly describe variation in the genus in terms of Latin binomials. My contention is that species are complex systems that need to be understood as highly variable elements that can respond to environmental differences and changes. H. rossouwii is already known to have an unusually wide distribution range as far as Southern Cape species were concerned. It is known from north northwest of Bredasdorp and is now known to us at 8 localities southwest of Heidelberg. Essie Esterhuyzen is reported to have found it at Voorstekop close to the N2 about 90km from the most southwestern population.
I relate “H. elizeae” to H. rossouwii. This was probably first recorded by Derek Tribble from the Bromberg mountain near Stormsvlei, about 40km north of the Bredasdorp populations of this species. There it is in sandstone, unlike the ferricrete and shale habitats the species is found in elsewhere. The Bromberg is also about 90km west of Morning Star. The similarity of the two populations is mostly in respect of size and the flowering time (without suggesting that this is inarguably evidence for sameness). The Morning Star plants are less proliferous in habitat at least, and this may simply be because slope and substrate so strongly relate to vegetative proliferation. The plants may get to as much as 50mm diameter under the protection of bushes, but in the exposed pebbles they average between 20 and 30mm. The colour seems to be darker green than the yellowish shade of the more general H. rossouwii but we saw plants in their stressed summer garb where there is a strong shade of purple in the plants. It will be interesting to see what the plants look like in the winter.
That this Morning Star population can so readily be allied with the Bromberg population, for me strengthens the view that the ‘systems’ approach to which I more strongly lean, is correct. What does worry we is that the formality of the nomenclatural systems, both botanical and horticultural, becomes very problematic. Essentially the infraspecific categories of subspecies, variety and forma are group categories derived in an era, and for plants, where less variability was known. In many species it has been relatively easy to add horticultural names and this may be because the flowers convey the interest and significance of difference. Where this is transferred to vegetative differences that are so dramatically affected by growing factors it will be very difficult to police. There is of course the additional problem of a taxonomy that is so personalized. Will anyone actually be able to establish when a published name is/was effectively a single plant description and hence a cultivar name as opposed to a group name? My view is that for the system to work, variability has to be limited and furthermore there is a complication where one now relies on vegetative characters that are so dependent on growing conditions – unlike the flowers of conventional horticultural cultivars. In Haworthia we may have an unusual problem in that many very similar looking species are involved and cultivars (single clones, bred or selected from nature) from different species may be very similar. I cannot personally see that anyone is going to successfully, meaningfully and usefully disentangle group and individual plant names. In H. rossouwii there might not be a problem as there is not a great deal of difference among the plants of the main body of populations that would excite the taxonomist. There is no doubt that there is enough variability to invite selection by the ardent grower who may develop a special interest in the species. The variant ‘elizeae’ and now ‘Morning Star’ (an informal and workable way of distinguishing the plants) may present problems in that the only way that they might be distinguished is if the collecting data remains attached, and with, any plant cultivated.
Acknowledgement I am very grateful to Coetzee and Sarita Uys of Morning Star, to access Diepkloof, and to Gerhard Marx for useful comment and opinion. ♦
In a recent set of articles published by the Haworthia Society I wrote the following in connection with H. floribunda… “MBB7738 H. floribunda ‘major’. Swellendam: These plants were in fact small when first collected and in cultivation grew so large that I coined the name ‘major’ for them. They do still exist in a very small and disturbed area close to gum trees but curiously in moss free of leaf litter. I did also find them a little further away in a more grassy area where they are/were more typically small and dark coloured. I should note that I also recorded this ’dentata’-like version within the Bontebok Park close to where H. mirabilis occurs and I am still committed to again finding that population in the light of this new material”.
In connection with H. mirabilis, I wrote … ”The Dankbaar plants are small versions of this and of course tie up with both older and newer (MBB7704) records for the Bontebok National Park. 2. MBB7743 H. mirabilis. Bontebok Park: Having written that, we did in fact locate still another population and of course it looked different as the area where it occurs had been recently burned and being on a northwest aspect the plants were very exposed and even more cryptic than usual.”
Since those finds and comments, we have again been back to the Bontebok Park. It is of course a large area of largely recent geological origins and therefore mostly ferricrete and riverine gravels. The lower lying areas are seasonally flooded. Being unsuitable for cropping or grazing, the area was handed over to the conservation cause (thank goodness for non-arable land). H. venosa occurs on the exposed Bokkeveld Shale cliffs along the Breede River, and H. minima is present in small scattered populations. H. marginata has virtually disappeared from thePark.
It is quite difficult to identify suitable Haworthia habitats as even in the seasonally flooded areas there is much gravel and consequently good drainage. It was in such a site that I seemed to remember seeing the record for H. floribunda cited in my Revision. Our searching has been very limited as it is quite difficult to explore the park with the restraints of accompanying rangers however willing or enthusiastic and knowledgeable. This is because I personally need mental space and physical freedom to walk where my intuition takes me. On this occasion we first went to an old entrance to the Park from the town where we thought H. marginata was first recorded. I was very disappointed to find that the hillside there had suffered the same mindless engineering fate that hit the gravel hills around Worcester. These were ”strip-mined” to obtain road materials. Of course this may have been necessary, but not the mindless extent of the surface scraping when some consideration for simply good housekeeping would have limited the desecration of the landscape and demanded some degree of resortaion.
We moved some way west to avoid the disturbed area and eventually found H. mirabilis (see MBB7805 Figs 1) about 500m east of the MBB6513 record of 1996 when I was writing the Revision, and illustrated elsewhere. That record came from a more distinct interface with the riverine shale while the new record was in more sandstone gravels from an old river bank. The plants were rather smaller than in 7704 with quite slender leaves which were also fairly toothed. There was little evidence of the flattened rounded leaf tips that characterize H. floribunda. We drove around the park trying while I was wracking my memory banks for where I might have seen H. floribunda so many years ago and eventually was drawn to where Tineke Kraaij had on a previous visit shown us H. mirabilis on a burnt zone (see MBB7704 Figs 2). These are new images to the original published ones, because after 6 months the plants had largely lost the burnt look but also being in flower they were far more visible. The leaves were more elongate and pointed than suggested by the photographs we took on the first visit and the plants reminded me very strongly of what I named as H. floribunda var. dentata. I also add here images for H. mirabilis (see MBB7744 Figs 3) from another population approximately midway between the previous two. Here the area had also been burnt as part of the management plan for the Park, but the plants themselves do not seem to have been affected possibly because there was a lighter fuel load that at the 7704 locality at the time of burning. The plants were more “maraisoid” and similar to the very first record of mine for this species in the Park (MBB6513). That collection I often noted among a set of 4 or more populations extending from Riversdale westwards to Riviersonderend that confounded any distinction between H. magnifica, H. maraisii and H. mirabilis.
All things considered, it seems highly unlikely that H. floribunda could possibly co-exist with H. mirabilis other than in the present known configuration where we have H. floribunda var major close to and north of the N1 highway near the town and inland, whereas H. mirabilis is further south. This geographic and presumed taxonomic relationship seems to hold at Heidelberg and Riversdale with some anomaly at Albertinia down the Gouritz river. This is where H. mirabilis is absent (with consideration for the retusa/pygmaea conundrum) and where H. chlrorocantha or H. variegata may be confounded in H. floribunda (or vice versa!). ♦
If the name “H.enigma” applies to the plant (or plants) from east of Riversdale at Komserante, it is a name that I really do not advise to be taken seriously from a botanical point of view. It is useful at population level and to demonstrate the nature of classification difficulties but it is a minor problem in so far as those difficulties extend. The plants were first shown to me by J. Dekenah on the same day that he also showed me ”H. magnifica” in the Nature Reserve just south of Riversdale that is less than 3km away. My impression then was that it was the same element even if it did look a bit different. The plants are quite large (to 70mm diameter), fairly tubercled and often with lines in the upper retused area of the leaf face. While I originally classified “H. maraisii” under “H. magnifica”, I later separated them because it seemed so incongruous to include all the variants of the western “H. maraisii” with the few populations of “H. magnifica” then known. Also, as Essie Esterhuizen pointed out, “H atrofusca” as a variant of “H. magnifica”, seemed to be more dominant than had been realized. There were several other complications largely due to ignorance. Since my revision I have done so much more exploration and turned up so much new material that I have been forced to the conclusion that there is really one main element involved and that is H. mirabilis. This is where I believe the Komserante plants belong and the difference from the Nature Reserve population is due to a degree of infusion of H. retusa.
I revisited the site with Kobus Venter many years ago but did not look at a reported second population higher up the hill, taking it to be a little different based on plants I saw in Kobus’ collection. What was on my mind while we were recently exploring the area further east to examine the possible connection of H. mirabilis “magnifica” to “splendens” (and which we confirmed), was the fact I had never seen Kobus’ plants from Kruis Rivier northeast of Riversdale other than in Kobus’ collection. The plants I saw were also generally more robust than “H. magnifica” and more evenly tubercled. Kobus kindly took me to that Kruis River locality and much to my surprise the plants were in flower late October (see JDV92/65 Figs1). This is quite wrong for H. mirabilis, which is essentially a summer flowering species. I later went again to explore Komserante more thoroughly and to look at both the “magnifica” populations to which I believe the name “H. enigma” has been applied. The populations are in fact no more than 75m apart and cannot be considered to be genetically discrete at all (see MBB7778 Figs 2, and MBB7779 Figs 3). While it is true that the habitats are slightly different, this is reflected in the plants that at the upper slope of the hillside are vegetatively more robust and even clump forming, while those lower down in a bushier grassier habitat tend to be solitary and more withdrawn into the soil. These plants flower in summer and it is evident to me that there must have been some genetic exchange with H. retusa that grows approximately 200m away on the same hillside.
1. JDV92/65 H. mirabilis Kruisrivier.
2. MBB7778 H. mirabilis. Komserante.
3. MBB7779 H. mirabilis. Komserante.
Habitat preferences are very strict and it is curious to observe that there are four small buttresses forming the western slope of the low hillside. On the northern one we have the two H. mirabilis populations. On the second is H. retusa ’geraldii’ (see MBB7780 Figs 4) that is very clump forming, the third has no haworthias and on the fourth is a less clump forming H. retusa that is often referred to as ‘fouchei’ (see MBB7781 Figs 5) because the leaves are slightly longer and more upright than in ’geraldii’. This is not quite correct. Mr. Dekenah showed me what he took to be the true H. retusa ‘fouchei’, further south off the hillside and along the stream at Komserante. This habitat has since been cleared for farmland. The plants were very large and the leaves considerably more erect than is the case for the plants on the hillside. Curiously H. minima is present on the first and last buttresses. Flowering time for these H. retusa is spring but we did see vicarious flowering in summer so that definitely it is possible and probably a regular occurrence for out-of-season flowering to occur with the possibility of hybridization. I noted such hybrids between H. retusa and H. mirabilis in our exploration further east from Riversdale and I did think some of the clones in the Komserante plants evidenced this as well.
4. MBB7780 H. retusa. Komserante.
5. MBB7781 H. retusa cf. ‘foucherii’. Komserante.
The reality is that the Kruisriver and Komserante “magnifica” may flower at different times, but they are vegetatively very similar indeed. I consider that they belong in the same system as H. mirabilis and this conservative view of “species” as systems is the most sensible way to interpret and understand the plants. The approach can be taken further, as I tried to do in an earlier article concerning H. mutica ‘nigra’. In that article I dealt with a whole series of populations in and around Heidelberg, to show that there are three main elements viz. H. retusa (including H. turgida), H. mirabilis (including H. magnifica) and H. floribunda. I did not deal specifically with, nor resolve, the issue of H. mutica var nigra. I also noted populations along the lower Duiwenhoks river and suggested that H. floribunda is absorbed into H. mirabilis virtually south of the N2 east west highway, re-emerging on the northwestern corner of the Potberg in the south.
My most recent exploration was to check the flowering times of the original population of H. mutica var nigra at its origin at Kransriviermond and what I assumed to be the same thing on rather limited examination further north at Morning Star. Both these populations are along the Duiwenhoks river southeast of Heidelberg and in a continuum of four populations of H. retusa extending from just east of Heidelberg to still further east at an eastern Diepkloof locality (I think it has been referred to as Droekloof and Droerivier too, and the name ‘chromatica’ attached to the plants – see MBB7794 Figs 6). Less than 10km away is a population of H. retusa at Pienaarsriver (see MBB7776 figs in Chapter 3) that is very near to the whole array of populations of Kiewietskraal that I discussed in my writings about H. mutica var nigra. Two of those were distinctly H. retusa and so is the Pienaarsriver population that is different in that the plants are mostly quite dark in colour.
6. MBB7794 h. retusa ‘chromatica’. SE Heidelberg.
In those same writings I mentioned several populations of H. mirabilis along the Duiwenhoks river as well as populations of H. retusa var turgida. Here I am unashamedly relating “turgida” to “retusa” as I believe that when one calls all the populations now known to mind, as well as the variants they contain, this is the correct position of the plants in relation to a sensible and rational species concept. It is evident that H. floribunda is merged in H. mirabilis south of Heidelberg and the same thing occurs south of Swellendam. Thus I see no problem in now dealing with the population at Kransriviermond to which I applied the name H. mutica var nigra. I attached the varietal name to H. mutica because this is what G.G.Smith had done, and I believed that it was the link between H. retusa in the east and H. mutica in the west, when there were still huge gaps in the known distribution records for both species.
I have mentioned a population of ostensibly H. mutica var nigra at Morning Star, which is about 5km north of the Kransriviermond origin of the plants so named. This Morning Star population (see MBB7221 Figs 7) is very significant because while it includes virtually the same range of variants as the Kransriviermond plants, it flowers in summer as opposed to spring for Kransriviermond. Not only that, but it is 300meters away from a population of H. mirabilis (previously this would have been H. magnifica) that flowers and seeds at the same time. The habitat is marine plane ferricrete that overlies Bokkeveld shale but there are subtle differences between the two habitats.
7. MBB7221 H. retusa ‘nigra’. Morning Star.
The Kransriviermond plants (see MBB7804 Figs 8) are down in the Duiwenhoks valley in exposed Bokkeveld shale and I consider that the dark colour of the plants in the two populations is due to infusion of H. mirabilis. The rounded leaf tip would be derived from the “atrofuscoid” element of H. mirabilis while some plants also have the tuberculate roughness that that element often has.
8. MBB7804 H. retusa ‘nigra’. Kransriviermond.
Further south and west is the Slangrivier valley where there is also a suite of populations that includes smaller forms of H. mirabilis (H. heidelbergensis var. toonensis) as well a H. retusa var turgida also in an unusual form that supports the taxonomic position I now adopt.
The conclusion is that the Morning Star and Kransriviermond ‘nigra’ can be regarded as H. retusa within the same broader concept that includes ‘turgida’. The problem now arises from still another new discovery. This is from along the Buffeljags River (see MBB7801 Figs 9) about 10km southeast of Swellendam and 12km from the nearest obvious H. retusa at Goedverwagting further east; and from the less obvious H. retusa (that I have elsewhere noted as H. mirabilis) south of the Tradouw Pass. The nearest H. mutica are within about 15km to the west at Napky and Luiperdskop. The plants are the now customary “variable”. The leaf tips are very rounded. Some plain, some lined, some with curious submersed pinkish dots. The surfaces can be quite smoothish or else the surface cells terminate in minute spines that give the surfaces a glistening appearance – evident in H. mutica too. These plants flower in summer whereas both H. mutica and H. retusa are spring flowering. Note needs to be taken of the habitat. It is an extensive and old river gravel embankment and terrace, and therefore another variant on the interface of recent ferricrete deposits and the older Bokkeveld shales. The vegetation is grassy Karooid bushveld, unlike the dry fynbos to the northwest or the renosterveld (mainly cultivated) to the east.
Kobus Venter saw the photographs and suggested that the plants might be H. mirabilis’ and reminded him of ‘atrofusca’, perhaps also by virtue of the flowering time. This is of course what my “enigma” is all about. It is compounded in this case because these Buffeljags ‘muticas’ are rather odd in that the leaf ends tend to be flattened almost as though H. floribunda has also entered the fray. Kobus was much happier with the H. mutica connection when he saw the plants in the field.
The story here closes with H. mutica MBB7741 from Dankbaar about 12km west of the Breede River bridge at Swellendam. The plants here also have very glistening surfaces and thus seem to confirm the very original perception of G.G. Smith’s that the Duiwenhoks plants and the plants west of the Breede River were connected. It is certain that he never envisaged the complexity of the interrelationships between three main elements viz. H. mirabilis, H. floribunda and H. retusa.
Footnote. It is just assumed that perhaps the name “H. enigma” has been used for the Komserante H. mirabilis, because that population is indeed enigmatic as this article explains. Gerhard Marx informs that “enigma” applies to a H. mirabilis population further west where the more atrofusca variants occur. Since writing this chapter yet another similar population to the Kruis River and Komserante populations has been found about 15km SE Riversdale. ♦
Among a plethora of new names, are two really interesting items.
Firstly there is a really useful formalization of a host of old names by Gordon Rowley. This is something that I understood to be taken as unwritten truth and logic. While so sensible and practical it almost widens the gulf between the ordinary way we use names to communicate, and the unreal world of formal taxonomy. I will just give the one example to demonstrate this viz. H. coarctata subsp. coarctata, var. grandicula ‘Baccata’, or H. coarctata subsp. coarctata ‘Grandicula’ ‘Baccata’. This is a probable botanical truth where in the end there is a population (yet to be formally identified) in which all the plants despite any variability are ‘grandicula’ and something in cultivation (and not wholly improbably in some similar incidences, two quite different clones) that on the available evidence seems to have been drawn from the same population, which is ‘baccata’.
The second item is the name H. schoemanii. Described by M. Hayashi in Haworthia Study (9:14, 2003), it was said to have one of two close allies viz. H. woolleyii and an H. crausii. This latter ‘species’ I did not know, or of. Gerhard Marx kindly sent me the picture that appears in Haworthia Study (4:8, 2000) and I really do not know why this is compared with H. woolleyii. I would say that it also collapses in the same way and into the same place as ‘Schoemanii’. Most of the other new names I have been able to assign with very little stress into my own understanding of the system of plants that falls in Haworthia. However, the illustration of this plant and the comparison with H. woolleyii, again stretches the limits of credibility. It was said to have been collected on a hill near Dwyka Station. I must confess that the report that Hayashi found this species by himself is one I find very hard to accredit, being familiar with his modus operandi and field familiarity. When I was at Dwyka station myself I was under the impression that the plants had been collected by Paul Schoeman for whom it is named. Had I known the facts I would have been even more skeptical that it had in fact been collected there. It is said to have thinner and shorter leaves than H. woolleyii and the stem not to elongate as in H. granulata. I consider this to be fairly threadbare stuff in as much as the number of leaves is not brought into the equation and neither is the variability of H. granulata. At Skitterykloof, Avondrust and Karooport, the plants do not form stems as they do in Verlatenkloof or the Koedoesberg. At Patatsriver the plants are stemless too and the leaves are fairly thin and erect (This may be the source of ‘Crausii’). A Harry Hall collection from Skitterykloof was of stemless plants with very long and thin leaves. I would not say that H. woolleyii makes dense clumps by rhizomes as Hayashi states. But it does have many long thin leaves broadening of course at the base. It does cluster, but by offsetting aboveground and very close to the crown of the plant as does H. attenuata for one. This can barely be stated to be rhizomatous, which implies an elongated self-rooting horizontal stem with food storage. The term stoloniferous is applicable to the underground trailing stems or even root-like structures that eventually manifest as new rosettes and that occur in several Haworthia species. Plants with elongated stems can also be found in H. venosa subsp. tessellata and stoloniferous forms are also common.
Anyone else who really and genuinely wants to understand plants in the context of relationships and a realistic classification is now faced with the same problem as myself and one of some magnitude. That magnitude is of course only measurable in terms of individual knowledge and experience of the South African vegetation and geographical reality.
Where now is Dwyka Station? It is between Beaufort West and Laingsburg and centre to an area that barely raises a blip in the context of the species rich South African flora. What happens there geologically, geographically, historically, botanically? I am not going to attempt to answer the question in any detail except to comment that there is absolutely nothing in my own database to suggest that there could be any new species of Haworthia there – of any kind, – let alone one that looks so like ‘woolleyii’ by author statement and by my own imaging of the illustration.
Firstly one has to consider that ‘woolleyii’ occurs as a single known population east of Springbokvlakte far to the east and south. It has other species associations there. Secondly it is vested in the classification of H. venosa for the very reason that it has an uncomfortable resemblance to the subsp. ‘tessellata’ (the fact that this classification of subspecies is in doubt is irrelevant and is only as valuable as the ‘looks like…’ observation).
Now I did not find anything in my very brief visit to Dwyka Station itself except to confirm my view that there is absolutely nothing there to suggest an environment in which something as fundamental as a different Haworthia species could ever have evolved or survived in terms of vegetation and historical changes. Hayashi writes that he found five clones on “the mountain slope near Dwyka station”. The slope at Dwyka Station barely passes mention as that of any mountain and is mostly contained in rail reserve that is severely impacted on. I should add that we could only identify Dwyka station by virtue of the abandoned ganger’s cottages and there is a possibility that Hayashi was referring to the power substation about a kilometer away where we also looked. There may be a few plants there at either place but it was extremely dry and the absence of any other vegetation of any interest at all discouraged anything else but a fairly brief but intensive search by the three of us. In my experience, Haworthias never occur in such depauperate environments. So we went a little further west to near Koup Station, rather than spend time trying to access ownerships and nearby hillsides that could have been more fruitful than immediately Dwyka Station. Koup is of course the location for H. arachnoidea var. scabrispina, fig. 6, and also for H. nortieri (Oh dear!) var. pehlemanniae ‘Albispina’ (?) or should I have written H. nortieri var. globosiflora ‘Pehlemanniae’ ‘Albispina’. Remember that H. nortieri ‘deVriesii’ (in the bounds of a more realistic language) occurs at Prince Albert to the southeast.
I was accompanied by my wife Daphne, whom I mention extremely reluctantly in the crucible of a fairly dubious lot of writers, literature and connotation. She has a mind and eye uncluttered by taxonomic perturbations and has developed a vegetation penetrating and interference defying Haworthia locating radar. So at Oskopvlakte west of Koup she found plants that must surely explain H. schoemanii. What is as unpeculiar about Oskopvlakte and Dwyka Station is that they are on the boundary of the winter rainfall biome and the edge of the Succulent Karoo domain within that. (I deliberately correct the formal botanical concept of the biome status of the Succulent Karoo where history and causation seem to be confused). While Daphne was finding haworthias I was looking at Conophytums, Pelargoniums (the obligate winter rainfall growers) figs. 6-7, as well as other species familiar to me such as Astroloba bullulata, and thinking “Yes, this is Tanqua Karoo stuff”. Then Daphne called me. What is the plant she found? Wholly reminiscent of H. venosa subsp. granulata, figs 1-5, that is known to me from only about 12 widely scattered populations on the periphery (refugial and mountaineous) of the Tanqua Karoo. The nearest populations to Koup (Oskopvlakte) east of Laingsburg are at Avondrust southeast of Touws River, and Patatsriver WNW of Matjesfontein. Distance can be said to be a factor, but not in view of the overall distribution of H. venosa and its allies in any taxonomic dispensation. To my jaundiced eye ‘Crausii’ could equally easily be explained by these Oskopvlakte plants.
So for the collector I would suggest the label as simply H. ‘Schoemanii’, without the ‘H’ and even without the capital ‘S’, and most importantly of all, without any derogation of the late Paul Schoeman who was well known to me and against whom I have no ill-will of any kind. My only complaint is that his name has been dragged into the realm of botanical reality in the way that it has.
I actually have no interest whatsoever in taxonomy and nomenclature apart from their evil necessity. I just felt it was incumbent on me to make known some of the obvious problems of identification and use of names in the sphere of plants that I like. So this foray out of retirement is simply necessitated by an original motivation that can never go away any more than that the name ‘schoemanii’ will, and this is reasonably so in terms of human interest.
Acknowledgement Dr. Gerrit Visser, who made this excursion possible, Daphne whose company I value, Mr. Adriaan Botes of Oskopvlakte, who so graciously and kindly gave us access to his farm. Gerhard Marx kindly sent me the copied description of ‘Schoemanii’ and ‘Crausii’ (The latter wholly in Japanese). He also kindly commented and offered some corrections to the manuscript. ♦
I wrote “What is Haworthia schoemanii? in reference to the statements I have made that I did not want to write any more. But also kind of intimating that really we need to find some sense in all this “namenklutter”, while at the same time I am drifting away from the community that controls it and isolated from that which generates most of it. My note was about the names ‘schoemanii’ and ‘crausii’; and their perceived improbable existence as biological systems. This is then ust a very short note to elaborate on a record of mine of H. venosa subsp. granulata that I gave as from Patatsriver Road. This was partly because I did not want to advertise too openly where the site was and because I was not even sure of the farm name. Nowadays, while I fully appreciate there are some not very nice people who do and will abuse the situation; I do not believe there is any merit whatsoever in secrecy. It seems to me that this simply exacerbates the whole situation just as does the insistence that conservation laws that so effectively exclude people from participation, contribute to conservation.
When I transferred to the Department of Agriculture in 1987, I became involved in vegetation regeneration. This had been initiated in the Western Cape as early as 1936. Without a single indication of success I was tasked with the job of continuing the great work. The department had used a site at Bantamsfontein in the southeastern Tanqua Karoo that is accessed via the Patatsriver Road that links Ceres by a country road directly with Matjesfontein that is on the main highway Cape Town to Laingsburg and on to the north. The site can best be described as a rather homogenous stand of Ruschia spinosa on a low-lying and flat area with very little surface rock. It had been used for a reseeding experiment. The only present evidence, after nearly 30years is a few scattered plants of Atriplex nummularia outside the treated plots. Our visit in 1988 was to attempt sowing of Osteospermumj sinuatum into the Ruschia as nurse plants using a rather inventive handseeder. I am sure it could only have been this close attention to those nurse plants that drew my attention to the very cryptic plants of ‘granulata’. (I must mention here the abhorrence that has been expressed at this violation of some sort of ethic that requires the use of virtual the full Latin set of epithets for every mention. I strongly suspect that this is only to vindicate a wholly false belief in a reality that the names seldom have).
The plants I saw were seldom stemmed and what I grew and propagated were smallish stemless plants generated by rhizome (underground shoot). The habitat is/was quite unlike the high rocky slopes that I would have expected to find these plants in and I often wondered about this issue. Unfortunately my activities in vegetation ecology, while allowing me so much time in the field, required me to be where the animal action was and focused on general vegetation rather than the sort of exploration and observation I would have liked to do in terms of my own interest. The consequence is that I learned very little about Haworthia in those years.
After my recent experience with ‘schoemanii’ and Steven Hammer’s expressed interest in Conophytum distribution, I took the opportunity of going back to Bantamsfontein. The farm is owned by Jan du Toit, a leading light in the fruit farming industry of the Koue Bokkeveld in the Ceres District. The farmers there traditionally also owned farms in the Tanqua/Ceres Karoo, and stock would be driven, or later transported, there for grazing during the winter months. Production has declined dramatically over the last 60 years and many farms are now owned by people who have “invested in a seaside cottage in the Karoo”. Mr. Du Toit does still continue there with sheep though his business is 98% fruit. The vegetation is in excellent condition.
Daphne and I started searching in the old experimental site and quickly found plants, few and scarce as they were. Our usual response in a situation like this is to ask “Now where are they really?” In other words, to try and establish where the preferred habitat actually was. We crossed to the west side of the road and walked a long way without finding any more plants. We only saw a few H. arachnoidea ‘tanqua’. Then we drove a short distance northwards to a rocky ridge that looked more probable, and as surely there were the plants in abundance. Mr. Du Toit was our host for the night and the next day we searched south of the farm house, finding ‘granulata’ and ‘arachnoidea’ over a wide area.
The pictures will illustrate the variation. The plants may be stemless or stemmed and propagate by short offsets or by longish rhizome. The leaves may be quite slender and acuminate or short and squat; unmarked on the inner upper surface or plain.
I have tried to understand the geology of the area and get some idea of the significance of parent materials in relation to soils and the distribution of plant species. I am quite convinced that this is really important, but I just cannot formulate why this is. Just as the SW Cape area from George to Caledon can be seen to vary from east to west, so is it that the Karoo is divided. In the latter case it is more easily explained by the fact that the parent sediments of the Karoo Supergroup come from essentially two different sources. The smaller southwestern area was fed in the times of Gondwanaland by fairly large particle
sediments from what is now South America (the Parana basin), while the larger northwest was fed by finer turbidites from northern and western (the Karoo Basin). Dwyka Tillite of earlier glacial origin is exposed on the western and southern boundaries, Dolerites of later origin dominate the north while the eastern boundary of the Tanqua karoo is contemporary deposition. In skeletal soils it seems obvious that source material strongly dictates mineralogy, derived soil texture and consequently vegetation. Overriding this geology is topographical feature and climate pattern.
I find it very odd, considering the short span of human life, that those events from as long ago as 545m years may be so relevant to the present and to plant species whose history seems to be so recent. It is also very odd, for me at least, to understand why so many of the Tanqua Karoo endemics are in the oldest formation of the Ecca Geological Group i.e. Tierberg Formation, and not in the overlying three.
Acknowledgement Mr. Jan du Toit for gracious hospitality and kindness. Dr. De Ville Wickens for information on the geology of the Tanqua Karoo. ♦
This chapter is based on recent field exploration and embroiders around many aspects of Haworthia species discussed in earlier chapters. What should be striking is that new populations follow the very predictable geographic pattern that all my earlier exploration has exposed and in my estimation confirm in every way what I consider a sound and satisfactory taxonomic solution and help explain its limitations.
1. Haworthia minima.
Two new populations of this widespread species are recorded:
MBB7807 Swartrug, SE Heidelberg. This is a steepish east facing slope. The plants are small, the leaf length barely exceeding 50mm and with a tendency to form clusters in the shallow clay banks that rift the area. There are many plants in quite a small area and it curious to see them within very large clumps of Euphorbia tridentata. This Euphorbia itself has a strange geographic distribution being known at Riversdale, Calitzdorp and then at Cradock in the Eastern Cape. (see Figs 1)
MBB7813 NW Skeiding, E Tradouw Pass. This is a high rainfall area and we were expecting to find H. mirabilis in the lower pressure bursts and white clay that can be seen from the farmhouse far away. But instead we came across H. minima at the highest point of the hill among sparse grass. They are rather green in colour and it is interesting to note in the one plant pictured that it has non-tuberculate upper leaf surfaces and white cartilaginous margins and keel of the leaves. (see Figs 2)
2. Haworthia rossouwii.
MBB7803b Morning Star, S Heidelberg. Morning Star was known to J. Dekenah but the area has been very developed since those years. It was very surprising to find this small form in abundance at the top of a high hill in ferricrete pebbles and virtually no vegetation ground cover. However on this subsequent visit we crossed through an internal farm fence and found the plants in abundance about 150m further away and over quite a large area. We later looked at a lower promising hillside and small area nearer the Morning Star homestead and found the plants again. (see Figs 3)
Looking from Morning Star to Kransrivermond 097
3. Haworthia mirabilis.
This item reports on 6 populations, three of which are new. They confirm the continuity down the Duiwenhoks River and also southwards and westwards from east of Riversdale through to west of Robertson and Greyton and Napier to the south of that. Attention is drawn to south of Heidelberg where the ‘enigma’tic relationship of this species with H. retusa is suspected.
MBB7808 Swartrug, SE Heidelberg. This farm is well known as the source of ‘Haworthia chromatica’ that I dismissed as an element in the complex surrounding the e H. retusa (mutica) ‘nigra’. The names Droekloof and Doringkloof have been used while the map reference gives Diepkloof for a large area extending to west of the Duiwenhoks River. The plants are on an east facing boulder terrace in a stony low bush site with some grass. The plants are small and show the expected wide range of variation. They are fairly similar to the plants in the lichen patch just east of Heidelberg that I have always mistakenly taken to be the origins of H. heidelbergensis, but here are subtle differences that mark a population difference, but I will come back to this. (see Figs 4)
Looking from Swartrug towards Riversdale.
MBB7809 Koeisekop, SE Heidelberg. This is on the same farm and a similar ferricrete hillside southwards. There are several hilltops by the same name in the general area and I cannot even speculate on the origins of the name (“cowshead”). The plants are larger than in the previous population and very abundant. The similarity to populations in the Swellendam and Riviersonderend area is in my opinion striking. Odd clones are miniature images of H.mirabilis from the southwestern populations west of Napier. (see Figs 5)
MBB7220 Morning Star, S Heidelberg. I have discussed this population before and I repeat it here because I did not have many field photographs. It is only 300m from the Morning Star MBB7221 H. retusa ‘nigra’’ population and they flower and seed virtually in unison. Therefore they probably also reflect on the strange interaction between the two major systems. What I have surmised is that H. floribunda gets absorbed into H. mirabilis at the southern limits of its distribution and in these pictures there is clear evidence of the rounded and flattened leaf tips of that species. Therefore we may actually here have a three way species interaction. Reinforced by the presence of three different things at Kransriviermond, covered later under 4. H.retusa ‘turgida’. (see Figs 6a and b)
MBB7221 Morning Star
MBB7811Kransriviermond, S Heidelberg. Populations further down the Duiwenhoks river have been reported on and this one fall within that mould and the same one as the previous Swartrug and Koeisekop populations. The population is smaller and the plants too are quite small. (see Figs 7)
MBB7814 Skeiding, W Heidelberg. This area is also reported on but digital photography makes it so much easier to record the enormous variability that such populations exhibit. Some of the plants are typically ‘atrofusca’ while others have leaves ended in an awn. Leaf surface may be smooth and there is even an indication of smooth leaves. What is notable is the absence of H. floribunda from what is the sort of habitat it can be associated with and can one can speculate its absorption into H. mirabilis with leaf shapes that demonstrate this. H. floribunda does appear again about 15km to the southwest in association with H. retusa and in the absence of H. mirabilis. (see Figs 8)
The Skeiding homestead
MBB7233 Die Plotte, Heidelberg. “H. heidelbergensis” correctly has its origin in this near vicinity and these small summer flowering elements are certainly part of a large assemblage of populations in the central area of what I now come to see as all H. mirabilis. Here discussion must pass to the following. (see Figs 9)
4. Haworthia retusa‘turgida’.
A difficulty that I experience is in the need to dismiss the impression and general idea we have that there are tiered ranks. H. retusa has many populations and past treatment has placed most of the variants in H. turgida. To suggest that these are really two separate systems is not realistic and confounds any explanation for a few anomalies that exist.
MBB7810 Tierkloof, SE Heidelberg. Tierkloof is quite a deep valley that pushes out of the Duiwenhoks to the east. We found this huge population of large clump forming plants and several such populations are known just north of Heidelberg itself. I have applied the epithet ‘longebracteata’ to these and this is no more satisfactory than generating names for each of a very wide range of populations. There are populations along the Duiwenhoks that are far more inclined to the ‘turgida’ element and Essie Esterhuizen wrote a very good account of the host of variable populations that he encountered. (see Figs 10)
H. retusa ‘turgida’ at Tierkloof, Swartrug
MBB7812 Kransriviermond, S Heidelberg. We only saw three plants at this locality and these plant mirror the preceding, if a bit smaller. The problem here is that we also have the element I myself described and named as H. retusa ‘mutica’ and I originally linked this “variety” with H.. mutica following what G.G. Smith had done and for predictive reasons. The fact that ‘turgida’ now occurs with it is a huge hurdle to cross. Think carefully. (see Figs 11)
MBB7804 Kransriviermond.I have written about his population in the chapter “Haworthia enigma”. This explains that the plants are not all leaden-black as perceived. They are spring flowering – as ‘turgida’ is. However, the range of forms and colour virtually replicates the plants in MBB7721 at Morning Star (pictures in the “enigma” chapter) where they are spring flowering. Across the river from both these populations is a weak population of small plants of H. mirabilis (7811 above) summer flowering. We know that this major difference in flowering season is not a total barrier to hybridization. (see Figs 12)
MBB7234 Die Plotte, Heidelberg. This is a population of large clump forming plants on a north slope about 300m south of MBB7233 H. mirabilis ‘heidelbergensis’. It is spring flowering. When I first saw these plants I dismissed (actually I never dismiss anything – I maintain reservations as I do now) these plants as large variants of ‘heidelbergensis’, and actually collected one magnificent specimen that looked to me just like H. mirabilis, say, from west Napier. Looking at the range of variation one is compelled to concede that here again is an infusion of H. mirabilis into a population that is primarily H. retusa in that vast shadowland it casts with ‘turgida’. (see Figs 13)
Conservation I add these comments for several reasons surrounding the poor image that collectors have in the eyes of conservationists, and the very rickety fence I therefore occupy. There is a note reprinted in a recent issue of the newsletter To the Point. This taken from Times magazine and seems to be a rather hyped up condemnation of widespread destruction based on a view of internet websites. These are also monitored by RSA law enforcement that is rightfully disturbed by the high number of locality and collector citations that are indicative of illegal collecting. I personally no longer apply for a “collecting” permit for the prime reason that I am concentrating on photographic record, as a herbarium record no longer serves any better practical short term purpose. The herbaria are overburdened and I prefer living plants to dead ones. After 70 years of field activity I feel treated like a pariah and reduced to subservience under less-informed inexperienced people operating under a rigid and oppressive legislation aimed at acquiring a prosecution rather than wisely trying to direct activity in a positive direction. This activity of collection is, and I have no doubt about this either, a human right imbedded in the very reason for creation and our place in it. Conservation has its roots in the passion that people have for nature and it is nurtured by experience, contact and knowledge of its varied components. It is just ridiculous that one can go to any animal or plant centre here in South Africa and see plants and animals from anywhere else in the world and none indigenous. It is Ok to, say, keep Madagascan Chameleons in captivity where it is someone else’ responsibility to secure them. But keeping local ones is taboo? It is OK to freely sell cacti that do not occur here but are now being dug out in tons in (if that is really true). Recently it has become illegal to so much as possess Encephalartos latifrons and even the National Botanic gardens can no longer propagate and sell this plant. While we talk “plant indigenous” on all public fronts, the fact is that the acquisition, cultivation and sale of indigenous plants is viewed in the most negative light – a viewpoint encouraged by conservators.
This is control by law, a large degree of pure ignorance and by improper application of power derived from legislation. This is not the way to go. It is quite evident from the Times magazine article that legislation has the negative effect of creating a false value and glamour to discovery, Latin names, acquisition and possession. I believe that knowledge and understanding is true power, not legislation and enforcement.
The collecting numbers, localities and information provided here is to promote understanding and knowledge of plants, and thereby to promote their continued existence against all the vagaries of nature that “threaten” them, least of it which may be actually human greed.
Acknowledgement I am glad to acknowledge the company, comments and interest of Gerhard Marx and whose organization sceptisism I also value. Landowners always impress me with their generosity and graciousness in allowing us access. On this occasion they include Mr. and Mrs. Nelie and Wimpie Jacobs of Diepkloof, Mr. Cloete and Belinda Fourie of Melkboom (B&B), Mr. and Mrs. Coetzee and Sarita Uys of Morning Star, Messrs. Christo and Pieter van Deventer of Doornvlei, Mr. and Mrs. Neels and Anne-Lise Uys of Skeiding (B&B) and then Ms. Jennifer Steggie of Heidelberg. In view of the comments on conservation noted above, I note that there are several farms that now offer accommodation and an outdoors experience and the opportunity to view these plants in their extraordinary natural surroundings. ♦
Alsterworthia produced a special edition (No.7) in 2004 to publish new species and combinations subsequent to the publication of Haworthia Revisited. I was given a copy because of my own contributions in respect of primarily new combinations. I had the previous year done some exploration along the Duiwenhoks River south of Heidelberg and found several Haworthia populations notably MBB7227 Witheuwel and MBB7229 Somona. I discussed these in Chapter 6 of my Update Vol 2. dealing with the complexity of the element H. retusa (mutica) var. nigra and the problematic nature of H. mirabilis as it occurs around and south of Heidelberg. So when I saw the picture of H. jakubii I merely glanced at the description to see the words Duiwenhoks River to think this was another of those weird armchair products to befuddle the enthusiast and add another name from an endless production belt. There was nothing about the illustration that suggested anything new to me so it is really fascinating to now only read what the author had to say “When the author first saw them, he thought they were something new because of their features.” This is a very subjective statement and I have no doubt that the author could be misled into thinking that other plants from the same population could also be “something new”. Why “something new” should be allied to a Latin binomial is intrinsic to “namenklutter” and the disrepute into which taxonomy has fallen.
I must dwell on that statement a little because it helps to demonstrate, falling very far short of explaining, why plant classification becomes such a hopeless mess. In 1986 I wrote an article published in Excelsa 12:91 entitled “Confusion reigns again” where I stated that it had become seemingly impossible to write about Haworthia without casting doubts on one’s own, or someone else’s, integrity or sanity. This was because of the apparent lack in the literature of any sense of discrimination and recognition of what might be right or wrong about any written piece. The more painful aspect of course is that it meant that I could write nothing myself that would be believable. I have to state that since 1986, the situation has not improved one iota and I regret that I am not tactless, aggressive or unkind enough to list a set of prominent people, including botanists, who have done nothing more than fuel the fire of enthusiastic writers. So we have “because of their features”. What features? I presume these same unlisted features are what made me sigh and think “Oh dear! – another name for H. mirabilis”.
I did not give the element anymore thought until I happened to see Jakub Jilemicky’s website and e-mailed him to query the principles he was adopting for recognition of “species”. From him I learned that the origin of “jakubii” was in fact the Goukou (ex Kafferkuils) River north of Stilbaai and Jakub kindly also sent me map details. Because there are virtually no records of H. mirabilis further south east of Riversdale than in the Frehse reserve close to town, I suspected that perhaps we had to do with a H. retusa ‘turgida’ variant or possibly a hybrid between that and H. variegata. While not generally prone to concede oddities to hybridization, I have become much more circumspect in this respect after learning a great deal more about hybridization between species despite big discrepancies in flowering time. Jakub’s map was a bit problematic because the designated spot was quite far from the Goukou River and in the middle of cultivated land. Furthermore the farm name Melklhoutkraal is cited and the boundary lay east of the designated spot. However, my wife and I came to the designated place where a road serves two farms south of the road. The first farm deserves special mention in respect of the negative aspects of agriculture where an over zealous new owner has decimated the landscape with no consideration for conservation whatsoever. The invasive Acacia cyclops (Rooikrans) has been bulldozed away and what may have been cultivated before certainly is now, and this also extends into that fragile zone between the productive shales and the non-productive calcrete. The owner initially offered, being busy at the time, to show us a hill that was being “conserved” and we made an appointment to do so. However, when we later arrived back we were informed that here had been a change of mind and we would not be allowed there. The reason will become apparent.
We went on to the second farm where the owners were delightful and allowed us to explore at will. The farm borders the river, but we stayed away from that as we were fairly sure that H. retusa ‘turgida’ would be all there was to see on the steep banks. So we explored the calcrete that is so reminiscent of H. paradoxa and Vermaaklikheid. We found a small and very cryptic population of H. variegata that rather strengthened our suspicion that perhaps ‘jakubii’ was of hybrid origin. Then we went further north to continue some exploration we had started there and we found a strong population of H. mirabilis about 15km SE Riversdale (MBB7818 Windsor – figs 8-10). We connected this population to plants at KomseRante east of Riversdale and also Kruisriver further east and north and suggested the possibility that ‘jakubii’ was then a Duiwenhoks River image of H. mirabilis.
Back home and back onto the internet. Jakub sent a better description of the locality much nearer to the river and on the farm Klipfontein rather than Melkhoutkraal. An error and something I might have worked out had I dug out the publication and recognized that the stated geographical facts were as unreliable as the taxonomic claims. Kobus Venter was thankfully able and anxious to get into the field in connection with other finds, and he kindly offered to drive all the way to Stilbaai to follow newest information. Gerhard Marx was also with us as a most welcome guest, and we then found the evasive ‘jakubii’ growing in a very narrow band right alongside the road and extremely vulnerable to any road works (see MBB7820). Our over zealous farmer had not only erected a new fence and deposited all the Acacia branches on the road reserve, but had also concentrated stock on the river bank on the farm side of the fence. The result was what is to be expected when there is no sympathy for the environment or for the stock. Animals under pressure will eat anything and everything and the concept of selective grazing and suitable stocking rate can fall away. One may as well cultivate the soil for the devastation too many animals in too small an area for too long causes. It is fortunate then that we have the road reserve. We found about 40 plants that appeared from the dry capsules and stalks to have flowered about 6 weeks previously in early February in close synchrony with H. variegata nearby and also with the Windsor population of H. mirabilis. We took Kobus and Gerhard to see the Windsor plants and they confirmed the similarity of individuals there to ‘jakubii’. Among Figures 8 are plants that demonstrate this similarities but also the wide range of variation so evident in most populations of H. mirabilis, thus H. mirabilis ‘jakubii’- a significant population and well worthy of a name outside of formal botany, which brings me back to my near opening remarks. The formal nomenclatural system works very well as no doubt does the horticultural code. However, there are serious limitations that severely affect Haworthia. The first is that few people actually have much experience with plants in the field and the true nature of natural variation. There are too few field-familiar competent taxonomists and the whole field of taxonomy is seriously flawed by any form of peer review outside of the nomenclatural rules and procedures. Another serious flaw is simply the absence of any general and common understanding of what a Latin binomial is actually supposed to be appended to. The same is true of lesser taxonomic ranks. Gordon Rowley many years asked (of me) “I would love to know what is to become of the luckless hybrids and cultivars if they are to be deprived of the rank of genus and species.” I am now not sure if he thought through that question very well but I understand what he means and in recent literature he has highlighted the value of names for the sphere of horticulture in which “collectors’ operate. Short of microchipping, many cultivated plants are going to be deprived of names simply through loss of physical labels. I am not sure, and underlined by that very curious statement about the features that made ‘jakubii’ into something new, that any formal system of cultivar recognition is going to work in Haworthia. The problem is that we are not dealing with flower characteristics and colours of virtually a single species as happens in most horticultural groups. Nor are we dealing with any controlled breeding program. We are dealing with vegetative characters of perhaps 30 species where there is much evidence of gross misidentifications by experienced professionals (botanists) at species level. Also there are any numbers of potential new cultivars by virtue of the simple facts of natural variation. How are we going to ever arrive at descriptive matter and even illustrations that will allow correct identification when growing conditions and season influence the vegetative characters of the plants concerned and we cannot even be sure to what species they belong.
The author of ‘jakubii’ does end by saying that the taxon is intermediate between H. mirabilis ‘paradoxa’ and H. mirabilis ‘magnifica’, listing shape, surface, coloration and solitary growth as reasons. This may seem reasonable on the surface of things. But it is totally unreasonable if one considers that one could take representatives from the same and different populations of these and use the same argument to justify many more names within those two elements as variable components of a much greater cohort of variants. The description itself simply cannot be taken literally. A plant is illustrated with leaves having from 3-9 face lines however one chooses to count them, while the description states 5-7. The colour is given as grass-green (in poor light), but we saw most plants in semi-shaded and poor light microhabitats and none were “grass-green”. The leaf surfaces are also described as “smooth” and this was not evident in the plants we saw. The converse is true and also in respect of either ‘paradoxa’ or ‘magnifica’. In fact the statement underscores the (poetic) license that is taken with descriptions that end up unworkable. In H. mirabilis, the leaves may indeed be smooth (e.g. ‘badia’) but in any population there are plants that have incipiently rough surfaces extending to prominent tubercles and even prominent hair-like spines. So, indeed, if Latin names are said to have a reality they do not in fact possess, this certainly is the case here.
Acknowledgement. Dr. an Mrs. Lindley Kok of Klipfontein, Ms.Sonya Walmsley of Melkhoutkraal (?), Mr. Johannes van Eeden of Windsor. Kobus Venter and Gerhard Marx, for participation and company. I particularly appreciate the interest and assistance of Jakub Jilemicky in resolving an issue his name is associated with. ♦
7818 Windsor 56647818 Windsor 56697818 Windsor 56777818 Windsor 5688Fig. 4a. MBB7820 H. mirabilis ‘jakubii’. Goukou Bridge, Klipfontein. 5746Fig. 4b. MBB7820 H. mirabilis ‘jakubii’. Goukou Bridge 5750Fig. 4c. MBB7820 H. mirabilis ‘jakubii’. Goukou 5752Fig. 4d. MBB7820 H. mirabilis. Goukou 5755Fig. 4e. MBB7820 H. mirabilis. G
Gerhard Marx’ article in respect of H. marumiana and its associations is very welcome as he is one of the very few persons who make any constructive and useful observations that do not further tarnish the image of plant classification. But the article does require a response from me because it challenges my own observations and to a degree I think it misrepresents my decision making. Furthermore it suggests some kind of rift in which it is possible to make a better decision by looking more closely at less. What is this truth that has no respect for us humans? Gerhard writes his taxonomic priority list in which geographic distribution is placed last and then goes on to suggest that the flower has been too often ignored. This is simply not true and ironically this contention may be why we are in the situation we are in. Darwin stated that geographic distribution was the primary key to understanding species and if one looks at classification problems, in Haworthia at least, they can often be shown to weak decisions based on detail and superficial difference. This is despite the classic tenet of experienced taxonomists who were known to state “look for similarities rather than for differences”. Gerhard perforce has to evade the very problem he describes relating to the time and effort needed to make a meaningful study of all this floral detail. Gerhard goes on to examine differences between ‘archeri’ and ‘marumiana’, but he does not say what specimens from what populations he uses to generate the comparisons he makes.
Gerhard’s observations are not new to me and he does not venture to try and explain well enough why I felt compelled to add ‘archeri’ into ‘marumiana’ admitting myself that this is not easy or even correct. All the new populations unearthed from Beaufort West and westwards did not lead me to conclude that there was only one species. The word “conclude” is a misrepresentation for a far more conservative term, “hypothesize”. There is simply not a choice one makes between looking at a big picture and looking at detail and I do not doubt for one moment that ‘dimorpha’ may be better allied to ‘nortieri’ – if more was known. When seeing the “big picture” by virtue of physical time and effort one is required to visualize the still “bigger picture”. What happened to me in the process was to recognize that detail from a small sample was actually not very helpful and somehow one has to short circuit the data collecting process to arrive at a decision. There is no sound fence to sit on when it comes to formalizing a classification. If there is, it is a wire fence and the wire is barbed.
Gerhard does not elaborate on the distribution of ‘archeri’ nor give any indication as to geographically where ‘archeri’ and ‘marumiana’ may separate. This is of course where the problem is rooted as I could not, and cannot, do this either. It was evident to me that the area concerned was simply unexplored and it was mainly P. V. Bruyn exploring for Stapeliads that unearthed populations in inaccessible places. My own field exploration yielded the single population northwest of Langberg (Merweville) for which I simply do not have any detail to lean on. Similarly this applies to the population that Bruyns found in the upper Moordenaarskaroo northeast of Laingsburg at Klipfonteinberg. Using criteria of detail this latter one would probably have to be separated out taxonomically. The plants are very small, relatively plain, highly stoloniferous and thus extremely proliferous, but as cantankerous and sensitive to over watering as is ‘dimorpha’.
The only time I set out purposefully to look for H. marumiana west of Beaufort West was in May 2008 when my wife and I climbed a mountain northwest of Laingsburg, Lospersberg at Josefskraal, and found many plants of ‘archeri’ at high altitude (Figs. 2-9). The plants flowered in cultivation in February, which is correct for ‘archeri’, but the upper perianth lobes in the two clones that flowered for me were curved upwards which is incorrect. How does that help clarify anything? Especially when I chanced to look at a last remaining flower and observed that its upper petals had remained straight.
There a number of other problems such as the ‘marumiana’ on the Tierberg northeast of Prince Albert that I did not find easy to grow. In fact it may no longer be in cultivation as far as I know. Then there are the few populations from the mountains south of Prince Albert assigned tentatively to ‘viridis’ (see MBB7376 – the plants at Malvadraai brought to my attention by Dr. P. Forster). It would be foolish of me to suggest they support my “hypothesis” while it does seem that the problem out there in the very extensive areas of the Karoo is a more elaborate version of the problem I have in the Southern Cape (with say H. mirabilis and H. retusa where sampling is far more intensive. It is curious that Ernst van Jaarsveld assigned plants of H. monticola from the mountain west of Willowmore to H. marumiana. I do not know of any exploration of the mountains between there and the Swartberg where H. vlokii was first found.
There are other parts of the picture, either big or small, that Gerhard does not venture into. It is not helpful to speculate about a relationship with H. nortieri nor H. arachnoidea in the context of either ‘archeri’ or ‘dimorpha’, valid as it is, without properly recognizing all the subtleties of their variation or the problem populations that impose themselves on the problem. It is not helpful to latch onto my weak statement that the ‘nortieri’ population at Kunye southeast of Citrusdal was related to ‘dimorpha’, as I would today having seen the plants have no hesitation whatsoever in placing them firmly with ‘nortieri’. Also of course the vast variability within each of those complexes that could generate a score of contentious statements derived from detail of small samples. Added hurdles are:
1. A single record, again by Bruyns, of a population southeast of Laingsburg that suggests the entry of H. mucronata into the fray on a greater scale than Gerhard’s reference to H. arachnoidea var. nigricans suggests. The latter is in any case a stated array of variant populations that confound the separation of these two species.
2. Several populations in the Hex River Valley that do not suggest either single element or integration of H. nortieri and H. arachnoidea. This is an unopened can of worms that may indicate that even my conservative approach is not conservative enough.
It is in this latter set that ‘dimorpha’ may fall. The area is east of Touws River from where only H. pulchella is known. ‘dimorpha’ was first indicated by a record and specimen in the Compton Herbarium made by Harry Hall when he was curator of succulent plants at Kirstenbosch. His record stated simply that the plants were found under proteoid leaf debris in the vicinity of Konstabel Station. I made several attempts to find plants there but to no avail. Eventually I did find plants that seemed to match the original collection at a point between Konstabel and JandeBoers station where there were very few Protea. I disclose this locality now for two reasons. Firstly I find secrecy to be something that is just reflected back on myself and in total contradiction to the principles of science. Secondly it obfuscates understanding. The glamorization derived from secrecy gives the plant added value and collectors in any case seek to find plants. Often this is successful and new information obtained becomes similarly shrouded in secrecy and egocentricity. Hopefully knowledge of the habitat will rather inspire meaningful and ethical attempts to learn more about the plant elsewhere. What occurs between Konstabel and Pieter Meintjies? Is there anything in similar habitats south of the N1? What of westwards to Touws River and beyond? It is rugged unexplored terrain.
The plants are very cryptic and hidden on steep south facing rock slabs, hidden in dense black lichen. The fact that it is sandstone means the habitats are nutrient poor and may explain why in cultivation they grew unrecognizably to inspire the name “dimorpha” viz. ”twoformed”. These habitats are very dry in summer. In fact the debate ‘archeri’ vs. ‘marumiana’ is one that concerns the issue of a winter rainfall flora versus a summer rainfall flora, and plant species that have adapted to both. In a recent visit to monitor the population we found about 60 plants (see Figs. 16-20), very occasionally with more than one rosette of leaves, in a four-hour long search covering approximately a third of local available habitat. This is considerably more than we had seen on any previous occasion and confirms the established nature of the plants in a sustainable population.
Acknowledgement Mr. Johan Putter of Bijsteen Private Nature Reserve who granted reserved permission to enter the property and who is very mindful of the conservation status of his domain. I also acknowledge Gerhard Marx’s big contribution to a debate that might be needed to inspire ethical and accurate field exploration, reporting and interpretation. ♦
MBB7681
H. marumiana ‘archeri’
Josefskraal
MBB7681
H. marumiana ‘archeri’
Josefskraal
MBB7681
H. marumiana ‘archeri’
Josefskraal
MBB7681
H. marumiana ‘archeri’
Josefskraal
MBB7681
H. marumiana ‘archeri’
Josefskraal
MBB7681. H. marumiana ‘archeri’ Josefskraal
MBB7681 H. marumiana ‘archeri’ Josefskraal
MBB7681 H. marumiana ‘archeri’ Josefskraal
MBB7376 H. marumiana ‘viridis’ Scholtzkloof
MBB7376. H. marumiana ‘viridis’ Scholtzkloof
MBB7376. H. marumiana ‘viridis’ Scholtzkloof
MBB7376. H. marumiana ‘viridis’ Scholtzkloof
MBB7374. H. marumiana. ‘viridis’ S. Prince Albert.
JDV91-108. H. marumiana. ‘viridis’ Malvadraai, N. Swartberg Pass.
MBB 7819. H. marumiana.
‘dimorpha’ JandeboersMBB 7819. H. marumiana.
‘dimorpha’ JandeboersMBB 7819. H. marumiana.
‘dimorpha’ JandeboersMBB 7819. H. marumiana.
‘dimorpha’ JandeboersMBB 7819. H. marumiana.
‘dimorpha’ Jandeboers
My experience with Haworthia dates back to my childhood and on to nearly 70 years of observation. However, my interest was only able to properly manifest when I began work at the Karoo Botanic Garden in 1969 and it has since been through many phases. I wrote a formal taxonomic revision of the genus in 1999 and have spent a good bit of the last nine years adding to and verifying what I wrote. Haworthia has always been regarded as a problem child of botany to be avoided by professional taxonomists for various reasons including an apparent phobia of the many amateur collectors peering over the shoulder while at work. This has puzzled me because it seemed to me that if the need for good classification and identification was so strong there was an obligation on botany to provide the service. So my involvement has been largely by default. I was trained in an agricultural and entomological tradition with a totally different and unsophisticated approach to things like taxonomy, systematics and nomenclature. In the infant science that agriculture then was in South Africa, I can barely claim that my MSc is much more than an indication that I tried to learn something beyond normal schooling. While trained as an agricultural entomologist, my leaning was to plants and I eventually came to the Karoo garden to do what I liked best viz. exploring plants. Unfortunately the route is via identification and names and so I have walked a long road through the minefield that this is. Was this only in respect of Haworthia? No! This is a persistent misconception. Haworthia is only different because it has attracted such close and sustained amateur interest by so many for so long. I experienced failing classification in many other genera. To be fair I think the real reason is the lack of importance attached to the whole function of plant classification. It even seems as if many modern botanists pursue the study of plant relationship under the guise of systematics that is not committed to providing formal names and identifications.
The problem in the genus Haworthia begins with weak botany from the moment Linnaeus decided that there were four small plants that belonged together as one species in the genus Aloe. This alone generated a nomenclatural problem the dust of which is still to settle. A worse and obscured fact is that because of their small flowers these plants were and are still treated as one genus – one set. This is not true. The allied genera of the Alooideae of the Asphodelaceae are not comfortably classified and the three sets of plants that constitute the genus Haworthia are not only florally distinctive, they are also phytogeographically and behaviorally different. Thus while there was or is this huge flaw at this classification level, it is actually bizarre that sense can ever be thought to have reigned at the species level. The proliferation and confusion of names coupled with the associated argumentation about names has even led me to doubt my sanity, or even lose it as my critics will happily claim. My impression now as a non-botanist is that taxonomic botany somehow lost sight of the requirement that a biological classification should mirror and reflect the evolutionary processes or phylogeny that underlies our view of species and their origins. Worse, is the actual absence of a clear and unsullied definition of what a species is and hence what a Latin binomial as a supposedly formal scientific term of reference actually means. Cladistic methodology in plant taxonomy is in my opinion nothing more than a belated and even misguided attempt to arrive at or return to some semblance of scientific truth. Molecular biology is now being desperately seen as a holy grail that will summarily correct reluctantly conceded failures of the past.
There is no doubt that the mess in the classification of Haworthia is the product of non-botanists, but it is absolutely incorrect to exculpate the contributory role of taxonomic botany and professional botanists who are or were taxonomists. My interest has by no means been confined to Haworthia and the need for names has largely been driven by a perpetual curiosity about many different plants in the field environment. I am, by the route my life has taken me, unusually and almost uniquely experienced in the relation between botany, botanists and interest of the layman in plants, as it is reflected in the names we use for them and how we respectively do so. Consequently I think that I am now qualified to express some opinions on what Haworthia can teach us quite apart from what the hostilities that wage across the whole taxonomic front suggest.
I pointed out rather timorously that in Oxalis, morphological characters were not as indicative of species as revisions and identification keys generally seem to imply. The dramatic changes in our classification systems that the results of molecular biology now seem to threaten, is a manifestation of this simple fact rather than any great new understanding of what really goes on in the plant kingdom. My fear is that molecular biology is likely to fudge most of its promise because the basic tenets of classification, the mistakes of the past and the reality of the field are still not properly appreciated. This is what I think Haworthia has to tell us. I made use of Oxalis to test my observations that we can look at Haworthia as composed of elements that reflect the very nature of the habitats and environments where they are found. I did this because Haworthia plants simply do not have any array of characters by which species can be arrived at whereas Oxalis has a plethora of such apparent (taxonomically bewitching, tempting, and misleading) means. While the few facts that there are in respect of differentiating Haworthia into three groups (subgenera as presently constituted) has been lost and confounded in the broader classification of the Alooideae there is not much else to help verbally to define the species. It is not to say that modern technology has not been resorted to, to help clarify the issue. Both cladistic and molecular studies have been done but the results flawed by weak hypothesis and perhaps just weak science too.
How then do I think sense can be made of Haworthia? It is firstly by defining the species as dynamic fractal systems that illustrate the responses of plants to skeletal habitats and geological substrate in the wider context of landscape and topography across the total distribution area. Furthermore, there is an obvious correlation to be made with vegetation and other biological systems both plant and animal. I cannot pretend to know a great deal about any of these things, but also suggest that nothing more is needed other than awareness that this is what biological diversity and species are essentially about anyway.
It has taken me nearly 40 years to arrive at the big picture of Haworthia. It is as a non-taxonomist that I say categorically that the three main elements in the present genus need to be seen and registered as “different”. Then we can focus on the subgenus Haworthia where the greater difficulty lies and which I believe offers the greatest insight into what plant species are and what the difficulties are in recognizing and defining them. Also into what Latin binomials should or might mean.
For this I am going to take just two species systems and ignore some of the peripheral facts that might confuse the issue rather than illustrate the profound nature of species interactions. I take H. retusa (Figures 1 to 4) and H. mirabilis (Figures 5 to 10). In order to do this I have to bypass even my own conservative classification by considering H. turgida and its many variants to be the riverine and cliff dwelling version of H. retusa. H. mirabilis is more complex and I have to include three other species viz H. maraisii, H. magnifica and H. heidelbergensis for which spatial factors and major geological substrate are the prime differentiators. Both my two selected species are highly variable both within and between populations. It is in fact the intra-population variability that I underestimated. Growing plants from field collected seed can result in a hundred or more individual plants that are all different in appearance.
The two species share the same distribution range, but with an extension of H. mirabilis into the Worcester/Robertson Karoo, while H. retusa extends further east from Albertinia to Great Brak. H. mirabilis flowers in late summer and H. retusa flowers in spring. They never grow in direct association! The populations of each are highly localized and when in proximity are usually separated by hundreds of meters, and difficult to ascribe to habitat conditions given the ranges of habitat that respective populations may occupy. I have already noted and warned of the reality that there is evidence of interaction between these two clearly defined systems. In addition, somehow or other, the species H. pygmaea emerges as discrete in the eastern Mossel bay area, while H. mutica sits in an island between Swellendam and Riviersonderend in the west.
What is most striking is the variability within each system and this is where the crunch comes and what my pictures should illustrate. They are but a small sample of many hundreds and I have adopted an informal method of captioning the pictures as the formalities of nomenclature would need to be followed to depart as I have from the existing status. Not only are my pictures supposed to illustrate the problem of variability within species, figures 2 and 3 show variability within a population and figures 3 and 10 show two very similar plants from two quite different species systems. Note that the plant in figure 3 is in flower and that the similarity to figure 10 is rather exaggerated by photograph scale as in H. mirabilis ‘toonensis’ the plants are smaller.
Taken individually from the morphological extremes between and even often within populations, it will stretch the credibility of the most ardent taxonomic lumper to claim that any two of these are representatives of the same species. It is an unpalatable truth that things that look different are the same just as the converse is that things that look the same are different. When this reality is properly conceded by the professional taxonomist, perhaps we will succeed in demystifying and deglamorising the Latin binomial. Botanists who are not taxonomists will arrive at a better understanding of their plant world and consequently we who look to them for understanding will be better served. We need to be more realistic about plant description and identification and also correspondingly more conscious of the real nature of diversity. Conservation is not to be seen as maintaining a list of names for which there is a use. It is to be seen as maintaining the options for change and the conservation of a far greater diversity than any list of names can hold. It is botanists (quoted in Levin, 2000) who wrote “our system of names appears to achieve a reality which it does not in fact actually possess”. I think this implies that even the Latin binomials we so modestly, reluctantly or even portentously use do not convey what they should; but rather the product of our individual limited experience, knowledge and perception. My feeling is that species do have a reality and for Latin names to have any sense, they need to be more circumspectly applied. There is little merit in having a highly regulatory nomenclatural code and little value in what the names actually refer to. ♦
In Chapter 1 of Haworthia Update Vol. 2, I discussed H. chloracantha and H. parksiana in the context of H. floribunda. Fig. 8 in that publication is labelled “North of Herbertsdale” when in fact it is MBB7425 from the Wolwedans Dam north of Great Brak (see fig.1). This was deliberate and not seriously misleading as the plants from the two respective populations are virtually identical. The correct images for that “north of Herbertsdale” are in Haworthia Revisited and labelled JDV87/80 and 97/138.
Recently I had an opportunity to visit Herbertsdale in connection with chameleons and took the opportunity to visit a locality also north of Herbertsdale, but a lot closer (see Figs 2.1-2.11. MBB 7825) reported to me by Mr. Jaap Viljoen of Swellendam.
The plants there are smaller and much less clump-forming than JDV87/80=97/138 and are more reminiscent of H. parksiana. Unlike JDV97/136 (see figs. 3) that are solitary, small, dark-green plants growing in the lichen on a steep east-facing conglomerate slope, the plants are quite green in colour and often the tips of the leaves are slightly rounded and flattened as in H. floribunda. The habitat is west-facing on an old gravel river terrace and much drier and warmer than for JDV97/136, in sharp contrast to the very steep north-facing clay cliff where KDV87/80=97/138 occurs. In neither of these latter two sites are other plants significant, while at MBB7825 the plants are under renosterbush (Elytropappus) and related vegetation. I should mention that my wife on seeing the plants commented on the similarity to H. parksiana and this is not an observation that can simply be dismissed.
About 3km south of Herbertsdale (see fig. 4, MBB7827) we again found very similar plants, perhaps a little more spinose and slightly more clump forming. Neither of these two new populations fit comfortably in the existing formal varieties, as indeed is true for so many populations in most of the other species. I do not think it is rational to attempt formalizing names for them as there is quite substantial variation in each population. Others may do so if these plants ever get into general cultivation from vegetative propagation and onto lists.
From Herbertsdale we went to Klein Brak to investigate a set of four populations on the farm Rooiheuwel as indicated to me by Gerhard Marx. One of the species supposedly there was H. pygmaea and it was rather distressing to find that the Water Affairs Department has seen fit to run a bulldozer over the area with no indication that there was any need for doing so. However, we did find the other three species indicated by Gerhard. One is H. kingiana (see figs. 5.1-5.4 MBB7835).
The others were H. parksiana (see figs 6 MBB7828)
and H. chloracantha (see Figs 7 MBB7829), but this requires a little bending of the truth.
This H. parksiana site is about 400m southeast of an old locality of mine on an outcrop of rock virtually jutting into the road and now largely obliterated. At Gerhard’s site (viz MBB7828) the plants are small and while there is little doubt that they are generally alike to H. parksiana, there is definitely some degree of hybridization and some of the plants may actually be H. chloracantha. This is a rather difficult decision to make on the basis of stressed plants in habitat. This habitat is very restricted to a patch of fairly level aggregated conglomerate so that the plants are virtually in tiny shallow soil and lichen encrusted patches. A little distance further north there is another population to which we assign the name H. chloracantha. This is above a steep loose conglomerate cliff with a little more soil and immediate bush. The plants have rather abbreviated leaves and a darkish colour. There must be some doubt that they are discretely H. chloracantha. The habitat is not so restricted and one is compelled to ask why the plants then so are.
I try to avoid the unanswerable question of how these two species that flower together and seem to be able to hybridize quite freely, maintain their identity and come to occupy such specific, localized and widely separated sites? It would be nice to think that the technology is now seemingly available in DNA sequencing to begin probing these questions, and I will point, with reference to chameleons (see next chapter), to what I think is a disturbing reliance on that technology without the necessary knowledge and experience of what actually can be seen in the field. ♦
In some cases acknowledgements have already been made in respect of some of the populations discussed in these chapters. New acknowledgements are Mr. Artho Saayman of Platkop, Mr. and Mrs. Wilhelm Zietsman of Kruis Rivier, Mr. and Mrs. Coetzee Uys of Morning Star, Messrs. Chris and Pieter van Deventer of Kransriviermond, Johan Groenewald of Buffeljags, Clive and Duprecia Stramrood of Kadies Landing, the Uys brothers of Sandfontein and Meerlus, the Steenkamps of Appelbos, Annatjie and Jaap Viljoen of Swellendam, Hector Odendaal of Dankbaar, Ryno Stander of KomseRandte, Tineke Kraaij and Carli Venter of Bontebok National Park, Anne Lise Vlok and Rhet Hisemann of W Cape Nature Conservation Board, Stiaan Conradie of Lower Breede Conservancy. Others such as to Mr. and Mrs. Anna and Arno Steenkamp of Anna’s Farm (Oudekraalkop) for their generosity and hospitality, Mr. and Mrs. Hennie van Deventer of Koppies, Mr. Dirk Papendorp of Voorstekop and Uitvlugt, Mr. Uys Willemse of Goedverwagting, need repeating. We are most grateful to Dr. Paul Taylor for an albeit failed excursion to the west bank of the Duiwenhoks River and similarly to Nico DeJager of Victoriasdale for an interesting but fruitless search of the Wankoe Randte and a little more success on Klipheuwel, Mr. H. Eksteen of Grootkloof and Mr. M. Dippenaar of Diptka for so kindly allowing us access to their properties. There are many other landowners and people we contacted with less significance in respect of places actually visited or in terms of plants found but no less in respect of the universal kindness and helpfulness of landowners.
There are many other peripheral and significant contributions from people (not necessarily agreeing with what I say nor how) such as Prof. Richard Cowling, Dr. Syd Rhamdani, Diederik van den Abbeele and even people who have no connection to Haworthia, but who share the same passion for nature and for plants … Mrs. Hettie Conradie of Worcester who makes us feel as though it is we who own her home and Mrs. Anso LeRoux who may both be housewives, but biologists by nature. Gordon Rowley has demonstrated the value of humour in a field where there seems naught else to do but tear hair out. Etwin Aslander in his quiet and effective way has been a great friend.
Then there is Gerhard Marx, whose dogged refusal to blindly accept anything at face value, has been most helpful in so many respects. Kobus Venter has always been a great friend and mentor in relation to the ways of the world and I am extraordinarily grateful for his unwavering support and kinship. He and Mirna, in graciously allowing us the use of their holiday home at Stilbaai facilitated this work. Steven Hammer has been another remarkable personage in my life and I am most grateful for him being the sensitive, perceptive, empathic and remarkable man he is.
Lastly where I have used the term “we” I have included my wife, Daphne. This is actually with great reluctance because anything to do with taxonomy and nomenclature brings with it a cloud of derogation and negativity – what to say of the negative attitudes that surround the word “collection”. It seems that in past civilizations there was great respect for nature and many of the rites and rituals were an appeasement to nature for the benefits she bestowed in terms of what she provided in the way of food, clothes and shelter – the basic needs of man. Daphne has provided me with the trust and support of a true companion and I regard her very dearly. There is no need for her to remind me, as she seldom does, that few other housewives would traipse so blithely and happily through the wilderness with someone who does not seem to know where he is going or what he is doing next.
I feel a need to explain again that I became a taxonomist by default where there are also systematists and nomenclaturalists and I would not wish to be any of these. We as individuals are all inclined to make bold and general statements based only on what we know and ignore the very much greater body that we do not know at all. I would say that it is depressingly sad that information is processed and distributed as knowledge often from a very limited experience and limited data base. After 45 years of this I actually am painfully aware that there is still a lot that I do not know about. I make no apologies for a revision that is precisely the sort of product that comes from constant testing of a hypothesis. I take it very remiss of a publisher that did not honour the integral part of a contract to further research and validate the work. I am extremely grateful to Harry Mays for publishing a vast series of articles that were originally intended for a home based publication but mindlessly lost in a mire of conflicting interests.
While Alsterworthia so kindly published the Update Volumes 2, 3 and 4, I continually feel that this is not the environment I am comfortable in. Bill Keen was one of the first to publish anything of mine and is the epitome of server to a group. My wish is to see and serve a user group such as the Haworthia Society and see it go from strength to strength. ♦