Seven chapters in four parts:
Chapter 1 – Haworthia retusa ‘nigra’ – Another grand finale
Chapter 2 – Further exploration in Haworthia. Further to finale
Chapter 3 – A field trip to the Potberg area
Chapter 4 – What is typical Haworthia mutica?
Chapter 5 – Still more Haworthia mutica and Haworthia mirabilis
Chapter 6 – Field trip to Van Reenes Crest and Niekerkshek
Chapter 7 – More on Haworthia mirabilis and H. mutica from east of Bredasdorp
Bruce Bayer’s Haworthia Update Essays on Haworthia Volume 7 is A4 size, printed on A3 paper, folded and stapled. Part 1. has 63 A4 pages, Part 2. 83, Part 3. 57 and Part 4. 48. The field work is profusely illustrated with plant photographs, maps and a pie chart. Part 1 has over 330 photos, Part 2 over 280, Part 3 over 330 and Part 4. over 160. Recommended retail price is £45.50 + p & p. It may be ordered from good book sellers.
I wonder. I have written so many words purporting to be my last that my credibility here too must be under stress. Two very recent articles of mine in Alsterworthia deal essentially with that issue, although they also cover the discovery of Haworthia mutica (Buffeljags) (= H. groenewaldii Breuer). They do not cover my subsequent thoughts on actually reading the description of this new “species” by Breuer, Marx and Groenewald. I hope that the present manuscript will explain why I reject this as a Latin binomial although anyone who is in the least familiar with my writing should already know. Spurred on by that discovery, I instigated a search in another area of the Buffeljags valley adjoining the Bontebok Park accompanied by Jannie Groenewald who informed me of what he had found in still another area I had long wanted to explore. So I instigated another search there too and again with Jannie. A discussion of these new finds is submitted to Cactus and SucculentJournal where I trust it will be published. The essence is already in Alsterworthia and this article is written to widen the readership, submit more pictures and maintain continuity with the 6 volumes of HaworthiaUpdate that Harry Mays has been so conscientiously and determinedly publishing. This is all writing that may not otherwise have seen the light of day. I am personally extremely grateful for that as I have had a mania since writing my revision Haworthia Revisited and Update Vol. 1 (both Umdaus), to set the record straight and explore all the unknowns, or at least some of them. So on with Haworthia retusa ‘nigra’.
The second area opened to me by meeting Jannie, was a farm southeast of Tradouw Pass, Heuningklip. In the Tradouw Pass, H. retusa ’turgida’ is found on the steep cliffs at the southern end. The road has been rebuilt since I first saw the plants and the population is greatly reduced from what it was. I simply do not have a picture handy to show what those plants are like, but that is not the need here and it does appear in Haworthia Revisited. Immediately south of the pass is another population of plants that puzzled me from the moment I saw them until now. In Haworthia Revisited there is a picture of one clone under J.D. Kobus Venter’s number 93/35. It is named as H. magnifica var. magnifica. The plants are present in a tension zone between Shale and River gravel, and vegetationally between what was Karoo Valley Bushveld and grassy Renosterveld. In April 1997 I collected seed and grew countless seedlings from there, which were all different. Only one picture, as MBB6666) is included in my long treatise on what I discussed under the title of “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.That picture is also under the name H. magnifica var. magnifica but it was not intended to stay that way. It must be noted that my use of names in the Update volumes was not in a decision-making process so much as a learning and informative one. I read my conclusion to the treatise and note that I made no formal name changes. It is only as further field exploration yielded more and more information that I came to realize that H. magnifica and H. mirabilis, classification-wise, are inseparable. The Tradouw collection still worried me because of the close proximity of H. retusa ‘turgida’. These plants at Tradouw and all the things I referred to as H. mutica ‘nigra’ are in fact better understood as variants of H. retusa in the complexity of its relation to H. mirabilis.
This article thus covers exploration of an area between Swellendam and Heidelberg that I have neglected for too long. It is particularly applied to the problems of classification and naming that have persistently clouded my life and my relations with other writers. But as importantly to me is the unclouding of the way taxonomy and classification is useful and necessary for those interested at all in plants and how it is viewed by them.
Consideration and use of formal names Here I need to explain the naming style I now adopt. I have dropped the use of any rank below that of the species name. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable herbarium specimen that is used to anchor the Latin name. The scientific ranks are aggregates and the terms variety and forma are so too. There is simply too much variation between populations and within populations to enable the certain identification of almost any plant that the binomial system suggests. There are no clear entities and ranks and this article should demonstrate that. There is a separate code of nomenclature that covers cultivars. Note that I do not drop names. I only drop the suggestion of status that additional names have, be they single clones or population references for all the different plants within them. I have used names in Haworthia for 70 years and intensively for the last 40 years and I am very familiar with the difficulties of any system. For the field and all the populations and plants I have seen, I can do no better than offer the list of names I updated in February 2009 in Haworthia Update 5.2:192. This, and the set of Updates, comprises a near total overview of the genus and effectively a revision. The attempted revisions by others fall short for me because they do not cover the range of populations that I have seen nor what history tells me, although they may include populations not seen by me. That is not strange because the area to explore is still vast. Another problem with even trying to be formal, is that which G.G. Smith encountered (Mrs. L. Bolus did too) where several people were generating new names at the same time and trying to keep track formally with concurrent name changes would not have been, is not, possible. The point I should make is that in the last ten years, my field work has confirmed the predictive nature of my classification and these latest finds are in extraordinary agreement with that kind of expectation. What I have found essentially agrees with what I would expect. This is dramatically so for H. mirabilis based on a find by J. Jelimicky at Sandhoogte midway geographically and visually between H. mirabilis ‘bobii’ Hayashi (my nom. nuda ‘pilosa’, ’velcro’, Steven Hammer’s ‘Shaggy dog’) along the Breede river, and a collection of mine further south in the DeHoop reserve, MBB7886 H. mirabilis, discussed (p11) and illustrated (p33) in Haworthia Update 5.2.
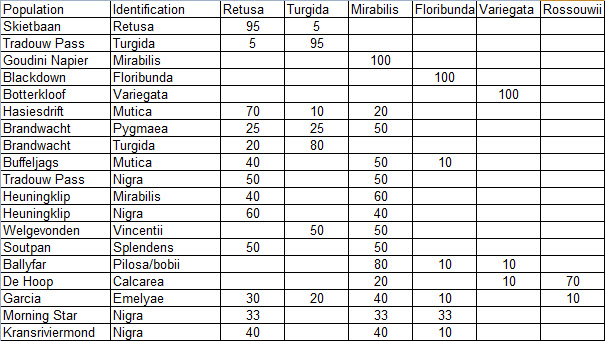
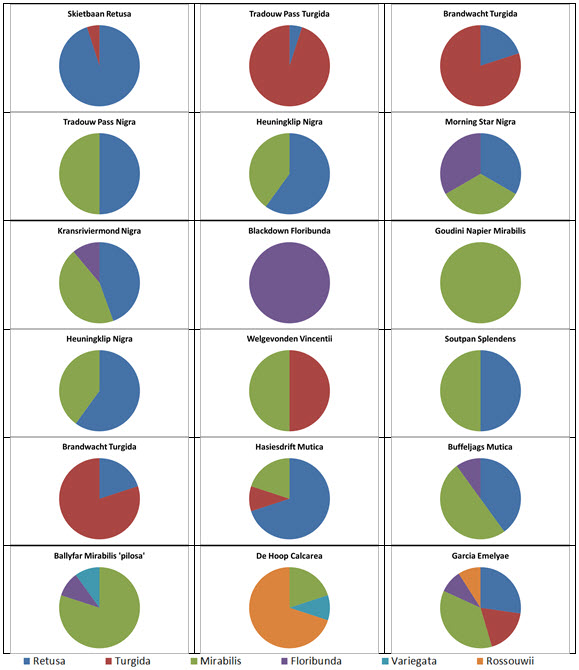
Prior to extending exploration in the Buffeljags area, I had also been down to the Haworthia populations in the lower Breede River area. This is where H. mirabilis seems to assume a wide range of forms and H. retusa ‘turgida’ and H. variegata’ are also present. I also, in the company of Kobus Venter and Lawrence Loucka, revisited Kransriviermond and Morning Star south of Heidelberg where H. floribunda is in the mix with H. retusa ‘turgida’ and H. mirabilis (in my broad new sense that includes H. maraisii,H. magnifica and H. heidelbergensis). Thus many of those images were very fresh in my mind when I went to Heuningklip southeast of Tradouw. Looking at all the plants and in editing my pictures I was truly impressed with the reality of continuity. The essence of H. retusa and H. mirabilis is in all those plants. A mobius strip is a strip of paper twisted and joined at the ends to produce a shape that only has one surface. What we have in the Southern Cape, and covered or touched on in this article, are interlinking mobius strips with one main surface as well as loose ends that present identifiable end-points. I tried once to illustrate the continuities between populations with single or multiple lines joining the different populations to express degrees of similarity. I can also visualize a pie-chart in which each population is represented by a circular chart with the contribution of main elements drawn as slices proportional to the strength of contribution. Table 1 is a schematic and very crudely suggested enumeration for such a set of charts. I have put the elements retusa and turgida as separate under an H.retusa heading for no other reason that I want to emphasize turgida as a variant at the two geographic extremes. One is eastwards where H.retusa ‘retusa’ is absent (Brandwacht, Mossel Bay) and one westward where it is present (in mutica, Hasiesdrift, Bredasdorp). H. retusa ‘turgida’ is present both east and west. But there are many populations contributing to the H. retusa pie in all the ranges of the two parts, from 0-50% thus making up the continuity from retusa to turgida.
Consideration and characters.
a. The pie chart Measuring the similarity of any two plants is extraordinarily difficult. Doing this for a population is even more so. Doing so for many populations, including those unseen in order to fulfill the predictive requirement of a proper hypothesis, is next to impossible. Hence all the disputation and name abuse. In table 1, I speculate an arrangement to illustrate the problem. The pie is constructed from the figures in the chart for just that set of genetic characters that are directly relevant to the set of plants in question. The pie is incredibly big and it should be understood that there must surely be pie-parts that spread their influence beyond the restrictions of the slices I conceive. So my table is restricted to the few species that are immediately relevant in respect of geographic association and direct interaction (by possible cross-pollination?). A pie-chart is drawn for each individual population. In the chart I assume that there is a set of genetic information that programs for H. retusa ‘retusa’. I have also suggested a different set for H. retusa ‘turgida’. If one considers only these two elements and all the known populations, my suggestion is that there is a set of charts that moves smoothly from one set to the next with no interruption. There is no discontinuity and the sets average 50:50 for the information. Thus it is one species and H. turgida is noted as H. retusa ‘turgida’.
Table 1. A schematic distribution of similarity
Table 2 is the complete set of pie-charts that should convey the possible relationships of the different populations and from which can deduce and understand to what degree H. retusa and H. mirabilis can be related. Also it will indicate the similarities between, and the difficulties of, trying to explain H. pygmaea in the east and H. mutica in the west. Both arise from the same genetic source. The problem is compounded by the infiltration of H. floribunda in the area midway between Heidelberg and Swellendam with the re-emergence of H. floribunda in the south at Potberg. H, variegata also enters the picture but from the south and also in the Potberg area. H. emelyae is the very probable combined entity entering the little Karoo. H. rossouwii is almost independent except in the very south-west at Bredasdorp and it may possibly link to other elements such as H. herbacea and H. reticulata.
Table 2. Pie Charts
At Heidelberg there is interaction between an element that is a 50/50 retusa/turgida with a variant of H. mirabilis cf ‘heidelbergensis’. The latter name suggests that H. mirabilis is the general name for all the plants of that species, and ‘cf’ is an abbreviation for ‘compare with’ ‘heidelbergensis’ to refer to the small plants in populations previously regarded as independent species.
b. What really constitutes the pie The pie is the totality of the genetic material, the DNA that determines all life forms. It is the genes, the materials of characters and inheritance, and the repository of all genetic information. The pie is very large indeed and it is said that the DNA pie of an elephant only differs by 3% from that of a mouse; that of a man by 1% from a chimpanzee. In Haworthia one species from another ‘species’, I could guess at 0.001% or less regardless of whose concept I follow. This difference does also not reside in one consolidated slice that can be reduced to a single small pie as I have done. In DNA analysis, certain segments of DNA from cellular chloroplasts, mitochondria or the chromosomes themselves are isolated and the sequence of the paired amino-acid is analyzed. It requires a formidable statistical process to arrive at a two-dimensional tree (phyllogram) that illustrates a branching relationship with a measure of branch separation and lengths. This can perhaps also be used to generate pie-charts with an innumerable number of slices. I personally consider the phyllogram generated in the DNA sequencing process grossly inadequate.
c.) The characters This is extraordinarily difficult to specify no matter how obviously the plants seem to differ from one another. Number, size, shape, colour, arrangement, surfaces, spination and attitude of the leaves in the spiral rosette constitute the main things that determine how we see the plants. These are influenced by growing medium and growing conditions so these are complicating factors. Flowers are used in the same way as the leaves but far less so because they are so similar in all the species. Flowers are also not always present either which reduces their usage. Time of flowering is a major factor because it is linked to the reproductive process. It is also difficult to capture any firm picture of the flower because of the aging process in the 3-5 day life of the individual flower.
d.) The relevance here It simply is not necessary to parade the obviousness of this and the way in which will be attempted anyway in the discussion below. It is flowering time that may be critical beyond, or linked, to the facts of spatial geographic relationships of populations. It is traditionally flower structure that has formed the basis of classification of plants because of the reproductive significance and the role of reproduction in distribution of genetic material between plants and populations. Pollination of Haworthia seems to be primarily by a Solitary bee species. These bees make individual nests and are not communal like the Honey bee. They make tunnels in the soil as nests during the summer months. I have observed them as adults in the winter but then they are less numerous and less active. Few Haworthia flower at the height of winter and do so in two main time frames viz. spring and late summer. The implications are that breeding between isolated populations apart from any other consideration of compatibility, is a question of distance and time. A nest bound Honey bee can forage to 13km from its nest. A free flying Solitary bee can be speculated to cover a considerably greater distance. All these things play on the frequency and power of inheritance. Genetic drift is acknowledged as the genetic movement within a population away from the norm for a species because of isolation, while genetic flow is perhaps the holding together of similarity over geographic distance by a travelling pollinator. Thus the statistical probabilities of breeding across distance are impossible to derive.
I cannot quantify any of these things nor suggest any greater knowledge but this suggests to me, given the freedom with which the various species hybridize in cultivation and even in the field, we need to consider geographic relationships. In my extensive experience with other genera, it appears to me that a key consideration is if things grow together and maintain their difference, or not. Haworthia species are invariably in small isolated populations and seldom do different species grow intermixed. They are also influenced by geology of the substrate and particularly so because they favour the lowered competition from more vigorous and larger plants on the rocky sites or shallow soils they favour. That can be stated in the converse, Haworthia are favoured by conditions poor for other plants.
e.) Flowering time and growing together. The existence as discrete species can be judged from the way in which populations share geographic space. I discussed this in some detail in Update 2.1:76. If two sets of plants grow close together and flower at the same time, while still maintaining obvious group differences, then they are most probably different species. If they flower at different times while still maintaining difference then the question of being different species is taken as certain. If the groups are separated and flower at different times while the differences are small and apparently within the experienced range of variation observed between populations, then it may be problematic. One would need to resort to the extended distribution and variability of the groups in question. Generally in Haworthia, species belonging in the same sub-genus do not share habitat and this is indicative of fairly rapid absorption of introduced genetic material (by pollinators). Hand-pollination and the ease with which plants of different species hybridize in cultivation suggest that there is practically no breeding isolation between them. In the following discussion, use is thus made of general distribution of species and proximity of populations in assessing to which species they best belong. It should be borne in mind that variability may be clinal. This means that the plants change along a geographic gradient. The direction of the gradient may also not be of increasing distance i.e. linear. It may also be circular so that ends of a gradient may meet again as two populations that appear wholly different, but linked by an extended set of other populations. A much more complex situation may actually exist where these clinal gradients may be multidirectional and thus involve more than one apparent species. My observation is that populations do differ and differences follow a pattern that becomes predictable i.e. gradual genetic difference over geographic area. This is not just a result of environmental gradient, but also linked to the question of genetic drift (the tendency of isolated populations) to move away from the general norm. It also results from the genetic flow between populations where the change of genetic material between populations at the distribution extremes is only through other populations.
f.) The species definition This is a subject I have covered many times. My view is that species are dynamic chaotic systems oscillating between different character states with time and with different rates of mutational change. Closely related species may not be recognizable on the basis of single character states and the spatial geographical distributions and local juxtapositions of groups of plants may be the only way to understand what the situation is and so make sensible decisions.
g.) Habitat geology I contend that species richness in South Africa is largely due to its varied geology, skeletal soils and comparatively mild climate. This present discussion deals with species that are associated with a geological phenomenon that is the Tertiary deposits of the Southern Cape. I am no geologist and am required to make the pretention I am one to explain why plants are distributed the way they are and what influences difference among them. In this case the vegetation at Buffeljags, Swellendam has been described (by Breuer et al. cited below) as on silcrete, and the general vegetation as by vegetation scientists as Ruens Silcrete Renosterveld. But there are two problems. The first is that vegetation classification is not infallible and I have never found it really useful in dealing with a wide range of species that occupy skeletal habitats. Secondly the substrate geology is definitely not simply silcrete nor is the clay soil either simply bentonite or kaolinite. There are many problems in the classification of clays. Bentonite in common terms seems to be the yellowish clay mined in large quantity from Heidelberg eastwards and there is a transition to ochre bearing clay. Bentonite may be aluminium, potassium, sodium or calcium dominated. As a montmorillomite clay it should have expansive cracking properties. Kaolinite is popularly regarded as the very white clay that is commonly seen under the rocky top layer of the sharp inselbergs that rise above the wheatfields west of Heidelberg. It seems to be less mineral dominated clay and does not have the expansive/contractive properties of bentonite. The distinction between ferricrete and silcrete seems to be that in ferricrete, iron oxides are infused into the petrified water deposited layers. Both the kaolinitic and bentonitic clays seem to be derived from the weathering of Bokkeveld shale that underlies the silcerte/ferricrete layer. It does not require any stretch of the imagination to see now why any one inselberg may be a little different from the next, and sometimes a lot different. What we do not know is how rapidly the landscape has moved from a broad white silcrete/ferricrete underwater plain to the remnant condition of that rocky plain today. Neither do we know what it has meant for the plant species that are linked to, and dependent on, the habitats generated in the erosion processes.
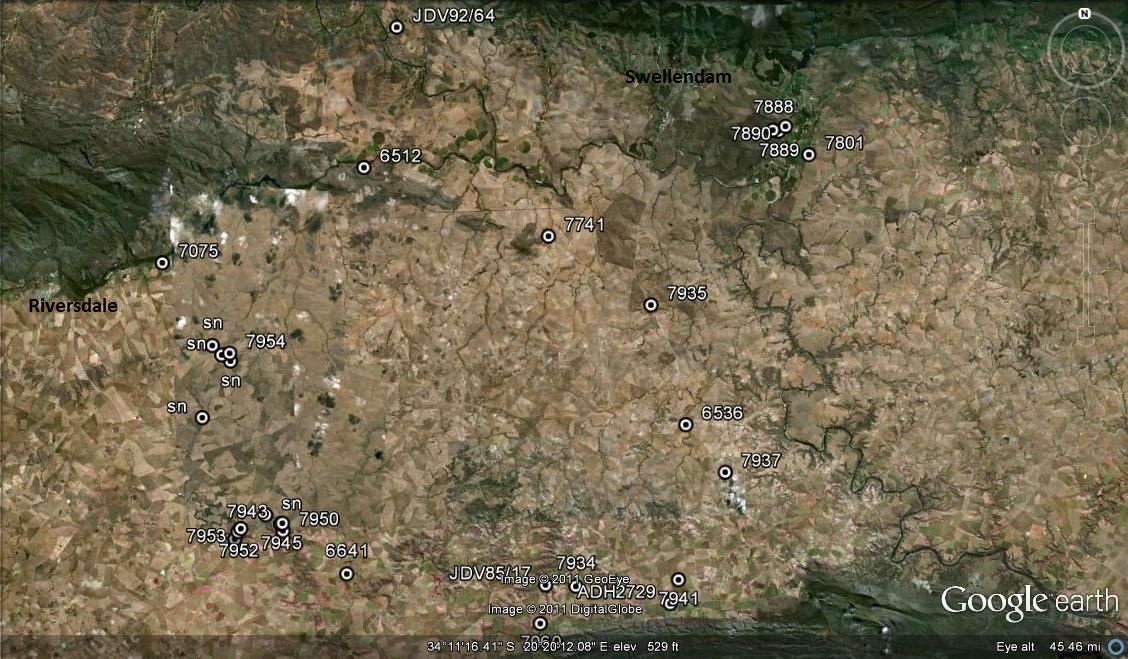
Map of Swellendam to Suurbraak area.
Map Legend – east of Swellendam.
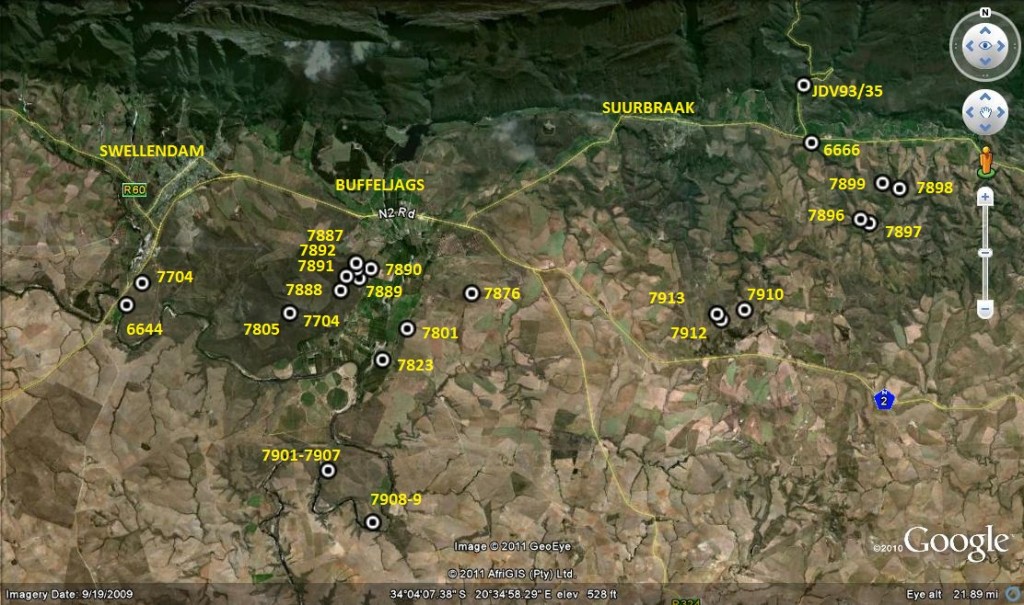
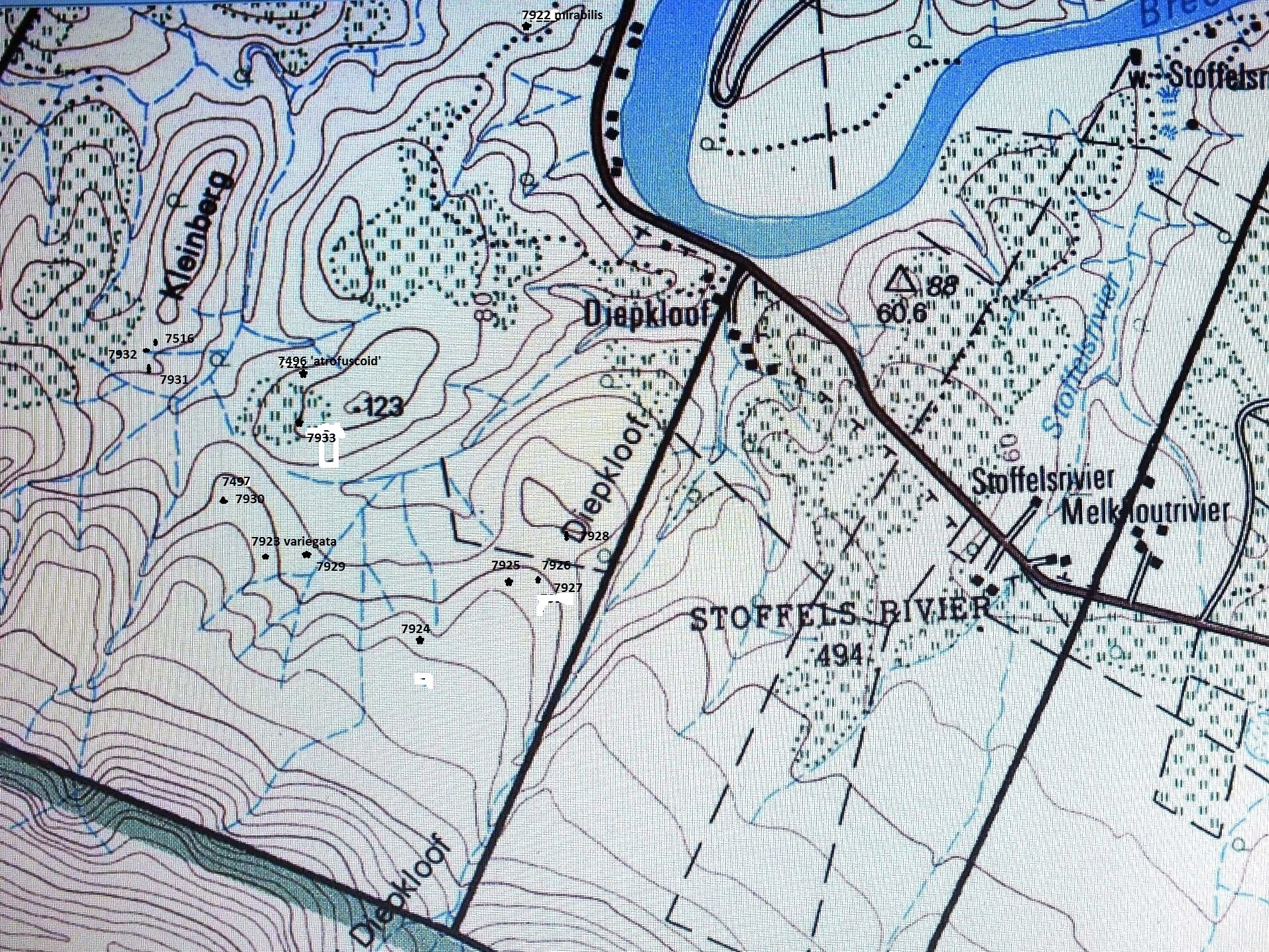
1. MBB7899 H. retusa ‘nigra’. Heuningklip. 2. MBB6666 H. retusa ‘nigra’ ↔ H. mirabilis. S Tradouw Pass. 3. MBB7896 H. retusa ‘nigra’. Heuningklip. 4. MBB7897 H. retusa ‘nigra’. Heuningklip. 5. MBB7898 H. retusa ‘nigra’. Heuningklip. 6. MBB7888 H. mutica. Rotterdam. 7. MBB7889 H. mutica. Rotterdam. 8. MBB7890 H. mutica. Rotterdam. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags. 10. MBB7741 H. mutica. Dankbaar. 11. MBB7892 H. marginata. Rotterdam. 12. MBB7891 H. minima. Rotterdam. 13. MBB6644 H. mirabilis. SW Swellendam. 14. MBB7704 H. mirabilis. Bontebok Park. 15. MBB7805 H. mirabilis. Bontebok Park. 16. MBB7823 H. mirabilis. Klipbult. 17. MBB7887 H. mirabilis. Rotterdam. 18. MBB7876 H. mirabilis. Disselfontein. 19. MBB7901 H. mirabilis. Crodini E. 20. MBB7902 H. mirabilis. Crodini E. 21. MBB7903 H. mirabilis. Crodini E. 22. MBB7904 H. mirabilis. Crodini E. 23. MBB7905 H. mirabilis. Crodini E. 24. MBB7906 H. mirabilis. Crodini E. 25. MBB7907 H. mirabilis. Crodini E. 26. MBB7908 H. mirabilis. Sandkraal W. 27. MBB7909 H. mirabilis. Sandkraal W. 28. MBB7910 H. floribunda. Rietkuil. 29. MBB7912 H. mirabilis. Rietkuil. 30. MBB7913 H. mirabilis. Rietkuil. 31. JDV93/35 H. retusa ‘turgda’. Tradouw Pass.
The populations
Set 1 – THE TRADOUW PASS AREA (H. retusa ‘nigra’) This first section deals with populations that complement those in the long discussion under the title “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.There I ended with the words … ”The habitat photo (of H. mutica Hasiesdrift) raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.” Added were these words … ”There the saga for the moment must rest.” What I had in mind was exploration that no one seems very anxious to do. This has now been done and I would lay the matter to rest as follows:
1. MBB7899. H. mirabilis, Heuningklip. See figs 1-30 Fig. 1 is a scenic view of the area to show the countryside and the type of inselberg the plants are often found on. Fig. 2 is a closer picture of the erodible clay held in place by grass clumps at the base of which the plants often are. Occasionally they are simply out in the open sun. I term these places pressure bursts because water build-up in the soil causes collapse of the small banks and erosion can be quite fast. Several times I have found plants lying in collapsed soil. Such a soil bank near Heidelberg has retreated by nearly 2 meters in about 15 years albeit accelerated by road maintenance, but it does relate to any attempt to explain how much suitable habitat may have disappeared with time.
Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – bFig. 2. A photograph of the habitat of MBB7899.
Figs 3 to 30 is a selection of plants at the site where one is almost compelled to examine each plant separately and easily generate another 28 practically different images. Fig. 3 is of a plant with the surfaces roughened by small tubercles. SEM (scanning electron microscope) images of similar leaf surface often show incipient tiny spines on the tubercles and they appear as spines or hairs in populations like H. mirabilis ‘pilosa’ at Ballyfar, or H. mirabilis at Windsor Riversdale. This surface roughness and all its degrees is fairly characteristic of H. mirabilis but is does occur in H. retusa too. Plants illustrated in Figs 4 and 16 has one muttering Kruisrivier and Komserante, northeast and east of Riversdale respectively, and which I would have allotted to H. magnifica in the early period of my exploration no doubt have Breuer and Hayashi names. Now I simply say H. mirabilis and add the locality to place the plant to at least give it a geographic reality that is better than using the formal Breuer/Hayashi in hanging commas as I also suggest. Fig. 6 is apparently a seedling and demonstrates the leaf-shape of H. floribunda that led me to suggest that H. floribunda is a perpetuated juvenile form. Of course the evidence suggests to me that H. floribunda coalesces with H. mirabilis in this wider area although it does appear at Swellendam to the west of my map as the large form and at the extreme SE corner of the same map as a convincingly normal form. Fig. 9 demonstrates longer pointed leaves and Fig. 12 short more obtuse leaves. Fig. 17 is of a plant that has relatively smooth leaf surfaces rather like H. retusa and the leaves are conspicuously lined. In Fig. 20 there is a suggestion of muticate (without a point) leaves. Fig. 22 is again quite strongly lined. Fig. 22 shows a leaf with a single line. Fig. 25 demonstrates what is termed rotate leaves where the points of the leaves point around as though to encircle the plant. Fig. 28 made me recall plants of H. mirabilis at Melkhoutrivier in the south west near Malgas and further to Ballyfar. The flowering time is late summer.
Fig. 3. MBB7896 H. retusa ‘nigra’. Heuningklip.Fig. 4. MBB7897 H. retusa ‘nigra’. Heuningklip.Fig. 5. MBB7898 H. retusa ‘nigra’. Heuningklip.Fig. 6. MBB7888 H. mutica. Rotterdam.Fig. 7. MBB7889 H. mutica. Rotterdam.Fig. 8. MBB7890 H. mutica. Rotterdam.Fig. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags.Fig. 10. MBB7741 H. mutica. Dankbaar.Fig. 11. MBB7892 H. marginata. Rotterdam.Fig. 12. MBB7891 H. minima. Rotterdam.Fig. 13. 13. MBB6644 H. mirabilis. SW Swellendam.Fig. 14. MBB7704 H. mirabilis. Bontebok Park.Fig. 15. MBB7805 H. mirabilis. Bontebok Park.Fig. 16. MBB7823 H. mirabilis. Klipbult.Fig. 17. MBB7887 H. mirabilis. Rotterdam.Fig. 18. MBB7876 H. mirabilis. Disselfontein.Fig. 19. MBB7901 H. mirabilis. Crodini E.Fig. 20. MBB7902 H. mirabilis. Crodini E.Fig. 21. MBB7903 H. mirabilis. Crodini E.Fig. 22. MBB7904 H. mirabilis. Crodini E.Fig. 23. MBB7905 H. mirabilis. Crodini E.Fig. 24. MBB7906 H. mirabilis. Crodini E.Fig. 25. MBB7907 H. mirabilis. Crodini E.Fig. 26. MBB7908 H. mirabilis. Sandkraal W.Fig. 27. MBB7909 H. mirabilis. Sandkraal W.Fig. 28. MBB7910 H. floribunda. Rietkuil.Fig. 29. MBB7912 H. mirabilis. Rietkuil.Fig. 30. MBB7913 H. mirabilis. Rietkuil.
These images can now be compared with MBB6666 at Tradouw Pass to establish if my assessment is reasonable. My contention is that this MBB7899 population is midway between the other three Heuningklip populations I am about to illustrate and Tradouw Pass. This similarity is repeated widely and eventually contributes to my contention that H. mutica has been generated by drift away from its two contributing parent species viz. H. mirabilis and H. retusa.
2. MBB6666 H. mirabilis. Tradouw Pass See figs 31-71. This population is at the mouth of the Tradouw Pass and relatively low-lying in respect of the previous population. The habitat is quite different being partially coarse river gravels, weathered shale and at one point is an un-weathered shale ridge where the plants are wedged in the cracks. The habitat is of course very different to the steep sandstone cliffs within the pass itself where H. retusa ‘turgida’ occurs. I initially thought these plants fitted with H. magnifica. A decision derived from a very small sample available at the time. Now I looked for the images and photographs in my records under H. mirabilis where they should have been since my inclusion there of ‘magnifica’, and also under H. retusa where I found them in the folder ‘nigra’. The flowering time is late summer. In this respect similar to the previous set MBB7899 H. mirabilis but the plants are more robust. Figs 31-45 were taken in 2009; figs 46-53 in 2011 and figs 54-71 are from seedlings grown by myself. I tried to reduce the pictures and eliminated a few. But I really think they all need to be presented to show again how serious the difficulty of circumscribing them as a single recognizable key-able entity really is. While looking at my pictures I seriously thought that the set taken in 2011 differed from those taken in 2009. The cultivated plants of course are not directly connected to their source and the variation is self-evidently wide. But it is equally so in field plants. There are the smoother surfaces of H. retusa and some very rough surfaces for H. mirabilis. My decision became that they fitted best with the ‘nigra’ populations where my organizing had taken them despite flowering time, but here MBB7899 shows absolute geographic position between H. mirabilis and MBB6666. I consider this key evidence for the basic continuity for H. retusa and H. mirabilis that I have conjectured elsewhere and I would not be able to defend an argument that MBB7889 plants belong better in H. mirabilis and this means MBB6666 belongs there too. I would simply dispute that there is no better solution so stick with history as far as possible and that means H. mirabilis. If Breuer or Hayashi have a name for a variant from this population intended as an aggregate name, I recommend the use in inverted commas. I would actually like the use of the equilibrium diacritic of a double arrow pointing between two names viz. H. mirabilis↔H. retusa ‘nigra’, although this available symbol is not quite the thing.
Fig. 31 6666 H. retusa ‘nigra’
I come to this vexing question of “typical” that is bandied about even by myself when I liken population to population. There is the same range of variables in MBB6666 noted for MBB7899, but the plants do not darken as much and they are larger. So we need now to look at the other three Heuningklip populations asking what significance is attached to their presence in exactly the same habitat as MBB7899, but geographically separated and by perhaps 500m and 2km respectively. The plants are so variable that it is not possible to arrive at a characterization that separates any two plants, or any two populations in any geographic set for that matter.
3. MBB7896 H. retusa ‘nigra’, Heuningklip 1 See figs 72-89. The plants in this and the next two populations were flowering in September as does H. retusa and they are relatively large, green in colour and generally smooth. Of course both vary enormously due to both intrinsic genetic difference and extrinsic local situation. One plant within an older decaying grass tuft was nearly 200mm diameter, and a healthy green colour, another plant in the open white clay was less than 50mm diam. and dark in colouration.
Fig. 72 7896 H. retusa ‘nigra’
4. MBB7897 H. retusa ‘nigra’, Heuningklip 2 See figs 90-105. The first two pictures show the general landscape and the local habitat where the plants occur. The clay is highly erodible and often there are no plants at all in the clay. Some of the plants have very acuminate pointed leaves while in others they may be relatively obtuse and rounded.
Fig. 90 The countryside looking towards the Tradouw Pass.Fig. 91 The ‘white clay pressure burst’
5. MBB7898 H. retusa ‘nigra’, Heuningklip 3 See figs 106 -132. The great variability is again evident and in this population the plants were observed to be generally without much venation or lines as in fig.112. Then one plant, fig. 119, was so striped as to stand out from all the populations seen.
Fig. 106 Silcrete hilltopsFig. 107 7898 H. retusa ‘nigra’.
Set 2 – THE BUFFELJAGS AREA (H. mutica) This second section follows a recent manuscript submitted to Alsterworthia yet to be published entitled “Some last closing thoughts” written in the hope of finding the closure I seek and that will not come. That article is about a population that has been named as a species viz H. groenewaldii, in an article authored by Breuer, Marx and Groenewald (Alsterworthia 11.2:15) that I only saw after the submission of my manuscript. I also only saw the article after the exploration I would have asked for to verify opinions. The description seems to have been prompted by Marx who refers to similarities to H. springbokvlakensis and to H. magnifia var. atrofusca and raises many issues that my manuscript does not cover. I simply find the whole discussion unrealistic and especially so when no further exploration was done to examine anything. One should refer to Breuer’s World of Haworthias Vol 1 to see that the naive promises in the foreword and introduction of exploration and field study, was never achieved. It is true that the plants have an unusual surface texture and it is also true that the habitat is quite unlike any other H. mutica occupies. But it is Marx’s comment that disturbs me viz. “Unfortunately Bruce Bayer keeps on refusing to see it as something new and calls it simply H. mutica ‘Buffeljags”. This is a distortion and misrepresentation, and whose misfortune this is, not specified. My reticence is reflected in the preceding discussion and Gerhard does not seem to follow, or agree with, what I have been writing about and trying to explain in this general context for so many years. In fact in an unpublished (to my knowledge) manuscript he dismisses all my writing as “pretentious” that no doubt it is, because I have never fancied my self as a taxonomist in the tradition of taxonomists. I was as fascinated as Marx by the plants and am even more anxious than he is to see just how they connect to the probable geographic counterparts in the context of the species definition that I apply. I was also very curious to know what the connection to H. mutica and the H. retusa ‘nigra’ issue, only now laid to rest in the preceding discussion, actually might be here. This is why I use the word “instigated” in reference to exploration in the very valley where H. mutica ‘groenewaldii’ occurs and to the obvious habitat for similar plants. It is surely unfair iniquitous to so demand that I see this population as “something new” in an article that rather disregards the history of the subject and what I have been trying to understand of it.
I also recently submitted a manuscript to Cactus and Succulent Journal (US) where I wrote of “H. groenewaldii Breuer”… “I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February/March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time”. I have in many places discussed the problem of “hybridization” and it must surely be well-known from my writing that H. floribunda merges into H. mirabilis. Less well-known will be the fact that evidence for H. variegata doing the same is also available. In respect of floral characters, I wrote a chapter in Haworthia Update Vol.3.2:88 explaining the problems of using floral morphology for species recognition. I would challenge Breuer et al. to produce comparative results from studies of the flowers in any of these problematic species to demonstrate difference. Regarding the curious shiny leaf surface, I refer to Haworthia Update Vol.2.2:141, Chapter 15 where I discuss electron scanning photographs of leaf surfaces. This also demonstrates how problematic leaf surface is. It would be tiresome indeed to parade all the evidence in support of these contentions of mine. In Haworthia Update 5.2:108 I also briefly discussed H. mutica from Buffeljags (‘groenewaldii’) and submitted a large number of photographs explaining the habitat and what the possible connection was to the H. retusa ‘nigra’ conundrum. I may also have written about the issue in Haworthiad. Suffice to say that the authors of H. mutica ‘groenewaldii have certainly not checked or considered “all the facts” as the principal author claims. The very opposite seems to be the case.
Considering my oft repeated observation that geological substrate plays a major role in shaping species appearances, the Breuer, Marx and Groenewald description of the habitat for H. mutica ‘groenewaldii’ is a little misleading (see considerations above). Nowhere else is H. mutica on identical substrate and this would just support my contention of ecotypic differentiation that does not justify a Latin binomial. There is considerable doubt about the actual difference between these river boulder situations and the widely distributed terrace gravels elsewhere in the area, notably the Bontebok Park near where the following populations were found:
6. MBB7888 H. mutica, Rotterdam 1 See figs 133-169.
Fig. 133 7888 H. mutica.
7. MBB7889 H. mutica, Rotterdam 2 See figs 170-188.
Fig. 170 7889 H. mutica.Fig. 185
8. MBB7890 H. mutica, Rotterdam 3 See figs 189-191 The three populations above are separated spatially by several hundred meters and they are all on remnant river gravels on a more gently sloping bank than the Buffeljags east bank. The vegetation is closer to conventional fynbos as in the adjoining Bontebok Park, but grassy. The pictures are self-explanatory and the leaf surfaces of the plants are also generally as for the Buffeljags population. What is significant is that I cannot say I saw a single plant in these populations that resembles the one chosen and pictured as typical for this highly dubious species, H. groenewaldii. It makes more sense to me to identify plants from these populations by collecting number than to generate a supposedly descriptive Latin epithet from an imagined typical representative.
Fig. 189 7890 H. mutica.
9. MBB7801 H. mutica ‘groenewaldii’, Mullershof, Buffeljags See figs 192-215 The following set of photographs has surely seen the light of day somewhere and I repeat them because as far as I am concerned they are H. mutica with the shiny surface claimed for “groenewaldii’. Dankbaar is some 16km west of these Buffeljags populations and the substrate is Table Mountain Sandstone. The flowering time is just normal for H. mutica. Fig. 215 is a photograph of a plant apparently collected by Jannie Groenewald and it is quite uncharacteristic of the population and would suggest that it is a product of hybridization with H. mirabilis in the vicinity (e.g. Klipbult).
Fig. 192 7801 H. mutica ‘groenewaldii’.Fig. 215 7801 H. mutica ‘groenewaldii’.
10. MBB7741 H. mutica, DankbaarSee figs 216-235. My observations in respect of these two sets of plants confirms my widely written convictions about Haworthia taxonomy and the state of classification in the absence of a definition for the concept of “species”, and the intrinsic value of Latin binomials for rational and sensible communication. Of course this Dankbaar population is not identical to other H. mutica populations and I am not driven by fixation on any one character that defines this species. It is a geographic set and the various sets are not simply separable. But I am not finished yet.
Fig. 216 7741 H. mutica.
Set 3 – THE BUFFELJAGS AREA (Robustipedunculares) The western Buffeljags area also produced a population of H. marginata and one of a very curious H. minima, that both complement occurrences in the Bontebok Park a little further west. The conventional small, blue-green H. minima is present in the park as well as an unusually green population.
11. MBB7892 H. marginata, Rotterdam See figs 236-243. This population was quite exceptional and one of the strongest populations known to me. But the plants were in flower in September whereas the flowering time for the species is normally late summer; and the flowers and seed capsules exceptionally large. I do not consider that these factors command another Latin binomial.
12. MBB7891 H. minima, Rotterdam See figs 244-252 and Bontebok Park figs 253-4. These were also extraordinary plants, larger than those seen in the Bontebok Park population where the plants were also green viz. figs 253-4. Fig. 252 confuses the issue because, while the other plants seemed alike to H. marginata and distant hybrids, this one plant appears to be a recent hybrid. This occurrence cannot be overlooked in respect of other introgressive (or seemingly introgressive) populations of Robustipedunculares species. The plants were not in flower and again we have a suggestion or direct evidence of hybridization across a difference of seasonal flowering.
Fig. 244 7891 H. minima.Fig. 254 sn H. minima, Bontebok Park
Set 4 – THE GREATER BUFFELJAGS AREA (H. mirabilis) This is a set of 15 populations of H. mirabilis that occur east, west, south north of the H. mutica ‘groenewaldi’ related populations. I exclude from the discussion and illustrations the several populations of H. floribunda ‘major’ populations to the northwest, while I repeat the statement that H. floribunda contributes to the general gene pool of H. mirabilis and together with H. retusa, they all contribute to the gene pool of H. mutica ‘groenewaldii’. The huge presence of populations more readily assignable to H. mirabilis (sensu lato – in the broadest sense) is used by me to validate my opinion that the “mutica” elements draw most genetic resource from the H. retusa element, and hence why I refer those plants to H. mutica.
The sequence of populations 7901-7908 is located in a horseshoe of the Breede River, southwest of Buffeljags in very promising habitat. Only H. mirabilis was seen. I perhaps need to point out that these populations would have been extraordinarily difficult to name under any previous dispensation of mine where I was still trying to uphold H. magnifica,H. maraisii and H. heidelbergensis as species. These I have since merged in H. mirabilis and I reject the introduction of a group name like “aggregate” as Breuer seems to have done, as a complete fabrication that does nothing to solve the problem other than to highlight it.
The following 6 populations are all associated with tertiary and river gravels. There are differences between most of the populations but of an order that I cannot articulate. It is also not possible to really encapsulate the appearances of the plants in the two photographs for each population that I submit. At the same time it does not seem to me possible to make any communicable distinctions.
13. MBB6644 = 6860 H. mirabilis, SSW Swellendam. See figs 255-256.
14. MBB7704 H. mirabilis, Bontebok ParkSee figs 257-258.
15. MBB7805 H. mirabilis, Bontebok Park See figs 259-260.
16. MBB7823 H. mirabilis, KlipbultSee figs 261-262.
On selecting these two pictures I was reminded of my observation that there was a significant H. floribunda appearance to the plants. I had believed that I had seen H. floribunda in the Bontebok Park nearby many years ago but subsequent exploration has only produced H. mirabilis albeit with some similarities to H. floribunda. My reaction was to suspect that H. mutica ‘groenwaldii’ most probably also then needed to be considered as a product influenced by H. floribunda.
17. MBB7887 H. mirabilis, RotterdamSee figs 263-264.
18. MBB7876 H. mirabilis, DisselfonteinSee figs 265-266. These plants were a little more robust than the others in this set and rather similar to MBB7901.
The following populations are almost entirely in Bokkeveld shale but on a very close interface with tertiary gravels. In some places there was also some wind-blown sand.
19. MBB7901 H. mirabilis, Crodini 344, S Swellendam See figs 267-278.
20. MBB7908 H. mirabilis, Sandkraal, S SwellendamSee figs 279-282.
Set 5 – BACK TO TRADOUW PASS – AS IT HAPPENED In the process of compiling this manuscript I was bothered by the ever present question of what is still out there that might influence the outcome. My wife, Daphne, was planning an outing to see chameleons at Swellendam while simultaneously contacting landowners for me around the Potberg. Always anxious to get two birds with one stone, I asked her to find out who owned the land west of Heuningklip that we had just explored with Jannie Groenewald. The result was that we ended up spending a day with Odette Curtis of Custodians of Rare and Endangered Wildlife, south of Suurbraak and finding three populations that, for me, put the proverbial cherry on the pie.
All three were on the slopes of silcrete inselbergs and almost the last of this habitat westward from Heidelberg to Buffeljags. The first inselberg was different to the second and from the Heuningklip habitats because it was dominated by Aloe arborescens and A. ferox. Aloe brevifolia was also present in an unusually small form. The habitat was very impacted by grazing animals and there is an outside possibility that H. retusa ‘nigra’ may once have been present although I personally doubt it. To our surprise we found…
21 MBB7910 H. floribunda, Rietkuil, Suurbraak See figs 283-288.
There were many plants but severely trampled and disturbed. Their survival was obviously aided by the fact that much of the plant is below soil-level. But my experience of this species is that it struggles to maintain its identity in the presence of either H. mirabilis or H. retusa. They never share immediate habitat. There was no hybrid present and this also adds to the problem of interpreting flowering time and distance apart from other species as part of the process. The plants were not the large green variant of the species present to the west at Appelbos, Swellendam; and the leaves were both with and without marginal spines. Eastwards I know the species to be close only at Blackdown north of Heidelberg and at Goedverwagting south of the N highway about 15km east southeast. It definitely seems to be absorbed into H. mirabilis west and north respectively of those two populations. So while this is a slight contradictory element of surprise, it is the next two that really excited me. These were again inselberg habitat but without the strong silcrete upper layer of the former habitat and without the Aloe element. The second ‘population’ was about 200m from the first and comprised four groups of plants along a longer north-facing ridge.
22. MBB7912 H. mirabilis, Rietkuil, Suurbraak See figs 289-306.
23. MBB7913 H. mirabilis, Rietkuil, SuurbraakSee figs 307-331 I am very hard-pressed to say much more than that these plants met all my expectation for the very variable set of plants and populations that I ascribe to the one single species H. mirabilis. Who could possibly articulate the differences of the many populations of this species from each other and from this one? Variable becomes a done-to-death word. The individual plants, in two populations quite unlike any others I have seen, were generally robust with large specimens up to 100mm. diam. The colour is dark, although green when shaded within or under grass. The leaves were usually sharply pointed but there was always evidence of one or two leaves or odd individuals in which the leaves were muticate i.e. without points. In some plants the leaves were notably lined in others not. The colour also varied from chestnut to deep reddish-brown when the plants were more exposed. The surface texture was generally tubercled but there was evidence of smoothness of the leaves in many plants. Flowering time was obviously late summer gauged by the absence of flower at the time of the visit viz. September. Except for one oddity. This was a single plant with a weak flower spike. I photographed this for the useful information the flower-bud provides. Where Breuer et al. claim to have considered all the facts, this is a character that is conveniently overlooked when claims are made for the significance of floral characters when paraded to substantiate yet another claim to species significant difference. The bud-tip in all the Southern Cape species of subgenus Haworthia with the exception of perhaps H. rossouwii have this very obvious “fish-tail” bud tip. The tips of the upper two petals are flattened and spreading to resemble a fish’s tail. It is most marked in H. herbacea and H. reticulata where the bud tip curves downward from the semi-erect position of the body of the flower and then up again. I am fearful that my inability and reluctance to consider these plants as “new” may be the motive for someone to rush forward and apply a Latin binomial.
My opinion is that these two populations mirror the paradox in the east where H. pygmaea ‘fusca’ is nearer to H. retusa ‘retusa’ andwhere H. retusa ‘turgida’ is nearby. H. mirabilis ‘splendens’, in the doubt about its affinities, in my opinion remains in the context of H. mirabilis. This is a conclusion I reached after exploration of the area between Riversdale and Albertinia where the same elements noted in this article repeat themselves. I do not for one moment concede that better explanation is available by resort to a multitude of Latin binomials and by an artifact like “aggregate”.
Conclusion I have been deeply disappointed that so little progress on the classification front has been made among the fraternity writing about Haworthia. In communication with Lawrence Loucka, he wrote …”You might consider saying that a genus revision is needed and that until amateurs depart no proper botanist will make the investment. Until then chaos will rule, names will proliferate.” This is a useful and insightful comment and generally summarizes what I have often said. Unfortunately the position has been rather exacerbated when proper botanists have made their presence known and I can recount many instances to support this statement. My own view has been that botanists have not been able to confront genera that are so publicly and popularly known, and garlanded with names. Thus amateurs, including myself despite at least an iota of formal training in science, simply have a field day generating names like rabbits out of a hat.
Unless some proper thought is given to this whole question of what the names are supposed to mean apart from catalogue items, taxonomy (classification) will never lift out of the disinterest, disrepute and scorn that collectors justifiably seem to have for it. Latin binomials do have a real value and to demean this by one-dimensional derivation and rationalization is intellectually fraudulent. My writing is replete with examples of the variability and interaction of populations over large area. Taking a single population and one flower, and often a single plant, to generate a species description and a Latin binomial is grossly simplistic.
The one change that this article generates is the confirmation that the population MBB6666 from immediately south of Tradouw Pass is better identified by name as an H. mirabilis variant, and not as H. retusa ‘nigra’. MBB7801 at Buffeljags and now also at Rotterdam, remains as H. mutica and the epithet generated by enthusiasts can be added to make the trinomial H. mutica ‘groenewaldii’. For formality, the name for the form illustrated as typical would be H. mutica ‘Groenewaldii’. A few additional names will be needed for similar clones and a lot more for the cultivars that will be derived from this population set.
My conclusions here make no substantial departure from anything that I have observed or written over 50 years. While the subject has been comprehensively covered in the field and in writing, there is a huge amount of exploration that still needs to be done if the genus is to be still better known. My prediction is that the answer will conform very closely to the species-list I finalized in 2009 (see Alsterworthia 5.1:192). The disclaimer is that botanical classification may eventually reach the point where the real nature of species and their purpose comes to be known. In that case what we now may see and persuade ourselves are discrete species, are not. Instead these highly complex chaotic systems may in fact be quantum units of consciousness and whole elements such as some genera and sub-genera we now recognize will be the species.
Acknowledgement I want particularly to acknowledge Jannie Groenewald whose interest and enthusiasm are boundless. Then Allan Jeptha, an environmental law practitioner with great vision for the Suurbraak community; Odette Curtiss, a consultant for the conservation of Renosterveld vegetation; Lawrence Loucka whose patient guidance and comments I value and who generated the set of pie charts; Kobus Venter and Steven Hammer who have always been supportive and contributory; Max Coetzee of Felix Unite and Samie Lategan of Buffalo Breeding Programme for access to Sandkraal West and Crodini East; Jaap Viljoen for his contribution; my wife Daphne, who trusts me and whom I deeply trust. ♦
The writing of my grand finale was inspired by several things. One of these was another item of a mind-numbing foray into the classification of Haworthia. So I asked that deep thinker and observer, Gerhard Marx, for a devil’s advocate (abbrev. DA) point of view which he has done with the same competence he has as an artist. I have many times in my writing addressed the issue of a species definition and produced one too. Not surprisingly the first thing the DA does is dismiss my definition without producing one of his own. Simply being able to say that an indeterminate number of plants from some population are sufficiently different in respect of a character or two from other haworthia, is motivation enough for the generation of a new name?
The case of H. groenewaldii Breuer, described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald is the case in point. It presents the description of this supposed new species from Buffeljags east of Swellendam. The article is written in the first person (Breuer) who quotes extensively from Gerhard’s e-mails, and includes a piece by Jannie Groenewald under the heading “Description of the Vegetation type and distribution”. The overall impression is of an article that conforms to the style of a forgotten era and it is not possible or sensible to attempt a rational dismissal. Who is actually responsible for the article and how does one correct misleading statements without giving offence?
There is probably only one main issue that can be made level-headed sense of. This is the statement in the article…”another (it is not clear what the other was) striking feature was the fine but rough leaf texture…” Then quoted again…”the typical H. mutica grows only 20km to the west at Dankbaar”. But the Dankbaar plants also have the same shiny leaf surface. The word “typical” is generally overused in the article and I would counter claim that the Dankbaar plants are typically H. mutica and reinforce my argument for the similarity to the Buffeljags plants. But this is not the main issue, nor is it the least of many rectifications the article almost demands. In the new-age freedom of communication there are virtually no secrets anymore and this adds to my discomfort because the article only touches on some of the issues clouding Haworthia classification. The DA says I should meet this following argument in support of an H. groenewaldi’…” it is consistently smaller than H. mutica and reminds superficially actually of H. bruynsii or springbokvlakensis with its rounded leaves. Kobus Venter’s feeling was that it is closest to H. magnifica var atrofusca and I agree 100%.” These are subjective statements and I cannot agree about the size as I have seen many H. mutica populations that I do not think supported the opinion. The roundness of the leaves is also subjective. The element H. mirabilis ’atrofusca’ in any case has plants with variably rounded leaves, so which do you mean?. The name ‘mutica’ means leaves without a point and rounded and it is no co-incidence that Col. Scott initially identified H. springbokvlakensis as H. mutica until I pointed out to him that I thought H. mutica was actually the plants (note plural) in the Bredasdorp area. Scott plagiarized my words describing them alongside old wagon tracks that suggested their discovery in earlier times.
DA lists several more indeterminates…”all the unique morphological features like small size, rounded rough-textured and uniquely flecked leaves”. Of course there are not several more as I have already dismissed two and this leaves the third viz. the “uniquely flecked leaves”. Firstly not all the plants have flecked leaves and I have shown that these occur in H. mutica at Klipport too. The main issue is flowering time and I agree this is problematic. But this is actually so in respect of the whole Southern Cape Haworthia complex where it is quite evident that there are two flowering periods, summer and winter and that despite this difference there is hybridization. What happened in the past?
DA writes…”MBB 7801 flowers…with the typical thin tender peduncles and delicate flowers identical to that of H. maraisii and some H. magnifica and H. mirabilis. Normal H. mutica flowers in spring (Sept-Oct) and has more robust peduncles and flowers.” This word “typical” again, does not convey the truth. There are no floral characters by which one can separate the three “species” the sentence suggests. I have also seen small H. mutica (Crodini) with very small weak inflorescences and comment on this as a general variable. See fig. 149, a recent picture of peduncles of H. mutica from Grootvlakte, Riviersonderend. There is a population of H. mirabilis (I presume Breuer and Marx would use the name H. magnifica var. magnifica) from south Riversdale MBB6651. The plants are very small and cryptic and when I collected seed it was from small slender inflorescences with 3-4 seed capsules. In cultivation I raised a large number of plants that were all different and some were huge. The inflorescences were robust as well with 15 -20 capsules. So this reference to floral characters is simply a loose statement and I do not believe that a proper overview of the flowers in H. retusa and H. mirabilis will reveal any dramatic differences. I have taken good note of flowers and in particular the strange fish-tail bud-tip that characterizes the larger set of populations in the southern Cape.
DA states that I… ”made several blatant errors by ignoring the importance of flower features in his studies”. It this one supposed error or several? I have not ignored flowers at all and it is a blatant error to say so. I simply did not and still do not find them useful. It is fraudulent to suggest otherwise. That fish-tail bud is highly significant and I made a great effort to quantify the differences between H. reticulata and H. herbacea. That the attempt failed was only because the sample required to get to statistical difference became too large. Nevertheless I then tackled a good number of H. mirabilis populations that also have the same bud tip. I concluded that there was a difference between the populations south of the Langeberg and those north and then had to discard that as erroneous because the same problem of variability occurred as with the rosette of leaves. Even the general flowering times I gave for the area was quite wrong – flowering time ranged from November to late April. It is odd that the fish-tail bud tip does actually influence the tips of the upper tepals in that the margins tend to be flattened together. But there is a huge difference in that in H. herbacea and H. reticulata the upper tepals are held far out from the mid-axis (hence ‘subregularis’ or ‘cowboy-hat’) while in H. mirabilis the tepals may meet above the middle upper tepal.
More by the DA… ”So, only careful observation and comparison of diagnostic features, starting with flowers, is the solution.” This is a weak sop because it is something that I started to do and found quite unhelpful. The history is that the small aloid genera were virtually recognized by the fact that the flowers were small. The differences between the subgenera were dismissed by Dr L.A. Codd with the statement that “small differences” were being ignored. DA overlooks a single major binding factor between H. retusa and H. mirabilis viz. bud similarity. I have also discussed in detail the whole question of diagnostic features and went to the genus Oxalis to substantiate my observations that small detail is not helpful. It comes back to the issue of what is a species? I maintain that they are not these artificial elements circumscribed by morphological detail. They are systems set in a geographical framework. The species, whatever they are, have surely not arisen from independent sources. My field work is very extensive and I have shown over and over again that the populations are linked so that when one finds a single population like at Buffeljags, one asks how it fits. It is not any misconstrual by G.G. Smith that a sample from a population south of Heidelberg was likened to H. mutica. In his time it was not known where H. mutica even occurred and hence he made a mistake in describing H. otzenii. DA is defending a methodology that failed miserably and is not going to provide any solutions in the future.
Well, well, well one might say. The DA maintains that Bayer has lost his way, to support an argument that he had stopped “doing taxonomy”. DA is not strong on logic but it is intended that way and neither are the DA’s observations necessarily correct. DA writes this … “Bruce’s dilemma is that he spent a lifetime and hundreds of thousands of Rands doing his research and while he did indeed do an invaluable contribution, he is now faced with the fact that he can’t do that full and neat integration of it all because 1) there is more variety in the wild than what his dispensation recognizes and he feels overwhelmed by the idea of having to try and do a real truth-reflecting revision at this ripe age and 2) he made several blatant errors by ignoring the importance of flower features in his studies.” Point two I have already dismissed, but DA cannot be serious when he suggests that there is some kind of dilemma involving money and time spent. This is an issue that I will have to address when I meet the Angel of Death.
I do recognize that there is great ‘variety in the wild” quite contrary to DA’s claim that here is more variety than what my dispensation recognizes. The main gist of my writing has been this extraordinary variation. I am not overwhelmed by it at all and I maintain that my dispensation is the only way to rationally express it. To say it confuses me is not an overstatement because it is clarification that I have been seeking. Now Steven Hammer puts it very well… ”it’s almost as if you are being blamed for nature’s complexities” and I said the same thing in different words long ago. The Latin binomial system fails and I have argued and shown evidence for the need of a new and different model What is more I have proposed such a model too. Anyone who contemplates at any age, the “truth reflecting revision” the DA demands will be faced with a daunting task of citing the synonymy of all the species described or recognized by Breuer and Hayashi (let alone cite all the collecting records). That alone could occupy a lifetime. Not only that, but to also cover the usages and references by the many authors who have written about the plants would be a humungous task. I suspect that DA is implying that I have become senile with a “ripe old-age” comment. My response is that at least my senility was preceded by some degree of clarity in contrast to what I am forced to conclude about contemporary Haworthia taxonomists! I see no reason whatsoever to depart from a logical argument about species definition and the significance of geographical distribution from my species list of February 2009. Effectively this is a revision and it just does not have literature and specimen citations that meet the needs of “a full and neat integration”. Let me state categorically that there is not the slightest hope of such a pie-in-sky solution.
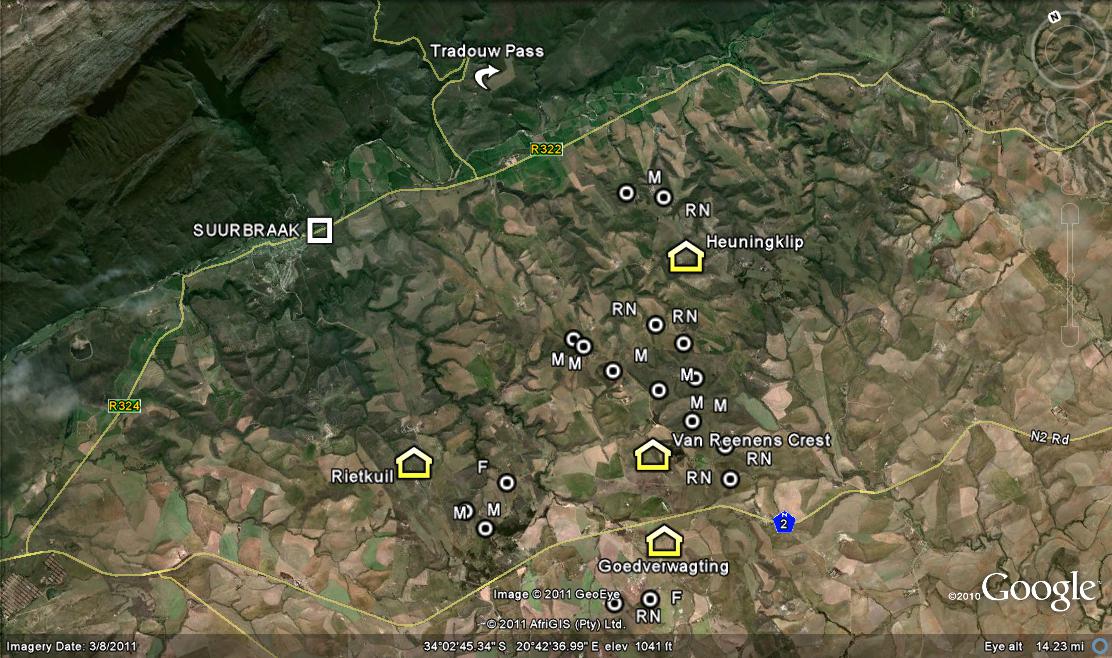
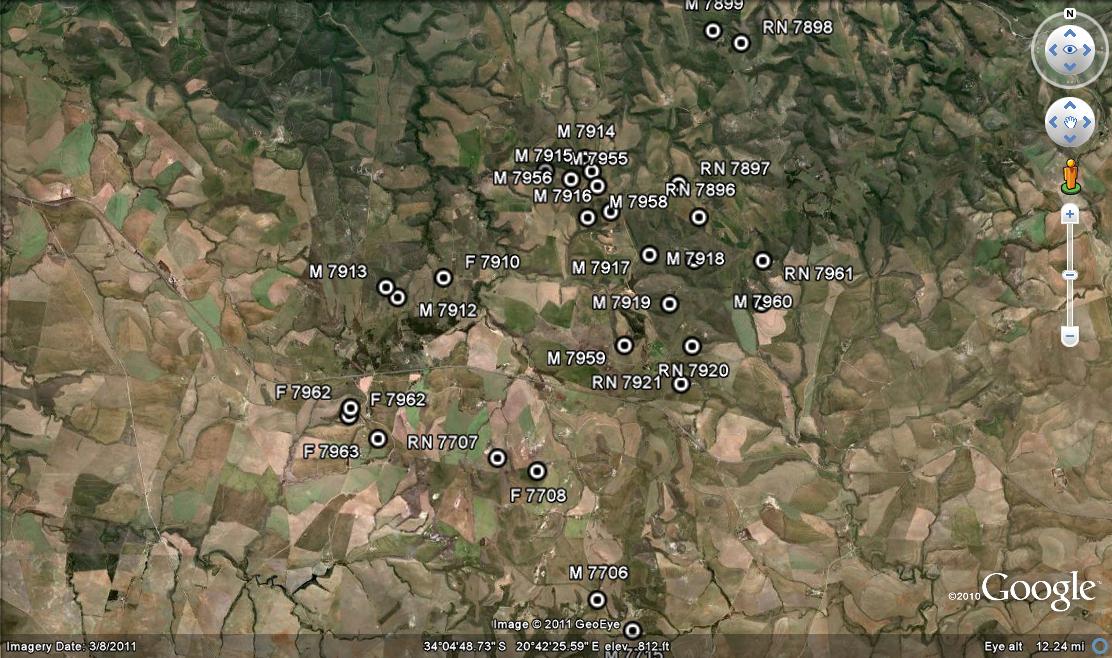
So we ask Lawrence Loucka’s question… ”How best to separate facts and knowledge from personalities and ego?” First present the facts. I want to add to the information that I presented in my article on “Haworthiaretusa ‘nigra’ – A grand finale”. There I discuss and illustrate plants from the wider general area of Buffeljags and especially from the Tradouw area to the northeast. Covered are four populations from the farm Heuningklip, and three from the farm Rietkuil. Here I want to discuss and illustrate eight more populations from the farm Van Reenens Crest that lies between. The gist of my argument is that classification of Haworthia is far beyond this nonsensical and simplistic generation of new names in exactly the same way that a miasma of names was generated in the years before 1948.
It is not just chance that the area I have recently been exploring has been so neglected for so long. It is a high-lying and fairly featureless area and very grassy in comparison to the Karoid Broken Veld, Karoid Valley Bushveld remnants, and the rocky hills, streams and river beds of the areas north east and west. The vegetation is a grassy Renosterveld and the species diversity seems extraordinarily low. There is not much there to attract a succulent plant enthusiast and explorer compared to far more enticing and rewarding countryside elsewhere. My exploration there is a direct result of my having exhausted most other options available to me. An added consideration is that of prediction when long experience has proven that connections between populations exist and need to be found.
So it is not surprising that we found 8 new populations, 6 in the context of H. mirabilis as I circumscribe it, and 2 for H. retusa ‘nigra’ in the same vein of personal circumscription. These populations were all new to me and very exciting. Nevertheless I would not dream of seeing them as new species anymore than I did for H. mutica ‘Buffeljags’. The Van Reenen’s Crest populations fit a pattern and need to be considered in the context of the bracketing populations at Heuningklip and Rietkuil discussed in the preceding chapter. The collections and illustrations are mapped in an accompanying map and are identified as follows:-
The populations.
Set 1 – H. mirabilis. The plants in the following six populations are collectively different from those at either Rietkuil or Heuningklip. Using the erroneous or absent logic that characterizes the description of “H. groenewaldii”, there is no doubt that at least one new Latin binomial should be generated. Far from being confused by the huge variation I have seen in Haworthia and guided by a systems view of species, I see these as being representative of one species. It would be useful if proponents of some other dispensation take these pictures individually and arrange them according to the characters they so diligently use to denote their many different species. The question of flower character I laugh off because I have tried to use it and know it will fail. I will come to flowering time again. Plant size is barely significant because in such clay soils the plants do not grow to the size that they will in cultivation. Generally the plants in these populations fall in the category 35mm to 800mm diam. But it is difficult to gauge age and the direct effects of niche favourability that will favour growth and size.
1. MBB7914 H. mirabilis, Van Reenens Crest See figs 1-19. This was a small population as can be judged from fig.1 where the pressure burst and exposed white clay area barely exceeds 100sq m. The plants were only on the perimeter of the area. This is what makes it quite difficult to assess what the population structure may have been in the era before farming and cultivation. Very seldom indeed have I found plants anywhere in pristine arable or even semi-arable landscapes. Fig. 2 is a view looking across to Heuningklip and the locality of MBB7896 H. retusa ‘nigra’.
Fig. 1 View of locality MBB7914.Fig. 2 View from locality MBB7914 to Heuningklip.Fig. 3 7914 H. mirabilis
2. MBB7915 H. mirabilis, Van Reenens Crest See figs 20-27. Fig. 20 is a view looking south to a very large area that we did not explore because we chose instead a similar valley immediately beyond the one pictured. The MBB7915 locality was rather odd. In fact the whole exercise was in the respect that we kept being reminded how similar the situation was to Kiewietsvlakte where H. mirabilis and H. retusa play out a similar show. In this case there was no actual pressure burst. In this case there was less white clay and there were the characteristic platform-like ridges as if made by grazing animals. There were few plants and while rather scattered, were still confined to what I can only describe as low-biomass bearing area i.e. shallow gravelly soil that does not support dense vegetative cover.
Fig. 20 View from locality MBB7915.Fig. 21 7915 H. mirabilis.
3. MBB7916 H. mirabilis, Van Reenens Crest See figs 28-48. Fig. 28 is a view of the white clay in an area very like that pictured in Fig. 20. Very heavily utilized by grazing animals, it is surprising that there were so many plants. The variability is quite evident from the pictures and some plants stand out as very different.
Fig. 28 The clay soils in which the plants often grow. Locality MBB7916.Fig. 29 7916 H. mirabilis
4. MBB7917 H. mirabilis, Van Reenens Crest See figs 49-70. This was a dramatic population because the plants were not quite where we expected them to be. Fig. 49 shows a more typical habitat in the distance. Here the plants were rather in a very brown gravelly situation and absent a short distance away where there was more white clay. As for all these populations there was a wide range of surface textures and also leaf shapes and endings. Often the leaves had a very distinct mucro (end-awn or point). Equally often the leaves were without any point and rounded and this was not true for all the leaves on any single plant. It is very problematic to make statements about the plants because when we were looking at Rietkuil plants we were reminded of a host of more easterly populations and plants. In this case even variants of H. emelyae came to mind.
Fig. 49 View from locality MBB7917 to that of MBB 7919.Fig. 50 7917 H. mirabilis.
5. MBB7918 H. mirabilis, Van Reenens Crest See figs 71-87. This population was unusual in that the plants were generally small and proliferous. Sometimes there were as many as 20 rosettes in a clump.
Fig. 71 7918 H. mirabilis.
6. MBB7919 H. mirabilis, Van Reenens Crest See figs 88-116. One needs to refer to the map to see how the populations now relate to one another. These were again very variable as I have come to expect in the whole H. retusa/H. mirabilis collective – to coin yet another adjective to do what I do not think the formal system allows. Note carefully fig.100 in which a young bud is clearly visible.
Fig. 88 7919 H. mirabilis.Fig. 100
Set 2 – H. retusa ‘nigra’ We found just these two populations.
7. MBB7920 H. retusa ‘nigra’, Van Reenens Crest See figs 117-137. These plants were in full flower and setting seed as one expects for the summer-flowering H. retusa elements. In fig. 128 a plant is shown with well-developed end-awn and fig. 134 a plant with rounded leaf tips. Fig. 137 is simply a view of the flower that does not even depict the more normal fish-tail bud tip – see next!
Fig. 117 7920 H. retusa ‘nigra’.Fig. 128Fig. 134Fig. 137
8. MBB7921 H. retusa ‘nigra’, Van Reenens Crest See figs 138-148. Fig. 138 is a plant that could have come from an H. retusa ‘retusa’ population from around Riversdale, while Fig. 146 could have come from H. retusa ‘turgida’ from somewhere around Heidelberg. Fig. 147 clearly shows what I mean by the fish-tail buds. Fig. 148 shows that there has been quite extensive exploitation of the white clay (kaolin). Fortunately (?) unlike the bentonite that is mined on large scale, the kaolin pockets are often small and restricted and apparently not commercially viable.
Fig. 138 7921 H. retusa ‘nigra’.
Conclusion The plants in the above populations cannot be categorized without reference to the totality of populations and I will try and summarize my point of view. These populations occupy a position very central to the distribution of both H. retusa and H. mirabilis. I need to quote DA again who wrote … ”His (Bayer’s) emphasis is upon phylogeny and understanding the evolutionary processes at work in Haworthia and he loves getting involved with very complicated arguments and theories which (frustratingly or delightedly) to him nobody quite follows or is really interested to follow.”
DA is most fortunate to be able to dismiss the basis of classification and what all biologists involved in classification try to do. It is quite wounding to say that I enjoy this theorizing. I would be far happier if there was no contradiction and I could wander out in to the veld and be assured that there were smart neat boxes and simple ways of filling them from a manageable list of names and unequivocal identifying characters. My observation is that all this so-called “taxonomy” practiced by too many writers is so flawed because the very basis of biological variation is wholly ignored and forgotten. It does not take rocket science to imagine that science was supposed to set us free from superstition and blind belief and allow us to individually question the nature of creation.
Here we have a real problem that is overridden by a basic problem in formal classification. If I could start now afresh with a clean slate and knowing what I now do, I would still be handicapped by the restrictions of the nomenclatural code. This is that I do not believe that the name H. mirabilis as typified by a plant that comes from an outlying population truly represents the species. Perhaps this is a point not understood by the collector community, and in fact even by botanists in other disciplines. The “type” is only there to serve as a reference point for the NAME and may not be remotely typical of the species that this name is applied to. What is unfortunately true as with the crop of taxonomists who have muddied the classification of Haworthia in more ways than one, is that more often than not the application of names is not to species at all but to an odd assortment of plants that meet no criterion for a basic unit of life forms. In the case of the name and type of H. retusa there has been curiously no problem and the vast variation there is forgotten simply because of familiarity. The interesting thing is that the name H. mirabilis was not even in use before I resurrected it in about 1976, so anything that remotely departs from an unfamiliar norm must be “new”? For H. retusa ‘nigra’, I must have got the name ‘nigra’ from the old literature and reference to the Kransriviermond collection that Smith informally listed as H. mutica. The point is that in my acknowledged ignorance I attached the name to H. mutica because of the absence of an end-awn and a relatively rounded leaf tip to the plants seen. (Looking back to all my pictures I see that in some plants there is definitely an end-awn.) The dark-colour has resulted in a name that is not appropriate for the populations that can be considered to constitute the taxon. Neither is the population at a geographic end-point and as a probable hybrid entity representative. This statement also only scratches at the surface of a real problem of origins and changing faces of the plants with time.
Reflecting about relationship as a taxonomist is required to do, and with due respect to DA and the weird views of my preoccupation with relationships the invitation to him required; it seems to me that the unseen and unknown element, now only evident (H. retusa ‘nigra’) as a widespread and common element in this central area, is important. It may be the precursor to the real way in which H. retusa and H. mirabilis have developed in close synchrony and association in both easterly and westerly directions.
The really strange occurrences are evident from the map herewith (fig. 150). The two species do not grow in direct close association in the area under consideration. There are places where they do e.g. Skeiding about 20km eastwards; KomseRante, Riversdale; Soetmelksrivier, E Riversdale. Why is this? There is no observable difference in habitat or associated vegetation, and still there is pattern in the distribution. It appears to me that they are separated by flowering time (that does not preclude hybridization as field hybrids show) and in character by colour and a tendency to greater surface roughness of various kind and degree in H. mirabilis. The mystery deepens when H. floribunda appears in two places but again as sole occupant of habitat seemingly favourable to either H. retusa or H. mirabilis. Pattern is broken here because the evidence seems to be absorption into H. mirabilis in other places where the two species do closely meet. The conclusion has to be that I have come no nearer to a neat solution. The H. retusa ‘nigra’ populations cited above will attract no takers, but the H.mirabilis populations will. I create no new name because my personal view of Haworthia does not call for one.
Fig. 149 Peduncles of H. mutica, E RiviersonderendMap of Tradouw South
Acknowledgement I particularly want to thank Gerhard Marx for his considered points of view he has crafted as my Devils Advocate. He is an extraordinary grower and observer, and an artist of the highest order. He had driven me to more carefully and thoroughly explore and document what I see and think. I am extremely grateful to him. Lawrence Loucka has been a great help as an advisor, and so has Steven who brings something special to the whole field in the way he “communicates” with plants and about them. Kobus Venter is ever supportive. Access to Van Reenens Crest was through kindness of Trevenan Barry unsoured by the pressures of harvest time. ♦
These field trips, to the Potberg this time, are always made with some objective in mind in respect of new exploration. In this case I wanted to get more pictures of H. mutica as it is a species that I have few digital images of. There were also localities that I remembered from the days when I was sweeping the countryside at a fairly coarse scale and was not much bothered by detail. I confidently expected the number of real species conforming to that in other fields of botany and zoology, to be in the region of 33. I never dreamed that such divergent views would, or even could, arise from less information than even then available to me. So while 450 names were whittled down to the mid-hundreds by me, students of the genus have in recent years pushed that up to the 600 mark. My opinions have been couched in quite conservative terms but it is a problem of the nomenclatural system that an identification in respect of a Latin name evokes a reality that does not exist. I maintain that the problems we experience in Haworthia are no different to that which exists in many animal and plant genera. I think that primarily this is because of the absence of insight into, and understanding of, the actual nature of species and the two dimensional model we use to relate them. Species are very variable systems because they have to be to survive the constantly changing world they occupy. In this article I am just going to present images of plants within populations of four different species viz. H. variegata, H. minima, H. mirabilis and H. mutica.
Set 1 H. variegata H. variegata has an odd distribution in that it is primarily in the lower Goukou River drainage system southeast of Riversdale. Plants of H. monticola from the Uniondale and Willowmore areas, and even those of H. angustifolia from the Eastern Cape, can be very similar. So the distribution south and west of the lower Breede River is quite remarkable. Two populations, one high on the western end of the Potberg Mountain and another northwest of this at Luiperd’s Kop are very similar to those in the Goukou area. Chris Burgers of Cape Nature brought me plants from the lower north slopes of the Potberg that were quite different. Small single plants with outcurving leaf ends barely protruding above the pebbles the plants were growing in. I named this H. variegata ‘hemicrypta’ but not without reservation in the same way as I also named the plants in another population on the Potberg and at Kathoek as H. variegata ‘modesta’.
Chris Burgers did take me to his site many years ago and it is one of those places I cannot recall in the pebbly flats and Fynbos vegetation of that area. So the finds here seem to represent that same element but with subtle differences in respect mainly of variegation and spination of the leaf margins. I have mentioned elsewhere that there is an interaction with an element I refer to as H. floribunda although it is not clear if this indeed that species, or if it is an emergent polytopic element of H. mirabilis in consideration of the variation and distribution of these two apparently different species. We may have a truth for one area that is different from a truth in another. Polytopic means similar looking things arising in different places from a common genetic pool.
1. Map of the Diepkloof area (fig.1)
Map of the Diepkloof area.
2. MBB7923 H. variegata S Diepkloof. (figs 2)
Fig. 2.1 7923 S Diepkloof
3. MBB7924 H. variegata SE Diepkloof (figs 3)
Looking west to the Kleinberg
4. MBB7925 H. variegata SE Diepkloof (figs 4)
Fig. 4.1 7925 SE Diepkloof
5. MBB7927 H. variegata SSE Diepkloof (figs 5)
Fig. 5.1 7927 SSE Diepkloof
6. MBB7928 H. variegata SSE Diepkloof (figs 6)
View looking east to 7927.View looking southwest with Kleinberg off to the right.
7. MBB7515 H. variegata Stoffelsriver (figs 7)
Fig. 7.1 7515 Stoffelsrivier.
Applying the available varietal name, I should refer to these populations as H. variegata var. hemicrypta. However, I think this implies fixity of opinion and a certainty or reality that are neither not strictly true. I have added the Stoffelsriver two pictures just as a reference because while the first 5 populations are associated with recent Tertiary deposits and Renosterveld vegetation, the last is in sandstone gravels and in Fynbos.
Set 2 H. minima This species of course belongs with three others in the Robustipedunculares and in my opinion more closely related to Astroloba than to Haworthia. It is widespread and even occurs in the Little Karoo. None of these populations are significantly different from the general run for the species, but we were awed by the beauty of the plants in their natural settings where exposure results in compactness and beautiful colouration. They can occur in quite different environments and habitats. There are a few aberrant populations and hybrids with other members of the sub-genus.
8. MBB7496a H. minima Diepkloof S (figs 8)
Fig. 8.1 7496a Diepkloof S
9. MBB7926 H. minima SE Diepkloof (fig. 9)
Fig. 9 7926 Diepkloof S
10. MBB7929 H. minima SE Diepkloof S (figs 10)
Fig. 10.1 7929 SE Diepkloof
11. MBB7930 H. minima S Diepkloof (figs 11)
Fig. 11.1 7930 with 7947 SE Diepkloof minima
12. MBB7932 H. minima S Kleinberg (figs 12)
Fig. 12.1 7932 S Kleinberg
Set 3 H. mirabilis H. mirablis is an extremely complex group of many populations and plants and distributed from Albertinia in the east to near Caledon in the west, and from the foothills of the Langeberg to the coast. The relationship with H. retusa hangs on the slender thread of flowering time, with H retusa flowering in late spring and H. mirabilis in late summer. The relationship of these two species to H. pygmaea in the east and H. mutica to the west has been mooted by me elsewhere. A similar scenario can be suggested for the species recognized north of the Langeberg.Included in the following sets is the population at Die Kop that I surmise has its origins in interaction between H. mutica and H. mirabilis.
There are three populations in these sets (nos 13-15) that are suggestive of H. mirabilis ‘atrofusca’, while others have distinct resemblances to H. floribunda (e.g. 16). It is curious that nearly all the populations of H. mirabilis that occur in the same general geographic area as does H. mutica are of smallish plants. These I originally regarded as a separate species viz. H. heidelbergensis. This should be seen as one of those decisions based on inadequate sampling and knowledge of the field. This phenomenon now suggests to me that H. mutica is occupying middle ground between H. retusa in the ‘turgida’ form and H. mirabilis.
13. MBB7496 H. mirabilis Diepkloof S (figs 13)
13.1 7496 Diepkloof SView overlooking habitat 7496.View looking west to north of Kleinberg.
14. MBB7497 H. mirabilis S S Diepkloof (figs 14)
Fig. 14.1 7497 S S DiepkloofView looking north.
15. MBB7933 H. mirabilis S Diepkloof (figs 15)
Fig. 15.1 7933 S 7496 Diepkloof
16. MBB7922 H. mirabilis Diepkloof (figs 16) A locality established by Jakub Jelimicky. The similarities to H. floribunda is only apparent in some of the plants where the leaf tips are twisted, slightly flattened and rounded at the ends.
Fig. 16.1 7922 Diepkloof
17. MBB7931 H. mirabilis S S Kleinberg (figs 17)
Fig. 17.1 7931 SS Kleinberg.
18. MBB7531 H. mirabilis S Kleinberg (figs 18)
S Kleinberg slope. Enthusiasts at play.Figs 18.1 7531 S Kleinberg
19. MBB7935 H. mirabilis Langvlei (figs 19)
7935.1 LangvleiLangvlei, the plants are in a tiny bit of rock just beyond the dumps.
20. MBB7938 H. mirabilis SE Stoffelsriver (figs 20) This locality was recorded when Gerhard Marx and Jakub Jelimicky misunderstood my directions. The plants are incredibly cryptic and very hard to see in a not very well defined habitat. One can only follow my opinion on the classification in the context of 8 other populations in the sandstones and Fynbos, and then relate these to populations in shales, tertiary deposit and transitional vegetation.
Fig. 20.1 7938 SE StoffelsrivierSE StoffelsrivierSE Stoffelsrivier View looking east to Sandhoogte
21. JDV86/2 H. mirabilis Haarwegskloof (figs 21) These are small plants in very close association with H. mutica as discussed above and are in quartz outcrops rather than in the shale ridges where H. mutica is found.
Fig. 21.1 JDV862 Haarwegskloof
Set 4 H. mutica H. mutica is not a species that is well-known and this is probably because it is not proliferous. The name mutica means without a point and “typically” plants of H. mutica should and do have no point (end-awn). The main object of the trip was to obtain more images of this species to convey a better impression of the many variants and how it can be viewed in relation to other species.
22. MBB7075 H.mutica Grootvlakte (figs 22) I am not certain of the origins of H. otzenii G G Smith and the locality that I traced was approximately 8km east of Riviersonderend. That site is now very degraded and I could not find plants on a visit many years ago but where I did see them on my first visit there. This Grootvlakte population occupies a similar habitat in an exposed shale ridge south of the Riviersonderend River.
Fig. 22.1 7075 Grootvlakte7075 View of site at Grootvlakte
23. MBB7935 H. mutica Crodini (figs 23) The name Crodini may have been used for the following collection at nearby Witklipkop, and I say this because I did once get seed from there and Jakub Jelimiky informs he has a plant labeled ‘Crodini’. I only saw two plants here at Crodini itself when the site was severely grazed, and we found only 8 plants on this occasion. The plants appear to be in quartz, but this is a bit of an illusion as there are white quartz bands in the shale and the plants prefer the shale.
Fig. 23.1 7935 CrodiniView across site. Grootvlakte
24. MBB6536 H. mutica Witklipkop (figs 24) This is not a robust population in shale and also marginal to quartz.
Fig. 24.1 6536 Witklipkop, Napky
25. MBB7937 H. mutica Platkop, S Napky (figs 25) The substrate here seems to be silcrete but there is no underlying kaolinite and the area is cultivated, which suggests shale. So that is a bit of an unknown.
Fig. 25.1 7937 N Platkop SE LangvleiN Platkop SE LangvleiN Platkop SE Langvlei
26. ADH2729 H. mutica Ouplaas, DeHoop (figs 26)
Fig. 26.1 ADH2729 Ouplaas, De Hoop
27. MBB7934 H. mutica Wolwefontein (figs 27)
In early 1970 I took Peter Brandham and David Cutler of Kew to look at Haworthia in the field. In fig.27.12 the view is southward to Kathoek and the cottage in the foreground was occupied. It must have been abandoned soon after and the access road obscured. The result was that the place was later difficult to find, but the plants are still there. It is not far from the problematic Die Kop population to come.
Fig. 27.1 7934 WolwefonteinWolwefontein
28. JDV85/17 H. mutica Haarwegskloof (figs 28) This record goes back to 1985. We did not have landowner permission here as we did not know exactly where to go. The area was also unfenced. So we were confronted by the farm manager and never took as many pictures nor explored more widely. It certainly would have been problematic had we not simply being photographing. Anybody else may have ended up at the police station. It is a case of farm land too hilly and rocky to cultivate and bought by apparently wealthy city people with an eye to conservation. This may result in the place being game-fenced, a lodge built for the up-market; and made perhaps even less accessible.
Fig. 28.1 JDV85-17 Haarwegskloof
29. MBB6641 H. mutica Hasiesdrift (figs 29 JDV97/26) This was one of my early encounters in 1969 with Haworthia, where I fortuitously stopped at a river crossing, lifted a branch of a shrublet and saw a magnificent plant just like that illustrated and serving as the type for the name H. mutica (see fig.29.1for an example)). A year later a new road was built and if there had been a population there it was no more. In 1996 when I was writing Haworthia Revisited, I went back there and after a diligent search found a very small group of plants at the base of a small plant of Aloe ferox. The two species seldom grow among each other so this really was odd. I have periodically been back to see what was happening because there were many Aloe seedlings. The very small population has waxed and waned and the mothering Aloe seedling has died. There has been a small spread of plants to occupy an area of about 2 square meters. I know here are more plants about 2km away but have not explored the wider area more thoroughly.
Fig. 29.1 JDV97-26 Hasiesdrift.
30 MBB7500 H.muticaXmirabilis Die Kop (figs 30) So we come to the problem child that has acquired the species Latin binomial H. hammeri. I think this is unfortunate as Steven Hammer deserves a great deal better than this. It should not be necessary to have to also manage personal issues in an inquiry into what the name means or if it is a valid statement in science. This population is one I found when I was exploring widely for an understanding of H. mirabilis. It was not growing in conventional habitat and populations occur at many places in the wider vicinity. So I do not comprehend at all why this single population with absolutely nothing to support its designation as a species in respect of anything but the fact that the plants look different from something else. The plants are also very variable as the set of pictures should show, and the plant suggested by the picture accompanying the description is not typical. The proximity of H. mutica and the nuances of these plants with this species and also with that of other nearby populations of H. mirabilis, suggest to me that this population is of hybrid origin. I also use the term hybrid in the sense of genetic similarity because hybridity itself implies separate existence. My feeling is that H. mirabilis and H. retusa have separated in time from a common gene pool and that the process is not complete and nor will it be. My species definition requires that we see all these populations as one system that is spread in geographic space and is changing with time. The rate of change is a fourth dimension.
I suppose one should comment on the shiny leaf surface (fig. 30.3 especially) that is present in H. mutica, as this character is also described as a diagnostic for H. groenewaldii that is as contentious as H. hammeri.
Fig. 30.1 7500 Die Kop
Conclusion The objectives of the excursion were fully met and I am sure these pictures will contribute to a better view of how these four species vary. Perhaps this will illustrate also the illogic of Latin binomials for every variable, however desirable these names are for traders and collectors.
Acknowledgement Bennie Viljoen of Grootvlakte, Koos Badenhorst of Langvlei, Maree and Elizabeth Prinsloo of Stoffelsriver, Ferdie of Diepkloof Security Services, Adele and Vlooi DuToit for Die Kop, Martin Mijnhardt of Haarwegskloof, Neil Giliomee of Hasiesdrift. In some cases we simply could not track landowners and trust they will forgive us our trespasses remembering that invasion of ones territory is close to an unforgivable sin. ♦
A supposed new species of Haworthia viz. H. groenewaldii Breuer, is described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald. It presents the description of this supposed new species from Buffeljags east of Swellendam that I would simply have identified as another variant of H. mutica. This is not because I am confounded by the variability among the plants in the genus, or even in any one species. I recognize the species as systems of populations in which the individuals vary from one another as one would expect in any group of living things that maintains the flexibility to adapt to constantly changing world conditions. In this even time becomes a variable. I think species are very important elements if the whole of creation and not just for taxonomist and collector activity. Other people have other ideas of what species are, so my disagreement is hopefully forgivable..
Although H. mutica was described by Haworth in 1821 it was not allied to a South African field population until recognized by Col. Scott in 1985. G.G. Smith had failed to recognize it when he described his H. otzenii in 1945. The type by which the name is supported is a Kew illustration reproduced here as Fig. 1. This then is what one would expect a typical representative of the species to look alike. Now the ever present problem in Haworthia, is that no two plants in a population may look quite the same. Hence my problem with the description in which the word “typical” is rather bandied about. It falls into the first aspect of taxonomy.
Fig. 1 H. mutica. Kew herbarium. The type.
Firstly a plant is illustrated on the front cover of the respective Alsterworthia that, presumably the authors, state is a typical specimen of H. groenewaldii. Secondly, Marx is quoted as saying that the “typical H. mutica” grows only 20km west at the farm Dankbaar. Statements like these are used to strengthen and support opinions and generate a reality that Latin binomials sadly lack. It so happens that I know both these populations quite well and these statements are news to me. I do not think the specimen on the front cover is by any means typical of the population at Buffeljags, and certainly not at three sites recently discovered nearby. The plants at Dankbaar also do not in my opinion fully meet the imputed similarity to fig.1. See fig 2. for an image of a plant representative of the Dankbaar population. I would be very hesitant to say that this is typical of Dankbaar plants.
Fig. 2 MBB7741 H. mutica. Dankbaar. A representative.
There is a curious problem here in that Scott does not use the type illustration in his Revision and does not state any origin of the plant he uses to illustrate the species i.e. H. mutica. As far as I am aware the type of a synonym that G.G. Smith described viz. H. otzenii, came from east of Riviersonderend, but this is for an Otzen collection no. 6. The type was cited by Scott as Otzen 10 but this is not in the Compton herbarium where it should be. So I am not sure offhand where that came from. However, this is not really relevant to the discussion. I just want to state that finding a plant that matches the type is no mean feat and that it was by sheer chance that in 1969 I came across a population of plants at Hasiesdrift that did. See fig. 4 and 5. I selected one image and then realized that it did not have round enough leaf tips to meet need, so I selected another. In the first picture the leaves tend to have a “mucro” – a small point to the leaf that looks different to a well developed end-awn (bristle) that the leaves can also have.
Fig. 3 JDV97-26. H. mutica. Hasiesdrift. Representative.Fig. 4 JDV97-26 H. mutica. Hasiesdrift. More representative.Fig. 215 7801 H. mutica ‘groenewaldii’.
Another issue is that Gerhard Marx once argued with me that the mooted H. groenewaldii was much nearer to H. mirabilis than to H. mutica as I had suggested. What he luckily is able to ignore is my observation of the similarity of some plants of H. mutica to H. mirabilis see figs 5 and 6. I never saw a plant quite like fig. 5 in all my exploration at Buffeljags but Jannie Groenewald collected this one there. Fig. 6 is not typical of the population either and I used this same figure somewhere else in my writing to comment on the similarity to H. mirabilis ‘badia’ variants at Sandfontein (east of the “typical”). Gerhard is still more fortunate to be able to ignore the similarity of H. mutica to H. retusa see figs 7 and 8. I even surprised myself when in looking for a suitable picture, from many, I picked this fig. 7 and find it is also Hasiesdrift albeit a cultivated version. (There is such an interesting story around the Hasiesdrift site). Fig. 8 is a representive of H. retusa from Pienaarsriver pictures and I feel sure that readers will agree that the names could be switched. It was very difficult to ignore pictures from Pienaarsriver that I could have used with figs 5 and 6 in the context of H. mirabilis.
Fig. 6 JDV96-16 H. mutica. S Napky.Fig. 7 JDV97-26 H. mutica. Hasiesdrift, N Bredasdorp.Fig. 8 MBB7776 H. retusa. Pienaarsrivier.
It becomes still more interesting (I would have said complicated but my critics maintain that this variation confuses me and it is actually possible to get it all tidy and neat) when one further compares a plant of H. retusa ‘nigra’ ( fig. 9) with both H. mutica (fig. 7); and then H. retusa ‘nigra’ (fig. 10) with H. retusa (fig. 11). The latter is in fact from the population where the variant ‘geraldii’ originates.
Fig. 9 MBB7920 H. retusa ‘nigra’. Van Reenens Crest.Fig. 10 MBB7921. H. retusa ‘nigra’. Van Reenens Crest.Fig. 11 MBB7781 H. retusa. Komserante.
A last point I can make is that the leaf flecks said to characterize “H. groenewaldii” do occur in H. mutica at Klipport (see figs 12 & 13). One cannot simply ignore the extraordinary chain of similarities that extends across the entire distribution area producing only a slightly different situation at either extreme.
Fig. 12. MBB6512 H. mutica. Klipport.Fig. 13. MBB6512 H. mutica. Klipport.
Floral difference is a great issue that is misused to force an opinion. The flower is extremely difficult to study because the differences across the entire subgenus are so small. There are complications where, as an example, flowers of H. pulchella ‘globifera’ are indistinguishable from those of H. cymbifomis var. incurvula. In the subg. Hexangulares there is an incredible problem where floral differences within species exceeds that between species e.g. a flower of a plant of H. limifolia may more closely resemble that of H. coarctata rather than that of another plant of H. limifolia. It is easy to draw conclusions from small samples of a few flowers from a few populations but one very soon finds that with increasing sample size the most carefully constructed table of differences becomes senseless. Just when is an ever-aging flower on a stalk at the precise same stage of a flower you want to compare it with? How many flowers from how many plants are needed to make a valid statement? Do not forget that the observations must be made on plants on the basis of random selection too. This is a requirement mostly totally ignored when the more serious question of a species difference is being debated. That of course brings us back to this casual use of the word “typical” and its intractability.
The hardest problem to deal with is that of flowering time and on the face of it a winter flowering time versus a summer flowering time can be taken to suggest significant difference. Yet if one considers that a species needs to harbour genetic variation to ensure adaptation to any kind of environmental change and so survival, a different flowering time may be an extremely useful resource. Then we do have the reality of hybrids between species that do flower at these different times. So obviously and self-evidently populations of species may exist that has arisen from such hybridization between plants that flower at different seasons. The capacity of plants and animals to synchronize breeding periodicity is well-known.
I close with my observation that H. mutica is an assorted group of plants that seem to fall in some middle zone between H. mirabilis and H. retusa. Hence in the west we have H. mirabilis/ H. mutica/H. retusa ‘nigra’, while in the east we have H. pygmaea (=H. mirabilis+H.retusa ‘retusa’) and H. retusa ‘turgida’. In the area between there is vast variation of H. mirabilis and H. retusa that get a bit of H. floribunda thrown in too.
Map showing known distribution of H. mutica.
So am I confused, or have I confused you? As Steven kindly put it…’it’s almost as if you were being blamed for nature’s complexities’. Of course the ultimate reality is that we each have our own idea of what “species” are, and here I have used my version! ♦
It so happens that Heidi Hartmann first visited the Karoo Garden more than 35 years ago and it has been very difficult for me to pay attention both to her mesembs and all my other plant interests. In the last few years she has been working on Acrodon. This is a small genus of only 5 to 6 species that occurs in the Southern Cape with much the same distribution and habitat requirements as Haworthia. She had had some second thoughts on a species she had described as Acrodon calcicola and intimated that she needed photographs to show what proves to be detaching fruits (capsules). So off we went to get that northeast of Bredasdorp at Rooivlei. But Nick Helme had about a year before sent me an intriguing picture of a greenish soft looking plant from near the De Hoop Reserve entrance road to the east. I had considered that it might be an equivalent of the H. muticaXmirabilis population at Die Kop (MBB7500) that Ingo Breuer usefully described as H. hammeri. I use the name with great trepidation because to say what is correct usage is difficult. It could pass as a cultivar name, a varietal name or a form name. I am quite sure it has its origins in the interaction of two species and that is what a botanical name should reflect that; thus H. muticaXmirabilis or however else the nomenclaturists may require. So these journeys are never without distractions as Rooivlei itself is a remarkable site. I find that I have few images of the populations of Haworthia that occur there. The product is nearly all pictures/images.
Set 1. MBB7941 H. mutica Ouplaas, Wydgelee (figs 1-45)
This population is just 3km west of the Die Kop MBB7500 population. There are two intervening H. mirabilis populations; one small, one large and scattered, and a third a little to the north of east and more to the north. But that species is well covered in an Update article and my attention here is on H. mutica. Why Nick Helme’s plant was so green and soft-looking is a bit odd because the population of H. mutica there at Ouplaas is quite one of the largest and most dramatic of any that I have seen of the species. Daphne and I spent hours examining, reexamining and exclaiming at the sheer beauty and variation of the plants. There were plants squeezed into cracks and flattened so that one could not see any centre, others in the flat clayish areas between the exposed vertical shale ridges, but mostly in the shale. The site is north-facing and so is very hot and dry in summer. The plants had just finished flowering.
There was one particular image (fig. 24) I have used to show five different leaf-end shapes on the same plant.
Fig. 1 7941 OuplaasFig. 44 7941 View west to HaarwegskloofFig. 45 7941 View east to Die Kop
Set 2. MBB7943 H. mutica E Klipbankskloof (figs 46-54)
I saw plants and photographed one (MBB7246) south of Klipbankskloof in a valley mapped as Verbrandskloof. There is another farm, also named Klipbankskloof further west where Jakub Jelimicky recorded H. mutica too. I wanted to see that locality and registered the two Klipbankskloof places as the same. Not so. So I intended up exploring further east where we first found this in a west facing ridge. There was no sign of fruits or flower.
Fig. 46 7943 E KlipbankskloofFig. 54 7943 View west to farmhouse
Set 3. MBB7945 H. mutica SE Klipbankskloof (figs 55-80)
This was on a north facing slope and at first we were a bit taken aback because the plants we first observed had flowered poorly or some time previously. But that changed as from north facing slope we entered a very minor valley with both northeast and northwest aspects (more to follow). Here there were many plants and all just finishing flowering. There were some outstanding, dramatic plants. I wish I was a more accomplished and better equipped photographer. Some of the images would have been stunning.
Fig. 55 7945 SE KlipbankskloofFig. 80 7945 View lookin across depression to west facing slope
Set 4. MBB6638 H. mirabilis Rooivlei (KoeiseKop) (figs 81- 92)
It was fortunate for my story that I took pictures here. As in most of the greater area occupied by H. mutica,H. mirabilis occurs as these smaller, often (how often is often?) dark rough versions of H. mirabilis. Of particular note is the first image (fig. 81). This is of a plant with much smoother textured leaves and had there been and H. mutica in the near vicinity I would surely (mistakenly?) taken this for a hybrid because I know from experience elsewhere that summer and winter flowers can and do hybridize. But wait! Rooivlei is a unique site because there is a population of H. retusa ‘turgida’ nearby that is prominent in my whole explanation of relationships that my critics so dislike. Outside chance is that hybridisation is an explanation.
Fig. 81 6638 Rooivlei
Set. 5 MBB7944a H. mirabilis SE Klipbankskloof (figs 93-101)
I have numbered this as ‘a’ and ‘b’ for a reason. The two ‘populations’ are less than 200m apart. In fact the two H. mutica populations are not all that much more distant and there would be some debate as to whether they constitute populations at all. My argument would be that they occupy discrete separated habitats. This mirabilis population falls into middle ground. It is the upper slope where aspect changes from south to north. It is moister and the southern aspect of the small shale ridges provides mirabilis habitat. It is as if H. mirabilis is found where what I can describe is an escape route is present. These dark plants do not readily tolerate a full days exposure to direct sun and I speculate that this is the same kind of thing that happens on north-facing mountain slopes as around MacGregor. Plants are usually where the aspect allows escape from a full days worth of sunlight. The plants are much like at Rooivlei.
[ed.] images mislabeled as 7943, should be 7944.
Fig. 93 7943a SE Klipbankskloof
Set. 6 MBB7944b H. mirabilis SE Klipbankskloof (figs 102-110)
The different number is because the plants are now sharing both habitat and niche with H. mutica. I do not know any other site where this happens. The two species do occur in the close general area but that is then different habitat and the distance between ranges from about 50m (Haarwegskloof) to 400m (Rondeheuwel). What is now fascinating is that some of the plants resemble fig 1 of Rooivlei and we are faced with the probability that there is exchange of genetic material between H. mutica and H. mirabilis. This is what is suggested at Rooivlei too for H. retusa ‘turgida’ and H. mirabilis (see above).
Fig. 102 7943b SE Klipbankskloof
Conclusion None. This is purely instructive. What I gained from the trip was the realization that H. mutica would be a very good subject by which to explore variability and change with time. Also to look at conservation and impact of land-use on populations. Over to Jakub Jelimicky.
Acknowledgement Again a list as long as an arm. Francois Uys for access to Rooivlei. Nick Helme for information in respect of Ouplaas. Odette Curtis for interest in Renosterveld conservation and contact for Nick. Johan Dutoit of Vleitjie and a friendly combine mechanic for access to Ouplaas. Wynand Dutoit of Klipbankskloof for access to what is his wife, Madrẽ’s farm. ♦
The objective was to explore some likely habitats previously observed at Van Reenens Crest and nearby. We extended the scope to include further exploration for Haworthia mutica as I am still questioning the place of this species in the greater scheme of things. Thus here are four sets of populations that I report on viz. H. mirabilis, H. retusa ‘nigra’, H. floribunda and H. mutica. See maps Figs 1 and 2 for geographical position.
Van Reenens CrestFig. 1 Van Reenens Crest. M = H. mirabilis, RN = H. retusa ‘nigra’, F. = H . floribunda.Fig. 2 Known distribution of H. mutica
H. mirabilis
Set 1. MBB7955 Van Reenens Crest – mid west. (figs 3-11)
The plants are moderate in size to about 50mm diam. and growing on an erosional slope below tertiary gravels where there is white clay. They vary from densely tubercled to relatively smooth.
Fig. 3 7955 Van Reenens Crest
Set 2. MBB7956 Van Reenens Crest – west 7955. (figs 12-34)
Despite apparently favorable habitat these plants were about 300m west of the previous set. Variation is again apparent in respect of presence or absence of surface roughness. Generally there are plants in which some leaves are without a point (see Fig 1) while others are acute tipped (mucronate – with a short hard point) or even have a long end-awn. Margins may be smooth or very lightly spined. This seems to be more apparent in young plants.
Fig. 12 View looking east to Set 1.Fig. 13 7956 Van Reenens Crest
Set 3. MBB7957 Van Reenens Crest – west 7956. (figs 35-41)
Again the plants are very variable with dense surface tubercles to virtually none. The leaf shapes vary and in fig.39 that appears to be a group of seedlings there is a remarkable series of different leaf ends and margins. In some plants there is distinct spination of the leaf margin.
Fig. 35 View looking south to Tradouw PassFig. 36 7957 Van Reenens Crest
Set 4. MBB7958 Van Reenens Crest – south west 7957. (figs 42-51)
Fig. 43 shows two rosettes of a plant with a “crocodile-skin” surface. The surface tubercles seem to be flattened and the leaves are mucronate. The following photograph, fig.44, is of a plant with acuminate leaves ending in long end-awns, and the surfaces are relatively smooth – retusoid? Unlike the previous records for the area that are north-facing, this is a south-facing “pressure-burst” of white clay. A nearby habitat of vertical shale that might have been considered favorable for H. mirabilis elsewhere was devoid of plants.
Fig. 42 7958 Van Reenens Crest
Set 5. MBB7959 Van Reenens Crest – west homestead 7956. (figs 52-64)
Figs 53-55 are smallish plants with rounded leaf ends and relatively opaque surfaces. Fig.58 is remarkably like H. retusa, say, at Pienaarsrivier with very awned leaf ends and a high degree of translucence in the leaves.
Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS.Fig. 53 7959 Van Reenens Crest
Set 6. MBB7960 Kruiskloof, E Van Reenens Crest. (figs 65–78)
This population departs from a very broad norm for Van Reenens Crest and picks up the continuity with populations north, east and south. This is most noticeable in the surface texture where the tubercles are smaller and denser. The leaves can be highly rotate, see fig.68. A feature seen occasionally in the Van Reenens Crest populations and as far afield as the Diepkloof (Malgas) area much further south, is the occasional plant with a very stellate (five –pointed star) arrangement of the leaves that are quite horizontal see fig.70. Among the many oddities is the plant in fig.72 with floribundoid leaf ends and a leaf surface where the tubercles seem to be flattened. In fig.73 the tubercles seem to be spined. In fig.75 tubercles are almost absent.
Fig. 65 7960 Kruiskloof
H. retusa ‘nigra’
Set 7. MBB7961 Kruiskloof, E Van Reenens Crest. (figs 79-90)
These plants reinforce my perception that H. retusa and H. mirabilis are uncomfortably closely related. Figs 79 and 80 have the same general form as H. mirabilis ‘sublineata’. Fig.82 appears to me as possibly a hybrid as leaf opacity is deeper and there are marginal spines.
Fig. 79 7961 KruiskloofFig. 82
H. floribunda
Set 8. MBB7962 Niekerkshek. (figs 91-99)
Fig. 91 7962 Niekerkshek
Set 9. MBB7963 Niekerkshek. (figs 100-111)
The plants vary in size and to some degree in colour. This variation in colour from a deep purple to dark grey is very evident in fig.107. The leaves are much flattened and an occasional leaf is pointed. The plant in fig.102 is unusual in having white spots on the leaves. Marginal spination can be absent or very coarse. The leaf surfaces show no venation or translucens, as is expected in this species.
Fig.100 View looking south to Potberg.Fig. 101 7963 Niekerkshek
H. mutica
Set 10. MBB7950 DeDraai Klipbankskloof. (figs 112-123)
Fig. 112 – 7950 Die Draai, E Klipbankskloof
Set 11. MBB7951 Klipbankskloof West 1. (figs 124-126)
Fig. 124 – 7951 Klipbankskloof West 1
Set 12. MBB7952 Klipbankskloof West 2. (figs 127-129)
Fig. 127 – 7952 N Klipbankskloof West 2
Set 13. MBB7953 Klipbankskloof West 3. (figs 130-144)
Fig. 130 – 7953 Klipbankskloof West 3 View westwardsFig. 131 – 7953 N Klipbankskloof westward view of habitatFig. 132- 7953 N Klipbankskloof West 3
Fig. 175 JDV92-64 Very localised.Fig. 176 JDV92-64 Sanddrift
Set 16. JDV85/17 Haarwegskloof. (figs 188-213)
Fig. 188 JDV85-17 Haarwegskloof
There is a very wide range of variants among these 7 population sets. Leaf colour and shape vary enormously and it can only be said that generally the leaves are muticate. Almost invariably any one plant has a leaf or leaves that are also mucronate and in some cases even awned. The leaves may be abbreviated and short and squat or drawn out and elongated. Venation is also varied from many to few or only a single vein on the upper leaf surface. These veins may be straight and clearly separated or they may anastomize and link togther in a reticulate pattern. I did include two pictures of flowers, figs 173 and 174 just to indicate that simply taking two flowers at random one can see obvious differences.
Attention can be paid to figs 145 and 146 of Rondeheuwel where I consider these two plants indicate hybridization with H. mirabilis that does occur about 1km to the northwest. Fig. 147 shows a plant with a rather wrinkled surface.
Conclusion My opinion is that consideration has to be given to the odd way in which the “species” do not share habitat. It is extremely difficult to rationalize an ‘average’ plant for each population and so establish any norm for proper comparison. The populations vary by degree. It is also clear that if one examines the distribution of the species across the study area, it can be observed that H. mirabilis where it occurs with H. retusa ‘nigra’(at Heuningklip and Kruiskloof in the east), is not the same as plants to the west where H. retusa ‘nigra’ is absent. Consideration has to be given to absorption into H. mirabilis. Why H. floribunda should make a re-appearance at Rietkuil and Niekerkshek in the west, when everything points to its absorption into H. mirabilis in the east at Skeiding, is a mystery. The fact that at Niekerkshek it also has radically different forms to those found anywhere else adds to the difficulty.
I will conclude by acknowledging that the populations of H. mutica are all summer flowering but I do not think that this is as significant as the authors of “H. groenewaldii” will contend. The Van Reenens Crest complex demonstrated dramatically that H. retusa and H. mirabilis share a very close common history and an equally close contemporary relationship. My conjecture is that H. mutica is linked to that relationship in the same way the complex of populations are that I assign to H. pygmaea. It is painfully obvious that if floral differences that are so easily detected within populations, expecting to find more evidence in the flowers than exists in the vegetative structures across the genera, and especially the subgenus Haworthia is an absolute pipedream. Floral differences between species are minimal and if anything will provide support for a reduction of species rather than add to the. If any small difference is suggested to separate species, then equal weight must be given to similarities that should then unite species. The contention then that flowers are offering characters that I as an author have ignored, while already wholly untrue, is additionally nonsensical.
Acknowledgement Mr. and Mrs. Trevennan Barry of Van Reenens Crest. Mr. N. Swart of Poststal, Mr. J. duToit of Tarentaal and Wydgelee, Mr. W. duToit of Klipbankskloof, Mr. J. Mathee of Klipbankskloof West, Mr. Vlooi du Toit of Die Kop, Mr. M. Mynhardt of Haarwegskloof, Mr. and Mrs. Nelius Smith of Volmoed, Rondeheuwel, Mr. Adrian Steyn of Wankie Boerdery. Thank you to Kobus Venter and Lawrence Loucka for company and shared observation. ♦
The area concerned is the long and wide contact zone between the Limestone stretching from Bredasdorp to Potberg, and the Bokkeveld shale north of that. The soils and vegetation of the two areas are grossly different. The limestones are agriculturally almost useless, while the shales are prime wheat and pasturage producing soils although relatively low yielding. The vegetation of the shales is Renosterveld and there are very few patches left. Large areas resemble ecological deserts with nothing of the original surface intact. Here and there are shale banks and associated quartz outcrops and also some remnants of tertiary deposits that overlie the shale. Under this deposit layer the shale has decomposed to kaolin and in places there are gravel sheets of fine quartz on white clay. The skeletal nature of these remnants is the saving grace but it is unbelievable to what lengths farmers must have gone to make fields arable. Enormous amounts of stone that have been carted away and dumped to make cultivated lands. Sadly the stone is often dumped on exposed rock and prime Haworthia habitat. The remnants are still under threat and a mindset that has developed in the road construction and maintenance arena is that roads must be clean and scraped fence to fence. Similarly there are farmers who want every square inch under control and in subservience to their production needs. Dense vegetation is abhorred and burnt to control predation of sheep by jackal and lynx. Vegetation adjoining crops is treated with weedkiller to minimize crop contamination. Crops are also grown in conjunction with animal production. When crops are in, the animals are on fallow land and on whatever is left of natural vegetation. It is the harsh reality of conservation.
Haworthias exist on those remnants. I have often pondered that maybe they have not been so severely suffered by habitat destruction but now I am not so sure. There are many remnants where nothing is to be found, and others are species rich for no obvious reason. It is therefore difficult to assess.
This chapter explores the occurrences of two species at 11 localities in the Ouplaas vicinity east of Bredadsorp, and records some of their oddities.
The area in question:
This is shown in the map Fig 1 and I need to re-capitulate what I observed on Die Kop because I do not think I have given proper account for the populations I saw and the variance that I encountered in my first exploration there. This is most unfortunate as the name H. hammeri has since been applied to one or more of the variants and without context. Changes have taken place there too, that now compound the issue. I have in Vol. 3 Pt. 1 given an account of H. mirabilis in the wider area and will not repeat all of it, restricting myself just to those few populations that impact directly in these new observations. What is considerably more relevant is the chapter 13 in Part 2 where the main role players are listed and now again emerging in stark reality..
I have changed the arrangement from that in previous accounts to deal with local groups. Thus three sets, H. mirabilis at Die Kop – 5 populations and re-visits, H. mirabilis at Spitzkop, Haarwegskloof and Langvlei – 5 populations, and H. mutica at Spitzkop,- 3 populations.
Fig 1. Map of area and points covered
SET 1 – Haworthia mirabilis Die Kop
Subset 1 MBB7500 Die Kop E. Figs 2-8
Considering the wide array of populations and variants in the greater area, I concluded that these are most probably the result of interaction between H. mirabilis and H. mutica. The surfaces vary from shiny minutely tubercled to smooth, leaf tips rounded or pointed and flowering time is summer. This particular population is in a deposition area off the Silcrete inselberg.
Fig. 2 7500 Die Kop E
Subset 2 MBB7502 Die Kop N. Figs 9-10
When I first explored here in 2006 I skirted north of the first hill on Die Kop (see fig.1 Map) and observed an array of plants that were more like H. mirabilis and I dismissed them as such. However Jakub Jelimicky and Gerhard Marx both indicated that their observations contradicted mine.
Fig. 9 7502 Die Kop W
Subset 3 MBB7973 Die Kop N. Figs. 11-27
We went further north on this excursion passing through the area touched on in 2006 that was now dramatically different. The previous shrubbery was absent and the area was trampled and grassy with also Bobartia, a large grass-like Irid. It seemed unlikely that Haworthia previously there could have survived, but we had seen them before still further west and they may still be present. This new area we saw was densely populated with plants similar to but smaller than MBB7500.
Fig. 11 7973 Die Kop N
Subset 4 MBB7503 Die Kop mid. Figs. 28-34
My memory of these plants, east of those above, was that they inclined more to H. mirabilis than plants from 7500. Fig. 18 is almost identical to a clone photographed at 7500.
Fig. 28 7503 Die Kop mid
Subset 5 MBB7974 Die Kop mid. Figs. 35-46
Change due to heavy rain and erosion was more apparent here. There was less influence of H. mirabilis and the resemblance to 7500 nearly complete although the plants were generally smaller. This may be because the substrate was more skeletal and eroded than at 7500. What was really curious was the plant pictured figs 45-6 with narrow slender leaves. We did not observe any other species there that could have generated this as a hybrid but we did not have opportunity to comb the entire area small as it is.
Fig. 36 7974 Die Kop mid
Subset 6 MBB7504 N Die Kop. Figs 47-55
This population is about 1km north on an isolated inselberg. The plants are small and more typically mirabiloid with rough surfaces and spined margins and keel. Fig. 55 shows a plant with characteristics of the 7500 plants, but otherwise I would say this population perhaps relates better to Tarentaal MBB6539 (see Updates Vol 3.1 note 17).
Fig. 47 7504 N Die Kop
SET 2 – Haworthia mirabilis Haarwegskloof and Spitzkop.
Subset 7 JDV86-2 Haarwegskloof. Figs 56-63
This subset with subset 11, are included as a reminder of what other populations in the area are like. Originally when so much less was known, this was classified as H. heidelbergensis var. scabra and subsequently so many variants have been found that naming each is not deemed practical by me. It is probably 3-4km west of the following three populations and similar populations occur about the same distance further south at Kathoek and Beyersdal. There are other populations known to me that I have never sampled or photographed and the following task is to explore a large area between Bredasdorp and Spitzkop.
The plants at Haarwegskloof, at the time the pictures were taken in late spring, were both with translucence and without. The plants were small and with leaves erect.
Fig. 56 JDV86-2 Haarwegskloof
Subset 8 MBB7964 Spitzkop West. Figs 64-81
Fig. 64 indicates how difficult it is to find these small plants until you know exactly what you have to look for. One starts the search walking and eventually end upon all fours! Fig. 65 also shows how cryptic the plants can be. The plants are some of the smallest I have ever seen. Fig.72 shows an unusual plant with rounded tips to the leaves and I will show this again in another subset. Why this is so significant is because if the H. floribunda similarity in so many of the populations in the general area. The photographs disappointment me in that my first reaction on seeing the plants, was the association with images of H. parksiana. The pictures do no seem to fully reflect this. Kobus Venter has related his experience of finding H. parksiana-like plants among seedlings of H. mirabilis ‘magnifica’.
The leaves are reflexed as in H. parksiana as they of course are in many populations of H. mirabilis ‘maraisii’. But it is a curious reminder of the floribunda-like leaves in so many populations and the problem of look-alikes.
The plants were on a south slope in an area with white quartz and we had identified this as the most likely place to find them out of the direct north or northwest aspect. But we then did find them there too.
Fig. 64 7964 Spitzkop West
Subset 9 MBB7965 Spitzkop West. Figs 82-86
The plants were again hard to find over quite an extensive area. Fig. 83 again shows the floribundoid leaf tips.
Fig. 82 7965 Spitzkop West
Subset 10 MBB7966 Spitzkop West. Figs 87-103
It is curious how these small localized quartzitic areas can be so different. This was about 500m further north. There was more kaolin present as well as fine gravel. At an intervening similar habitat we found only Acrodon demidiatus, absent from the previous habitats and also from this one where Gibbaeum austricola now appeared. Again all the plants were small with dark green leaves, virtually no translucence and surfaces ranging from mildly spiculed to only minutely tubercled or just rough. Figs 99-101 are of plants with distinctly floribundoid leaves.
These are in white quartz and Aloe ferox is also present. There is moss present which is not the case at Spitzkop but true of Haarwegskloof. Curiously the pinched leaf ends as in fig. 106 have been observed in some plants of H. mutica too.
Although a small area of quartz in a cultivated field, the plants are densely concentrated in small areas. There were some dense clumps in the rocks and this is unusual for H. mutica. It is the variation in these plants that deserves close attention. Some plants have very rounded leaves, they can be quite elongate or short and squat, others have an end-awn, the leaves may be opaque or even translucent; the surfaces may be shiny and minutely tubercled or smooth. Colour may be reddish, golden, or green. Most striking is perhaps the venation. There may be anything from 3-13 straight lines or very anastomizing (veins joining). There is no sign of either true turbercles or surface spination and also none of marginal spines.
Fig. 109 7967 Spitzkop (quartz rock)Gasteria carinata is often severely animal grazed.Looking SE towards Die Kop and PotbergAlso looking SE towards Die Kop and PotbergViewing site from the north
Curiously there is a shale ridge (fig.149) between this population and the first that one would think would be suitable habitat for this species as it is very like the Ouplaas habitat. But they were absent, perhaps because the ridge was lower down the slope. But a little higher where the shale was also exposed in smaller ridges the plants were also absent. Where they did occur was among small quartz pebbles to which the quartz may fragment. Fig. 157 shows a plant that was under the protection of small shrub and shaded.
Plants absentPebbles and plants presentThis is not how one expects to see this species.Fig. 152 7968 Spitzkop (pebbles)Showing the small shale ridgesLooking to the previous habitat of larger quartz rock
Subset 14 MBB7969 Spitzkop East. Figs 160-161
This locality is about 3km east of the previous two at a slightly lower altitude. There is very little quartz but well exposed shale creating apparently suitable habitat as at Ouplaas now a short distance away. The area is well utilized by grazing animals and was very weedy, which may account for the fact that we found only two plants and these were not in the shale where we would have expected them.
Fig. 160 7969 Spitzkop East
Conclusion I must here again relate the story in respect of KG35/70 of which there is only an herbarium specimen. That was found more than 40 years ago at Verfheuwel about 15km eastwards from Spitzkop. I was scrambling up a rough animal track to the top of the Verfheuwel hill when I saw some dried seed stalk less than 10cm high. The plants were tiny, greyish leaved with marginal teeth and rounded tips. I simply did not know what to do with them and identified them tentatively along with a few other populations from that wider area as H. maraisii. I went back to look for them several times with no luck. In 1996 I was with Kobus Venter and we found a strong population of larger plants not far away on the same hill. I recognize now that those first plants I saw were floribunda-like and that this similarity is commonly seen in populations of H. mirabilis in this area.
I speculated in Update 3 about the origins of H. mutica in the context of H. mirabilis and I do contend that these three H. mutica populations show characteristics that support this view. The Die Kop populations suggest to me a more recent amalgamation of H. mutica with H. mirabilis, to virtually reverse the process. The floribunda-like characters and even the parksiana resemblance also support this image of very close common origin and that particularly H. floribunda is simply a polytopic phenomenon in this south western area (polytopic – similar elements arising independently from the same common gene pool) and its genetic resources are embedded in H. mirabilis particularly. This seems to be definitely the case in the greater Swellendam and Heidelberg areas.
What is also so confounding is that now we have H. mutica occurring virtually undifferentiated in several different kinds of habitat. This is quite unlike the Van Reenens Crest area where similar habitat does not even have the same species. The H. mirabilis at Spitzkop west is unlike the other small plants in populations of H. mirabilis in the area and does seem to be differentiated according to habitat.
Acknowledgement: Mr. Johannes and Pat Uys of Die Kop, Mr. Vlooi and Adele duToit of Die Kop, Mr. Gawie de Wet of Spitzkop (West), Mr. Franko de Wet of Spitzkop (East), Mr. Koos Badenhorst of Langvlei and Mr. Martin Mynhardt of Haarwegskloof. ♦