The ‘retuse’ haworthias are among the most fascinating of the genus and therefore Colonel Scott’s recent revision is of particular interest. This work…‘A revision of the genus Haworthia, Section Retusae” was published in ALOE, Dec.1973 and forms a new taxonomic record of the species involved. The object of the present paper is to examine the validity of this record particularly in the light of past history of Haworthia. A considerable literature on Haworthia is now available much of which points to the problems likely to be encountered in the group. Much of it also points to the consequences of inadequate species concepts for the group.
Printed in Excelsa 5:83 (1975).
(This article was subsequent to the revision of the Retusae by C.L.Scott).
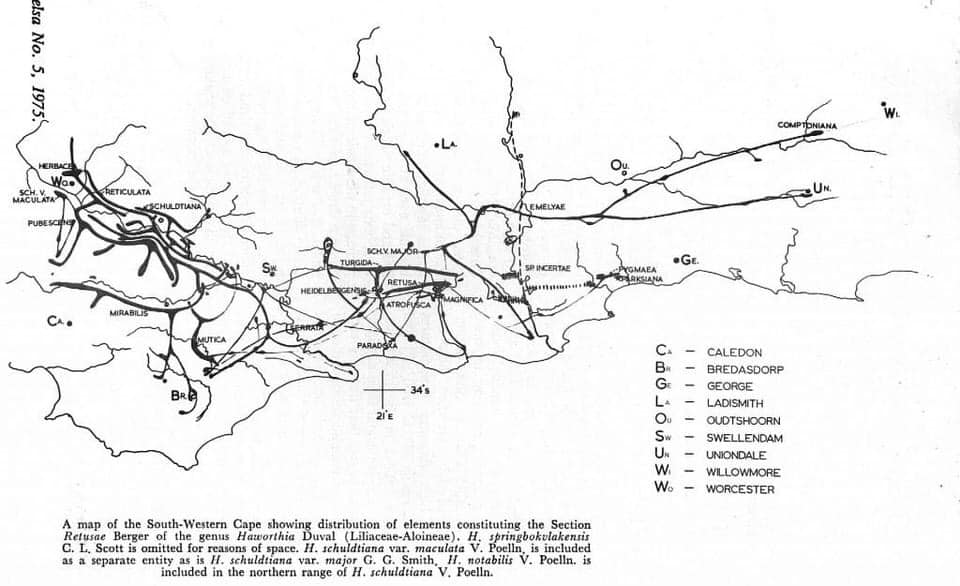
The 162 named species in the genus Haworthia Duval have been split into 20 sections of which Retusae Haworth is one. The clearest and most obvious subdivision in Haworthia is into three subgenera which are based on both floral and vegetative characters. The Retusae fall into the subgenus Haworthia which contains the soft- leaved stemless species in which the three outer petals of the flowers completely enclose the inner petals. Division within this subgenus is the most difficult of the three and thus the composition of the section Retusae is understandably problematic. The type species of the section is H. retusa (L.) Haw., so named on account of the fat, bent, thumblike leaves. In theory this type of leaf should characterise all the other species in the section. However, in the field it soon becomes apparent that apart from the difficulty in recognising and identifying “species” related to H. retusa, some of these related elements do not have retused leaves. What then is the composition of the section Retusae and how can the individual species be recognised? So far the only really satisfactory criterion we have is the geographic distribution and relationship of a large series of rather localised populations. The individual plants in each of these populations differ to a greater or lesser degree from each other, and in similar manner, the populations differ from each other. Thus we have variability within and between populations. The composition of the Retusae is based here on visual assessment of the discontinuities of this variability, on geographic distribution, and on habitat.
32. Haworthia retusa (L.) Duval, Pl.Succ.Hort.Alenc. :7(1809). Haw., Syn.Pl.Succ. :95(1812). Bayer :150(1976). Bayer in Excelsa 8:46(1979). Bayer :53(1982). pp Scott :112(1986). Aloe retusa L. Sp.Pl. :322(1753). Haw., Trans.Linn.Soc. 7:9(1804). Type: icon, 2:t6 Commelin, Hort.Amstel.(1701): H. foucheii V.Poelln., Succulenta 22:28(1940). Type: Cape, Riversdale district, Grootvlei, Fouche. Not preserved. Neotype (designated here): CAPE-3421(Riversdale): Near Riversdale, Grootvlei (-AB), Dekenah 212 (NBG): H. retusa var. multilineata Smith, JS.Afr.Bot. 12:3(1946). H. multilineata (Smith) Scott :135(1985). Type: CAPE-3421(Riversdale): 3km N. Riversdale, J. Dekenah 83 in Smith 5383 (NBG): H. retusa var. solitaria Smith, JS.Afr.Bot. 12:5(1946). H. solitaria (Smith) Scott, Aloe 11:37(1973). Type (designated here): CAPE-3421(Riversdale): along Corrente River, 10km N Riversdale, Dekenah 5 in Smith 5373 (NBG): H. retusa var. densiflora Smith, JS.Afr.Bot. 12:7(1946). Type: CAPE-3421(Riversdale): Riversdale district, Venter 106 in Smith 5056 (NBG): H. geraldii Scott, JS.Afr.Bot. 31:123(1965). Scott, Aloe 11:22(1973). Scott :132(1985). Type: CAPE-3421(Riversdale): 5km east of Riversdale, Scott 72 (PRE).3
retusa: with leaf-tips bent back thumb-like.
Rosette stemless, slowly or seldom proliferous, to 12cm φ. Leaves 10-15, turgid, rigid, with pronounced retused end-area, pointed tips, variously lined and windowed. Without surface spination and usually without spination on margins and keel. Colours brownish or green and seldom purpling. Inflorescence simple, robust, to 30cm. Flowers compacted on inflorescence, white with greenish-brown veins.
1982 – H. retusa is very closely associated with H. turgida, and an eventual re-evaluation of this association could lead to a total upheaval of species concepts in this group of haworthias. H. retusa is taken to be an assemblage of forms in the Heidelberg and Riversdale areas, with clearly defined end areas, and points, to the leaves. The various forms may or may not be proliferous, for example the var. solitaria was largely solitary, whereas the form described as H. geraldii is very proliferous. The forms vary in cultivation and both dark and light green forms may occur. The darker forms may have some relationship to H. magnifica, and the light forms to H. turgida. The species is not well known in the area between Heidelberg and the Breede River and hence it is not known whether or not H. mutica and H. retusa intergrade. H. mutica may simply be a blunt‑leaved form of H. retusa. The var. dekenahii from Albertinia is now thought to be represented by several populations in which plants have leaves very silver‑spotted ‑ a phenomenon which occurs in H. turgida too. This variety may eventually be shown to link H. retusa and H. pygmaea. There is still an apparent break in the distribution of these two species between Mossel Bay and Albertinia. The var. acuminata tends to be darker green and the leaves are very acuminate. It occurs in the south‑east of the distribution range of the species and may also be expected in the area southwest of Albertinia ‑ still unexplored.
1999 – This revised treatment fulfils the predictions of 1982 amid the realisation that the views expressed there are correct, but stopping short of uniting H. retusa with H. turgida. H. longebracteata Smith is now here regarded as a variant of that ubiquitous species and strictly H. geraldii Scott should be treated similarly. However, the key issue is that H. retusa, as perceived here, is the robust, generally solitary forms which occur only in the Riversdale area. Scott’s treatment is rather fortuitous in that he typifies the name in the same way, but applies it to the smaller clump-forming elements which comprise H turgida. This is evident from his synonymy but totally compromised in his discussion where:- 1. he says it is restricted to one locality at Riversdale and two to three in the Little Karoo, and 2. the distribution map where localities at Little Brak and at Tradouw pass seem to be indicated. Breuer and Metzing’s argument for the creation of an epitype, and also their choice, is also unfortunate. The Commelin illustration could hardly be more distinctive and apart from Col Scott who muddled H. retusa and H. turgida, there has never ever been any sign of doubt about its application. Furthermore, they select a specimen from the source of H. fouche. The Commelin, illustration depicts the acute leaf-tips of the species very well and there is little chance of confusion with H. mutica.
In the species concept for the genus, co-occurrence and consequent interaction, or lack thereof, are criteria for recognising species. In the field it has become obvious that the interaction between species revolves around H. turgida as a main role-player and not H. retusa. There is no interaction between the latter two named elements as they do not co-occur. The three species recognised by Scott viz. geraldii, fouchei and multilineata give a very good impression of the variation of this one species in the Riversdale area. The var. solitaria may represent interaction with H. magnifica, and the vars. acuminata and dekenahii are both transferred to that species. H. dekenahii var. argenteo-maculosa is treated as a variety of H. pygmaea. The problem plants around Heidelberg are associated with H. mutica.
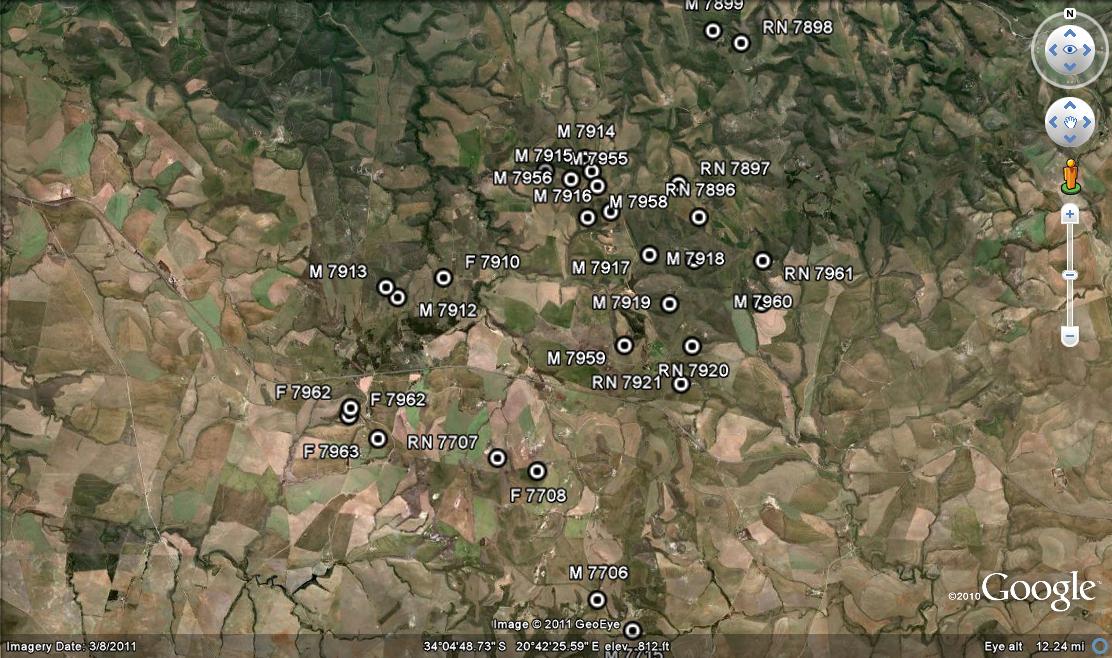
Distribution: 3421 (Riversdale): 8km W. Riversdale (-AA), Smith 5443 (NBG); Along Corrente River, 10km NW. Riversdale (-AA), Dekenah 5 in Smith 5373 (NBG), Dekenah in KG305/71 (NBG); 4km N. Riversadel (-AA), Smith 5493 (NBG); SW. Riversdale (-AA), Smith 5387 (NBG); Near Riversdale, Grootvlei (-AB), Dekenah 212 (NBG), Smith 4955 (NBG), Fourcade 263 (NBG), Bayer in KG627/69 (NBG), Bohnen 9058 (NBG); Blinkbonnie (-AB), Smith 6086 (NBG), Venter in KG 156/71; Zeekoegat (-AB), Smith 6089 (NBG); Ferguson Drive (-AB), Smith 5380 (NBG, PRE); 3km N. Riversdale (-AB), J. Dekenah 83 in Smith 5383 (NBG), Smith 5374, 5488 (NBG, PRE), Smith 6796 (NBG); 5km E. Riversdale (-AB), Scott 72 (PRE), Smith 5377 (NBG); Vet River Road (-AB), Smith 5383, 5387 (PRE); 14km E. Riversdale (-AB), Smith 5749, 7320 (NBG); Bolus 2868 (PRE); Commonage (-AB), Smith 5377 (PRE); Dekenah 11 (PRE); Riversdale (-AB), Fouche in PRE 34869; 3km E., Grootvlei (-AB), J. Scott 1778 (PRE); 3km E. (-AB), J. Scott 93 (PRE);
Inadequately located: Riversdale district, Venter 106 in Smith 5056, Smith 3432, 3850, 3919, 3962, 5043, 5056, 5599 (NBG), Bolus 11390 (BOL), Helm in NBG1746/32, Malherbe in NBG399/40, ex hort, Wilman in PRE 34904.
Haworthia retusa JDV93/57 east of Riversdale. Some forms are reminiscent of H. magnifica.Haworthia retusa JDV89/26 south of Riversdale. Others are enormous and range from brown to yellow.Haworthia retusa JDV92/122 southeast of Riversdale. A motly assemblage where some are conspicuously veined.Haworthia retusa JDV92/122 southeast of Riversdale. Some clones remain very compact.Haworthia retusa JDV92/122 southeast of Riversdale. Despite the size there is an attempt at concealment.Haworthia retusa JDV92/69 southeast of Riversdale. A cultivar referred to as ‘Jolly Green Giant’ probably originated in this area.Haworthia retusa JDV87/5 east of Riversdale. The well known fa. geraldii, which is not accepted in this treatement although the name could be retained for practical and historical reasons.Haworthia retusa JDV87/5 east of Riversdale. This form is only known from one small site where it forms massive clusters.Haworthia retusa JDV92/125 east of Riversdale. This is the fa. fouchei, occuring a stone’s throw from geraldii but also not upheld for the same reasons.Haworthia retusa JDV92/125 east of Riversdale. As with H. magnifica var. magnifica, there are two populations of this H. retusa variety on the same ridge. This is also the fa. fouchei, recognized by its erect leabes.Haworthia retusa JDV92/125 east of Riversdale. The leaves are more erect and there is continuity down the Kafferkuilsrivier with H. turgida.
[ed. Bayer subsequently resurrected some of the varieties.]
H. retusa (Linné) Duval (Pl. Succ. Horto Alencon., 7, 1809). Type: [lecto — icono]: Commelin, Horti Med. Amstelod. 2: 11, t. 6, 1701. — Distr: RSA (Western Cape); Fynbos vegetation.
H. retusa var. longibracteata (G. G. Smith) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type: RSA, Western Cape ( Dekenah 18 in Smith 5378 [NBG, PRE]). — Distr: RSA (Western Cape). I: Scott ( 1985: 127, as H. longibracteata).
≡ Haworthia longibracteata G. G. Smith (1945) ≡ Haworthia retusa fa. longibracteata (G. G. Smith) Pilbeam (1983) ≡ Haworthia turgida var. longibracteata (G. G. Smith) M. B. Bayer ( 1999).
Differs from var. retusa: L erect to suberect, ovate-lanceolate.
H. retusa var. nigra (M. B. Bayer) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type: RSA, Western Cape ( Smith 5753 [NBG]). — Lit: Bayer ( 2004a); Bayer ( 2004c); Bayer ( 2005); all as H. mutica var. Distr: RSA (Western Cape: Heidelberg area).
≡ Haworthia mutica var. nigra M. B. Bayer ( 1999) ≡ Haworthia silviae var. nigra (M. B. Bayer) M. Hayashi (2000); incl.Haworthia chromutica M. Hayashi (2000) ( nom. inval., ICN Art. 39.1, 40.1); incl.Haworthia quimutica Breuer ( 2011) ( nom. inval., ICN Art. 38.1a).
Differs from var. retusa: Ros more proliferous; L greener and more translucent.
H. retusa var. retusa — Distr: RSA (Western Cape: Riversdale). I: Bayer ( 1982: Fig. 33a).
Incl.Haworthia fouchei Von Poellnitz (1940) ≡ Haworthia retusa fa. fouchei (Von Poellnitz) Pilbeam (1983) ≡ Haworthia retusa var. fouchei (Von Poellnitz) Breuer (2016); incl.Haworthia retusa var. densiflora G. G. Smith (1946); incl.Haworthia retusa var. multilineata G. G. Smith (1946) ≡ Haworthia retusa fa. multilineata (G. G. Smith) Pilbeam (1983) ≡ Haworthia multilineata (G. G. Smith) C. L. Scott (1985); incl.Haworthia retusa var. solitaria G. G. Smith (1946) ≡ Haworthia solitaria (G. G. Smith) C. L. Scott (1973); incl.Haworthia geraldii C. L. Scott (1965) ≡ Haworthia retusa fa. geraldii (C. L. Scott) Pilbeam (1983) ( nom. inval., ICN Art. 41.5); incl.Haworthia retusa var. quimutica Hayashi (2001); incl.Haworthia subretusa Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a).
Ros stemless, rarely slowly proliferating, to 12 cm ∅; L 10–15, turgid, rigid, with pronouncedly retuse end-areas, 8 × 2 cm, brownish or green and rarely with purplish hue, variously lined and windowed, surface and usually also margins and keel without spines or tubercles, tips pointed; Inf robust, to 30 cm; Fl 20–30, closely spaced, white with greenish-brown veins.
Possible hybridization with H. mirabilis is mentioned by Bayer ( 2012d).
H. retusa var. suberecta (Von Poellnitz) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type [neo]: RSA, Western Cape ( Bayer s.n. in Karoo Garden 631/69 [NBG]). — Distr: RSA (Western Cape). I: Scott ( 1985: 126, as H. dekenahii).
≡Haworthia turgida var. suberecta Von Poellnitz (1938) ≡ Haworthia turgida fa. suberecta (Von Poellnitz) Pilbeam (1983) ≡ Haworthia suberecta (Poellnitz) Breuer ( 2010); incl.Haworthia turgida var. subtuberculata Von Poellnitz (1938); incl.Haworthia turgida var. pallidifolia G. G. Smith (1946) ≡ Haworthia turgida fa. pallidifolia (G. G. Smith) Pilbeam (1983) ≡ Haworthia pallidifolia (G. G. Smith) M. Hayashi (2010) ≡ Haworthia suberecta var. pallidifolia (G. G. Smith) Breuer (2016); incl.Haworthia pseuda Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a); incl.Haworthia reflexa Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a); incl.Haworthia rodinii Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a).
Differs from var. retusa: L strongly mottled, tips slightly truncate and rounded.
H. retusa var. turgida (Haworth) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type [neo]: RSA, Western Cape ( Bayer 2420 [NBG 132378]). — Distr: RSA (Western Cape). I: Bayer ( 1982: Fig. 40, as H. turgida).
Differs from var. turgida: Ros partially stemless, proliferous, 5–10 cm ∅; L 20–40, ovate-lanceolate, 4 × 1.2 cm, turgid, often as thick as broad, recurved or slightly retuse, margins and keel lightly spined.
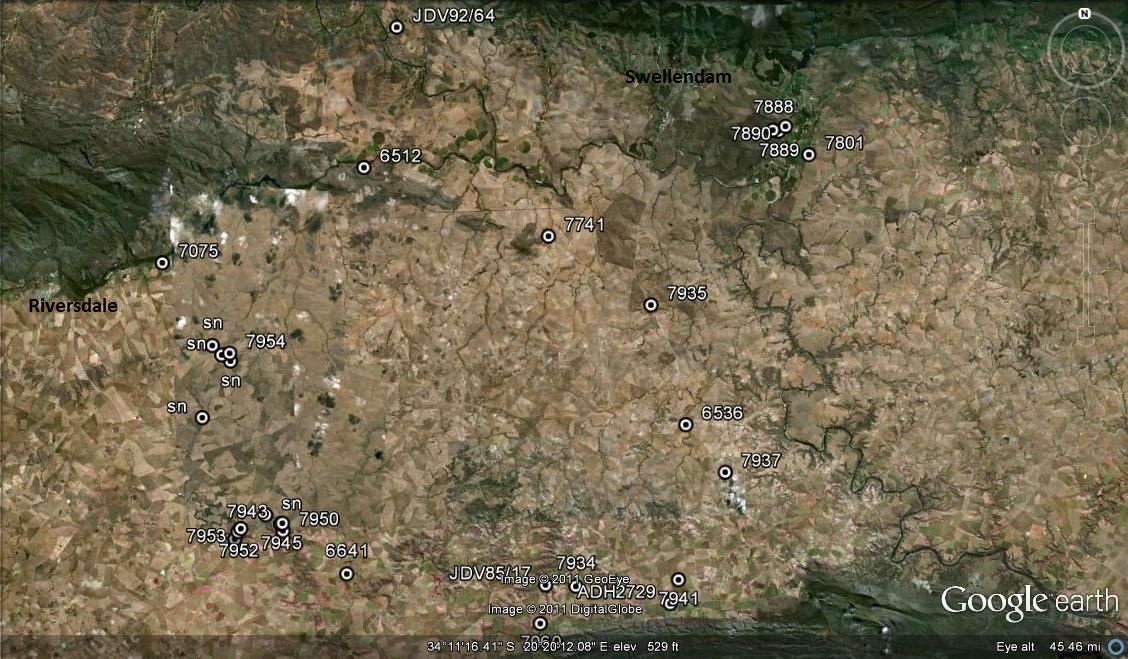
It has long been my stated contention that H. turgida is in fact a rock face ecotype as opposed to the solitary flat growing H. retusa. Thus one should expect the multiplicity of forms that are found between, and consequently superfluous to say within each, these two primary types. There are problems outside of this and I will deal with those in the Chapter Haworthia enigma. Here I am simply going to present pictures representing plants in nine populations of the species. Most of these populations are of the “typical” solitary form and they all demonstrate variation to greater to lesser degree. Perhaps some special mention should be made of the element H. mutica var. nigra. I have written at length about this and in doing so strayed widely into H. magnifica and its var. atrofusca (both falling now under H. mirabilis). This is because it is quite certain that there is an element of interaction in the field between the prime elements H. retusa and H. mirabilis that this summation is intended to expose. The first known H. mutica var nigra from Kransriviermond is possibly the product of such interaction, whereas all the subsequent collections from northwards and westwards are now perceived by me to be variants of H. retusa and H. retusa ’turgida’ (to use a more informal and flexible way of communicating).
1.MBB7776 Pienaarsrivier. This is a population just west of Kweekkraal and the agglomeration of populations that I dealt with in a series of articles covering the diffuse nature of ‘nigra’. The population was first brought to my attention by Etwin Aslander but finally shown to me by Kobus Venter. It is extraordinary that it is not better known in the literature and again a vivid demonstration of how thoroughly one has to explore to uncover the mysteries of what constitutes species. It has always been difficult to adequately illustrate plants and probably the very weak appreciation we have for variation is precisely due to the fact that often species ere even described without any accompanying illustration. If there was one it was taken to be the exact image of many such that could all go by the same one name and this a Latin binomial. This is the paradigm that needs to change.
Pienaarsrivier – truffeling.
At Pienaarsrivier, the habitat can be described as an erosion product of the ferricreteinselbergs that are so characteristic of the lower areas of the Southern Cape. However, it is an extremely complex situation as there are high and low level gravel terraces as products of direct erosion off mountain sides, river gravels as a product of countryside drainage and then the iron rich ferricretes themselves as the products of deposition along groundwater movement. Here at Pienaarsrivier there is dense Renosterbos and grass among fairly large sized stones. The plants are abundant and they flower in October. I have elsewhere shown pictures of H. retusa from both eastwards at Kweekkraal (two populations) and from Heidelberg itself (many). The Pienaarsriver plants tend to have a very purplish hue and often the leaf tips are very rounded (mutica!) ♦
2. MBB7754 H. retusa ‘turgida’. Brakkekuil. What is most significant about this population is its whereabouts that highlights the overwhelming importance of distribution and geography. The drainage systems (or parts of them) of the Southern Cape drain southwards from the mountains to the sea e.g. Gouritz, Goukou, Duiwenhoks and Lower Breede. These are important especially when it comes to the habitats in the way of exposed rock and steep faces that favour plants requiring skeletal soils. Brakkekuil is on the Slang River that drains southwestwards from near Heidelberg to flow into the Breede River near Malgas. H. retusa ‘turgida’ has not been reported for this entire river system before, while it is present on the Breede River and even westwards at Bredasdorp. So the Brakkekuil population is significant and also significantly different. The plants are neither strictly solitary nor greatly clump-forming and it is not really surprising as this mirrors what happens with H. cooperi in the Eastern Cape in situations that are neither fully cliff face nor plain. The Brakkekuil plants are on the surface of a rocky shale knoll with plants enduring direct exposure to northwestern sun as well as obtaining refuge in the more vegetated and protected slightly southern aspect. It is quite difficult to make reference of individual plants to Latin names, in that variation is already ensconced in the existing system viz. ‘longibracteata’. I gladly concede that all the old names, as Rowley has suggested, can be paraded out again and made use of. In fact I have also said that this is how the contribution of Breuer and Hayashi can be fruitfully used. For my reality this population is H. retusa ‘turgida’ Slangrivier. It is quite the most variable population of the ‘turgida’ side of H. retusa that I have ever seen and there are plants that resemble the more sandstone associated variants (‘caespitosa’) at, say, Tradouw Pass as well as individuals that compare with some of the other populations I will cover from the ferricrete inselbergs. Another very significant observation is the similarity of some plants to those that can be found in H. mirabilis ‘paradoxa’ that is not very far away to the southeast at Vermaaklikheid. There is no doubt that if a full and real understanding of natural systems is to be found it will lie in the realization that even my suggestion favouring a “super species concept” may be conservative. It is actually curious how my treatment of that has been met by readers who have been kind and considerate enough to communicate with me on the issue. The ‘super species” proposal actually comes from Prof. Canio Vosa. It is and was not, any attempt to confound anyone or obfuscate the issue. Prof. Vosa is directly addressing the issue that we have a classification that is a sorry marriage of scientist and layman user groups – both ignorant of the full extent of the field situation.
3. MBB7556 H. retusa ‘longibracteata‘. Dassieklip. There are ‘turgida’ populations all along the Duiwenhoks River. Some of these could be ‘turgida’ and some could be the variant ‘longibracteata‘. These at Dassieklip are close to the latter and are reminiscent of some of the large slender leaved plants one finds close to Heidelberg itself
However, the two plants illustrated are a few hundred meters west of the river and survivors of road materials quarrying. There are many more plants along the river itself and also at Brakrivier between Dassieklip,and Vermaaklikheid. But south of Dassieklip the river drainage passes from shale to calcrete and something really interesting may occur on the interface of these two geological formations northwest of Vermaaklikheid. Curiously no H. retusa is recorded at all between Brakrivier and Stilbaai to the east and this may be due to the calcrete substrate and the absence of suitable habitat.
4. MBB7758 H. retusa ‘retusa’. Skietbaan. Riversdale. This is a very well known population and one of many in the general area extending slightly south and then west rather than east, although there is a population in the rocks of the Goukou river itself to the east. The plants are on a shale substrate and huge with again the expected variation that has been so obscured by the limited illustrative material of the historical past. The habitat is highly disturbed and it is in fact surprising that it has survived here so successfully. A road and a fence cross the site that follows the scale of the relatively confined areas the plants are usually found in. There are a heap of epithets that could be mobilized to personalize these plants and even ‘solitaria’, ‘geraldii’ and ‘fouchei’ could probably find application despite being out of true context.
5. MBB7240 H. retusa. Skeiding. This population seems to be very relictual and I did not see many plants. I include it just to indicate that these ferricrete associated forms are here and they are trespassing into the series of populations that I discussed in the ‘mutica var. nigra’ story. That I now could refer to these as H. retusa ‘nigra’ should be seen as evidence of the flexibility and pragmatism that is really required to allow acceptance of a nature that we can understand.
6. JDV93/57 H. retusa. Plattekop. There are several populations in the ferricrete inselbergs. They are inclined to shiny yellow colours and rather longer, more erect leaves than one would expect in H. retusa. In fact one can see a drift from the latter to ‘longibracteata’ in the broader context that I originally used the name to really suggest populations that were neither ‘retusa’ nor ‘turgida’. This particular habitat is unusual in that the rock is larger than usually accepted by the plants and there is quite an infusion of the white clay of decomposed shale. The plants seemingly, and consequently, take on a rather powdery and milky appearance.
7. MBB7772 H. retusa. Platkop Annex. Where Skeiding is west of Heidelberg, Plattekop and Platkop Annex are east of Riversdale. These and the next two populations form an important extension of the distribution of the species eastwards and to perhaps emphasize a distinction between H. retusa as it grows on the inselbergs as opposed to H. retusa ‘turgida’ that is riverine.
8. MBB7765a H. retusa. Plaatjieskop. These plants are very similar to the two preceding. A similar population was shown to me by Japie Dekenah at Rooikoof to the north. What is significant here is the presence of hybrids with H. floribunda (fig. 74) and that in turn by the fact that H. retusa flowers in spring and H. floribunda in summer. Such hybrids are known at Cooper siding between H. pygmaea ‘argenteomaculosa’ and H. floribunda and also at Blackdown, N. Heidelberg between H. retusa ‘turgida’ and H. floribunda. Because I consider that there is a deep-seated ‘problem’ of interaction like this between major role players, I will discuss this again elsewhere.
7766H.retusaXfloribunda. Plaatjieskop JDV 0682Habitat H. retisa. Plaatjieskop JDV 0711
9. MBB7771 H. retusa. W Soetmelksrivier. There are seemingly always surprises in Africa (“always something new”) and this is one. Not only does H. retusa grow in direct association with H. mirabilis cf. ‘splendens’ but the habitat encroaches on the highly ferruginous rocks that form the hardest layer of the ferricrete inselbergs. It is really only two populations in the Kweekkkraal area where haworthias can be said to be truly in this kind of rock. ♦
If the name “H.enigma” applies to the plant (or plants) from east of Riversdale at Komserante, it is a name that I really do not advise to be taken seriously from a botanical point of view. It is useful at population level and to demonstrate the nature of classification difficulties but it is a minor problem in so far as those difficulties extend. The plants were first shown to me by J. Dekenah on the same day that he also showed me ”H. magnifica” in the Nature Reserve just south of Riversdale that is less than 3km away. My impression then was that it was the same element even if it did look a bit different. The plants are quite large (to 70mm diameter), fairly tubercled and often with lines in the upper retused area of the leaf face. While I originally classified “H. maraisii” under “H. magnifica”, I later separated them because it seemed so incongruous to include all the variants of the western “H. maraisii” with the few populations of “H. magnifica” then known. Also, as Essie Esterhuizen pointed out, “H atrofusca” as a variant of “H. magnifica”, seemed to be more dominant than had been realized. There were several other complications largely due to ignorance. Since my revision I have done so much more exploration and turned up so much new material that I have been forced to the conclusion that there is really one main element involved and that is H. mirabilis. This is where I believe the Komserante plants belong and the difference from the Nature Reserve population is due to a degree of infusion of H. retusa.
I revisited the site with Kobus Venter many years ago but did not look at a reported second population higher up the hill, taking it to be a little different based on plants I saw in Kobus’ collection. What was on my mind while we were recently exploring the area further east to examine the possible connection of H. mirabilis “magnifica” to “splendens” (and which we confirmed), was the fact I had never seen Kobus’ plants from Kruis Rivier northeast of Riversdale other than in Kobus’ collection. The plants I saw were also generally more robust than “H. magnifica” and more evenly tubercled. Kobus kindly took me to that Kruis River locality and much to my surprise the plants were in flower late October (see JDV92/65 Figs1). This is quite wrong for H. mirabilis, which is essentially a summer flowering species. I later went again to explore Komserante more thoroughly and to look at both the “magnifica” populations to which I believe the name “H. enigma” has been applied. The populations are in fact no more than 75m apart and cannot be considered to be genetically discrete at all (see MBB7778 Figs 2, and MBB7779 Figs 3). While it is true that the habitats are slightly different, this is reflected in the plants that at the upper slope of the hillside are vegetatively more robust and even clump forming, while those lower down in a bushier grassier habitat tend to be solitary and more withdrawn into the soil. These plants flower in summer and it is evident to me that there must have been some genetic exchange with H. retusa that grows approximately 200m away on the same hillside.
1. JDV92/65 H. mirabilis Kruisrivier.
2. MBB7778 H. mirabilis. Komserante.
3. MBB7779 H. mirabilis. Komserante.
Habitat preferences are very strict and it is curious to observe that there are four small buttresses forming the western slope of the low hillside. On the northern one we have the two H. mirabilis populations. On the second is H. retusa ’geraldii’ (see MBB7780 Figs 4) that is very clump forming, the third has no haworthias and on the fourth is a less clump forming H. retusa that is often referred to as ‘fouchei’ (see MBB7781 Figs 5) because the leaves are slightly longer and more upright than in ’geraldii’. This is not quite correct. Mr. Dekenah showed me what he took to be the true H. retusa ‘fouchei’, further south off the hillside and along the stream at Komserante. This habitat has since been cleared for farmland. The plants were very large and the leaves considerably more erect than is the case for the plants on the hillside. Curiously H. minima is present on the first and last buttresses. Flowering time for these H. retusa is spring but we did see vicarious flowering in summer so that definitely it is possible and probably a regular occurrence for out-of-season flowering to occur with the possibility of hybridization. I noted such hybrids between H. retusa and H. mirabilis in our exploration further east from Riversdale and I did think some of the clones in the Komserante plants evidenced this as well.
4. MBB7780 H. retusa. Komserante.
5. MBB7781 H. retusa cf. ‘foucherii’. Komserante.
The reality is that the Kruisriver and Komserante “magnifica” may flower at different times, but they are vegetatively very similar indeed. I consider that they belong in the same system as H. mirabilis and this conservative view of “species” as systems is the most sensible way to interpret and understand the plants. The approach can be taken further, as I tried to do in an earlier article concerning H. mutica ‘nigra’. In that article I dealt with a whole series of populations in and around Heidelberg, to show that there are three main elements viz. H. retusa (including H. turgida), H. mirabilis (including H. magnifica) and H. floribunda. I did not deal specifically with, nor resolve, the issue of H. mutica var nigra. I also noted populations along the lower Duiwenhoks river and suggested that H. floribunda is absorbed into H. mirabilis virtually south of the N2 east west highway, re-emerging on the northwestern corner of the Potberg in the south.
My most recent exploration was to check the flowering times of the original population of H. mutica var nigra at its origin at Kransriviermond and what I assumed to be the same thing on rather limited examination further north at Morning Star. Both these populations are along the Duiwenhoks river southeast of Heidelberg and in a continuum of four populations of H. retusa extending from just east of Heidelberg to still further east at an eastern Diepkloof locality (I think it has been referred to as Droekloof and Droerivier too, and the name ‘chromatica’ attached to the plants – see MBB7794 Figs 6). Less than 10km away is a population of H. retusa at Pienaarsriver (see MBB7776 figs in Chapter 3) that is very near to the whole array of populations of Kiewietskraal that I discussed in my writings about H. mutica var nigra. Two of those were distinctly H. retusa and so is the Pienaarsriver population that is different in that the plants are mostly quite dark in colour.
6. MBB7794 h. retusa ‘chromatica’. SE Heidelberg.
In those same writings I mentioned several populations of H. mirabilis along the Duiwenhoks river as well as populations of H. retusa var turgida. Here I am unashamedly relating “turgida” to “retusa” as I believe that when one calls all the populations now known to mind, as well as the variants they contain, this is the correct position of the plants in relation to a sensible and rational species concept. It is evident that H. floribunda is merged in H. mirabilis south of Heidelberg and the same thing occurs south of Swellendam. Thus I see no problem in now dealing with the population at Kransriviermond to which I applied the name H. mutica var nigra. I attached the varietal name to H. mutica because this is what G.G.Smith had done, and I believed that it was the link between H. retusa in the east and H. mutica in the west, when there were still huge gaps in the known distribution records for both species.
I have mentioned a population of ostensibly H. mutica var nigra at Morning Star, which is about 5km north of the Kransriviermond origin of the plants so named. This Morning Star population (see MBB7221 Figs 7) is very significant because while it includes virtually the same range of variants as the Kransriviermond plants, it flowers in summer as opposed to spring for Kransriviermond. Not only that, but it is 300meters away from a population of H. mirabilis (previously this would have been H. magnifica) that flowers and seeds at the same time. The habitat is marine plane ferricrete that overlies Bokkeveld shale but there are subtle differences between the two habitats.
7. MBB7221 H. retusa ‘nigra’. Morning Star.
The Kransriviermond plants (see MBB7804 Figs 8) are down in the Duiwenhoks valley in exposed Bokkeveld shale and I consider that the dark colour of the plants in the two populations is due to infusion of H. mirabilis. The rounded leaf tip would be derived from the “atrofuscoid” element of H. mirabilis while some plants also have the tuberculate roughness that that element often has.
8. MBB7804 H. retusa ‘nigra’. Kransriviermond.
Further south and west is the Slangrivier valley where there is also a suite of populations that includes smaller forms of H. mirabilis (H. heidelbergensis var. toonensis) as well a H. retusa var turgida also in an unusual form that supports the taxonomic position I now adopt.
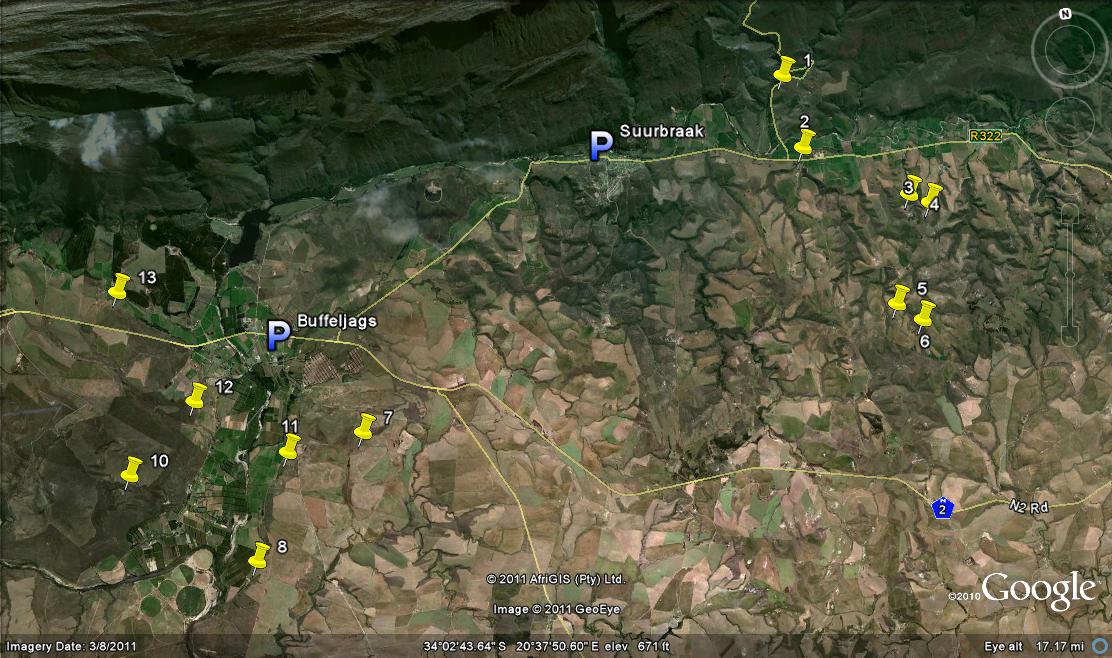
The conclusion is that the Morning Star and Kransriviermond ‘nigra’ can be regarded as H. retusa within the same broader concept that includes ‘turgida’. The problem now arises from still another new discovery. This is from along the Buffeljags River (see MBB7801 Figs 9) about 10km southeast of Swellendam and 12km from the nearest obvious H. retusa at Goedverwagting further east; and from the less obvious H. retusa (that I have elsewhere noted as H. mirabilis) south of the Tradouw Pass. The nearest H. mutica are within about 15km to the west at Napky and Luiperdskop. The plants are the now customary “variable”. The leaf tips are very rounded. Some plain, some lined, some with curious submersed pinkish dots. The surfaces can be quite smoothish or else the surface cells terminate in minute spines that give the surfaces a glistening appearance – evident in H. mutica too. These plants flower in summer whereas both H. mutica and H. retusa are spring flowering. Note needs to be taken of the habitat. It is an extensive and old river gravel embankment and terrace, and therefore another variant on the interface of recent ferricrete deposits and the older Bokkeveld shales. The vegetation is grassy Karooid bushveld, unlike the dry fynbos to the northwest or the renosterveld (mainly cultivated) to the east.
Kobus Venter saw the photographs and suggested that the plants might be H. mirabilis’ and reminded him of ‘atrofusca’, perhaps also by virtue of the flowering time. This is of course what my “enigma” is all about. It is compounded in this case because these Buffeljags ‘muticas’ are rather odd in that the leaf ends tend to be flattened almost as though H. floribunda has also entered the fray. Kobus was much happier with the H. mutica connection when he saw the plants in the field.
The story here closes with H. mutica MBB7741 from Dankbaar about 12km west of the Breede River bridge at Swellendam. The plants here also have very glistening surfaces and thus seem to confirm the very original perception of G.G. Smith’s that the Duiwenhoks plants and the plants west of the Breede River were connected. It is certain that he never envisaged the complexity of the interrelationships between three main elements viz. H. mirabilis, H. floribunda and H. retusa.
Footnote. It is just assumed that perhaps the name “H. enigma” has been used for the Komserante H. mirabilis, because that population is indeed enigmatic as this article explains. Gerhard Marx informs that “enigma” applies to a H. mirabilis population further west where the more atrofusca variants occur. Since writing this chapter yet another similar population to the Kruis River and Komserante populations has been found about 15km SE Riversdale. ♦
This chapter is based on recent field exploration and embroiders around many aspects of Haworthia species discussed in earlier chapters. What should be striking is that new populations follow the very predictable geographic pattern that all my earlier exploration has exposed and in my estimation confirm in every way what I consider a sound and satisfactory taxonomic solution and help explain its limitations.
1. Haworthia minima.
Two new populations of this widespread species are recorded:
MBB7807 Swartrug, SE Heidelberg. This is a steepish east facing slope. The plants are small, the leaf length barely exceeding 50mm and with a tendency to form clusters in the shallow clay banks that rift the area. There are many plants in quite a small area and it curious to see them within very large clumps of Euphorbia tridentata. This Euphorbia itself has a strange geographic distribution being known at Riversdale, Calitzdorp and then at Cradock in the Eastern Cape. (see Figs 1)
MBB7813 NW Skeiding, E Tradouw Pass. This is a high rainfall area and we were expecting to find H. mirabilis in the lower pressure bursts and white clay that can be seen from the farmhouse far away. But instead we came across H. minima at the highest point of the hill among sparse grass. They are rather green in colour and it is interesting to note in the one plant pictured that it has non-tuberculate upper leaf surfaces and white cartilaginous margins and keel of the leaves. (see Figs 2)
2. Haworthia rossouwii.
MBB7803b Morning Star, S Heidelberg. Morning Star was known to J. Dekenah but the area has been very developed since those years. It was very surprising to find this small form in abundance at the top of a high hill in ferricrete pebbles and virtually no vegetation ground cover. However on this subsequent visit we crossed through an internal farm fence and found the plants in abundance about 150m further away and over quite a large area. We later looked at a lower promising hillside and small area nearer the Morning Star homestead and found the plants again. (see Figs 3)
Looking from Morning Star to Kransrivermond 097
3. Haworthia mirabilis.
This item reports on 6 populations, three of which are new. They confirm the continuity down the Duiwenhoks River and also southwards and westwards from east of Riversdale through to west of Robertson and Greyton and Napier to the south of that. Attention is drawn to south of Heidelberg where the ‘enigma’tic relationship of this species with H. retusa is suspected.
MBB7808 Swartrug, SE Heidelberg. This farm is well known as the source of ‘Haworthia chromatica’ that I dismissed as an element in the complex surrounding the e H. retusa (mutica) ‘nigra’. The names Droekloof and Doringkloof have been used while the map reference gives Diepkloof for a large area extending to west of the Duiwenhoks River. The plants are on an east facing boulder terrace in a stony low bush site with some grass. The plants are small and show the expected wide range of variation. They are fairly similar to the plants in the lichen patch just east of Heidelberg that I have always mistakenly taken to be the origins of H. heidelbergensis, but here are subtle differences that mark a population difference, but I will come back to this. (see Figs 4)
Looking from Swartrug towards Riversdale.
MBB7809 Koeisekop, SE Heidelberg. This is on the same farm and a similar ferricrete hillside southwards. There are several hilltops by the same name in the general area and I cannot even speculate on the origins of the name (“cowshead”). The plants are larger than in the previous population and very abundant. The similarity to populations in the Swellendam and Riviersonderend area is in my opinion striking. Odd clones are miniature images of H.mirabilis from the southwestern populations west of Napier. (see Figs 5)
MBB7220 Morning Star, S Heidelberg. I have discussed this population before and I repeat it here because I did not have many field photographs. It is only 300m from the Morning Star MBB7221 H. retusa ‘nigra’’ population and they flower and seed virtually in unison. Therefore they probably also reflect on the strange interaction between the two major systems. What I have surmised is that H. floribunda gets absorbed into H. mirabilis at the southern limits of its distribution and in these pictures there is clear evidence of the rounded and flattened leaf tips of that species. Therefore we may actually here have a three way species interaction. Reinforced by the presence of three different things at Kransriviermond, covered later under 4. H.retusa ‘turgida’. (see Figs 6a and b)
MBB7221 Morning Star
MBB7811Kransriviermond, S Heidelberg. Populations further down the Duiwenhoks river have been reported on and this one fall within that mould and the same one as the previous Swartrug and Koeisekop populations. The population is smaller and the plants too are quite small. (see Figs 7)
MBB7814 Skeiding, W Heidelberg. This area is also reported on but digital photography makes it so much easier to record the enormous variability that such populations exhibit. Some of the plants are typically ‘atrofusca’ while others have leaves ended in an awn. Leaf surface may be smooth and there is even an indication of smooth leaves. What is notable is the absence of H. floribunda from what is the sort of habitat it can be associated with and can one can speculate its absorption into H. mirabilis with leaf shapes that demonstrate this. H. floribunda does appear again about 15km to the southwest in association with H. retusa and in the absence of H. mirabilis. (see Figs 8)
The Skeiding homestead
MBB7233 Die Plotte, Heidelberg. “H. heidelbergensis” correctly has its origin in this near vicinity and these small summer flowering elements are certainly part of a large assemblage of populations in the central area of what I now come to see as all H. mirabilis. Here discussion must pass to the following. (see Figs 9)
4. Haworthia retusa‘turgida’.
A difficulty that I experience is in the need to dismiss the impression and general idea we have that there are tiered ranks. H. retusa has many populations and past treatment has placed most of the variants in H. turgida. To suggest that these are really two separate systems is not realistic and confounds any explanation for a few anomalies that exist.
MBB7810 Tierkloof, SE Heidelberg. Tierkloof is quite a deep valley that pushes out of the Duiwenhoks to the east. We found this huge population of large clump forming plants and several such populations are known just north of Heidelberg itself. I have applied the epithet ‘longebracteata’ to these and this is no more satisfactory than generating names for each of a very wide range of populations. There are populations along the Duiwenhoks that are far more inclined to the ‘turgida’ element and Essie Esterhuizen wrote a very good account of the host of variable populations that he encountered. (see Figs 10)
H. retusa ‘turgida’ at Tierkloof, Swartrug
MBB7812 Kransriviermond, S Heidelberg. We only saw three plants at this locality and these plant mirror the preceding, if a bit smaller. The problem here is that we also have the element I myself described and named as H. retusa ‘mutica’ and I originally linked this “variety” with H.. mutica following what G.G. Smith had done and for predictive reasons. The fact that ‘turgida’ now occurs with it is a huge hurdle to cross. Think carefully. (see Figs 11)
MBB7804 Kransriviermond.I have written about his population in the chapter “Haworthia enigma”. This explains that the plants are not all leaden-black as perceived. They are spring flowering – as ‘turgida’ is. However, the range of forms and colour virtually replicates the plants in MBB7721 at Morning Star (pictures in the “enigma” chapter) where they are spring flowering. Across the river from both these populations is a weak population of small plants of H. mirabilis (7811 above) summer flowering. We know that this major difference in flowering season is not a total barrier to hybridization. (see Figs 12)
MBB7234 Die Plotte, Heidelberg. This is a population of large clump forming plants on a north slope about 300m south of MBB7233 H. mirabilis ‘heidelbergensis’. It is spring flowering. When I first saw these plants I dismissed (actually I never dismiss anything – I maintain reservations as I do now) these plants as large variants of ‘heidelbergensis’, and actually collected one magnificent specimen that looked to me just like H. mirabilis, say, from west Napier. Looking at the range of variation one is compelled to concede that here again is an infusion of H. mirabilis into a population that is primarily H. retusa in that vast shadowland it casts with ‘turgida’. (see Figs 13)
Conservation I add these comments for several reasons surrounding the poor image that collectors have in the eyes of conservationists, and the very rickety fence I therefore occupy. There is a note reprinted in a recent issue of the newsletter To the Point. This taken from Times magazine and seems to be a rather hyped up condemnation of widespread destruction based on a view of internet websites. These are also monitored by RSA law enforcement that is rightfully disturbed by the high number of locality and collector citations that are indicative of illegal collecting. I personally no longer apply for a “collecting” permit for the prime reason that I am concentrating on photographic record, as a herbarium record no longer serves any better practical short term purpose. The herbaria are overburdened and I prefer living plants to dead ones. After 70 years of field activity I feel treated like a pariah and reduced to subservience under less-informed inexperienced people operating under a rigid and oppressive legislation aimed at acquiring a prosecution rather than wisely trying to direct activity in a positive direction. This activity of collection is, and I have no doubt about this either, a human right imbedded in the very reason for creation and our place in it. Conservation has its roots in the passion that people have for nature and it is nurtured by experience, contact and knowledge of its varied components. It is just ridiculous that one can go to any animal or plant centre here in South Africa and see plants and animals from anywhere else in the world and none indigenous. It is Ok to, say, keep Madagascan Chameleons in captivity where it is someone else’ responsibility to secure them. But keeping local ones is taboo? It is OK to freely sell cacti that do not occur here but are now being dug out in tons in (if that is really true). Recently it has become illegal to so much as possess Encephalartos latifrons and even the National Botanic gardens can no longer propagate and sell this plant. While we talk “plant indigenous” on all public fronts, the fact is that the acquisition, cultivation and sale of indigenous plants is viewed in the most negative light – a viewpoint encouraged by conservators.
This is control by law, a large degree of pure ignorance and by improper application of power derived from legislation. This is not the way to go. It is quite evident from the Times magazine article that legislation has the negative effect of creating a false value and glamour to discovery, Latin names, acquisition and possession. I believe that knowledge and understanding is true power, not legislation and enforcement.
The collecting numbers, localities and information provided here is to promote understanding and knowledge of plants, and thereby to promote their continued existence against all the vagaries of nature that “threaten” them, least of it which may be actually human greed.
Acknowledgement I am glad to acknowledge the company, comments and interest of Gerhard Marx and whose organization sceptisism I also value. Landowners always impress me with their generosity and graciousness in allowing us access. On this occasion they include Mr. and Mrs. Nelie and Wimpie Jacobs of Diepkloof, Mr. Cloete and Belinda Fourie of Melkboom (B&B), Mr. and Mrs. Coetzee and Sarita Uys of Morning Star, Messrs. Christo and Pieter van Deventer of Doornvlei, Mr. and Mrs. Neels and Anne-Lise Uys of Skeiding (B&B) and then Ms. Jennifer Steggie of Heidelberg. In view of the comments on conservation noted above, I note that there are several farms that now offer accommodation and an outdoors experience and the opportunity to view these plants in their extraordinary natural surroundings. ♦
My experience with Haworthia dates back to my childhood and on to nearly 70 years of observation. However, my interest was only able to properly manifest when I began work at the Karoo Botanic Garden in 1969 and it has since been through many phases. I wrote a formal taxonomic revision of the genus in 1999 and have spent a good bit of the last nine years adding to and verifying what I wrote. Haworthia has always been regarded as a problem child of botany to be avoided by professional taxonomists for various reasons including an apparent phobia of the many amateur collectors peering over the shoulder while at work. This has puzzled me because it seemed to me that if the need for good classification and identification was so strong there was an obligation on botany to provide the service. So my involvement has been largely by default. I was trained in an agricultural and entomological tradition with a totally different and unsophisticated approach to things like taxonomy, systematics and nomenclature. In the infant science that agriculture then was in South Africa, I can barely claim that my MSc is much more than an indication that I tried to learn something beyond normal schooling. While trained as an agricultural entomologist, my leaning was to plants and I eventually came to the Karoo garden to do what I liked best viz. exploring plants. Unfortunately the route is via identification and names and so I have walked a long road through the minefield that this is. Was this only in respect of Haworthia? No! This is a persistent misconception. Haworthia is only different because it has attracted such close and sustained amateur interest by so many for so long. I experienced failing classification in many other genera. To be fair I think the real reason is the lack of importance attached to the whole function of plant classification. It even seems as if many modern botanists pursue the study of plant relationship under the guise of systematics that is not committed to providing formal names and identifications.
The problem in the genus Haworthia begins with weak botany from the moment Linnaeus decided that there were four small plants that belonged together as one species in the genus Aloe. This alone generated a nomenclatural problem the dust of which is still to settle. A worse and obscured fact is that because of their small flowers these plants were and are still treated as one genus – one set. This is not true. The allied genera of the Alooideae of the Asphodelaceae are not comfortably classified and the three sets of plants that constitute the genus Haworthia are not only florally distinctive, they are also phytogeographically and behaviorally different. Thus while there was or is this huge flaw at this classification level, it is actually bizarre that sense can ever be thought to have reigned at the species level. The proliferation and confusion of names coupled with the associated argumentation about names has even led me to doubt my sanity, or even lose it as my critics will happily claim. My impression now as a non-botanist is that taxonomic botany somehow lost sight of the requirement that a biological classification should mirror and reflect the evolutionary processes or phylogeny that underlies our view of species and their origins. Worse, is the actual absence of a clear and unsullied definition of what a species is and hence what a Latin binomial as a supposedly formal scientific term of reference actually means. Cladistic methodology in plant taxonomy is in my opinion nothing more than a belated and even misguided attempt to arrive at or return to some semblance of scientific truth. Molecular biology is now being desperately seen as a holy grail that will summarily correct reluctantly conceded failures of the past.
There is no doubt that the mess in the classification of Haworthia is the product of non-botanists, but it is absolutely incorrect to exculpate the contributory role of taxonomic botany and professional botanists who are or were taxonomists. My interest has by no means been confined to Haworthia and the need for names has largely been driven by a perpetual curiosity about many different plants in the field environment. I am, by the route my life has taken me, unusually and almost uniquely experienced in the relation between botany, botanists and interest of the layman in plants, as it is reflected in the names we use for them and how we respectively do so. Consequently I think that I am now qualified to express some opinions on what Haworthia can teach us quite apart from what the hostilities that wage across the whole taxonomic front suggest.
I pointed out rather timorously that in Oxalis, morphological characters were not as indicative of species as revisions and identification keys generally seem to imply. The dramatic changes in our classification systems that the results of molecular biology now seem to threaten, is a manifestation of this simple fact rather than any great new understanding of what really goes on in the plant kingdom. My fear is that molecular biology is likely to fudge most of its promise because the basic tenets of classification, the mistakes of the past and the reality of the field are still not properly appreciated. This is what I think Haworthia has to tell us. I made use of Oxalis to test my observations that we can look at Haworthia as composed of elements that reflect the very nature of the habitats and environments where they are found. I did this because Haworthia plants simply do not have any array of characters by which species can be arrived at whereas Oxalis has a plethora of such apparent (taxonomically bewitching, tempting, and misleading) means. While the few facts that there are in respect of differentiating Haworthia into three groups (subgenera as presently constituted) has been lost and confounded in the broader classification of the Alooideae there is not much else to help verbally to define the species. It is not to say that modern technology has not been resorted to, to help clarify the issue. Both cladistic and molecular studies have been done but the results flawed by weak hypothesis and perhaps just weak science too.
How then do I think sense can be made of Haworthia? It is firstly by defining the species as dynamic fractal systems that illustrate the responses of plants to skeletal habitats and geological substrate in the wider context of landscape and topography across the total distribution area. Furthermore, there is an obvious correlation to be made with vegetation and other biological systems both plant and animal. I cannot pretend to know a great deal about any of these things, but also suggest that nothing more is needed other than awareness that this is what biological diversity and species are essentially about anyway.
It has taken me nearly 40 years to arrive at the big picture of Haworthia. It is as a non-taxonomist that I say categorically that the three main elements in the present genus need to be seen and registered as “different”. Then we can focus on the subgenus Haworthia where the greater difficulty lies and which I believe offers the greatest insight into what plant species are and what the difficulties are in recognizing and defining them. Also into what Latin binomials should or might mean.
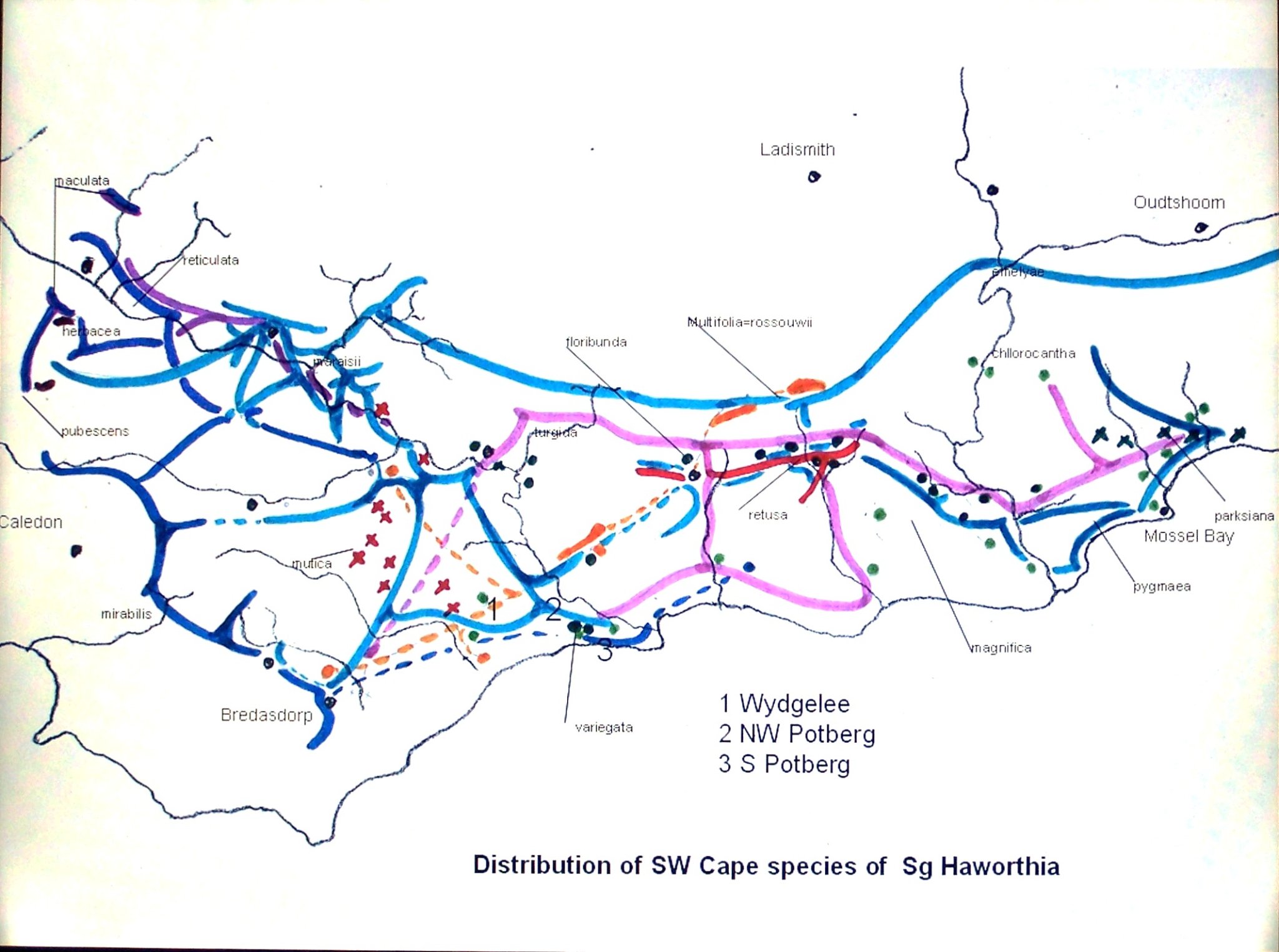
For this I am going to take just two species systems and ignore some of the peripheral facts that might confuse the issue rather than illustrate the profound nature of species interactions. I take H. retusa (Figures 1 to 4) and H. mirabilis (Figures 5 to 10). In order to do this I have to bypass even my own conservative classification by considering H. turgida and its many variants to be the riverine and cliff dwelling version of H. retusa. H. mirabilis is more complex and I have to include three other species viz H. maraisii, H. magnifica and H. heidelbergensis for which spatial factors and major geological substrate are the prime differentiators. Both my two selected species are highly variable both within and between populations. It is in fact the intra-population variability that I underestimated. Growing plants from field collected seed can result in a hundred or more individual plants that are all different in appearance.
The two species share the same distribution range, but with an extension of H. mirabilis into the Worcester/Robertson Karoo, while H. retusa extends further east from Albertinia to Great Brak. H. mirabilis flowers in late summer and H. retusa flowers in spring. They never grow in direct association! The populations of each are highly localized and when in proximity are usually separated by hundreds of meters, and difficult to ascribe to habitat conditions given the ranges of habitat that respective populations may occupy. I have already noted and warned of the reality that there is evidence of interaction between these two clearly defined systems. In addition, somehow or other, the species H. pygmaea emerges as discrete in the eastern Mossel bay area, while H. mutica sits in an island between Swellendam and Riviersonderend in the west.
What is most striking is the variability within each system and this is where the crunch comes and what my pictures should illustrate. They are but a small sample of many hundreds and I have adopted an informal method of captioning the pictures as the formalities of nomenclature would need to be followed to depart as I have from the existing status. Not only are my pictures supposed to illustrate the problem of variability within species, figures 2 and 3 show variability within a population and figures 3 and 10 show two very similar plants from two quite different species systems. Note that the plant in figure 3 is in flower and that the similarity to figure 10 is rather exaggerated by photograph scale as in H. mirabilis ‘toonensis’ the plants are smaller.
Taken individually from the morphological extremes between and even often within populations, it will stretch the credibility of the most ardent taxonomic lumper to claim that any two of these are representatives of the same species. It is an unpalatable truth that things that look different are the same just as the converse is that things that look the same are different. When this reality is properly conceded by the professional taxonomist, perhaps we will succeed in demystifying and deglamorising the Latin binomial. Botanists who are not taxonomists will arrive at a better understanding of their plant world and consequently we who look to them for understanding will be better served. We need to be more realistic about plant description and identification and also correspondingly more conscious of the real nature of diversity. Conservation is not to be seen as maintaining a list of names for which there is a use. It is to be seen as maintaining the options for change and the conservation of a far greater diversity than any list of names can hold. It is botanists (quoted in Levin, 2000) who wrote “our system of names appears to achieve a reality which it does not in fact actually possess”. I think this implies that even the Latin binomials we so modestly, reluctantly or even portentously use do not convey what they should; but rather the product of our individual limited experience, knowledge and perception. My feeling is that species do have a reality and for Latin names to have any sense, they need to be more circumspectly applied. There is little merit in having a highly regulatory nomenclatural code and little value in what the names actually refer to. ♦
In Haworthia Update 4 I wrote an essay about the haworthias east of Albertinia in which I discussed their relation to H. retusa and H. mirabilis, while generally lumping them largely in H. pygmaea. There are of course real ‘turgida’ populations as far east as near Mossel Bay, so I argued the case for an interplay of the two former species that over the whole distribution range generated two ‘species’ in the east viz. H. pygmaea and H. retusa (to include ‘turgida’), and three ‘species’ in the west adding H. mutica to H. mirabilis and H. retusa.
Recently I had the pleasure of meeting Gregory Nicholson who is studying botany at University, Cape Town. He surprised me by telling me that there was a Haworthia on his parent’s property west of Herbertsdale. It was not in fact so much surprising as confirmation of the belief I formed on a trip a short while before that there must be haworthias in the very suitable terrain of the Jakkals River valley 6km west of Herberstdale. The surprise came when Greg indicated the position of the plants much deeper into the mountains.
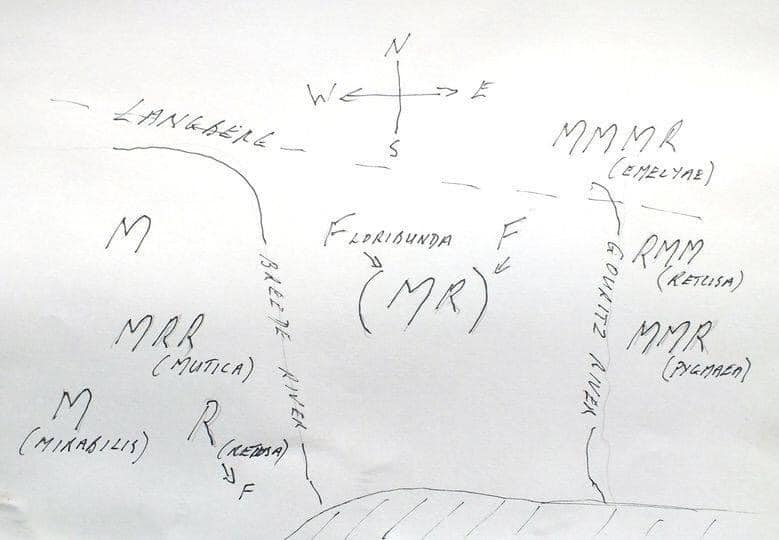
We visited the site while exploring H. emelyae north of the Langeberg Mountains and I will report that in an additional short article. We over-nighted east of Barrydale and a puncture made us detour via Riversdale rather than take the northern dirt road. It was raining heavily at Albertinia, but we could see sunshine on the Langeberg in the direction of Herbertsdale. We traveled the dirt road through the upper Gouritz, because we also wanted to gauge the condition of the gorge through the mountain, as Greg reports the presence of H. retusa ‘turgida’ some distance in. Towerlands is some 8km on and it was still just sunny when we arrived. We walked immediately the several 100m to Greg’s site along a good path among true Fynbos vegetation and found the plants very easily in a ferruginous sandstone ridge sloping into the river. Then the rain came and made picture taking a bit difficult (see MBB7489 figs. 1-16).
The plants are quite small and there was not much clustering. Unfortunately the weather restricted us so we could not look for better habitat around the corner where Greg had actually pointed. Greg’s photographs had made me lean to H. pygmaea and indeed I do not think one can deny the reality of this if one also considers the odd plants at Herbertsdale itself (JDV97/135) that I assign to H. pygmaea. However, the similarity to ‘turgida’ is just as striking and Daphne and I concluded that we could recognize similarities to ‘dekenahii’ and even ‘mutica’. To make it even more confusing, I felt, and still do, that the plants had similarities to H. emelyae. This latter was confirmed in my mind when we continued our trip via Cloetes Pass back to the Little Karoo to just west of the Gouritz gorge and a population of H. emelyae known to us from Gerhard Marx. I will submit that in a subsequent short note.
I include a habitat scene (fig. 1 – the arrow marks the locality) and note that looking south east one can see Albertinia and Aasvoelberg in the distance. We know that ‘turgida’ is present in the sandstones of the Langeberg, but this find suggests it is far more abundant and widespread than present records suggest. We need to now what the plants in the upper Gouritz look like.
Acknowledgements Gerhard Marx has been extremely helpful and informative and I am most grateful to Gregory Nicholson for his contribution. ♦
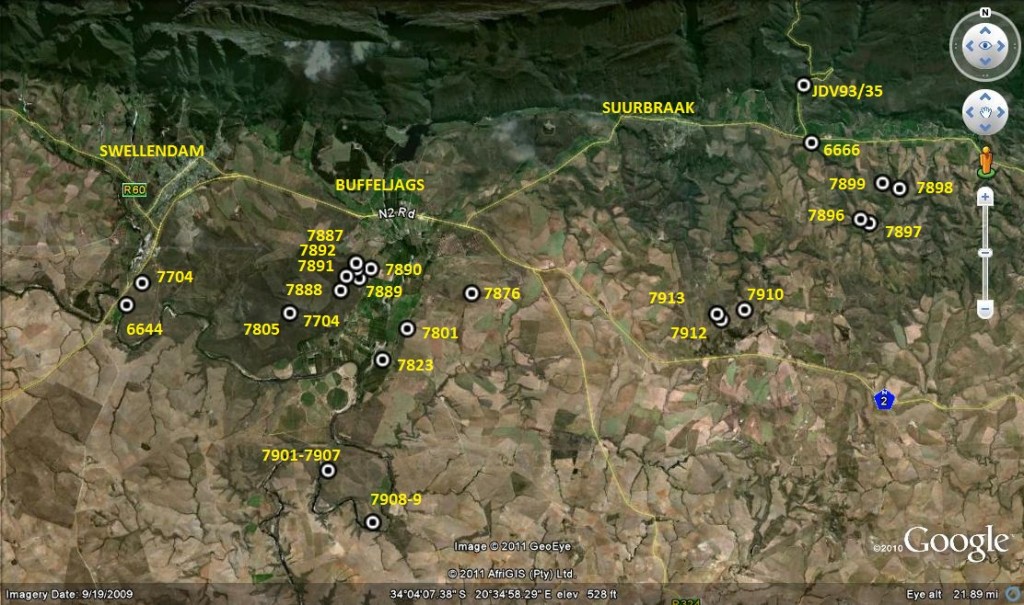
I wonder. I have written so many words purporting to be my last that my credibility here too must be under stress. Two very recent articles of mine in Alsterworthia deal essentially with that issue, although they also cover the discovery of Haworthia mutica (Buffeljags) (= H. groenewaldii Breuer). They do not cover my subsequent thoughts on actually reading the description of this new “species” by Breuer, Marx and Groenewald. I hope that the present manuscript will explain why I reject this as a Latin binomial although anyone who is in the least familiar with my writing should already know. Spurred on by that discovery, I instigated a search in another area of the Buffeljags valley adjoining the Bontebok Park accompanied by Jannie Groenewald who informed me of what he had found in still another area I had long wanted to explore. So I instigated another search there too and again with Jannie. A discussion of these new finds is submitted to Cactus and SucculentJournal where I trust it will be published. The essence is already in Alsterworthia and this article is written to widen the readership, submit more pictures and maintain continuity with the 6 volumes of HaworthiaUpdate that Harry Mays has been so conscientiously and determinedly publishing. This is all writing that may not otherwise have seen the light of day. I am personally extremely grateful for that as I have had a mania since writing my revision Haworthia Revisited and Update Vol. 1 (both Umdaus), to set the record straight and explore all the unknowns, or at least some of them. So on with Haworthia retusa ‘nigra’.
The second area opened to me by meeting Jannie, was a farm southeast of Tradouw Pass, Heuningklip. In the Tradouw Pass, H. retusa ’turgida’ is found on the steep cliffs at the southern end. The road has been rebuilt since I first saw the plants and the population is greatly reduced from what it was. I simply do not have a picture handy to show what those plants are like, but that is not the need here and it does appear in Haworthia Revisited. Immediately south of the pass is another population of plants that puzzled me from the moment I saw them until now. In Haworthia Revisited there is a picture of one clone under J.D. Kobus Venter’s number 93/35. It is named as H. magnifica var. magnifica. The plants are present in a tension zone between Shale and River gravel, and vegetationally between what was Karoo Valley Bushveld and grassy Renosterveld. In April 1997 I collected seed and grew countless seedlings from there, which were all different. Only one picture, as MBB6666) is included in my long treatise on what I discussed under the title of “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.That picture is also under the name H. magnifica var. magnifica but it was not intended to stay that way. It must be noted that my use of names in the Update volumes was not in a decision-making process so much as a learning and informative one. I read my conclusion to the treatise and note that I made no formal name changes. It is only as further field exploration yielded more and more information that I came to realize that H. magnifica and H. mirabilis, classification-wise, are inseparable. The Tradouw collection still worried me because of the close proximity of H. retusa ‘turgida’. These plants at Tradouw and all the things I referred to as H. mutica ‘nigra’ are in fact better understood as variants of H. retusa in the complexity of its relation to H. mirabilis.
This article thus covers exploration of an area between Swellendam and Heidelberg that I have neglected for too long. It is particularly applied to the problems of classification and naming that have persistently clouded my life and my relations with other writers. But as importantly to me is the unclouding of the way taxonomy and classification is useful and necessary for those interested at all in plants and how it is viewed by them.
Consideration and use of formal names Here I need to explain the naming style I now adopt. I have dropped the use of any rank below that of the species name. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable herbarium specimen that is used to anchor the Latin name. The scientific ranks are aggregates and the terms variety and forma are so too. There is simply too much variation between populations and within populations to enable the certain identification of almost any plant that the binomial system suggests. There are no clear entities and ranks and this article should demonstrate that. There is a separate code of nomenclature that covers cultivars. Note that I do not drop names. I only drop the suggestion of status that additional names have, be they single clones or population references for all the different plants within them. I have used names in Haworthia for 70 years and intensively for the last 40 years and I am very familiar with the difficulties of any system. For the field and all the populations and plants I have seen, I can do no better than offer the list of names I updated in February 2009 in Haworthia Update 5.2:192. This, and the set of Updates, comprises a near total overview of the genus and effectively a revision. The attempted revisions by others fall short for me because they do not cover the range of populations that I have seen nor what history tells me, although they may include populations not seen by me. That is not strange because the area to explore is still vast. Another problem with even trying to be formal, is that which G.G. Smith encountered (Mrs. L. Bolus did too) where several people were generating new names at the same time and trying to keep track formally with concurrent name changes would not have been, is not, possible. The point I should make is that in the last ten years, my field work has confirmed the predictive nature of my classification and these latest finds are in extraordinary agreement with that kind of expectation. What I have found essentially agrees with what I would expect. This is dramatically so for H. mirabilis based on a find by J. Jelimicky at Sandhoogte midway geographically and visually between H. mirabilis ‘bobii’ Hayashi (my nom. nuda ‘pilosa’, ’velcro’, Steven Hammer’s ‘Shaggy dog’) along the Breede river, and a collection of mine further south in the DeHoop reserve, MBB7886 H. mirabilis, discussed (p11) and illustrated (p33) in Haworthia Update 5.2.
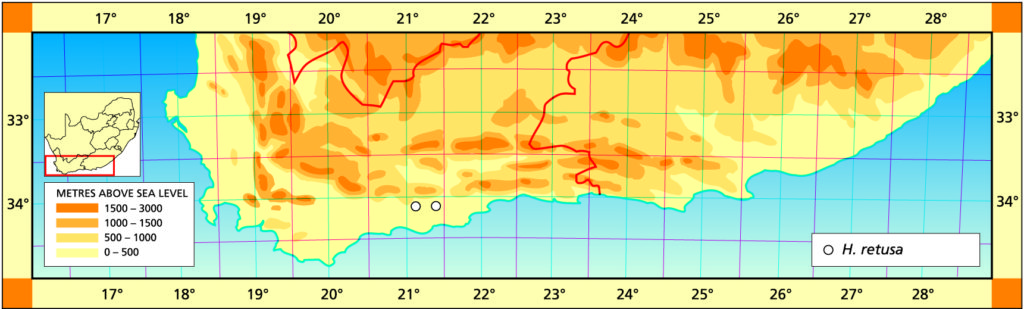
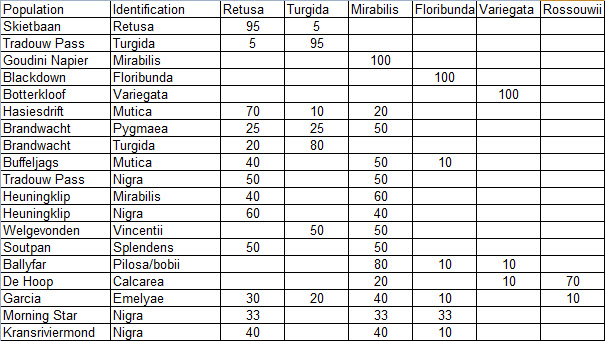
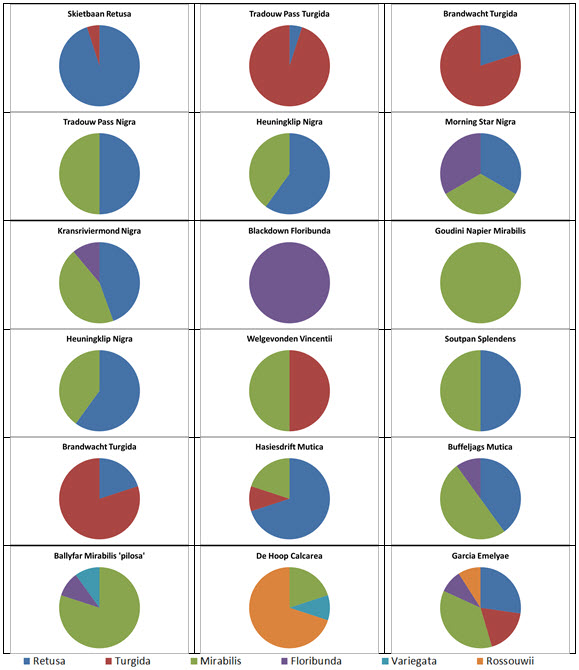
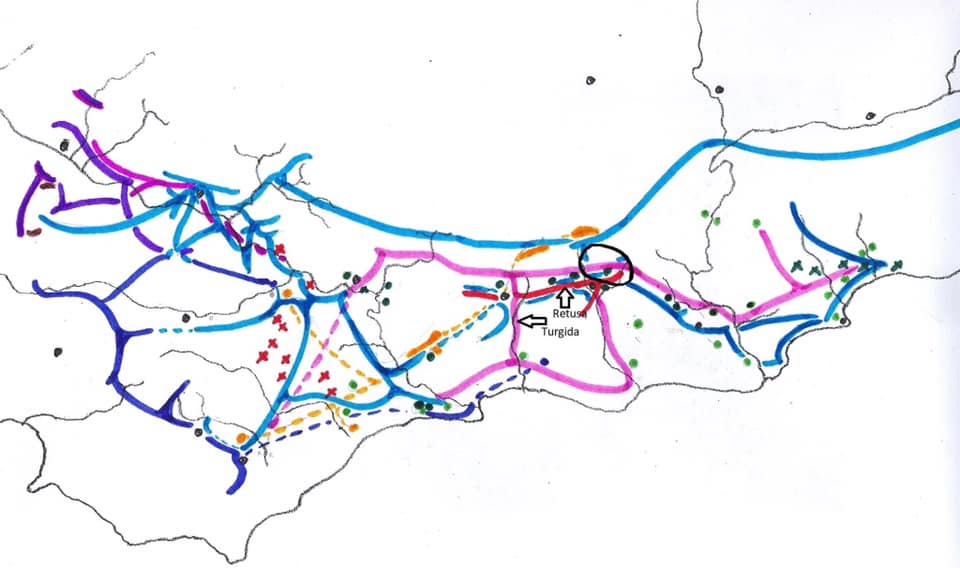
Prior to extending exploration in the Buffeljags area, I had also been down to the Haworthia populations in the lower Breede River area. This is where H. mirabilis seems to assume a wide range of forms and H. retusa ‘turgida’ and H. variegata’ are also present. I also, in the company of Kobus Venter and Lawrence Loucka, revisited Kransriviermond and Morning Star south of Heidelberg where H. floribunda is in the mix with H. retusa ‘turgida’ and H. mirabilis (in my broad new sense that includes H. maraisii,H. magnifica and H. heidelbergensis). Thus many of those images were very fresh in my mind when I went to Heuningklip southeast of Tradouw. Looking at all the plants and in editing my pictures I was truly impressed with the reality of continuity. The essence of H. retusa and H. mirabilis is in all those plants. A mobius strip is a strip of paper twisted and joined at the ends to produce a shape that only has one surface. What we have in the Southern Cape, and covered or touched on in this article, are interlinking mobius strips with one main surface as well as loose ends that present identifiable end-points. I tried once to illustrate the continuities between populations with single or multiple lines joining the different populations to express degrees of similarity. I can also visualize a pie-chart in which each population is represented by a circular chart with the contribution of main elements drawn as slices proportional to the strength of contribution. Table 1 is a schematic and very crudely suggested enumeration for such a set of charts. I have put the elements retusa and turgida as separate under an H.retusa heading for no other reason that I want to emphasize turgida as a variant at the two geographic extremes. One is eastwards where H.retusa ‘retusa’ is absent (Brandwacht, Mossel Bay) and one westward where it is present (in mutica, Hasiesdrift, Bredasdorp). H. retusa ‘turgida’ is present both east and west. But there are many populations contributing to the H. retusa pie in all the ranges of the two parts, from 0-50% thus making up the continuity from retusa to turgida.
Consideration and characters.
a. The pie chart Measuring the similarity of any two plants is extraordinarily difficult. Doing this for a population is even more so. Doing so for many populations, including those unseen in order to fulfill the predictive requirement of a proper hypothesis, is next to impossible. Hence all the disputation and name abuse. In table 1, I speculate an arrangement to illustrate the problem. The pie is constructed from the figures in the chart for just that set of genetic characters that are directly relevant to the set of plants in question. The pie is incredibly big and it should be understood that there must surely be pie-parts that spread their influence beyond the restrictions of the slices I conceive. So my table is restricted to the few species that are immediately relevant in respect of geographic association and direct interaction (by possible cross-pollination?). A pie-chart is drawn for each individual population. In the chart I assume that there is a set of genetic information that programs for H. retusa ‘retusa’. I have also suggested a different set for H. retusa ‘turgida’. If one considers only these two elements and all the known populations, my suggestion is that there is a set of charts that moves smoothly from one set to the next with no interruption. There is no discontinuity and the sets average 50:50 for the information. Thus it is one species and H. turgida is noted as H. retusa ‘turgida’.
Table 1. A schematic distribution of similarity
Table 2 is the complete set of pie-charts that should convey the possible relationships of the different populations and from which can deduce and understand to what degree H. retusa and H. mirabilis can be related. Also it will indicate the similarities between, and the difficulties of, trying to explain H. pygmaea in the east and H. mutica in the west. Both arise from the same genetic source. The problem is compounded by the infiltration of H. floribunda in the area midway between Heidelberg and Swellendam with the re-emergence of H. floribunda in the south at Potberg. H, variegata also enters the picture but from the south and also in the Potberg area. H. emelyae is the very probable combined entity entering the little Karoo. H. rossouwii is almost independent except in the very south-west at Bredasdorp and it may possibly link to other elements such as H. herbacea and H. reticulata.
Table 2. Pie Charts
At Heidelberg there is interaction between an element that is a 50/50 retusa/turgida with a variant of H. mirabilis cf ‘heidelbergensis’. The latter name suggests that H. mirabilis is the general name for all the plants of that species, and ‘cf’ is an abbreviation for ‘compare with’ ‘heidelbergensis’ to refer to the small plants in populations previously regarded as independent species.
b. What really constitutes the pie The pie is the totality of the genetic material, the DNA that determines all life forms. It is the genes, the materials of characters and inheritance, and the repository of all genetic information. The pie is very large indeed and it is said that the DNA pie of an elephant only differs by 3% from that of a mouse; that of a man by 1% from a chimpanzee. In Haworthia one species from another ‘species’, I could guess at 0.001% or less regardless of whose concept I follow. This difference does also not reside in one consolidated slice that can be reduced to a single small pie as I have done. In DNA analysis, certain segments of DNA from cellular chloroplasts, mitochondria or the chromosomes themselves are isolated and the sequence of the paired amino-acid is analyzed. It requires a formidable statistical process to arrive at a two-dimensional tree (phyllogram) that illustrates a branching relationship with a measure of branch separation and lengths. This can perhaps also be used to generate pie-charts with an innumerable number of slices. I personally consider the phyllogram generated in the DNA sequencing process grossly inadequate.
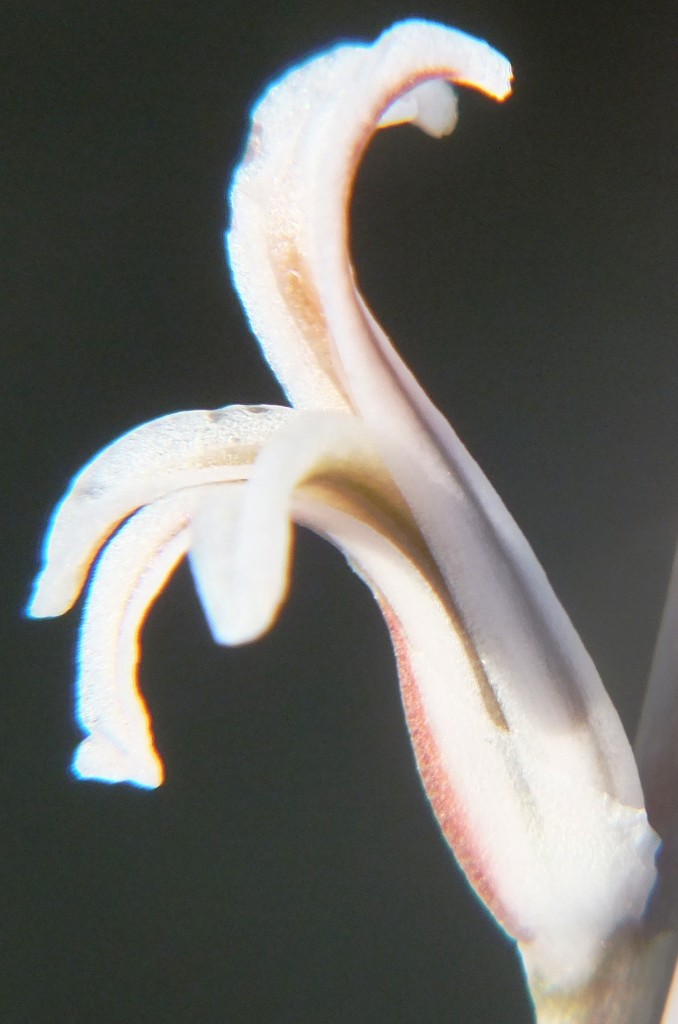
c.) The characters This is extraordinarily difficult to specify no matter how obviously the plants seem to differ from one another. Number, size, shape, colour, arrangement, surfaces, spination and attitude of the leaves in the spiral rosette constitute the main things that determine how we see the plants. These are influenced by growing medium and growing conditions so these are complicating factors. Flowers are used in the same way as the leaves but far less so because they are so similar in all the species. Flowers are also not always present either which reduces their usage. Time of flowering is a major factor because it is linked to the reproductive process. It is also difficult to capture any firm picture of the flower because of the aging process in the 3-5 day life of the individual flower.
d.) The relevance here It simply is not necessary to parade the obviousness of this and the way in which will be attempted anyway in the discussion below. It is flowering time that may be critical beyond, or linked, to the facts of spatial geographic relationships of populations. It is traditionally flower structure that has formed the basis of classification of plants because of the reproductive significance and the role of reproduction in distribution of genetic material between plants and populations. Pollination of Haworthia seems to be primarily by a Solitary bee species. These bees make individual nests and are not communal like the Honey bee. They make tunnels in the soil as nests during the summer months. I have observed them as adults in the winter but then they are less numerous and less active. Few Haworthia flower at the height of winter and do so in two main time frames viz. spring and late summer. The implications are that breeding between isolated populations apart from any other consideration of compatibility, is a question of distance and time. A nest bound Honey bee can forage to 13km from its nest. A free flying Solitary bee can be speculated to cover a considerably greater distance. All these things play on the frequency and power of inheritance. Genetic drift is acknowledged as the genetic movement within a population away from the norm for a species because of isolation, while genetic flow is perhaps the holding together of similarity over geographic distance by a travelling pollinator. Thus the statistical probabilities of breeding across distance are impossible to derive.
I cannot quantify any of these things nor suggest any greater knowledge but this suggests to me, given the freedom with which the various species hybridize in cultivation and even in the field, we need to consider geographic relationships. In my extensive experience with other genera, it appears to me that a key consideration is if things grow together and maintain their difference, or not. Haworthia species are invariably in small isolated populations and seldom do different species grow intermixed. They are also influenced by geology of the substrate and particularly so because they favour the lowered competition from more vigorous and larger plants on the rocky sites or shallow soils they favour. That can be stated in the converse, Haworthia are favoured by conditions poor for other plants.
e.) Flowering time and growing together. The existence as discrete species can be judged from the way in which populations share geographic space. I discussed this in some detail in Update 2.1:76. If two sets of plants grow close together and flower at the same time, while still maintaining obvious group differences, then they are most probably different species. If they flower at different times while still maintaining difference then the question of being different species is taken as certain. If the groups are separated and flower at different times while the differences are small and apparently within the experienced range of variation observed between populations, then it may be problematic. One would need to resort to the extended distribution and variability of the groups in question. Generally in Haworthia, species belonging in the same sub-genus do not share habitat and this is indicative of fairly rapid absorption of introduced genetic material (by pollinators). Hand-pollination and the ease with which plants of different species hybridize in cultivation suggest that there is practically no breeding isolation between them. In the following discussion, use is thus made of general distribution of species and proximity of populations in assessing to which species they best belong. It should be borne in mind that variability may be clinal. This means that the plants change along a geographic gradient. The direction of the gradient may also not be of increasing distance i.e. linear. It may also be circular so that ends of a gradient may meet again as two populations that appear wholly different, but linked by an extended set of other populations. A much more complex situation may actually exist where these clinal gradients may be multidirectional and thus involve more than one apparent species. My observation is that populations do differ and differences follow a pattern that becomes predictable i.e. gradual genetic difference over geographic area. This is not just a result of environmental gradient, but also linked to the question of genetic drift (the tendency of isolated populations) to move away from the general norm. It also results from the genetic flow between populations where the change of genetic material between populations at the distribution extremes is only through other populations.
f.) The species definition This is a subject I have covered many times. My view is that species are dynamic chaotic systems oscillating between different character states with time and with different rates of mutational change. Closely related species may not be recognizable on the basis of single character states and the spatial geographical distributions and local juxtapositions of groups of plants may be the only way to understand what the situation is and so make sensible decisions.
g.) Habitat geology I contend that species richness in South Africa is largely due to its varied geology, skeletal soils and comparatively mild climate. This present discussion deals with species that are associated with a geological phenomenon that is the Tertiary deposits of the Southern Cape. I am no geologist and am required to make the pretention I am one to explain why plants are distributed the way they are and what influences difference among them. In this case the vegetation at Buffeljags, Swellendam has been described (by Breuer et al. cited below) as on silcrete, and the general vegetation as by vegetation scientists as Ruens Silcrete Renosterveld. But there are two problems. The first is that vegetation classification is not infallible and I have never found it really useful in dealing with a wide range of species that occupy skeletal habitats. Secondly the substrate geology is definitely not simply silcrete nor is the clay soil either simply bentonite or kaolinite. There are many problems in the classification of clays. Bentonite in common terms seems to be the yellowish clay mined in large quantity from Heidelberg eastwards and there is a transition to ochre bearing clay. Bentonite may be aluminium, potassium, sodium or calcium dominated. As a montmorillomite clay it should have expansive cracking properties. Kaolinite is popularly regarded as the very white clay that is commonly seen under the rocky top layer of the sharp inselbergs that rise above the wheatfields west of Heidelberg. It seems to be less mineral dominated clay and does not have the expansive/contractive properties of bentonite. The distinction between ferricrete and silcrete seems to be that in ferricrete, iron oxides are infused into the petrified water deposited layers. Both the kaolinitic and bentonitic clays seem to be derived from the weathering of Bokkeveld shale that underlies the silcerte/ferricrete layer. It does not require any stretch of the imagination to see now why any one inselberg may be a little different from the next, and sometimes a lot different. What we do not know is how rapidly the landscape has moved from a broad white silcrete/ferricrete underwater plain to the remnant condition of that rocky plain today. Neither do we know what it has meant for the plant species that are linked to, and dependent on, the habitats generated in the erosion processes.
Map of Swellendam to Suurbraak area.
Map Legend – east of Swellendam.
1. MBB7899 H. retusa ‘nigra’. Heuningklip. 2. MBB6666 H. retusa ‘nigra’ ↔ H. mirabilis. S Tradouw Pass. 3. MBB7896 H. retusa ‘nigra’. Heuningklip. 4. MBB7897 H. retusa ‘nigra’. Heuningklip. 5. MBB7898 H. retusa ‘nigra’. Heuningklip. 6. MBB7888 H. mutica. Rotterdam. 7. MBB7889 H. mutica. Rotterdam. 8. MBB7890 H. mutica. Rotterdam. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags. 10. MBB7741 H. mutica. Dankbaar. 11. MBB7892 H. marginata. Rotterdam. 12. MBB7891 H. minima. Rotterdam. 13. MBB6644 H. mirabilis. SW Swellendam. 14. MBB7704 H. mirabilis. Bontebok Park. 15. MBB7805 H. mirabilis. Bontebok Park. 16. MBB7823 H. mirabilis. Klipbult. 17. MBB7887 H. mirabilis. Rotterdam. 18. MBB7876 H. mirabilis. Disselfontein. 19. MBB7901 H. mirabilis. Crodini E. 20. MBB7902 H. mirabilis. Crodini E. 21. MBB7903 H. mirabilis. Crodini E. 22. MBB7904 H. mirabilis. Crodini E. 23. MBB7905 H. mirabilis. Crodini E. 24. MBB7906 H. mirabilis. Crodini E. 25. MBB7907 H. mirabilis. Crodini E. 26. MBB7908 H. mirabilis. Sandkraal W. 27. MBB7909 H. mirabilis. Sandkraal W. 28. MBB7910 H. floribunda. Rietkuil. 29. MBB7912 H. mirabilis. Rietkuil. 30. MBB7913 H. mirabilis. Rietkuil. 31. JDV93/35 H. retusa ‘turgda’. Tradouw Pass.
The populations
Set 1 – THE TRADOUW PASS AREA (H. retusa ‘nigra’) This first section deals with populations that complement those in the long discussion under the title “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.There I ended with the words … ”The habitat photo (of H. mutica Hasiesdrift) raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.” Added were these words … ”There the saga for the moment must rest.” What I had in mind was exploration that no one seems very anxious to do. This has now been done and I would lay the matter to rest as follows:
1. MBB7899. H. mirabilis, Heuningklip. See figs 1-30 Fig. 1 is a scenic view of the area to show the countryside and the type of inselberg the plants are often found on. Fig. 2 is a closer picture of the erodible clay held in place by grass clumps at the base of which the plants often are. Occasionally they are simply out in the open sun. I term these places pressure bursts because water build-up in the soil causes collapse of the small banks and erosion can be quite fast. Several times I have found plants lying in collapsed soil. Such a soil bank near Heidelberg has retreated by nearly 2 meters in about 15 years albeit accelerated by road maintenance, but it does relate to any attempt to explain how much suitable habitat may have disappeared with time.
Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – bFig. 2. A photograph of the habitat of MBB7899.
Figs 3 to 30 is a selection of plants at the site where one is almost compelled to examine each plant separately and easily generate another 28 practically different images. Fig. 3 is of a plant with the surfaces roughened by small tubercles. SEM (scanning electron microscope) images of similar leaf surface often show incipient tiny spines on the tubercles and they appear as spines or hairs in populations like H. mirabilis ‘pilosa’ at Ballyfar, or H. mirabilis at Windsor Riversdale. This surface roughness and all its degrees is fairly characteristic of H. mirabilis but is does occur in H. retusa too. Plants illustrated in Figs 4 and 16 has one muttering Kruisrivier and Komserante, northeast and east of Riversdale respectively, and which I would have allotted to H. magnifica in the early period of my exploration no doubt have Breuer and Hayashi names. Now I simply say H. mirabilis and add the locality to place the plant to at least give it a geographic reality that is better than using the formal Breuer/Hayashi in hanging commas as I also suggest. Fig. 6 is apparently a seedling and demonstrates the leaf-shape of H. floribunda that led me to suggest that H. floribunda is a perpetuated juvenile form. Of course the evidence suggests to me that H. floribunda coalesces with H. mirabilis in this wider area although it does appear at Swellendam to the west of my map as the large form and at the extreme SE corner of the same map as a convincingly normal form. Fig. 9 demonstrates longer pointed leaves and Fig. 12 short more obtuse leaves. Fig. 17 is of a plant that has relatively smooth leaf surfaces rather like H. retusa and the leaves are conspicuously lined. In Fig. 20 there is a suggestion of muticate (without a point) leaves. Fig. 22 is again quite strongly lined. Fig. 22 shows a leaf with a single line. Fig. 25 demonstrates what is termed rotate leaves where the points of the leaves point around as though to encircle the plant. Fig. 28 made me recall plants of H. mirabilis at Melkhoutrivier in the south west near Malgas and further to Ballyfar. The flowering time is late summer.
Fig. 3. MBB7896 H. retusa ‘nigra’. Heuningklip.Fig. 4. MBB7897 H. retusa ‘nigra’. Heuningklip.Fig. 5. MBB7898 H. retusa ‘nigra’. Heuningklip.Fig. 6. MBB7888 H. mutica. Rotterdam.Fig. 7. MBB7889 H. mutica. Rotterdam.Fig. 8. MBB7890 H. mutica. Rotterdam.Fig. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags.Fig. 10. MBB7741 H. mutica. Dankbaar.Fig. 11. MBB7892 H. marginata. Rotterdam.Fig. 12. MBB7891 H. minima. Rotterdam.Fig. 13. 13. MBB6644 H. mirabilis. SW Swellendam.Fig. 14. MBB7704 H. mirabilis. Bontebok Park.Fig. 15. MBB7805 H. mirabilis. Bontebok Park.Fig. 16. MBB7823 H. mirabilis. Klipbult.Fig. 17. MBB7887 H. mirabilis. Rotterdam.Fig. 18. MBB7876 H. mirabilis. Disselfontein.Fig. 19. MBB7901 H. mirabilis. Crodini E.Fig. 20. MBB7902 H. mirabilis. Crodini E.Fig. 21. MBB7903 H. mirabilis. Crodini E.Fig. 22. MBB7904 H. mirabilis. Crodini E.Fig. 23. MBB7905 H. mirabilis. Crodini E.Fig. 24. MBB7906 H. mirabilis. Crodini E.Fig. 25. MBB7907 H. mirabilis. Crodini E.Fig. 26. MBB7908 H. mirabilis. Sandkraal W.Fig. 27. MBB7909 H. mirabilis. Sandkraal W.Fig. 28. MBB7910 H. floribunda. Rietkuil.Fig. 29. MBB7912 H. mirabilis. Rietkuil.Fig. 30. MBB7913 H. mirabilis. Rietkuil.
These images can now be compared with MBB6666 at Tradouw Pass to establish if my assessment is reasonable. My contention is that this MBB7899 population is midway between the other three Heuningklip populations I am about to illustrate and Tradouw Pass. This similarity is repeated widely and eventually contributes to my contention that H. mutica has been generated by drift away from its two contributing parent species viz. H. mirabilis and H. retusa.
2. MBB6666 H. mirabilis. Tradouw Pass See figs 31-71. This population is at the mouth of the Tradouw Pass and relatively low-lying in respect of the previous population. The habitat is quite different being partially coarse river gravels, weathered shale and at one point is an un-weathered shale ridge where the plants are wedged in the cracks. The habitat is of course very different to the steep sandstone cliffs within the pass itself where H. retusa ‘turgida’ occurs. I initially thought these plants fitted with H. magnifica. A decision derived from a very small sample available at the time. Now I looked for the images and photographs in my records under H. mirabilis where they should have been since my inclusion there of ‘magnifica’, and also under H. retusa where I found them in the folder ‘nigra’. The flowering time is late summer. In this respect similar to the previous set MBB7899 H. mirabilis but the plants are more robust. Figs 31-45 were taken in 2009; figs 46-53 in 2011 and figs 54-71 are from seedlings grown by myself. I tried to reduce the pictures and eliminated a few. But I really think they all need to be presented to show again how serious the difficulty of circumscribing them as a single recognizable key-able entity really is. While looking at my pictures I seriously thought that the set taken in 2011 differed from those taken in 2009. The cultivated plants of course are not directly connected to their source and the variation is self-evidently wide. But it is equally so in field plants. There are the smoother surfaces of H. retusa and some very rough surfaces for H. mirabilis. My decision became that they fitted best with the ‘nigra’ populations where my organizing had taken them despite flowering time, but here MBB7899 shows absolute geographic position between H. mirabilis and MBB6666. I consider this key evidence for the basic continuity for H. retusa and H. mirabilis that I have conjectured elsewhere and I would not be able to defend an argument that MBB7889 plants belong better in H. mirabilis and this means MBB6666 belongs there too. I would simply dispute that there is no better solution so stick with history as far as possible and that means H. mirabilis. If Breuer or Hayashi have a name for a variant from this population intended as an aggregate name, I recommend the use in inverted commas. I would actually like the use of the equilibrium diacritic of a double arrow pointing between two names viz. H. mirabilis↔H. retusa ‘nigra’, although this available symbol is not quite the thing.
Fig. 31 6666 H. retusa ‘nigra’
I come to this vexing question of “typical” that is bandied about even by myself when I liken population to population. There is the same range of variables in MBB6666 noted for MBB7899, but the plants do not darken as much and they are larger. So we need now to look at the other three Heuningklip populations asking what significance is attached to their presence in exactly the same habitat as MBB7899, but geographically separated and by perhaps 500m and 2km respectively. The plants are so variable that it is not possible to arrive at a characterization that separates any two plants, or any two populations in any geographic set for that matter.
3. MBB7896 H. retusa ‘nigra’, Heuningklip 1 See figs 72-89. The plants in this and the next two populations were flowering in September as does H. retusa and they are relatively large, green in colour and generally smooth. Of course both vary enormously due to both intrinsic genetic difference and extrinsic local situation. One plant within an older decaying grass tuft was nearly 200mm diameter, and a healthy green colour, another plant in the open white clay was less than 50mm diam. and dark in colouration.
Fig. 72 7896 H. retusa ‘nigra’
4. MBB7897 H. retusa ‘nigra’, Heuningklip 2 See figs 90-105. The first two pictures show the general landscape and the local habitat where the plants occur. The clay is highly erodible and often there are no plants at all in the clay. Some of the plants have very acuminate pointed leaves while in others they may be relatively obtuse and rounded.
Fig. 90 The countryside looking towards the Tradouw Pass.Fig. 91 The ‘white clay pressure burst’
5. MBB7898 H. retusa ‘nigra’, Heuningklip 3 See figs 106 -132. The great variability is again evident and in this population the plants were observed to be generally without much venation or lines as in fig.112. Then one plant, fig. 119, was so striped as to stand out from all the populations seen.
Fig. 106 Silcrete hilltopsFig. 107 7898 H. retusa ‘nigra’.
Set 2 – THE BUFFELJAGS AREA (H. mutica) This second section follows a recent manuscript submitted to Alsterworthia yet to be published entitled “Some last closing thoughts” written in the hope of finding the closure I seek and that will not come. That article is about a population that has been named as a species viz H. groenewaldii, in an article authored by Breuer, Marx and Groenewald (Alsterworthia 11.2:15) that I only saw after the submission of my manuscript. I also only saw the article after the exploration I would have asked for to verify opinions. The description seems to have been prompted by Marx who refers to similarities to H. springbokvlakensis and to H. magnifia var. atrofusca and raises many issues that my manuscript does not cover. I simply find the whole discussion unrealistic and especially so when no further exploration was done to examine anything. One should refer to Breuer’s World of Haworthias Vol 1 to see that the naive promises in the foreword and introduction of exploration and field study, was never achieved. It is true that the plants have an unusual surface texture and it is also true that the habitat is quite unlike any other H. mutica occupies. But it is Marx’s comment that disturbs me viz. “Unfortunately Bruce Bayer keeps on refusing to see it as something new and calls it simply H. mutica ‘Buffeljags”. This is a distortion and misrepresentation, and whose misfortune this is, not specified. My reticence is reflected in the preceding discussion and Gerhard does not seem to follow, or agree with, what I have been writing about and trying to explain in this general context for so many years. In fact in an unpublished (to my knowledge) manuscript he dismisses all my writing as “pretentious” that no doubt it is, because I have never fancied my self as a taxonomist in the tradition of taxonomists. I was as fascinated as Marx by the plants and am even more anxious than he is to see just how they connect to the probable geographic counterparts in the context of the species definition that I apply. I was also very curious to know what the connection to H. mutica and the H. retusa ‘nigra’ issue, only now laid to rest in the preceding discussion, actually might be here. This is why I use the word “instigated” in reference to exploration in the very valley where H. mutica ‘groenewaldii’ occurs and to the obvious habitat for similar plants. It is surely unfair iniquitous to so demand that I see this population as “something new” in an article that rather disregards the history of the subject and what I have been trying to understand of it.
I also recently submitted a manuscript to Cactus and Succulent Journal (US) where I wrote of “H. groenewaldii Breuer”… “I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February/March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time”. I have in many places discussed the problem of “hybridization” and it must surely be well-known from my writing that H. floribunda merges into H. mirabilis. Less well-known will be the fact that evidence for H. variegata doing the same is also available. In respect of floral characters, I wrote a chapter in Haworthia Update Vol.3.2:88 explaining the problems of using floral morphology for species recognition. I would challenge Breuer et al. to produce comparative results from studies of the flowers in any of these problematic species to demonstrate difference. Regarding the curious shiny leaf surface, I refer to Haworthia Update Vol.2.2:141, Chapter 15 where I discuss electron scanning photographs of leaf surfaces. This also demonstrates how problematic leaf surface is. It would be tiresome indeed to parade all the evidence in support of these contentions of mine. In Haworthia Update 5.2:108 I also briefly discussed H. mutica from Buffeljags (‘groenewaldii’) and submitted a large number of photographs explaining the habitat and what the possible connection was to the H. retusa ‘nigra’ conundrum. I may also have written about the issue in Haworthiad. Suffice to say that the authors of H. mutica ‘groenewaldii have certainly not checked or considered “all the facts” as the principal author claims. The very opposite seems to be the case.
Considering my oft repeated observation that geological substrate plays a major role in shaping species appearances, the Breuer, Marx and Groenewald description of the habitat for H. mutica ‘groenewaldii’ is a little misleading (see considerations above). Nowhere else is H. mutica on identical substrate and this would just support my contention of ecotypic differentiation that does not justify a Latin binomial. There is considerable doubt about the actual difference between these river boulder situations and the widely distributed terrace gravels elsewhere in the area, notably the Bontebok Park near where the following populations were found:
6. MBB7888 H. mutica, Rotterdam 1 See figs 133-169.
Fig. 133 7888 H. mutica.
7. MBB7889 H. mutica, Rotterdam 2 See figs 170-188.
Fig. 170 7889 H. mutica.Fig. 185
8. MBB7890 H. mutica, Rotterdam 3 See figs 189-191 The three populations above are separated spatially by several hundred meters and they are all on remnant river gravels on a more gently sloping bank than the Buffeljags east bank. The vegetation is closer to conventional fynbos as in the adjoining Bontebok Park, but grassy. The pictures are self-explanatory and the leaf surfaces of the plants are also generally as for the Buffeljags population. What is significant is that I cannot say I saw a single plant in these populations that resembles the one chosen and pictured as typical for this highly dubious species, H. groenewaldii. It makes more sense to me to identify plants from these populations by collecting number than to generate a supposedly descriptive Latin epithet from an imagined typical representative.
Fig. 189 7890 H. mutica.
9. MBB7801 H. mutica ‘groenewaldii’, Mullershof, Buffeljags See figs 192-215 The following set of photographs has surely seen the light of day somewhere and I repeat them because as far as I am concerned they are H. mutica with the shiny surface claimed for “groenewaldii’. Dankbaar is some 16km west of these Buffeljags populations and the substrate is Table Mountain Sandstone. The flowering time is just normal for H. mutica. Fig. 215 is a photograph of a plant apparently collected by Jannie Groenewald and it is quite uncharacteristic of the population and would suggest that it is a product of hybridization with H. mirabilis in the vicinity (e.g. Klipbult).
Fig. 192 7801 H. mutica ‘groenewaldii’.Fig. 215 7801 H. mutica ‘groenewaldii’.
10. MBB7741 H. mutica, DankbaarSee figs 216-235. My observations in respect of these two sets of plants confirms my widely written convictions about Haworthia taxonomy and the state of classification in the absence of a definition for the concept of “species”, and the intrinsic value of Latin binomials for rational and sensible communication. Of course this Dankbaar population is not identical to other H. mutica populations and I am not driven by fixation on any one character that defines this species. It is a geographic set and the various sets are not simply separable. But I am not finished yet.
Fig. 216 7741 H. mutica.
Set 3 – THE BUFFELJAGS AREA (Robustipedunculares) The western Buffeljags area also produced a population of H. marginata and one of a very curious H. minima, that both complement occurrences in the Bontebok Park a little further west. The conventional small, blue-green H. minima is present in the park as well as an unusually green population.
11. MBB7892 H. marginata, Rotterdam See figs 236-243. This population was quite exceptional and one of the strongest populations known to me. But the plants were in flower in September whereas the flowering time for the species is normally late summer; and the flowers and seed capsules exceptionally large. I do not consider that these factors command another Latin binomial.
12. MBB7891 H. minima, Rotterdam See figs 244-252 and Bontebok Park figs 253-4. These were also extraordinary plants, larger than those seen in the Bontebok Park population where the plants were also green viz. figs 253-4. Fig. 252 confuses the issue because, while the other plants seemed alike to H. marginata and distant hybrids, this one plant appears to be a recent hybrid. This occurrence cannot be overlooked in respect of other introgressive (or seemingly introgressive) populations of Robustipedunculares species. The plants were not in flower and again we have a suggestion or direct evidence of hybridization across a difference of seasonal flowering.
Fig. 244 7891 H. minima.Fig. 254 sn H. minima, Bontebok Park
Set 4 – THE GREATER BUFFELJAGS AREA (H. mirabilis) This is a set of 15 populations of H. mirabilis that occur east, west, south north of the H. mutica ‘groenewaldi’ related populations. I exclude from the discussion and illustrations the several populations of H. floribunda ‘major’ populations to the northwest, while I repeat the statement that H. floribunda contributes to the general gene pool of H. mirabilis and together with H. retusa, they all contribute to the gene pool of H. mutica ‘groenewaldii’. The huge presence of populations more readily assignable to H. mirabilis (sensu lato – in the broadest sense) is used by me to validate my opinion that the “mutica” elements draw most genetic resource from the H. retusa element, and hence why I refer those plants to H. mutica.
The sequence of populations 7901-7908 is located in a horseshoe of the Breede River, southwest of Buffeljags in very promising habitat. Only H. mirabilis was seen. I perhaps need to point out that these populations would have been extraordinarily difficult to name under any previous dispensation of mine where I was still trying to uphold H. magnifica,H. maraisii and H. heidelbergensis as species. These I have since merged in H. mirabilis and I reject the introduction of a group name like “aggregate” as Breuer seems to have done, as a complete fabrication that does nothing to solve the problem other than to highlight it.
The following 6 populations are all associated with tertiary and river gravels. There are differences between most of the populations but of an order that I cannot articulate. It is also not possible to really encapsulate the appearances of the plants in the two photographs for each population that I submit. At the same time it does not seem to me possible to make any communicable distinctions.
13. MBB6644 = 6860 H. mirabilis, SSW Swellendam. See figs 255-256.
14. MBB7704 H. mirabilis, Bontebok ParkSee figs 257-258.
15. MBB7805 H. mirabilis, Bontebok Park See figs 259-260.
16. MBB7823 H. mirabilis, KlipbultSee figs 261-262.
On selecting these two pictures I was reminded of my observation that there was a significant H. floribunda appearance to the plants. I had believed that I had seen H. floribunda in the Bontebok Park nearby many years ago but subsequent exploration has only produced H. mirabilis albeit with some similarities to H. floribunda. My reaction was to suspect that H. mutica ‘groenwaldii’ most probably also then needed to be considered as a product influenced by H. floribunda.
17. MBB7887 H. mirabilis, RotterdamSee figs 263-264.
18. MBB7876 H. mirabilis, DisselfonteinSee figs 265-266. These plants were a little more robust than the others in this set and rather similar to MBB7901.
The following populations are almost entirely in Bokkeveld shale but on a very close interface with tertiary gravels. In some places there was also some wind-blown sand.
19. MBB7901 H. mirabilis, Crodini 344, S Swellendam See figs 267-278.
20. MBB7908 H. mirabilis, Sandkraal, S SwellendamSee figs 279-282.
Set 5 – BACK TO TRADOUW PASS – AS IT HAPPENED In the process of compiling this manuscript I was bothered by the ever present question of what is still out there that might influence the outcome. My wife, Daphne, was planning an outing to see chameleons at Swellendam while simultaneously contacting landowners for me around the Potberg. Always anxious to get two birds with one stone, I asked her to find out who owned the land west of Heuningklip that we had just explored with Jannie Groenewald. The result was that we ended up spending a day with Odette Curtis of Custodians of Rare and Endangered Wildlife, south of Suurbraak and finding three populations that, for me, put the proverbial cherry on the pie.
All three were on the slopes of silcrete inselbergs and almost the last of this habitat westward from Heidelberg to Buffeljags. The first inselberg was different to the second and from the Heuningklip habitats because it was dominated by Aloe arborescens and A. ferox. Aloe brevifolia was also present in an unusually small form. The habitat was very impacted by grazing animals and there is an outside possibility that H. retusa ‘nigra’ may once have been present although I personally doubt it. To our surprise we found…
21 MBB7910 H. floribunda, Rietkuil, Suurbraak See figs 283-288.
There were many plants but severely trampled and disturbed. Their survival was obviously aided by the fact that much of the plant is below soil-level. But my experience of this species is that it struggles to maintain its identity in the presence of either H. mirabilis or H. retusa. They never share immediate habitat. There was no hybrid present and this also adds to the problem of interpreting flowering time and distance apart from other species as part of the process. The plants were not the large green variant of the species present to the west at Appelbos, Swellendam; and the leaves were both with and without marginal spines. Eastwards I know the species to be close only at Blackdown north of Heidelberg and at Goedverwagting south of the N highway about 15km east southeast. It definitely seems to be absorbed into H. mirabilis west and north respectively of those two populations. So while this is a slight contradictory element of surprise, it is the next two that really excited me. These were again inselberg habitat but without the strong silcrete upper layer of the former habitat and without the Aloe element. The second ‘population’ was about 200m from the first and comprised four groups of plants along a longer north-facing ridge.
22. MBB7912 H. mirabilis, Rietkuil, Suurbraak See figs 289-306.
23. MBB7913 H. mirabilis, Rietkuil, SuurbraakSee figs 307-331 I am very hard-pressed to say much more than that these plants met all my expectation for the very variable set of plants and populations that I ascribe to the one single species H. mirabilis. Who could possibly articulate the differences of the many populations of this species from each other and from this one? Variable becomes a done-to-death word. The individual plants, in two populations quite unlike any others I have seen, were generally robust with large specimens up to 100mm. diam. The colour is dark, although green when shaded within or under grass. The leaves were usually sharply pointed but there was always evidence of one or two leaves or odd individuals in which the leaves were muticate i.e. without points. In some plants the leaves were notably lined in others not. The colour also varied from chestnut to deep reddish-brown when the plants were more exposed. The surface texture was generally tubercled but there was evidence of smoothness of the leaves in many plants. Flowering time was obviously late summer gauged by the absence of flower at the time of the visit viz. September. Except for one oddity. This was a single plant with a weak flower spike. I photographed this for the useful information the flower-bud provides. Where Breuer et al. claim to have considered all the facts, this is a character that is conveniently overlooked when claims are made for the significance of floral characters when paraded to substantiate yet another claim to species significant difference. The bud-tip in all the Southern Cape species of subgenus Haworthia with the exception of perhaps H. rossouwii have this very obvious “fish-tail” bud tip. The tips of the upper two petals are flattened and spreading to resemble a fish’s tail. It is most marked in H. herbacea and H. reticulata where the bud tip curves downward from the semi-erect position of the body of the flower and then up again. I am fearful that my inability and reluctance to consider these plants as “new” may be the motive for someone to rush forward and apply a Latin binomial.
My opinion is that these two populations mirror the paradox in the east where H. pygmaea ‘fusca’ is nearer to H. retusa ‘retusa’ andwhere H. retusa ‘turgida’ is nearby. H. mirabilis ‘splendens’, in the doubt about its affinities, in my opinion remains in the context of H. mirabilis. This is a conclusion I reached after exploration of the area between Riversdale and Albertinia where the same elements noted in this article repeat themselves. I do not for one moment concede that better explanation is available by resort to a multitude of Latin binomials and by an artifact like “aggregate”.
Conclusion I have been deeply disappointed that so little progress on the classification front has been made among the fraternity writing about Haworthia. In communication with Lawrence Loucka, he wrote …”You might consider saying that a genus revision is needed and that until amateurs depart no proper botanist will make the investment. Until then chaos will rule, names will proliferate.” This is a useful and insightful comment and generally summarizes what I have often said. Unfortunately the position has been rather exacerbated when proper botanists have made their presence known and I can recount many instances to support this statement. My own view has been that botanists have not been able to confront genera that are so publicly and popularly known, and garlanded with names. Thus amateurs, including myself despite at least an iota of formal training in science, simply have a field day generating names like rabbits out of a hat.
Unless some proper thought is given to this whole question of what the names are supposed to mean apart from catalogue items, taxonomy (classification) will never lift out of the disinterest, disrepute and scorn that collectors justifiably seem to have for it. Latin binomials do have a real value and to demean this by one-dimensional derivation and rationalization is intellectually fraudulent. My writing is replete with examples of the variability and interaction of populations over large area. Taking a single population and one flower, and often a single plant, to generate a species description and a Latin binomial is grossly simplistic.
The one change that this article generates is the confirmation that the population MBB6666 from immediately south of Tradouw Pass is better identified by name as an H. mirabilis variant, and not as H. retusa ‘nigra’. MBB7801 at Buffeljags and now also at Rotterdam, remains as H. mutica and the epithet generated by enthusiasts can be added to make the trinomial H. mutica ‘groenewaldii’. For formality, the name for the form illustrated as typical would be H. mutica ‘Groenewaldii’. A few additional names will be needed for similar clones and a lot more for the cultivars that will be derived from this population set.
My conclusions here make no substantial departure from anything that I have observed or written over 50 years. While the subject has been comprehensively covered in the field and in writing, there is a huge amount of exploration that still needs to be done if the genus is to be still better known. My prediction is that the answer will conform very closely to the species-list I finalized in 2009 (see Alsterworthia 5.1:192). The disclaimer is that botanical classification may eventually reach the point where the real nature of species and their purpose comes to be known. In that case what we now may see and persuade ourselves are discrete species, are not. Instead these highly complex chaotic systems may in fact be quantum units of consciousness and whole elements such as some genera and sub-genera we now recognize will be the species.
Acknowledgement I want particularly to acknowledge Jannie Groenewald whose interest and enthusiasm are boundless. Then Allan Jeptha, an environmental law practitioner with great vision for the Suurbraak community; Odette Curtiss, a consultant for the conservation of Renosterveld vegetation; Lawrence Loucka whose patient guidance and comments I value and who generated the set of pie charts; Kobus Venter and Steven Hammer who have always been supportive and contributory; Max Coetzee of Felix Unite and Samie Lategan of Buffalo Breeding Programme for access to Sandkraal West and Crodini East; Jaap Viljoen for his contribution; my wife Daphne, who trusts me and whom I deeply trust. ♦
The objective was to explore some likely habitats previously observed at Van Reenens Crest and nearby. We extended the scope to include further exploration for Haworthia mutica as I am still questioning the place of this species in the greater scheme of things. Thus here are four sets of populations that I report on viz. H. mirabilis, H. retusa ‘nigra’, H. floribunda and H. mutica. See maps Figs 1 and 2 for geographical position.
Van Reenens CrestFig. 1 Van Reenens Crest. M = H. mirabilis, RN = H. retusa ‘nigra’, F. = H . floribunda.Fig. 2 Known distribution of H. mutica
H. mirabilis
Set 1. MBB7955 Van Reenens Crest – mid west. (figs 3-11)
The plants are moderate in size to about 50mm diam. and growing on an erosional slope below tertiary gravels where there is white clay. They vary from densely tubercled to relatively smooth.
Fig. 3 7955 Van Reenens Crest
Set 2. MBB7956 Van Reenens Crest – west 7955. (figs 12-34)
Despite apparently favorable habitat these plants were about 300m west of the previous set. Variation is again apparent in respect of presence or absence of surface roughness. Generally there are plants in which some leaves are without a point (see Fig 1) while others are acute tipped (mucronate – with a short hard point) or even have a long end-awn. Margins may be smooth or very lightly spined. This seems to be more apparent in young plants.
Fig. 12 View looking east to Set 1.Fig. 13 7956 Van Reenens Crest
Set 3. MBB7957 Van Reenens Crest – west 7956. (figs 35-41)
Again the plants are very variable with dense surface tubercles to virtually none. The leaf shapes vary and in fig.39 that appears to be a group of seedlings there is a remarkable series of different leaf ends and margins. In some plants there is distinct spination of the leaf margin.
Fig. 35 View looking south to Tradouw PassFig. 36 7957 Van Reenens Crest
Set 4. MBB7958 Van Reenens Crest – south west 7957. (figs 42-51)
Fig. 43 shows two rosettes of a plant with a “crocodile-skin” surface. The surface tubercles seem to be flattened and the leaves are mucronate. The following photograph, fig.44, is of a plant with acuminate leaves ending in long end-awns, and the surfaces are relatively smooth – retusoid? Unlike the previous records for the area that are north-facing, this is a south-facing “pressure-burst” of white clay. A nearby habitat of vertical shale that might have been considered favorable for H. mirabilis elsewhere was devoid of plants.
Fig. 42 7958 Van Reenens Crest
Set 5. MBB7959 Van Reenens Crest – west homestead 7956. (figs 52-64)
Figs 53-55 are smallish plants with rounded leaf ends and relatively opaque surfaces. Fig.58 is remarkably like H. retusa, say, at Pienaarsrivier with very awned leaf ends and a high degree of translucence in the leaves.
Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS.Fig. 53 7959 Van Reenens Crest
Set 6. MBB7960 Kruiskloof, E Van Reenens Crest. (figs 65–78)
This population departs from a very broad norm for Van Reenens Crest and picks up the continuity with populations north, east and south. This is most noticeable in the surface texture where the tubercles are smaller and denser. The leaves can be highly rotate, see fig.68. A feature seen occasionally in the Van Reenens Crest populations and as far afield as the Diepkloof (Malgas) area much further south, is the occasional plant with a very stellate (five –pointed star) arrangement of the leaves that are quite horizontal see fig.70. Among the many oddities is the plant in fig.72 with floribundoid leaf ends and a leaf surface where the tubercles seem to be flattened. In fig.73 the tubercles seem to be spined. In fig.75 tubercles are almost absent.
Fig. 65 7960 Kruiskloof
H. retusa ‘nigra’
Set 7. MBB7961 Kruiskloof, E Van Reenens Crest. (figs 79-90)
These plants reinforce my perception that H. retusa and H. mirabilis are uncomfortably closely related. Figs 79 and 80 have the same general form as H. mirabilis ‘sublineata’. Fig.82 appears to me as possibly a hybrid as leaf opacity is deeper and there are marginal spines.
Fig. 79 7961 KruiskloofFig. 82
H. floribunda
Set 8. MBB7962 Niekerkshek. (figs 91-99)
Fig. 91 7962 Niekerkshek
Set 9. MBB7963 Niekerkshek. (figs 100-111)
The plants vary in size and to some degree in colour. This variation in colour from a deep purple to dark grey is very evident in fig.107. The leaves are much flattened and an occasional leaf is pointed. The plant in fig.102 is unusual in having white spots on the leaves. Marginal spination can be absent or very coarse. The leaf surfaces show no venation or translucens, as is expected in this species.
Fig.100 View looking south to Potberg.Fig. 101 7963 Niekerkshek
H. mutica
Set 10. MBB7950 DeDraai Klipbankskloof. (figs 112-123)
Fig. 112 – 7950 Die Draai, E Klipbankskloof
Set 11. MBB7951 Klipbankskloof West 1. (figs 124-126)
Fig. 124 – 7951 Klipbankskloof West 1
Set 12. MBB7952 Klipbankskloof West 2. (figs 127-129)
Fig. 127 – 7952 N Klipbankskloof West 2
Set 13. MBB7953 Klipbankskloof West 3. (figs 130-144)
Fig. 130 – 7953 Klipbankskloof West 3 View westwardsFig. 131 – 7953 N Klipbankskloof westward view of habitatFig. 132- 7953 N Klipbankskloof West 3
Fig. 175 JDV92-64 Very localised.Fig. 176 JDV92-64 Sanddrift
Set 16. JDV85/17 Haarwegskloof. (figs 188-213)
Fig. 188 JDV85-17 Haarwegskloof
There is a very wide range of variants among these 7 population sets. Leaf colour and shape vary enormously and it can only be said that generally the leaves are muticate. Almost invariably any one plant has a leaf or leaves that are also mucronate and in some cases even awned. The leaves may be abbreviated and short and squat or drawn out and elongated. Venation is also varied from many to few or only a single vein on the upper leaf surface. These veins may be straight and clearly separated or they may anastomize and link togther in a reticulate pattern. I did include two pictures of flowers, figs 173 and 174 just to indicate that simply taking two flowers at random one can see obvious differences.
Attention can be paid to figs 145 and 146 of Rondeheuwel where I consider these two plants indicate hybridization with H. mirabilis that does occur about 1km to the northwest. Fig. 147 shows a plant with a rather wrinkled surface.
Conclusion My opinion is that consideration has to be given to the odd way in which the “species” do not share habitat. It is extremely difficult to rationalize an ‘average’ plant for each population and so establish any norm for proper comparison. The populations vary by degree. It is also clear that if one examines the distribution of the species across the study area, it can be observed that H. mirabilis where it occurs with H. retusa ‘nigra’(at Heuningklip and Kruiskloof in the east), is not the same as plants to the west where H. retusa ‘nigra’ is absent. Consideration has to be given to absorption into H. mirabilis. Why H. floribunda should make a re-appearance at Rietkuil and Niekerkshek in the west, when everything points to its absorption into H. mirabilis in the east at Skeiding, is a mystery. The fact that at Niekerkshek it also has radically different forms to those found anywhere else adds to the difficulty.
I will conclude by acknowledging that the populations of H. mutica are all summer flowering but I do not think that this is as significant as the authors of “H. groenewaldii” will contend. The Van Reenens Crest complex demonstrated dramatically that H. retusa and H. mirabilis share a very close common history and an equally close contemporary relationship. My conjecture is that H. mutica is linked to that relationship in the same way the complex of populations are that I assign to H. pygmaea. It is painfully obvious that if floral differences that are so easily detected within populations, expecting to find more evidence in the flowers than exists in the vegetative structures across the genera, and especially the subgenus Haworthia is an absolute pipedream. Floral differences between species are minimal and if anything will provide support for a reduction of species rather than add to the. If any small difference is suggested to separate species, then equal weight must be given to similarities that should then unite species. The contention then that flowers are offering characters that I as an author have ignored, while already wholly untrue, is additionally nonsensical.
Acknowledgement Mr. and Mrs. Trevennan Barry of Van Reenens Crest. Mr. N. Swart of Poststal, Mr. J. duToit of Tarentaal and Wydgelee, Mr. W. duToit of Klipbankskloof, Mr. J. Mathee of Klipbankskloof West, Mr. Vlooi du Toit of Die Kop, Mr. M. Mynhardt of Haarwegskloof, Mr. and Mrs. Nelius Smith of Volmoed, Rondeheuwel, Mr. Adrian Steyn of Wankie Boerdery. Thank you to Kobus Venter and Lawrence Loucka for company and shared observation. ♦
No conclusion is written to this appendix. It is evident to me that there just is no way for me to generate a composite picture for each set of flowers by which one can adequately compare. In general the summer flowering cf. H. retusa set have flowers that are correspondingly lighter coloured than those of the darker flower of the correspondingly darker H. mirabilis set. While gathering the plant pictures from files it struck me how similar plants are from quite different sets across both groups and persuading me that H. retusa and H. mirabilis are in fact one integral system and H. mutica and H. pygamea are western and eastern offshoots.
Acknowledgement I must again appreciate the kindness and helpfulness of landowners in allowing access. Neels and Suzanne Smit of Volmoed; Hector Odendaal of Dankbaar; Jon and Cindy Webber of Klipport; Tom,Trevennan and Hesphia Barry of Van Reenens Crest, and Coetzee and Surieta Uys of Morning Star. The present owner of the Bromberg site could not be established. The other plants photographed are not field plants and neither can they be seen as any more than flowers of one or few cultivated specimens with provenance already acknowledged.
The following additional populations and plants were observed:-
1. MBB7954 H. mutica, Volmoed. 2. MBB6512 H. mutica, Klipport. 3. MBB7741 H. mutica, Dankbaar. 4. & 5. MBB7920 and MBB7921 H. retusa (nigra), Van Reenens Crest. 6. MBB7803 H. rossouwii, Morning Star, S. Heidelberg.
In addition I include pictures from plants in cultivation:-
7. MBB7758 H. retusa, Skietbaan, S Riversdale. 8. MBB7780 H. retusa (geraldii), Komserante, E Riversdale. 9. MBB7781 H. retusa (foucheii), Komserante, E Riversdale. 10. MBB7776 H. retusa, Pienaarsrivier, W. Riversdale. 11. MBB6747 H. pygmaea, Vleesbaai. 12. MBB33 H. comptoniana, Georgida. [ed.] MBB33 doesn’t exist in Bayer’s Accession, correct reference is most likely KG114/72
The population H. rossouwii (elizeae) at Bromberg, Stormsvlei was visited but the plants were only in very early bud.
The Heidelberg H. rossouwii is a small solitary plant and only known to me from two populations. H. rossouwii (elizeae) is also small, proliferous and very problematic. It is so far only known to me as one very restricted population. I doubt if the eastern end of the Riviersonderend Mountains have ever been adequately explored to exclude it from there. Even the Bromberg alone could yield more populations.
MBB7954 H. mutica, Volmoed.
2. MBB6512 H. mutica, Klipport.
3. MBB7741 H. mutica, Dankbaar.
MBB7920 Van Reenens H. retusa (nigra), Crest.
MBB7921 Van Reenens H. retusa (nigra), Crest.
6. MBB7803 H. rossouwii, Morning Star.
7. MBB7758 H. retusa, Skietbaan, S Riversdale.
8. MBB7780 H. retusa (geraldii), Komserante, E Riversdale.
9. MBB7781 H. retusa (foucheii), Komserante, E Riversdale.
10. MBB7776 H. retusa, Pienaarsrivier, W. Riversdale.
There seems to be so much harping about my departure from the International Code of Botanical Nomenclature (ICBN) that I obviously need to try and explain myself better. The real issue is that we are dealing with a group of plants that is largely appreciated for its vegetative characters and not for its small and unexceptional flowers. Because the plants are small and so commonly grown by collectors the numbers of plants in cultivation and close observation is large. The plants also do vary in respect of leaf morphology, arrangement, and surfaces to a greater extent than in many other genera. Furthermore, the variation is also exaggerated by growing conditions. The fact that flowers are not used in the classification process beyond the level of sub-genera means that there is an almost total reliance on vegetative characters for classification. The nomenclatural system in botany tends to be a typological one, which means that reliance is placed on descriptions very often derived from single specimens. This is particularly so when the nomenclatural types are simply old illustrations that have been used to arrive at identifications and names by consecutive authors for decades. Thus use of those identifications and names, and their continuing re-interpretation causes a great deal of either grief or great personal satisfaction depending on just who is being affected by the process. The fact that the names should indicate “species” is lost from sight and totally obscured by the additional absence of any good universally accepted explanation for what a species is or might be.
Explanation in and around all this has been a large part of my writing since I started this in 1962. Thus I will not enlarge on the subject but rather try to explain again this way using a species name generated by Col. C.L. Scott with whom I had some considerable altercation. It must be understood that Col. Scott was not a biologist and it is just a simple fact that the problems that the above brief remarks expose, trouble many professional taxonomists to this day. I do not condemn Col. Scott and express my respect and admiration for him and am very grateful to him for the friendship he later extended to me before he died.
The example I will take is that of his species Haworthia geraldii. It comes from a small hillside east of Riversdale running south to north, named Komserante. This is an Afrikaans term meaning the ridge around a small geographic basin. In the way that haworthias are assembled in small habitat defined and localised populations, there are three recognizable populations of plants (of the subgenus Haworthia) along this one ridge about 1km long. The southern population used to be complemented by still another that occurred where the stream bed left the basin at the southern end. A plant from that southern population was named as H. foucheii by K. von Poellnitz. The northern population is a bit problematic and I initially included it in my then concept of H. magnifica. Since then however, it has become clear to me that my idea of species was too conservative and that H. magnifica as described by von Poellnitz from south east of Riversdale (now the Frehse Reserve) is part and parcel of a huge complex that I regard as H. mirabilis. Thus this northern population is seen by me as H. mirabilis but complicated by the fact that it is largely influenced by hybridization with the next population or populations south. A plant (or perhaps, and doubtfully, plants) have been given the name H. vernalis by the Japanese writer M Hayashi. The name H. geraldii is attached to the second population southwards. The plants are very proliferous, form large clumps and the leaves are usually quite strongly retused (“bent back like a thumb”). The name H. foucheii is simply attached to the third because the plants there tend to be solitary and the leaves are also fairly erect and spreading as described and pictured for the original from the fourth population that was off the ridge.
The problem is now that we have descriptions and plants in each population that do not match the descriptions. While I assign the northern population and all the plants in it to H. mirabilis, other writers use the names H. vernalis, H. magnifica and even the confused name H. asperula. What they mean is by no means clear and in actuality a lot more names and descriptions would then be required to name each of the countless variants there for what might be an original pure species and variants or for first second or third generation hybrids with H. geraldii or H. foucheii as those names were applied.
But the problem is far more extensive than just these four populations. After years and years of field exploration it has become evident to me that like H. mirabilis, H. retusa is also highly diverse and I see it to include all the variants of H. turgida. In fact I suggested a long time ago that H. retusa is simply the solitary large form of H. turgida in low-lying level areas as opposed to the more common and extensive clumping cliff dwelling forms of that species. This is also why I object to the requirements of the ICBN that require the name H. retusa to take precedence for the species for historical reasons when it would be far more realistic to take it as a variant of H. turgida for biological reasons.
It has always been simply evident to me that H. geraldii and H. foucheii are variants of H. retusa. The problem now is that the plants in the two populations vary so much that, while the names are indeed useful for commercial and collector use, there can be problems that the use and application of the names will be confused.
Look at it like this. In respect of H. geraldii; it came from one population at Komserante, Riversdale and not all the plants are the same. I think this one population and all those plants in it belong to the species H. retusa, so I called it H. retusa var. geraldii. (var. = variety). Because not all the plants look the same that means that only some are truly geraldii and only the plants that meet Scott’s description are actually var. geraldii. Because the plants multiply vegetatively and Scott only took a piece, the original plant may still be there. This means that we should recognize it as H. retusa var. geraldii forma geraldii and in cultivation as H. retusa ‘Geraldii’ or just as H. ‘Geraldii’. What do we do with the other 100 plants or more in the population that are not H. retusa ‘Geraldii’? I do not think they can all get names so I take away the capital letter, leave out “var.” and use ‘and’ (inverted commas) to show that the identification is, and will, be a bit uncertain unless you know from a label that the plant comes from that particular population at Komserante. The same applies to H. retusa var. foucheii. In this case the original population is gone and I am not sure if H. ‘Foucheii’ is still in cultivation. So I use the name H. retusa ‘foucheii’ for a second population on Komserante. Some of the plants in this population look like some of those in the H. retusa‘geraldii’ population but not quite like H. ‘Geraldii’ itself. So for me the answer is MBB7780 H. retusa ‘geraldii’, Komserante and MBB7781 H retusa ‘foucheii’, Komserante.
It can be seen that a complication comes in when more than one population is involved. The problem then is that you can put them in a line so that some plants in the first population look like plants in the second, some in the second look like plants in the third, some of those look like those in the fourth until at the end none of the plants in the last population look like any in the first. The trouble in the field is that the line is not straight and it can also go off in different directions. Some of those directions may end up where they started. One simply cannot be truly confident about many of those names that are given to plants. Serious and proper consideration must be given for how different they may be even within the populations. Not to say of the shared similarities and still added variation from geographically adjacent populations. It may happen that I may write H. ‘retusa’ (and add number and place name) for a population that may better (not necessarily correctly) be identified as H. retusaXmirabilis. Other writers who are not biologists and have other considerations in mind may want to give a new name altogether. That new name and description may be again really only for one plant, or maybe a few more in a population, that have real or imagined novelty value and attraction. But they take no account of all of the less attractive variants or other populations that comprise the biological whole by close proximity if for nothing else.
The above discussion is integral to the rationalized list of names that Dr John Manning helped me produce. There are some species names there that I have deliberately presented as synonyms or variants of the species I recognize, because the authors of those names fail to convince me that they have any understanding of the situation at all. The word “species” is apparently simply a convenient naming system for oddities and novelties and not any scientific construct to explain the natural phenomena of living systems and their parts.
To summarize; I have written a report on flower characters in which I refer to the Komserante populations and I have used the following names and my own convention as follows:-
Please note that the way I have omitted the word variety from the following names and also used inverted commas …
7779 H. mirabilis, Komserante (the northern population – I could add ‘vernalis’ but it carries all the baggage of doubt about actual status). 7780 H. retusa ‘geraldii’, Komserante. 7781 H. retusa ‘foucheii’, Komserante. 7920 H. retusa ‘nigra’, Van Reenens Crest.
The omission of the word variety is for two reasons…
1. Economy
2. To convey the idea that the actual indication of status is not certain as I have used the name to indicate a population or populations rather than a single described plant. The prime and overriding uncertainty is that we cannot know what a species is that I have described for Haworthia. Thus how can we possibly rank populations at levels below?
The use of inverted commas reinforces what I want to convey. This is that the individual plants in the populations are variable and it may not be easy to always identify the plants (individual or population) according to a more formal classification.
Any departure from the ICBN or the way the names are treated in formal botany conveys the difficulty that I personally find in trying to reconcile formal nomenclature with names that are so often tied to single plants.
Previously published Cact. Succ. J. (Los Angeles) 84(1): 41-50.
Map Legend – east of Swellendam. 1. JDV84/75 Haworthia retusa ‘turgida’. 2. MBB6666 H. retusa ‘nigra’↔ H. mirabilis. 3. MBB7898 H. retusa ‘nigra’. 4. MBB7899 H. retusa ‘nigra’. 5. MBB7897 H. retusa ‘nigra’. 6. MBB7896 H. retusa ‘nigra’. 7. MBB7871 H. mirabilis. 8. MBB7823 H. mirabilis. 9. MBB7909 H. mirabilis 10. MBB7805 H. mirabilis. 11. MBB7801 H. mutica ‘groenewaldii’. 12. MBB7886-7889 H. mutica ‘groenewaldii’, H. mirabilis, H. minima, H. marginata. 13. MBB7722 H. floribunda ‘major’
1. Haworthiamarginata and H. minima
The Robustipedunculares is quite a distinctive group within the currently recognized genus. While the four species in the group are generally quite distinct, there are some remarkable complexities that rival that elsewhere in the genus. Recognizable and obvious hybrids are found between H. marginata, H. pumila and H. minima, but there are instances of whole populations that appear to consist of such in-between forms e.g. H. Xmortonii. I have speculated that perhaps rather than have just hybridized, the species have never ever really truly separated in the supposed evolutionary process. There is a vast body of variants that still link them in intimately as this piece will show.
I long ago observed that a small remnant population of ostensibly H. minima just south of Swellendam flowered in November as opposed to the general rule for the Robustupedunculares as late summer flowering. A vicarious population at Brandrivier north of the Langeberg (H. minima ‘opalina’) also flowers in November and both populations have fairly large and white flowers for the species.
I have recorded the normal bluish-green H. minima within the Bontebok Park at Swellendam as well as a very green variant. But in very recent exploration to the south-east we found an even more divergent group of plants that, while varying among the plants, seemed to be hybrids of H. marginata and H. minima (fig. 2 as a single sample and not representative of all).It was September and there were no signs of old or new flower spikes. Kobus Venter, who was present, remarked that the first plant seen was reminiscent of the plants once present south of Swellendam. The plants were large and in exposed situations even colored brownish as does H. pumila. No flowers were present and their color may have shown if H. pumila could have been directly involved at all, while it is essentially its distribution restricted eastwards from the Robertson Karoo by some 20km that reduce the possibility.
What makes the situation more interesting is that nearby was a population of H. marginata that was flowering and the flowers were also large and very white for the species (figs 3 & 4). Added to the fun was a smaller probable hybrid (fig. 5).
Differences and complexities like this do not really surprise me because it is what I have come to expect in my many wanderings in the field. The problem is that it certainly makes classification and any agreement on a set of names very difficult. I just accept it as a fact that plant species can exhibit greater differences between individuals of the same species than between individuals of different species, ridiculous as it may sound. This is because I perceive species as systems of individuals in populations with a very strong geographic component. To actually make a decision it is frequently necessary to determine just what else is growing in the vicinity in respect not only of the genus in question, but also that of other plants. Even the habitat factors need to be considered.
In the case of the plants pictured with this, the Bontebok Park terrain is mostly tertiary gravels, while the habitat we found the plants in was more recent riverine boulders. It is very curious that in the description of H. groenewaldii, the habitat is implicitly described as Ruens Silcrete. I do not think this is true. It is in the true Ferricrete – Silcrete that we found the next and it seems to be these differences in substrate geology that play a large role in generating variation and consequent controversy.
Fig. 2 MBB7891 cf H. minima, RotterdamFig. 3 MBB7892 H. marginata, RotterdamFig. 4 MBB7892 H. marginata, RotterdamFig. 5 MBB7891 H. minimaXmarginata, Rotterdam
2. Haworthia retusa ‘nigra’
I first allied this element with H. mutica because this is how G.G. Smith referred to his collection from Kransriviermond south of Heidelberg. Since then there have been many new collections from which can be gathered that this population is a hybrid one between H. retusa ‘turgida’ and H. mirabilis. There is another collection north of this at Morning Star that appears to have the same parentage but also including H. floribunda. Then there are populations continuing up the Duiwenhoks and then Klip rivers to northwest of Heidelberg, a population between Heidelberg and Tradouw Pass further west and also a population at Goedverwagting south from there. Apart from the Morning Star population (February – March) these are all September/October flowering. There is a population at the southern entrance to Tradouw Pass of the same ilk that is February/March flowering. Then there is quite a distance between these known populations and a remarkable population at Buffeljags south of Tradouw Pass. It is remarkable for the fact that it is lauded as a new species viz. H. groenewaldii when I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February – March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time.
To explore the realities of the situation we undertook two expeditions, one was to Buffeljags to explore west of H. groenewaldii and the other was to the Tradouw Pass area to explore H. retusa ‘nigra’. The first exploration yielded three populations of ‘groenewaldii’, which convince me that despite its flowering time as for H. marginata above, is simply H. mutica in another guise. I also think far too much is made of superficial and trivial differences that are as much characteristic of the variation in the one original population as they are within the four populations and for H. mutuca in its full sense. I consider it significant that H. mirabilis in its more normal non-retuse and dark green form is present in discrete populations both at and west of Buffeljags.
The second expedition was nearly as fruitful. It showed the Tradouw Pass population to be February/March flowering (see figs 6 & 7), while three new populations we discovered between there and the previous records in the easterly Heidelberg direction were September – October flowering. They link up to the populations elsewhere that I assign to H. retusa ‘nigra’. An additional find by Jannie Groenewald, for whom that H. mutica viz. H. groenewaldii, variant is named, also took us to a population of what is clearly H. mirabilis (see map) as I know it in its many disguises in the white kaolinic/bentonite clays of the silcrete – ferricrete inselbergs throughout the low-lying Southern Cape. There is unquestionably an overlap of characters between what I assign to H. mirabilis and H. retusa and I consider inarguable that H. mutica is a reflection of a shared gene pool.
Fig. 6 MBB6666 H. retusa ‘nigra’, Tradouw PassFig. 7 MBB6666 H. retusa ‘nigra’, Tradouw Pass
What this demonstrates again, as does the Kiewietsvlakte populations between Heidelberg and Riversdale, that H. retusa and H. mirabilis are closely intertwined from east to west. There is an added complexity that H. floribunda is admixed along the northern populations and H. variegata along the southern. The admixture of the two species produces H. retusa ‘turgida’ and H. pygmaea in the east and H. mirabilis, H. mutica and H. retusa ‘turgida’ in the west. This is complicated by the other interactions along the northern and southern areas. In the Potberg area it appears that the genetic material of all five “species” is evident in the populations that I have seen there.
While I would like to explain the situation around H. groenewaldii I. Breuer that I interpret as a variant of H. mutica, this should be left for another occasion as too many images are required to support any argument. As it is, the issue of H. mutica ‘nigra’ occupies 29 pages and 79 illustrations in my book Haworthia Update Vol.2 pt 1:50, where all the above mentioned variants are discussed and illustrated. The naming of Haworthia is highly contentious because the species consist of aggregates of small fairly isolated populations that may differ to large or small degree. The populations are in turn also aggregates of plants that can all be identical from vegetative propagation, similar because of low genetic difference or very different from each other because of large genetic differences. Therefore figures 6-13 simply show just a sample of the variation within these four populations in the Tradouw Pass area. The plants vary quite considerably in size too and the one in Fig 8 is nearly 200mm diameter. I have added to the map locations of the only significant other populations that I know of in the area including H. retusa ‘turgida’ and H. floribunda ‘major’, excluding those within the Bontebok Park viz. H. minima, H. mirabilis and H. marginata.
Fig. 8 H. retusa ‘nigra’, HeuningklipFig. 9 H. retusa ‘nigra’, HeuningklipFig. 10 H. retusa ‘nigra’, HeuningklipFig. 11 H. retusa ‘nigra’, HeuningklipFig. 12 H. retusa ‘nigra’, HeuningklipFig. 13 H. retusa ‘nigra’, HeuningklipFig. 14 The typical ‘pressure burst’ of white kaolinic soil from under more solid ferricrete where H. retusa and H. mirabilis are commonly found.
Perhaps I should close by explaining that I have dropped the use of any rank below that of the species name. I simply am suggesting that we recognize the need for a trinomial system without the pretension of status, and more greatly honour the binomial as an entity of a greater significance than we may know. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable specimen that is used to anchor the Latin name.
Acknowledgement. Any proper excursion into Haworthia territory always requires acknowledgement of landowners and I thank Jaap Viljoen and Jannie Groenewald for organizing that and for their company. I was also glad to have Kobus Venter along who had persuaded me to show him some of populations known to me on promise of new exploration.
Note. Cross seasonal hybrids observed are-
H. retusa turgida X H. floribunda Blackdown, Heidelberg.
And also Platjieskop, Riversdale.
H. pygmaea X H. floribunda Coopert Siding, Albertinia. H. mirabilis X H. retusa Soetmelksrivier, Riversdale H. mirabilis X H. variegata Stoffelsriver, Swellendam.
References. I need to record that Harry Mays of Alsterworthia kindly undertook the non-profitable publication of 5 volumes of Updates (2-6) between 2006 and 2009. Vol. 1 was published by Umdaus in 2001. These volumes were the product of research to validate or correct what appeared as a formal revision in Haworthia Revisited, published by Umdaus in 1999. The description of H. groenewaldii appears in Alsterworthia 11.2:15 (2011). ♦
I recently wrote an essay on the situation between Haworthia retusa and Haworthia mirabilis at Komserante east of Riversdale. The essay was entitled “My view of names” and is posted on the HaworthiaUpdates.org web site. Etwin Aslander posted some pictures from what he called Kruisrivier. These caught my eye because they did not look like the plants I know from a place of the same name. My known population is JDV95/62 and generally these plants have the dark colour and rough surface texture of H. mirabilis. The issue is that they are spring flowering whereas H. mirabilis is generally considered and observed to be late summer flowering. Etwin indicated to me where he had found his plants and I duly went to look.
In the process I incidentally called on a well known H. retusa population at the Skietbaan locality south of Riversdale. There has been a dramatic turnabout in the appearances of these plants since I last looked there 2 years ago. Whereas there were then huge clones well above ground level, the plants were now again smaller and drawn into the ground. I experienced this dramatic shift in plant appearances just west of the Frehse Reserve many years ago when there were giant size plants as opposed to my first visit when the plants were really small and withdrawn.
Kobus accompanied Daphne and I to Kruisrivier where the owners Wilhelm and Mandi Zietsman were extremely helpful. They told us also of a neighbor, Gert van Rensburg, who had also seen the same plants on his farm to the west. Mandi accompanied us on a jaunt to find that farmer and failing that we explored north of the original Kruisrivier locality. There we found another population of plants as well as H. floribunda (see Set 1 MBB7998). These two species H. retusa and H. floribunda were occupying different habitat and spaced about 100m apart. The H. floribunda was numerous and rather smooth leaved as well as paler green in colour than I expect from that species. The H. retusa-like plants were much smoother in surface texture than the original known population and they were in bud (see Set 2 MBB7999). We went back to the older population just to confirm that they were in bud too as we expected. Just so and the buds were just emerging from the rosettes. The plants were generally smaller than they were at a previous visit (see set 3 JDV95/62).
We parted company with Mandi Zietsman, and went off westwards intended to explore the Klein Kruisrivier area that seemed to better fit Etwin’s site indicator. By good fortune we ran into Gert van Rensburg of Wegwysersrivier. He eyed us very suspiciously indeed and obviously very reluctant to show anyone the plants. However, he very kindly relented, took us to the spot and left us to freely photograph and explore (see set 4 MBB8000). The plants can be described as midway between the generally rougher surfaces of JDV95-62 and the smooth surfaces of MBB7999. What was more dramatic is that there were six flower spikes so that flowering is possible as early as July 6th.
We returned via another route regretting leaving distant habitat unexplored. But we did find another population of H. floribunda, a little more toothed and perhaps brighter green than at Kruisrivier.
I also note that I long ago confirmed Smith’s record for H. retusa ‘turgida’ at Klein Kruisrivier in the upper Wegwysersrivier Gorge. This is the small spinose proliferous version known elsewhere from the Langeberg Mts.
Digesting this new information is a bit difficult in view of the very opposed views of what names mean and how they should be applied. Taking all the populations that I have explored and written about, my perspective is further to a view expressed in Haworthia Update 7. This is that H. retusa and H. mirabilis are uncomfortably close. The only thing that appears to separate them is the yellowish green and smooth tendency in H. retusa and the darkish green and surface rough tendency in H. mirabilis. Further to that is of course the question of spring flowering versus late summer flowering. But I have already reported several case of hybridization across this divide as well as the Komserante situation. Here we now have plants in three populations that occupy middle ground and one of these populations has a significant degree of a winter flowering capacity. The identification should perhaps utilize the chemical equilibrium symbol. This is not quite it “↔” as the better symbol comprises halved arrows pointing in opposite directions.
I wish to add that in the case of plants I attribute to H. ‘turgida’ at Towerlands, I commented on the very real possibility of a close connection to H. emelyae. There is also evidence for this elsewhere. I use the name ‘turgida’ like this because of the uncertainty of it really being H. retusa var. turgida or perhaps H. pygmaea.
My experience in other situations viz. H. limifolia, H. herbacea/H. reticulata, H. arachnoidea/H. mucronata, H. cymbiformis/H. cooperi, Kiewietsvlakte etc. all suggests to me that the view of species is grossly distorted in the splitter direction. It is clear to me, if to no one else, that H. retusa and H. mirabilis form a very cohesive entity with ramifying oddities the length and breadth of the distribution range. I do not cover this issue here, but there is the added complication of the involvement of H. floribunda. It seems to be very discrete in most places, whereas at others it seems to get lost mainly (only?) in H. mirabilis. This may be because the introgression is favoured by the same flowering season. H. retusa and H. mirabilis are drifted apart by the difference in flowering season but it is by no means anything more than a general observation.
I have added the images of the available flowers as well as that of a bud to show the flared fishtail bud-tip that the southern Cape species tend to have. The flowers are variable and it is difficult to make a statement that characterizes them i.e. no composite image forms.
Acknowledgement I would like to acknowledge Etwin Aslander’s input. Wilhelm and Mandi Zietsman were extremely helpful. Gert van Rensburg was surprisingly well informed about the plants too and very generous in his attitude to us. Part of the pleasure of field work is meeting people like this.
Set 1. MBB7998 H. floribunda Kruisrivier
Set 2. MBB7999 H. retusa Kruisrivier
Set 3. JDV92/65 H. retusa Kruisrivier
Set 4. MBB8000 H. retusa Wegwyserivier
MBB8000 flower faces
MBB8000 flower profiles
MBB8000 flower bud
Addendum To demonstrate the problem of similar looking plants that appear in different populations, I take 3 plants from the original Kruisrivier population (JDV92/65) see figs 1 to 3. Fig. 1 is obviously a mirabiloid plant and if this population flowered in late summer it would probably be identified as H. mirabilis. The figs 2 and 3 are more retusoid. I leave out plants from the newer Kruisrivier population (MBB7999) because none of the plants have the rougher mirabiloid leaf surfaces. I add a Wegwysersrivier (fig. 4 MBB8000) plant that is again mirabiloid and like Fig. 1 except that it appears to be a spring flowering population with a significant number of plants in flower in early July. From there I take a plant from Komserante (MBB7779) that flowers in late summer but is apparently generally hybrid with H. retusa. Moving eastwards from Riversdale and impinging on H. mirabilis splendens, I show a plant MBB7762 from Platkop (fig. 6) where both H. mirabilis and H. retusa occur with occasional hybrids. Fig. 7 is MBB7818 H. mirabilis Windsor SE Riversdale, where the plants frequently have a frosted appearance because of minute surface spines.
There is a significant geographic jump with fig. 8 MBB7850 H. emelyae north of the Langeberg at Aasvoelvallei. This is a population that I have noted elsewhere that highlights the probable relationship of H. emelyae with the H. retusa turgida and pymaeaoid elements from Herbertsdale eastwards. Fig. 9 is a plant of MBB6666 Tradouw Pass that I recognize as a hybrid population H. mirabilisXretusa. Inland from there are several populations, MBB7899 is H. mirabilis, Heuningklip (fig. 10) and MBB7896 H. retusa nigra also Heuningklip (fig. 11). East of that are three populations of H. mirabilis, MBB7912 and MBB7913 Rietkuil and MBB7919 Van Reenens Crest (figs 12 to 14).
As only single plant comparisons, it seems fairly safe to say that, bar flowering time and figs 2 and 3, they are all similar. However, the variability in each of these populations is great and this has been reported elsewhere in the Update volumes. If one had to now take figs 2 and 3 and look for similarities in other populations, it would be very easy to demonstrate a complete gradation from what could be construed as typical H. mirabilis through to typical H. retusa through a large array of populations.
Fig. 1. JDV95-62.9 H. retusa, KruisrivierFig. 2. JDV92-65 H. retusa, KruisrivierFig. 3. JDV92-65 H. retusa, KruisrivierFig. 4. MBB8000.5 H. retusa, WegwysersrivierFig. 5. MBB7779 H. mirabilis. KomseranteFig. 6. MBB7762, H. mirabilis, W Platkop 4616Fig. 7. MBB7818, H. retusa, WindsorFig. 8. MBB7850, H. emelyae, AasvoelvalleiFig. 9. MBB6666 H. mirabilisX retusa, TradouwFig. 10. MBB7899 H. mirabilis, HeuningklipFig. 11. MBB7896 H. retusa nigra, HeuningklipFig. 12. MBB7919 H. mirabilis, Van Reenens CrestFig. 13. MBB7912, H. mirabilis, RietkuilFig. 14. MBB7913 H. mirabilis, Rietkuil
It has long been my contention that there is no separation between Haworthia retusa and Haworthia turgida. It is one very variable system viz H. retusa, with a larger fairly non-proliferous plants tending to level areas and then smaller proliferous plants on steeper habitats. There is huge variability among members of any one population and of course much more between populations. Over and above this is the relationship of this apparently one single system, with H. mirabilis that is probably even more complex and varied. If one takes all the known populations and variants into consideration it become necessary to ask if H. retusa and H. mirabilis are also not just elements of one system, and one species. If all the considerations are summed and referral is made to vegetation and speciation drivers; what constitutes an area of endemism, then I am sure the answer will be “Yes”! What seems to have happened is a natural sequence. As sampling has progressed so has there been recognition of differences. The logical outcome is that sampling progression should lead to understanding and synthesis by reduction. Unfortunately there will be diehards that stay with the differences syndrome and cannot see the similarities.
There is surely no longer any doubt that Haworthia classification has been confounded. There are several factors. One is the historical one of sampling and naming. Sampling dates from the seventeenth century and description based on a few words and weak illustration. The second problem has been a nomenclatural system that revolves around the single types and assumes that departures from that still allow comfortable accommodation of all other departures (variants) under the primary name. The third problem is the absence of a species definition deriving from a lack of knowledge and hence understanding of what species are. The fourth problem is enthusiasts, writers, collectors, editors, reader etc. who generate and propagate within the confines of their own needs, limitations, knowledge and understanding. Too often they are not adequately informed to undertake something that really should be the task of professional botanists. Of course it is also true that professionals have not proved to be faultless either, as simply the lack of a species definition alone indicates.
From my personal point of view, I consider the sampling history and the nomenclatural constrictions of priority and automatic creation of type elements, among the main obstacles to a classification solution. Perhaps it is only secondary to the human factor where writers become anxious to establish their own opinions, based on who knows what, crowing from the top of a metaphorical farm dungheap. I have paid a lot of attention to an element that I name as H. retusa ‘nigra’. It is based on a very unrepresentative specimen from Kransriviermond where it seems as if it is a product of hybridization between H. retusa and H. mirabilis. But this situation, and the name, is inextricably involved in a series of populations grouped in areas like Van Reenens Crest (Swellendam), Klip River (W Heidelberg) and Kiewietsvlakte (W Riversdale). These populations do not seem to figure in any earlier exploration and if looked at objectively may now perhaps be seen as the mother lode from which H. retusa, H. mirabilis, H. pygmaea, H. mutica and even H. emelyae as species may emerge. The reality is that all these may not in fact even be discrete species.
In the forerunning article I discussed some new populations at Kruisrivier northeast of Riversdale and where they occur along the Kruis River. This river partially follows the interface of the Cape Sandstone Fold Mountains in an east/west direction and the situation is replicated to some measure elsewhere between Worcester and George. Most relevant is the similarity of the Kruis River valley and its south banks to the situation along the Klipriver west of Heidelberg. But it is not in the scope of this discussion to cover that now and I would in any case need to research the geology of both areas to do so. I will just present images to cover firstly the area, then to allow creation of a composite image of what the small sandstone H. retusa looks like, then images from just two populations of H. retusa to show how shadowy our image of this species is; and then I will show the images of plants from five points down a distance of about 4km along the south bank of the Kruis River to a point south and west of the better known Kruisrivier plants.
Setting the scene. The Kruis River Valley is slightly east of north from Riversdale and the Kruis River is a headwater of the Goukou River that enters the sea at Stilbay. The views I have (figs. 1-4) are of a satellite image of the Valley from where the Kruis River exits the Langeberg Range to where it joins the Goukou. Three images are from the south bank looking northwest and northeast. The area is very heavily impacted by alien vegetation. The grazing animals are mainly cattle (beed and dairy) and hoof impact on the slopes is severe. The underlying shale along the south bank is not exposed and there is both boulder deposit, river alluvium and even some marine terrace in evidence. Nevertheless the area is not so dramatically varied geologically as is the area around Heidelberg to the west. Hence the transitions and interactions between species and their variants are both similar but very different. The initial conditions will be very different too.
Fig. 1 An overview of the Kruis River Valley.
Fig. 2 Some indication of the topography of the south bank.Fig. 3 View northwest from a central position.Fig. 4 View to the northwest across the flood plain.
The populations. The first three sets are to briefly review H. retusa. The general perception of H. retusa is very restrictive and similarly the case with what was H. turgida. I consider them together. Nevertheless I have to sympathize with the splitter approach because I have to make a concession for the sake of discussion. Thus…
Set 1 MBB7895 H. retusa ‘caespitosa’, Diepkloof, Heidelberg (figs 5-9). I explored the upper Kruis River in early 1970 and found the small sandstone version of what was then recognised as H. caespitosa. Later seen as H. turgida, it is known at many places from the Tradouw Pass, north of Heidelberg, Garcia Pass and Kok’s peak. Small and very proliferous, its leaves are also quite spinose on margins and keel and also flecked with reticulate and longitudinal translucence. The summer coloration tends to reds and yellows. I do not have images of this actual form but show five from a population south of Heidelberg that is very similar. But it should not be assumed that the transitional flow in the Heideberg area equates that at Kruis River. The variants down the length of the Klip and Duiwenhoks Rivers is another profound story.
Set 2 MBB7758 H. retusa, Skietbaan, S Riversdale (figs 10-19 ). I show images of this species very recently taken. This is because I do not think that as a species H. retusa is properly appreciated for all its variants. It is this reality that we are confronted with when considering populations as at Komserante and now Kruisrivier where there is an evidence that it cannot any longer be seen as completely removed from H. mirabilis.
If images 10a and b appear to be different from the rest of this wet, they should. They are plants from the same locality but they were taken 4 years ago when most of the plants were very much larger. They illustrate a very useful point – the plants do NOT necessarily look the same all of the time.
Set 3 MBB8010 H. retusa, 8km S Riversdale (figs 20-28 ). Primarily one should consider surface roughness and colouration as main separators from H. mirabilis. Flowering time is the prime differentiator and this is what is now to be seen in another light.
Set 4 MBB8004 H. retusa↔mirabilis, Bloekombos, Kruisriver (figs 29-42). Reference can perhaps be made to the three sets illustrated in the preceding article. Here is will just re-capitulate to say that there is some doubt in respect of plant appearance and colouration, to examine the view that the represent H. retusa or H. mirabilis. (I dismiss totally and absolutely the notion that a third species needs to be fabricated to accommodate differences impossible to itemize or catalogue across the range of variants within and between populations.) I do not think that the similarity of surface roughness to that of the H. mirabilis ‘atrofuscoids’ should be overlooked – even some of the leaves show the tendency to roundness at the ends. This particular population was in full flower at the end of July.
Subset 4 The flowers (figs 43-45). I do not see anything to distinguish these flowers from those photographed for the Wegwysersrivier population and given in the preceding article. It is difficult to imagine that the flowers may offer any distinctive features in the direction of difference that are not already offered by vegetative and geographic considerations. There may of course be direction towards similarity.
Set 4 H. retusa-mirabilis, MBB 8004 Bloekombos, Kruisriver – Subset 4a Flower profiles
Set 4 H. retusa-mirabilis, MBB8004 Bloekombos, Kruisriver – 45. Subset 4b Flower faces
Set 4 H. retusa-mirabilis, MBB8004 Bloekombos, Kruisriver – 45. Subset 4c
Set 5 MBB8003 H. retusa↔mirabilis, W Bloekombos, Kruisriver (figs 46-73). Curiously this population was only in early bud although only a few hundred meters away from the preceding. I do think there is significant difference in the appearances of the plants with more translucence of narrower leaves. However, plants from this and the preceding population could very easily be said to identify with those in any of the H. mirabilis populations southeast, south and west of Riversdale. It is also obvious that there is a degree of similarity to the rougher plants of H. retusa illustrated in sets 2 and 3, and also to the Komserante H. retusa↔mirabilis plants.
Set 6 MBB8007 H. retusa↔mirabilis, 600m W Bloekombos, Kruisriver (figs 74-81). The banks west of set 5 are less steep and the terrain not clearly differentiated into discrete habitats. Heavy cattle trampling would severely impact on plants. Nevertheless we did find a small colony and are confident more would be found if more time was spent searching. The area is quite extensive and we did not consider several north/south valleys that could well harbor plants. The same is true for quite large hinterland to the Kruis River Valley and Etwin Aslander’s plants, similar to the ones we saw, are surely present back there. This small colony reflects further transition to H. retusa ‘turgida’. The leaves are smoother and more translucent. There are some floral differences muted by the small sample and by the fact that these features such as the absence of frilled base to the bract and the presence of a short pedicel, are variations observed within populations elsewhere.
Set 7 MBB8008 H. retusa↔mirabilis, 1km W Bloekombos, Kruisriver (figs 82-83). Also barely, registering as a population, these odd plants support the predicted presence of a direct transition from the Kruisriver mirabiloid-like H. retusa to the known sandstone forms of H. retusa ‘turgida’. Hoof action has severely impacted this area and it sems very probable that there was a larger population at some time or another. The slope militates against the solitary ground-level growth form of the more usual H. retusa but not steep or rocky enough to be home to the turgidoid cliff growing forms.
Set 8 MBB8005 H. retusa ‘turgida’, S Wegwysersrivier (figs 84-98). The use of the equilibrium sign for the preceding is more informative than accurate. It reflects the problem of decision making where it is not clear if H. mirabilis is even in the area. This of course impacts on the situation further east when the reality of H. pygmaea and all its associated variants are brought into the picture. I am sure it is inarguable that we are now confronted with plants that are truly similar to H. retusa ‘turgida’. Curiously the flower is much the same as in MBB8003 and 8004 to the east but there is a difference in that the peduncles were softer and shorter than of plants to the east and suggesting the shorter lax habit of the cliff dwellers. The habitat is unlike the preceding in that the Tertiary deposit is more alluvial and the rock is more boulder-like. Trampling is still a severe problem. Note the two pictures of a H. retusaXfloribunda hybrid.
Set 9 MBB8006 H. floribunda, S Wegwysersrivier (figs 99-107). The presence of this species is no surprise. It was quite abundant although slightly separated from where we found the H. retusa ‘turgida’. It depends on rocks for protection from trampling. This species of course generally flowers in late summer but once again here is a hybrid present with the other parent flowering a good 6 months later. This has been observed in several other places where these same two species occur in close association. [photo file names are mistakenly named MBB8005, they are in fact MBB8006 – ed.]
Conclusion The product of this exploration was based on prediction and I would wish that cognizance was taken of the fact that this could and can be done. Using an aggregate or group category and a host of Latin binomials is not botany and obfuscates things. Perhaps to the benefit of traders, but certainly confusing to the collector who make wish to know more about plants than simply having a label.
Acknowledgement I have already acknowledged the help of Wilhelm and Mandi Zietsman; and Gert and Lynette van Rensburg. We were now also assisted by Wessie Wessels, a retired airline pilot who owns the Bloekomsbos Farm. Kobus Venter came to see my pictures and I really value his interest and useful comment. I wish there was more similar interested and informed comment. ♦
A review of the plants photographed recently around Albertinia … I only know of 23 populations in this area that relate in different ways to the names Haworthia turgida, H. retusa, H. mirabilis, H.emelyae, H. pygmaea, H. splendens, H. vincentii, H. acuminata, H. fusca, H. dekenahii, H. argenteo-maculosa, H. pygmaea, and H. esterhuizenii. I still need to get pictures of H. splendens, H. vincentii, and H. turgida on the Wydersriver.
In essence there are two variants – the clumping cliff dwellers (H. turgida) and the mostly solitary flats dwellers (H. retusa). The flat dwellers are far more variable than the cliff dwellers. If all the variation is considered against the differences in each of the habitats, it seems very unreasonable to suggest that these populations are anything but a single species (H. retusa) expressing itself differently according to local dictates and demands. This is same issue of variability occurs in these plants all the way westwards.
97. 2020.03.04 – Esterhuizenii – Aasvoelberg. I was concerned that I am not sure anymore of flowering time because essentially retusa and mirabilis are separated by a late summer vs a spring flowering. None of the populations I saw now in late February showed any sign of buds or flower. Only the Humor plants had just two old inflorescence of about two months age. The fact it is a SYSTEM. The retusa elements are lighter coloured signified by having an element “pallidifolia”, while the mirabilis element has the element “nigra”. Recognising species as systems, variation and differences can be seen in a more realistic way. This does help to overcome the pressures that ignorance, egocentrism, emotion, commercialism and intellectualism bring to bear on classification. H. retusa ‘esterhuizenii’?
The map is useful although it does raise the question of how confidential should we be about locality. Part of this quest was to examine how populations have fared in the years since “discovery” and also in the 50 years since I have been directly involved. My observation is that the main threat is excessive animal pressure – and/or combined with drought. The Aasvoelberg population seems to be very small and a secondary threat is alien weed infestation. But I really do not know the actual size and health of the population. I did see plants further west long ago, but it will be a real mission to explore there properly. An important point about conservation is that it is really only as systems that species can cope with natural and even man-imposed change. It is only with a proper and true definition of the term “species” that we can even discuss them sensibly. “Adaptive” must surely be part of that?
98. 2020.03.05 – This is H. retusa (turgida) at Buisplaas on the Gouritz River between the Gouritz Bridge and Herbertsdale. This form was informally known as H. rodinii and is known from the Gouritz Bridge and also south of that. As a cliff dweller it is normally clumping. The first time I saw such a plant it was a specimen reputed to have come from Die Hell much further north in the Swartberg Mts. VERY unlikely, but H. retusa should be sought in the Gouritspoort where the river passes through the Langeberg Mts. There are many populations of these cliff dwellers and countless variants. I do not and cannot, dispute the fact that names present a problem for suppliers and collectors, but that is not a problem that can be settled by science. Association of a botanical binomial and place is the only solution that makes sense to me. Taking out the geographical detail diminishes the sense of any name. But this series is to explain and expand on this issue.
These are also Buisplaas but in a totally different habitat several hundred meters from the cliff dweller. One picture shows a very proliferous clone. This same juxtaposition of clumping and non-clumping occurs elsewhere and is a pronounced feature of the H. cooperi and H. cymbiformis problem in the Eastern Cape. These Buisplaas populations validate my conviction that the Western cape retusoids are in fact a single species system. There has been a predictive element in all my exploration since the revision of 1999 Haworthia Revisited and this population simply fits a pattern. Comparing all the pictures of this series I am now showing will demonstrate this too. H. retusa ‘pygmaea’. I write the name like this because in the absence of a species definition, formal lower ranks have to be meaningless. Furthermore the nature of local variation is such that it is generally impossible to circumscribe ranks. A very good example of this is the persistent reference to H. mirabilis var. magnifica which can absolutely not be supported on the basis of a type specimen representing an original group of such similar or like plants.
99. 2020.03.07 – Despite the drought and animal activity (and human predation presumably) – the Cooper Siding plants are in great health. I am posting all my pictures (of Haworthia – there are many other plant species of significance present) because the reality of diversity within populations gets zero attention in citizen science and little more in academia. But do not get me wrong. I write “citizen science” not citizen cognition. Also I do not ignore the fact that “citizen science” also features in academia. These plants (a plant?) was described as H. dekenahii var. argenteo-maculosa. In my Handbooks 1976, 1981), I placed the two under H. retusa. Was this wrong? No – because moving them to H. pygmaea as I did in my revision (1999) highlights the reality that this is a single system and not just spatially isolated populations that can be treated as different.
In writing about Albertinia and the conservation state of these Haworthia populations (that really bothers me), some very serious thoughts come to mind. Do we really want to know the problem and the answer(s)? The problem is environmental deterioration – climate change. This is coupled with the decline in moral values across the counter. But do we want to acknowledge this and really want to know what is true. That all of existence is actually a grand illusion that we may choose to be locked in to. I should not have suggested that climate change is the main factor – although Albertinia has been experiencing the worst drought in living memory. Livestock pressure also definitely impacts negatively on vegetation, and our dietary addictions prevent us recognizing this fact.
100. 2020 03.08 – Dekenahii is at Draaihoek on the Valse River northeast of Albertinia. Juxtaposed again on a nearby rocky cliff is “turgida pallidifolia”. A floribunda / chlorocantha variant is in close proximity in its own preferred space. A different form of the turgida retusoid used to be on north facing cliffs a little further north at Weltevrede. Keep in mind that I am considering a bigger picture in which differences within populations negate differences between them, and especially when habitat is considered. I also juggled dekenahii between retusa and pygmaea and whatever you do with it, the best explanation I can offer as a binomial is H. retusa ‘pygmaea’. Better still is to add the unwritten meaning that it is actually Helgaard Oosthuizen’s discovery of H. retusa as a group of plants at Draaihoek circa 1943 given to Smith by Dekenahii. Names standing alone may be good for business but the interest lies in the information attached to them and which they point to.
This is the “turgida” form of H. retusa at Draaihoek. These plants have more pointed leaves and are more colourful than the “pallidifolia” that I remember 59 years back. But the precise locality may have been different. I remember plants more like those at Tweekuile further east along the river that I will still post pictures of. At Weltevrede further upstream the plants had longer leaves. Steven Molteno coincidentally reminded me of H. marumiana and the exact same conundrum I am trying to explain here, but magnified manifold over the vastness of the Karoo and our ignorance of both marumiana and the Karoo.
101. 2020.03.10 – Tweekuile is north of Albertinia and just a few km east of Draaihoek on the Valse River. This is what H. retusa ‘turgida’ looks like there. It occurs to me that sensible taxonomy, like prevention and cure of disease, is not a sustainable business model 😅
Without any simple and practical measure, how does one deal with an endless array of slightly different looking things affected by age, exposure to direct sun and an also seemingly endless lot of agencies affecting appearance. Compare these two Cooper Siding and Humor pygmeas with the Tweekuile turgidas.
102. 2020.03.11 – Have a good look at the rest of Humor. I suppose it is still fashionable to dismiss variation like this as “hybrid swarm”. Problem here is what parents where and when. To be strictly correct, is there really any option but to say H. retusa ‘pygmaea’? ♦
103. 2020.03.11 – Melkhoutfontein. The weird history of “acuminata” is not very flattering. It started with me when I described this as a variety of H. retusa. But I did say some interesting things in 1981 that lead to my present point of view. One is that I said that acuminata may be eventually shown to link H. retusa and H. pygmaea. This is exactly what the present thread is about without any conscious effort to support what I might have predicted. I think I may have changed acuminata to a variety of H. mirabilis in Haworthia Revisited? Weird too, but I would not be ashamed. That would reflect the reality – it is one system and countless opinions can be woven around the evidence that supports the idea. The original population of acuminata was destroyed and although I did find plants nearby I did not think such a strong population as this still existed so close by. This population is at Melkhoutfontein on the Gouritz River southeast of Albertinia. The habitat is old river terrace. There were several smaller populations to the north. Coming are populations to the east that inspire further support for the kind of continuities that occur between practically all Haworthia populations.
104. 2020.03.16 – Johnson’s Post plants are also something else. Large clumps on clay pedestals on an ancient river bank? An amazing sight.
105. 2020.03.17 – These plants are at Vleesbaai and I have no doubt that a species name has been coined for them without condemning the need for a name. It is the formal species part that bothers me. From my perspective these are H. retusa ‘pygmaea’. Steven Molteno can give a good habitat description. The conservation aspect is interesting because it is a small rocky area in an expanse of cultivated and heavily stocked lands. The real threat to the plants is the severe stocking pressure and the irony is that grazing is also necessary to keep vegetation under control. I will, I’m sure, come back to the conservation issue.
Steven Molteno – If these are the plants I’m thinking of (they look like it) then they’re all growing on a lone outcrop of quartzitic sandstone of the Skurweberg formation (Ss), Table Mountain Grp. This exposure is a rather eroded ridge very near Vleesbaai, surrounded by a lot of calcrete and alluvium I think.
There’s another (much larger) exposure of this same formation (Ss), not far away to the north. It’s that line of tall hills that runs parallel to the N2, south of the road, and east of Albertinia. Interesting that this larger Skurweberg formation ridge is also home to some unusual mirabiloid-retusoid confluences (the ‘vincentii’ & co. towards the western end of that ridge of hills; pygmaea ‘argenteo-maculosa’ towards its eastern end).
I think this correlation with a specific geological formation is very suggestive of broader connections across this whole Gouritz region.
Soils are very rudimentary on these outcrops. Lithic leptosols, plants growing very close to (or in) bedrock. Eroded grit from quarzitic Skurweberg formation is probably rather acidic, especially with the contribution of broken-down organic material that’s pretty much its only supplement!
Up near Johnson’s Post it’s a completely different story though.
There, the geology is very complex, with lots of funny “slivers” of different formations, all parallel to each other but zig-zagging across the flats.
At Johnson’s Post though, the river’s carved out a series of little kloofs in the terrain. The central valley floor is quite wide and filled with alluvium. The haworthias grow along +-north-facing slopes of Kirkwood formation (Kk). It’s often reddish, but here it’s a light-brown (almost a little ‘greenish’?) siltstone with greyish shales and bit of sandstone sometimes. This is what was called the “variegated marls”, and it erodes into a heavy, fine-grained clayey loam. Very muddy if wet. The same formation (Kk) is the main home for H.pygmaea in the Mossel bay and Brakke regions to the east. There is a bit up near Herbertsdale, and a long strip of it going east-west through Riversdale and Heidelberg. Another patch is just east of Swellendam, and I think there’s probably an exposure at Klaasvoogds but its very eroded. Everywhere except Klaasvoogds, it’s inhabited by the floribunda/chloracantha complex. Sometimes also these funny mirabilis-retusa integrades like near Heidelberg.
To return to Johnson’s Post, above the Kirkwood formation you get this pebbly calcarenite (De Hoopvlei fm I think), but it’s on the flatter area on top and I don’t know of Haworthia there. It’s also mostly ploughed though.
Landtypes at Johnson’s Post are also interesting. Eastern part is Fc15. Fc15-39 are lime-rich river valleys, networking across most of the Overberg, that are always inhabited by retusa ‘turgida’ and by one of the floribundoids. Usually not a mirabilis though.
Western part of Johnson’s Post is landtype Ae133 (obviously these aren’t strict boundaries though, they grade). The south-eastern extremity of the same landtype (Ae133) is where those Haworthia at Vleesbaai live – i.e. Ae133 connects these two populations.
There’s a similar landtype, Ae113, just north of the Breede River mouth (!)
Vegtype at Johnson’s Post is what Jan Vlok calls: Southern Cape Valley Thicket. It’s a thin belt that winds along the Gouritz and its tributaries, with a strip along the lower Goukou too. It nearly always has some sort of toothed-leaved floribundoid/chloracanthoid (along the lower Goukou its H. variegata) as well as H. retusa turgida. The correlation in distribution is quite strong. These haworthias almost seem to trace out this vegtype on the map. I think the correlation with a specific geological formation is very suggestive of broader connections across the whole Gouritz region. These sandstones are very much older than the surrounding sediments.
106. 2020.03.18 – This is Paulsfontein just west of Albertinia. A small population in an intensively grazed and weed encroached area. A reminder to keep in mind within and between population variation. This has the Latin epithet ‘fusca‘ and I personally fuddle over whether to address it as pygmaea or mirablis. So I suggest H. retusa because these populations simply mirror what I was showing for populations to the west where names like mutica, groenewaldii, badia, joleeniae, bobii, hammeri etc. are collector useful. Conservation is poking me in the ribs but I am not finding the right ambit for that discussion.
A very powerful supporting argument for my thesis of H. retusa as a seemingly massive conglomerate, is the close association with H. floribunda. H. chloracantha, H. parksiana and H. variegata; as well as in the continuities one sees among those ‘species’.
Essie Esterhuizen wrote (31 July 2021) … Populations in that area confuse me. I am not sure whether I deal with the western end of Haworthia pygmaea or eastern end of Haworthia retusa.
Bruce Bayer replied … Delighted to hear Essie’s voice. Essie I do not know what you have followed of mine and I have no idea of where your thoughts have taken you. My conclusions are that we have all got things wrong about botanical classification and that Haworthia is just showing us. So I think species are vast fractal systems. At the least H. retusa (incl. turgida) mirabilis, mutica, pygmaea and emelyae are a single system?
I am so glad Essie has commented because I have just felt so in need of expressing my present feelings about the aloids generally. There was a congress on Aloe classification about 10 years ago that was hailed as a breakthrough for the subject. I saw it as a total farce because the essential nature of species was ignored. My experience and slight knowledge of the DNA methodology applied in the latest solution for the genera (and species), is that it is supremely cringeworthy. All it gets right is the three Haworthioid genera where sequencing was not needed to reach that conclusion. Part of my fears for the worst are based on the fact that two of the major sequencing studies were initiated to explore the phylogeny of just Haworthia species. That the results were written up to totally avoid discussion of the initial hypothesis will never cease to confound me. My confidence in aloid classification is at zeropoint.
107. 2020.03.19 – I wish I had pictures from my visit to this site about 20 years ago. The place? Lodewykstenk east of Albertinia and east of Aasvoelberg – esterhuizenii. There were many plants. This time I really struggled to find only these 3. Why? Certainly not collectors. Drought and severe animal pressure. The plants have the name H. vincentii. But really. Can you name plants in casual fashion like this without a knowledge of what the name is directed at? A basic life form. I consider that it fits comfortably with all these plants I have been dealing with here.
108. 2020.03.19 – Flat or cliff? What the thesis is that these plants belong in a single gene pool and the enormous variability is due to the demands of habitat as much as anything else. Cliff is vertical rock. A level habitat may have the same rock to soil ratio. Whatever the vast range of non-steep rock sites, most significant is that animals (livestock) cannot graze or trample a vertical rock wall? This is then the cliff hanging form of H. retusa east of Tweekuile, north of Albertinia, but a bit off the Valse River. A fortuitous new record to demonstrate just how much exploration is still needed. ♦
109. 2020.03.22 – Soutpankoppies. What is there to do? The Albertinia area is really absorbing. Westwards the gene pool (retusa/mirabilis/turgida/mutica) is confused as it tries to express itself in so many different ways, whereas at Albertinia it simplifies to two and, in my opinion, then one. At the same time there is a second gene pool comprising floribunda/chloracantha/variegata/parksiana. The area is not well explored and there are clearly small islands of vegetation that may still hold some secrets. The photographs here are of ‘splendens’ at Soutpankoppies that I perceive to be the most eastern expression of ‘mirabilis’. Retusa in its typical form ends a little to the west. Further exploration in the Albertinia area will surely expose the facts about these two hypothesized gene pools.
“Splendens”. Not for nothing is it specially protected in an enclosure. But this underlines the real problem of conservation – a huge subject. This particular enclosure is of interest for several reasons. When I first heard of it I assumed it was where the plants were seen in ca 1996. But it is not. So what has happened to that small population? There was also a third ‘population’ in the immediate vicinity of an old small farmstead. Local disturbance and alien weed invasion seem to have annihilated that one. The splendens enclosure is hugely successful in respect of the short term health of the specific population and even satellite imagery demonstrates that. The impact of livestock activity and weed invasion outside of the enclosure are obvious. But the total effect is confounded by the presence of surface rock (ferricrete?). The splendens population would have been as safe in the absence of any fence. The other two populations were far more vulnerable. It is simply not practical to enclose vulnerable ‘species’ and this was a hot topic in professional conservation as far back as 1970 if not before. We cannot ignore the problem of taxonomy, because the essence of conservation is surely the preservation of diversity. At what level now does one do this? The current trend in conservation is the identification of “red-data” species. I think this is complete nonsense because it amounts to plum-picking in which the end-product is the maintenance some kind of museum record of what was?
110. 2020.03.23 – An enthusiast commented that Haworthia magnifica made sense to him. Well here are pictures of plants at the type locality (Riversdale Commonage or now known as the Werner Frehse Reserve) as was indicated to me by the collector of the original material i.e. J. Dekenah. If you want confirmation that the name, as a species name, makes sense it is worth using google to locate the type and see the images that come up. What would be useful is for that enthusiast to set out to demonstrate why this particular name makes sense to him against all the other names that may do the same.
This is probably what H. magnifica is imagined to be, based on my experience with the name, the trade and the literature. But the fact is the population at the Frehse Reserve is substantially fragmented and the plants are highly variable. Not only that, anyone who has seen a retusa/mirabilis hybrid will recognise that influence in the pictures I’ve posted.
111. 2020.03.25 – Here is a set of pictures of plants propagated from seed from a H. mirabilis population (MBB6651) 2km further south from the Frehse Reserve and what was purported to be H. magnifica. The habitat is a bit different because here the plants are in white clay (kaolinite) whereas at Frehse they are in shale. What is common to these plants that makes them different to an impossible to define commonality of H. magnifica???? It is a problem throughout ALL the mirabiloids and retusoids of the Southern cape and extending to the emelyoids of the Karoo. Examine the last picture and you will see the ultimate trickster. The floribundoids (variegata, chloracantha and parksiana) have a show-in too.
The coloration in this plant is what we see in the old ‘mutica var. nigra‘ population. When one talks of magnifca one cannot dismiss a series of mirabilis populations that occur from Frehse Reserve at Riversdale westwards to Riviersonderend. ♦
Lawrence suggested I finish what I had to say about the retusa combobulation. Actually I find myself just plain exhausted trying to explain why I name things the way I do when so many otherwise knowledgeable enthusiasts are in real ignorance about the fundamental problems of Haworthia classification and identification. I was just writing about populations of plants in a very wide area between Steytlerville and Oudtshoorn, explaining that it was nigh impossible to decide whether they are decipiens, mucronata, bolusii or cooperi once the easy conclusion was made that they were not arachnoidea. This all reduces argument in the retusoids to absurdity. Someone tell me what these are and why?
I felt the need to say something about how, at the end of my useful life, what my feelings are about (my) classification of Haworthia. It so happens that I was extraordinarily fortunate to have had the confidences of some very distinguished people as they closed their lives down. People like Prof. R H. Compton, Dr R.A. Dyer, Dr Alan Codd, Mrs A. Obermeyer, Dr Courtney-Latimer, Dr O. Hilyard and others in botany. also in other sciences like Dr H.K. Munro, Dr B.K. Petty, Dr B. Smit from entomology. Col. C.L. Scott came to the same place. The common factor was an admission that really they did not know very much. I thought a word that would cover the state of mind would be “pragmatism”. That is until I checked the definition of the word, “Perceptual inattention to the specific functions comprising inquiry led realists and idealists alike to formulate accounts of knowledge that project the products of extensive abstraction back onto experience.” What about the product of my experience “experience” – the knowledge that you acquire by the time you do not need it anymore) projecting abstraction??? The funny thing is that the rules of nomenclature confound the purpose of classification. The purpose of classification is to identify all the basic life forms and to organise them in the system and order of their historic origins. Just as pragmatism can be left unexplained and confounded in many words and pages of incomprehensible text, so is my classification of Haworthia. A point is “groenewaldii”, where I suggest it really should be H. mutica var. groenewaldii. This is a conceptual trap! Combobulation cannot be avoided. All the evidence I see is that there is a single system that what we call turgida is the strongest possible contender for origin. The problem is that nomenclatural rule is that if retusa has its origin in turgida, the name retusa must be upheld for the system. Do you follow that??? In my explanation of the “species” mutica, I explain it as emanating from the interaction of mirabilis and retusa. So it is quite wrong to use the combination of mutica var. groenwaldii. It should be retusa var. groenewaldii and still more correctly H. turgida var. groenewaldii. I am presently really just enjoying sharing my thoughts about some “species” that will surely trouble anyone wanting certainty in identification and names. ♦
Here are maps attempting to illustrate relationships and a reality of “species”. The BW map was the one presented in Salisbury 1976. The colour map was done a few years ago and I may (need to) work on the legend along the lines of the MTR model. But I do feel disturbed by Gerhard’s unnecessary and unkind remarks. The fact is that I said long ago that Haworthia as was, would not be understood until the generic problem was resolved. At least PRE accepts that. But there is a bigger and deeper problem that I cannot get through to anyone despite the obvious. We have a classification system that is antiquated and dysfunctional based on a philosophy and methodology that does not work for Haworthia and MANY other genera. There is no general and precise species definition and there cannot be while we are restricted to a mechanistic world view that wholly ignores the fact that the physical world we occupy is a projection from a far greater metaphysical reality that we are programed to forget. Species are not just mechanistic products of a whimsical nature. Books like Sheldrake’s Morphogenesis, or Lovelock’s Gaia Hypothesis, and Capra The Turning Point (many more) are not sucked out of thin air. Neither are the Bible, The AdiGranth, the Baghavad Gita and also many more. But we do not need to go to that level to realise that we are getting nowhere pursuing a mindless argument. We do not even have to know the metaphysical reality while we think we can use a two dimensional clado- or phylo-gram to illustrate a three-dimensional issue of time and space.
For the sake of clarity, red is retusa and pink is turgida but do not make a mistake and think there is a clear separation.
The dotted lines are really a guess at a situation before the Younger Dryas? 12,000 years ago They suggest a southern connection of the mirabiloids were I have no doubt that mirabilis ‘bobii’ directly connects to mirabilis ‘paradoxa’. I would love to paste my updates stuff all over again. Trouble is I only got a digital camera about 2006 and as it is, posting one population a day would need about 3 years.
I understand there are people who are offended by my opinions and the frustrations that I generously express. But there are real problems that commercial and lesser needs prefer to wish away. Seeing H. fusca did not require for me to wait months to see any connection to H. retusa. It simply confirmed what I have been tediously trying to explain to an audience that has an element reluctant to listen. Or reluctant to read and think objectively. Why do these people want to lead enthusiasts away from what may be true? The fact is that DNA sequencing has not been able to resolve the nature of species in these retusoid/mirabiloid populations. I do not think that next-generation sequencing is going to change anything either. Here is a very rough map I drew to try and explain the situation of a single gene pool (one system = one species). I use R in the map for retusa for nomenclatural priority when it is actually turgida (thus T) that dominates. True retusa is centered on Riversdale and Heidelberg in the centre. Rather than throw rocks at me, I would be grateful for constructive ideas based on observable and quantifiable facts. If I add a smile will it alkalise the dyspepsia?
Here is a map to show the phytogeographic regions of the South African southwest – the Fynbos area. The fact is that vegetation classification like this is no easier or better than plant classification. It is full or errors and omissions. I wish I had the time, the skills, the knowledge to produce an accurate map of the distribution of the aloid species as an overlay. The map here used in a book on the Cape bulbs, comes from a book for the Fynbos as a whole. The discussion reads that these areas are primarily determined by rainfall but it sure is more complicated than that. The areas demarcated all lie within a greater area that is actually a winter rainfall one. Temperatures are never truly continental, nights are generally mild and actually there seem to be two main non-growing times viz July/August when its cold, and then Jan/Feb when it is hot and dry. What I do know is that the Haworthia genera and species can be mapped and understood in exactly this way. Nature has not just haphazardly sprinkled stuff about and had a randomly orchestrated ball.
From my point of view, you need to understand that retusa is not a separate unit. It is part of this one system = one species, Look at my coloured map and ask what names go to what colours? In fact the colours merge into each other and it is difficult to say where a name stops and starts. Again it is a problem that you might not have been able to see much of the literature where this is explained. It was about 1972 I discussed the sequence of populations from N Heidelberg from the sandstone mountains (turgida caespitosa) all the way to the sea then east and up to Riversdale where it was retusa. Atrofusca is not a separate unit but you can use atrofuscoid because there are populations you can characterise as atrofusca. You cannot do that for magnifica for example. ?? It is not funny to see what the relationship is between atrofusca and floribunda that I have also described in the literature. ??
MBB7998 – Even if this IS a species, how do you collate all this variation into a single meaningful, usable species description and which plant do you select as a “type”. G.G. Smith fell at this hurdle long before (fortunately he had finished his manuscript) Except some never learned nothing and continued in the same vein – taking one plant to cook up a description?
MBB7999 Kruisriver. These plants all fall on an interface of turgida and mirabilis.
MBB8000 Kruisriver. I am sure this is formally named somewhere. If these guys are honest it is several times. Fabulous place and people. To our amazement they had a pet chameleon and also a pal with a dozen pet tarantulas. Our one regret is having met so many great people who were so friendly and hospitable that it was awful to experience this as a kind of one-off and never see them again!
Goodness me a floodgate of thoughts and a spider waiting to catch me in its web ?? This is turgida ‘pallidifolia’ growing nearly cheek by jowl, with dekenahii (var. of retusa or pygmaea?). The classic cliff face/flat terrain situation that marks the difference between the generally clump-forming turgida and the solitary flatter withdrawn retusa? But this is not the only place. Throughout the Southern Cape we have these juxtapositions of M R and T. Old methods have not solved these problems and old ideas will not either. If critics or doubters would please just kindly and coolly, look in an unprejudiced or jaundiced way at the published information. I have tracked all of mirabilis in a really large scale operations like at Kruisriver, Komserante, Kewietsvlakte, Vermaaklikheid, Potberg, Malagas, Haarwegskloof, Napier, VanReenens Crest etc. (North of this locality by about 1km used to be another turgida population on a cliff face was that more retusoid than this one).
I would so love to post each population in the spider web map of relationships. It would take months/years at a population/day. My good friends fellow Haworthia name appliers and commercial outlets hate me for this. You need to know that Daphne, Kobus and I explored the greater area thoroughly and found about 15 populations or more. Only in one other location did we find two species and they were also contiguous but floribunda and retusa – also a cross season hybrid observed. How other Haworthia pundits evade these realities and the classification issues would need Einstein or Heisenberg or Jung or Kinsey to explain. Note Kinsey was a famous psychologist of a more recent era.
MBB8003 from the area NE Riversdale as circled on map. Yes you guessed right. I did not know whether to file these as H. retusa or H. mirabilis because it is in a dramatic series that forms a continuum turgida ‘caespitosa’ (=mirabilis) to a more mirabiloid thing.
Yes you guessed right, again. I did not know whether to file these as H. retusa or H. mirabilis. Why for example is H. vernalis seen as a more useful name than H. turgida ‘vernalis’ or H. retusa ‘vernalis’ (to observe rules priority). It is because it is more profitable I guess.
H. retusa and H. mirabilis are a dramatic series that form a continuum of turgida ‘caespitosa’ (=mirabilis) to a more mirabiloid thing.
A real problem is that people just can’t keep up. Sometimes they form strong opinions but are unable it seems, to maintain a discussion or keep relevant. We have seen 8003 and 8005 and now 8004. still in that small circled area NW Riversdale I refer to as Kruisriver. I have these under retusa but would be equally happy with them in mirabilis and with the best of intention, very confounded if someone suggested some other name. Thank goodness for emojis where people read venom dripping fangs where there is simply a plea for common sense.
These are upstream from that last lot in the same circled area but to the west 2km? Note the floribunda hybrid! I filed this under turgida!!! But I am immensely frustrated by Emile’s excellent post. Take my map for example. It has been around since 1976 and not a soul of all these potential collaborators has so much as whispered a syllable to suggest where it might be right or wrong. That has not stopped them from littering the web with names with no explanation of how they might fit and why? The name “magnifica” makes sense!!! I cannot even use it as “magnificaoid” – I will get to post pictures of this ? “species” “variety” “hodge-podge “. If you give me time. Emile can you tell where there are discontinuities in this lot – even where floribunda or rossouwii are concerned? Who do consider an authority on Lithops? Is there a communal solution or is there a truth? It is said that the camel is the product of consensus.
Retusa Travels was first published as a series of Facebook posts from May and June 2022.
Here are pictures of Kweekkraal Haworthia mirabilis MBB8064 – some plants can be deemed to be atrofuscoid, others magnificoid and even heidelbergensoid. Kweekkraal has come up before in my exposé of “H. mutica var. nigra” and at this stage of my journey I conclude that the “nigra” issue was just one small issue showing that there is a single system that unites retusa (including turgida) and mirabilis.
More MBB8064 … Quite a lot on the mind with a mind boggling amount of new material. Talking to Lawrence and Essie about utilizing Haworthiaupdates.org more intensely as publication is very time consuming and also very limiting in the sense of illustration that tells most of the story. Kweekkraal is some 10km west of Riversdale and the area is the focus of my first article on the H. mutica ‘nigra’ anomaly. There is so much feeling about the disclosure of locality that I think is counterproductive. These plants are just as much under stress from porcupines, grazing animals, other farming activity, while there are clear laws in place about trespass and plant removal. My number for this collection is MBB8064, a new and recent ‘location’ and it was incredibly difficult to find the plants at all. People need to switch to camera capture, and I will post more photographs of this site to demonstrate variability. One could say H. mirabilis “atrofuscoid”. i.e. many if the plants are reminiscent of what was described as H. atrofusca. The name can be kept and used in that sense while technically the actual Latin binomial refers to a very complex and large SYSTEM with many variables. So a formal name is really just a matter of convention and agreement.
Essie brought this population of mirabiloids to my attention. It is also Kweekkraal as part of Kiewietsvlakte that figures in my first essay on “How to understand H. mutica var. nigra“. H. floribunda occurs as discrete and also lost in the milieu. H. retusa is present. H. mirabilis occurs as the atrofuscoid form in the east, a magnificoid cocktail in the middle and heidelbergensoid to the west. Kweekkraal is a Guest Farm and there is a large area of conserved Renosterveld. My reference number for these is MBB6085.
More of MBB6085 at Kweekkraal west of Riversdale. A mirabilis potpourri. Do look at the shrinkage of the plants. This is at the end of the dry season and there would normally have been rain by now. Plants can look very different as the seasons come and go. Who can tell me the difference between a low-end intellectual enjoying plants in the field, and a top-end intellectual in a white coat sitting at the controls of the Large Hadron Collider trying to manipulate dark matter? There is a lot in this question!!
The last of MBB8065 – a point of interest is porcupine activity. I have seen this again and again and also where the damage is extensive. I reported it to conservation management who replied quoting a “scientific” report on porcupine activity at Nieuwoudtville where there is very little threat of land transforming to agriculture, The conclusion is that Porcupines are an essential part of the ecosytem. When they do become a nuisance in urban areas they are trapped and moved to conservation areas. Well I agree. Under natural conditions porcupines and nature go together. But enter man and his greedy and destructive ways; the situation is different.
This retusa (retusoid) is another of Essie’s finds and is very close to the splendens (mirabiloid) localities which would suggest that they might be discrete systems. This is not the only situation that does this, but looking at all our known populations, we still conclude that it is the nature of the beast as it is drawn and quartered by nature! Incidentally, we struggled to find plants where many had been reported. Why? Immense porcupine activity. There used to be floribunda at this place too. Since degraded by agricultural activity and is destined to be totally cultivated in the near future.
With Essie near a retusa pilgrimage spot. Essie has explored so much of this area and here he is indicating what he has seen and where. The plants here were in bad shape compared with some previous visits and porcupines have also been very busy. Sometimes chewing off the leaves and the plants survive, but more often grubbing them out.
Re those silver flecks. I feel constrained to post this here and now. There has been and is so much ill-feeling and misunderstanding in Haworthia classification that I have been accused of initiating while also bearing the brunt of it. Here is a quote from a great Haworthia personality who finds me unbearable …”the silvery flecks in the leaves of H. jakubii suggest connection to H. magnifica and H. pygmaea.” That is interesting because in the photograph MBB7818 Windsor 2, there are clearly silver flecks that this exceptional man denies it is determination to prove me wrong about the species status of H. jakubii. I do not find them in any of my magnifica (mirabilis) pictures from the type locality or in MBB6651 from nearby, but I do find them in H. retusa. I would be so grateful if this was shared around and brought to the attention of all Haworthia enthusiasts. Essie has been caught up in the mess too and I am enlisting his help to solve the problem. We have together written two articles in the process and are stuck in the third seemingly because of the wish to avoid further injury.
Essie showed me these retusa at the Riversdale town edge. Municipalities universally feel it is necessary to have a great swath of vandalistic dumping to signify their dominance over nature and make human presence known. So its a bit painful to walk on the margins and bid what will surely be a goodbye. But do also watch for flecks and see if you can decide how you would describe H. retusa.
Daphne Bayer exploring.
More retusa from west of Riversdale. This species also has that enormous range of variation that makes classification so awkward and contentious. Some of those quirky leaf shapes and tips occur in H. mutica too.
I could quite happily go back to Riversdale for another 20 pictures of these captivating plants. I was on about “white flecks”. Us mad taxonomists are always looking for characters by which to make outrageous claims, but some of these things like flecks and windows can be pretty behavioral beyond just variables in a population.
The significance of the flecks is that it is critical in relation to the recognition of jakubii as a species in view of the arguments put forward by proponents. The argument is made that jakubii has these flecks whereas my collection at Windsor a little further is supposed to not have them. Further argument is made that my photographs are just fleeting images and bear no relation to close observation in cultivation. So I can show that these are plants at Windsor that do have flecks, not only that, they appear in more species where this antagonistic proponent implies their absence. Both Essie and I simply observe that jakubi is in the continuum of H. mirabilis, and the fleck argument is void.
This is a photo of a pygmaea from Humor. These plants were named argenteo-maculosa because of the silver markings. There are actually two kinds. The narrow sharply defined and the more diffuse larger markings along the veins. The former could be oxalate crystals and the latter aggregates of chloroplast filled cells. It should be understood that there are more than 20 features in leaf anatomy that some scientists feel could actually be used alone to identify down to species level. My experience and opinion is that it only leads to very emphatic opinions than are not always true. In fact generally quite false. An example is my experience where I was called in as a practical field observer to support results obtained for surface studies where atomic absorption photography had been used to make big decisions. Why was that necessary at all?
More retusa – it is quite instructive to examine windowing in the individual plants. It would be very difficult to quantify. Flecking either as chlorenchema aggregates or oxalate crystals if that is what they are – is relatively absent.
This diagram of interconnectedness and continuity has its roots in a first draft drawn in 1966 when it was pretty clear that there were clinal gradients from population to population of Haworthia. This has been demonstrated time and time again. Essie has independently come to exactly the same conclusion from the same careful and considered exploration and observation. All these elements on the map are somehow connected and are best understood in this context. This has nothing to do with simply providing labels and names for collectors – it is about providing a basis for a deeper appreciation of the plants and their origins. In fact, of nature itself.
The blue is the mirabiloids, pink the turgidoids, red retusoids, orange rossouwoids, red crosses muticoids. There are species names in the map. Just remember the lines connect discrete localised populations. Ideally each populations should be “identified” and individually mapped.
What now? I think I am going to carry on with retusa as the red-line drawn on that map. It is totally implicated in turgida as a complementary element. The two are really different in respect of habitat occurrence and degree of proliferation AND not always! I have had these all published and I have posted before, but I want to write with new emphasis supported by what Essie has to say. He is posting on his page… Haworthia veld memories and we are busy writing together as a team. Together we can make this map more significant. These are Essie’s “close east of Riversdale” retusa.
Another West of Riversdale population with pictures taken by Warwick. Take note that populations differ from one another just as the plants within them do too.
These are about the most southerly of retusa.
I have often taken a pilgrimage to this site S Riversdale and the population always seems a bit different. I posted a few pictures of plants from here a few days ago taken May this year. The present pictures were taken 1st Aug 2012. Bud are just pushing. Wait until you can compare these with pictures taken 2008 and 2020. Habitat pics should be time-stamped to take into account the shifting dynamics of field populations. Perhaps it would be better to say that some research ought to be undertaken to explore population level changes. Just note that many of these populations are not somewhere you just walk out the back door to see. With modern photography I would suggest a minimum of 20 good photographs for any one habitat – which means take about 50! The underlying truth is that taxonomy has to move away from tinkering with names, to considering the impact of time and conditions.
Sept 2008 S Riversdale. Plants in flower and looking great?
January 2020 and the plants colouring up in the sun.
This is another of Essie’s discoveries and particularly interesting because it highlights the reality of the relationship between the recognised species. I regard this population as Mirabiloid retusa because over the whole Southern Cape there is ample evidence that despite gross differences in flowering time of the two “species” involved, they do “hybridise”, but more significantly that they are part and parcel of one system, which calls for a better definition of what the word “species” actually means.
This is close to east Riversdale where there used to be four populations of this plant. The southern one was gobbled up by cultivation and was the origin of the name “fouchei” that was an erect leaved non-proliferating form of H. retusa. Semblances of that form occur in the next population that is some 300m north …
A few hundred meters north is another very well known population H. retusa “geraldii”. When I first saw these plants ca 1971 the plants were in huge flat clusters over 400mm diam. With time the clusters broke up and were far less impressive. Retusa is normally a fairly non-proliferating element.
There is direct continuity between turgida and retusa. They do not “mix”. I also think now that it is incredibly more complex and that we only have the two groups … the very dark green late-summer flowering mirabilis and the light green/yellow spring flowering turgida. These two match and mix to produce a host of variants of which the main are mutica, pygmaea and emelyae. I do not know quite how herbacea/rerticulata/maculata/pubescens come out in the west.
I wrote in 1981 …”The species problem very seldom arises where species grow together, but rather from the difficulty in associating populations strung out in isolated populations.” This in itself is a rather weak explanation. Here I have just posted two H. retusa populations but there is a third population that I ascribe to H. mirabilis that others say is H. asperula. Curiously I took two sets of pictures for that because I found there to be two sub-populations closely adjacent … this is one of them…
In contrast. About 30m away in vegetation unprotected from grazing animals for the past 80+years, thus impacted vegetation and skeletal soil, is this bunch of variables.
by J.M. “Essie” Esterhuizen and M. Bruce Bayer. While Bruce Bayer is the writer of this piece is by convention prime author, Essie Esterhuizen is cited as such for the following reason; it honours an astonishing amount of field work and profound insights that many taxonomic botanists never acquire. It is instigated by Essie’s more recent exploration and comments he has made that reflect the writer’s own conclusions. Thus this article is intended simply to mirror the thoughts of open minds looking for answers in a domain dominated by academics.
Introduction and general preamble The two of us have been long interested in nature and particularly Haworthia. We have a combined experience of at least 120 years of exploring the subject that we would like to share. Neither of us makes any claims to some or other authority or qualification that gives us more authority than anyone else. We have simply had an enormous privilege to have spent so much time in the field learning about these fascinating plants. Just like anyone interested in anything, we have needed a vocabulary and words or names to understand, organize and communicate our thoughts and words. This is either as internal private mental processes or as external communication with other people. The most important words we need are designations and names for the very objects of our interest. Perhaps for our own purposes we do not need words at all, but for external communication we certainly do. Then it becomes problematic because agreement, definitions and all kinds of conventions and guidelines exist that may not correlate with our individual capabilities, character, belief or value systems. In fact all this may clash with what our deepest individual understanding tells us.
The facts are that we have an organized system of information gathering and exploration we term ‘science’. Within that we have various disciplines of which botany exists for the plant kingdom. Within that we have a sub-discipline (taxonomy) of description, identification and classification. To guide this process international rules of nomenclature are agreed upon to provide a powerful framework of names that Essie and I now try to evaluate as a framework for our observations in that group of plants we know as Haworthia. Already there is a problem because Haworthia have been known in history as Aloe at the birth of the Linnaean system before being separated as Haworthia. At present there is one faction that maintains this arrangement and another that expounds Haworthiasenso stricto, Haworthiopsis, and Tulista as a formalization of three distinct sub-groups (subgenera) that both of us have no problems with.
This all seems very good and well until one faces the harsh reality that science as a discipline is concerned with physical creation. Rather than serving the purpose of information gathering across every single aspect of sentient life, it is in fact restricted to what has been described as mechanistic materialism. This is where explanations are sought for anything and everything by processes of experimentation, measurement, analysis, reason and logic. Plant taxonomy, at its base, rests on the principle that there are basic life forms (species) as in the animal world and that there are basic discrete life forms kept apart by virtue of their closed reproductive systems. This is derived or adapted from zoology and tacitly transferred to botany when there is enough evidence to demonstrate that fertility can extend across generic barriers. There is in fact no species definition. There is also no recognition of metaphysical realities or even the possibilities of their existence. Curiously a formidable intellectual who has edited and largely authored a massive six volume encyclopedia of mystic terms by John Davidson M.A. (Cantab) concluded that the definition of the species concept as a basic life form was not the same for science and mysticism. This is patently absurd.
Endless debate can ensue, but Essie and I, in our quest to understand Haworthia, have simply wanted to know what these basic forms are in practical terms so that we can organize and communicate our interests and thoughts with each other or with anyone else that may have common interest. In our quest we have simply used the hundreds and hundreds of names that have been coined for Haworthia species to come to know what these tell us and how useful they are to us in understanding them. Rationally each name is based on the tacit perception that it applies to a group of plants that are, or potentially are, interbreeding. This is blatantly untrue and no good definition actually exists to our knowledge that in any way regulates what is formally described as a species. An example could be cited where a single plant specimen has been selected to represent a hypothetical, fictitious species. No name in mentioned to avoid hurt. So we (the authors) have labored under this handicap where no one knows what a name even means, in trying to use and apply the many names sensibly. Both of us have written extensively about our field experience and what we will do now is collaborate in this article to explain how we have used what is available to explain the relationship of various populations. This is with the recognition and realization that the plants occur in highly localized limited localities, are highly variable and yet somehow seem to be integrated and related according to a geographic modality.
It is simply not rational that we may only confidently apply one or other of the many names if we know where a plant comes from. Lawrence Loucka has done an incredibly good job in creating a compendium of Bayer’s writing in a website Haworthia Updates (https://haworthiaupdates.org/). This culminates in a set of what were Facebook postings (in Haworthia Shucked ) under the title “Mysteries”). In these, particularly “part 36”, it is posited that named species such as retusa, turgida, mirabilis, mutica, pygmaea and emelyae are in fact one and the same species. All Bayer’s writing since the publication of his Haworthia Revisited in 1999 is in pursuit of the goal that Essie independently undertook circa 1980 viz. Just what constitutes Haworthia in respect of different life forms that intrigue all of us even it we do not cognitively know this. The section “Mysteries” in Haworthia Updates deals specifically with Bayer’s exploration in the Albertinia area while Esterhuizen has evidence from an additional six localities that provide the very evidence that Bayer was expecting and looking for.
Together we thus consider that the six species are in fact one single system according to a definition of plant species that we agree on and repeat here … A species is a complex dynamic conscious fractal system of living organisms that are metaphysically, and physically (genetically, morphologically, physiologically, behaviorally) continuous in space and time. It seems logical that these systems must be open as it the same one consciousness that pervades all of creation, and surely that is incontestable. Furthermore there is constant change and not necessarily at all in an interminably slow Darwinian selective way. Both of us have observed dramatic changes within our time spans of observation.
The evidence The evidence for our conclusions is in fact already out there in Haworthia Updates and also in Essie’s more recent Facebook Page Haworthia Veld Memories (https://www.facebook.com/Haworthia-Veld-Memories-110859041540965/). Here we simply post a selected set of 54 pictures of plants from 27 populations from between Riversdale and Albertinia to as far east as Great Brak, east of Mossel Bay. The published names, primarily of species (sic!) rank our present discussion covers are the six already named viz. retusa, turgida, mirabilis, mutica, pygmaea and emelyae, and then fusca, splendens, dekenahii, argenteomaculosa, acuminata, rodinii, pallidifolia, suberecta, esterhuizenii and vincentii. There are no doubt many more names that sprout from a need that should not be met by resort to formal botanical taxonomy or nomenclature. No further discussion of those names is deemed necessary because that is what the literature on the subject already does.
Very important in consideration of these pictures, is the question of randomness and bias. The pictures we have selected were based on selecting what is reasonable from those that are available. Unfortunately the range of pictures available is generally very limited for most populations. Secondly it is difficult to avoid any kind of bias when photographing. Populations can be extremely variable and one can select pictures to either prove similarity or difference. This needs then to be adequately weighted to geographical relationships and habitat. As an example of the latter point is that it the contention of the authors that turgida is really a cliff hanging variant of a flats dwelling retusa. Just how does one formulate a single image from a mélange of different ones? This is probably where the difficulty really lies in reaching agreement because we may simply be doing so differently.
Essie’ perspective. (I have adapted Essie’s direct written contribution to this third person version to try and get a better fit …) Understanding Haworthia is like putting together a giant jig-saw of many thousands of pieces. Only nature knows the full picture, and we will only see that if we can complete the puzzle.
The difficulty we truly struggle with is this. If we try and apply the many names made available to us for plant populations we do know, to further new populations we come across in my field jaunts, we end up nonplussed and confused. In fact sometimes a name applied to some or other population does not seem appropriate even for many of the plants in those same populations.
Essie grew up in Heidelberg so it was just logical that H. heidelbergensis would be the first species he came to contend with. It was described as coming from under one bush just west and practically within Heidelberg itself. What he found there did not match what Smith or anyone else wrote or said. Later he found a second population nearby in which there was nearer agreement at least with the original description. But he also looked at about 10 other populations in the general area and could not imagine how one could apply the criteria of the description to all the variants he was seeing. As originally selected and described, H. heidelbergensis is not even representative of the population from which it came. He had a similar experience with H. turgida that occurs abundantly in and around Heidelberg. It just seemed irrational to formally recognize all the variations. His exploration further east of Heidelberg gradually lead him into a swamp of variables and names that included amongst many more H. heidelbergensis, H. magnifica, H. retusa and H. atrofusca. He could not see or decide where one started or stopped or how the descriptions could possibly explain the situation.
Widening his exploration still further to the east and then west, south and north simply added to his dilemma. Just where did a “species” stop or start? H. dekenahii was a particular problem for him because he saw it change status from a discrete species to a variety of either H. mirabilis, H. pygmaea or H. retusa. He ended up in a situation that he could not give a name to a plant unless he knew where it had originated. This is not something any of us would like to admit to because there are four reasons for the unfortunate truth of the issue. Firstly there is the extraordinary intrinsic genetic variation of the plants themselves, secondly the variation coupled to age and growing conditions (in the field or in cultivation), thirdly the extraordinarily idiosyncratic way in which names are generated and imagined to have meaning, and lastly what we all do have to admit to viz. personal perceptions.
Concluding remarks No formal taxonomic alterations are suggested. The communities and persons involved and interested in these plants need names and we still use names just as if they never disappeared from a formal revision and new classification. Bayer suggested in his very first Handbook (1976) that it is quite unnecessary to consider names as lost just because a taxon is no longer regard as formally valid. It seems foolish to us to expect formal taxonomy to submit to a hobbyist interest, even though it should definitely underlie whatever names are used there.
More important is the idea put forward here that H. retusa and H. mirabilis are a single system that cannot be subdivided into any set of names with equal rank. It does not offer any solution in respect of better delimitation of other species. We have not presented any evidence regarding H. emelyae for example, or how species like H. herbacea and H. reticulata relate to the system we have covered here. The article does not touch on a definite and incredible role that H. floribunda plays as part of the puzzle. Nor on the intricate close relationship this species has with H. chlorocantha, H.variegata and H. parksiana. Furthermore we are both acutely aware that the same complexity is evident among other acknowledged species. So even having a species definition is not resolving the issue of names.
Dr. Davidson has suggested that perhaps the metaphysical element needs to be explored and understood. What this means is that we cannot to continue believing, as “science” intends us to do, that consciousness has evolved in creation. Instead it is creation that has evolved from consciousness. Rather than cite any references to this concept we suggest searching the internet on the topic. The conclusion really seems to us that the systems, organizations and individuals (tasked or self-anointed) to describe and name plant species and their variants, are simply not up to the task. This is largely because of a paradigm of science that is self-limiting.
End note While drafting this article I learned that Davidson has in fact authored a book ”Natural Creation or Natural Selection”. I would have asked “Intentional Creation or Accidental Creation”. Unfortunately too few of us are intellectually up to arguing the subject and it seems to me that our whole society has been dumbed down and fooled into believing that we human beings are the central and most important entities in creation viz. anthropocentrism. There is another belief paradigm that all forms of life have intrinsic value viz. biocentrism. Personally I favour this point of view, even going so far as to say that it underlies this curious preoccupation and brotherhood I feel with Haworthia! There are other authors such as Capra, Sheldrake and Meyer that emphasize a creation that stems from consciousness rather than the converse.
Acknowledgement Both of us have an enormous debt to the incredible number of people who we have touched on our journey and the best we can do is just adopt an attitude of great gratitude for all the interactions great or small, our last hurrah.
Suggested literature Capra, F. The Web of Life, Anchor Books. 1996 Davidson, J. Natural Creation or Natural Selection, Vega Books. 2012 Roads, Michael. Talking with Nature, Journey into Nature, Six Degrees Publishing Group. 2018 Sheldrake, Rupert. A New Science of Life, J.P.Tarcher. 1982 Meyer, Steven. Darwin’s Doubt, Harper One. 2010
Introduction. Apart from the vexing question of species delimitation, there is a truly curious phenomenon in how the species interact spatially and also behaviorally. The authors here believe in a systems approach as the only rational approach to Haworthia. With H. floribunda as part of one system (viz. with H. variegata, H. chlorocantha and H. parksiana), H. retusa is another (viz. with H. turgida, H. mirabilis, H. pygmaea, H. mutica and probably H. emelyae). Within these two systems there is secondary interaction and we are trying to unravel the role of the two prime elements with H. mirabilis (as a secondary in the H. retusa system). Bayer has discussed floribunda in HaworthiaUpdates.org and it is not necessary to repeat that here. In this article we are only dealing with single populations of H. floribunda and H. retusa that occur contiguously at a locality SW of Riversdale where they were located by Essie. H. mirabilis does occur 600m to the south-east as a discrete population in a kaolinite exposure site as opposed to the rocky more stable nature of the H. floribunda/H retusa site. It is important to note two things:
This locality is southwest of multiple discrete populations of H. mirabilis as well as H. retusa, while …
It is the only known occurrence for H. floribunda south of the N2 highway from west of Heidelberg to beyond Albertinia.
Preamble. This is neither a taxonomic and nomenclatural debate nor any attempt to explain how these things came about. It is simply the synthesis of the experience of two field enthusiasts and explorers trying to rationalize what they observe in the field. Trying to make sense of many latin names. It is Essie’s suggestion that H. floribunda and H.retusa interact at a deep level and that H. retusa becomes rougher and darker where the two are in close proximity. They do flower at different seasons with a general and approximate 4 month time barrier. Bayer on the other hand suggests that the two species definitely do produce hybrids despite difference in flowering time (observed in several places as isolated individual hybrids). The apparent roughening and darkening observed in H. retusa is more likely to be consequent to a deeper direct connection with H. mirabilis that may have nothing to do with hybridization. As observed elsewhere, hybridization seems to be a convenient excuse to explain phenomenon that have no other obvious origin. What then happens when H. retusa and H. floribunda virtually share habitat here SW Riversdale.
To understand the situation better we have to look at that H. mirabilis population mentioned to the south (MBB6651). I personally have no photographs because I saw the population before I had a digital camera. But I grew them from seed and the accompanying pictures show the incredible variation that suggests an interaction of the retusoid, the mirabiloid and the floribundoid. Essie made a special excursion to obtain 3 field photographs.
The plants. Multiple photographs of the two species accompany this discussion. H. floribunda can be seen to be H. floribunda with perhaps one notable exception that could be dismissed as a hybrid with the greener coloured H. retusa. The H. retusa plants on the other hand do show some variation. The plants are smaller and generally greener than is the case with other nearby populations.There are indeed some plants on the much darker side and this is where an important distinction may be made between the retusoid and the mirabiloid factions (also spring vs late summer flowering) of a single system. The retusoid system is characterized by light coloration with yellows greens and pinks, while the mirabiloid faction is darker green to blackish green with rusty reds and browns. Two plants are shown from an essentially retusoid population about 33m north.
Conclusion. We do not draw any conclusion from this single location that simply adds fuel to our belief in system and that far less formal names are needed to explain the incredible complexity and high degree of variation. (Note to writer … “Keep it simple, stupid”.) We are further suggesting that all these varied populations are elements of just two complex species systems, and not the fragmented cherry-picked elements that the many Latin names suggest.
Footnote by MBB. It is necessary to write something about Essie Esterhuuizen because whether he knows it or not, he has made an incredible contribution to Haworthia by field exploration that is beyond extraordinary and exceptional. I asked him of he could explain his dedication and commitment to Haworthia and this is what he wrote …
“I was fortunate to have grown up in Heidelberg in the Western Cape. As children we spent most of our time playing in the veld around the town. From a very early stage I was thus exposed to Haworthia and needless to say it formed part of the plants on our rockery. Because of their small size I kept 8 different plants in my so called “collection” which I regarded as a collection of notice. Around 1982 I visited the Karoo Botanical Gardens to see if I could find names for the “small aloes” on the rockery when Bruce at that time the Curator of the garden showed me the garden’s collection. Suddenly I realized that if I want to continue with my hobby – the challenge is humongous.”
Now Essie is Afrikaans speaking and Afrikaans is a language very dear to me because it is what I refer to as a heartfelt language – a language of real people and not like English that is used and abused by many cultures and castes. So I asked Essie to rather write in his home language and leave it to me to try to extract the really personal essence of his interest and opinions. So translated by me, this is what he says…
“I am not a scientist, a collector, or a nurseryman – just a plain field observer. In my first field excursion involving the name heidelbergensis, I was shocked to find that what was known and written about these plants was just a drop in a bucket. Just judging and identifying plants on the basis of what is written, or in collections, does not reflect what the situation is in the field. On the other hand, to explain what one observes in the field verbally or in writing is extremely difficult. Field observation becomes like a treasure hunt. When you think you have found some sort of answer, you find another clue to lead you on.”
The crux of the matter is that it is extremely difficult to articulate what is meant in this last sentence. By putting our heads together we agree that there is pattern in this very complex system, as we have just explained in the article “Last Hurrah”. We have planned to collaborate in presenting the evidence for the continuity we say exists within and between two species systems. Essie’s final sentence reads …”For me my observations will never come to an end … the challenges are too great, continuous and wide ranging”.
Fig. 1 H. mirabilis 6651JME SW RiversdaleFig. 2 H. mirabilis 6651JME SW RiversdaleFig. 3 H. mirabilis 6651JME SW RiversdaleFig. 4 H. mirabilis 6651JME SW RiversdaleFig. 5 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 6 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 7 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 8 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 9 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 10 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 11 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 12 H. mirabilis 6651 (cult. exSW Riversdale)
A note on a H. floribunda and H. retusa interaction was written by Bruce Bayer based on the opinions, observations, and exploration of Essie Esterhuizen.