Haworthia Revisited: A Revision of the Genus. (out of print) Umdaus Press, Hatfield, South Africa, 1999 ISBN-10: 1919766081 Taxonomic revision of the genus. Comprehensively illustrated with photographs of both habitat and cultivation. Hardcover. 250 pages 21 x 26 cm.
Foreword
In 1977 Dr D.R. Hunt, formerly of Kew Gardens, London, wrote in the Cactus and Succulent Journal of Great Britain that M.B. Bayer “has emerged in recent years as the leading researcher on Haworthia“. Since the publication of the Haworthia Handbook, now a rare item of Africana, many more books and a vast number of articles have been written on Haworthia, with the popular journal Haworthiad leading much discussion on issues of topical interest. Twenty years later it is without exaggeration to say that Bayer remains the leading expert on Haworthia. His experience of these plants in the field and in cultivation is without parallel and it is consequently a great privilege for me to introduce his new and revised classification of Haworthia.
The eighteen years that have elapsed since the New Haworthia Handbook appeared has been a period of unprecedented botanical exploration of southern Africa, with more and more remote areas becoming accessible and being investigated. Succulents have received their fair share of the attention. Bayer’s previous two books greatly stimulated interest in Haworthia among collectors. To some extent this has caused a “Haworthia-craze” on a smaller but similar scale to the “Aloe-craze” brought on in this country by the monumental works on Aloe by G.W Reynolds. In some cases, this had a very negative impact on plants in the wild but it has also had the effect of stimulating a new ethic of propagation from small numbers of field-collected plants. The present book will keep this interest alive. With the vastly increased success of cultivation and propagation of succulents it is hoped that the destructive collecting of large numbers of plants from their natural habitat is a thing of the past. It is my hope that this book will stimulate competent growers to new pinnacles of achievement on propagating Haworthia, rather than renewed efforts at pillaging their natural habitats.
The great increase in enthusiasm for Haworthia among collectors has brought with it plenty of controversy over names. There is now a plethora of opinions, often strongly and not always wisely held, nor necessarily based on observable facts. Bayer has steadfastly maintained that species must be related to continuity in characters and continuity in distribution of populations in the field and that any other arbitrary selection of individual plants or populations for recognition as species, without placing them properly in their geographical context, is worthless. This rational view enabled him to reduce the “species” from about 150 to 68 in 1976. Since then, the steadily accumulating body of knowledge of these plants in the field has resulted in the reduction of the number of species to 61 in this new book, including 6 new species. It is particularly noteworthy that it has been possible to fit most of the newer collections into the scheme first proposed in 1976. In the new book he sets out to justify the changes made by presenting a much increased photographic record of each taxon and basing his conclusions on material from the now reasonably extensive herbarium record. In addition he has maintained many names at varietal rank so that unusual populations are distinguished taxonomically for the use of the enthusiast. My own experience of these plants in the wild has clearly demonstrated how complex the relationships between and within the different species are.
Bruce Bayer is to be applauded for creating some order in this maze of complexity and crystallizing out of this a further, far more detailed and much expanded account of the genus. This should once more set the standards for the group for many years to come, and also provide plenty of food for thought for those who take the trouble to read carefully what he has written.
Dr P.V Bruyns Mowbray Cape Town
Preface
This book is written for my friends and at their instigation. It is also for those who genuinely like these fascinating plants. I have been extremely fortunate to have been able to spend so much time in the field, and still there is so much to find and see. Despite this, I am confident that this book presents a very sound hypothesis and it contains many lesser and secondary postulates. It is nested within a very wide experience and knowledge of other groups of organisms. The book rests comfortably on the 1978 and 1982 works, and is a very sound foundation for further construction. It is based on a firm definition of what a ‘species’ is, rather than on a personal construct.
Discussion and comment on the issue of the names Haworthia pumila and H. margaritifera could probably form a book in its own right and I could quote a number of other examples too. At the time Scott (and Dr L.A. Codd) worked out typification and application of the name H. pumila. I said I thought they were correct, possibly for the wrong reason. The obvious thing to do seems to me to opt for conservation of the name. It was clear to me at the time that Linnaeus had used the name Aloe pumila to cover five different elements, and I think there should be an Haworthia pumila. By choosing and citing Commelin’s figure 10 to represent his concept of Aloe pumila, Burman (1768) implicity typified Aloe pumila. Haworth (1804) selected Boerhaave’s figure 131 to represent Aloe pumila and, in so doing followed Aiton (1781), who chose the same illustration to represent his Aloe arachnoidea var. pumila. Aiton had created a new illegitimate name Aloe pumiia and he was followed by Haworth and then by Duval. Scott (1985) recognised Haworthia pumiia (L.) Duval (with the support of Codd) ahead of the then fashionable H. margaritifera (L) Haw. Unfortunately Scott overlooked Burman and implicitly used the Aition illegitimate pumila (either as a new name or a Linnaean one). The fact has also been overlooked that H. margaritifera (L) Haw. is based on a Bradley figure which is indisputably H. minima. I had intended to use the name H. maxima (Haw.) Duval as the first and indisputable reference to a species everybody knows by apparently the wrong name. At this very late stage in writing the preface, I have chosen to revert to Scott and to Burman. This is to use the name H. pumila (L) Bayer as a new combination to equate H. pumila (L) Duval sensu Scott. I believe someone has decreed the Linnaean varieties to be typified in some fashion. It is more than probable that this someone is no less fallible than several authorities who have gone before, have made similar mistakes, and also stand correction.
I came to write about Haworthia because of my fascination with plants and because I saw a real deficiency there in the available literature. I have concern that my own honesty; and the accompanying difference and reservation in my writing will not be perceived and respected by readers and other writers. Socrates wrote -The living word of knowledge has a soul of which the written word is no more than an image’. King Thamus used the words ‘only a semblance of the truth’. I have tried my best to come as close to the truth as I can, and in doing this try to observe a fundamental principle of science.
Bruce Bayer
Acknowledgements
For a work of this kind, herbarium record is an absolute foundation and necessity. The work can be no better than the evidence upon which it is built, tested and tried. No opinion in a work of this kind really carries weight unless the evidence can be examined and corroborated. Therefore I owe everything to the institutions and custodians whose work it is to curate botanical specimens and literature. These are the National Herbarium: Pretoria (PRE), the Compton Herbarium (NBG) at Kirstenbosch, and the Bolus Herbarium (BOL), University of Cape Town: Dept. of Botany – and staff. These are people who care about what they do and who set high standards. I am grateful for their tolerance and patience.
Acknowledgement is also made to those who have contributed material and particularly assisted me in the collection process, including particularly Dr P.V. Bruyns and Mr J.D. Venter. Many others have assisted in one way and another and I am deeply grateful, in respect of material contribution or otherwise, to even the least of these.
This work and its completion owe a great deal to: 1. the eternal cheerful optimism and friendship of Mrs. W. Schwegmann of Sheilam Nursery, Robertson; 2. the resolute and steadfast encouragement and support of Kobus (J.D.) Venter, his integrity, and his interest in Haworthia; 3. Steven Hammer for his interest and collaboration, and for the editing which he did on the manuscript; 4. Peter Bruyns for editing and correcting the text; 5. the publisher, Umdaus Press, and specifically François Steffens for his meticulous editing and Kotie Retief for his patient co-ordination; 6. my wife Daphne, and our children, who have managed to endure and even come to share some of my absorption in another world, without asking anything in return.
Readers should please note that the plants on the photographs are denoted by JDV accession numbers. Some of these plants are from collections made by other collectors.
These include:
Dr V.B. Reddi, Dr P.V. Bruyns, J.G. Marx, C. Marais, C. Rowe, V. de Vries, E.J. van Jaarsveld, J. Louw, J. Vlok, C. Burgers, T. Visser, C. McDowell, H. Gie, J. Bouwer, P. Fourie, C. Grobbelaar, J. Meyer, G. Lombard, D. & T. Rossouw, D. Tribble, R. Kent, P. Bosch, E. Dunne, M. Parisi, D. Cumming, D. de Kok, E. Aslander, R. Kratz and E. Esterhuizen. Their cooperation and enthusiasm is greatly appreciated. The contribution of Sheilam Nursery, the Karoo Botanic Garden and Kirstenbosch Botanical Gardens are also acknowledged.
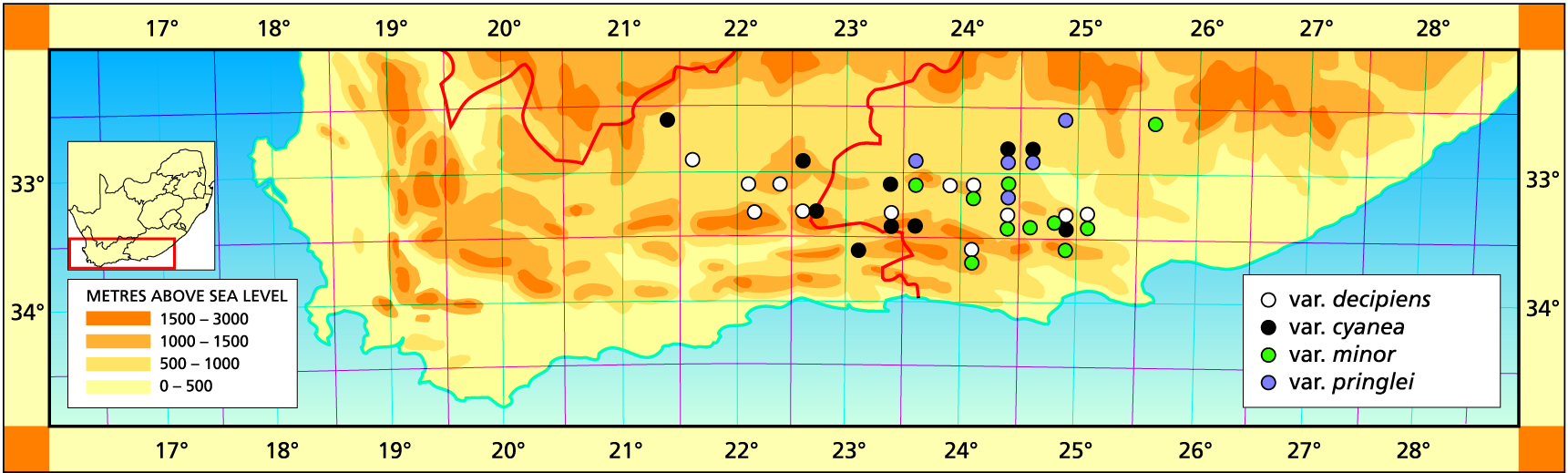
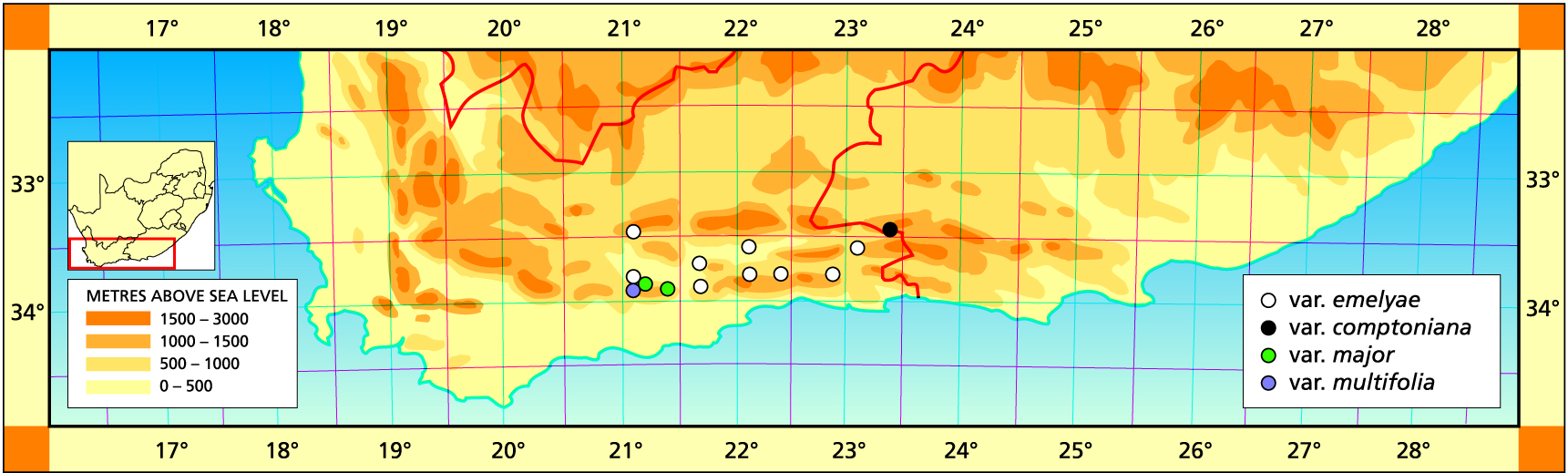
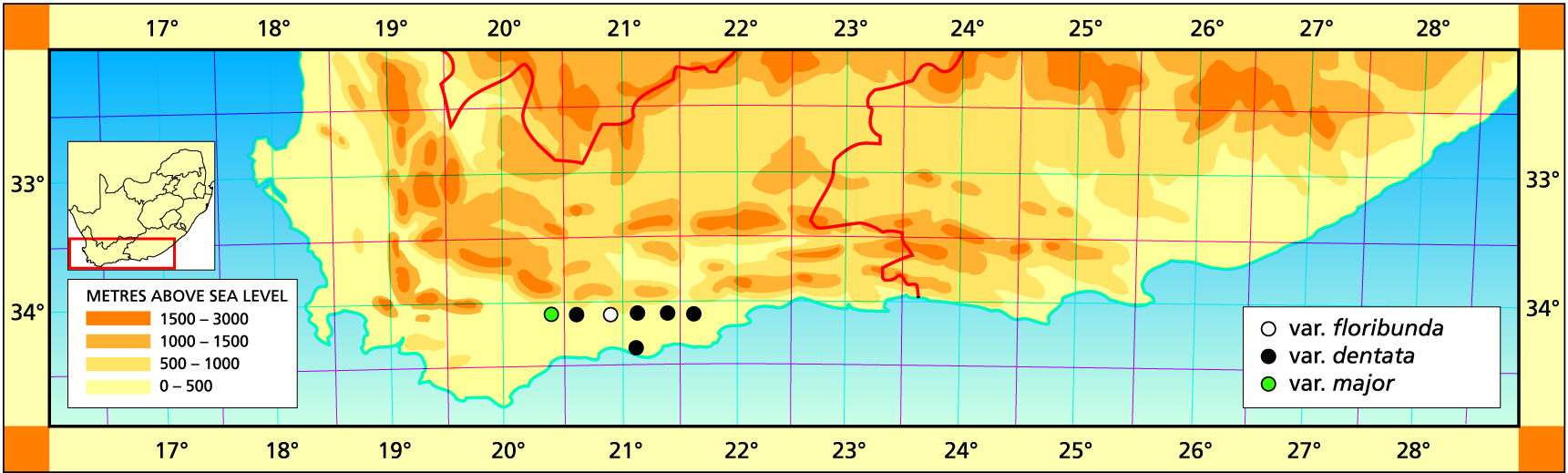
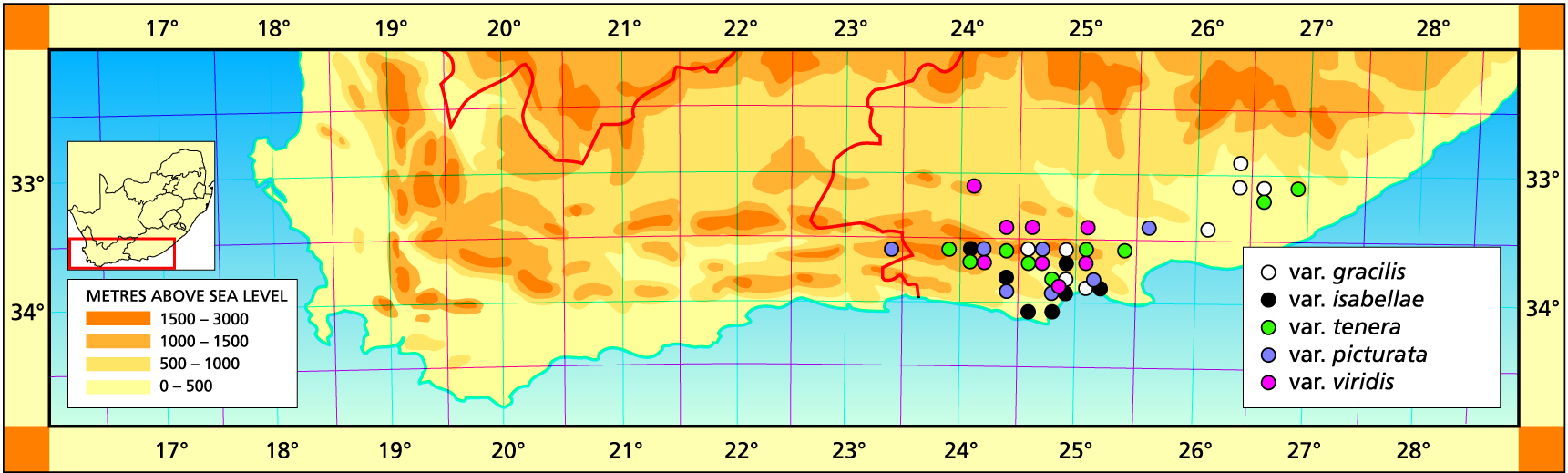
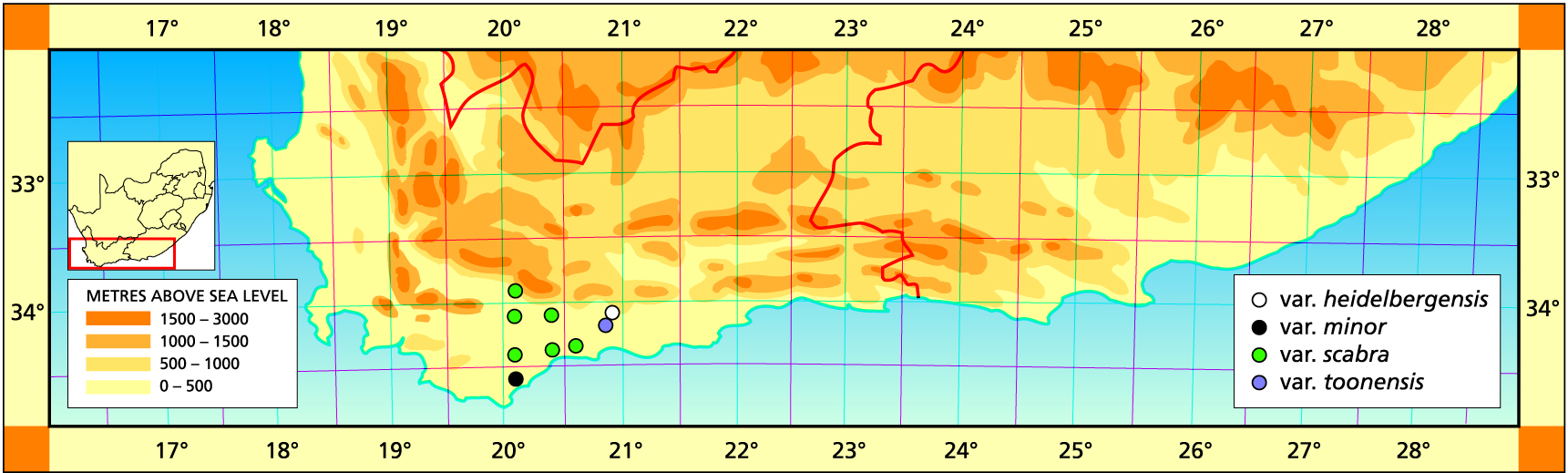
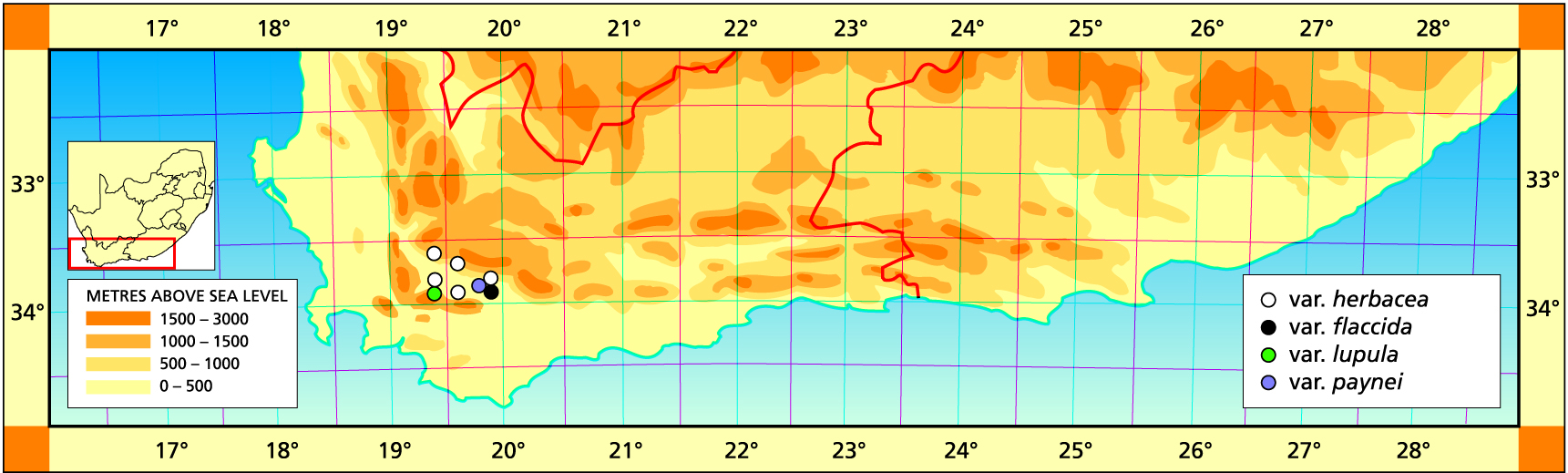
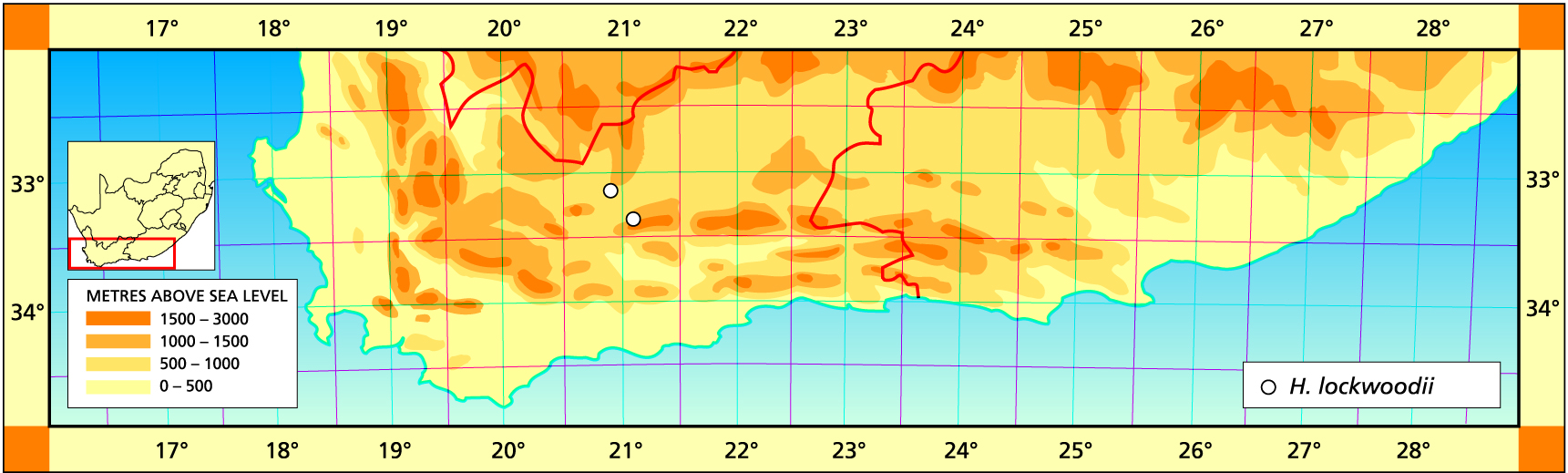
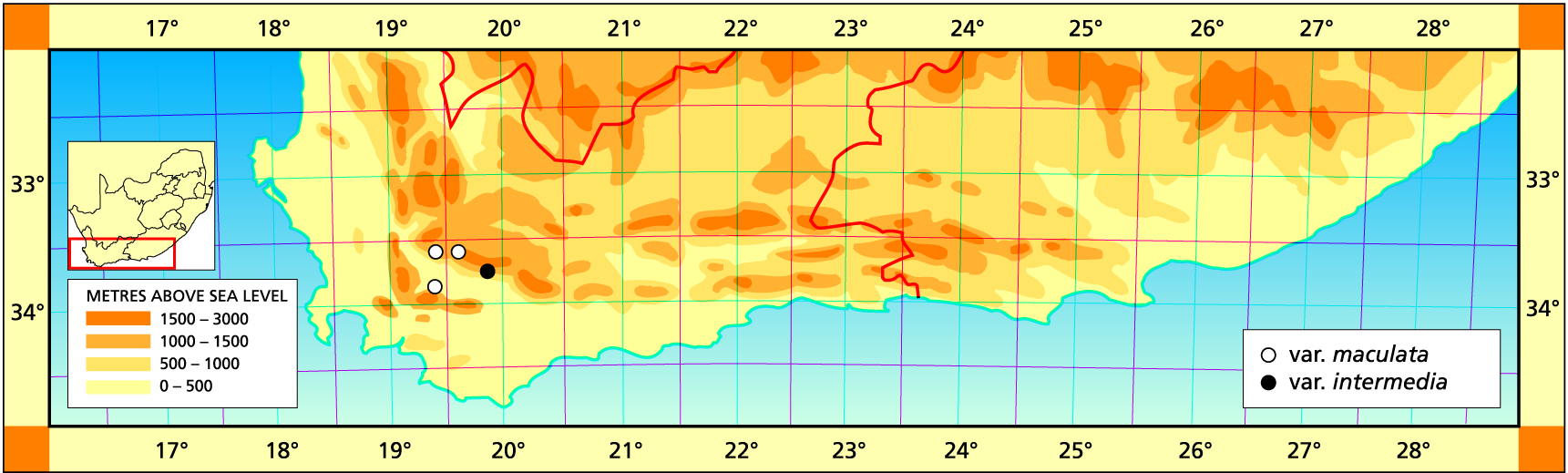
Herbarium specimens are cited according to the system of Edwards and Leistner (PRE), and mapped to the centre points of the quarter-degree blocks, except in a few cases where the centre point represented the wrong aspect or the wrong distance between collection points.
Introduction to The Haworthia Handbook – 1979 Version
Interest in Haworthia has been very largely an amateur one, and this interesting succulent genus has a large following among the ranks of collectors. Names are critical to collectors as they are indeed to any attempt to observe and record information concerning the plants. There is an extensive literature to Haworthia and over 400 different names have been used at one time or another to refer to species, varieties, etc. The present publication is an attempt to review the history and literature of the genus and present a list of names which will be of practical value in dealing with it. This is by no means a definitive work and the writer does not pretend that it is anything but a simplistic view of a complex situation. It is however based on an extensive knowledge of the plants in the field derived from the records compiled by G.G. Smith and from the writer’s personal experience. The work was in fact made possible largely by the systematic and dedicated contribution made by G.G.Smith. The illustrations are all from photographs taken by the writer of living plants mostly from the original type localities where these are known.
In many cases more than one photograph is required to adequately illustrate a species and even colour photography cannot adequately demonstrate differences between species. The illustrations thus do not reflect the real diversity in the genus, and similarly the checklist does not express this diversity adequately either. Apart from the difficulties in applying the ranks of subspecies, variety, forma consistently, there are several populations known to the writer which cannot be confidently included in any given species and thus could merit independent species status. The rank of forma has no particular significance other than to express a particular form in a species where there may in any case be a high degree of variability. Many of the names rejected in this work could easily be resuscitated at this level. An attempt has been made to use varietal rank to denote populations with some geographically associated differences, and subspecies where such a distinction is greater.
The keys to the species must be regarded only as approximate particularly in the subgenus Haworthia where many of the couplets are indeed weak. This is particularly true towards the end of the key. The nature of the available superficial characters and their variability make it virtually impossible to construct any infallible key. The nuances of form, colour and armature and changes brought about by differences in growing conditions preclude an accurate and really worthwhile key. It is believed that the photographs will serve a better purpose than the possibly misleading keys. No attempt has been made to divide the subgenera into sections for much the same reason. The subgenera Hexangulares and Robustipendunculares are small enough to handle without sections, and in the case of the former the natural divisions may fragment it excessively. In the subgenus Haworthia there are several small natural sections leaving a residue of irreconcilable species.
A proper understanding of the genus Haworthia will have to be based on anatomical and ecological grounds, and this account is presented in the knowledge that a more objective technical account is desirable. In all, 68 species are recognized together with 47 subspecies, varieties and forms.
Introduction to The new Haworthia Handbook – 1982 Version
The original title of the Haworthia Handbook should perhaps have read ‘Illustrated Index of Haworthia names’. However, the Handbook was intended to serve a specific purpose in bringing all the names and literature of previous years together. The purpose of the second edition is to present an up‑to‑date version with necessary corrections and alterations in a new format. It is based on the G.G. Smith collection from the East London Museum acquired on loan and now housed in the Compton Herbarium at Kirstenbosch. Dr M.Courtenay‑Latimer was instrumental in obtaining quite a considerable quantity of papers, drawings and photographs from G.G. Smith’s estate. The collation of all this information and herbarium records has provided an ideal opportunity to review species concepts in Haworthia, and establish where problems exist. This was followed by many field excursions and new collections, which have all tried and tested the writer’s application of names. C.L. Scott has published several further papers which have contributed some useful ideas and called for further explanation. Notable here is the application of the older names H. pumila and H. altilinea. The latter name is not accepted in this work, but the former is. A further consequence of Scott’s work is that the name tessellata is resurrected in the place of recurva, but still as a subspecies of H. venosa.
Recent experience and observation have not led to any real solutions to the problem of recognising species in Haworthia. They mostly tend to confirm the views expressed in the first edition, in the chapter on genus and species concepts. This includes the view that species intergrade across geographic barriers and vary across ecological barriers. Very few of the more widely distributed species can as yet be clearly circumscribed to establish infallible identifications. For example, H. cooperi clearly intergrades with H. bolusii. H. bolusii has small forms which are to all intents and purposes similar to H. aranea. The small forms of H. bolusii lead, with little imagination, to H. translucens ssp. tenera. Similarly H. bolusii apparently transforms into H. translucens in the mountains near the southern boundary of its distribution range.
It has long been clear that a taxonomic solution may be possible for species together in one locality, and wrong in another. Thus H. herbacea and H. reticulata grow together and hybridise northwest of Worcester, but southwest of Robertson there are populations which cannot confidently be ascribed to one or the other species. Similarly this complex of H. reticulata/herbacea intergrades into H. turgida. The intergradation unfortunately is not through a continuous series of plants, but leaps across from isolated population to isolated population, creating an irregular pattern difficult to unravel into separate species.
The relationship between H. turgida and H. retusa is possibly more complex than the above, and is perhaps a contest of small/proliferous versus large/solitary plants. In other species too, there is variation coupled with distribution, and variation coupled with habitat. In an area of broken terrain and diverse geology this will be aggravated, particularly in low rainfall areas and on skeletal rocky soils. In some cases this is easily demonstrated in the field, e.g., H. turgida north of Heidelberg, H. habdomadis west of Ladismith, H. bolusii east of Jansenville, H. cooperi at Chalumna and at Kayser’s Beach. In other cases the presence of other species, or more dramatic differences, confuse the issue. H. decipiens, H. bolusii, H. habdomadis, H. cymbiformis and H. cooperi all diffuse into the mountains of the Baviaanskloof, and the Groot and Klein Winterhoek, merging with H. translucens and presenting something of a nightmare. Dramatic vegetative differences occur in the case of H. starkiana and H. scabra which are both closely related and practically sympatric; H. marginata and H. pumila occur practically sympatrically as do glabrous and hairy forms of H. translucens ssp. tenera. H. radula and H. attenuata grow mixed together and may in fact be only forms of one species. H. magnifica can sometimes be confused with H. retusa, which may be a true or false situation, i.e., there may be plants or populations which really are intermediate and neither one nor the other species. The same applies to H. magnifica and H. mirabilis.
Field hybridisation is definitely not the serious problem it has been made out to be. The number of known field hybrids is really minimal and the writer knows only the following:
H. pumila X Astroloba muricata (= Astroworthia bicarinata) H. pumila X H. marginata H. minima X H. marginata H. turgida X H. floribunda H. graminifolia X H. arachnoidea H. herbacea X H. reticulata H. cymbiformis X H. angustifolia H. scabra X H. viscosa H. viscosa X H. longiana H. truncata X H. arachnoidea
There are, of course, a number of difficulties such as H. fasciata fa. browniana, H. glauca var. herrei fa. armstrongii, and H. angustifolia fa. baylissii, which may possibly be the consequence of hybridisation. The low incidence of hybridisation is a situation to be expected in a relatively young genus, as it would be difficult in the first place for species to arise until some form of isolation becomes entrenched. Hybridisation is more of a secondary mechanism operating where established species meet. In Haworthia there is definitely no sound basis on which problems of variability and identification in the field can be attributed to hybridisation.
This second edition, like the first, makes no attempt to meet the need of the collector in covering all those odd names which have arisen over the years. Many clonotypes of species described by Von Poellnitz, Uitewaal, Resende and Smith are still cherished in collections. There are also many plants bearing names by Haworth, Salm‑Dyck and Baker which may even be reasonably accurately identified. Descriptions and illustrations are available in the literature and wait for another enthusiast to collate and present.
In this revised addition, three new species replace three species which cannot be further upheld, and there are several changes in the lesser ranks. Altogether 68 species are again recognised, with 47 subspecies, varieties and forms. These species are here grouped into 3 subgenera where they are arranged alphabetically and not in any phylogenetic order.
Introduction to Haworthia Revisited – 1999 Version
The Handbook was first published in 1973 and revised in 1982. It now seems timely to revise the work again, especially in view of the very extensive interest in Haworthia, and all the collecting which has been done since then. This has led to new perceptions, and the object will be to try and synthesise these in a consistent way in a new arrangement. It appears to me that much of the new material and evidence substantiates the robust nature of the old Handbook treatment, although many of the new finds cannot be accommodated comfortably in the existing nomenclatural framework.
The Handbook was originally written to ‘present a list of names which will be of practical value in dealing with it’ (i.e., communication – the history, literature and cultivation of the genus). There were a number of very substantial differences between my treatment and that of Col C.L. Scott and this is an honest attempt at reconciliation. Generally these differences will be treated in the text as they arise and there is no associated intention to be belittling in the process of doing this. Some differences are purely due to the fact that my work did not address some tricky nomenclatural problems. In some cases they are real differences of opinion and perception, and in others there does not seem to be any good explanation for gross divergence of view other than intransigence on both sides. I hope that readers will appreciate that their own perceptions as expressed in multifarious different journals, newsletters, round robins, etc. are very often also far removed from truth. These expressions are probably driven by the same misguided feelings and fervour common to us all.
I certainly erred in not recognising the differences between the Uniondale Haworthia bayeri and the westerly H. emelyae. I was aware of difference in flowering time but was so preoccupied in trying to establish H. emelyae as a valid entity (as opposed to ‘picta’ and ‘correcta’) which also took account of H. comptoniana that I just missed the obvious. The debate still continues. Which goes to show that a lot of effort is wasted in unresolved conflict, while the need for stable reference points comes second. I did come to see that there were two species present at the locality for H. comptoniana near Georgida, and David Cumming also astutely intimated that there were two species. There are a number of other instances where I really was not certain about what path to take, and the reasons will, I hope, become clearer on close reading of the chapter on species concepts – and perhaps in the main text also.
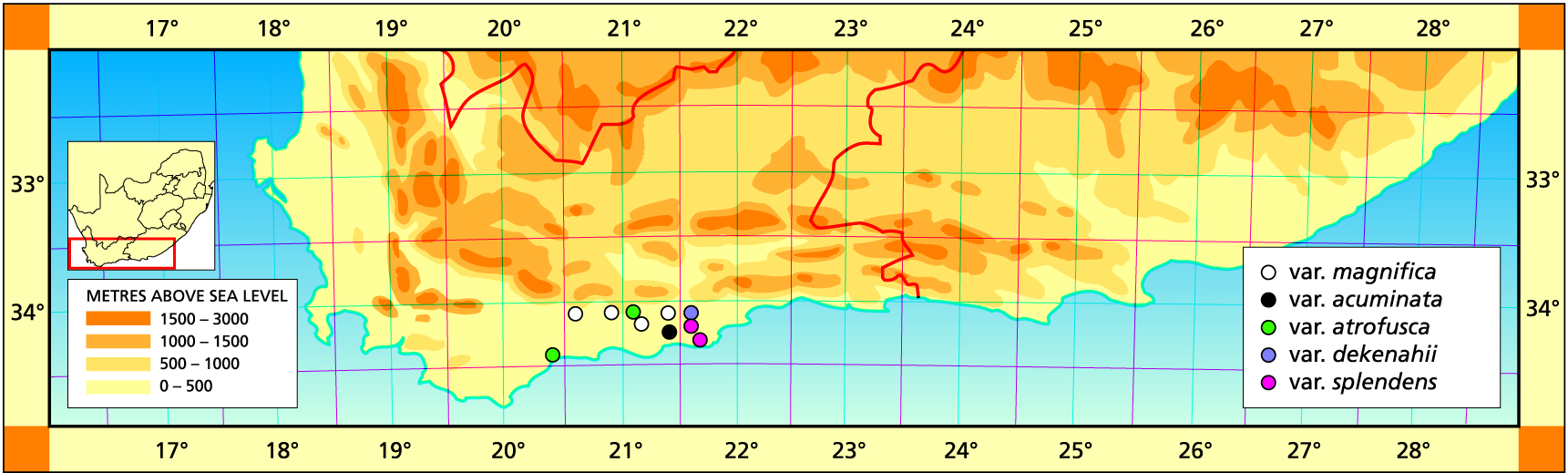
I made a number of other errors too and these are much more obvious in hindsight. A particular case is with regard to H. dekenahii Smith and H. argenteo-maculosa Smith. In reducing them first to synonomy and then to varietal rank, I unnecessarily confounded a real problem. As pointed out above, the distinction between H. retusa and H. magnifica is problematic and the two varieties mentioned are in the trouble zone. Subsequent to the Handbook I also may have generated some problems by guessing at the identity of some troublesome finds.
When I worked on Haworthia there were very few fellow collectors. Much of my effort was directed at establishing what had in fact been collected and where. I was, and still am, more comfortable with generalising than with specifying. I was sometimes quite comfortable with tentatively identifying a collection and adding a locality to indicate that I was really uncertain eg, H. emelyae var. ‘montana’ – Swartberg Mts, which is now H. vlokii. Similarly, quite a few of my collections were identified with a two-way arrow because I could not be sure to which of two species the plants could belong. Since then the quality and extent of collecting has indeed been remarkable, although it is probably only P.V. Bruyns who has followed the proper route of herbarium documentation. J.D. Venter’s is the next best properly documented collection which, while frequently lacking the formality of the herbarium record (for which lack of herbarium space is the main reason) will at least ensure that information is preserved for posterity. These additional collections have helped a great deal in clarifying some points.
However, it appears to me that identification is always going to be a problem in Haworthia, and a sad truth that a workable key for Haworthia is an unattainable ideal. Taxonomy of succulent plants is often criticised and the amateurs who indulge are scorned for their efforts. However, there are probably many great taxonomic works by revered botanists which never receive a fraction of the critical examination that the popular genera do.
There has been a large degree of excessive, illegal, and disconcertingly self-interested collecting which has made and will make no contribution to the scientific and herbarium record. Whatever growers and collectors have to say about nomenclature and much of the necessity and nonsense that goes with it, this is what provides the reference framework for all their communication.
I have to pretend to have some knowledge of taxonomy and deep insights into genetics and speciation. These I do not have. My understanding is based on my life experience and is limited by my intellect, so it would be naive to think that everyone will agree with my perceptions. It has been disappointing to me that people better equipped than myself have often not come nearly up to my expectations. There seem to be other factors than training, education and intellect which determine the arrival at mutual and then good understanding and truth. Many decisions are by the nature of things subjective. It takes some effort to surrender one’s own opinions, especially when no good reason appears to substantiate the alternative decision. This kind of difference is illustrated in the description of H. reddii, where the population was well-known to me and referred to in the Handbook.
The Haworthia Society based in England first published its Newsletter in 1986, and this has continued as Haworthiad. This small magazine has done much to recapture the interest in Haworthia and bring it to the level it appears to have enjoyed during the years preceding World War II. My wish is to see the interest and skills of my collaborators combined as a tribute to a delightful group of plants, for whatever reason they attract the attention and interest that they do.
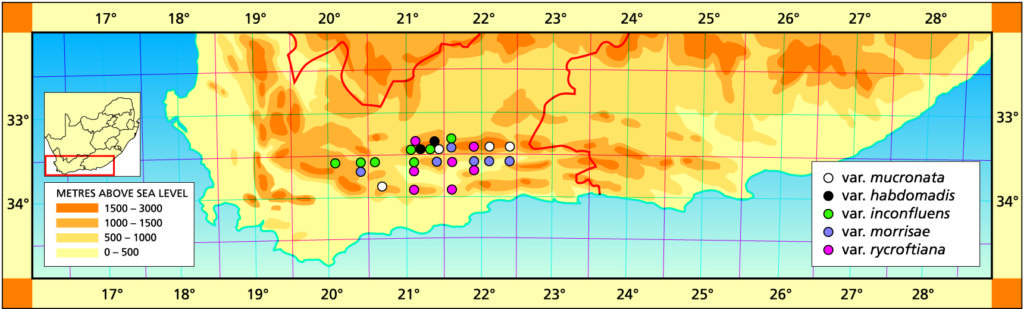
The list of natural hybrids can be extended to include: H. sordida X H. woolleyi H. blackburniae X H. arachnoidea H. viscosa X H. fasciata H. viscosa X H. glauca var. herrei H. monticola X H. mucronata var. inconfluens
Many commentators in the general literature on Haworthia have since commented on the ease of hybridization across even the sub-genera. More important to me are those rare discerning commentators who have used this field to substantiate that the three groups are real and not just a product of Uitewaal’s or my own wishful thinking.
In this newer revised addition, 6 new species names appear while 13 are no longer upheld, and there are many changes in the lesser ranks. Altogether 61 species are recognized, with 105 lesser ranks. These species are again grouped into the 3 subgenera, arranged alphabetically within the subgenera.
Historical Sketch
1982 – The first written record concerning Haworthia appears to be that of Oldenland, superintendent of the Dutch East India Company garden at the Cape Colony in 1695. This record includes a list of 28 ‘aloe’ species and is found in Valentyn’s Beschrywinge van der Kaap der Goede Hoop (1726). These descriptions appear verbatim in Commelin’s Praeludia Botanica (1703) with amplified descriptions, and also illustrated. Four of Oldenland’s ‘aloes’ are now placed in Haworthia; these are:
’23 … Aloe africana arachnoidea’ (Commelin, fig. 27) now Haworthia arachnoidea (L.) Duval, although this name has not been previously associated with any field population. It is considered that the name is linked with H. setata Haw.
’25 … Aloe africana folio non nihil reflexo, floribus ex albo et rubro variegatis’ (Commelin, fig. 29) now H. venosa (Lam.) Haw. despite mention of white and red variegated flowers.
’26 … Aloe africana folio in summitate triangulari, rigidissimo marginibus albicantibus’ (Commelin, fig. 30) now H. marginata (Lam.) Stearn.
’27 . .. Aloe africana erecta, triangularis et triangulari folio viscosa’ (Commelin, fig. 31) now H. viscosa (L.) Haw. Commelin, in an earlier work of his, Horti medici Amstelodamensis (1701) described and illustrated four other Haworthia. These are:
‘Aloe africana brevissimo crassissimoque folio flore subviride’ (fig. 6) now taken to be H. retusa (L.) Duv. ‘Aloe africana folio glabro et rigidissimo flore subviride’ (fig. 9) which is perhaps best regarded as H. marginata also.
The two descriptions:
‘Aloe africana folio in summitate triangulari margaritifera flore subviride’ (fig. 10) and ‘Aloe africana margaritifera minor’ (fig. 11)
– are here both regarded as H. pumila (L.) Duval. Commelin distinguished the latter by the presence of tubercles on both leaf surfaces and being half the size of the former. These differences are not incompatible with known variability in H. pumila. The differences in the taste of the sap of the two elements is inexplicable as no real difference can be readily detected in the sap of H. pumila and H. minima (Ait.) Haw., as the only species which can be considered in this context. The statement that the two elements grow together suggests that but one species is involved.
Reynolds’ (1950) conclusion that Commelin used Oldenland’s Kruidboek verbatim regarding Aloe species, is thus not true for Haworthia. Neither H. retusa nor H. pumila are listed by Oldenland, and Commelin did not consider Oldenland’s n.26 to be the same as his fig. 9 (H. marginata). In the case of H. retusa and H. marginata, Commelin stated that at least the seed capsules were drawn from a book by (or compiled for) Nicolaas Witzen, Mayor of Amsterdam and Director of the Oriental Institute in about 1692.
Notable additions to this total of 6 species are in Commelin’s Praeludia Botanica (1715) where ‘ … margaritifera minima’ is taken to be the first real reference to H. minima; and Boerhaave’s ‘ … minima, atroviridis, spinis herbaceis numerosis ornata’, n.40 in his Index alter Plantarum (1720). This latter description is now interpreted as H. herbacea. These eight species were treated in various ways but without addition until De Candolle described Aloe rigida (1799) and Jaquin A. radula (1804). That so much confusion could ensue from such an elementary breakdown becomes understandable when it is realised that at least ten distinct segregates of Haworthia can be found in the Robertson Karoo, and a further eight in the South‑western Cape area alone. The Commelin and Boerhaave figures were probably regarded as a complete portrayal of the smaller Aloineae and subsequent collections identified accordingly.
The value of historical records of the early exploration of the Cape in elucidating taxonomic problems in Haworthia must be regarded as dubious. Even Reynolds, despite an elaborate account of early botanical exploration, cannot provide earlier references to Aloe microstigma Salm‑Dyck (1854) and A. speciosa Baker (1880), which are both abundant in relative close proximity to the Cape Peninsula. Conversely Reynolds could not relate the record of A. humilis (L.) Mill. by Commelin in 1701, to exploration. Both this aloe and H. viscosa (in Oldenland’s Kruidboek) occur at their nearest point to the Peninsula, at Calitzdorp and Barrydale respectively, both situated in the Little Karoo. From available accounts, the area as far east as Swellendam was well‑known by 1676, while Cruse had already travelled overland from Mossel Bay to Cape Town by 1668. If historical records are of any value, there is no good explanation for the appearance of H. mirabilis Haw. as late as 1804, or for that matter Poellnitzia rubriflora (Bolus) Uitewaal in 1920. Specimens of H. mirabilis may, of course, as already suggested, simply have been placed under H. retusa, while P. rubriflora could very well have been Oldenland’s ’28 … Aloe africana, folio parvo et in acumen rigidissimum exeunte’. The description of H. reticulata Haw. very common in the Robertson Karoo, as late as 1812 also emphasizes the disparities between chronological progress and botanical discovery. The suggestion by Scott (1970) in his description of H. springbokvlakensis, that H. mutica Haw. was probably collected by Bowie at Mossel Bay, also demonstrates the fallibility of this kind of historical evidence. A species (eco‑type?) barely distinct from the Mossel Bay element is common in the south‑western Cape, and at two localities, adjoins deeply rutted wagon transport routes between the Cape and Swellendam. The basis of Haworth’s nine new species described in 1804 is apparently to be found in the collecting by Thunberg and Masson. However, propagation from seed was practised from the inception of the Company garden at the Cape, and Miller (Gardener’s Dictionary, 1768) referred to a variety of H. herbacea raised from seed. It is thus certain that artificial hybrids were intruding on the taxonomic scene at an early stage. Willdenow described H. fasciata (Willd.) Haw. in 1811, Salm‑Dyck H. papillosa in 1817, and Haworth added another 13 species in 1819. Most of these additions by Haworth were of plants from Kew Garden, as were the six species added in 1821. In 1824 Haworth added a further six species resulting from collection by Bowie (1816‑1822), who undertook four journeys and travelled as far afield as Grahamstown eastwards, and Graaff-Reinet to the north. Bowie’s contribution extended to three more species by Haworth in 1825 and one more in 1828. Several of Haworth’s species and varieties are referable to Salm‑Dyck too, whose Monographia was published in seven parts between 1836 and 1863. No significant additions were made until Baker published some 17 new species in 1880, all stemming from collecting done by Cooper. Cooper was in South Africa from 1859 to 1862 and Reynolds briefly records that Cooper resided at Worcester before moving to Algoa Bay, inland to Graaff-Reinet and then through the Orange Free State to Natal. There does not seem to be much hope for determining the precise geographic origin of Baker’s species and hence elucidating some of the problems these involve. Baker was responsible for the revision of the whole genus in Thiselton‑Dyer’s Flora Capensis (1896). This was followed by Berger’s monograph of Aloe in Engler’s Das Pflanzenreich (1908). New species of Haworthia were published by Wright (1‑1907), Marloth (2‑1908, 1912), Schonland (2‑1910, 1912), Archibald (2‑1940, 1946), Barker (1‑1937), Resende (5‑1938, 1946), Fourcade (1‑1932), Scott (4‑1965, 1968, 1970, 1979), Hardy and Mauve (1‑1967) and Bayer (6‑1972, 1973, 1981). More extensive contributions were made by Von Poellnitz from 1929 to 1940, Smith, 1943 to 1950 and Uitewaal, 1939 to 1948. J.R. Brown published many commentaries on Haworthia and in 1957, in “A brief review of the genus Haworthia”, noted the existence of 160 species and 210 varieties! An attempt to catalogue these names was made by Jacobsen in his Handbook of Succulent Plants vol. II, published first in German in 1954 and in English in 1960. This was followed by a review of the sections and the species comprising these by the same author in Kakteen und andere Sukkulenten (1965). In the now vast and often inconsequential literature on Haworthia, mention can be made of the Haworthia Review, a roneoed publication of ‘The Succulent Liliaceae League of America‘. This first appeared in 1946, was edited by J.W. Dodson and one number was accompanied by a comprehensive check‑list. Also of passing interest are the publications by Farden and Zantner, and more recently by Fearn, Pilbeam (including booklets, The first 5O Haworthias and The second 50 Haworthias), Speechley, Scott and Bayer in publications of English, American and Australian cactus and succulent societies.
The genus has not lacked interested researchers so much as good observation of the plants, their affinities and distribution in their native habitat. It is particularly unfortunate that G.G. Smith discontinued his work at a critical stage when he perhaps could have made a major contribution. In 1947 he wrote ‘Views on the naming of Haworthia’ published in the Journal of South African Botany, in which he presented some of the difficulties arising from lack of authoritative field work. If anything this was a plea to be allowed to work to a conclusion, unhindered by duplication and developments elsewhere. Resende took violent and personal exception to Smith’s comments and replied in acrid tones in ‘Variability and taxonomy in the genus Haworthia’ in Portugaliae Acta Biologica (B) (1949). This attack undoubtedly led to Smith’s retirement from the field. Despite being responsible for several errors of the same magnitude criticised in his paper, Smith did leave an unrivalled legacy of herbarium specimens, photographs, field notes and literature compilation. Von Poellnitz was killed during the allied bombing of Berlin in 1945, when a straying bomber pilot mistook his baronial castle for an industrial target. Familiarity with his work arouses some sympathy for the errors which he made. Obviously open‑minded, with a willingness to concede misinterpretation and change, it is conceivable that he could eventually have revised the genus adequately. Most of his material was destroyed during the war, but fragments are still available in the herbarium at Berlin (see Appendix I). Flavio Resende was a highly regarded Portuguese botanist who published several papers in Portugaliae Acta Biologica, Broteria, Boletim da Sociedada Broteriana, and Memorias da Sociedada Broteriana. Nearly all his new species and varieties were of unknown or garden origin and included polyploid hybrids. His work is thus respectfully discounted in this review.
Collectors and Contributors
1976 – The role played by private collectors in adding to information on Haworthia has been a considerable one although not ever fruitfully collated. The revival of interest in the genus stems from Dr Karl von Poellnitz working at his home at Oberlodla in Germany, coupled with the collection by Hans Herre at Stellenbosch, F.R. Long at Port Elizabeth, J.R. Brown in America and Wilhelm Triebner in South West Africa. Most of the plants received by Von Poellnitz were routed through Herre, Long and Triebner. Herre’s contribution in the field of South African succulents generally, is very well known and does not need elaboration. As an enduring record, very little remains of an accessioned and labelled Haworthia collection at the Stellenbosch University Garden. The accessions are rather too sparsely recorded to be of much value in tracing localities, which are with the wisdom of hindsight, more important than then appreciated.
Triebner was a nurseryman at Windhoek ‑ an indefatigable collector and enthusiast ‑ his contribution is notoriously inaccurate and unreliable. Many collections were confused, localities and collectors also frequently being incorrect. Plants distributed by Triebner were on many occasions misnamed, although it is quite certain that Triebner was not entirely at fault. Many independent observers relate the experience of submitting plants to so many recognised authorities and receiving that number of different identifications. J.R. Brown, in correspondence with Long, noted that names of one‑third of plants sent for re‑identification did not agree with previous naming by Von Poellnitz. Long was superintendent of parks at Port Elizabeth from 1930 to 1940. Taking a keen interest in Haworthia, he built up an enormous collection which he did not administer himself. In a protracted and sporadic correspondence with Brown, he made very little contribution in the way of constructive and informative comment. In fact, apart from a compilation of photographs and descriptions, the written record of his activities over a period of 25 years is disappointing. Had it not been for Dr H.G. Fourcade, forester, surveyor and botanist, it is likely that Long’s contribution would have gone completely unrecorded. Dr Fourcade saw Long’s collection in 1938 and recognised the importance of a collection containing co‑types of nearly all Von Poellnitz’s species. With the help of Miss Eily Archibald, a check-list was prepared and Fourcade took 230 photographs of plants in Long’s collection. Six sets of photographic prints were eventually prepared and, as intended by Dr Fourcade, form a useful reference in further study of the genus. With the outbreak of the second world war, Long became involved in the construction of airfields and his plants were left in the care of Mr F.J. Stayner (later Curator of the Karoo Botanic Garden at Worcester).
After the war, the entire collection was handed to Mr G.G. Smith, engineer, businessman and naturalist at East London. Smith collected extensively and contributed many new species and varieties. Despite the vast record handed by Smith to Kirstenbosch, together with his collection in 1957, there was very little to indicate in what direction his concepts of Haworthia were developing. Several notes indicated that he had new species and varieties in mind for publication, and also a few deletions. It was only with the recent acquisition of Smith’s papers, that it was possible to see that he was contemplating a whole series of species and varieties. He was proposing 13 new species, 18 varieties and 2 subvarieties in the H. cymbiformis complex alone, while in the H. viscosa complex he contemplated 4 new species and 14 varieties. In all there were 18 new species, 65 varieties, 4 subvarieties and 1 forma in draft manuscript. Contributors to Long’s collection included nearly all the persons commemorated in Von Poellnitz’s species names. G.J. Payne of McGregor, a pianist, church‑organist and collector of sheet‑music, sent plants mostly to Triebner, and later to Long and Smith. Mr Rossouw was a policeman stationed at Napier who collected with Payne. P.L. Meiring was a speculator in land and a horticulturist, who also sent plants to Triebner, Herre and Long. Hurling and Neil were two bachelors who owned the Winton nursery at Bonnievale and who submitted many plants to Kirstenbosch, Triebner and Long, for naming. Mrs Dora van der Bijl was president of the now defunct South African Succulent Plant Society, and lived at Great Brak. Mrs E.D. Helm was also a house‑wife at Great Brak near Mossel Bay and was responsible for the discovery of H. parksiana V. Poelln. Mrs Emely Ferguson, wife of a surveyor, lived at Riversdale. Miss G.V. Britten was the curator of the Albany Museum herbarium at Grahamstown, where Miss G. Blackbeard also lived. Mr C.H.T. Woolley resided at Addo, Mrs K.E. Jones at Steytlerville, Mrs H. Blackburn at Calitzdorp, W.A. van Ryneveld at Somerset East, M. Stiemie at Kirkwood, Mrs E.B. King and her son Gordon at Port Elizabeth, Mrs E. Braun at Kendrew, W.R.B. Marais at Beaufort West, Mrs L. Lategan, Mrs G. Morris and L. Smith lived at Oudtshoorn, Miss A. de Klerk at Tulbagh and Major H. Venter collected in the South‑western Cape. Several collectors operated fairly independently and these include M. Otzen, whose localities were fortunately recorded by Dr J. Luckhoff. A.J. Joubert was a teacher at Ladismith, as was C. Beukmann who retired at Bonnievale. G. Mclaren appears to have been involved in a nursery operation with L. Leighton near Stutterheim and may have been responsible for some of the localities dubiously recorded at Grahamstown by Von Poellnitz. S. Lockwood‑Hill was for a period magistrate at Laingsburg and W.E. Armstrong, stationmaster at Adelaide. As Smith took over Long’s collection, naturally many of these persons are also involved in Smith’s personal records. Here Dr M. Courtenay‑Latimer’s contribution was certainly considerable although not recorded to any degree. J. Dekenah, photographer and naturalist at Riversdale, also helped Smith considerably and his localities are the best recorded of all. Dr Nortier of Clanwilliam sent in plants from the Western Cape as did Miss E. Esterhuysen of the Bolus Herbarium, Cape Town. Mrs A. Taute lived at Molen River between Oudtshoorn and Uniondale. Prof. R.H. Compton was, of course, Director of the National Botanic Gardens at Kirstenbosch, where H. Hall was curator of succulent plants. Mr L.A. Malherbe, owner of the Sheilam nursery at Robertson, was specifically interested in Haworthia and submitted the original specimens of H. comptoniana Smith. Captain D.R. Keith of Stegi, Swaziland, also sent plants to Smith.
Several overseas collectors built up good collections from which relatively little useful information has ever flowed. This, together with the failure of local collections to provide any permanent and reliable record, should be a clear indictment of collecting per se as a corollary of preservation and scientific record. A specimen without a locality record is a liability in taxonomy, and Brown records his disappointment at Von Poellnitz refusing to accept plants from America for certainly that reason. Any plant considered out of its natural habitat and without reference to the population from which it comes, particularly in Haworthia, has to be regarded with caution. Thus even J.R. Brown’s admirable commentaries and photographs are at most a reflection on the contemporary taxonomy and interchange between collectors, rather than a contribution to further order in the genus. J.T. Bates, tramway conductor of Hounslow, England, was in a better position as many of Von Poellnitz’s plants found their way into his collection. Whether his collection will have retained its value as a privately administered record of Von Poellnitz’s work is doubtful. Brown felt that even the Royal Botanic Gardens at Kew had failed in providing any enduring record of Baker’s work, and Von Poellnitz must have felt this too regarding Wright’s H. pearsonii. This role of a botanic garden in relation to preservation of types is a vital one where herbarium material cannot (by ordinary methods) be depended on as a certain clue to identity. However, the conservation of a species with specific and restricted habitat preferences will depend on the preservation of habitats as the only effective and realistic method. In recent times Col. C.L. Scott has published widely, often expressing opinions at great variance from those in this work. All these have been considered and incorporated wherever possible.”
1999 – A great deal should be added here to account for the huge volume of additional information that has accumulated in the informal literature, and among collectors and growers. Seed propagation and selection are far advanced. Many collectors are very knowledgeable and authoritative comment is almost the order of the day. Only one thought needs expression and repetition: a revision like this is only an hypothesis built on the experience and knowledge of the author. It stands and falls on information. This is very largely the dry and seemingly uninteresting herbarium record. However, without that tangible physical evidence, identifications become hearsay and idle talk.
There is a lot of evidence to suggest that field-collected plants are being commercially exploited. Two kinds of people are responsible – those who physically remove the plants from the field, and those who eagerly and irresponsibly buy or receive such plants. I came early to the point where I could really only justify my own aquisitiveness by going to work for an organisation whose function, it seemed to me, was to collect and conserve plants. However, I was disappointed to find that such an ideal is also too subject to human failing and organisations seldom have the motivation and skills that personal interest generate. However,by far the greatest change and losses that I have observed in the field are those directly attributable to physical damage and destruction of habitat. Nevertheless, the potential for such loss should never be construed as an excuse to remove plants from habitat that simply ensures non-survival. Cut collecting can be done in such a way that there is no detriment to natural populations. Offsets, seeds and moribund plants are often available.
There is no doubt that a work of this kind stimulates interest and leads to further collection and potential exploitation of plants. However, the positive side is awareness of those plants and their survival status that also develops. It is ridiculous to suggest that there should be no collecting when life subsists on life, and we harvest from nature in so many other ways. But we need to tread the world lightly and learn circumspection and conservatism.
Cultivation, Propagation and Plant Health
1982 – In nature, haworthias are very particular as to habitat. Where several species occur together, they very seldom share the same ecological niche. Species also change as habitats differ and so it is surprising to find how easy they are in cultivation. It is generally recognized that haworthias can be grown in nearly any potting mixture based on a loamy soil. The essential requirement is good drainage, although this is not necessarily what they always get in the field. Most species occur naturally under the protection of other plants or in rock crevices and crannies. They very seldom occur under a closed canopy of larger plants and in the field one looks for haworthias in stable habitats, in rocky situation or shallow soils. Established plants can tolerate full exposure and it is in such plants that really beautiful colouring occurs. In some species the leaf tips naturally die back and it is unusual for H. longiana for example, to have tips on the older leaves. In H. lockwoodii and H. semiviva the leaf ends die back to afford the plants a protective umbrella of dry white leaf tissue. The complete dying away of the roots that so often occurs in cultivation is quite unknown in the field. Occasional large moribund clumps may occur where the centre rosettes begin to die but this is not the same condition as that of the cultivated plant losing its roots. This root loss is due to poor drainage and a wrong watering schedule. A plant may tolerate a heavy soil if it is watered less frequently. Mortality in the field seems to be very low whereas in cultivation plants may show signs of aging and frequently die. The development of a stem in the characteristically stemless species is a bad thing. Root development occurs immediately under the lowest living leaves and the stem beneath these roots is of no value to the plant. In cultivation this superfluous stem may remain alive to the detriment of the plant and a good root system. When re-potting plants care must be taken to trim away excess stem tissue. It is then often best to remove all roots and so encourage a completely new system. It appears that nearly all species will grow from a whole leaf cutting if the leaf is taken with some remnant of stem tissue – and if the timing is right.
Haworthias are essentially winter-growing plants and all species occurring west of at least Port Elizabeth respond to a winter growth cycle. January, February and March are too hot and dry for haworthias to grow. These statements will be contested by growers in summer rainfall areas and naturally by growers in the northern hemisphere where the seasons are reversed. However, successful cultivation also requires intuitive skills and there is no doubt that different approaches will work for different people. Propagation of haworthias is more often than not by vegetative offset, and the most common species represented in collections, are represented by proliferous, adaptable clones. Not all clones of the same species respond in the same way, and certainly a species considered to be non-proliferous may have individuals which are. Cultivation is an artificial situation. There is a selective factor in favour of easy-to-grow clones. The presence of self-sown seedlings and the speed and vigour at which they grow is evidence of this. Plants which do not normally offset can be forced to do so by damaging the growing point. Alternatively the entire plant may be cut into separate pieces, or only whole leaves used for propagating purposes. Roots may be treated the same way as leaves, although there is no documented success story. Various authors have now suggested or shown (Majumdar and Sabharwal, 1870; Wessels et al.,1976; and Shargi et al., 1980) that tissue culture is a feasible proposition.
Because of the penchant to hybridise in cultivation, seed propagation has been greatly neglected. Indications are that the species are all self-sterile which means that genetically different clones must be used for cross-pollination. The parent plants must also be isolated from bird and insect pollinators. Pollination is very simply effected by inserting a fine bristle into the flower tube immediately under the upper-middle lobe. The bristle is then inserted similarly into the flowers of the second plant. Pollen is released from the youngest flowers and so pollen should be collected from the upper open flowers of the one parent and transferred to the lower unwilted flowers of the other. The quantity and quality of pollen shed is not consistent and seems to be associated with weather conditions and plant vigour. The seed capsules begin to form immediately after successful pollination and may ripen within a few weeks. Not all species and clones are equally fertile and results can sometimes be disappointing. Failure of pollination is indicated by the early abscission of the pedicels. Very little is known concerning either how soon seed should be sown after the capsules open, or viability of seed. Indications are that seed should be collected only from dry, splitting capsules and sown at least within a few months of collection. Flowering time is not a good indication of when to sow seed as it varies very much from species to species. In the strictly winter rainfall region there are species in which the seed ripens in October, December and February. No documented results are available to indicate the best sowing times. At Worcester seed has been germinated successfully at all times of the year as it becomes available after flowering. A well-drained soil mixture with some compost is used and pebbles of 3-5mm diameter are scattered over the prepared sowing surface. The seeds are lightly covered with the same soil mixture and the tray finally sprinkled with smaller pebbles. These both support the seedlings, prevent washing away of the seeds, and provide a surface mulch. Watering can be done by standing the entire seedling tray in a water bath, by fine sprayer from above, or by watering onto a piece of paper laid over the pebble surface. The tray must be kept moist for the first few days and in semi-shade. A glass covering to maintain moisture may also be used early on. The seed should begin to germinate within 7-10 days and if not up after 4 weeks can reasonably be considered a failure.
Haworthias are as a rule not particularly prone to either diseases or insect pests. The Robustipedunculares are more susceptible than other species to mealy-bug attacking the heart of the plant. In outdoor cultivation there is also a weevil which attacks the plants in the same way as a bigger version attacks aloes. The only other pest of note is a white scale-insect. All these problems are very easily controlled by physical destruction. If the infestation is already out of control resort has to be made to any of the multitude of insecticide/fungicide combination dusts available. Diseases are also seldom encountered but the softer species of the subgenus Haworthia can develop a devastating heart rot and be lost before one is aware of a problem. However, this is a problem preceded by loss of the roots. If the disease has recurred, plants must be regularly checked for firm root anchorage. Excess dying of the lower leaves is also symptomatic. The cause of the problem is a fungal disease initiated by unsuitable growing conditions such as poor drainage, poor ventilation, excess moisture or humidity and perhaps an other-wise unsuitable soil mixture. Again recourse can be made to a suitable fungicide for watering onto the plants. However, it would be better to first use a dust formulation and allow the plant to dry off. If poorly rooted then the whole plant must be cleaned of all dead and moribund tissue, dusted with a fungicidal compound and carefully replanted in fresh soil.
1999 – Contribution by S.A.Hammer
Haworthia is a diverse genus in many respects, not the least in the varied horticultural requirements of its species. Most of these can adapt to widely different styles of growing, and almost all of them are easy to grow badly. A few seem universally fussy about what they need in order to grow well. It is therefore unsatisfactory to generalize, but perhaps a few points can usefully be made here. Some of the practices I discuss here are idiosyncratic, but they do work for me and can be assessed against, or added to, other methods which also work. It is fortunate that there are now so many keen people working with the genus horticulturally.
Roots are at the heart of the matter. I sometimes tell myself that I do not grow plants, I foster root systems; and while this may be only semantic, it does encourage me constantly to consider root health. Roots are of course hidden from view, but an experienced cultivator can glance at a potted haworthia and obtain a good idea of root health.
Healthy roots anchor the plants firmly and symmetrically; they absorb large amounts of water quickly; and they do not rot, although they die off periodically and systematically. Excessive root loss is taxing, but it is true that haworthias can regenerate roots with amazing speed. Not all haworthias have the same type of roots; consider the fat, stilt-like, long-lasting roots of H. truncata versus the finer stringy roots of H. venosa.
Roots are affected by many factors: the richness, acidity, tannic content, texture and aeration of potting media; the watering regime (depth and rhythm); and heat. The latter factor is critical. If certain species need heavy shade (H. pulchella is a good example), it is not so much that their leaves cannot endure bright light as that their roots cannot function well if they dry out quickly, as happens, of course, in bright and hot situations. Some plants have trouble establishing new roots in a hot environment. For example, a repotted H. angustifolia will re-establish very well below the bench, and can then be introduced to higher light; but it will have great difficulty if introduced to strong light before root formation is secure.
Why not grow them all in the shade? In a sense that would be safer, and I have tried it, but the results are generally unattractive; leaves bloat and turn a dull camouflage green, compactness is impossible, and (with some species) rot is more likely. Even worse, the species begin to look alike! Cultivation should emphasize the individuality of plants, not mask it. So it is usually good to give haworthias as much light as they can tolerate without risking root loss, tip burn, or the desperately self-shading bunched-in look that says: help! A bright and cool ambience is ideal. Bright = light strong enough to keep lithops from looking like newly-risen toadstools; cool = under 30°C, though plants will certainly tolerate far higher temperatures. Some species will tolerate frost, but it is better to avoid temperatures below 5°C. In any case good air movement is vital.
One of the main difficulties with haworthias is that they do not readily wilt or wrinkle; they do not give one an immediate signal of their needs. This delayed response always puts one at a remove from the plant’s condition, it is like reading last year’s newspaper. It is true that a heavy watering is quickly followed by obvious swelling and glossiness, but the effects (which are often cumulative) of subtler factors – misting, light feeding, adjustment of fertilizer ratios, slight changes of position, repotting, leaf removal, and, especially, root loss – take longer to manifest themselves. A virtuoso grower, Bob Kent, once told me that last month’s sunlight = this month’s leaf colour. It took me a long time to appreciate what he meant. Colour change can be so gradual as too imperceptible to an ardent daily viewer, and it can be positive (an increase in the reddish tints which attract most of us) or negative (reddish tints are also often indicative of stress).
Haworthias burn in an obvious way only if they are suddenly introduced to a grossly brighter environment. They may, however, darken quickly if they are moved from dull quarters to a situation in which more ultraviolet is available; they may turn nearly black overnight. This is not burning, exactly, but it can compromise plants all the same. Certainly it slows them down.
As a corollary to Kent’s observation, I would add that last year’s feedings = this year’s beauty; the effects on symmetry (and on the expression of leaf patterns) are that long-lasting. Should one feed at all? That partly depends on one’s patience. But there is a sense in which feeding= strengthening. And there is no virtue in waiting four years for a starved seed1ing to mature, when a compact look can be obtained in one-quarter the time, via dilute but frequent doses of low nitrogen fertilizer! Such feeding will also improve flowering and seed-set on mature plants. It will, however, encourage offsetting, which can be a bit excessive.
Soil mixtures have caused infinite anxiety. Many growers change their mixtures every few years and find that nothing works equally well for everything. Nothing can: roots and needs are too diverse, but one can adjust proportions here and there. A light mixture (i.e. fine-textured humus plus inert drainage material like pumice, gravel, perlite, or clay aggregate) is open in texture and will require frequent wetting. A heavy mixture (i.e., loam + sand + drainage material) stays wet for a long time, which can be an advantage in a dry warm climate, a fatal liability in a damp one. People who love the act of watering, for whom watering is synonymous with plant enjoyment, should either use a light mix, or employ a hand mister frequently and avoid the hose. But it remains true that haworthias will adapt to a wide range of potting media as long as the grower adapts along with them.
Whatever the soil one uses, a top dressing of grit, gravel, or pebbles is normally beneficial and always attractive. In a hot greenhouse, dark pebbles can actually cook plant tissue, so pale ones are better; they also show off the plants more effectively. Mulches do conceal the condition of the soil, but one learns to judge that simply from the state of the leaves.
I have avoided the difficult question: how often to water; it is too individual. However, for plants in full growth, a cycle of: 1 – 2 days of saturation 3 – 4 days of even moisture 1 – 2 days of near-dryness gives a rhythm which is effective in arid climates. In damp areas, and during the dampest periods anywhere, the cycle will be radically slower. In any case, small shallow pots dry out much faster than one imagines; deep pots, on the contrary, have hidden and dangerous reserves. Whatever the pot depth, one can consider an alteration of infrequent deep watering, in which water runs copiously out of the drainage holes, and frequent shallow watering – almost a splashing on the surface – which amounts to a light refreshment, perhaps mostly for the grower.
Too-frequent deep watering will lead to root loss and stem rot; it will kill some haworthias and turn others into green behemoths. Slightly overwatered plants tend to develop cracks or crevasses on their leaf surfaces (curiously, the upper, not the lower surface, is subject to this); such cracking is unsightly though not injurious. I might note that some clones crack even if they are watered gingerly; as a matter of good horticultural practice these should not be propagated. The syndrome is genetically based and cannot be wholly eradicated. In my experience H. magnifica var. atrofusca,H. truncata and H. emelyae var. comptoniana are particularly prone to this.
Many species grow continuously; their life is a perpetual spring. Has anyone ever killed, or even discouraged, H.cymbiformis? Others have obvious bursts of growth which should be observed and encouraged: one should run with the tide. Generally such tides flow in late winter and early to late spring. I will even water hyperactive plants twice in a row, on successive mornings, but only if the air is warm and dry.
Some Haworthia species have unmistakable dormancy syndromes, possibly because they are adapted to a winter rainfall regime: H. lockwoodii and H. semiviva are the most obvious of these. Summer is the time for their papery sleep; during this period they need only light watering, enough to prevent complete desiccation, not of the leaves, which are already desiccated, but of the roots and meristematic core. Other species turn inert and unresponsive for long erratic periods (e.g., H. bruynsii and H. springbokvlakensis, those unrelated behavioural twins). Generally this torpor occurs in winter, when light misting is sufficient to maintain a sluggish viability, but it may also occur in high summer, when extra shading is the safest response.
Pollination is now taken seriously by many growers. It might be helpful to mention that seed production is best on well-fed plants and that flowers are most receptive a day or two after anthesis (first opening). I generally pollinate a flower twice a day, morning and evening, two days in a row. I spray my hands with alcohol when shifting from one species to another to prevent accidental transfers, because the barriers between species – particularly in the subgenus Haworthia – are slight or absent.
Some species are usually difficult to pollinate. H. limifolia is a good example. Other species may elude one grower and not another; for me H. koelmaniorum is recalcitrant, for an excellent grower in torrid Arizona it makes masses of seeds, but then his plants look better and healthier than mine. He successfully uses a fine fibre of nylon as a pollinating tool; very narrow camel’s hair brushes are ideal but expensive.
Clones of most species exhibit some self-fertility but it is difficult to prove the absence of any animal vector. Often one will notice that the topmost flower of a neglected inflorescence ripens into a fat fruit; it is always worth sowing the contents. (By the way, the offspring will not necessarily be uniform.) In other cases, the whole inflorescence seems to ripen at once in a rush, skipping no fruits; this is certainly an instance of self-fertility and the offspring will usually be self-fertile as well.
Haworthias can be propagated by seeds, leaves, offsets, stolons, coring, tissue-culture, some even from roots. Seed raising is the best method for anyone who enjoys the promulgation of variability; tissue-culture is more efficient but it results in a massive sameness.
For seed work, start with good seeds! Much seed in the trade has a low endospermatic content – the seeds are well-formed but small – and the resultant weakness can cause much trouble. Seed germinates best when a few weeks old, better than when newly ripe, but it will keep for a few years, up to seven in my experience.
Seed can be sown at any time of the year. Preferably one should avoid the hottest and coldest periods unless one sows indoors under lights; indeed I sow all seeds under lights, which gives me far greater control of quasi-sterility, insects, and temperature, ca. 20 – 25°C being ideal. The lights can be the standard “cool” tubes, but those work best when paired with broad-spectrum “grow-light” tubes.
For sowing I use a commercial sowing medium (peat moss treated to improve its wettability + a little sand + vermiculite) which I cut with an equal part of fine-screened pumice (>2mm diameter) or perlite. I sterilize the mixture in the microwave and bleach the pots; this is tedious but it pays off. I fill the pots, tamp firmly, sow the seeds, and cover with a thin layer of grit. I sow the seeds in small pots, up to 100 seeds per pot. I prefer very small plastic pots (30 x 30 x 30mm); these dry out quickly, which means that I can water them frequently and thus administer frequent doses of fertilizer. It is important not to sow so thickly that seeds cover each other, leading to a tangled chaos.
I soak the sown pots in distilled water, draining them after being thoroughly saturated (an hour or so). The pots are placed ca. 15 cm under fluorescent lights which are never turned off, and are covered with tight-fitting plastic. The next day I make sure that the plastic shows a beaded film of moisture; if it does not, I mist the pots. After four days I remove the plastic, misting the pots twice a day thereafter. Germination should occur within a few more days, 6 – 14 from the time of sowing. After germination I mist the pots once a day but I avoid saturation; aeration in the soil is critical. A small fan kept on perpetually (along with the lights) helps to prevent damp-off and algal growth; the latter can also be stemmed by progressively mulching the seedlings.
I transplant seedlings at the 4 – 5 leaf stage which usually occurs for me when they are ca. two months old; the unspotted seedlings are soaked in an hour-long bath of distilled water to which I add some Vitamin B1. I use the same medium as I used for sowing (though sterilization is unnecessary and particle size can be larger). If newly transplanted seedlings dry out, they will have a hard time re-rooting, and of course the soil mix may be difficult to re-wet.
Not all species have the same rate of growth, either as cuttings or seedlings. H. bruynsii is always slow, taking at least four years to reach flowering size from seed under my conditions. H. sordida, H. scabra and H. koelmaniorum are also slow although the latter can expand quite rapidly once it passes the juvenile stage. The retuse species are much faster and most can be in flower at 12 – 18 months. H. bayeri and its ally H. truncata var. maughanii are quite slow, both pass through a tediously prolonged (and similar) juvenile stage in which they are not quite themselves.
With all species, extra shading will promote seedling growth. Though this is at the expense of colour, that can quickly be (re-)gained. One should experiment with placement of seedlings; full morning exposure and afternoon shade are the obvious poles but many gradations between these states are possible. Seedlings (and adults) are highly responsive to changes in light and many troubles can be cured or courted by a simple shifting of pots.
Most haworthias will root from leaves. Thin leaves, e.g., those of H. arachnoidea, are very difficult to root, and firm ones, e.g. H. sordida, are slow, taking many months. Plump leaves and a bit of stem should be removed from depotted plants, dusted with fungicidal rooting powder, and placed in a sowing medium in a dim corner of the greenhouse. I mist them once a day, lightly; otherwise I ignore them. They should start to callous in a few weeks and root soon thereafter, but differentiation can take months. Incidentally, leaf removal is probably the least damaging way to collect material in the field if the mother plants are not disturbed by it.
Offsets also amount to propagation from leaves, but the plant itself does the work, forming plantlets at the base of leaves which are nearing the end of their term of duty. The plantlets can be removed as soon as they have an independent root system. Coring plants – scooping out the meristem, like removing the eyes on a potato – often results in proliferation from the basal leaves, but it is an ugly method. In another rather brutal method, which I mention only as an experiment, plants are subjected to a few hours just below freezing; this may kill them, or it may result in enormously increased offsetting.
Hybridization is not much touched on in this book, which focuses on species as they occur in nature. Here I will only mention that hybrids have a great potential for horticultural beauty and taxonomic confusion, possibly for some elucidation as well. Hybridization includes, of course, pollinating plants from different populations. When making hybrids, keep meticulous records (noting which species is the pollen parent, which is the unorthodox father), use the most attractive parents, pollinate for three or four days in a row, and don’t give up if only a few seeds form.
To date the most attractive hybrids (attractive to me, anyway) have involved H. emelyae, H. magnifica and H. bayeri, but thousands of possible combinations have not been tried, especially when one considers those involving three or more already hybridized parents! Only a few of the hybrids in cultivation have great horticultural merit; the rest should probably be eaten or otherwise recycled.
Allied to hybridization in its artificiality, but more palatable to many growers, is the practice of selection. The plants are so variable in nature that they offer limitless scope. One can pollinate the most attractive examples of a species, the seedlings can be selected and back-crossed with the parents, and in this way many (any?) desired characteristics of form or colour can be reinforced. Of course, it can not be assumed that seedlings cultivated from material from a specific locality will represent those which would have survived selection processes in their natural state. Nor is it clear how long-lived haworthias can be, either in nature or cultivation, though well-favoured plants can probably live for centuries. The little amount of this work that I have done to date is most promising. It is simply a way of beating the natural odds.
Whether or not these “artificial” beauties should still bear the data their grandparents came with is a good question; my own opinion is that they have crossed over from nature into art. Fortunately a good collection of Haworthia can represent both realms!
The Genus and Species Concept
1982 – Since Berger’s monograph in Engler’s Das Pflanzenreich, the only real contributor to an understanding of the genus as a whole, and its relation to the Aloineae has been A.J.A. Uitewaal. This is expressed in 2 series of papers published in Desert Plant Life (1947), Cactus and Succulent Journal of Great Britain (1947), Succulenta (1948) and Sukkulentenkunde (1951). Originally Haworth recognised only Linnaeus’ genus Aloe and it was Duval (1809) who erected Haworthia to contain the species in Haworth’s section Parviflorae. According to Uitewaal, Haworth and Willdenow independently applied the same name Apicra to the genera now recognised as Astroloba and Haworthia respectively. Apicra Willd. is thus a synonym of Haworthia Duval, and the name Apicra Haw. for a different genus is a later homonym and invalid. This is the reason for the creation of the name Astroloba Uitewaal. Actually Haworthia is already a nomen conservandum in favour over the earliest name Catevala used by Medicus (1786).
Apart from Salm-Dyck, no authority with any reasonable knowledge of the Aloineae has suggested that the genera do not fairly reflect the diversity of the group. Parr has published (African Succulent Plant Society, 1971-2) what purports to be a revision of the genus Astroloba in which he includes these species together with Poellnitzia Uitew. as the section Quinquefariae in Haworthia. The wisdom and intent of such a change is dealt with by Bayer (National Cactus and Succulent Journal, 1972) and it is rejected for a number of reasons including that re-alignment of genera when the elements within them remain poorly defined, is unquestionably premature. Existing genera do portray natural groups despite weak rationalisation in terms of morphological and other criteria. The independent recognition of the different status of Haworthia and Astroloba by Haworth and Willdenow is indicative of the ‘naturalness’ of the two genera. It is true that there was some confusion as to the relation of H. viscosa (L.) Haw. to Astroloba prior to Haworth. Mrs P. Roberts-Reinecke in an unpublished revision of Astroloba (M.Sc. thesis, Department of Botany, U.C.T.) also concludes fairly convincingly that Apicra aspera Haw. is in fact synonymous with H. nigra Haw. This is nevertheless easily attributed to poor observation of floral characters and doubtful evaluation of vegetative characters. The question of incorporation of Astroloba in Haworthia needs consideration in light of recognition of three subgenera in Haworthia. This first finds expression under Salm-Dyck, later Berger, then Uitewaal (1947) and then Bayer (1971). The distinction between Haworthia and Astroloba on the basis of bilabiate as opposed to stellate flowers is extremely weak and Uitewaal’s usage of ‘approximately’ (plus/minus) in his key is very apt.
Mrs. A.A. Obermeyer-Mauve in Bothalia (11:119, 1973) included Chortolirion Berger (one species only) in Haworthia, and Chamaealoe Berger in Aloe L. She at the same time endorses Parr’s inclusion of Astroloba and Poellnitzia in Haworthia. The sole reason given here is also that the latter genera have regular flowers while some haworthias do too, and the claim is made that there are no correlated distinctions. This is quite a fallacious argument for reasons expressed elsewhere (Bayer, 1971, 1974). Mrs Mauve likens Chortolirion to H. graminifolia Smith when in fact the latter is not the least bulbous and neither is there the degree of similarity in the leaves which she suggests. The flower of Chortolirion is like that of the Haworthia subgenus Hexangulares and cannot be confused with that of the subgenus Haworthia where H. graminifolia belongs. In The Genera of Southern African Flowering Plants by R.A. Dyer (1976), Astroloba, Poellnitzia and Chortolirion are all included in Haworthia. For the purpose of this book the traditional generic states are maintained.
The species concept in Haworthia is discussed briefly by both Uitewaal and Resende who stress the view that speciation in the genus is still in progress, and that species are poorly defined. To a greater or lesser degree this must be true in the majority of plant genera, and is certainly not true for all species, even in Haworthia. Each species needs to be considered in terms of distribution, variability and isolation. Speciation and the evolution of discrete segregates (species, subspecies, varieties, etc.) is self-evidently a function of isolationary mechanisms. Whether anyone can honestly claim to have an adequate proven explanation for sympatric distribution of so many Haworthia species in so many areas, each with specific habitat requirements and consequent discontinuities is very doubtful. The isolationary mechanisms operative where species are growing in the same near vicinity can be of several kinds. Either the plants are not cross-fertile due to physiological or genetic barriers; or the fertile seed does not produce a seedling which can survive ecological selection pressure; or flowering times are staggered. Experience suggests that hybridisation is a common phenomenon, at least in cultivation, and many ‘species’ are, by admission, garden hybrids. Although hybridisation in the field is far less common than this suggests, it does occur and it can be implied that species have arisen from such hybrids too. Selection as a result of ecological requirements is in the writer’s opinion, a highly acceptable explanation for the development of Haworthia species whether breeding barriers occur or not. Flowering time also differs consistently enough in the field to suggest that it is a good isolating mechanism. However, the species problem in Haworthia very seldom arises where species grow together, but rather from the difficulty in associating populations strung out in isolated localities. These populations can be mostly recognised as species complexes, but populations do occur which cannot be confidently assigned to either one or more of their geographically obvious counterparts. The complexity of what species are is beyond the object of this book, beyond providing some clue to the concept of a species accepted in the checklist. The species have been largely defined in terms of geographical distribution, on the assumption that selection across ecological gradients is mainly responsible for their origin. It is very comforting for the laymen to know that even the most learned botanists cannot define a species in simple terms. It is important to realise, though, that a species is more than a simple expression of differences between plants which the ordinary man in the street can recognise. Two plants may appear to be quite different in size, colour, markings and flower, yet easily recognised as the same kind (species) when seen growing together with a number of other plants sharing all the differences of the first two. If one or several plants still stand out from such a group, then they can be recognised as distinctive forms. Normally a plant species can and should be considered to consist of several groups (populations) growing at different sites (localities). If one of these groups is considered to be sufficiently different from the others, then it may earn recognition as a variety. If several groups are different in this way, then they may be referred to as subspecies. In each case the degree of difference is important and different people may not share a decision to recognise differences by naming forms, varieties, subspecies or even species. In this way classification and taxonomy can become distorted by the judgement of the individual applying himself subjectively to the naming and renaming of plants. The more information available, and the more thoroughly and objectively the work is done, the less likely is the final product to be a reflection of individual idiosyncrasy and personal opinion. The differences between plants of the same kind may result from external factors such as soil type, exposure to light, availability of water, etc., or from internal factors such as age and genetic make-up. Uitewaal expressed the view that it would be possible to classify Haworthia even without a full understanding of how these factors actually affect the plants. The author agrees with this view because experience shows that under moderate conditions of cultivation, plants can be expected to remain consistent. Uitewaal in 1951 was perhaps less optimistic that floral characters would be useful than he had suggested in 1947, but the importance of using flowers as a source of characters is not diminished thereby. Most authorities have claimed that floral characters are too uniformly the same to be of any use in classification. Such a conclusion may have arisen only because similarities occurred where differences were expected and vice versa, based on the a priori presumption that classification by vegetative characters was correct. Vegetative characters have always been the predominant basis for classification in Haworthia in complete contrast to other genera. Far too much weight has been attached to differences which may occur even between plants of the same kind at the same locality. Examples are the presence or absence of tubercles or spines, or the number of veins showing on semi-transparent leaf faces. Smith’s descriptions are models of detail and uniformity, but rather than circumscription of species and varieties, are descriptions of single plants. Many other descriptions include few diagnostic characters by which the identity of a plant among near relatives can be determined with any certainty. The descriptions are padded with measurements which have no reference to the range of dimensions in the entity, or by characters equally applicable to other, or all, members of the genus. The state of Haworthia classification is easily assessed by the past arrangement of the species in 20 sections compared with their natural disposition in the three subgenera. If even sections have not been defined correctly to include related species let alone variants of one species, there can be little confidence that the species themselves are all correct.
1999 – As far as genera are concerned, the attempt to relegate Astroloba, Poellnitzia and Chortolirion to Haworthia appears to have failed, so that good sense can be said to be prevailing. Dr G.F. Smith has written many papers relating to Aloaceae which generally project the view that the genera are adequately different. Emphasis on the differences within Haworthia at a corresponding level is still sadly lacking and Dr L.E. Codd’s comment that ‘small differences are going to be ignored’ is sadder still. Small in size does not equate to small in significance. There can be no doubt that Haworthia is composed of three very distinctive sub-groups, recognisable on floral characters alone. I have previously commented on the similarity of the Chortolirion flower to that of the Hexangulares. I would like to say forcefully that while I doubt if I could distinguish many of the species in the subgenus Haworthia on the basis of the floret, I would confidently attempt to recognise Chortolirion among them, and among any of the other subgenera too. Nevertheless a taxonomic revision is only an hypothesis which cannot be proved right. If tested and it is infirm, a new hypothesis should replace it. Thus there will be an on-going attempt to re-define the genera, and one can only hope for some balanced approach where due recognition is given to the foundations of the old, before an equally, or even more greatly flawed alternative is put forward. The subdivision into lower groups (Sections) is very difficult and if the prime divisor is disregarded (i.e. the flower), likely to be impossible. If the subgeneric characters are disregarded, on what kind of characters are the species to be recognised? Simple characters which one would expect to separate species, and form sectional characters, do not exist. For example tuberculate versus smooth leaf surfaces or leaves spineless versus spined, frequently occur within species. Even windowing or coloration of the leaves is not a certain clue to identification. Ultimately the test is distribution and co-occurrence.
Why the taxonomy of these genera in the Aloaceae has been considered more confused than elsewhere in the plant kingdom is a bit of a mystery to me. I see a similar situation in the Cactaceae (although I do not know it very well); and have experienced the same problems in Asparagus, Oxalis, Pelargonium, several other genera which I came to know quite well, the Stapelieae, and also strangely enough in Noctuid moths. Very high collector interest and low academic involvement may be seen at the root of the problem. I am not confident that intellect as such will necessarily produce more truthful answers. The concept of truthfulness I use here is not that of an honesty of expression, but that of objective factual correctness. Anything subjected to the intense interest of collectors, is bound to generate controversy. It is salutory to see what wars have been waged in very quiet backwaters of botanical interest, between protagonists barely known to any but each other. I would like to repeat what I have already said in the introduction where I credit collectors with as much acumen as any botanist who claims that title by virtue of qualification rather than from interest. Both groups rate from low to high on the various and many scales by which intellect could or should be measured, and with an unfortunate poor correlation to how the individuals may assess themselves.
To understand Haworthia, it seems to me that James Gleick’s book on ‘Chaos: Making a New Science‘ is useful reading. Even if intellectually incomprehensible, the gist of infinite variability, period doubling and bifurcation diagrams will help how one thinks about species. The nature of variation in plant species – the nature of genetic material, the way it is ordered, and the way in which this is expressed in the appearance of plants – can be said to be a phenomenon of chaos theory: disorder which is highly ordered. We look for regularity and order in plant species and populations and it simply is not visibly there. How many of us have said that our classification system does not match the facts?
Vavilov’s law of homology presents the view that variation in one element of a group is parallelled in other elements of that group. He said that: ‘In closely related species and genera, similar series of heritable variations occur. These are found so regularly that knowing the series of forms within the limits of one species, we can predict the occurrence of parallel forms in other species and genera’.
Our attempt to assign position to things is really to locate them on a kind of dendrogram. No two individuals in a sexually propagating population are exactly the same, and the same can be said of vegetatively propagated plants, with certain provisos. Thus at what time will we recognise that a species has split into two? We have to do it on some kind of statistical basis or make an ‘educated’ guess. A split may be the consequence of a difference of only a tiny fraction of the total genetic material. A dendrogram indicates the split, but the appearance of the individuals in the now-divided species may not. On the other hand a single gene character may induce a massive change in appearance which might occur frequently or infrequently within a population. So it is logical and reasonable that individuals of closely related species will tend to resemble one another, and that variation within a species may exceed that between species. Some individuals of different species may resemble each other more closely than they resemble individuals within their own ranks.
This book has of necessity set out to honestly bypass technical nicety and complexity and to practically identify nodes in a complex interlinked web. I do not believe that there is any way that the vast variation that does exist can possibly be accounted for in any written or pictorial record. In many instances there are no real boundaries and we have adopted the view:
that species are a group or groups of interbreeding or potentially interbreeding individuals varying continuously, genetically and morphologically, in both time and space.
There is an island-like situation in Haworthia which is both real in terms of the isolation of habitats, and also artificial due to destroyed habitats. The latter are significant to us only in present time because they affect what we can observe. The observation and interpretation (measurement and evaluation) of continuity is limited by inaccessibility to the past and by the sheer physical problem involved in locating and seeing everything. It is the nature of creation that diversity exists in space and change is brought about in time. Chaos theory must apply.
In this new treatment I have tried not to get involved in the rigours of systematic nomenclature for which I am not adequately proficient nor sufficiently orthodox. It has been proved to me that it is subject to gross error and personal antagonism as any other field of human endeavour. Some people are able to indulge in a semantic and legalistic forest of words for that reason and pleasure alone. I respect the purpose, but it is worth recounting briefly how senselessly it has been applied to Haworthia. In my very first encounter with the problem of the name H. margaritifera vs H. pumila I put the problem to a Professor and two of his doctoral graduates. None would even brave an opinion. Then Dr Codd offered a view applied by Col. Scott, which was up-ended by Dr Wijnands. Since then that has been brought into question too, and I have become convinced that argument is directed to the end-point desired. Rationalization follows decision. In truth it appears they have all missed the mark because the element they are most frequently arguing about is most probably what we commonly refer to as minima and not pumila. Thus the correct interpretation is perhaps that pumila should be scrapped and that maxima should be used for this larger species, and margaritifera for the littler minima. An attempt in this direction will be apparent in this work, with an apology to the cognoscenti who are most active in critical retrospect, rather than pro-actively helpful when needed.
We have the case of the argument against the retention of emelyae based on doubts as to origin of the type, despite evidence which exists to show the contrary. I was no doubt wrong in avoiding the use of atrovirens for H. magnifica var. maraisii, and am also argued to be wrong about integra. In my view these are cosmetic problems which should not be allowed to distract anyone until such time as someone can come along with a truly better understanding of the things they are trying to classify. There seems little sense in getting all the names right while the elements to which they refer remain totally confused. There is a real problem in trying to typify the species on the basis of old herbarium specimens and also on the basis of old illustrations, because quite simply it is often very difficult to even identify living flowering material without direct reference to origin. In this respect it should be noted that the whole question of typification has been addressed by I. Breuer and D. Metzing and shown to me in manuscript form as I finished this manuscript. I assume that their typification is authoritative and where I have depended and used their typification it is acknowleged by the letters “B&M” as the source for designation of the type. I do not consider their typicication to be based on any real knowledge of the genus, and their selection of neotypes and epitypes is often unfortunate. This is apart from the lecotypification of photographs in the Berlin Herbarium which were mostly published with the original descriptions. Otherwise I have considered the types to be as designated in this work, and representing the historical record and the traditional interpretation and understanding of the names.
It has not been necessary for me to have the view confirmed that species of Haworthia can only be reasonably defined in terms of geographical distribution and interpreted on the grounds of co-occurrence. The nature of the interlinking web has been enormously strengthened by the collections I have seen in the past 15 years, without seriously weakening the artificial structure of names. It is this ‘structure’ which I would claim has facilitated communication and observation. The discussion under the individual species will deal with new collections and the insights which they bring.
The main point is that there are alternative decisions that can be made or could have been made – and if good sense is maintained there will be no problem. Problems arise from unseen mistakes as we have in the confusion of H. arachnoidea sensu Scott which is the same species as H. herbacea sensu Bayer. This is not just the product of nomenclatural nicety. There is also the mistaken identity of H. serrata by Col Scott where he has assumed that a population of H. mucronata is that species. In this revised edition, an attempt will be made to specify a type for each name, following Breuer and Metzing as closely as possible.
Other difficulties arise from completely different perceptions about what constitute species. Unless a good strong base is laid in the geographic component, it just becomes impossible to handle the close resemblances within Haworthia and the ‘chaotic’ nature of transitional populations. In this treatment we find difficulty in supporting, say, H. reddii as a species because we do not regard it as substantial enough to hold that rank. We think a species should be more substantial and have a range of distribution and variation across that range. We are, I think, a little pretentious in the use of the many ranks of subspecies, variety and form. In this revision I have reverted to mostly the use of varietal rank. However, this does have its own problems because as soon as a subordinate rank is created, it implies that the species is composed of two discrete elements. This is probably not problematic where subspecies are concerned and the concept of extended variability is retained. However, when a variety is selected to illustrate or communicate a specific population which is unusual in one or other respect the balance of variability is loaded towards the typical and we have incongruity. This is the view adopted here i.e. there is not a typical variety in the sense demanded by the nomenclatural system. Rather there are variable species which intergrade, and among them are consistencies which are identified as distinctive varieties. The convention of citing locality is by far the most suitable way of adequately communicating where identification becomes doubtful, as it inevitably will. The ideal of consistency is very difficult to attain because the plant populations seem to be aligned at different grades of similarity.
I recognise that if my initial view of the species is reasonable, then the formal recognition of varieties may ease the understanding of the nature of the variability and existing or potential inter-relationships between the species. This will be particularly evident in the way variation of the old H. retusa varieties are now treated and also in the many varieties recognised here, particularly in H. archeri and H. heidelbergensis, which were relatively unknown in 1983. Similarly while several new species appear in this work as a product of new finds, new insights, corrected mistakes, and new mistakes, some old species have been relegated to varietal level for the same reasons. Lesser ranks should not be taken too seriously as they also represent a view which changes with familiarity and with time. A revision may present a new set of names and ranks, but does not erase the history of those names and they remain there for the edification of all.
Synopsis of Taxonomic Changes
The main changes in this revised edition are as follows:-
H. aranea A.Berger = H. arachnoidea var. aranea (A.Berger) M.B.Bayer.
H.archeri Barker ex M.B.Bayer = H. maru.miana var. archeri (WF.Barker ex M.B.Bayer) M.B.Bayer.
H. batesiana Uitewaal = H. marumiana var. batesiana (Uitewaal) M.B.Bayer.
H. comptoniana G.G.Sm. = H. emelyae var. comptoniana (G.G.Sm.) J.D.Venter &: S.A.Hammer.
H. correcta of Scott (pro parte H. emelyae V.Poelln.) = H. bayeri J.D.Venter & S.A.Hammer.
H. divergens M.B.Bayer = H. monticola Fourc.
H. graminifolia G.G.Sm. = H. blackburniae var. graminifolia (G.G.Sm.) M.B.Bayer.
H. habdomadis V.Poelln. var. habdomadis = H. mucronata Haw. var.habdomadis (V.Poelln.) M.B.Bayer.
H. habdomadis V.Poelln. var. inconfluens (V.Poelln.) M.B.Bayer = H. mucronata Haw. var. inconfluens (V.Poelln.) M.B.Bayer.
H. habdomadisV.Poelln. var. morrisiae (VPoelln.) M.B.Bayer = H. mucronata Haw. var. morrisiae (V.Poelln.) M.B.Bayer.
H. maughanii V.Poelln. = H. truncata var. maughanii (V.Poelln.) Fearn.
H. magnifica var. major (G.G.Sm.) M.B.Bayer = H. emelyae var. major (G.G.Sm.) M.B.Bayer.
H.magnifica var. paradoxa (VPoelln.) M.B.Bayer = H. mirabilis var. paradoxa (V.Poelln.) M.B.Bayer.
H. poellnitziana Uitewaal = H. minima var. poellnitziana (Uitewaal) M.B.Bayer.
H. pumila (L.) Duval sensu C.L.Scott = H.pumia (L.) M.B.Bayer.
H. radula (Jacq.) Haw. = H. attenuata var. radula (Jacq.) M.B.Bayer.
H. retusa var. acuminata M.B.Bayer = H. magnifica var.acuminata (M.B.Bayer) M.B.Bayer.
H.retusa var. dekenahii (G.G.Sm.) M.B.Bayer = H.magnifica var. dekenahii (G.G.Sm.) M.B.Bayer – pro parte.
H. retusa var. argenteo-maculosa (G.G.Sm.) M.B.Bayer = H. pygmaea var. argenteo-maculosa (G.G.Sm.) M.B.Bayer – pro parte.
H. rycroftiana M.B.Bayer = H. mucronata var. rycroftiana M.B.Bayer.
H. starkiana VPoelln. = H. scabra var. starkiana (VPoelln.) M.B.Bayer.
H. translucens Haw. sensu M.B.Bayer = H. gracilis V.Poelln.
H.unicolor (V.Poelln) unicolor = H.mucronata Haw. var. mucronata.
H. unicolor var. helmiae (V.Poelln.) M.B.Bayer = H. mucronata var. mucronata Haw.
H. unicolor var. venteri (VPoelln .) M.B.Bayer = H. arachnoidea var. nigricans (Haw.) M.B.Bayer.
H. wooI!eyi V.Poelln. = H. venosa subsp. woolleyi (VPoelln.) M.B.Bayer.
H. xiphiophylla Baker = H. arachnoidea var.xiphiophylla (Baker) M.B.Bayer.
New species included are:
H. aristata Haw.
H. bayeri J.D.Venter & S.A.Hammer.
H. maraisii V.Poelln.
H. outeniquensis M.B.Bayer.
H.pungens M.B.Bayer.
H. vlokii M.B.Bayer.
Species desclibed since publication of The New Haworthia Handbook and their treatment :-
H. joeyae CL.Scott = H. cooperi Baker var. dielsiana (V.Poelln) M.B.Bayer.
H. magnifica VPoelln. var. splendens J.D.Venter & S.A.Hammer.
H. mcmurtryi C.L.Scott = H. koelmaniorum C.L.Scott var. mcmurtryi (C.L.Scott) M.B.Bayer.
H. pehlemanniae C.L.Scott = H. nortieri var. pehlemanniae (C.L.Scott) M.B.Bayer.
H. pringlei C.L.Scott = H. decipiens V.Poelln. var. pringlei (C.L.Scott) M.B.Bayer.
H. reddii C.L.Scott = H. cymbiformis (Haw.) Duval var. reddii (CL.Scott) M.B.Bayer.
H. venusta C.L.Scott = H. cooperi var. venusta (C.L.Scott) M.B.Bayer.
In my view they are not substantial elements with variation and range, and can be accommodated within species already extant. The illustrations in this work will include similar elements in other species and still fall short of accommodating quite substantial variation in many of the species.
Key to the Subgenera
1982 – Uitewaal (1947) presented a Latin diagnosis for two sections, Triangulares (type species Haworthia translucens Haw.) and Hexangulares (type species H.coarctata Haw.). The latter section he again divided into the Robustipedunculatae and Gracilipedunculatae, partly derived from Berger (1908). Uitewaal did not relate this subdivision to then existing sections and stated specifically that it was only of a tentative nature. The following exposition (Bayer, 1971) takes Uitewaal’s division to its logical conclusion with the recognition of three subgenera. In the process, the subgenus containing the type species for the genus repeats the name of the genus:
1. Perianth at base triangular or rounded-triangular, style upcurved, tube obclavate ………. subgenus Haworthia Type species H. arachnoidea (L.) Duval – Perinath at base hexangular or rounded-hexangular, style straight, tube obcapitate ………………….
2. Perianth gradually narrowing to junction with pedicel (substipitate), tube curved…….. subgenus Hexangulares Uitewaal ex M.B.Bayer Type species H. coarctata Haw.
3. Perianth abruptly joined to pedicel (non-stipitate), tube straight ………………. subgenus Robustipedunculares Uitewaal ex M.B.Bayer Type species H. pumila (L.) Duval
This construction of subgenera would necessitate re-organisation of the section Margaritiferae as composed by Jacobsen (1965), as several of those species belong naturally with the Coarctatae. H. papillosa (Salm Dyck) Haw. was placed in the section Coarctatae by Baker on account of its elongated stem. This was totally incorrect as caulescence does occur in the Margaritiferae where the plant can obtain sufficient water and nutrient to maintain an extended growing point. More often the central parent stem dies away and the plant is perpetuated by several acaulescent offsets. However, there are many other anomalies which can be quoted in demonstrating the inaccuracy of the sectional divisions up to Jacobsen (1965). The real problems and difficulties in Haworthia are best appreciated by referring to synonymies of species like H. reticulata and H. herbacea. Here it is seen how authors have described several species from what can be proven to be a single entity.
1999 – A key to species seems to be the summum bonum of revisionary work and it would be really an achievement to provide such a key by which the species could be readily identified. In Haworthia I do not think a key will provide a probability for correct identification of existing material much better than 50% of the time. For new collections from poorly explored or poorly collected areas, it may even be worse. This may confirm that in reality there are many fewer species, which I would not dispute.
The most common and practical way of identification is by trial and error using visual comparisons. In my experience there have been and are misidentifications with the simplest material and even the best of keys and explanatory notes. There are enough cases where even photographs and illustrations are hopelessly misidentified.
There have been several attempts to create sections in Haworthia and these have nearly all been across the subgeneric boundaries. One might expect that sub-division would be easier if the subgeneric limits were first accepted, but the subgenus Haworthia seems to defy resolution. Whatever way is attempted is eventually confounded by affinities across demarcated boundaries. Some links and affinities are clearly stronger than others. It is clear to me that as exploration and collecting yield more material, so will the complexity of the problem. Hence the hope of finding a simple solution will recede with time and not as we like to think, improve.
Flowers of the three subgenera. Diagram by J.G.Marx
1. Haworthia angustifolia Haw., Phil.Mag. 46:283(1825), Baker, JLinn.Soc. 18:210(1880). Non Jacobsen 2:536(1954). Bayer :97(1976). Bayer :26(1982): Bayer, Nat.Cact.Succ.J 37:31(1981). Scott :54(1985). Type: Not preserved. Neotype (designated here): icon, 13:f2, Salm-Dyck, Monogr. Epitype: CAPE-3326 (Grahamstown), Highlands (-AD), Cooke in NBG68977, non B&M Bruyns 1653 (NBG): Aloe stenophylla Roem. et Schult. Syst.Veg. 7:641(1829). Salm-Dyck, Monogr. 13.t2(1849). Type: As for H. angustifolia: H. angustifolia var. grandis Smith JS.Afr.Bot. 9:105(1943). Type: Cape, Albany Div, Smith 5216 (PRE): H. albanensis Shonl., Rec.Albany Mus. 2:256(1912). H. angustifolia var. albanensis (Shonl.) V.Poelln., Feddes Repert.Spec.Nov. 41:194(1937). Type: Cape, Grahamstown, Britten s.n. sub Triebner 835 (not preserved).
angustifolia: narrow leaved.
Rosette to 40mm φ, proliferous, stemless. Leaves 10-40, slender, erect to 100 X 10mm, lanceolate‑acuminate, sub-flaccid, brownish to dark green, margins and keel finely denticulate. Inflorescence to 200mm, lax. Flowers 8‑10, white to dull pinkish‑white.
1982 – The introduction of this species to Kew in 1824 is attributed to Bowie. It is a small clump-forming species with long narrow leaves, which has frequently been confused with H. chloracantha. However, it is a softer species and the marginal teeth are smaller and more closely spaced. The range of variation in both species is fairly similar. Smith’s var. paucifolia was based on a miniature form from Coombes, east of Grahamstown. Here the plants are miniaturised and growing within clumps of Restio among proteas. Across the valley, in grassveld, the plants are much more robust and this is the origin of var. grandis. Small forms are also found in grassveld along the edges of the lower Fish River Valley where only the short leaf-tips are visible above the soil in exposed rock cracks and crevices, sometimes shaded by grass. The Fish River seems to be the eastern limit and from there H. angustifolia extends westwards to the Zuurberg mountains. At Riebeeck East the leaves are particularly slender and this is probably the most attractive form to grow.
Haworthia angustifolia var. angustifolia JDV92/86 Coombs. Leaves are usually erect and straight.
H. baylissii was described from the farm Oudekraal not far from the Zuurberg Pass. It was first described as having broad, highly recurved leaves but plants collected by Bayliss at the same time and place, and grown at Kirstenbosch, were typical of normal H. angustifolia. Plants from the type collection also do not retain their character in cultivation although the leaves remain very deltoid and spreading. This prompted the idea that H. baylissii was only an odd form or hybrid of H. angustifolia. A visit to the type locality in 1979 confirmed this view (of form) as vast numbers of H. angustifolia were found growing on a rocky sandstone knoll overgrown with fynbos species (Protea spp., Restio spp.). This knoll was above a steep sided valley with the rocky faces clothed with succulents. H. glauca and H. cooperi were both present and seen in the immediate area. H. angustifolia must be considered to be a species of the False Macchia associated with acid quartzitic soils and in a moderate rainfall area. The False Macchia peters out west of the Zuurberg and occurs in small isolated patches in the Klein Winterhoek Mountains and Groot River Heights, apart from the main body further to the southwest. These areas have not been fully explored and it is possible that a relationship may extend between H. angustifolia, H. zantneriana and even H. divergens. H. angustifolia has been known to hybridise with H. cymbiformis at Howiesonspoort near Grahamstown, and the hybrid was described as H. perplexa V.Poelln.
H. angustifolia is easy to grow and the ease with which some forms proliferate may be an embarrassment. Under shaded conditions the leaves may also spread too much and lose the very dark colour that well-grown plants should have.
1999 – Scott selected a specimen to represent his concept of the species without specifically designating it as the type. The illustration in Salm-Dyck is available as the oldest and most generally used reference for the name, despite Fourcade’s discounting Baker’s description of H. angustifolia (see under H. monticola). The type selected by Breuer and Metzing is the specimen cited as representative for the species. It is an unfortunate choice because it represents a variation and is not typical of the species.
There have been no new collections to suggest any changes to the concept of this species. G. Marx has commented on the leaf length changing with favourable growing conditions and Cumming also correctly states that the var. paucifolia loses its distinctiveness in cultivation. Despite this it has been decided to recognise the following varieties:-
a. var. angustifolia This is most commonly found in the environs of Grahamstown and has been collected as far west as the Zuurberg (NE. Kirkwood) where it occurs together and appears to intergrade with the var baylissii. It is not known east of the Fish River nor further north than Riebeek East. Scott refers to records from the south-western districts but these are unquestionably mis-identifications of either H. chloracantha or H. divergens. The latter both to a degree have markings on the leaves and in neither do the leaf-tips incline inwards as they do in H. angustifolia.
Distribution: 3325 (Port Elizabeth): Uyepoort, Zuurberg (-AD), C.H.F. Woolley (NBG); Somerset East to Zuurberg (-BB), Holland in NBG18/25 (BOL); Kommadagga Stn. (-BB), Stayner in KG688/71 (NBG); Annsvilla to Uitenhage (-BD), Smith 2540 (NBG). 3326 (Grahamstown): Albany(-AB), Britten in PRE 34949; Alicedale (-AC), Stayner in KG251/70 (NBG); Wireless Stn. (-AD), Highlands (-AD), Smith 7270, 7449 (NBG), Cooke in NBG68977; Fort Collingwood (-AD), Smith 639 (NBG); Howiesonspoort (-AD), Smith 911, 7440 (NBG); Timm (NBG); Grahamstown (-BC), Breijer in TRV 23226 (PRE); Mt. Drive (-BC), Dyer 2280 (PRE); Mt. Drive (-BC), Smith 5630 (NBG), Barnes (BOL), Dyer 5; Hospital Hill (-BC) Smith 5656 (NBG); Near Coombes (-BD), Smith 5216 (NBG, PRE); Trappes Valley (-BD), Britten (BOL); Bathurst (-BD), NBG 68060; Lincoln Siding (-DA), Smith 3839 (NBG). 3227(Kingwilliamstown): Paradisekloof, Britten 5676 (NBG);
Inadequately located: Grahamstown, Whitworth in NBG 901/23 (BOL); Bolus (BOL), van der Merwe 115 (BOL).
Haworthia angustifolia var. angustifolia JDV89/38 Woesthill Pass, east of Grahamstown. May develop large mats under shade.
Haworthia angustifolia var. angustifolia JDV97/67 east of Grahamstown. Purplish shades develop in the leaf colour.
b. var. altissima Bayer var.nov. Type: CAPE-3326(Grahamstown): Riebeek East to Carlisle Bridge (-AA), Smith 5220 (NBG, Holo.).
altissima: tallest
Rosette proliferous. Leaves slender, erect to 150 X 10mm. Finely denticulate along the margins and keel. Leaf colour tending to greyish green rather than darker green. (A var. angunstifolia foliis griseo-viridibus erectioribus gracilibusque differt).
The var. altissima is created to accomodate a more distinctive taller element with more upright slender leaves than the var. grandis of Smith. The leaves in that variety are not particularly long for the species and neither this nor the distribution are as significant as may have been originally thought.
Distribution: 3326 (Grahamstown): Riebeek East to Carlisle Bridge (-AA), Smith 5220 (NBG); Sidbury (-AC), Smith 7450 (NBG); 32km Grahamstown to Alicedale (-AD), Bruyns 1653 (NBG).
Haworthia angustifolia var. altissima JDV97/66 Riebeek East. Note the greyer coloura of the leaves and the more conspicuous marginal spines.
c. var. baylissii (Scott) Bayer comb.nov. H. baylissii Scott, JS.Afr.Bot. 34:1(1968). Bayer :102(1976). Bayer, Nat.Cact.Succ.J 37:31(1981). Scott :101(1985). H. angustifolia fa baylissii (Scott) Bayer :26(1982): Type:- CAPE-3325 (Port Elizabeth): Oudekraal, Zuurberg (-BC), Bayliss in Scott 796 (PRE).
baylissii: for Col R.D.A. Bayliss.
P.V. Bruyns collected at the Oudekraal locality in 1993 and found again the forma baylissii, while independently Ernst van Jaarsveld recorded plants similar to the ordinary var. angustifolia from the same locality. The account in the New Handbook states that the Bayliss type collection represented at Kirstenbosch was atypical of the form described in that the leaves were not recurved. My own collection from Oudekraal was also not at all typical for Scott’s species as the leaves of the plants were not at all recurved and only slightly broader that in plants of the Grahamstown area. Neither this collection nor van Jaarsveld’s included any really broad leaved plants (nor with recurving leaves) which resembled true baylissii. Bruyns’ collection was made some 2-3km lower down the river valley where the plants were growing on rather sheltered rock ledges. The leaves of the plants were broader than either my or van Jaarsveld’s collections. The contention is that Scott’s species has neither the distribution, variability nor co-occurrence (other than with H. glauca and H. cooperi var. pilifera), to justify that status. Varietal status is propose because of the Bruyns re-collection. The concept is broadened somewhat to include plants with both narrower and non-recurved leaves.
Distribution: 3325 (Port Elizabeth): Zuurberg, Oudekraal (-BC), Bayliss in Scott 796 (PRE); Oudekraal (-BC), Scott in KG 416/70 (NBG); Oudekraal (-BC), Bayer 2052 (NBG).
Haworthia angustifolia var. baylissii co-type, Oudekraal. Only recorded from the Zuurberg at one locality, and most plants in the population have much narrower leaves than as described. This plant was grown at Kirstenbosch.
d. var. paucifolia Smith, JS.Afr.Bot. 14:48(1948). Type:- CAPE-3326 (Grahamstown): Frazers Camp (-BD), Smith 6819 (NBG).
paucifolia: with few leaves.
This variety is fairly abundant along the lower Fish River around the old Chalumna Police Station. This is a different grassland habitat to that at Frazers Camp where it was first recorded in Fynbos. This variety is not proliferous and the leaves flex outward. It is very small, with few leaves, seldom exceeds a height of about 7cm and the body of the plant is usually well drawn into the soil. These differences are not maintained in cultivation.
2. Haworthia arachnoidea (L.) Duval, Pl.Succ.Hort.Alenc. :7(1809). Haw., Syn.Pl.Succ. :96(1812). nec Pole Evans, Flow.Pl.S.Afr. 7:t.248(1927). Bayer :97(1976). Bayer :27(1982). nec Scott, Cact.Succ.J(U.S.) 49:205(1977). nec Scott, Aloe l6:41(1978). nec Scott :66(1985): Aloe pumila var. arachnoidea L., Sp.Pl. 1:322(1753): Aloe arachnoides Thunb., Diss. :311(1785). D.C., Pl.Gr. :f.50(1799). Ker-G., Bot.Mag. :t756(1802). Haw., Trans.Linn.Soc. 7:10(1804). S.D., Monogr. 12:t.2(1840). Lectotype: icones :78,t27, Comm., Prael.Bot. :78(1703). Epitype (B&M): CAPE-3319 (Worcester) Buitenstekloof (-DB), Bayer 153 (NBG). H. arachnoidea var. minor Haw., Suppl.Pl.Succ. :52(1819). Type: Not preserved.
arachnoidea: spider-like.
Rosette stemless, variable in size from 60 to exceptionally 120 mm φ, solitary or forming small clusters. Leaves 25-80 dense incurving, uniformly light to dark-green, no translucence and only occasionally faintly reticulate, flattened and often drying grey‑white to brownish at the tips forming a protective cover for the leaves. Triangular‑ to ovate‑lanceolate, 20‑70 X 10‑15mm, keeled, margin and keels bearing translucent spines to 12 mm in length, apex acuminate‑aristate. Inflorescence to 300mm. Flowers 20-30, perianth white.
1982 – There is some controversy and confusion over the application of the name H. arachnoidea. The difficulty stems from the interpretation of an illustration dating back to 1703. However, the interpretation arrived at by Salm-Dyck appears reasonable and in the light of his illustration it is difficult to understand why H. setata was not earlier reconciled with this species. H. arachnoidea is recognised by its uniformly plain green leaves. The ‘cobweb’ suggested by the name, is derived from the many white hairs on the leaf margins and keel, which in a healthy well-grown specimen do indeed resemble a spider-web. However, Ker-Gawler (1804) did not like this name ‘cobweb aloe’ given by Miller (1768) because he considered that the spined leaves more resembled the legs of a spider and hence the name. The plants are variable in size even within populations. Normally they do not exceed 60 mm in diameter but in some populations magnificent specimens of over 120 mm diameter may occur. The distribution does not coincide distinctly with any of its likely relatives and a great deal more information will be required before its relationship to any other species is better known. H. arachnoidea occurs in Namaqualand and in the Ceres Karoo. It is replaced on the Roggeveld Plateau near Sutherland and then eastwards, by H. semiviva. Plants from the Merweville area suggest an affinity with H. decipiens but are without the rather broader, flatter leaves and more deltoid spines characteristic of that species. H. arachnoidea occurs recognisably with H. archeri at Matjesfontein. Forms further to the southwest have marked leaves and are a deeper purplish-green so that there may be some difficulty in reconciling them with H. arachnoidea. There is no difficulty in recognising H. arachnoidea in the Worcester/Robertson Karoo, where it is quite common. It is poorly known in the Montagu area apart from at Cogmanskloof, Baden and the Ouberg Pass. A rather depauperate form is found in the Tradouw Pass south of Barrydale, and in the wider Ladismith area it is replaced by either H. unicolor or by H. habdomadis. However, near Ladismith itself H. arachnoidea is small and densely white-spined – more coarsely so than in the Oudtshoorn area where the plants are often small and the spination is finer and a little softer too (see H. aranea). A weaker spined but very proliferous form occurs in Meiringspoort, east of Oudtshoorn. The distribution eastwards from here is not clear. H. decipiens occurs at Klaarstroom north of the Swartberg mountains and it is possible that odd collections east of Uniondale, attributed to H. arachnoidea may in fact be either this species or H. unicolor. The illustration in Flowering Plants of South Africa (1927) is clearly of H. cooperi. H. arachnoidea is generally not proliferous and offsets very slowly if at all. Its distribution is clearly that of a winter-growing species and its occurrence on south slopes and shaded under bushes suggests the treatment it should get in cultivation.
1999 – The 1703 illustration referred to above is the illustration t27 in Commelin’s Praeludia Botanica. Bayer (in Excelsa 12:91, 1986) discusses the problem of interpretation surrounding this illustration and the associated confusion, pointing out that Scott’s analysis of the picture does not support his argument. Thus Scott uses this illustration to typify his concept of H. arachnoidea which is the same as Bayer’s H. herbacea in this work. Kerr-Gawler disliked Miller’s name ‘cobweb’, preferring the likeness to the spined leg of a spider rather than to the woolliness of its web. General usage seems to have favoured the use of ‘arachnoidea’ in terms of woolliness rather than simply spiny. If one looks at the illustration Botanical Magazine t.1417 that is used by Scott to typify his concept of H. translucens (viz. Aloe arachnoidea var. translucens (Haw.) Ker-G.) the situation is further confounded. The illustration in black and white indicates a species vegetatively fairly similar to the t27 of Commelin. Scott applies the epithet to what, in terms of its reported location, can only be one of three species viz. H. arachnoidea (H. setata sensu Scott), H. pehlemanniae or H. marumaina var. archeri (H. marumiana sensu Scott). The herbarium specimen referred to by him is certainly of the latter, while the illustration of his H. translucens appears to be of H. arachnoidea. It is not clear what Haworth actually intended in his Revisions when he created the section Arachnoideae and omitted mention of H. arachnoidea. I assume that he simply subsumed it under his H. setata. When Salm-Dyck discussed Aloe setosa he seemed to suggest that it was a remnant not adequately covered by Haworth’s description. Typification and usage of these old names is problematic and the scheme followed here does not have a flawless rationale. However, Salm-Dyck’s illustrations of both Aloe arachnoides and Aloe setosa are certainly not of H. herbacea as perceived in this work. In ascribing the type to the Worcester/Robertson Karoo there is an inherent problem in that elsewhere there will be a residue which may not be well accommodated in other varieties. However, the name setata is taken up for the more centrally situated variety which will better do that.
The varieties are largely geographic entities. The discovery of H. nortieri var. pehlemanniae (Scott) Bayer, as well as that of variants of H. marumiana var. dimorpha, which are vegetatively practically inseparable from H. arachnoidea, create problems. Var. pehlemanniae grows in very close proximity to H. arachnoidea var. scabrispina from which it quite distinctive, but it is very difficult to distinguish it from some of the other varieties without flower or habitat information. Possibly the former has leaves which tend to be more flaccid and thicker when fully turgid. We already know that H. mucronata var. mucronata may probably be the origin of arachnoidea-like plants in the Tradouw Pass. A similar transition of a deviant population of H. mucronata from north of Montagu, to the Cogman’s Kloof, also occurs. In the widest context, H. arachnoidea is impinged on by all three varieties of H. nortieri, as well as by H. marumiana var. archeri, H. mucronata, and H. decipiens. It is quite probable that correctly interpreted, more than one species may be involved in the concept of H. arachnoidea given here. However, there is no reason why over its whole range it should not exhibit the same degree of variation that for example H. magnifica or H. turgida exhibit over their comparatively restricted ranges. The species does hybridise with H. blackburniae var. blackburniae and var graminifolia, as well as with H. truncata.
a. var. arachnoidea The form common in the Worcester/Robertson Karoo is taken as the typical variety although the choice is arbitrary. In this area there is no confusion with other species and although it grows in close association with a number of other species, there is no evidence of hybridisation. This variety often occurs on southern slopes where it is embedded in moss and lichen, and thus very moist in the winter months. The plants can reach as much as 18cm in diameter. The basic leaf coloration is darkish-green and the marginal spines are white. The terminal bristles tend to be blackish.
Distribution: 3319 Worcester): Hex River (-BD), Bruyns 2411 (NBG); Tonnel Stn. (-BD), Bayer in KG36/71 (NBG); Worcester (-CB), Fouche 14 (PRE); 8km NE. Worcester (-CB), Bayer in KG 640/69 (NBG); Worcester (-CB), Marloth 12572 (PRE), Venter 3 (BOL); Effata (-DA), Bayer in KG 118/70 (NBG); Buitenstekloof (-DB), Bayer 153 (NBG).
Haworthia arachnoidea var. arachnoidea JDV86/109 east of Worcester. The typical variety occurs in the Worcester/Robertson Karoo and the end-awn tends to be blackish.
b. var. aranea (Berger) Bayer comb.nov. H. bolusii var. aranea Berger, Das Pflanzen. 33:114(1908). Scott, :72(1985), not as illustrated. H. aranea (Berger) Bayer :98(1976). Bayer :29(1982). Lectotype: icon :114,t39, Berger, Das Pflanzen. 33(1908). Epitype (B&M): CAPE-3322 (Oudtshoorn): Moeras River (-CA), Bolus 12372 (BOL).
aranea: cobweb.
1982 – This species is described and illustrated in Berger’s revision. The first comparable plants seen by the writer were from the fynbos vegetation between Oudtshoorn and Uniondale. The uniform green coloration and soft canopy of hairs, as well as the existence of larger similar forms near Oudtshoorn itself, suggested an affinity with H. arachnoidea rather than with H. bolusii. Since then a much smaller proliferous form of H. bolusii has been discovered by Peter Wilkins east of Jansenville which could well be closely related to H. aranea. Here the plants have the typical bluish green colour and translucence of the upper leaves as in H. bolusii. They occur in a flaking blue shale. Plants close by in compacted brown shale are larger and with fewer firmer hairs. It appears more likely that Berger’s plants originated from the Oudtshoorn area. Until the complete picture of species distribution is available H. aranea is being retained.
1999 – No new information suggests any change here and it has long been known that it intergrades with H. arachnoidea var. setata around Dysseldorp and De Rust as well as in Schoemanspoort. Thus H. arachnoidea in itself is so variable that the pressure for consistency seems to call for varietal rank. Col Scott maintains this element as a variety of H. bolusii and cites a specimen which is apparently not that species at all, disregarding the comments made above. Scott’s illustration also does not accord with the type of var. aranea species but nevertheless appears to represent H. arachnoidea. The earlier comment ‘until the complete picture…’ is nonsense and it is unlikely that there can be new evidence to prove the original species closest to either bolusii or arachnoidea; nor likely or even possible that distribution data will clarify a position clouded by history. This variety is characterised by its small size (to 6cm diam.) and very dense soft spination. Despite the comments made in 1982 concerning colour and translucence, there opacity of the leaves of this variety (and the species) is often forgotten.
Haworthia arachnoidea var. aranea JDV90/59 Vergelegen. Always with very narrow leaves and soft spines.
Haworthia arachnoidea var. aranea JDV97/108 north of Schoemanspoort. The accumulation of old black leaves is very characteristic.
c. var. namaquensis var.nov. Type: CAPE-2917 (Springbok): Karrachabpoort, Richtersveld, Bayer 1674 (NBG, Holo.).
namaquensis: from Namaqualand.
Rosette as for species, to 60mm φ. Leaves paler green. (A var. arachnoidea foliis subviridibus et in terra valde submersa differt).
The var. namaquensis has not been previously accommodated. It is essentially recognised for its geographic association and applies to populations in Namaqualand, Bushmanland and the Ceres Karoo. The spination is usually moderate and the plants are usually lighter coloured than elsewhere in the species. It also differs from the typical variety in being pale green and well-drawn into the soil.
Inadequately located: Little Namaqualand, Britten in PRE 34983.
Haworthia arachnoidea var. namaquensis JDV91/43 north of Bitterfontein. Invariably in the shade of bushes or stones.
Haworthia arachnoidea var. namaquensis JDV94/24 east of Springbok. The plants are usually withdrawn into the soil like this.
Haworthia arachnoidea var. namaquensis JDV91/155 Umdaus. This variety seems to flower in early spring.
d. var. nigricans (Haw.) Bayer comb. nov. H. setata var. nigricans Haw., Revis. :56(1821). Type: Not preserved. Neotype (designated here): CAPE-3321(Ladismith): SW. Vanwyksdorp, Bayer 2419 (NBG): H. helmiae V.Poelln., Feddes Repert.Spec.Nov. 41:201(1937). Scott :99(1985). H. unicolor var. helmiae (V.Poelln.) Bayer : 121(1976), Bayer 59:(1982): Type: Cape, Heidelberg, Smithers in Triebn. 901, Gt Brak River, Mrs Helm in Triebn. 901. Not preserved. Lectotype (B&M): icon, (B). Epitype (designated here): CAPE‑3322 (Oudtshoorn): Schoemanspoort (‑AC), Bayer 171 (NBG). H. venteri V.Poelln., Cactus.J 8:19(1939). H. unicolor var. venteri (V.Poelln.) Bayer :166(1976). Bayer :59(1982): H. aristata sensu Scott :110(1985) Type: Cape, Barrydale Distr. Major Venter (not preserved). Lectotype (designated here): CAPE-3320 (Montagu): Barrydale to Lemoenshoek (-DC), Smith 3994 (NBG).
nigricans: blackish, for the dark hues.
1982 – The type locality for H. venteri was recorded by Smith as ” 3/4m. Barrydale‑Lemoenshoek”, although Major H. Venter’s localities are very broadly stated in publication and practically valueless. Only Smith’s notes give any better clue to these but in this case there is still no certainty. However, there is a very common species extending from Barrydale to Oudtshoorn from which this variety is clearly drawn. At Barrydale itself the plants are rather pale green with elongated sparsely setate leaves – this is regarded here as the species H. unicolor. Eastwards the plants are more robust and often glabrous, usually distinguished by particularly dark‑purplish coloration towards the leaf tips. The further eastwards one progresses the more setate the plants become until at Oudtshoorn, south of the Cango Caves, the plants are comparatively small and more softly setate (H. unicolor var. helmiae).
It is not in the least clear what the relationship is between H. unicolor and H. arachnoidea. They do not appear to cohabit which suggests that they may be conspecific and only differentiated on an ecotypic basis. This is borne out by the pale arachnoidea‑like population at the northern entrance to the Tradouw Pass, south of Barrydale. However at Ladismith H. arachnoidea is present as a very distinct heavily setate form, and similarly south of the Cango Caves it occurs near to H. unicolor var. helmiae as a softer haired but also highly setate form. The species H. unicolor is therefore not well‑known or properly understood and this solution must be regarded as suspect. Von Poellnitz’s citations of localities for the var. helmiae are highly confusing and in Feddes Repert. Spec. Nov. 44:223(1938) are totally disparate. The type is cited in 1938 as “Great Brak River, Mrs Helm” and three Triebner numbers are added to this. Mrs Helm’s strong personal recollection (private communication, and acknowledgement to Col C.L. Scott) is that the original plants were collected at a specific site south of the Cango Caves, Oudtshoorn. The photograph extant in the Botanical Museum Dahlem (which must serve as the type) is of a very poor specimen, but it can be reasonably allied with plants from the site given by Mrs Helm. It is concluded that it is an extension of the complex to which it is here referred. J.R. Brown’s illustrations in Cact.Succ.J(U.S.) 18:39(1946) are not of this variety.
1999 – The incorporation of H. unicolor var. venteri under H. arachnoidea as the var. nigricans of Haworth represents a major change. Smith does state that his collection is from the locality to which he was directed by Col Venter. The type selected by Breuer and Metzing for the name venteri, is doubtful because of the locality cited. The initial problem was to relate unicolor (= H. mucronata) to a substantial species body, which I could not previously do. How this problem is resolved is also discussed under H. mucronata. It is clearer now that var. nigricans is better associated along its northern and western borders with var. arachnoidea and there are areas where it is not possible to make a distinction between these two varieties. This realisation suggests the re-application of Haworth’s original name which means blackish-green. From records of Dekenah’s collections it appears that var. venteri does interface with H. mucronata in the southwest and I can confirm this from my own collection from south of Vanwyksdorp.
Regarding H. helmiae: neither of the three numbers given by von Poellnitz in 1938 agrees with Heidelberg, Smithers in Triebner 901 as in the original description, to which was added a collection of Mrs Helm’s from Great Brak. In addition Von Poellnitz cites several quite disparate and improbable localities between Worcester in the west and Avontuur in the east. Where H. helmiae was reputed to have been actually collected (Schoemanspoort), fairly glabrous forms occur together with softly spinose ones. These are small dark elements with very reduced spines and often without spines at all (they do not resemble the softish translucent plants figured by Brown as mentioned above). They must be closely associated with the var. nigricans (previously unicolor var. venteri) and the change of interpretation here makes it very much easier to accommodate a very wide range of forms which tend to be glabrous and without any spines.
Southwest of Oudtshoorn var. nigricans can be robust with heavy spines and it is probably here that H. ferox var. armata V.Poelln. originated. The point is made that H. arachnoidea does have glabrous variants and they are generally accommodated in this variety.
It is distinguished by the purplish coloration towards the tips of the leaves and the generally darker colour. The leaves tend to be broader and the spines larger and sparser than for the species generally. Glabrous forms with pronounced keels commonly occur and the more delicate softer versions of this can perhaps be ascribed to H. mucronata var. integra. These variants of H. mucronata are distinguished by their translucent margins and keel. The interaction between that species and H. arachnoidea is an interesting field for investigation.
Distribution: 3320 (Montagu): Jagerskraal (-AB), Bayer 3612 (NBG); Kareevlakte (-AD), Archer (BOL); SE. Konstabel Stn.(-BC), Bruyns 2448 (NBG); Elandskloof (-BD), Bruyns 2418 (NBG); Prinspoort (-BC), Venter 91/114 (NBG); Keurfontein (-BD), Bruyns in KG18/76 (NBG); 24km Montagu to Ladismith (-CB), Esterhuysen (BOL); Bellair Dam (-DA), Venter 5 (NBG); E. Barrydale (-DC), Bayer 4671 (NBG), Stanford (BOL); W, Barrydale (-DC), Smith 5770, 7310 (NBG); Brandrivier (-DD), Smith 3995 (NBG); Near Barrydale (-DD), Smith 5672, 3994 (NBG); E. Lemoenshoek (-DD), Hurling & Neil (BOL), Bayer in KG126/72 (NBG), Stayner in KG15/67 (NBG); Warmwaterberg (-DD), Bayer 1704 (NBG); W. Warmwaterberg (-DD), Bayer 4575 (NBG). 3321 (Ladismith): N. Calitzdorp (-BC), Bayer in KG441/75 (NBG); Winkelplaas (-CA), Bayer 2716 (NBG); S. Ladismith (-CA), Smith 5506 (NBG); Ockertskraal (-CA), Smith 4004, 5510 (NBG); W. Ladismith (-CA), Smith 7140 (NBG), Bayer 1624 (NBG); Die Eike (-CA), Laidler 314 (NBG); SW. Ladismith (-CA), Bayer 1615 (NBG); Kareebosch (-CA), Laidler 481 (NBG); Adamskraal (-CA), Smith 4002 (NBG), Bayer 1617 (NBG), Ferguson 5 (BOL); Ladismith to Vanwyksdorp (CB), Bayer in KG573/51 (NBG); Springfontein (-CC), Smith 5657, 7325 (NBG), Bayer in KG 166/71 (NBG); W. Springfontein (-CC), Smith 5379 (NBG), Bayer in KG167/71 (NBG); Muiskraal (-CC), Smith 3989, 7136 (NBG), Bohnen 8416 (NBG); Riversdale to Ladismith (-CC), Smith 537 (NBG); Muurkeeskraal (-CC), Smith 4000, 3976, 3995, 3997, 7138 (NBG); W. Vanwyksdorp (-CD), Bayer 2418 (NBG), Stayner in KG57/67 (NBG); S. Vanwyksdorp (-CD), Bayer 2419 (NBG); S. Calitzdorp (-DA), Bayer in KG126/72 (NBG), Bayer in KG125/73 (NBG); Oudtshoorn to Calitzdorp (-DB), Bayer 1616 (NBG); Warmwaterbron (-DB), Bayer in KG115/71 (NBG). 3322 (Oudtshoorn): Schoemanspoort (‑AC), Bayer 171 (NBG); N. Oudtshoorn (-AC), Bayer 4499 (NBG). Volmoed (-CA), Bayer 4656 (NBG), Zeekoeigat (-CA), Gie (NBG); Mt Hope (-CB), Bruyns in KG437/75 (NBG).
Inadequately located: Near Doornrivier, Helm in Fourcade 4740 (BOL); Caledon, Venter 9 (BOL);
Haworthia arachnoidea var. nigricans JDV south of Calitzdorp. In close proximity to H. mucronata var. rycroftiana,
Haworthia arachnoidea var. nigricans JDV92/52 west of Ladismith. This area has produced the var. setata and H. mucronata var. inconfluens.
Haworthia arachnoidea var. nigricans JDV94/31 west of Muiskraal. Glabrous forms grow intermingled with setate forms.
Haworthia arachnoidea var. nigricans JDV89/57 Anysberg. This is one of the few places where the var. setata and H. mucronata are also present.
e. var. scabrispina var.nov. Type: CAPE-3320(Laingsburg): Baviaans (-BA), Bayer 2105 (NBG. Holo.).
scabrispina: rough-spined.
Rosette roundish and raised above ground level. Leaves with firm rigid brownish spines. (A var. arachnoidea spinis rigidis bruneolis differt).
The var. scabrispina is created to accommodate the southern Karoo elements which have very hard stiff spines. The plants tend to be raised and the leaves and spines are fairly rigid so that the plants form quite rounded spheres rather than flattened rosettes. There are populations in which the plants have the spines produced from translucent raised bases and also in which the upper petals do not curve upward. This seems to suggest some affinity with H. marumiana var. archeri.
Distribution: 3220 (Sutherland); 35km N. Laingsburg (-DC), Bayer 2124 (NBG). 3221 (Merweville): Schoppelmaaikraal (-CD), Bruyns in KG30/76 (NBG). 3320 (Montagu): N. Baviaans Stn. (-BA), Bayer 2105 (NBG); Whitehill Ridge (-BA), Barker 13368 (NBG), Archer (BOL); Laingsburg (-BB), Lewis & Barker in NBG2772/32 (BOL); Nougaspoort (-CA), Bayer 1963 (NBG); 20km W. Ladismith (-CA), Bayer 4462 (NBG).
Haworthia arachnoidea var. scabrispina JDV91/111 Rooinekpas. There are several collections of this dark form which has similar flowers to H. marumiana var. dimorpha.
Haworthia arachnoidea var. scabrispina JDV89/59 south-west of Lainsburg. Grows very near to H. nortieri var. pehlemanniae, but is usually on shadier south-facing slopes.
Haworthia arachnoidea var. scabrispina JDV95/31 south-east of Laingsburg. With very rounded rosettes and brown end-awns.
f. var. setata (Haw.) Bayer comb. nov. H. setata Haw., Suppl. Pl.Succ.:52(1819). Brown, Cact.Succ.J(U.S.) 16:3(1944). Scott, Aloe 16:42(1978). Scott :68(1985). Type: not preserved. Neotype (see Scott, 1985): icon, artist unknown, specimen received from Dr Mackrill ex Cape, (K): H. setata var. media Haw., Revis. :56(1821). Type: Not preserved: H. setata var. major Haw. ibid. Type: Not preserved: H. gigas V.Poelln., Feddes Repert.Spec.Nov. 31:84(1932). H. setata var. gigas ibid. 44:224(1938). Type: Cape, Amalienstein, Stellenbosch 6692 (not preserved). Neotype (designated here): CAPE-3321(Ladismith): Amalienstein, Smith 7390 (NBG): H. minima var. major V. Poelln., Kakt.u.and.Sukk. :39(1938). H. tenera var. major Uitew., Sukkulenta :52(1948). Type (designated here): Cape, Derust, Mrs Helm (not preserved). Neotype: CAPE-3322(Oudtshoorn): Meiringspoort (-BC), Bayer in KG 160/72 (NBG): Aloe setosa Roem. et Schultes, Syst.Veg. 7:641(1829). Salm-Dyck, Monogr. 12:f3(1841). Type: Not preserved. Neotype (designated here): icon, f3 Salm Dyck, Monogr. 12(1840).
setata: bristled.
This variety tends to have dense white spines which do not blacken on drying, and the texture of the species is always softer than the var. scabrispina. It is quite a variable element and like the var. nigricans is also closely interwoven with H. mucronata. Although usually moderate in size and only upto about 8cm in diam., some robust specimens occur east of Ladismith. Two specimens are cited below from the Jansenville district and it is more probable that they belong with H. decipiens var. cyanea.
Distribution: 3224 (Graaff Reinett): Jansenville (-DC), Long 1321 (PRE); Jansenville (-DC), Marloth 9384 (PRE). 3320 (Montagu): Keurkloof (-BC), Barker 1913 (NBG); Elandskloof (-BD), Bruyns 2259 (NBG); Buffelsrivier Pass (-BD), Bruyns 2564 (NBG); Cogmans Kloof (-CA), Esterhuysen (BOL), Hurling & Neil (BOL), Long 1124 (PRE), Smith 7224 (NBG), Stayner in KG38/67 (NBG), Otzen in NBG 1316/35, Barker 8266 (NBG), Smith 3989, 7490 (NBG); Prins River (-DB), Bruyns 2462 (NBG); Barrydale (-DC), Smithers (BOL); Tradouw Pass (-DC), Hurling & Neil (BOL), Smith 3991, 6782, 6789 (NBG), Thompson 664 (NBG). 3321 (Ladismith): Amalienstein (-AD), Herre in Stell.6639 (BOL), Smith 7390 (NBG); Huis River Pass (-BC), Smith 7389 (NBG); Opsoek (-BC), Smith 6886 (NBG); S. Ladismith (-CB), Bayer in KG 160/72 (NBG), Smith 5507 (NBG), Joubert 10 (BOL); 5km S. Ladismith (-CB), Smith 5652 (NBG); 6km S. Ladismith (-CB), Smith 5729, 5733 (NBG), Stayner in KG42/70 (NBG); 14km E. Ladismith (-CB), Bayer in KG593/71 (NBG); W. Vanwyksdorp (-CB), Bayer in KG535/71 (NBG); Groenfontein (-DA), Smith 6888 (NBG); Calitzdorp (-DA), Fouche 53 (PRE); E. Vanwyksdorp (-DA), Bayer 5753, in KG388/71 (NBG). 3322 (Oudtshoorn): 32km S. Prince Albert (-AC), James (BOL), Taute (BOL); Boomplaas (-AC), Moffett 141 (NBG); Schoemanspoort (-AC), Bruyns in KG92/77 (NBG), Smith 5230 (NBG); Meiringspoort (-BC), Bayer in KG160/72 (NBG); Oudtshoorn (-CA), Smith 5785 (NBG), Bolus 12375 (BOL); N. Dysselsdorp (-CB), Bayer & Venter 6608 (NBG). 3324 (Steytlerville): N. Steytlerville (-AC), Bayer & Venter 6610 (NBG).
Inadequately located: Ladismith, Jordaan (BOL), Joubert 11 (BOL); Sundays River Estate, Heatlie in NBG254/25 (BOL).
Haworthia arachnoidea var. setata JDV94/86 Dyseldorp. There are also populations of vr. aranea in the area.
Haworthia arachnoidea var. setata JDV93/23 east of Steytlerville. A population north of Steytlerville flowers in September at the same time as the var. aranea. Considered to be tending towards the var. xiphiophylla which also flowers early.
Haworthia arachnoidea var. setata JDV92/93 east of Anysberg. Co-occuring with the var. nigricans. It does not have the purplish tips to the leaves.
Haworthia arachnoidea var. setata JDV90/70 De Rust. There are several populations like this which tend towards the var. aranea.
Haworthia arachnoidea var. setata JDV92/100 south-west of Ladismith. This very white-spined form is included in the var. setata.
g. var. xiphiophylla (Baker) Bayer comb.nov. Haworthia xiphiophylla Baker, Fl.Cap. 6:354(1896). Baker, Curtis’ Bot.Mag. t.7505(1896). Bayer :168(1976). Bayer :61(1982). Scott :73(1985). H. setata var. xiphiophylla (Bak.) V.Poelln., Feddes Repert.Spec.Nov. 44:225(1938). Type: Cape, Uitenhage, Howlett (K): H. longiaristata V. Poelln., Kakteenkunde 9:134(1937). Type: Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): N. Coega Station (-DA), Mrs E.B. King (NBG).
xiphiophylla: sword‑like leaves.
1982 – This is a bright green species with broad but short, well‑spaced marginal teeth. The leaves are long and slender. H. xiphiophylla is found in the vicinity of Uitenhage and Coega. It has been regarded as a variety of H. arachnoidea but this is highly unlikely from the viewpoint of distribution only. A number of anomalous populations at Coega, Dead Man’s Gulch and Kirkwood confuse the issue. Thus H. translucens and H. xiphiophylla pose an intriguing and interesting problem for the field worker.
1999 – The latter sentence is also fairly typical of the loose words used when the herbarium record is inadequate. This is a bright green variety with broad but short, well‑spaced marginal teeth. The leaves are long and slender. The incorporation into H. arachnoidea is to include the variables along the southern Swartberg mountains from Anysberg through to Steytlerville where the species meets with the Eastern Cape variants. One of the populations alluded to is represented in this work by the ‘re-discovered’ H. aristata. From the collections by J.D. Venter, it seems that var. xiphiophylla does merge directly with var. arachnoidea at its westerly limits. I would have expected a relationship with the greener, braoder leaved H. decipiens var pringlei as discussed under that species. However, var. xiphiophylla does not develop translucent windows and the leaf spines tend to be separate and narrow at the base. The flowering time appears to be early summer ahead of typical var. arachnoidea which flowers in the summer.
Distribution: 3324 (Steytlerville): 30km E. Steytlerville (-BC), Venter 85/68 (NBG). 3325(Port Elizabeth): Mentz Dam (-AA), Venter 91/122 (NBG); S. Mentz Dam (-AC), Venter 91/117 (NBG); Mannetjie (-CA), Venter 94/19 (NBG); Maraishoop (-CA), Smith 3583 (NBG); Bauerskraal (-CB), Venter 93/75 (NBG); Sandfontein (-CB), Archibald in NBG 1326/32; Uitenhage (-CB), Britten in PRE 34951; De Rust, 2km NW. Couga Kop (-DC), Branch 5 (NBG); Couga Kop (-DC), Smith 3546 (NBG), Britten (BOL).
Haworthia arachnoidea var. xiophiophylla JDV93/75 north-west of Uitenhage. Longer more slender leaves and relatively larger and more widely spaced spines.
Haworthia arachnoidea var. xiophiophylla JDV96/88 west of Coega. The plants are seldom solitary like this and dense stands may occur.
Haworthia arachnoidea var. xiophiophylla JDV94/98 Coega. The leaves are generally more yellow-green and the spines shorter and broader than in the other varieties.
3. Haworthia aristata Haw., Suppl.Pl.Succ. :51(1819), Rev.Pl.Succ. :58(1821). Non V.Poelln., Feddes Repert.Spec.Nov. 43:92(1938). Non Jacobsen 2:537(1954). Non Scott, Natn.Cact.Succ.J 35:12(1980). Non Scott :110(1985). Type: Cape, ex hort Kew. Not preserved. Lectotype (designated here): icon (K). Epitype (designated here): CAPE-3325(Port Elizabeth: Deadmans Gulch (Soutkloof) (-DA), Smith 3550 (NBG): H. denticulata Haw., Rev.Pl.Succ. :58(1821). Baker, JLinn.Soc. 18:213(1880). V.Poelln., Feddes Repert.Spec.Nov. 41:199(1937). non idem. 45:168(1938). Type: Cape, ex hort. Kew. Not preserved. Lectotype (designated here): icon, Kew library.
aristata: furnished with an awn.
Rosette stemless, proliferating slowly, to 6cm φ. Leaves slender, erect, incurved, dark-green, margins and keel entire or finely spined, with little translucence and faintly reticulated. Inflorescence simple, lax. Flowers 10-15, white.
In the first editions of the Handbook these two names were rejected as insufficiently known, and I did not agree with von Poellnitz’ use of aristata for his later H. unicolor. I mentioned too with regard to H. denticulata, that Von Poellnitz had consistently allied that name to the Hankey H. translucens (now gracilis) complex. However, von Poellnitz cites a number of improbable associates, further lists it as a variety of H. altilinea, and also describes it as having translucence. I do thus do also not agree with Scott’s 1985 interpretation where he overlooks the very translucent margins and keel of the species (see H. mucronata) to which he applies the name. It is only recently when I re-examined collections in the Compton herbarium, and a collection received from W.R. Branch from Addo, that it became apparent that Haworth’s two species can be allied to collections which have previously been hidden in H. gracilis. As applied here, it is indeed to a small dainty species which has very little translucence to the leaf. It is odd that the description as it appears translated in Scott (1980), reads ‘resembles a dark green cuspidata…’. Haworth’s description refers to Sempervivum cuspidatum and not to the big presumed hybrid Haworthia of that name. The late F.J. Stayner had in private communication commented on this odd species from Cougakop, east of Port Elizabeth. He collected specimens for the Karoo garden and these are still in cultivation. It was treated as a variant of H. xiphiophylla because it was not as translucent as most of the small elements in that area are, and because it was not substantial in terms of botanical record. Cougakop is now virtually quarried away. H. aristata is represented by about four collections from that area and also by three collections by P.V. Bruyns from further north and west. Here the plants do show some translucence and also tend to bluish-green, so it is possible that it may link up with small forms of H. decipiens. It should not be assumed that this element is really subtstantial enough as recent attempts to establish its validity have not been very successful. The collection from Mt Stewart cited below is probably H. decipiens var, minor.
Distribution: 3324 (Steytlerville): S. Mt. Stewart (-AB), Bruyns 1812 (NBG). 3325 (Port Elizabeth): Stonefountain (-BA), Bruyns 1654 (NBG); E. Verdun (-BB), Bruyns 1630 (NBG); Near Kommadagga (-BC), Smith 5890 (NBG); Soutkloof (-DA), Swart (NBG), Stayner in KG7/76 (NBG), Branch 459 (NBG); van Jaarsveld 6902 (NBG); Deadmans Gulch (Soutkloof) (-DA), Smith 3550 (NBG).
Inadequately located: Somerset East, Herre in STE6612 (BOL).
Haworthia aristata JDV87/53 Soutkloof. This is a photograph of a plant collected by W. R. Branch. Although there are several records of this element it has not been clearly shown to be a valid species.
Haworthia aristata JDV96/90 Soutkloof. This collection seems to more closely resemble an intermediate between H. gracilis and H. cooperi.
4. Haworthia bayeri Venter & Hammer, Cact.Succ.J. (U.S.) 69:75(1997). H. emelyae p.p. Bayer, figs. 10 & 11 in Natn.Cact.Succ.J 34:28(1979). p.p. Bayer :109(1976). Bayer :38(1986): H. willowmorensis sensu Scott, Aloe 11:8(1973). H. correcta sensu Scott :74(1985). Type: CAPE-3323 (Willowmore): Hills S. Uniondale (-CA), F.J. Stayner in KG164/69 (NBG).
bayeri: for M.B. Bayer.
Rosette stemless, 15-20 leaves. Leaves: retused, dark brownish-green to blackish- green, slightly scabrid, keels and margins with minute spines, or smooth, tip rounded and not pointed, the end-area opaque, cloudy transparent, with sparse reticulate patterning or longitudinal lines. Inflorescence to 300mm. Flowers 15-25.
This species is in some respect an embarrassment to me. The fact that it now bears my name may be a tribute but it also points to fallibility. It must surely be one of the more spectacular of the retuse-leaved species and my own two illustrations in 1979 indicate how clearly different it is to H. emelyae. It is only after the publication of the revised Handbook that I saw this species growing in the very near vicinity of H. emelyae var. comptoniana. I was also aware that a Dysselsdorp collection of mine flowered at a different time to H. emelyae. Dr W.R. Branch of Port Elizabeth had also sent me a plant from near the western end of the Baviaanspoort near Willowmore which was more like H. bruynsii than H. emelyae. David Cumming of Australia had also commented in his astutely observant way that there were two species involved. Robert Kent’s discussion of this species in Haworthiad (7:15, 1993) under the title H. emelyae, deals with the original description of H. correcta. His view is that while Mrs Blackburn may have collected H. bayeri at Uniondale as Scott contends, it was not the element described and illustrated by von Poellnitz. The photograph in Kakteenkunde is not the species from Uniondale and shows the leaves as pointed as they are in H. emelyae. My collections of H. bayeri were grown rather hard and were never free of surface encrustation, and neither did my specimens of H. emelyae ever reach a pristine condition. This further obfuscated already poor observation on my part. There is a reliable report of a collection of this species in the Rooinek Pass south of Laingsburg which constitutes a considerable extension of its range.
H. bayeri always has a rounder leaf-tip than does H. emelyae and in cultivation develops a dark green coloration. The translucence of the leaves is deeper and there is no flecking in the leaves as is the case in H emelyae. There is no significant difference in the habitats where the two species are found. H. bayeri is a small degree eastward from H. emelyae, north of the Kamanassie Mountains, and does not go much further west than Oudtshoorn. This species has the potential to produce the same stunning range of selected cultivars that Japanese growers have produced in H. truncata.
Distribution: 3321 (Ladismith): Rooinek Pass (-BB), Venter sn. (NBG). 3322 (Oudtshoorn): S. Oudtshoorn (-CA), Peers in NBG1940/37, Smith 5780, 5781 (NBG), Bayer in KG111/72 (NBG); De Rust (-BC), Smith 2061, 2062 (NBG); 3km S. De Rust (-CB), J. Scott 243 (PRE); Doringkloof (-CB), Bayer in KG146/72 (NBG); E. De Rust (-DB), Smith 2062, 4009 (NBG), Rossouw 404 (NBG). 3323(Willowmore); SW. Willowmore (-AC), Smith 5206 (NBG); E. Willowmore (-BC), Viviers 908 (NBG); S. Uniondale (CA), Stayner in KG164/69 (NBG).
Haworthia bayeri JDV91/152 De Rust. At the western end of the range the plants tend to be more scabrid.Haworthia bayeri JDV84/13 north-west of Dysseldorp. The only markings on the end surface are the leaf veins, in the field the plants themselves are difficult to see.Haworthia bayeri JDV93/26 south of Dysselsdorp. This is where it occurs in close proximity to H. emelyae.Haworthia bayeri JDV90/13 west of Georgida. Growing near H. emelyae var. comptoniana. The flowering times of the two species are staggered only by a few weeks across their ranges.
Roots fusiform. Rosette stemless, fibrous, 10-15mm φ at base, clumping. Leaves 10-15, long slender, to 400 X 1.5 to 5mm, channeled on the upper surface, margins glabrous or finely spined, colour bright green to brownish green or dark greyish green. Inflorescence simple raceme, to 300mm. Flowers white, green veined, 15-20.
a. var. blackburniae The typical variety shows considerable variation over its range. The leaves may be very long and slender, or even be squat, short and with a tendency to flex downward and sideways. Leaf coloration bright green.
1982 – This is one of two unusual species with long slender canaliculate grass-like leaves, fibrous dry leaf bases, and thick fusiform roots. It is common in quartzitic rock in the Rooiberg mountains southeast of Calitzdorp and extending to about 32 km west of Ladismith. The full extent of its distribution and hence also its variability, is not known. Originally found growing singly and exposed, it generally prefers a cooler protected southern aspect where it forms dense clumps. In some forms the leaves are only 60 to 80mm long but as much as 3mm wide. In others the leaves may be 200mm or more long and only 2mm wide. H. blackburniae is very slow and difficult in cultivation. The Little Karoo is technically a summer rainfall area but with rainfall peaks in March and September/October. The hot dry summers discourage plant growth and many of the indigenous plants respond best to the cool wetter winters.
1999 – Many new collections have precipitated the decision to now include H. graminifolia here. H. blackburniae has in recent times been collected northeast of Calitzdorp as well as much further west in the Touwsberg and Anysberg. There are variants with more slender leaves which are toothed. Most significant is the collection by Jan Vlok in the De Rust area which is given varietal status here. Mention of the bluer-green coloration of the leaves of var. graminifolia was omitted from my previous discussions. This colour difference seems to be the only significant character which can really contribute to the decision to recognise three varieties. There is speculation about the affinities of this species. The fibrous nature of the stem as well as the broad insertion of the leaf onto the stem suggest a close relation with H. wittebergensis, which is very similar in these respects. This relationship is confirmed by Dr M. Hayashi (priv. comm.) on the basis of similar karyotypes. Interestingly Smith had, in his records, proposed a variety ladismithensis for plants collected southwest of Ladismith which flowered two months ahead of the Calitzdorp collections. Thus while flowering time is significant it may also vary quite dramatically within a species.
Distribution: 3320 (Montagu): Prinspoort (-BC), Bruyns (NBG). 3321 (Ladismith): W. Ladismith (-AC), Bayer in KG100/74 (NBG); Hartmansberg (-BC), Bruyns (NBG); SW. Ladismith (CA), Smith 5505 (NBG); Assegaaibos (-DA), Bayer 4429 (NBG); Rooiberg (-CB), Acocks 20395 (PRE); Rooiberg (-DA), Smith 2062 (NBG), Bayer 4428 (NBG); 12km SW. Calitzdorp (-DA), Blackburn in Reynolds 1842 (PRE); Calitzdorp (-DA), Blackburn in NBG1174/36 (NBG); Blackburn Valley (-DA), Barker 5094 (NBG, PRE) Warmbaths (-DB), Bayer in KG100/740 (NBG) Inadequately located: ex hort. Thudichum in NBG130/43 (NBG); Calitzdorp, Blackburn in BOL21933, Barker 5094 (BOL).
Haworthia blackburniae var. blackburniae JDV92/51 west of Ladismith. The leaves can be very short and broad.Haworthia blackburniae var. blackburniae JDV90/20 south of Calitzdorp. Here clump-forming with quite long leaves.Haworthia blackburniae var. blackburniae JDV92/92 Anysberg. The plants here are not clump-forming.
b. var. derustensis var.nov. Type: CAPE-3222 (Oudtshoorn): West of De Rust (-BC), J. Vlok in J.D. Venter 93/24 (NBG, Holo.).
derustensis: from Derust.
Rosette robust, to 18mm φ. Leaves very long, to 450 X 3mm wide, brownish green at base, green above. (A H. blackburniae var. graminifolia foliis bruneo-viridibus validissimis et subtiliter striatis differt).
This variety was discovered by Jan Vlok east of Oudtshoorn in the De Rust area. It differs in that it is far more robust than the species, while the colour of the leaves has a brownish tinge and the leaves tend to be more erect. It occurs on the north upper slopes of conglomerate hills together with Euphorbia enopla. The brownish coloration is more pronounced in the old leaf bases. The seeds are surprisingly large. According to S. Hammer, the leaves of seedlings are characteristically striped. There is a difference in the flowers and the flowering time is also earlier than in the other two varieties. This robust variety also is a vigorous grower in contrast to the other two which are generally slow in cultivation.
Distribution: 3222 (Oudtshoorn): West of Derust (-BC), J. Vlok in Venter 93/24, (NBG).
Haworthia blackburniae var. derustensis JDV93/24 west of De Rust. A more robust variety with brownish leaves, which flowers considerably earlier than var. graminifolia.Haworthia blackburniae var. derustensis JDV93/24. The plants generally grow within other species and can be difficult to see.
c. var. graminifolia (Smith) Bayer (stat. nov.): H. graminifolia Smith, JS.Afr.Bot. 8:247(1942). Bayer :120(1976). Bayer :40(1982). Scott :3(1985). Type: CAPE 3322(Oudtshoorn): Schoemanspoort ( AC), M. Courtenay Latimer in Smith 5222 (NBG).
graminifolia: grass like leaves.
1982 – When first collected, the plants were presumed to be a new species of Aloe and given to G.W. Reynolds. It was only when they flowered that they were seen to be a new Haworthia species. The leaves can be over 300 mm long but are seldom more than 1,5 mm wide. They are more channelled than is the case in H. blackburniae, and the margins are armed with minute white teeth. The leaf bases are amplexicaul and fibrous to papery not at all swollen or bulb like. The roots on the other hand are very thickened and fleshy. The narrow attachment to the stem is very easily broken or damaged and this must be guarded against when moving plants. H. graminifolia is only known in Schoemanspoort, north of Oudtshoorn. A smaller form has been reported on the rocky slopes above the Cango Caves, and collections have also reputedly been made from further west. Although separated from H. blackburniae on the basis of the narrower leaf and pronounced marginal teeth, this distinction may fail as the species becomes better known. H. graminifolia occurs on cooler, higher south slopes and is winter-growing. It is not as proliferous as H. blackburniae although J. Dekenah spoke of huge clumps overhanging the road between Oudtshoorn and the Cango Caves. This has never been confirmed, and H. graminifolia remains very scarce.
1999 – I have no indications that var. graminifolia is any commoner than I have suggested. The very slender leaves and their coloration were all that influenced the decision to maintain it earlier at species level. Leaf channeling is as much a characteristic in the typical variety, as is the presence of marginal leaf spines. Curiously I seem to have confounded the initial response to the collection of this variety by Miss Courtenay-Latimer. Scott I think correctly states that it was in fact H. blackburniae which was first considered to be an Aloe species. I clearly recall Miss Latimer also recounting her reaction to seeing the plant for the first time and her response was also to consider it an Aloe rather than an Haworthia. If she collected it in the absence of flowers, she must have indeed been very observant to have identified it among the many grass-like look-alikes, as being of interest to G.G. Smith. A collection by Peter Bruyns in the Gamkapoort is very similar to the original from Schoemanspoort, having perhaps a less blue coloration to the leaves, which are also slightly broader.
Distribution: 3321 (Ladismith): Hartmansberg (-BC), Bruyns sn. (BOL). 3322 (Oudtshoorn): Schoemanspoort ( AC), M. Courtenay Latimer in Smith 5222 (NBG).
Haworthia blackburniae var. graminifolia JDV97/107 Schoemanspoort. Plants may be solitary or clump-forming.Haworthia blackburniae var. graminifolia JDV97/107 Schoemanspoort. Note the channeled leaf and pronounced white marginal spines.
Rosette from 40-150mm φ, slowly proliferating. Leaves oblong-lanceolate, incurved, not acuminate or truncating, translucent bluish green, with spines on margins and keel >2mm long. Inflorescence robust, simple. Flowers broad and flat across upper basal part.
1982 – The previous concept of H. bolusii was based on plants occurring around Graaff-Reinet. Here the plants are 50-60 mm in diameter and the white bristly leaves are tightly closed over the centre of the plant. The essential difference between this species and H. arachnoidea is the colour. H. bolusii has translucent bluish-green or greyish-green leaves. Further investigation in the field has shown that it is not possible to clearly separate H. bolusii and the previously recognised and larger H. blackbeardiana. Thus H. bolusii is now here considered to be a widely distributed species with a fair degree of variability. It extends from Middelburg to Sterkstroom and southwards to Graaff Reinet and Cradock. The distribution extends further southwestwards in the eastern Karoo to Klipplaat and Jansenville, where very small forms occur in some habitats. It is probable that the small, relatively hairless plants south of Klipplaat are extreme forms of H. bolusii. Similarly there are aberrant, small forms with rather fewer but larger leaves than normal in the extreme northwest near Barclay East. At Middelburg the plants are also very large (up to 150 mm diameter) as they can also be at Cradock. There is no justification for upholding H. batteniae in which the supposed distinguishing characters, viz. spiralled florets and raised leaf venation, are wholly fortuitous. The problem really lies in separating H. cooperi and H. bolusii as discussed under the former species. H. bolusii is technically in the summer rainfall region, but responds to winter growing conditions. Good light is necessary if it is to be grown to handsome natural appearances.
1999 – Col Scott’s perceptions of var. blackbeardiana, H. mucronata, H. cooperi and H. batteniae perhaps reflect the problem in this melange of species. Scott’s H. mucronata seems to include the relatively glabrous elements of three different species. His 1983 discussion of that species includes what is quite obviously the spined H. cooperi var. pilifera. My perception is that H. cooperi is a main element and includes several of Scott’s species. It is in fact difficult to exclude H. bolusii if the var. blackbeardiana is placed with cooperi; unless a distinction is made between elements which tend to have at least partially truncated leaf ends. My dilemma is that blackbeardiana could have been placed as a variety of H. cooperi as there is unquestionably a close alliance. However, this is largely a problem of the nomenclatural system. Had H. cooperi not been described, it would have not been presented here as a species but rather as a variety of a more substantial species. I concluded that larger elements with the more lanceolate leaves and longer spines from the higher altitudes of the Eastern Cape formed a more logical and coherent entity. In the northern Transkei (higher altitude) there appear to be elements with truncated leaves which I would assign to H. cooperi var. obtusata).
A similar problem arises in the wider Port Elizabeth area where a similar transition to H. decipiens occurs. I have experienced difficulty with plants collected east of Pearston but have never had adequate material on which to pass an opinion.
a. var. bolusii. This typical variety is a smallish element in which the spination is dominant. The spines are fine and fully cover the plant so that it appears like a white ball. There is no clear integration with H. semiviva, but near Murraysburg and also at Victoria West the plants do show some tendency for leaf tips to die back as in that species. The collection from Jansenville cited below has understandably been confused with H. arachnoidea var. aranea because of the fine spination. It does have the bluish coloration of bolusii. A short distance away, on dolerite soils as opposed to tillite, the plants are larger and more typical of this species too.
Distribution: 3123 (Richmond): E. Murraysberg (-DD), Bayer 2389; Murraysburg (-DD), Banks (BOL). 3124 (Hanover): W. Graaff- Reinet (-CC), Bayer 2380 (NBG). 3224 (Graaff-Reinet): W. Graaff-Reinet (-BA), Smuts (NBG); Graaff-Reinet (-BA), Bolus 158 (BOL, K), Stayner in KG 232/62, (NBG), Ryder in NBG3152/34 (NBG); Bayer 2022, 2072 (NBG); Adendorp (-BC), Stayner in KG284/62 (NBG); Lootskloof (-DD), Bayer 2071 (NBG). 3225 (Somerset East): NE. Ashbourne (-AC), Bayer & Bruyns 6566 (NBG); NE. Pearston (-CA); Bayer 173 (NBG). 3323 (Willowmore): NE. Fullarton (-BB), Bayer 4160 (NBG); Eensaam (-BB), Marx 56 (NBG).
Haworthia bolusii var. bolusii JDV97/71 south of Graaff-Reinet. A small compact plant with fine spination.
Haworthia bolusii var. bolusii JDV87/186 south-west of Graff-Reinet. As one moves further south and eastwards, the plants seem to lose their compactness and dense spination.
b. var. blackbeardiana (V.Poelln.) Bayer Bayer :103(1976). Bayer :31(1982): H. blackbeardiana V.Poelln., Feddes Repert.Spec.Nov. 31:82(1932). Type: Cape, Bowes Farm near Queenstown, Miss G. Blackbeard. Not preserved. Lectotype (B&M): ex cult. V.Poelln. 1932 (B): H. blackbeardiana var. major V.Poelln. 41:196(1937). Type: Cape, Halesowen, Cradock, R. James in Parks 1483/36. Not preserved. Neotype (designated here): CAPE-3225 (Somerset East): beyond Halesowen station (-BA), Smith 2301 (NBG): H. inermis idem. 31:85(1932): H. altilinea var. inermis idem. 41:194(1937). H. altilinea var. limpida fa inermis idem. 49:29(1940). Type: Cape, Halesowen, Cradock (-BA), Stell. Not preserved. Neotype (designated here): CAPE-3225(Cradock): Halesowen (-BA), Smith 2289 (NBG): H. batteniae Scott, Cact.Succ.J(U.S.). 51:268(1979). Type: CAPE-3225(Cradock):, 35km west of Cradock (-AA), C.L. Scott 5272 (PRE): H. cooperi Bak. pp. Scott ibid 53:70(1981) as to Cradock and Thomas River. Scott :104(1985): H. mucronata Haw. pp. Scott ibid. :80(1986) as to Cradock specimen.
blackbeardiana: in honour of Miss G. Blackbeard.
This variety is generally larger than the typical form and the spination is sparser. However it is more densely spined than is H. cooperi, and the spines are longer, exceeding 2mm. The definition is otherwise geographic and the interface with H. cooperi is from south of Cathcart in a southwesterly direction to at least Jansenville. In the general Rippon Station area there are two adjacent populations, one of which is attributable to this species and the other to H. cooperi. The Patryshoogte/Baviaanskranz collection is also this species growing in the same general area of H. cooperi var. dielsiana. The interpretation in this work of H. pringlei Scott as a variety of H. decipiens may be erroneous as it may perhaps be better placed with var. blackbeardiana. This is also suggested by the Patryshoogte specimens.
There are forms of var. blackbeardiana from the Queenstown area which have very few or sparse short spines, and similarly on the Hogsback mountain. It has been reported that var. blackbeardiana has been collected in the southern Orange Free State and that its distribution extends considerably westward from Middelburg. I have no confirmation of this. There are also reports that it intergrades with H. semiviva in the far west of its range, and the sparse herbarium record suggests that this may be true.
Distribution: 3023 (Britstown): Die Puts (DD), Fuller 193 (BOL). 3024 (Philipstown): Hanover to De Aar (-CD), Stayner in KG307/70 (NBG); 3026 (Aliwal North): Elandshoek (-DA), Smith 6828 (NBG); SE. Aliwal North (-DD), Smith 7471 (NBG). 3123 (Richmond): Richmond (-AC), Stayner in KG591/69 (NBG); Richmond (-BD), Wohlman in KG581/69 (NBG). 3124 (Hanover): Cypherwater (-AD), Bruyns 3010 (NBG); Bethesda Road to Goliadskraal (-BA), Smith 3640 (NBG). 3125(Steynsberg): Thebus (-BC), Smith 3635 (NBG); 8km E. Steynberg (-BD), Smith 3634 (NBG); Tafelberg (-CA), Smith 3828 (NBG), Stayner in KG693/71 (NBG). 3126 (Queenstown): Bowes’ farm (-DD), Britten 160 (BOL), Bayer in KG 312/70 (NBG); Finchams Nek (-DD), Smith 7212 (NBG); Nonesses Nek (-DD), Smith 7216, 7217 (NBG); Longhill (-DD), Bursey in KG393/70 (NBG). 3127 (Molteno): Cofimvaba (-CD), Bruyns 4385 (BOL). 3224 (Graaff Reinet): Kendrew (-DA), Rossouw 183 (NBG). 3225 (Somerset East): 35km west of Cradock (-AA), C.L. Scott 5272 (PRE); Doornberg (-AA), Herre STE6632 (BOL); Denmark (-AB), Bruyns 1769 (NBG); Post Chalmers (-AB), Smith 7444 (NBG); 24km SE. Fish River Stn. (-BA), Smith 5791 (NBG); beyond Halesowen station (-BA), Smith 2301 (NBG); Halesowen (-BA), Smith 2289, 2301, 5340 (NBG), Herre STE6668 (BOL); Cradock (-BA), Smith 2263, 2289 (NBG), Britten (BOL); Swagershoek Pass (-BC), James 474 (BOL), Fourcade 40 (NBG), Bruyns 1627 (NBG); Daggaboer (-BC), James (BOL); Baviaanskranz, Patryshoogte (-DB), Bayer & Bruyns 6561 (NBG); NW. Rippon Stn. (-DD), Bayer & Bruyns 6556 (NBG). 3226 (Fort Beaufort): Huntley Glen (-AC), Smith 7493 (NBG); Austrey (-AD), Branch 370 (NBG); Katberg (-BC), Luckhoff in NBG407/34 (NBG); Waterdown Dam (-BD), Bayer & Bruyns 6569 (NBG); S. Estrelle (-BD), Bayer & Bruyns 6570 (NBG); Hogsback (-DB), Smith 375 (NBG). 3227 (Kingwilliamstown): Imvani (-AA), Bayer in KG311/70 (NBG); 24km N. Cathcart (-AA), Smith 3633 (NBG); N. Waku (-AA), Smith 5718 (NBG); 2km N Goshen (-AA), Branch 20 (NBG); Turnstream (-AB), Bayer & Bruyns 6571 (NBG); New Haven (-AB), Branch 18 (NBG); W. Cathcart (-AC), Smith 5744 (NBG); 16km W. Cathcart (-AC), Smith 38, 358 (NBG); SW. Cathcart (-AC), Barker 3427 (NBG), Bayer in KG310/70, KG393/70 (NBG); Inverbolo (-BC), Bruyns (NBG).
Haworthia bolusii var. blackbeardiana JDV91/102 soth of Cradock. This variety is widespread and the spination is coarser and sparser, or occasionally absent.
Haworthia bolusii var. blackbeardiana JDV91/99 south of Cathcart. Here the plants are often sparsely spined and easily confounded with H. cooperi.
Haworthia bolusii var. blackbeardiana JDV97/62 Aliwal North. The variety is not well-known at its north-eastern limits.
7. Haworthia chloracantha Haw., Revis. :57(1821). Bayer :106(1976). Bayer :32(1982). Scott :52(1985). Aloe chlorocantha Roem. et Schultes, Syst.Veg. 7:641(1929). Salm-Dyck, Monogr. 13:f1(1836). Type: Not preserved. Neotype: icon, 13:f1 Salm-Dyck, Monogr. Epitype (ex B&M, designated here): N. of Herbertsdale, Bayer in KG411/75 (NBG).
chloracantha: green-thorned.
Rosette from 25-40mm φ, proliferous. Leaves: erect spreading, firm to slightly scabrid, triangular in cross-section, spines on margins and keel. Inflorescence simple raceme, lax. Flowers small.
1982 – H. chloracantha is a fairly localised species occurring in the Herbertsdale, Mossel Bay and Great Brak area. It occurs as three main varieties. Firstly the variety chloracantha from north and west of Herbertsdale which is a relatively robust light green form. The var. denticulifera is a smaller, usually purplish-green form found in and around Mossel Bay, while var. subglauca is a more robust waxygreen form from the granitic soils at Great Brak. Von Poellnitz, particularly, confused this species with H. angustifolia but it is slightly more scabrid and the marginal teeth are larger and wider apart. Although more robust, the var. subglauca may reach up to 60-70 mm tall with leaves up to 10 mm broad at their widest. The var. denticulifera may be as small as 30 mm tall with leaves less than 3 mm broad at their widest. The relationship between H. chloracantha and H. floribunda to the west is obscure. The Gouritz River valley effectively divides the two species but there are dubious populations north and south-east of Albertina which may suggest a relationship between these two species (see H. floribunda). However, it is unlikely that either H. floribunda, H. divergens or H. variegata will be confused with H. chloracantha.
1999 – This species was related to H. angustifolia even prior to the recognition of H. monticola ( H. divergens Bayer, 1982). H. chloracantha is more probably more directly related to H. floribunda and H. variegata, but it is also possible that the relationship of the varieties given here is incorrect and that the typical variety has weaker links with the southern Cape than the other two varieties. The new H. monticola var. asema from Calitzdorp, as well as the discovery of H. outeniquensis, also need to be taken into consideration.
It appears that the dark-green erect plants from southeast and east of Albertinia (Cooper Siding) should be regarded as H. chloracantha and not as H. floribunda. The population at Draaihoek to the north, includes plants which resemble H. parksiana and it may best be related to H. floribunda. Plants with erect leaves growing with H. parksiana at Groot Brak have been regarded as H. floribunda and it is more probable that they are in fact also H. chloracantha. It seems improbable that three similar species, in which the affinities with one another are in question, can co-occur.
a. var.chloracantha. The typical variety is taken to be the very proliferous green forms around Herbertsdale and along the Gouritz River as it passes through the Langeberg mountains. While Breuer and Metzing do not use the Salm Dyck illustration to typify this name, that illustration and the icon in Bergers revision in Fas Pflanzenreich, are really the main sources for the application of the name.
Haworthia chloracantha var. chloracantha JDV87/80 Hertertsdale. In cultivation, this variety often appears pale and chlorotic.
Haworthia chloracantha var. chloracantha JDV97/138 Herbertsdale. The typical variety occurs only in this area and is always a plainer and yellower green.
b. var. denticulifera (V.Poelln.) Bayer :112(1976). Bayer :32(1982). H. angustifolia var. denticulifera V.Poelln., Feddes Repert.Spec.Nov. 41:194(1937), ibid. 44:228(1938). Type: Cape, Montagu, Mrs Helm. Not preserved. Lectotype (B&M): icon (B).: H. angustifolia var. lilliputana Uitew. Sukkulenta 43(1953). Type: Not preserved. Neotype (designated here): CAPE-3422 (Mossel Bay): Hill above Mossel Bay (-AA), Courtenay-Latimer in Smith 5223 (NBG).
denticulifera: bearing small teeth.
This variety is dark-green and occurs around Little Brak and Mossel Bay. It also occurs along the lower Gouritz Valley where it has been previously assigned to H. floribunda. Von Poelnitz (1938) cited a number of improbable localities for this element as a variety of H. angustifolia. These included Montagu, Calitzdorp, Riversdale and Great Brak. It is generally concluded that the plants originated from the greater Great Brak area. Uitewaal’s smaller plants – var. lilliputana – are comparable with the population which occurs within the town of Mossel Bay itself. The specimen cited from the Duiwenhoeks causeway is also one with erect pointed leaves which draws the relation with H. floribunda into the discussion.
Distribution: 3420 (Bredasdorp): Duiwenhoks Causeway (-BD), Muller-Doblies 82/098 (NBG). 3421 (Riversdale): Cooper Siding (-BB), Bayer 3586 (NBG); 3km N Gouritzmond (-BD), Smith 7519 (NBG), Bayer 3586 (NBG). 3422 (Mossel Bay): 2km N. Mossel Bay (-AA), Smith 2896 (NBG); 1km N. Mossel Bay (-AA), Smith 3958 (NBG); Mossel Bay (-AA), Smith 5223, 5750 (NBG); Little Brak (-AA), Smith 5751 (NBG); Great Brak (-AA), Bouwer (NBG).
Inadequately located: ex hort. Armstrong in Smith 2830 (NBG).
Haworthia chloracantha var. denticulifera JDV88/17 Hartenbos. Usually a dark green plant with fine spination.
Haworthia chloracantha var. subglauca JDV87/79 west of Great Brak. Sometimes almost bicoloured with reddish tips above the dark green leaves.
c. var. subglauca V.Poelln., Kakteenkunde 9:135(1937). Bayer :106(1976). Bayer:32(1982). Type: Cape, Great Brak, Mrs Helm. Not preserved. Neotype (B&M): Great Brak, Hurling & Neil (BOL).
subglauca: nearly glaucous.
As stated above, this variety differs in coloration from the other two varieties and in the larger sparser spines. There is a specimen cited rather vaguely as from near Zebra which could perhaps be H. outeniquensis.
Distribution: 3422 (Mossel Bay): Great Brak (-AA), Fourcade 18 (NBG), Hurling & Neil (BOL), Ferguson 1 (BOL), Smith 2889 (NBG); E. Great Brak (-AA), Smith 2885, 3957 (NBG); Bayer in KG(98/71 (NBG); George (-AA), Malherbe in NBG304/40 (NBG), Fourcade in NBG2617/34 (NBG).
Inadequately located: ex hort, Pillans (BOL). Heyn’s farm near Zebra, van der Bijl 474 (BOL).
Haworthia chloracantha var. subglauca JDV84/58 south of Great Brak. Larger leaves which are a waxy bluish colour.
Haworthia chloracantha var. subglauca JDV94/3 north-east of Brandwacht. This population seems to be intermediate between vars. subglauca and denticulifera. Very dark coloured.
Haworthia chloricantha var. subglauca JDV84/58 south of Great Brak. Plants can be very withdrawn into the soil.
8. Haworthia cooperi Baker, Saund.Ref.Bot. 4:t.233(1871). Bayer :109(1976). Bayer :33(1982). pp. Scott :103(1985) as to Adelaide. H. arachnoidea (Haw.) Duv. sensu Pole-Evans, Flow.Pl.S.Afr. 7:t.248(1927). Type (B&M): Cape, Cooper (K): H. vittata Baker, loc.cit. :t.263. Type: Not preserved. Lectotype: icon t.234, Saund.Ref.Bot. cooperi: for Thomas Cooper.
Rosette to 120mm φ, often proliferous, stemless. Leaves 20-40, fleshy, swollen, oblong-lanceolate, quickly tapering, acuminate or truncating, marginal spines <2mm long if present. Bluish-green in colour, slightly translucent, with veins usually reddening and leaves developing purplish hues in exposed situations. Inflorescence compact, firm peduncle with many closely arranged flowers, to 20cm long. Flowers 20-30, perianth white.
1982 – This species is beautifully illustrated in Refugium Botanicum and two other species, H. pilifera and H. vittata, were also described and illustrated here. The name H. pilifera has been most commonly used for plants with relatively blunt leaves which occur in the Eastern Cape particularly around King Williamstown. The name H. vittata was similarly applied to the longer-leaved forms in the Thomas River and Cathcart areas which probably intergrade with H. bolusii. However, the name H. cooperi can equally be applied and has page preference over the other two names. H. cooperi is very common widespread and variable as the synonymy suggests. It is logical to suppose that in this case Haworth must have received specimens at some time or another. Uitewaal attempted to apply the name H. obtusa Haw. which has since been refuted by Bayer and Pilbeam (1974). Scott similarly implemented H. altilinea Haw. but uses the name in a restricted sense to exclude all three of Baker’s names, as well as names such as H. limpida and H. mucronata. The writer’s contention is that the name H. altilinea is a source of confusion and should for the present at least, be rejected. There are thus the four species with translucent bluish-, or greyish-green leaves as indicated in the key. H. cooperi is distinguished by its generally less hairy or shortly haired leaves. The leaves are obtuse to obtuse-ovate although forms with longer, more acuminate leaves also occur. These usually have thicker leaves than corresponding forms in H. bolusii and the leaves are also less hairy. Scott comments on the withdrawal of plants into the ground, and the question of exposure and soil substrate are responsible for much of the variability in the species. In ‘stayneri’ collected at Bethelsdorp near Port Elizabeth, and in several other populations, the leaf veins actually become necrotic and the leaf-ends truncated. In ‘gordoniana’ from Hankey this does not occur.
H. cooperi is a species of the dry grassveld areas of the Eastern Cape and occurs in the high-lying sourveld of the Zuurberg to the dry Valley Bushveld of the Fish River. Size varies enormously from small glabrous plants less than 40 mm in diameter southwest of Grahamstown, to enormous hairy specimens over 100 mm in diameter at Keiskamma. The variety leightonii is exceptional in that it occurs on the edges of exposed granite slabs west of East London. This variety is very proliferous and is characterised by the persistence of reddish coloration in the leaf veins.
1999 – One of the difficulties with this species is in its relationship with H. bolusii var. blackbeardiana, and here the typical variety is applied to that intermediate element in the Thomas River area. This does not represent the main body of the species at all, which is a product of the nomenclatural system. Also the previous Handbooks do not present any idea of the variation in this species. There is an additional problem in the association with H. cymbiformis. Uitewaal really exposed the problem with his recognition of the name ‘obtusa’ of Haworth for this species. Bayer and Pilbeam refuted this by suggesting that Uitewaal had misinterpreted Haworth’s description and the illustration associated with it. Scott similarly refuted Uitewaal and also applied the name as a variety of H. cymbiformis. However, he illustrated a blue-green variant which we regard as H. cooperi var. obtusata, a new name, in this work. There is clearly a problem in finding a point of origin for ‘obtusa’ as variant of H. cymbiformis rather than of H. cooperi, and that is answered under the former species. However, herbarium records indicate a problem in the broader Alicedale/Adelaide areas where truncation and abbreviation of the leaves of both species seems to be evident, together with the problem of green versus blue coloration. The essential difference between the two species – which generally have the same distribution range and very often co-occur – is that H. cooperi is an open ground species, whereas H. cymbiformis occurs on rocky shelves and cliffs. It is interesting that even a variety like leightonii has a tendency to resemble H. cymbiformis, by acquiring an opaque yellowish-green coloration as opposed to the required blue-green for the species cooperi. This is a common problem in trying to absolutely circumscribe the species. But even outside of that, there has been a massive problem in the plethora of names attached to either species. Both extend into the mountains north and west of Port Elizabeth and it is not clear just how this relates to other species which do the same. The following varieties are recognised.
a. var. cooperi. The typical variety occurs in the Thomas River area and is along the transition zone to H. bolusii var. blackbeardiana. The distinction is that the leaves are slightly more acuminate and the venation tends to acquire reddish tints, while general coloration tends to a purplish hue. The leaf spines are also generally less than 2mm long.
Distribution: 3225 (Somerset East): 30km S. Cradock (-BD), Smith 5194 (NBG); Bruintjieshoogte (-CB), Britten in PRE 34923, Bayer 2024 (NBG); 11km S. Somerset East (-DC), Smith 2841 (NBG). 3226 (Fort Beaufort): Bobbejaanrivier, Bedford (-CA), Smith 2244 (NBG); Koonap Bridge (-CD), Bayer & Bruyns 6563 (NBG); Elandskop, Adelaide (-CD), Smith 2687, 2797, 2799 (NBG); Adelaide (-CD), Krynauw in NBG270/43; Katberg (-DA), Herre in STE6607 (BOL), Read in BOL71291, Smith 2773 (NBG); Warfield (-DB), Venter 91/82 (NBG); Woburn (-DB), Smith 578 (NBG); Alice, Stewart Memorial (-DD), Smith 5211 (NBG). 3227 (Kingwilliamstown): Thorn River near Cathcart (-AC), Acocks 11003; W. Cathcart (-AC), Bayer in KG 392/70 (NBG); Thomas River (-AD), Smith 3631 (NBG), Scott 1720 (NBG), Stayner in KG402/61 (NBG); Tsomo (-BB), Branch 13 (NBG).
Inadequately located: ex hort. Stanford (BOL).
Haworthia cooperi var. cooperi JDV91/26 Koonap River. On a south-facing rocky slope.
Haworthia cooperi var. cooperi JDV91/98 Thomas River. In the Cathcart area there are also plants with less of the purplish colouration and which may also be without spines – confused with H. bolusii var. blackbeardiana.
Haworthia cooperi var. cooperi JDV91/26 Koonap River. Note the darker lines and purplish tints.
Haworthia cooperi var. cooperi JDV91/14 De Plaat, Kirkwood. Here there may be an interaction with H. gracilis.
b. var. dielsiana (V.Poelln.) Bayer comb. nov. H. dielsiana V.Poelln., Feddes Repert.Spec.Nov. 28:103(1930). H. pilifera var. dielsiana idem. 49:27(1940). H. obtusa var. dielsiana (V.Poelln.) Uitew., Succ. 29:50(1948). Type: Cape, Sheldon, H.H. Hutton 489. Not preserved. Neotype (designated here): CAPE-3325(Somerset East): Sheldon (-BB), A.J. van der Merwe in Smith 1140 (NBG). H. joeyae Scott, Bradleya 13:80(1995). Type: CAPE-3225( Somerset East): a few kilometers NE. of Cookhouse (-BD), Scott (GRA).
dielsiana: in honour of Prof. Diels of Berlin, Dahlem.
There appear to be two elements within the species with very truncated leaf ends. The first is this variety from the western area in which the leaves are variously truncated and obtuse. Smith records a wide range of forms with or without leaf spines and some with very rounded leaf-ends. Sometimes the leaf-margins are almost ridge-like. The leaf-tips do not appear to become necrotic as happens in the var. pilifera. The leaf awn is virtually absent and the leaves tend to have marked venation. The keel and margins tend to be rounded.
Distribution: 3225 (Somerset East): Sheldon (-BB), Smith 1140 (NBG), Hutton in Herre 489 (BOL); 5km E. Somerset East (-DA), Bayer & Bruyns 6565 (NBG); Glen Avon (-DA), Smith 5790 (NBG); Merantes Kloof (-DB), Smith 3492 (NBG); Eastpoort (-DB), Smith 3491, 3493 (NBG), Reynolds (BOL), Bayer & Bruyns 6558, 6559, 6560 (NBG); Little Fish (-DC), Smith 5550 (NBG). 3226 (Fort Beaufort): 10km N. Adelaide (-CB) Krynauw in NBG267/43, Krynauw in NBG687/41; 19km NE. Adelaide (-CB), Krynauw in NBG269/43 (NBG); Chancery Hall (-CB), Bayer & Bruyns 6564 (NBG); Kagasmond (-CC), Krynauw in NBG273/43; Paardefontein, SE. Adelaide (-CD); Brakfontein, Kroomie (-CD), Smith 5112 (NBG); Koonap (-CD), Smith 7131 (NBG); Adelaide (-CD), Smith 2687 (NBG); 30km S. Adelaide (-CD), Smith 2240 (NBG); Somerset East (-DA), Fouche in PRE 34919; Woburn; Tyumie river (-DB), Acocks 13575 (PRE); W. Fort Beaufort (-DC), Bayer & Bruyns 6592 (NBG); S. Fort Beaufort (-DC), Bayer & Bruyns 6591 (NBG); E. Fort Beaufort (-DC), Smith 3489 (NBG); 16km N. Cradock road (-DC), Britten in PRE 34918.
Inadequately located: Ross’ Mission, Smith 7492 (NBG); Zaysdorp NBG1019/25; ex hort Whitehill (NBG), Ross-Frames (NBG), Stanford (BOL).
Haworthia cooperi var. dielsiana JDV91/101 Eastpoort. This photograph is of a plant from south of Eastpoort, where they are slightly greener and the leaves less truncated than in the type of ‘H. joeyae Scott’.
Haworthia cooperi var. dielsiana JDV91/101 Eastpoort. This is a plant with very truncated leaves with virtually no end-awn.
c. var. gordoniana (V.Poelln.) Bayer comb. nov. pp H. cooperi (Bak.) Bayer :119(1972). pp. Bayer :33(1982). H. gordoniana V.Poelln., Feddes Repert.Spec.Nov. 42:269(1937). H. pilifera var. gordoniana V.Poelln. idem. 44:237(1938). H. obtusa var. gordoniana (V.Poelln.) Uitew., Succulenta 29:50(1948). Type: Zuurbron, Hankey, Long 814. Not preserved. Neotype (designated here): CAPE-3324 (Steytlerville): Patensie (-DD), Smith 3028 (NBG).
gordoniana: in honour of Gordon King.
There are many variations of small glabrous and spined plants in the Hankey/Patensie area and it is rather problematic to decide to which species they really belong. Von Poellnitz associated his species with H. cooperi as recognised in this work and it is thus the small blue-green element which is nearly identical to his H. stayneri. Unlike the latter which is made synonymous with H. cooperi var. pilifera the leaf tips do not truncate on exposure to direct sun. It differs from similar plants which are regarded as H. gracilis, in that the outer leaves stay incurved and are sparsely spined. Also the leaves are usually more thickly turgid than in that species. The plants are less proliferous and tend to be withdrawn into the soil.
Distribution: 3323 (Willowmore): Redcliffe (-BA), Bruyns 7062 (BOL); Uniondale Poort (-CA), Bayer 4404 (NBG); Damse Drif (-CA), Bruyns 1651, 1652 (NBG); Nuwekloof (-CA), Bruyns 1840 (NBG); Saptou (-CB), Bruyns 7079 (BOL); Studtjes (-DB), Bruyns 2190 (NBG). 3324 (Steytlerville): Enkeldoorn (-CB), Perry 1427 (NBG), Bean (NBG); Holgat Kloof (-CC), van Jaarsveld 6885 (NBG); Moordenaarskloof (-CD), Stayner in KG673/71 (NBG); Grootriver Poort (-DA), Bruyns 2202 (NBG); Quagga to Cambria (-DA), Smith 2912 (NBG); Cambria (-DA), Smith 2904 (NBG); Ouplaas (-DB), Bruyns 7043 (BOL); 2km E. Hankey (-DD), Rossouw 141 (NBG); Patensie (-DD), Smith 3025 (NBG); E. Patensie (-DD), Smith 2908 (NBG); S. Hankey (-DD), Smith 3186 (NBG); Hankey (-DD), Smith 2905 (NBG); 5km N. Hankey (-DD), Bayer in KG193/73 (NBG); N. Hankey (-DD), Smith 2883, 3687 (NBG); NE. Hankey (-DD), Bayer 4474 (NBG), Stayner in KG180/71 (NBG); E. Hankey (-DD), Smith 2597, 2893, 2980 (NBG); Stayner in KG180/71 (NBG); 7km N. Zuurbron (-DD), Smith 3671 (NBG); 5km N. Zuurbron (-DD), Bayer & Bruyns 6553 (NBG); 8km N. Zuurbron (-DD), Bayer & Bruyns 6554 (NBG); N. Joubertina (-DD), Venter 91/64 (NBG). 3325DC Coega Kop(-DC), Long 1132 (PRE). 3424 (Humansdorp): Jeffrey’s Bay (-BB), Bayer & Venter 6597 (NBG).
Inadequately located: Herre STE6609 (BOL).
Haworthia cooperi var gordoniana JDV91/8 Baviaanskloof. A smaller plant with non-truncating leaf-tips and often confused with H. gracilis.
Haworthia cooperi var gordoniana JDV94/76 north of Humansdorp. The bluish-green colour is usually a distinguishing character, and the rosettes are fairly globose.
Haworthia cooperi var gordoniana JDV96/92 Jeffrey’s Bay. Here the plants in the field are brownish and may be confused with H. gracilis.
d. var. leightonii (Smith) Bayer :128(1976). Bayer :34(1982). H. leightonii Smith, JS.Afr.Bot. 16:10(1950). Scott :107(1985). pp. H. cooperi Bak., Scott, Cact.Succ.J(U.S.) 53:70(1981). Type: CAPE-3327 (Peddie): Kayser’s Beach (-BA), Smith 6938 (NBG).
leightonii: for I. Leighton.
There are several populations of this variety and the circumscription is enlarged to include those from further to the northwest in which the leaves are also more lanceolate and untruncated. It is almost certainly an ecotype associated with the granitic slabs of the coastal area near Kayser’s Beach, where the plants are very proliferous indeed. This, and the strong purplish coloration, characterise the variety.
Distribution: 3327 (Peddie): Kaysers Beach (-BA), I.H. Leighton in NBG 68362, Smith 6938 (NBG), Bayer in KG6/72 (NBG), Venter 91/105 (NBG); SW. Paynes Hill (-BA), Smith 514 (NBG); Bayer 1621 (NBG).
Inadequately located: Kingwilliamstown, Taylor 3039 (NBG), Leighton in NBG662/34.
Haworthia cooperi var. leightonii JDV91/105 Kyser’s Beach. It is also very proliferous.
Haworthia cooperi var. leightonii JDV91/97 east of Chalumna. This variety is characterised by the strong reddish-pink colour which develops under stress, but here the plants are greener.
Haworthia cooperi var. leightonii JDV91/104 Kayser’s Beach. Associated with granite slabs.
Haworthia cooperi var. leightonii JDV91/105 Kayser’s Beach. A population in the area has plants which bear some similarity to H. cymbiformis.
e. var. pilifera (Baker) M.B.Bayer comb.nov. H. pilifera Baker, Saund.Ref.Bot. 4:t.234(1871). V.Poelln., Feddes Repert.Spec.Nov. 44:236(1938). Scott :104(1985). H. obtusa var. pilifera (Baker) Uitew., Succulenta 29:50(1948). Type: Not preserved. Lectotype (here designated): icon, :t.234, Saund.Ref.Bot.: H. stayneri V.Poelln. loc.cit. 42:270(1937). H. pilifera var. stayneri idem. 44:237(1938). H. obtusa var. stayneri (V.Poelln.) Uitew. loc.cit. Type: 14m Port Elizabeth to Uitenhage, F.J.Stayner. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): 13.8m Port Elizabeth to Uitenhage (-DD), F.J.Stayner in KG 2/70 (NBG): H. stayneri var. salina V.Poelln., Feddes Repert.Spec.Nov. 42:271(1937). H. pilifera var. salina idem. 44:237(1938). H. obtusa var. salina (V.Poelln.)Uitew. loc.cit. Type: Cape, Bethelsdorp Salt Pan, Mrs King. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): Bethelsdorp salt pan (-DD), Smith 5817 (NBG): H. pilifera var. dielsiana fa acuminata V.Poelln. loc.cit. 49:27(1940). H. obtusa var. dielsiana fa acuminata (V.Poelln.) Uitew. loc.cit. Type: 4m from Kingwilliamstown, F.A.Fouche. Not preserved. Neotype (designated here): CAPE-3227(Kingwilliamstown): 5km Kingwilliamstown to Pirie Dam (-CC), Smith 3913 (NBG): H. altilinea Haw. sensu Scott, Natn.Cact.Succ.J 34:53(1979). Scott :84(1985).
pilifera: with hairs.
This variety constitutes the main body of the species which is centred about Kingwilliamstown and Grahamstown. It is characterised by the obtuse acuminate end-area with a pronounced point to the leaf. The margins and keel are sharply angled. It is very closely drawn into the soil surface and, on exposure to direct sunlight truncates to form a necrotic end-area.
Distribution: 3128 (Umtata): Viedgesville (-DA), Rush in KG87/80 (NBG). 3225(Somerset East): Little Fish (-DA), Smith 5551 (NBG); Cookhouse, Patryshoogte (-DD), Long 1462 (PRE). 3226 (Fort Beaufort): 24km S. Bedford (-AC), Bursey in KG335/70 (NBG); Good Hope, Alice (-DD), Smith 5496 (NBG); Fort Hare, Cressey in NBG2132/26 BOL); Alice (-DD), Smith 5210 (NBG); Middledrift (-DD), Smith 5342 (NBG). 3227 (Kingwilliamstown): Peddie (-AA), Smith 5654 (NBG); Fort Murray Bridge (-BD), Bayer (NBG); E. Kieskammahoek (-CA), Smith 6065 (NBG); Kieskammahoek (-CA), Smith 5309, 7350 (NBG); Stayner (NBG); 5km Kingwilliamstown to Pirie Dam (-CC), Smith 3110, 3913 (NBG): (-CC), Dyer 2079 (PRE); Golf course (-CD), Scott 1980 (PRE), Crampton in NBG1096/28 (NBG), Taylor in NBG250/34 (NBG); Line Drift (-CD), Smith 5434 (NBG); Balazi (-CD), Smith 3574 (NBG); St Johns Drift (-DA), Bayer 1623 (NBG); Near Komgha (-DB), Flanagan 1116 (BOL, PRE); Brigadoon (-DC), Bayer 4460 (NBG); Buffalo River (-DC), Smith in NBG343/35 (NBG); Bridal Drift (-DC), Smith 386 (NBG); Cambridge (-DD), Grenfell in NBG872/35. 3325 (Port Elizabeth): Cookhouse to Zuurberg (-BA), Smith 3494 (NBG); Shenfield (-BB), Blackburn in BOL71305; Kommadagga (-BB), Bruyns 1651 (NBG); 18km N. Zuurberg Inn (-BC), Stayner in PRE 57676; Enon (-BC), Dyer 498a (PRE); Zuurberg (-BC), Stayner in KG685/71 (NBG); Addo Park (-BD), Branch 31 (NBG); Bauerskraal (-CB), Bayer & Venter 6559 (NBG); Despatch (-CD), Muir 13133 (PRE). 1km W. Uitenhage (-CD), Smith 5818 (NBG), Britten (BOL); 22km Port Elizabeth to Uitenhage (-DC), F.J.Stayner in KG 2/70 (NBG); E. Uitenhage (-DC), Smith 3582 (NBG); Bethelsdorp salt pan (-DC), Smith 5817 (NBG), Stayner in KG2/70; Redhouse (-DC), Paterson 431 (BOL). 3326(Grahamstown): E. Riebeek East (-AA), Smith 5219 (NBG); Willowfountain (-AA), Bayer 1622 (NBG); Blaauwkrantz (-AC), Dyer 2306 (PRE); NW. Salem (-AD), Bayer 4450 (NBG); 8km N. Grahamstown (-AD), Smith 5628 (NBG), Britten in BOL71290; The Fort (-BA), Smith 5065 (NBG); Keiskamma near Breakfast Vlei (-BB), Acocks 11871 (PRE); Debe Nek (-BB), Smith 3545 (NBG); N. Committees (-BB), Smith 5435 (NBG); E. Committees (-BB), Smith 5430 (NBG); Committees (-BB), Smith 2428, 5352, 5353, 5366 (NBG), 16km N. Cradock road (-BC), Marloth 12607 (PRE); Grahamstown (-BC), Whitmore in NBG1238/24 (NBG), Erens in PRE 34929; Bothas Hill (-BC), Smith 5355 (NBG); Frazers Camp (-BD), Smith 5215, 7413 (NBG); Peninsula (-DA), Bayer in KG383/70 (NBG); Brigadoon (-DC), Bayer in KG382/70 (NBG). 3327(East London): S. Kingwilliamstown (-AB), Krynauw in NBG684/41 (NBG); E. Chalumna (-BA), Smith 5773 (NBG).
Inadequately located: Cape ex hort, Marloth 5861 (PRE); Grahamstown, Dyer 1 (BOL), Britten in BOL71289; Leighton in NBG662/34, in BOL71301 (BOL); Kingwilliamstown, Sim 1030 (BOL), Herre in STE6619 (BOL); Mquakwebe, Leighton in BOL20734-6, in Smith 6938 (BOL).
Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. Not very proliferous and drawn into the soil where they are cryptic when it is dry.
Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. Growing very near a population of H. gracilis, and the difference is in the way this species has shorter stumpier leaves.
Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. The stumpy leaf-ends may be attractively veined and may become almost necrotic to exclude light.
e. var. truncata (Jacobs.) M.B.Bayer comb. nov. H,. obtusa var. truncata Jacobs., Nat. Catt. Succ. J. 10: 81 (1955). Type: Not preserved. Neotype: (B&M in ms.): icon in Jacobsen, Handbuch der Sukk. Pfl. 724, f644 (1956). Epitype: CAPE-3227 (Stutterheim): Runlets, Mgwali (-DA), Smith 5295 (NBG).
truncata: with obtuse leaves.
Rosette proliferous, to 70mm φ, Leaves 20-25, 20-25 x 8mm wide, pale blue-green, erect, truncated, translucent and lightly veined above. (A var. cooperi foliis parvioribus truncatis differt).
I am grateful to I. Breuer for drawing attention to the possible correct application of this name. The important arbiter would actually be colour, which Jacobsen does not stipulate. However, Breier suggests the plants from the Bolo Reserve distributed under I.S.I. No. 1762 represent Jacobsen’s variety; otherwise I would have incorporated it with H. cymbiformis var. obtusa. This is a smaller variety than the western var. dielsiana with similarly truncated leaves, and the distinction is largely on geographical grounds. Other differences are the smaller size, the rapid off-setting, and the almost unlined leaves of the var. truncata as opposed to var. dielsiana. The var. truncata is at the northeast of the distribution range of the species where it extends into the Transkei. Both Bayer and Pilbeam, and Scott have attempted to refute Uitewaal’s interpretation of H. obtusa Haw. as an earlier synonym for H. cooperi. This is a case in which the option is entirely an open one. How and why Uitewaal chose to depart from the general application of the name seems to be the norm for Haworthia.
Distribution: 3227 (Kingwilliamstown): Hunts Drift (-AC), Smith 5175 (NBG); Inerbolo (-BC), Bruyns (NBG); Mgwali (-BC), Smith 5193 (NBG); Runlets, Mgwali (-DA), Smith 5295 (NBG).
Haworthia cooperi var. truncata MBB 386/70 Mgwali, Sutterheim. Grows in a level rocky but grassy area, and forms large blue-green clumps.
g. var. venusta (Scott) Bayer comb.nov. H. venusta Scott, Bradleya 14:87(1996). Type: CAPE-3226 (Grahamstown): NE. Alexandria (-DA), Britten 781 (GRA).
venusta: charming, and for Miss Grace Britten. Her excellent herbarium sheet led to the recent rediscovery by Gerhard Marx.
This very handsome variety is, surprisingly, coarse white-haired, exaggerating the slightly hairy tendency foundin the other varieties. It offsets sparsely if at all, and remains quite small.
Distribution: 3326 (Grahamstown): NE. Alexandria (-DA), Britten 781 (GHS, NBG), Britten and Archibald 781 (BOL, PRE).
Haworthia cooperi var. venusta JDV94/5 south of Grahamstown. A really remarkable and dramatic variety on account of the short shiny hairs.
Haworthia cooperi var. venusta JDV94/5 south of Grahamstown. Known only at one locality. It seems to set seed easily and should become a favorite in collections.
9. Haworthia cymbiformis (Haw.) Duv., Pl.Succ.Hort.Alenc. :7(1809). Bayer :110(1976). Bayer :35(1982). Scott :91(1985). H. concava Haw. Revis. :58(1821). Aloe cymbiformis Haw., Trans.Linn.Soc. 7:8(1804). Sims, Bot.Mag. 1:21,t.802(1805). Salm-Dyck, Monogr. 11:t.1(1840). Type: Not preserved. Neotype (designated here): icon, t.802, Bot.Mag.. Epitype (ex B&M): Walmer, Port Elizabeth, Smith 2844 (NBG): H. planifolia Haw., Phil.Mag. 44:282(1825). H. cymbiformis var. planifolia (Haw.) Baker JLinn.Soc. 18:209(1880). Aloe planifolia (Haw.) Salm-Dyck, Monogr. 11:t.2(1840). Type: icon. t2, Salm-Dyck, Monogr. 1840: H. cymbiformis var. angustata V.Poelln., Feddes Repert.Spec.Nov. 45:166(1938). Type: Discovery region unknown, T.Foster. Not preserved: H. cymbiformis var. angustata fa subarmata idem. 45:166(1938). Type: Rocklands, Adelaide, W.E.Armstrong in Triebn. 1187. Not preserved. Neotype (designated here): CAPE-3226 (Fort Beaufort): Rocklands, W.E.Armstrong in Smith 2801 (NBG): H. cymbiformis var. compacta Triebn. idem. Type: Cape, west of Peddie, Mrs G. McLaren in Triebn. 1148. Not preserved. Neotype (designated here): CAPE-3327 (Peddie): W Woolridge, Peddie (-AB), Bayer 4648 (NBG). H. planifolia var. exulata V.Poelln., Feddes Repert.Spec.Nov. 43:93(1938). idem. 45:162(1938). Type: Cape, Ubi?, C.H.Woolley in Long 392. Not preserved. Neotype (designated here): Photogr. H.G.Fourcade of Long 392 (NBG): H. planifolia var. planifolia fa agavoides Triebn. et V.Poelln., Feddes Repert.Spec.Nov. 45:162(1938). Type: Cape, Fort Beaufort, W.E.Armstrong in Triebn. 1169. Not preserved. Neotype (designated here): CAPE-3226 (Fort Beaufort): Sulphur Baths (-DC), Bayer 4655 (NBG): et fa alta ibid. Type: Cape, Grahamstown, Mrs Helm in Triebn. 851. Not preserved: et fa olivacea ibid. Type: Cape, Quagga West, Mrs Helm in Triebn. 853. Not preserved: et fa robusta ibid. Type: Cape, Baakens Valley, Mrs I.King 100 in Triebn. 1066. Not preserved. Neotype (designated here): CAPE-3325(Port Elizabeth): Baakens Valley (-DC), Smith 3894 (NBG): et var. incrassata V.Poell. idem. 45:163(1938). Type: Cape, Kowie River, Mrs Archibald 335 in Long 446. Not preserved: et var. sublaevis V.Poelln., Kakteenk. 6:67(1938). idem., Feddes Repert.Spec.Nov. 45:163(1938). Type: Albany district, Mrs Britten in Triebn. 940. Not preserved: et var. longifolia Triebn. et V.Poelln. idem. Type: Cape, Grahamstown, Mrs Helm in Triebn. 864. Not preserved: et var. longifolia fa calochlora ibid. Type: Cape, Port Elizabeth, Mrs Helm in Triebn. 941. Not preserved: H. planifolia var. poellnitziana Res. ibid. 48:133(1940). Type: ex Hort., Hamburg. Not preserved: H. lepida Smith JS.Afr.Bot. 10:21(1944). Type: Cape, Albany district, between Carlisle Bridge and Fort Brown, Smith 5066 (NBG).
cymbiformis: boat-shaped.
Rosette to 130mm φ, partially stemmed, proliferous. Leaves broad ovate to lanceolate, flat to slightly concave, generally <1/3 as thick as wide, usually opaque, green turning yellowish to pink hued on exposure. Inflorescence to 250mm, 10-15 flowers, lax. Flowers white.
1982 – This name has been used interchangeably with H. planifolia for a very widespread and variable species growing in the Eastern Cape. This species has practically the same distribution range as H. cooperi but unlike that species is an opaque, truer green colour. It forms dense mats with a fairly superficial root system and grows on rocky slopes and krantzes along rivers and streams. H. planifolia Haw. was only described in 1825 and was stated to be less proliferous and with distinctly flatter leaves. Baker placed the two species together and a study of Von Poellnitz’s and Smith’s records plainly show that neither could clearly distinguish these two species. Von Poellnitz in fact described H. planifolia var. transiens and H. cymbiformis var. translucens from the same locality, and even stated that he did not know how to clearly separate the two species. Von Poellnitz in Feddes Repert. Spec. Nov. 41:199(1937) records H. cymbiformis from Graaff Reinett (Triebner 861) but in 1938 (ibid. 45:163) describes H. planifolia var. incrassata from the same collection. Berger, in a masterly piece of irrationality, justified separate sections for the two species. Smith transferred H. incurvula V.Poelln. from the section Muticae to the section Obtusatae, on the basis of observed continuity with H. cymbiformis. In Flowering Plants of Southern Africa (pl. 356, 1929) a plant identical to H. incurvula is illustrated under the name H. cymbiformis var. planifolia (Haw.) Baker. It can only be concluded that one name be retained for the entire complex and this is H. cymbiformis.
Uitewaal (1948) put forward the view that H. obtusa was not related to H. cymbiformis at all, but that is was an earlier name for H. pilifera (here a synonym of H. cooperi). He based his observation on a colour plate in the Kew herbarium, and goes as far as to say that Haworth’s original description is faulty. The strongest argument in favour of Uitewaal’s contention is the historical one. None of Haworth’s species can be referred to Baker’s H. cooperi and this is hard to concede knowing how widespread and variable that species is. The name H. obtusa is here considered to be probably synonymous with H. cymbiformis and should probably be rejected as superfluous and as a source of confusion. The distribution range extends from the Bashee River in Transkei, to Prince Alfred’s Pass in the west, and northwards to Fort Beaufort. Variation within individual populations is small but no two populations are quite the same. Thus there is scope for a good many more varieties than previously recognised. However, to stay within manageable limits, only the one really different form viz. fa ramosa with the elongated stem, and varieties not limited to single populations, are upheld. The var. incurvula is apparently restricted to Plutosvale which is a contradiction, but, because Smith discusses continuity with H. cymbiformis (atypical for Smith and not borne out by his field records), because of the possibility of a relationship with H. translucens subsp. tenera, and because of similar forms occurring in the Humansdorp area, it is maintained. The var. transiens is really only more translucent, and at the type locality also larger, than incurvula, otherwise they may have been considered synonymous. The var. umbraticola is a distinctive variety from the Swartwaterpoort west of Alicedale and northeastwards to Fort Beaufort. The leaves are very obtuse and round in cross‑section. The fenestrate blunt tips with shining pellucid areas separated by dark green lines, make it a most attractive variety. H. cymbiformis occurs primarily in the summer rainfall area and is very easy to grow in cultivation. It proliferates to an rapidly and should be exposed to at least some direct sunlight to prevent bloating and excessive softening of the plants. This species is also particularly prone to losing its roots with overwatering so it is also essentially a winter‑growing species which likes at least some resting period during the summer.
1999 – Sims in 1805 commented on the name H. cymbiformis, saying ‘Its name (with too much latitude by the way) is taken from its leaves’, which prompted Haworth to change the name to H. concava. Von Poellnitz repeatedly exclaimed at the poor coloration of these plants in Europe as opposed to the colours which the plants develop under good light, when they are indeed very attractive. The two illustrations in Salm Dyck’s Monograph of Aloe cymbiformis and Aloe planifolia are really very similar in relation to the variation within this one species. One would think thus that a more sensible and conservative approach to species would have been achieved a great deal earlier than it has. It is not obvious to which species H. cymbiformis is most closely allied, but it does seem to be closely associated with H. cooperi. Bayer and Pilbeam may have been in error in their treatment of Uitewaal’s re-appraisal of H. obtusa Haw. as it is not easy to find the obvious field counterpart. The solution suggested below may not be the most appropriate. Col Scott also regarded obtusa as a variety of H. cymbiformis but unfortunately seems to have illustrated the H. cooperi variant. A population on the Kat River near Fort Beaufort sampled by Scott, supports his argument and the decision taken here. The explanation regarding the var. incurvula in the 1982 Handbook is poorly constructed. What was meant, was the recognition of the variety despite its limited distribution, because of the evidence claimed by G.G. Smith, and because of its possible transitional nature towards H. gracilis. H. lepida is regarded as a variant of H. cymbiformis because it could not be re-located despite a detailed description of the one locality where it was recorded. A collection further to the east does not appear to differ dramatically from an already wide range of forms.
Breuer and Metzing nominate a specimen as a neotype when the early illustrations are excellent and are really the basis for the correct and historical application of the name. Haworth cited and accepted the illustrations in Botanical Magazine as well.
a. var. cymbiformis. The typical species is considered to comprise the main body of the species which occurs from Port Elizabeth, eastwards to East London and inland to Adelaide and Committees on the Fish River. Plants in this area tend to have broad, flat smooth leaves without spines.
Distribution: 3226 (Fort Beaufort): Near Alice (-DB), Smith 5635 (NBG); Kat River, W. Alice (-DB), Smith 105 (NBG); 13km S. Fort Beaufort (-DC), Smith 5617, 5617a (NBG); Rocklands (-DC), W.E.Armstrong in Smith 2801 (NBG); Sulphur Baths (-DC), Smith 2795, 3826, 7371 (NBG), Bayer 4655 (NBG); W. Sulphur Baths (-DC), Bayer & Bruyns 6593 (NBG). 3227 (Kingwilliamstown): Debe Nek (-CC), Britten in PRE 39472; Fort Murray Bridge (-CD), Smith 3111, 3317, 3576 (NBG); Bridal Drift (-DC), Smith 2806 (NBG); Umdanzini (-DD), Smith 5336, 5337 (NBG). 3325 (Port Elizabeth): Kranspoort, W. Patterson (-BC), Bayer 4549 (NBG); Slagboom Dam (-BC), Branch 37 (NBG); Below old Fort (-DC), Smith 5040 (NBG); Walmer (-DC), Smith 2790, 2844 (NBG); Baakens Valley (-DC), Smith 3894 (NBG), Paterson 155 (BOL). 3326 (Grahamstown): Fish River valley (-AA), Dyer 4549 (PRE); Carlisle Bridge (-AA), Smith 5567, 5597 (NBG); Cloudlands (-AB), Britten in BOL71308; Howiesonspoort (-AD), Smith 105a, 439, 909, 2843b, 5302, 5305 (NBG); The Fort (-BA), Courtenay-Latimer in Smith 5066 (BOL, PRE), Smith 5066 (NBG, PRE); Between Carlisle Bridge and Fort Brown (-BA), Smith 5066 (NBG); E. Fort Brown (-BA), Bayer 1620 (NBG); Ballinafad (-BB), Smith 3365, 5404a, 5405a (NBG), Bayer 4652 (NBG); Horseshoe (-BB), Smith 2765, 3124, 5311, 5312 (NBG); Committees (-BB), Compton 17834 (NBG), Smith 5071, 5404 (NBG); 3km W. Committees (-BB), Smith 3382 (NBG); S. Committees (-BB), Smith in NBG322/40; Giffords Bush (-BB), Smith 1997 (NBG); 30km E. Peddie (-BB), Smith 3515 (NBG); Grahamstown (-BC), Britten 218 (PRE); Grahamstown (-BC), Britten in PRE 34903; Grahamstown (-BC), Britten in PRE 39480; Grahamstown (-BC), Curator PRE Bot. Garden in PRE 26300; 24km S. Grahamstown (-BC); Stayner in KG254/70; Blaaukranz (-BC), Smith 5544 (NBG); Fernkloof (-BC), Smith 5629 (NBG); Mt. Drive (-BC), Britten (NBG), Dyer 6 (BOL); Kowie (-BC), Dyer 7 (BOL); Kariega (-DA), Branch 43 (NBG); Bussock Farm (-DA), Smith 768 (NBG). 3327 (East London): Peddie (-AA), Smith 3112 (NBG); Paradise, Wooldridge (-AA), Smith 2800, 5602, 5671 (NBG); Gqora (-AA), Smith 5777 (NBG); W Woolridge, Peddie (-AB), Smith 3113, 3115 (NBG), Bayer 4648 (NBG); Kapp-Fish confluence (-AC), Bayer 4654 (NBG); Kaffirdrift (-AC), Smith 655, 5255, 5261, 5262, 5263, 5274 (NBG); E. Fish River (-AC), Smith 5256 (NBG); Wesley to Falloden (-AD), Smith 3157 (NBG); Chalumna (-BB), Smith 571, 2786, 3089, 3391, 3392, 5131, 5313, 5400, 6199 (NBG).
Inadequately located: Cape, Marloth 6284 (PRE), Long in Smith 3892 (NBG), Smith 398, 2804, 3112 (NBG), Stellenbosch 3889, 5560; ex hort, Ross-Frames in NBG76/48; Zaysdorp, NBG101825, Warden (BOL); ex hort, Whitehill (NBG); Albany, Dyer in NBG1806/30, Britten in NBG 734/31, Luyt in NBG309/45.
Haworthia cymbiformis var. cymbiformis JDV92/89 Kleinmond. Can have remarkable colours under stress.Haworthia cymbiformis var. cymbiformis JDV90/54 Beverly Cove. In dense clumps on steep rock faces.
b. var. incurvula (V.Poelln.) Bayer :124(1976). Bayer :36(1982). Scott :94(1985). H. incurvula V.Poelln., Feddes Repert.Spec.Nov. 31:85(1932). H. cymbiformis var. planifolia Flow.Pl.S.Afr. 9:t356(1929). Type: Grahamstown, Plutosvale, Mrs E. Ferguson. Not preserved. Neotype (B&M): Plutosvale, Britten (BOL71307).
incurvula: curved inwards.
As stated above, there is not very much substance to this variety either. It is smaller than the norm and relatively narrow-leaved. It has been collected many times from the same locality. The flower is very similar to that of H. gracilis var. minima, but Smith nevertheless maintained that it is continuous with H. cymbiformis. What he said was this “At the type locality near the top of the slope of a very deep valley, this plant is hardly variable, but as one descends, the plant changes, and at a point in the valley about a mile from the type locality, they are in appearance approaching H. cymbiformis.” The correct way to establish this is with physical evidence, and this is absent.
Distribution: 3326 (Grahamstown): Fish River Ridge (-AB), Britten in PRE 34959; Plutosvale (‑BA), Smith in NBG340/35 (BOL), Smith 5402 (NBG), Britten 11 (BOL), Britten in PRE 34909, in PRE 39477, Long 1029 (PRE), Dyer 3 (BOL), Dyer 2082 (PRE), Fourcade 99 (NBG), Smith 915 (NBG); Road to Plutosvale (-BA), Reynolds 2948 (PRE); 16km from Grahamstown (-BA), Erens in PRE 34910; S. Plutosvale (-BA), Smith 5402, 5403 (NBG); S. Hunts Drift (-BB), Smith 5741, 6508 (NBG).
Inadequately located: Albany, Dyer in NBG1803/30 (NBG), Smith 1123 (NBG), Luckhoff in NBG 404/34, Smith in NBG340/35, Britten in NBG740/31 (BOL); ex hort, NBG705/30, NBG1110/36, Luyt in NBG302/45, in NBG306/45, Whitehill (NBG).
Haworthia cymbiformis var. incurvula JDV93/60 Plutosvale. The relationship of this variety with H. gracilis is uncertain.
c. var. obtusa (Haw.) Baker JLinn.Soc. 18:209(1880). Bayer and Pilbeam, Cact.Succ.J(U.S.)46:166(1974). Scott idem. 48:260(1976). Scott :93(1985). H. obtusa Haw., Phil.Mag. 46:282(1825). Type: Cape ex hort Kew. Not preserved. Lectotype (designated here): Icon Kew library: H. umbraticola V.Poelln., Kakteenkunde 9:134(1937). H. cymbiformis var. umbraticola (V.Poelln.) Bayer :164(1976). Bayer :36(1982). Type: Swartwaterpoort, near Adelaide, W.E. Armstrong. Not preserved. Lectotype (B&M): icon (B): H. hilliana V.Poelln., Desert Pl.Life 9:103(1937). H. umbraticola var. hilliana V.Poelln., Feddes Repert.Spec.Nov. 44:234(1938). Type: Cape, ex hort Kew. Not preserved: H. obtusa var. pilifera fa truncata Jacobs., Handb.Succ.Pl. 2:574(1960). Type: ?. Not preserved.
obtusa: with obtuse leaves.
As discussed under H. cooperi, there is a real problem in understanding the situation concerning the blunt leaved forms of that species and of H. cymbiformis. I have seen very dark green forms with brown venation in cultivation which I would not relate to H. cooperi. The highly translucent forms from Fort Beaufort are greener and seem to be continuous with H. cymbiformis through the Swartwaterpoort. Certainly the plant mentioned above from the Kat River, are very close indeed in appearance to H. cooperi var. obtusata except for the brownish-green coloration. It has been noted that plants of that variety from Inverbolo (Upper Kei River) tend to become greener under lower light intensities. The herbarium record shows that the typical form of H. cymbiformis also occurs along the Kat River. Very robust forms in fact occur 10km S. Fort Beaufort. H. cooperi var. pilifera is also present in very close proximity. At Kagasmond and probably at Olifantsbeen nearby, the plants are very similar to the rather longer leaved forms, still with obtuse tips, which are found in Swartwaterpoort near Alicedale. A Kagasmond collection is also cited under var. dielsiana, which is indicative of the difficulty in using dry herbarium material to make indisputable identifications. G. Marx has made a collection from Swartwaterpoort in which the plants have the coloration of H. cooperi but the boat-shaped leaves of H. cymbiformis.
Distribution: 3226 (Fort Beaufort): Kagasmond (-CD), Bayer & Bruyns 6562 (NBG); Olifantsbeen (-CD), Krynauw in NBG268/43 (NBG); S. Adelaide (‑CD), Krynauw in NBG 67996; Blinkwater (-DA), Smith 6195 (NBG). Scott 600 (PRE), Bayer 4651 (NBG); Kat River, 10km SE. Fort Beaufort (-DC), Scott 1065 (PRE); Rocklands, Adelaide (-DC), Smith 2801 (NBG). 3325(Port Elizabeth (-BB), Bayer 4653 (NBG). 3326(Grahamstown): Thornkloof (-AA), Bayliss in KG382/76; S. Alicedale (-AC), Bayer 4650 (NBG); Alicedale (-AC), Britten in PRE 34905.
Haworthia cymbiformis var. obtusa JDV91/100 north of Fort Beaufort Is very variable in terms of leaf-shape and colour.Haworthia cymbiformis var. obtusa JDV96/63 Kagasmond. The plants have a greyer-blue colour than the Fort Beaufort plants and the leaves are more marginate.
d. var. ramosa (Smith) Bayer comb.nov. H. cymbiformis fa ramosa (Smith) Bayer :149(1976). Bayer :34(1982). H. ramosa Smith, JS.Afr.Bot. 10:22(1940). Type: CAPE-3427 (Peddie): Wooldridge (-AB), Smith 3168 (NBG).
ramosa: branched.
Consistency is a difficult ideal and this variety does not conform well with the principle of substance. It is only known from a long crescent-like rock-face north of Woolridge where plants vary from the normal stemless to increasingly stemmed plants on a gradient from west to east.
Distribution: 3327 (Peddie): Wooldridge (-AB), Smith 3168 (NBG, PRE), Smith 3105 (NBG), Bayer 4648 (NBG); NW. Wooldridge (-AB), Smith 3168, 3169 (NBG).
Haworthia cymbiformis var. ramosa JDV85/53 Wooldridge. The population is quite extensive and not all the plants are as stemmed.
e. var. reddii (Scott) Bayer comb.nov. H. reddii Scott, Cactus Succ.J(US) 66:182(1994). Type: CAPE-3226( Fort Beaufort): Waterdown Dam, Cathcart(-BB), Scott 8968 (PRE).
reddii: for Dr V.B. Reddi.
Plants from this population at Waterdown Dam have been known for a long time and identified (Bayer,1982) as possibly intermediate between H. cymbiformis and H. batesiana. Col Scott similarly mentions both species names in his discussion and the matter appears rather problematic. The population was portrayed as a depauperate one, at least in my perception, with quite considerable variation between the few odd plants at the site. The few clones sampled in 1982 were not as robust as the one described by Scott, who also thought the population to consist of but a few individuals. A re-visit to the site by Bayer and Bruyns in 1996 revealed that the south-facing cliff alongside the dam is clothed with huge numbers of plants. The huge clumps are just like those of H. cymbiformis. Some of the plants have very distinctive translucent dots and lines, others unmarked and uniformly opaque. The floral characters mentioned by Scott are not definitive but the flowers do appear to have strongly colored veins. At the time (Bayer :30, 1982) the species batesiana was still upheld, although not positively associated with field populations. Since then the range of H. marumiana has been shown to extend to at least Queenstown and further north. The Cathcart population does not seem to belong there although the block patterning in the leaves does suggest this.
At the same time the known range of H. cymbiformis has been extended by P.V. Bruyns to the upper reaches of the Black Kei much nearer to Queenstown. This is on a south-facing cliff at the farm Turnstream. Here there are small forms which distantly resemble H. lepida as described and illustrated by Smith, and also suggesting the same possible link with H. marumiana and certainly with reddii. This is particularly so because the Waterdown Dam is on the upper reach of the Black Kei. A little to the southeast is also a population of larger plants on a very high west-facing cliff which are however with spined margins, and thus apparently belonging to the var. setulifera. The block-patterning in the leaves of these two populations is not as marked as the Waterdown plants. There is an old Galpin collection from even nearer to Queenstown. Col. Scott was not correct in his opinions about H. marumiana var. batesiana and it is evident from his book that H. marumiana was unfamiliar to him too. The population at the Waterdown Dam does not seem substantial enough in terms of viability nor range, to justify species rank. What is needed is cognitive exploration of potential new localities to substantiate already expressed opinions. In this case there are new records for var. batesiana as well as for H. marumiana and H. cymbiformis.
Haworthia cymbiformis var. reddii JDV92/113 Waterdown Dam. The translucent markings are quite variable.Haworthia cymbiformis var. reddii JDV96/68 Turnstream. A new collection north-east of Cathcart which confirms relationship with H. cymbiformis. Unfortunately this population is under-sampled and more information is required for a really good decision to be made.
f. var. setulifera (V.Poelln.) Bayer comb.nov. H. planifolia var. setulifera V.Poelln., Kakteenkunde 5:54(1938). idem., Feddes Repert.Spec.Nov. 45:163(1938). Type: East London, Stellenbosch 3332. Not preserved. Neotype (designated here): CAPE-3228(East London): Kwelegha bridge (-CC), Smith 5257 (NBG): H. cymbiformis var. obesa V.Poelln. idem. 45:166(1938). Type: Idutywa, Bashee River (-BA), G.W. Reynolds. Not preserved. Neotype (designated here): CAPE-3228(Butterworth): Xobo River, E. Idutywa (-BA), Smith 7796 (NBG).
setulifera: bearing small bristles.
In the earlier handbooks, the representative specimen for the species was poorly chosen from a Nahoon River specimen. It is apparent from Haworth’s acceptance of the Bot. Mag. illustration that the more western forms such as those at Baakens River or Howiesons Poort, would have been more in keeping with the original circumscription of either the typical species or H. planifolia. East and north of East London, H. cymbiformis begins to develop a thicker and shorter, more deltoid leaf and the teeth become markedly spined and von Poellnitz’ name is re-instated for this variety.
Distribution: 3128 (Umtata): Mquanduli (-DC), Walker in NBG2271/27 (BOL). 3227 (Kingwilliamstown): Highclere (AB), Bayer & Bruyns 6573 (NBG); Inverbolo (-BC), Bruyns (NBG); Bluewater (-DA), Smith 676 (NBG); Near Komgha (-DB), Marloth 6510 (PRE); Tangla River (-DC), Smith 3881, 3882a (NBG); Newlands Location (-DD), Smith 3510 (NBG); Pump Stn. (-DD), Smith 611, 2785, 3096 (NBG); Kings Farm (-DD), Smith 3071 (NBG); McCleantown (-DD), Smith 3126, 3127, 2883, 2884, 2885, 3882 (NBG); Slippery Drift (-DD), Smith 3122, 3125, 3134, 3134a (NBG); below Horseshoe (-DD), Smith 3097 (NBG); Fort Jackson (-DD), Smith 3133, 3388, 3389, 3389a (NBG). 3228 (Butterworth): Willowvale (-AD), Luyt in NBG180/42, in NBG17/46, in NBG57/46, NBG341/38; Xobo River, E. Idutywa (-BA), Smith 7796 (NBG), Smith in NBG341/35 (BOL), Reynolds 2850 (PRE), Reynolds 146 (BOL); Xobo (-BA), Reynolds in PRE 39470, in NBG660/38, Smith 2760, 2796 (NBG); S. Mooiplaas (-CC), Bayer 1706 (NBG); Gonubie (-CC), Smith 6826 (NBG); Kwelegha bridge (-CC), Smith 5257 (NBG); Kwelegha (-CC), Smith 5251, 7183 (NBG); Kei River (-CC), Holmes (BOL).
Haworthia cymbiformis var. setulifera MBB6573 Highclere. This is from the Black Kei north-east of Cathcart, very near to the Turnstream population of the var. reddii. The flowers and seed in this population seem to be very large.Haworthia cymbiformis var. setulifera JDV91/91 north-east of East London. Leaves spinescent and more triangular in cross-section.Haworthia cymbiformis var. setulifera JDV91/91 north-east of East London. ALways in mats on steep rock faces.
g. var. transiens (V.Poelln.) Bayer :162(1976). Bayer :36(1982). H. planifolia var. transiens V.Poelln., Feddes Repert.Spec.Nov.45:163 (1938). Type: Cape, Prince Alfred Pass, Archibald 327. Not preserved. Lectotype (B&M): icon (B): H. cymbiformis var. translucens Triebn. et V.Poelln. idem. 45:166(1938). Scott :94(1985). Type: Cape, Prince Alfred Pass, Lategan in Triebn.1137. Not preserved. Neotype (designated here): CAPE-3323(Willowmore): Prince Alfred Pass, Smith 5709 (NBG): H. cymbiformis var. multifolia Triebn. idem. 45:166(1938). Type: Uitenhage, W.E. Armstrong. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): Hellsgate, UItenhage (-CB), Smith 2794 (NBG): H. cymbiformis var. brevifolia Triebn. et V.Poelln. idem.: 165(1938). Type: Cape, Hellsgate, Uitenhage, Mrs I. King in Treibn. 1068. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth: Hellsgate, Mrs I. King in Smith 2756 (NBG).
transiens: changing into.
In the publication where Von Poellnitz published his two varieties, he actually expresses doubt about identifying three parent species. Yet he states that H. planifolia var transiens is between H. planifolia and H. cymbiformis, but nearer to the former ‘to look at’. Two pages further he describes a var. translucens of H. cymbiformis from the same locality. In this work the circumscription of var. transiens is widened to include the Uitenhage elements at Hellsgate. There are variants in the Gamtoos Valley (eg. Andrieskraal) which are reminiscent of this highly translucent form, and it is not certain just how these variants relate to either H. cymbiformis or H. gracilis. The element H. gracilis var. picturata is applied to those forms of that species which are very similar to H. cymbiformis variants. The ‘clear’ way (a comment made by Dr M. Hayashi) in which var. transiens is related to H. mucronata is symptomatic of the alternative solutions available in classifying Haworthias, which are not always clear. It is generally understood that the Little Karoo species are continuous with the Eastern Cape species and this is commonly expressed in Von Poellnitz identifications, and also a feature of Col. Scott’s species distributions.
Thus H. mucronata can be allied with equal facility to either H. cymbiformis or H. cooperi, when in fact in the field it is more intimately related to H. decipiens. The location of this note is a powerful reminder that distinctions between species are highly blurred and that alternative solutions are possible.
Distribution: 3323 (Willowmore): Prince Alfred Pass (-CC), Smith 5624a, 5709, 5624, 5624a (NBG), Taute (BOL), Fourcade 3490 (BOL); Oskloof (-DA), Bruyns 7077 (BOL); Luiskraal (-DA), Forrester 399 (NBG). 3324 (Steytlerville): Scholtzberg (-CA), Van Jaarsveld 7804 (NBG); Andrieskraal (-DA), Fourcade 176 (NBG). 3325 (Port Elizabeth): Hellsgate, Uitenhage (-CB), Smith 2794 (NBG); Hellsgate, Mrs I. King in Smith 2756 (NBG); Kemachs (-CB), Smith 905 (NBG); Near Port Elizabeth (-DC), Smith 3892 (NBG); Stayner in NBG46/56, Stayner in KG80/70 (NBG), Taute in NBG1283/36, in NBG468/37, Taute .
Inadequately located: ex hort, NBG486/30.
Haworthia cymbiformis var. transiens JDV86/110 Prince Albert Pass. Now known to be more widespread and plants in other populations can be greener.
10. Haworthia decipiens V.Poelln., Feddes Repert.Spec.Nov. 28:103(1930). Bayer :111(1976). Bayer :37(1976). H. pearsonii Wright sensu C.L. Scott, Aloe 18:7(1980). Scott :44(1985). Type: Cape, near Zwartberg mountains, Mrs van der Bijl. Not preserved. Neotype (designated here): CAPE-3322 (Oudtshoorn): Kleinsleutelfontein, E. Prince Albert (-AB), Bayer 5157 (NBG).
decipiens: deceiving.
Rosette stemless, slowly proliferous, to 20cm φ. Leaves broadly ovate sometimes acuminate, relatively thin, marginal spines sparse but broad at the base, bright green in colour. Inflorescence robust, simple. Flowers many, densely arranged, broad and flat across top.
1982 – Von Poellnitz recorded this species only once from the Zwartberg Mountain in the Prince Albert district and derived the name from the similarity to H. arachnoidea. As pointed out in the discussion on H. arachnoidea, there may be a close relationship between these species. Available distribution information indicates that they may be geographically complementary. However, at its most westerly point (Klaarstroom), H. decipiens has the broad flat yellowish‑green leaves of H. Iockwoodii but with sparse, short broad marginal spines. The leaf tips do not die back. H. decipiens may grow up to 120mm in diameter but it occurs in smaller forms too, only 30‑40mm in diameter. From Klaarstroom it extends eastward to Springbokvlakte in the south. Northwards it is found from Klipplaat to Pearston. However, in this area there is some deviation from the typical form and the plants tend to have translucent areas in the leaf with a tendency to be bluish‑green. There is thus a suggestion of intergradation with H. bolusii and H. cooperi on the grounds of both superficial similarities and distribution. The picture in the west is similarly obscure and the forms regarded as H . decipiens east of Uniondale also have leaf translucence, but tend to be a darker purplish‑green. In the Groot River area west of Campherpoort, H. decipiens occurs as a dwarfed form with notably incurved leaves.
1999 – Although von Poellnitz described this species in 1930, by 1938 he had still recorded only this single collection. However, in the absence of a preserved specimen and adequate locality data, there can be no certainty that the original description is correctly interpreted. In all its guises it is a very common species as recognised here; and the type, as selected here seems to represents the species as it is more generally known. The type selected by Breuer and Metzing is unfortunate as it is not geographically correct and is better related to the blue-green var. cyanea.
Col Scott has taken the view that H. decipiens is synonymous with H. pearsonii Wright but unfortunately does not consider or refer to H. decipiens sensu Bayer anywhere in his revision. It is as difficult to relate the Kew illustration (presumably the chosen lectotype) of H. pearsonii to a natural population. It apparently does not have translucent leaf faces. H. decipiens of von Poellnitz was a relatively small plant and if indeed it came from north of the Zwartberg mountains as the reference to the Prince Albert district suggests, then it could probably be only as interpreted here, or as a variant of H. marumiana. Von Poellnitz did not relate his H. decipiens to pearsonii so it is an improbable option. Scott’s illustration is not representative of either of these interpretations and, without reference to locality, could be of the var. cyanea here described (the blue-green form). It seems to compare very poorly with the Kew illustration. The species as recognised here is a very complex one with connections to H. bolusii, H. lockwoodii, H. mucronata, H. cooperi and also H. gracilis.
a. var. decipiens. It is concluded that von Poellnitz’ species did in fact come from the Prince Albert area where the species transposes to H. lockwoodii. The var. cyanea is also found in the same general area and it is smaller and with more incurved leaves. The species is most strongly represented in the Willowmore, Steytlerville area where the plants are large and robust. The leaves may recurve and are heavily spined with large spines, rather broad and flat at the base. Interspersed over the same area, and extending to the east, is a smaller form which tends to develop reddish venation.
Distribution: 3221 (Merweville), Prince Albert (-DC), Bolus 11657 (BOL, PRE). 3224 (Graaff Reinet): near Jansenville (-DC), Smith 3642 (NBG). 3322(Oudtshoorn): W. Prince Albert (-AA), Bayer 5182 (NBG); Kleinsleutelfontein, E. Prince Albert (-AB), Bayer 5157 (NBG); W. Klaarstroom (-BC), Bruyns in KG124/77 (NBG); S. Prince Albert (-CA), Bayer 5261 (NBG). 3323 (Willowmore): Skerpkop, E. Willowmore (-AD), Bayer in KG231/70 (NBG); Constantia (-BB), Bayer in KG140/72 (NBG). 3324 (Steytlerville): Campherpoort (-AA), Smith 3649, 3657 (NBG); Steytlerville (-AD), Rossouw 453 (NBG); Kleinpoort (-BD), Smith 7065 (NBG); Dam se Drif (-CA), Rossouw 478 (NBG), Bruyns 1842 (NBG); S. Steytlerville (-CA), Stayner in KG637/61 (NBG); 3325 (Port Elizabeth): 20km N. Glenconnor (-AC), Long (BOL).
Inadequately located: Kleinswartberg, Wood (BOL).
Haworthia decipiens var. decipiens JDV96/102 Tierberg. This is a smaller variant in which the venation is reddish.
Haworthia decipiens var. decipiens JDV94/93 Springbokvlakte. The range of variation is not well understood and some populations resemble H. bolusii var. blackbeardiana.
Haworthia decipiens var. decipiens JDV94/73 west of Klipplaat. There are transformations to the smaller var. minor and to this, the red-veined form mentioned.
Haworthia decipiens var. decipiens JDV87/169 south of Stytlerville. The better known forms are large with spreading open leaves which are strongly spined.
Haworthia decipiens var. decipiens JDV94/69 Kleinsleutelfontein. Considered to be fairly typical of a very variable complex.
b. var. cyanea var.nov. Type: CAPE-3324 (Steytlerville): Fairview, W. Jansenville (-CD), Bayer 4180 (NBG, Holo.).
cyanea: blue.
Differs in being smaller with incurving leaves and in the bluish-green coloration. (A var. decipiens foliis venetis incurvatis differt).
This variety occurs north of the mountains between Jansenville and occurs as far to the northwest as Merweville. It is distinguished from H. arachnoidea by its translucence in the upper leaf and by the more robust spines. Herbarium specimens will be even more difficult to separate and two specimens cited under H. arachnoidea from the Jansenville area, may in fact be specimens of H. decipiens var. cynaea It generally differs from H. bolusii var. blackbeardiana also on account of the robust spines but also for the broader and shorter leaves.
Distribution: 3221 (Merweville): De List, Merweville (-CB), Bayer 2377 (NBG). 3222 (Beaufort West): Trakas Kuilen (-DC), Bayer sn. (NBG). 3224 (Graaff Reinet): Fairview, west of Jansenville (-CD), Bayer 4180 (NBG); Meerlust (-DC), Bayer & Bruyns 6580 (NBG). 3322 (Oudtshoorn): E. Klaarstroom (-BC), Bayer 4440 (NBG). 3323 (Willowmore): S. Rietbron (-AB), Bruyns in KG 439/75 (NBG); Beervlei (-AB), Latti in KG56/78 (NBG); Georgida (-AD), Bayer in KG436/75 (NBG); Zuurberg near Georgida (-AD); Fourcade 4637 (NBG); Nahoogte (-BC), Van Jaarsveld 7865 (NBG); N. Uniondale (-CA), Bayer 2083 (NBG), Bruyns 2241b (NBG). 3324 (Steytlerville): Hanekam (-BD), Bayer 4657 (NBG).
Inadequately located: Georgida, Fourcade 4637 (BOL); Uniondale, Van Blerk (BOL).
Haworthia decipiens var. cyanea MBB6580 north of Jansenville. Very like H. archnoidea but with bluish-green leaves and translucence in the upper-leaf.
Haworthia decipiens var. cyanea JDV90/103 soth-west of Merweville. Notmany populations are known, but it may be more abundant in the southern Karoo than supposed.
c. var. minor var.nov. Type: CAPE-3324 (Steytlerville): Kleinpoort, Smith 3588 (NBG, Holo.).
minor: smaller.
Differs in being much smaller, to 6cm φ, with broad incurved leaves, and light green in colour. (A var. decipiens foliis valde parvioribus incurvatis et subviridibus differt).
Occurs in the Groot River valley between the ranges. It may be directly related to H. gracilis var. viridis, and the transition to that species can be followed southward through the Zeekoeinek Pass near Baroe. Northwards the transition is to the var. pringlei. A similar transition to H. translucens var. viridis occurs in the Perdepoort north of Sapkamma.
Distribution: 3225 (Somerset East):In valley behind Bosberg (-DA), Van Der Westhuizen 287 (PRE). 3323 (Willowmore): Redcliffe (-BA), Bruyns 7052 (BOL). 3324 (Steytlerville): Campherpoort (-AB), Smith 7061 (NBG); Campherpoort (-AA), Barker 5009 (NBG), Bayer in KG 315/70 (NBG); Grootriver, Mara (-AA), Bayer 2076 (NBG); Tuinskloof (-AC), Bruyns 3125 (NBG); NE. Steytlerville (-AD), Smith 3591 (NBG); Waaipoort (-AD), Bayer & Bruyns 6583 (NBG); NW. Die Bordjie (-BC), Bayer & Bruyns 6587 (NBG); Two Waters (-BC), Smith 7244 (NBG); Kleinpoort (-BD), Smith 3588 (NBG); Baviaanskloof (-CA), Wisura 1837 (NBG); Ouplaas (-DB), Bruyns 7040 (BOL). 3325(Port Elizabeth): Sapkamma to Perdepoort (-AC), Bayer & Venter 6618, 6619 (NBG).
Haworthia decipiens var. minor JDV91/118 west of Sprinbokvlakte. The plants grow larger in cultivation.
Haworthia decipiens var. minor JDV96/71 north of Campherpoort. Where the species occurs in sandstone it changes to this smaller form which varies continuously with H. gracilis (notably the var. viridis).
d. var. pringlei (Scott) Bayer comb.nov. H. pringlei Scott, Bradleya 12:103(1994). Type: 3224 (Graaff Reinet): Adelaide district (-DD), Scott in PRE8970.
pringlei: for Victor Pringle.
This bright green plant with contrasting white spines has been known for a long time and is represented by many collections from the general area of Jansenville, Klipplaat, Aberdeen, and Pearston. These are well represented in the Compton Herbarium. The leaves are incurved and erect to sub-erect and not as broad and ovate-deltoid as in the typical variety. The connection to Adelaide is by no means clear and there is very little to substantiate this as a discrete element in that area. This is because H. cooperi, H. bolusii var. blackbeardiana, H. gracilis and H. cymbiformis all present problems of their own there. I have a collection from Baviaanskrans which I have placed with H. bolusii var. blackbeardiana and it is to a degree very similar to this element.
Distribution: 3223 (Rietbron): S. Aberdeen (-DC), Perry 659 (NBG). 3224 (Graaff Reinet): Aberdeen Road (-CD), C.A. Smith 2806a (PRE); Oatlands (-CD), Smith 907 (NBG); Ebenezer (-DB), Smith 7245 (NBG), Bayer 2070 (NBG); Harefield (-DB), Smith 7244 (NBG); Welgelegen (-DC), Bayer & Bruyns 6581 (NBG); Jansenville (-DC), Stayner in KG188/62 (NBG); Adelaide district (-DD), Scott in PRE 8970. 3324(Steytlerville): Klipplaat (-AB), Branch (NBG); SE. Mt.Stewart (-AB), Bayer & Bruyns 6582 (NBG).
Haworthia decipiens var. pringlei JDV received from Dr. V.B. Reddi. There may be a closer relationship to H. bolusii var. blackbeardiana and there is a collection from south of Eastpoort which supports that possibility.
11. Haworthia emelyae V.Poelln., Feddes Repert.Spec.Nov. 42:271(1937). Bayer, Natn.Cact.Succ.J 34:28(1979). Bayer :115(1976). Bayer :38(1982). Type: Cape, locality unknown, Mrs E. Ferguson in Long 322. Not preserved. Lectotype (B&M): icon (B): H. blackburniae V.Poelln., Kakteenk. 9:132(1937). Nom. illegit. non Barker 1937. H. correcta V.Poelln., Feddes Repert.Spec.Nov. 43:103(1938). Type: Cape, Calitzdorp, Mrs E. Blackburn in Triebn. 978. Not preserved. Neotype (designated here): CAPE-3321(Ladismith): Rooiberg, E. Vanwyksdorp(-DA), Mrs Schnettler in KG 335/71 (NBG): H. picta idem. 44:133(1938). V.Poelln., Desert.Pl.Life :126(1939). Type: Cape, Moeras River, Mrs S. Blackburn in Triebn. 1062. Not preserved. Neotype (designated here): icon, Fourcade 72 (NBG).
emelyae: for Emely Ferguson.
Rosette to 10cm φ, seldom proliferous. Leaves 15-20, distinctly retused, pointed, barely translucent, with scattered elongate small flecks, with obscure raised tubercles, lined, reddish-brown hued. Inflorescence simple. Flowers 15-20, white.
1982 – H. emelyae is a widely distributed species occurring from Uniondale through the Little Karoo to west of Ladismith. It occurs in a variety of different geological formations and varies accordingly. The plants have very short recurved leaves with convex end areas usually flecked with pinkish markings. The forms east of Oudtshoorn usually have rounded end areas and are scabrid on the upper leaf faces. Westwards the leaves become pointed and smoother. However, there are clear signs of intergradation with H. magnifica in the Muiskraal area of Riversdale. Doubt has been cast on the origin of Mrs Ferguson’s plants but G.G. Smith’s records clearly indicate that they came from Van Wyksdorp and that they were collected there by a Mrs Le Roux. The var. multifolia has up to 60, more slender, suberect leaves; and was compared in the original description with H. serrata. Other forms of H. emelyae where the few leaves are shortly recurved and round‑tipped can similarly be compared with H. magnifica var. atrofusca. The eastern forms with the flatter, rounded leaf tips can be compared to H. bruynsii and H. springbokvlakensis. H. emelyae is generally non‑proliferous.
1999 – The 1982 discussion is fraught with error as two species are actually involved. The scabrid element referred to above has since been excerpted as H. bayeri and it is fairly obvious that there can be no extension to H. bruynsii which belongs in the Hexangulares. The problem is compounded by the fact that von Poellnitz described three species all of which are synonymous, and yet may have cited H. bayeri among these. The integration with H. magnifica was through the var. major which is now transferred to H. emelyae because of the intermediate populations mentioned above which occur just east of, and in the closer vicinity of, Muiskraal.
a. var. emelyae. The typical variety is rather strongly flecked and the leaves are always retused to horizontal and close to ground level. The neotype cited above is a specimen originating from near Vanwyksdorp and sent by the original collector (Mrs Le Roux) to G.G. Smith.
Distribution: 3321 (Ladismith): 20km W. Ladismith (-BD), NBG144768; Springfontein (-CC), Smith 5787 (NBG); 42km Calitzdorp to Vanwyksdorp (‑DA), Mrs Le Roux in Smith 5437 (NBG); Vanwyksdorp (-DA), Joubert (BOL);Rooiberg, E. Vanwyksdorp (-DA), Mrs Schnettler in KG 335/71 (NBG); SE. Vanwyksdorp (-DC), Bayer in KG257/77 (NBG). 3322 (Oudtshoorn): Mt Hope (-CA), Schnettler in KG582/69 (NBG); Moeras River (-CC), Smith 2447 (NBG); N. Robinson Pass (-CC), Bruyns in KG434/75 (NBG); Zebra (-CD), Smith 2905 (NBG); Erfpacht (-DD), Smith 5808, 6106 (NBG); 3323 (Willowmore): Uniondale to Avontuur (-CA), Smith 2935 (NBG).
Inadequately located: Ladismith, Pillans 858 (BOL); Oudtshoorn, Taylor (BOL).
Haworthia emelyae var, emelyae JDV93/28 north of Herold. Attractively marked end-areas make this a sought-after species.
Haworthia emelyae var, emelyae JDV90/120 south-east of Oudtshoorn. Plants in this population are very comparable with the var. comptoniana with much finer reticulation and a smooth leaf surface.
Haworthia emelyae var, emelyae JDV93/22 south-west of Calitzdorp. The plants get more scabrid and have small white markings in the leaves as does, for example, H. turgida.
Haworthia emelyae var, emelyae JDV84/11 Zebra, also fairly close to the population of H. bayeri.
Haworthia emelyae var, emelyae JDV90/10 south of Oudtshoorn. Usually associated with white quartz outcrops.
Haworthia emelyae var, emelyae JDV93/28 north of Herold. This population has been decimated by road construction and subsequent over collecting.
Haworthia emelyae var, emelyae JDV90/120 south-east of Oudtshoorn. Fairly glabrous plants which strengthen the link to the var. comptoniana.
Haworthia emelyae var, emelyae JDV97/133 south of Vanwyksdorp. One of three new collections which seem to confirm continuity with the var. major.
Haworthia emelyae var, emelyae JDV93/22 south-west of Calitzdorp. Several recent collections seem to confirm the relationship of the varieties.
b. var. comptoniana (Smith) Venter & Hammer Cact.Succ.J(US) 69:77(1997). H. comptoniana Smith, JS.Afr.Bot. 11:76(1945). Bayer 108(1976). Bayer :33(1982). Scott :128(1985). Type: CAPE-3323 (Willowmore): Georgida (-AD), M. Malherbe in Smith 3433 (NBG).
comptoniana: in honour of Prof.R.H. Compton.
1982 – The species with the flattened ends to the leaves are the most sought after, and H. comptoniana is particularly attractive. It occurs in quartz patches and the plants are often well hidden under the stones. The plants are bigger than in H. emelyae (the nearest relative), growing up to 120 mm in diameter in cultivation. The growth form parallels that of H. emelyae exactly and the difference is that it is a bigger and entirely smooth species. The leaf end area is slightly pellucid and reticulated. Where H. emelyae is generally tinted purplish-brown, H. comptoniana is green. Distribution and abundance is not fully known and H. comptoniana is considered very rare in the field. It has proved very easy in cultivation and good seed is easily produced by cross-pollination.
1999 – This variety grows in very close association with H. bayeri but it is not known if other populations exist other than at the type locality. Some forms which are pale coloured and in which there is little contrast between the reticulation and the background colour and translucence of the leaf, are very plain. Other plants are darker coloured or with more conspicuous specks and marked reticulation, and they are are much more attractive.
Distribution: 3323 (Willowmore): Georgida (-AD), M. Malherbe in Smith 3433 (NBG), Smith 3433 (NBG), Bayer in KG114/72 (NBG).
Haworthia emelyae var. comptoniana JDV90/8 Georgida. Recorded only from one population and severely threatened by collectors.
Haworthia emelyae var. comptoniana JDV90/8 Georgida. Also very cryptic in the summer months.
Haworthia emelyae var. comptoniana KG114/72 Georgida. The reticulation and relative length of the leaves are quite variable.
c. var. major (Smith) Bayer comb.nov. H. schuldtiana var. major Smith, JS.Afr.Bot. 12:1(1946). H. maraisii var. major (Smith) Bayer :132(1976). H. magnifica var. major (Smith) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). Type: CAPE‑3321 (Ladismith): Garcias Pass (‑CC), Smith 5370 (NBG).
major: greater.
1982 – The var. major has an unusually large flower similar to that of H. emelyae and there is evidence of intergradation of these two taxa.
1999 – Although in this variety the leaf tubercles are armed with a spine, the general form of the plants is that of H. emelyae, and populations east of Muiskraal confirm this relationship. The change in relationship forged here has its roots in the new collections of H. magnifica around Riversdale and in the new arrangement regarding H. maraisii. Another consideration is the position of var. paradoxa, mentioned in 1976, and that is its relationship with H. mirabilis. Similarly the following variety and its position need also to be considered.
Distribution: 3321 (Ladismith): Garcia’s Pass (‑CC), Smith 5370 (NBG), Garcia’s Pass (-CC), Dekenah 9 (PRE); Muiskraal (-CC), Smith 3458 (NBG), Bayer in KG118/71 (NBG); Riversdale (-CC), Muir in NBG164/25 (BOL); Sandvlakte (-CD), Bayer in KG138/72 (NBG).
Haworthia emelyae var. major JDV90/19 Muiskraal. A sandstone-dwelling variety with spines on the leaf surfaces.
Haworthia emelyae var. major JDV95/36 Muiskraal. The var. multifolia is now know to occur within a few kilometers of the var. major and these two may not vary continuously.
Haworthia emelyae var. major JDV95/23 Garcia’s Pass. A densely spined seedling grown by P. Bosch and coming from east of the type locality.
Haworthia emelyae var. major JDV90/22 east of Muiskraal. Several quite variable populations associated with shales.
d. var. multifolia Bayer Natn.Cact.Succ.J 34:31(1979). Bayer :39(1982). Type: CAPE‑3321(Ladismith): Springfontein, Riversdale (‑CC), Bayer 1558 (NBG).
multifolia: many leaved.
When this variety was described it was associated and compared with H. serrata. A new population is now known a little further to the west and collections by J. Dekenah suggest that it is possibly a little more widely represented in that general area. The plants have many more upright leaves and the relationship with H. serrata through H. heidelbergensis, which is also now better understood, becomes real. There is a strong suggestion that these links all also involve H. mirabilis and this is perhaps supported by von Poellnitz’ interpretation and citations of varieties of that species.
Distribution: 3321 (Ladismith): Springfontein, Riversdale (‑CC), Bayer 1558 (NBG); W. Springfontein (-CC), Smith 5769 (NBG); E. Springfontein (-CC), Smith 5768 (NBG), Smith 5389 (NBG, PRE), Bayer 1558 (NBG); Muiskraal (-CC).
Haworthia emelyae var. multifolia JDV87/162 Springfontein. Since located at two sites east and west of the type locality.
Haworthia emelyae var. multifolia JDV94/32 Brandrivier. In this population the plants are more robust.
12. Haworthia floribunda V.Poelln., Feddes Repert.Spec.Nov. 40:149(1936). idem. 44:228(1938). Bayer :117(1976). Bayer :39(1982). Scott :58(1985). Type: Cape, Heidelberg, Mrs E. Ferguson. Not preserved. Lectotype: icon (B). Epitype (B&M): Blackdown, N Heidelberg, Bayer 158 (NBG).
floribunda: many flowered.
Rosette stemless, up to 3cm φ, slowly proliferous. Leaves 20-30 dark green opaque, upto ovate-lanceolate, spreading, twisted with flattened, rounded tip, margins scabrid to dentate. Inflorescence simple, to 250mm. Flowers 10-15, greenish-white, few open together.
1982 – This is a very interesting small species with twisted lanceolate leaves with blunt rounded tips. It was described from plants collected north of Heidelberg (Cape) where they are all glabrous and where hybridisation with H. turgida also occurs. It has not been collected further west although there is a very old collection in the Botanical Research Institute (PRE) from Swellendam. Around Riversdale the plants may have scabrous leaves with denticulate leaf margins. There is a known population north of Albertinia in which the plants have more and shorter leaves, as well as another similar population near Gouritzmond. These two populations may suggest an affinity with H. chloracantha, as put forward by A.E. Speechley (unpublished). There may be such a relationship, but it seems likely that H. floribunda and H. parksiana are in fact related. They both tend to grow well‑shaded and in moss and lichen. Further exploration of the Gouritz River tributaries may produce an answer to this puzzle.
1999 – The Gouritz valley has provided no answers to the above puzzle but there have been a number of other significant collections. There is a population south of Swellendam of which specimens in cultivation have been very robust. They are relatively light green in cultivation and also individual plants are smooth as was the original type. Odd specimens have pointed leaves. Northwest of Swellendam is a similar but darker green plant also with pointed leaves and this is taken to be H. variegata. Further south from Swellendam, H. floribunda assumes the same form as around Riversdale. At Great Brak there is a population of plants growing with H. pygmaea which were assumed to be H. floribunda, but they are probably simply H. chloracantha var. denticulifera. This is true also of the populations south of Albertinia and also of one population south of Heidelberg which has pointed leaves. The flattened leaf-tip is not confined to this species, and appears in H. magnifica and H. maraisii. A relationship with H. variegata is probable through the two populations in the vicinity of Swellendam.
a. var. floribunda. The typical variety is not typical of the species at all and this glabrous variety is only known from the type locality at Heidelberg where it is very scarce. Hybrids with H. turgida are also present.
Distribution: 3420 (Bredasdorp): 6km N. Heidelberg (-BB), Smith 5545 (NBG, PRE), Bayer 158 (NBG); Heidelberg commonage (-BB), Ferguson in BOL20507.
Haworthia floribunda var. floribunda JDV88/22 north of Heidleberg. One of the few places where the plants have no spination.
Haworthia floribunda var. floribunda JDV89/22 north of Heidelberg.
b. var. dentata var. nov. Type: CAPE-3421 (Riversdale): W. Riversdale, Dekenah 90 in Smith 5502 (NBG, Holo.).
dentata: toothed.
Differs in being smaller, to 4cmφ, the leaves very dark green, slightly scabrid and with spined margins. (A var. floribunda foliis subtiliter scabridis et denti-marginatis differt).
This variety describes the smaller form which has distinct and widely spaced marginal spines. The leaves are very dark green and also slightly scabrid. It occurs from the Bontebok Park at Swellendam to northwest of Riversdale, and includes larger forms east of Riversdale also with pronounced marginal spines.
Distribution: 3420 (Bredasdorp): Bontebok Park (-BA), Bayer 3439 (NBG); E. Buffeljachts (-BA), Viviers 156 (NBG). 3421(Riversdale): W. Riversdale (-AA), Dekenah 90 in Smith 5502 (NBG): 5km N. Riversdale (-AB), Smith 5381 (NBG, PRE); 15km E. Riversdale (-AB), Smith 5758 (NBG); Dassieklip (-AC), Venter 92/31 (NBG); Wydersrivier (-BA), Smith 5491, 6781 (NBG), Bayer 2311.
Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Very difficult to find when dry.
Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Showing the more scabrid leaves of this variety.
Haworthia floribunda var. dentata JDV93/56 east of Riversdale. Also with the characteristic rounded leaf-tips.
Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Usually well-hidden and small.
Haworthia floribunda var. dentata JDV93/58 east of Riversdale. A large robust darkest clone in cultivation.
Haworthia floribunda var. dentata JDV93/58 east of Riversdale. In among stones and lichens.
c. var. viridescens var.nov. Type: CAPE-3420 (Swellendam): S. Swellendam (-AB), De Kok (NBG, Holo.).
viridescens: becoming green.
Very green plants with darker coloration at the basal leaf margins. Relatively glabrous and more robust in cultivation. (A var. floribunda habitu robusto et colore viridi vivido ad margines basales foliorum atranti differt).
This is a large robust plant in cultivation and includes two forms, one of which has pointed leaves. The coloration is greener than normal for even cultivated plants of the species.
Distribution: 3420 (Bredasdorp): Below Swellendam Stn. (-AB)., C.A. Smith 2724a (PRE); S. Swellendam (-AB), D.De Kok (NBG).
Haworthia floribunda var. major JDV93/3 south of Swellendam. Plants in the field are quit small.
Haworthia floribunda var. major JDV91/3 south of Swellendam. Leaves often without spines and tips sometimes pointed.
13. H. gracilis V.Poelln., Feddes Repert.Spec.Nov. 27:133(1929). idem. 41:201(1937). idem., Des.Pl.Life 9:90(1937). idem., Feddes Repert.Spec.Nov. H. translucens sensu Bayer :162(1976). Bayer ::56(1982). non Scott :69(1985). Type: Graaff-Reinet, Amalienstein, Willowmore, Stellenbosch. Not preserved. Neotype (designated here): CAPE-3326 (Grahamstown): Hellspoort (-BA), Britten (PRE).
gracilis: graceful.
Rosette stemless. to 6cm φ, proliferous. Leaves 30-40, lanceolate acuminate, incurved with slender short marginal spines, pale greyish-green, upper surface translucent between lines. Inflorescence simple to 200mm. Flowers white.
1982 – Reflection on the first edition of the Handbook showed that there were two main areas of uncertainty. The first of these was the relation of the blue‑green species, i.e. H. semiviva, H. bolusii, H. blackbeardiana and H. cooperi. The second was H. translucens. In the introduction it is stated that H. bolusii, H. cooperi, H. cymbiformis, H. habdomadis and H. decipiens diffuse into the mountains between Uniondale and Port Elizabeth. To these can still be added H. zantneriana, H. xyphiophylla and perhaps even H. arachnoidea, so that forms of each of these species may contribute to this concept of H. translucens. To the present day the composition of H. translucens is thus still very unclear. There are a considerable number of unlikely elements which can only be thrown together until more understanding of this complex problem can be gained. The ssp. tenera occurs in the middle Fish River valley east of Grahamstown. In Von Poellnitz’s type locality, Plutos Vale, both glabrous and haired forms occur in populations within 75 metres of one another. H. cymbiformis var. incurvula is only a little further away. H. gracilis was collected north‑east of Grahamstown at Hellskloof and here the leaves of the plants are longer, more erect, and glabrous. The flower is essentially the same. However, the home of H. translucens is really the Gamtoos Valley and in the first edition it was pointed out how difficult it was to reconcile such a disjunct population. Although not providing an entirely satisfactory explanation, the discovery of the small forms of H. bolusii in the Jansenville area do suggest how this occurs. Small translucent and fairly hairy forms occur in the conglomerates west of Uitenhage. Darker more opaque and less hairy forms occur at Dead Man’s Gulch and Coega to the east. Dr W.R. Branch has collected very large forms from the lower Krom River near Humansdorp. Many different collections are recorded from Loerie in the lower Gamtoos valley to as far west as Uniondale. H. cymbiformis is also distributed across the area and the var. transiens is found at Prince Alfred’s Pass in the extreme west of the distribution range. The similarity of some collections of this species (e.g. at Andrieskraal) to H. cymbiformis var. incurvula are further evidence of some relationship between the Gamtoos and the Fish River species.
1999 – Here some attempt is made to cast a little more light on a confused situation and it has to be admitted that this solution may be inadequate. In the first place it is apparent that the name ‘translucens’ as typified by the illustration in Curtis’ Bot. Mag., is applicable to Haworthia herbacea and not available for this species. In the second place the small dark more opaque forms referred to above are now regarded in this work as the missing H. aristata of Haworth. The name H. gracilis as used here now, is to refer to a wide range of populations extending from Grahamstown westwards to at least Uniondale. Considerable local knowledge will be required to properly evaluate the species as presented here. The recognition of varieties will hopefully present a realistic view of the variables involved and hopefully facilitate communication about these plants. The fact that the var. minima appears to re-occur around Uitenhage is curious. The var. picturata, which has been collected in the Baakens River Valley together with H. cymbiformis, is also odd. Its relationship with H. cymbiformis var. transiens needs to be explored. The var. viridis is also a poorly known entity with an improbable distribution. It is inextricably linked with H. decipiens var. minor. The distributions for the varieties are not very convincing at all, and as is the case elsewhere, they provide only the skeleton of an hypothesis which should be examined in the field. It seems that there is some interaction with both H. cymbiformis and H. cooperi, if not also with H. bolusii var. blackbeardiana. There is strong interaction with H. decipiens. It appears that there is often a transformation H. gracilis var. viridis to H. decipiens var. minor from steep, perhaps shady south-facing cliff to lower lying more horizontal and exposed sites. It will also be obvious from the specimens cited that in two case there is co-occurrence with H. decipiens. There are three major river valleys which need to be better explored. These are the Gamtoos, Swartkops and Sundays. The intervening mountain ranges are a formidable challenge and this is one species in which the varieties are not closely tied to geographical distribution.
a. var. gracilis. This variety is mostly known from northwest of Grahamstown and it is not certain how widely it may occur or with which species it may interact. There are a number of collections from the Eastern Cape which have the same narrow elongate and relatively blunt leaf-tip and the Gladhurst (Adelaide) citation is very close geographically to H. cymbiformis var. obtusa, and to H. cooperi. In the Hankey/Patensie area these are usually somewhat bluish-green and associated therefore with H. cooperi var. gordoniana. It is a difficult distinction to make as the latter may then be an ecotype associated with low-lying level areas where the plants tend to be single, are less proliferous and are withdrawn into the soil. Gerhard Marx collected a larger form from northeast of Grahamstown which is very similar to H. cooperi var. leightonii. At Jeffrey’s Bay it is difficult to decide if the small element there represents H. cooperi var. gordoniana or H. gracilis. Similarly towards Middleton there is a population which contains two elements, one is H. cooperi and the other could be either H. bolusii var. blackbeardiana or H. gracilis. This is an interesting comparison because it raises the possibility that the latter two are continuous.
Distribution: 3226 (Fort Beaufort): Gladhurst, Adelaide (-AC), Krynauw in NBG272/43 (NBG), Venter & Bayer (NBG). 3324(Steytlerville): Paul Sauer Dam (-DA), Swart (NBG); Mistkraal (-DA), Smith 7062 (NBG); Ferndale (-DB), Smith 3672, 3675, 3677, 3678, 6204, 7185 (NBG); 2km E. Hankey (-DD), Bayer 4476 (NBG); Hankey (-DD), Paterson 24 (BOL), Fourcade 3329 (BOL); Gamtoos Heights (-DD), King 90 (BOL); Longmore (-DD), Bayer & Bruyns 6855 (NBG). 3325(Port Elizabeth): Loerie (-CC), Britten (BOL). 3326(Grahamstown): Hellspoort (-AB), Dyer 4 (BOL), Long (BOL), Blackburn in BOL71332, Britten in PRE 34922, in PRE 34929, Smith in NBG326/34, Bayer in KG336/70 (NBG); 5km N. Grahamstown (-AB), Smith in NBG332/34; Howiesonspoort (-AC), James 549 (BOL); 7km NE. Grahamstown (-BA), Bayer & Venter 6603 (NBG); Brakkloof (-BA), Acocks 12046 (PRE); Fish River (-BA), Dyer 580 (PRE).
Inadequately located: Albany, Dyer in NBG1804/30.
Haworthia gracilis var. gracilis JDV89/42 north-west of Grahamstown, The leaves tend to be quite acute and incurved at the tips.
Haworthia gracilis var. gracilis JDV89/42 north-west of Grahamstown. Plants fairly solitary and less robust.
Haworthia gracilis var. gracilis JDV96/4 north-east of Grahamstown. Growing near to H. cooperi var. pilifera. The plants are quite robust and clump forming.
b. var. isabellae (V.Poelln.) Bayer comb.nov. H. isabellae V.Poelln., Feddes Repert.Spec.Nov. 44:226(1938). non Scott :76(1985). Type: Cape, near Port Elizabeth, Mrs I King. Not preserved. Neotype (designated here): CAPE‑3325 (Port Elizabeth): Humansdorp, Gamtoos bridge (‑CC), H. Hall in NBG 68799.
isabellae: for Mrs Isabella King.
This variety is recognised as the more traditional form of the species in which the outer leaves are spreading and the inner leaves more erect. It is very proliferous. The leaves are not as bluish-green, nor as swollen towards the tip as in H. cooperi variants which have often been assigned to this species. The marginal spines are quite dense and relatively long for the narrow-leaves, as compared to H. cooperi var gordoniana which also occurs in the Gamtoos Valley.
Distribution: 3324 (Steytlerville): Dam se Drif (-CA), Rossouw 483 (NBG); Moordenaarskloof (-CD), Stayner (NBG); Ferndale (-DB), Smith 3674 (NBG); Gamtoos (-DD), Stayner in KG367/62 (NBG); Hankey to Humansdorp (-DD), Smith 7267 (NBG); Gamtoos River (-DD), Smith 7439 (NBG); NE. Hankey (-DD), Bayer 4475 (NBG), Stayner (NBG). 3325 (Port Elizabeth): Loerie (-CC), Britten in PRE 34924; Gamtoos Ferry (-CC), Stayner in KG341/62; Vanstaadens to Loerie (-CC), Smith 3112 (NBG); Gamtoos bridge (‑CC), King 92 (BOL), H. Hall in NBG 68799, Smith 7266 (NBG); Longmore Forest (-CC), Bayer & Bruyns 6555 (NBG). 3424 (Humansdorp): Woodlands (-BA), Marais in NBG8/66, Krom River Estuary, Rippon (-BB), Branch 1 (NBG).
Haworthia gracilis var. isabellae WRB 1 Kromme River. The leaves are spined and tend to spread. At this locality the plants are quite large.
Haworthia gracilis var. isabellae JDV92/136 Elandsriver. Plants here can be densely spined. The area is not well explored.
Haworthia gracilis var. isabellae JDV90/71 Gamtoos Bridge. In this area the variety is on steep rocky slopes.
c. var. tenera (V.Pielln.) M.B.Bayer. H. tenera V.Repert. Spec. Nov. 31: 86(1932). Scott: 76 (1985). H. translucens ssp. tenera (V.Poelln.) Bayer :161(1976). Bayer :56 (1982). Type: Cape, Plutosvale, Grahamstown, Miss Blackbeard 15. Not preserved. Neotype (B&M): Glenelg, Smith 5416 (NBG). H. minima Baker, J. Linn. Soc. 18: 215 (1880). Nom. illeg. ype: Cape, imported by Tuck, in hort Kew. Not preserved. Lectotype (designated here): icon (K).
tenera: tender or delicate.
This variety passed without comment in the early editions and Scott is justified in regarding it as discrete against the ill-defined view of the species then presented. It is well-represented in the area northeast and east of Grahamstown and is quite variable. Entirely glabrous forms occur and Smith’s collection from north of the type locality is also markedly less greyish green, and also less prominently spined. Breuer and Metzing have unfortunately chosen this specimen as the type for H. tenera when many specimens from Plutosvale were available.
An additional and important consideration in including this distinct element with H. gracilis, is a range of collections from the Uitenhage area (Groendal in particular) where very similar growth forms occur. These differs from the typical variety in respect of the more compact incurved rosettes and off course, spination. There are forms of var. isabellae which are practically identical. To complicate the issue, it should be noted that H. gracilis var. gracilis also occurs near to Plutosvale, and its relation there with H. cymbiformia var. incurvula is not clear.
Distribution: 3323 (Willowmore): Spreeugat (-DB), Bruyns 1650 (NBG). 3324(Steytlerville): Wilgekloof (-CA), Branch 36 (NBG); Groot Kommando Kloof (-CB), van Jaarsveld 7704 (NBG); Ziewefontein (-CB), Fourcade 5471 (BOL); Kouga (-CB), Esterhuysen 7119 (BOL); Scholtzberg (-CB), van Jaarsveld 7792a (NBG); Ashoek (-DA), Smith 3679, 3680 (NBG); Kaan (-DD), Smith 3686 (NBG); 3325 (Port Elizabeth): Groendal wilderness, Nounek (-CA), Scharf 1061 (PRE); Groendal Dam (-CB), Bayer 1404a, Bruyns 1822 (NBG). 3326 (Grahamstown): Plutosvale (-BA), Britten in PRE 34942, Britten 12 (BOL), Smith 9, 5419 (NBG), Bayer in KG47/72, in KG47/72a (NBG), Dyer in NBG 802/30 (BOL), Dyer 2173 (PRE), Erens 435 (PRE), Hutchinson 1577 (BOL); Top Plutosvale (-BA), Smith 5417; Glenelg (-BA), Smith 5416 (NBG); Near Fletchers Farm (-BA), Smith 5418, 5420 (NBG); S. Hunts Drift (-BB), Smith 5679 (NBG); S. Committees (-BB), Smith 6509 (NBG).
Inadequately located: Uitenhage, Long in NBG1456/35, Britten in NBG741/31, Cook (BOL); Albany, Luyt in NBG305/45; Hankey, Coates in NBG369/39, NBG355/39; Patensie, Smith 3025 (NBG); Port Elizabeth, Arnold (BOL).
Haworthia gracilis var. tenera JDV91/103 Hunts Drift. Usually rounded and a neater plant than the typical variety.
d. var. picturata var.nov. Type: 3325(Port Elizabeth): Enon (-BC), Thode 21507 (NBG, Holo.).
picturata: variegated.
Differs from the type in having glabrous, bright green leaves, in which the translucent areas of the leaves contrast strongly with the dark green opaque reticulation. (A var. gracilis foliis glabris et bene notatis differt).
This variety has generally been overlooked as intermediate between typical H. cymbiformis and the var. transiens. However, there are localities where this variant reportedly grows together with H. cymbiformis, and it appears to be derived primarily from the gracilis element in the greater Baviaanskloof. The leaf-tips are either normally pointed, and thus very similar to the var. isabellae, or may be blunt and incurved. The main distinction is the distinct translucent reticulation contrasting with the opaque lower leaf surfaces.
Distribution: 3323 (Willowmore): 9km E. Haarlem (-CB), Smith 3669 (NBG). 3324 (Steytlerville): Geelhoutboskloof (-CA), Viviers 879 (NBG); Diepriver (-CD), Van Jaarsveld 15342 (NBG); N. Andrieskraal (-DA), Fourcade 176 ((NBG); Kleinwaterkloof (-DA), Smith 7102 (NBG); Grootwaterkloof (-DA), Smith 7103 (NBG); 3.5km W. Hankey (-DD), Stayner (NBG); Kleinrivier road (-DD), Smith 2931 (NBG). 3325(Port Elizabeth): Enon (-BC), Thode A2774 (PRE), Thode 21507, 26090 (NBG); Longmore Forest (-CC), Branch 369 (NBG).
Haworthia gracilis var. picturata JDV90/76 Andrieskraal. The leaf-tips are blunt and there is a strong colour contrast between the translucent and opaque areas of the leaf.
Haworthia gracilis var. picturata JDV90/76 Andrieskraal. Always on rock faces and seldom solitary.
e. var. viridis M.B.Bayer var.nov. Type: CAPE-3325 (Port Elizabeth): Perdepoort (-AC), Smith 6867 (NBG, Holo.).
viridis: green.
Differs from the type in the brighter green coloration and in its more northwesterly geographic distribution. (A var. gracilis foliiis viridibus vividuis et distributione geographica septentionali-occidentali differt).
Occurs to the northwest in the Winterhoek mountains and may have the form typical of the species, with longish, erect and glabrous incurving leaves, or be somewhat squatter with broader incurving leaves. One record from east of Hankey is attributed to this variety, and there are two records from the Steytlerville area where the small form of H. decipiens is also recorded. Field observations show a transition directly to H. decipiens var. minor and also to H. decipiens var. pringlei. As stated earlier, it is very difficult to detect any geographical patterns in the variation of the species and it is probably over-ridden by ecotypic variation in a topographically diverse region.
Distribution: 3324 (Steytlerville): Oulande, Steytlerville (-AA), Schoeman in KG22/84 (NBG); Campherpoort (-AA), Smith 3594, 3646 (NBG), Bruyns 1629 (NBG), Bayer 2074 (NBG); Die Poort (-AD), Branch 353 (NBG); Dorschfontein (-BC), Bayer 3375 (NBG), Bayer & Bruyns 6589 (NBG); Diepnekkloof (-CA), Branch 35 (NBG); Kouga Dam (-DA), Stayner in KG343/62 (NBG); Witrivier (-DA), Bayliss in KG376/75 (NBG); Ouplaas (-DB), Bruyns 7040b (BOL); Hankey to Patensie (-DD), Bosch in KG73/70 (NBG). 3325 (Port Elizabeth): Brakfontein (-AC), Bayer 4198 (NBG); Perdepoort (-AC), Smith 6867 (NBG), Branch 10 (NBG), Schoeman (NBG); E. Perdepoort (-AC), Smith 7337 (NBG), Bayer & Venter 6600 (NBG); Sapkamma (-AC), Bayer & Venter 6620 (NBG); Perdepoort Mt.(-CA), Swart in Bayer 970 (NBG).
Inadequately located: Baviaanskloof, Bayliss in KG379/76 (NBG).
Haworthia gracilis var. viridis MBB970 Paardepoort. Typically with brighter green incurving leaves with no spines. However, the continuity with H. decipiens var. minor has to be remembered.
Haworthia gracilis var. viridis JDV94/20 south of Perdepoort. On rocky slopes like the var. isabellae, but green rather than bluish in colour.
Haworthia gracilis var. viridis JDV96/82 south of Perdepoort. Plants more spinose.
14. Haworthia heidelbergensis Smith, JS.Afr.Bot. 14:42(1948). Bayer :121(1976). Bayer :41(1982). Scott :131(1985). Esterhuizen, Aloe 29:64(1992). idem. 33:15(1996). Type: CAPE‑3420 (Bredasdorp): W. Heidelberg (‑BB), J. Dekenah 230 in Smith 6566 (NBG).
heidelbergensis: from Heidelberg, Cape.
Rosette stemless, proliferous, to 8cm φ. Leaves many, erect to recurved, generally with small marginal and keel spines, usually dark green with reddish hues, end-area of leaf slightly translucent. Inflorescence simple. Flowers 10-15, white with brownish veins.
1982 – This is a small species occurring immediately east of Heidelberg. It grows deeply imbedded in moss and lichen and although the leaves have a distinct end area they are markedly acuminate. Known only from one locality, there is a related population at Matjestoon to the southwest in which the plants have longer recurving leaves, and another at Bredasdorp similar to the last. Together these three populations have no obvious affinities with any other species in the complex. Heidelberg is a very significant area for haworthias, as species such as H. turgida, H. magnifica, H. floribunda and H. retusa all occur nearby. H. heidelbergensis is not continuously variable with any of these. It is a winter‑growing species, invariably non‑proliferous and difficult in cultivation. The occurrence in moss and lichen suggests that it requires more shade than usual, as well as a distinct summer resting period.
1999 – This has proved to be the most surprising species among all the collections of the last ten years. Many collections have been made including at all the compass points immediately around Heidelberg, although those north are really considered hybrid with H. turgida. As Esterhuysen (1992) pointed out, the type locality is probably west of Heidelberg as recorded in Smith’s collecting records and on his type specimen. This is cited in the published description simply as ‘Swellendam District’. It extends further east towards Riversdale, westwards to Bonnievale and is common east and north of Bredasdorp. It has proved to be a major role player among the species in that area apart from its association with H. emelyae already dealt with. The populations around Heidelberg show evidence of direct introgression with H. turgida and cast new light on the relationship of that species with H. retusa. H. heidelbergensis is apparently the eastern counterpart of H. mirabilis and these two species never co-occur. H. serrata also cannot be excluded from discussion of this species (see H. serrata). At its westernmost limit at Bonnievale, it is confounded with H. maraisii var. meiringii. At the type locality it is rather atypical, an oxymoronic statement. Here the leaves are erect, but with a distinct end-area and there are not many leaves. Characteristic of the species is, however, the abundance of individuals at each site and their density in colonies usually on slightly southern slopes and in thick moss. The leaves may be smoothish and cloudily translucent or with translucence between veins on the upper leaf surfaces. The leaves tend to spread and recurve rather than being distinctly retused or with a swollen end-area as in the type. This is also one of the main feature separating H. mirabilis from H. maraisii, although it is by no means an absolute discriminant. The colour is also usually a reddish tinted greenish-brown. At Bredasdorp the population is greener than normal and the plants are like miniaturised H. mirabilis. Several varieties are distinguished to facilitate perception of this species.
a. var. heidelbergensis Resende in Feddes Repert.Spec.Nov. 48:114(1940) suggested a change in the rules of nomenclature so that if a name proved not to be the true stem form (in other words not central to the circumscription of the species) the type should be changed for one that was. This is a case where that rule would prove quite useful. A manuscript concerning the pivotal role this species plays in the Southern Cape has been submitted to ALOE. Despite the major role this species plays in relation to all the species in the Southern Cape, not one of the illustrations in Bayer (1980), ‘The story of Haworthia nitidula’ depicts this. Larger plants occurs around Heidelberg and mid-way to Riversdale. The actual locality of the collection is rather odd, as Dekenah pointed out the locality to the author as east of the town, whereas there are two collection of Smiths received from Dekenah, one east and one west. They are virtually identical and Esterhuizen (1992) confirms this.
Distribution: 3420 (Bredasdorp): W. Heidelberg (‑BB), J. Dekenah 230 in Smith 6566 (NBG); E. Heidelberg (-BB), Smith 5756 (NBG), Bayer 2550 (NBG); Koppie opposite sports grounds (-BB), Smith 5044 (NBG); S. Heidelberg (-BB), Smith 6989 (NBG).
Haworthia heidelbergensis var. heidelbergensis JDV93/54 G=Between Heidelberg and Riversdale. An extensive population where the plants are robust.
Haworthia heidelbergensis var. heidelbergensis JDV87/1 east of Heidelberg. The upper leaf surfaces are always a little translucent.
Haworthia heidelbergensis var. heidelbergensis JDV87/1 east of Heidelberg. Plants are smallish and grow in lichen and moss.
b. var. minor var.nov. Type: CAPE-3420( Bredasdorp): Rooivlei (-CA), Bayer in KG 36/70 (NBG, Holo.).
minor: smaller.
Differs in being small, to 3cm φ, and in being light yellowish-green and well spined. (A var. heidelbergensis foliis parvioribus flavo-virentibus pallidis et spinosis differt).
North of Bredasdorp the plants resemble miniaturised forms of H. mirabilis var. sublineata and have the same plain green colour. The leaf-tips are also notably translucent.
Distribution: 3420 (Bredasdorp): Rooivlei (-CA), Bayer in KG 36/70 (NBG), Smith 3903 (NBG).
Haworthia heidelbergensis var. minor JDV86/5 north of Bredasdorp. Leaves narrower and smaller than the typical variety.
c. var. scabra var.nov. Type: CAPE-3420 (Bredasdorp): Leeurivier (-AB), Bayer 1700 (NBG, Holo.).
scabra: rough.
Differs in being small, to 3cm φ and as tall. The colour is usually very dark green. The leaves erect or sub-erect, slightly scabrid especially along the margins and keel. (A var. heidelbergensis foliis recurvatioribus cum fenestra apicali distincta differt).
This is the commonest variety in terms of population distribution. The plants are quite small and very easily mistaken for erect-leaved forms of H. maraisii despite the general absence of ‘cloudy’ translucent windows in the leaves of the latter species. Although populations are often small, they are usually very dense.
Distribution: 3320 (Montagu): Drew (-CC), Fouche in PRE 34945; W. Bonnievale (-CC), Bayer 6509 (NBG), Lewis in NBG2457/33 (BOL). CAPE-3420 (Bredasdorp): Kliphoogte (-AA), Bayer 4677 (NBG); Leeurivier (-AB), Bayer 1700 (NBG); Brakfontein (-AC), Bayer 2547 (NBG); Beyersdal (-AD), Bayer 2556 (NBG); Haarwegskloof (-AD), Bayer 5101 (NBG); W. Kathoek (-AD), Bayer & Bruyns 6566 (NBG), Burgers 2581 (NBG); E. Kathoek (-AD), Bayer & Bruyns 6561 (NBG); N. Potberg (-BC), Bayer & Bruyns 6544 (NBG), Bayer & Bruyns 6545 (NBG).
Haworthia heidelbergensis var. scabra JDV86/3 north-west of Bredarsdorp. Occurs here in very dense clusters but very localized.
Haworthia heidelbergensis var. scabra JDV92/85 north of Infanta. The leaves are shorter and broader than typical.
Haworthia heidelbergensis var. scabra JDV87/4 east of Drew. Less translucent and the leaves are a little more rigid and scabrid than the other varieties.
Haworthia heidelbergensis var. scabra JDV87/4 east of Drew. This population was severely damaged by cultivation and has survived only in a few remaining small ridges.
d. var. toonensis var.nov. Type: CAPE-3420 (Bredasdorp): Matjestoon (-BB), Smith 6797 (NBG, Holo.).
toonensis: abbreviation for origin at Matjestoon.
Differs in the more recurved leaves with a distinct transparent end-area to the leaves.
15. Haworthia herbacea (Mill.) Stearn, Cactus J. 7:40(1938). Bayer, Nat.Cact.Succ.J 27:51(1972). Bayer :122(1976). Bayer :42(1982). Aloe herbacea Mill., Gardeners Dictionary :n18(1768). Type: icon, 130:t131 Boerhaave Index Alter Pl. (1720). Epitype (B&M): CAPE-3319(Worcester): N. Ribbokkop (-DC), Bayer 161 (NBG): A. atrovirens D.C., Pl.Gras. :f81(1799). H. atrovirens (D.C.) Haw., Revis. :57(1821). Type: as above: A. arachnoidea var. pumila Aiton, Hort.Kewensis 1:468(1789). Willd., Spec.Pl. 2:188(1799). A. pumila (Willd.) Haw., Trans.Linn.Soc. 7:10(1804). H. pumila (Willd.) Duval, Pl.Succ.Hort.Alenc. :7(1809). Haworth, Syn.Pl.Succ. :95(1811). Type: as above (Linnaeus, Aloe pumila var. E): Aloe translucens Haw., Trans.Linn.Soc. 7:10(1804). H. pellucens Haw., Syn.Pl.Succ. :96(1812). Aloe arachnoidea var. translucens (Haw.) Ker-G., Curtis’ Bot.Mag. t.1417(1811). H. translucens Haw., Suppl.Pl.Succ. :52(1819). H. arachnoidea (L.) Duv. sensu Scott, Cact.Succ.J(U.S.) 49:205(1977). Scott, Aloe 16:41(1978). Scott :39(1985). Type: Cape, Masson. Not preserved. Lectotype (designated here): icon. :t.1417 Curtis’ Bot. Mag.: H. pallida Haw., Revis.:56(1821). Type: Not preserved. Lectotype (designated here): icon (K): H. aegrota V.Poelln., Desert Pl.Life 11:193(1939). Type: Cape, Worcester, Swellendam etc. H. Venter. Not preserved. Lectotype (designated here): icon (B): H. submaculata idem. Type: Cape, Worcester, etc. H. Venter 5. Not preserved. Lectotype (designated here): icon (B): H. luteorosea Uitew., Cact.en Vetpl. 5:88(1939). Type: ex Hort, Holland. Not preserved.H. arachnoidea (L.) Duval sensu Scott, Cact. Succ. J. (US) 49:205 (1977).
herbacea: yellow-green.
Rosette stemless, proliferating, to 8cm φ. Leaves erect, incurved, scabrid, margins and keel with firm spines, greenish yellow in colour, reticulate patterning with translucent interstices. Inflorescence simple, to 300mm. Flowers large, beige, pinkish tips, buds bi-arcuate.
1982 – The choice of the name herbacea is possibly a poor one as this name is associated with a number of perhaps incompatible early illustrations. Scott’s selection of the name arachnoidea is equally dubious particularly as it also ignores the flower character. H. herbacea occurs only in the Worcester area extending in the southeast to east of McGregor, south into the mountains at Villiersdorp where a large flowered form occurs and to just northwest of Worcester itself. It is very close to H. reticulata and southwest of Robertson it is often difficult to know with which species one is dealing. However, H. herbacea usually has beige flowers, the plants are usually solitary, more deeply sunk in the ground, less proliferous and more hairy than H. reticulata. At De Wet the two species grow together and there are distinct hybrids. There are at least two other localities where both species grow in close association without hybridisation. H. herbacea flowers in late winter (September‑October) and has an unusually large flower. The Villiersdorp mountain form collected by E. Esterhuysen, has a particularly impressive flower but has not been introduced into cultivation. H. herbacea has a direct connection with H. maculata at the Brandvlei Dam south of Worcester. The size of the plant varies from 80mm in diameter to as little as 35mm near McGregor where the flowers may also be pinkish.
1999 – Nomenclature is indeed a slippery slope and this is a case in point. The name pumila which Duval applied in Haworthia, was based on Boerhaave’s early illustration as can be traced through a tortuous synonymy. It is in my opinion now invalid in the genus Haworthia, because the same type is used for the name herbacea. (see also Haworthia margaritifera). Apart from this nomenclatural tangle, the application of the name to a naturally occurring element is a problem. The Boerhaave illustration can perhaps be better related to Haworthia maraisii than to the species for which I have used the name. The earlier epithet atrovirens would have been a far better choice for that species had it been available. Rather than make this unfortunate change, and then also using the name translucens for this species, I am following a principle of conservation and letting sleeping dogs lie. It is enough that Col Scott has unfortunately confounded the issue by his use of the name H. arachnoidea for this species, without the acknowledgment in synonymy that may have clarified the issue.
The type nominated by Breuer and Metzing for H. translucens is based on the 1982 Handbook interpretation, where the name was applied incorrectly. It is apparent that H. translucens Haw. as perceived in 1819, includes the Botanical Magazine illustration cited above. This must be related to H. herbacea as understood here now.
a. var. herbacea. The typical variety is widespread and relatively homogeneous throughout its range.
Distribution: 3319 (Worcester): Karoo Garden (-CB), Dobay 62 (NBG), Barker 8766 (NBG); Veld Reserve (-CB)., Olivier 129 (NBG, PRE), Pamphlet 44 (NBG); W. Worcester (-CB), Searle in NBG162/84 (NBG), Bayer 2421 (NBG); Brandwacht (-CB), Bayer in KG166/70 (NBG); SE. Brandvlei Dam (-CB), Bayer 2422 (NBG); W. Doornrivier (-CD), Bayer 1995 (NBG), Leipoldt (BOL); Lemoenpoort (-CD), Bayer 1996(NBG); N. Lemoenpoort (-CD), Bayer 4439 (NBG); Patryskloof, Mauve & Oliver 243 (NBG); De Wet (-DA), van Heerden 1024 (NBG); Rabiesberg (-DA), Esterhuysen & Lewis in NBG2689/35 (NBG); Ribbokkop (-DC), Bayer 161 (NBG); Keerweerder, Jonaskop (-DC), Bayer 2697 (NBG); Nuy (-DC), Hurling & Neil (BOL); Mowers (-DC), Bayer in KG218/70 (NBG); Wansbek (-DC), Bayer 1997 (NBG), Moffett in KG181/70; Rabiesdal (-DD), Heunis 1 (NBG); Koningsrivier (-DD), Bayer in KG 329/70 (NBG).
Inadequately located: Marloth 8912 (PRE); Worcester, Smith 3910 (NBG), Hurling & Neil (BOL), Venter 4 (BOL), Joubert 1 (BOL); Worcester to Robertson, Venter 5, 7 (BOL); ex hort, Leipoldt (BOL), Logan (BOL), Starke in BOL24591, Venter in NBG215/39, Aryer’s nursery (BOL); Ross-Frames in NBG78/44; Bonnievale, Malherbe in NBG297/40, van der Merwe 174 (BOL); Riversdale, Ferguson (BOL).
Haworthia herbacea var. herbacea JDV84/5 Ribbokkop. Grows, and hybridises with H. reticulata at this site.
Haworthia herbacea var. herbacea JDV85/60 Bosfontein. The spination of the plants in this area is more pronounced.
Haworthia herbacea var. herbacea JDV85/12 Sandberg. Here it is growing in quartzitic rock.
b. var. flaccida var.nov. Type: CAPE-3319 (Worcester): Rooiberg (-DD), P.V.Bruyns 7114 (NBG, Holo.).
flaccidus: weak, soft.
Differs from the species in being small and delicate. There is considerable variation among the plants in habitat. (A var. herbacea foliis parvioribus et virellis differt).
The habitat of this variety is on steep north-facing rocks where the plants occur in small very dense clusters. They co-occur with H. maraisii and with H. reticulata. The former is on the same rock face but not as exposed, while the latter occurs on slopes both lower down and higher up on the mountainh. The locality is outside the known northeast distribution of the species. There are very few plants and the habitat is uncharacteristic of any of the species in the Southern Cape. The flower is outwardly identical to that of H. herbacea as is also that of H. pubescens. The flowering time co-incides with that of the latter but there is very little vegetative similarity.
Haworthia herbacea var. flaccida JDV96/31 Rooiberg. Still very poorly known from only four clones.
Haworthia herbacea var. flaccida JDV96/31 Rooiberg. This clone more closely resembles the typical variety.
c. var. lupula var.nov. Type: CAPE-3319 (Worcester): Boscheveld Mt., Wolfkloof, Villiersdorp (-CD), E. Esterhuysen (NBG, Holo.).
lupula: small wolf as reference to origin.
Distinctive for the larger pink flower, and for the broader, shorter leaves which are more finely flecked and slightly less scabrid than the typical.
This variety was first collected by Elsie Esterhuysen who specialised in the flora of the Cape mountains. It occurs in sandstones, which is not unique for this species as it is also in sandstones south of McGregor. The occurrence is at the southwestern limits of the species. This is one of the elements which recurs through the Cape mountains and which suggest a common ancestry from here for at least the subgenus Haworthia. The similarities to H. vlokii, H. mirabilis var. consanguinea, H. maculata, H. variegata var. modesta (on the Potberg Mountain) and to H. turgida at high altitude are notable.
Haworthia herbacea var. lupula JDV94/36 Villersdorp. A surprising occurrence in sandstone with Fynbos vegetation.
Haworthia herbacea var. lupula JDV94/36 Villiersdorp. The flowers of this variety are surprisingly large and pinkish.
d. var. paynei (V.Poelln.) Bayer comb.nov. H. paynei V.Poelln., Feddes Repert.Spec.Nov. 41:206(1937). H. pallida var. paynei V.Poelln., Cactus J 6:19(1937). Type: Cape, McGregor district, G. Payne. Not preserved. Neotype (designated here): CAPE-3319 (Worcester): Olifantsdoorn, McGregor (-DD), Bayer 4430 (NBG).
paynei: for G. Payne.
Mr George Payne accumulated an extraordinary collection of plants prior to the war and subsequently lost interest. This variety named after him is indeed different for its small size and for the bi-coloured flower which is pink above and white below. It is on the southeastern limits of the species and a still smaller proliferous form has been reported in the area.
Distribution: 3319 (Worcester): McGregor (-DD), Payne in PRE 27254, in PRE 34948, Bayer 695/69 (NBG); 1km S. McGregor (-DD), Bayer (NBG); Near McGregor (-DD), Smith 3978, 7196 (NBG); 3km E. McGregor (-DD), Scott 2203 (PRE); Olifantsdoorn, McGregor (-DD) Bayer 4430 (NBG).
Haworthia herbacea var. paynei JDV96/41 McGregor. A smaller plant and the flowers have pinkish upper lobes with traces of green, whereas the lower lobes are whiter
16. Haworthia lockwoodii Archibald, Flower.Pl.Afr. 20:f792(1940). Bayer :129(1976). Bayer : 43(1982). Scott :86(1985). Type: Cape, near Laingsburg (-BC), S. Lockwood-Hill 215 (GRA).
lockwoodii: after S. Lockwood‑Hill.
Rosette stemless, to 10cm φ, slowly proliferous, withdrawn into soil. Leaves many, incurved, broad, smooth, usually spineless and dying back at the tips, pale green and translucent above. Inflorescence simple, robust. Flowers many, large adpressed to stem and broad across upper surface.
1982 – This species is at its most attractive in the field when the dead, whitened leaf tips are closed in a tight umbrella‑like canopy over the plant. The plants then resemble dried‑off onions. They grow usually very well hidden by stones or under bushes. H. lockwoodii is relatively restricted to the area south of Laingsburg and its affinity lies most probably with H. decipiens. It differs in that the leaf margins are entirely smooth and in that the leaf tips die back so characteristically. A relationship with H. habdomadis is frequently suggested but H. lockwoodii always has a thinner and broader leaf. In cultivation it tends to etiolate and the leaves can be seen to be a clear pale‑green with the leaf tips without veins or chlorophyll. The Laingsburg area is in the summer rainfall zone but H. lockwoodii should receive very little water and preferably in the winter.
1999 – The alliance of this species is with H. decipiens as tentatively stated above. At Prince Albert this is confirmed by several populations which are intermediate. However, southeast of Laingsburg both species apparently interact with H. mucronata. This is at a farm Rouxpos. H. lockwoodii and H mucronata co-occur discretely in the Rooinek Pass, but to the south-west at Anysberg, seem to hybridise in an extensive population. This interaction should be seen in the light of the newer discussion concerning species concepts. These interactions are not in the context of ‘new’ contact, which is a notion the previous handbooks attempted to reject. In the application of chaos theory to species recognition, it is entirely to be expected that transitional or undifferentiated populations should occur. The Rouxpos populations seems to show that there is simply a continuum of the major elements such as H. lockwoodii and H. mucronata in the west to H. cooperi and H. cymbiformis in the east.
Distribution: 3320 (Montagu): W. Klein Riet River, Waterford (-BB), Wilmot & Hill 215 (PRE); Laingsburg (-BB), Wilmot in PRE 34946 (PRE); near Laingsburg (-BB), S. Lockwood-Hill 215 (BOL,GRA), Fourcade in Archibald 1459 (NBG), Otzen in Smith 3538 (NBG), Smith 3406 (NBG), Malherbe in NBG653/41; Ezelsfontein (-BB), Bayer 2425 (NBG); Floriskraal (-BB), Bayer 155 (NBG); 16km S. Laingsburg (-BB), Minnaar (BOL). 3321(Ladismith): W. Rouxpos (-AC), Venter 31 (NBG), Venter 86/96 (NBG).
Inadequately located: ex hort Whitehill NBG 68384; James (BOL), Archer (BOL).
Haworthia Lockwoodii JDV95/32 south of Laingsburg. The dead leaf-tips are very white and cover the plant completely in the dormant season.
Haworthia Lockwoodii JDV90/88 south-east of Laingsburg. The leaves are flattish and are not keeled.
Haworthia Lockwoodii JDV95/32 south of Laingsburg. Despite having such notable projection from the sun, the plants nevertheless are invariably hidden among stones or under bushes,
17. Haworthia maculata (V.Poelln.) Bayer :130(1976). Bayer :43(1982). H. schuldtiana var. maculata V.Poelln., Feddes Repert.Spec.Nov.49:25(1940). Type: Cape, Worcester, Swellendam etc. H. Venter 6a. Not preserved. Lectotype (B&M): Worcester, Venter 6 (BOL).
maculata: spotted.
Rosette stemless, proliferous, to 8cm φ. Leaves many, sub-erect to spreading, purplish-green, spotted, short spines on margins and keel. Inflorescence simple, slender. Flowers 15-20, few open, white, yellowish in throat, green veined.
1982 – The original locality for this species was not accurately recorded but it was collected by Major H. Venter, and so it is possible from Smith’s and Long’s records to trace it to the Brandvlei Dam south of Worcester. H. maculata flowers in October/November and the form of the flower is like that of H. herbacea and H. reticulata, rather than like that of H. magnifica (H. schuldtiana). There is a clear intergradation with H. herbacea in the area, and the plants are very similar to H. herbacea except that there are fewer and more turgid leaves. H. maculata also occurs further south in a similar quartzitic rock formation to that at Brandvlei Dam. A problem is the occurrence of populations apparently of H. maculata in the mountains to the north of Worcester (both high altitude ‑ Audenberg Peak, and low altitude ‑ at Sandhills). A similar element occurs eastward towards Robertson at Buitenstekloof, distinguished again from H. magnifica by an earlier flowering time and the wide spread of the tips of the upper perianth lobes.
1999 – It is difficult to know just what is present on the higher mountains. Few succulentophiles are also mountaineers, and besides the plants could be expected to be on rocky north faces which may not attract the conventional high altitude botanist. H. nortieri bears some resemblance to H. maculata and that species is found as far south as Opdieberg (Ceres). It is quite probable that populations may occur elsewhere in the area between Worcester and Citrusdal.
a. var. maculata This variety seems to be linked to H. herbacea with possible ties to H. nortieri. The similarity to other high mountain forms (eg H. vlokii and H. turgida) cannot be overlooked and this has been repeated at several places in this book.
Distribution: 3319 (Worcester): Brandvlei Dam (‑CB), Bayer 164 (NBG), Smith 3912 (NBG); Bayer in KG669/69 (NBG); NE. Brandvlei Dam (-CB), Bayer 2591 (NBG); Audensberg Peak (-CB), Esterhuysen 16706 (BOL), Bayer 1119 (NBG); Moddergat (-CD), Bayer 1145 (NBG); Keeromsberg, Boskloof (-DA), BOL28719; S. Sandhills (-DA), Bayer 1120 (NBG).
Haworthia maculata var. maculata JDV88/29 east of Brandvlei. Growing in quartzitic stones, the plants become very cryptic
Haworthia maculata var. maculata JDV86/46 Brandvlei. The plants usually have a deep purplish hue and are very cryptic.
Haworthia maculata var. maculata JDV86/47 Brandvlei. Each localized population is a little different from its neighbours.
Haworthia maculata var. maculata JDV88/29 east of Brandvlei. At the eastern end of the range the plants form clusters and turn quite bright red during the dry hot summer.
b. var. intermedia (V.Poelln.) Bayer stat.nov. H. intermedia V.Poelln., Kakteenkunde 9:133(1937). V.Poell., Feddes Repert.Spec.Nov. 44:233(1938). Type: Cape, Robertson, McGregor, G.J. Payne Not preserved. Lectotype (designated here): Epitype (designated here): CAPE-3319 (Worcester): Buitenstekloof (-DC), Bayer 4461 (NBG).
intermedia: between.
In the case of this variety, Payne (priv. comm.) did indicate the actual origin at Buitenstekloof west of Robertson. Von Poellnitz’ later citation for Scottburgh, Port Elizabeth, as well, is indicative of the close resemblance of even very different species and the difficulties which arise in trying to identify them consistently and correctly. In his discussion von Poellnitz concluded that while the plants had the long end-awn of H. mucronata, the reticulated patterning of the leaves was that of the H. reticulata group. The name suggests the difficulty in deciding just what to do with this element. It co-occurs with H. reticulata and with H. arachnoidea and bears a very close resemblance to the shale form of H. maraisii var. notabilis. As already noted it has a different flower and flowering time to that variety. Nevertheless it may be correct to place them together in one species as there is also a population recorded mid-way between the two at Agtervink. Possibly a more direct link with H. turgida should be sought as the plants do bear a close resemblance to the montane forms of that species. Certainly it is possible that there may be a connection somewhere in the mountains between Robertson and Swellendam.
Haworthia maculata var. intermedia JDV86/108 Buitenstekloof. The plants grow amongst Restionaceae. There is a close connection with H. maraisii var. notabilis.
Haworthia maculata var. intermedia JDV86/108 Buitenstekloof. The leaves are softer, more slender and upright than in the typical variety.
18. Haworthia magnifica V.Poelln., Feddes Repert.Spec.Nov. 33:239(1933). Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). H. maraisii var. magnifica (V.Poelln.) Bayer :131(1976). Type: Cape, Riversdale commonage, Mrs E. Ferguson. Not preserved. Lectotype (B&M): Riversdale, Ferguson (BOL).
magnifica: magnificent.
Rosette stemless, slowly proliferous, to 8 cm φ. Leaves spreading, retused to ground level, dark green to purplish, scabrid to finely spined margins, end-area slightly translucent between the veins, surfaces with small slightly raised tubercles. Inflorescence slender, to 40cm. Flowers brownish veined, few open, upper lobes pinched at tips.
1982 – Haworthia magnifica is an incredibly variable species and it will not be possible to gain any insight into probable affinities with H. turgida, H. retusa, H. emelyae and H. mirabilis without in‑depth study. It is characterized by its dark‑green colour, small size, short green‑lined buds and flowers, and ‘fish‑tail’ bud tips. It flowers in March ‑ April although this varies and only serves as a character to distinguish the species from H. mirabilis. Only the more notable and widely spread forms are recognized as varieties here. There are several other populations which could justly be named as varieties. The variety major has an unusually large flower similar to that of H. emelyae and there is evidence of intergradation of these two taxa. The var. paradoxa may constitute a south‑eastern link between H. magnifica and H. mirabilis, except that truer forms of H. magnifica var. maraisii intercede along the Breede River and at Bredasdorp. The var. maraisii at its type locality at Stormsvlei is quite a robust form which adopts many guises in the Robertson/Worcester area. South of Stormsvlei there is an aberrant population which appears intermediate between the var maraisii and H. mirabilis. However, just north of Stormsvlei, H. mirabilis occurs in a small, many‑leaved form. The var. meiringii appears vegetatively very like a smaller, darker green version of H. herbacea, until it flowers. Also to the west it intergrades into the more characteristic retuse‑leaved var. maraisii. The var. notabilis also has erect leaves which are darker green and more turgid than in the case of H. herbacea. The var. atrofusca was originally collected from west of Riversdale and is characterized by its rounded leaf tips. The var. magnifica has unusually long acuminate leaf tips and is restricted to a small area southeast of Riversdale, recurring again just west of Heidelberg. H. magnifica has been recorded north of both Montagu and Barrydale and as far south as Cape Infanta and westwards to Bredasdorp. Consideration of the variability of this species and distribution of variants is very helpful towards understanding variability in the genus as a whole.
1996 – The comment ‘in-depth study’ is quite inappropriate in the Haworthia literature which generally suffers from any formal objective study. What has happened since 1982 is much further collecting which has brought some new perceptions. The problems with this species are exemplified by Scott’s circumscription and synonymy of H. asperula where it is barely possible to separate all the diverse elements included in it. Esterhuizen wrote in ALOE (1996) that it was easier for him to separate the var. atrofusca from var. maraisii, than from magnifica. This perspicacious observation is all the more so for the new collections from east of Riversdale, and also for the changed view of H. heidelbergensis. It has been decided to separate the elements maraisii and magnifica because it appears that they relate to each other as H. heidelbergensis does to H. mirabilis. It is also more realistic that some of the varieties that were with magnifica are rather variants of H. maraisii. Thus H. magnifica is applied to the eastern populations which are rather more robust and in which the end-area of the leaf is longer and more pointed. If the end-area is shortened it is also rounded. H. magnifica is usually more greenish than the nearly black H. maraisii. Where previously considering H. retusa as a major element has led to problems, the perception that H. turgida and H. magnifica are the main role players makes it easier to understand what the different populations may represent. This should be apparent from the varieties recognised here, and in the corresponding discussion.
a. var. magnifica. Originally from only south of Riversdale. The name is now also applied to the population from south of the Tradouw Pass, and to that just east of Riversdale. These are quite robust plants with rather scabrid, sub-tuberculate leaves with denticulation of the margins at least. The surfaces may also be slightly scabrid with the tubercles bearing small spines. The plants are lighter green in colour than H. maraisii.
Distribution: 3420 (Bredasdorp): Tradouw Pass (-BA), Smith 6788 (NBG). 3421 (Riversdale): S. Riversdale at beacon (-AA), Smith 5372 (NBG), Bayer in KG83/71 (NBG); Reserve (‑AB), J. Dekenah 16 (NBG); E. Riversdale (-AB), Smith 5376, 5376a, 5748 (NBG), Bayer in KG92/71 (NBG); Riversdale (-AB), Dekenah 6a (PRE); (-AB), Smith 5372 (PRE); Riversdale (-AB), Muir 3553 (PRE).
Inadequately located: Riversdale, Smith 3900, 5057 (NBG).
Haworthia magnifica var. magnifica JDV93/35 Tradouw Pass. The plants here are very variable and usually much greener than typical.
Haworthia magnifica var. magnifica JDV93/13 south-east of Riverdale. Characteristic of these plants are the very dark colour and acuminate, retused leaf-tips.
Haworthia magnifica var. magnifica JDV97/131 Also north-east of Riverdale. Two adjacent populations which show translation to H. retusa.
Haworthia magnifica var. magnifica JDV94/6 north-east of Riversdale is very similar to plants at Tradouws Pass.
Haworthia magnifica var. magnifica JDV97/39 Barrydale. North of the Langeberg, these plants are more like H. maraisii. However, it seems probable that they will be shown to link up with H. emelyae var. major along the northern slopes of this range.
b. var. acuminata comb.nov. H. retusa forma acuminata Bayer :94(1976). H. retusa var. acuminata Bayer :53(1982). Type: CAPE‑3421 (Riversdale): N. of Gouritzmond (‑BD), Bayer in KG 311/7 (NBG).
acuminata: sharp pointed.
Previously under H. retusa, this variety has been transferred here because of the now restricted view of that species, and the new concept of H. magnifica. It is only known from the one locality as the original nearby locality appears to have been destroyed.
Distribution: 3421(Riversdale): N. of Gouritzmond (‑BD), Bayer in KG 311/7 (NBG), Bayer 2423 (NBG); S. Gouritz Bridge (-BD), Smith 5047 (NBG); 9.5km Gouritz to Albertskraal (-BD), Smith 3946 (NBG).
Haworthia magnifica var. acuminata JDV94/94 north of Gouritzmond. The leaf-tips are much more acuminate than in H. retusa, and the plants are darker green.
Haworthia magnifica var. acuminata JDV91/39 north of Gouritzmond. Plants are proliferous.
c.var. atrofusca (Smith) Bayer Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). H. atrofusca Smith, JS.Afr.Bot. 14:41(1948). Bayer :100(1976). Scott :130(1985). Type: CAPE‑3421 (Riversdale): (‑AA), J. Dekenah 225 in Smith 6169 (NBG).
atrofusca: very dark brown.
This variety was represented by a single small population to the west of Riversdale and characterised by the blunt rounded leaf-tips. This is in effect a single character which occurs in other populations and other species. The scope of the variety is widened to include the large element to the north and west of Riversdale collected mostly by C. Craib (unpublished). These are large brownish-green to blackish plants which are densely and finely tubercled against a scarcely translucent background. The original variety and populations to the west include forms with sharply pointed leaves. There is a very interesting population in the Potberg area to the west of the Breede River. The habitat is identical to that of the type locality. It is very unusual to encounter such a vicariant distribution record in the genus where isolated plants so closely resemble the type.
Distribution: 3420 (Bredasdorp): NW. Kathoek (-AD), Bayer & Bruyns 6549 (NBG). 3421(Riversdale): (‑AA), J. Dekenah 225 in Smith 6169 (NBG), Bayer in KG202/70 (NBG); Droerivier (-AA), Bayer 2665 (NBG).
Haworthia magnifica var. acuminata JDV90/91 west of Riversdale. Leaf-tips rounded.
Haworthia magnifica var. acuminata JDV97/146 west of Riversdale. Leaf-tips rounded. The ‘aubergine’ locality of C. Craib.
Haworthia magnifica var. atrofusca JDV90/91 west of Riversdale. The dark coloured species seem to need more protection from direct sunlight.
Haworthia magnifica var. acuminata JDV92/97 north-west of Riversdale. More robust plants with even more rounded leaf-tips.
d. var. dekenahii (Smith) Bayer comb.nov. H. dekenahii Smith, JS.Afr.Bot. 10:140(1944). H. retusa var. dekenahii (Smith) Bayer :53(1982). Type: Cape, on farm Draaihoek (-BA), J. Dekenah 86 in Smith 5489 (NBG).
dekenahii: for Japie Dekenah, a born naturalist.
This element as a species was completely discarded in the 1976 handbook and resurrected as a variety of H. retusa in 1982. Col. Scott upholds it as a species but illustrates H. turgida var. pallidifolia which co-occurs with it. The significant things about this variety are the raised tubercles on the leaves, the silver flecks, and the blunt rounded leaf-tips.
Distribution: 3421 (Riversdale): Draaihoek (-BA), J. Dekenah 86 in Smith 5489 (BOL,NBG,PRE).
Haworthia magnifica var. dekenahii JDV87/146 north-west of Albertinia. Plants are fairly scabrid, darkish-green and the leaf-tips are rounded.
Haworthia magnifica var. dekenahii JDV87/146 north-west of Albertinia. Plants do not proliferate and can be very cryptic.
e. var. splendens Hammer and Venter Cact.Succ.J(U.S.) in ms. Type: W. Albertinia (-BA), Venter (NBG).
splendens: splendid.
The name is indeed apt. I first saw this plant in about 1970 in a visit to Dr Hans Herre. I simply assumed the unlabelled pinkish-red plant with the shiny black raised tubercles was an unusual specimen of H. emelyae which itself was practically unknown to me at that time. The full story of this variety is better told by the authors whose persistence and tenacity led to its rediscovery at what is presumed to be a second locality – the first apparently having been destroyed. Strangely enough a further population was discovered fatefully, and perhaps fortuitously, by Mary Parisi and Ed Dunne to the east of Albertinia. The word fatefully are used with some deliberation because these two people deliberately avoided the contamination of plans aforethought in wanting to do their own exploration and discovery. This eastern population flowers earlier together with the var. acuminata and also with H. emelyae. There is obviously some significance to this which implies some cross-mountain connection and throws some doubt on the relation of the H. emelyae varieties.
Distribution: 3421 (Riversdale): E. Albertinia (-BA), Marx sn. (NBG); W. Albertinia (-BA), Venter (NBG).
Haworthia magnifica var. splendens JDV97/127 Albertinia. Spectacular glazed pieces of pottery.
Haworthia magnifica var. splendens JDV93/59 west of Albertinia. The tubercles on the leaves can be very dark coloured.
Haworthia magnifica var. splendens JDV97/127 Albertinia. Possibly this is the spot where Herre first collected this variety.
Haworthia magnifica var. splendens Kent sn. Originally collected by Dr. Hans Herre from west of Albertinia.
Haworthia magnifica var. splendens JDV93/59 west of Albertinia. Rediscovered by Venter, Kent and Marx. Flowers here in late summer.
Haworthia magnifica var. splendens JDV94/108 east of Albertinia. The leaf surface in this population are very mottled. Flowering time is in the spring although it seems as though the plants may also flower in late summer in cultivation.
Haworthia magnifica var. splendens JDV94/108 east of Albertinia. Plants from here have more acuminate leaves than in plants from west of Albertinia; also the leaf-tips are densely white-spotted.
19. Haworthia maraisii V.Poelln., Feddes Repert.Spec.Nov. 38:194(1935). idem. 43:104(1938). Bayer :141(1976). H. magnifica var. maraisii (V.Poelln.) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :44,106(1982). Type: Cape, Swellendam, Marais in Swellendam 6410. Lectotype (Bayer, 1976): Icon (B): H. schuldtiana V.Poelln., Feddes Repert.Spec.Nov. 41:211(1937). idem. 43:102(1938). idem. 49:23(1940). Type: Cape, Robertson, SW. McGregor G.J. Payne in Triebn. 903. Lectotype (Bayer, 1976): icon (B): H. schuldtiana var. robertsonensis idem. 49:25(1940). V.Poelln., Des.Pl.Life 9:101(1937). Type: Cape, Robertson, G.J. Payne in Triebn. 991. Not preserved: H. schuldtiana var. minor Triebn. et V.Poelln., Feddes Repert.Spec.Nov. 49:25(1940). Type: Cape, McGregor, G.J. Payne in Triebn. 1096. Not preserved: H. schuldtiana var. subtuberculata V.Poelln. ibid. 49:26(1940). Type: Cape, N. McGregor, G.J. Payne in Triebn. 1089. Not preserved: H. whitesloaneana V.Poelln., Desert.Pl.Life 9:102(1937). idem Feddes Repert.Spec.Nov. 43:107(1938). H. schuldtiana var. whitesloaneana V.Poelln., Feddes Repert.Spec.Nov. 49:26(1940). Type: Cape, McGregor, G.J. Payne in Triebn. 1021. Not preserved. Lectotype (Bayer, 1976): icon (B): H. schuldtiana var. sublaevis idem. 49:26(1940). Type: Cape, loc. unknown, Beukman in Long 690. Not preserved: H. schuldtiana var. simplicior idem. 49:26(1940). Type: Cape, Malgas, G.J. Payne in Triebn. 1112. Not preserved. H. schuldtiana var. unilineata idem. 49:26(1940). Type: Cape, N. McGregor, G.J. Payne in Triebn. 1089. Not preserved. Neotype (designated here): McGregor (-DD), Payne in PRE34897: H. sublimpidula V.Poelln., Cactus J 5:33(1936). idem. Feddes Repert.Spec.Nov. ibid. 41:212(1937). idem. 43:105(1938). idem. Beitr.Zukk.Pfl. 1:45(1939). Type: Cape, Swellendam, Hurling in Triebn. 847. Not preserved. Lectotype (Bayer, 1976): icon.B: H. triebneriana var. diversicolor Triebn. et V.Poelln. ibid. 47:9(1939). Type: Cape, Olifantsdoorn, McGregor, G.J. Payne in Triebn. 1092. Not preserved. Neotype: CAPE-3319 (Worcester): Olifantsdoorn Kloof (-DD), Payne in PRE34881: H. angustifolia var. subfalcata V.Poelln. Sukkulentenkunde 4(1951), nom. inval.
maraisii: for W.R.B. Marais.
Rosette stemless, slowly proliferous, 4-7cm φ. Leaves few to many, very dark green, opaque, usually retused, scabrid with small raised tubercles, tubercles occasionally spined, margins and keel with small spines. Inflorescence slender, to 30cm. Flowers outer upper lobes pinched, frequently yellow throated.
This species was previously treated under H. magnifica and it is now taken out as the smaller darker range of populations from Heidelberg westwards. It is a very common element which is seldom abundant at any one locality. It co-occurs with H. heidelbergensis, H. herbacea, H. reticulata, H. turgida, and H. mutica of the same sub-genus. It is found close to H. mirabilis but it does not co-occur with this species and there are two populations known which appear to be intermediate.
Breuer and Metzing note that a lectotype for H. maraisii was designated by Bayer (1982), when in fact a Berlin-Dahlem illustration was regarded as a type by virtue of the non-preservation of anything else. It is highly unlikely that there was any other preserved material. They nominate presumably the same illustration as a lectotype. Then they state that this illustrations is not original material in the sense of the code, but also that a lectotype must be chosen from the original material.
a. var. maraisii. The variety is typified by a rather robust form at Stormsvlei whereas it is generally a little smaller elsewhere. Note can be made of forms near Robertson which are rather similar to H. pubescens, forms near Bonnievale in which the leaves are rather guttate (spotted), and to near Eilandia which have short erect leaves. The Bonnievale plants are particularly difficult because of a number of populations which are uncharacteristic and vary from the recognisable H. maraisii var. meiringii to a small form which co-occurs with the typical variety. Even the possibility of interaction with H. heidelbergensis cannot be excluded.
Distribution: 3319 (Worcester): Trappieskraalkloof (-DC), Bayer 1210 (NBG); Langkloof (-DC), Bayer 1215 (NBG), Moffett (NBG); Dublin (-DC), Stayner in KG400/61 (NBG); S. Goudmyn Bridge (-DD), Bayer in KG163/70 (NBG); Goudmyn (-DD), Bayer 1216 (NBG); 18km Robertson to Bonnievale (-DD), Bayer in KG46/70 Langvlei (-DD), Scott 2211 (PRE); 5km E. McGregor (-DD), Payne & Scott 22 (PRE); 1.5km S. Robertson (-DD), Scott 2210 (PRE); Muiskraalkop (-DD), Hurling & Neil (BOL), Bayer 1707 (NBG); McGregor (-DD), Payne in PRE34883, Smith 3977, 5606, 5765, 5774 (NBG), Triebn. 1089 in Smith 5767 (NBG); W. McGregor (-DD), Bayer 4437 (NBG); McGregor (-DD), Payne in PRE34897; Vrolijkheid (-DD), UPE 3207 (PRE); SE. McGregor (-DD), Smith 3975 (NBG); S. McGregor (-DD), Bayer 1222 (NBG); Olifantsdoorn Kloof (-DD), Payne in PRE34881, Smith 5774 (NBG); Houtbaai Kloof (-DD), Payne in PRE 34887, Smith 5766 (NBG); Bayer & Stayner 2271 (NBG); W. Robertson (-DD), Bayer 1211, 1703, in KG628/69 (NBG); Bonnievale to Robertson (-DD), Smith 3980 (NBG); Skurweberg (-DD),Bayer 1221 (NBG); SW. Robertson (-DD), Smith 3987 (NBG), Bayer in KG688/69 (NBG); 9km W. Robertson (-DD), Bayer in KG630/69, in KG345/71 (NBG); Klaasvoogds (-DD), Smith 398, 2836 (NBG), Bayer 1220 (NBG). 3320 (Montagu): Dobbelaarskloof (-CB), Bruyns (NBG); N. Ashton (-CC), Bayer 1708 (NBG); Goedverwacht (-CC), Bayer 2177 (NBG); N. Drew (-CC), Bayer 1219 (NBG); Drew (–CC), De Kok 295 (NBG); Cogmanskloof (-CC), Littlewood in KG520/60 (NBG); 6km N. Drew (-CC), Smith 5615 (NBG); Bonnievale (-CC), Marloth 14186 (PRE); Barrydale (-DC), Smith 7353 (NBG), Bolus (BOL), Hurling & Neil (BOL). 3419 (Caledon): N. Napier (-BD), Venter 3 (NBG). 3420 (Bredasdorp): Stormsvlei (-AA), Smith 2700, 2367, 5158, 5641 (NBG); 32km Swellendam to Caledon (-AA), Smith 3250, 3251 (NBG); 7km N. Stormsvlei (-AA), Bayer 1213 (NBG); Rondeheuwel (-AA), Bayer in KG326/71 (NBG); 19km N. Bredasdorp (-AC), Smith 5476 (NBG), Bayer in KG35/70 (NBG); Juliusfontein (-AD), Bayer 1221 (NBG); SW. Heidelberg (-BB), Bayer in KG104/74 (NBG); Skeiding (-BB), Smith 7219 (NBG); Ziekenhuis (-BC), Bruyns in KG49/76 (NBG); Potberg (-BC), Burgers 2506 (NBG); Infanta (-BD), Malherbe in NBG673/41 (NBG), Smith 5477 (NBG).
Inadequately located: Swellendam, Marais in STE6410 (NBG), Smith 5055 (NBG); Stormsvlei, Payne (NBG); Robertson, Payne (NBG), Malherbe in NBG172/41; Swellendam, Malherbe in NBG299/40; Bredasdorp, Barker in NBG698/33, Venter 18 (BOL); Stormsvlei to Bonnievale, Lewis in NBG2456/32 (BOL); Robertson, Esterhuysen (BOL), Hurling & Neil (BOL), Herre in STE6379 (BOL); Bonnievale, van der Merwe 94 (BOL); Bredasdorp, Venter 20 (BOL), Hurling & Neil (BOL).
Haworthia maraisii var. maraisii JDV95/1 Rooiberg. The end-area is shorter and more deltoid than in H. magnifica.
Haworthia maraisii var. maraisii JDV95/15 west of Robertson. There is a wide array of differing populations around Robertson.
Haworthia maraisii var. maraisii JDV85/15 Napier. At this locality the plants are small and the leaves rounder at the tip.
Haworthia maraisii var maraisii JDV86/50 Rooiberg. The plants are invariably well hidden.
Haworthia maraisii var. maraisii JDV87/1 17 Stormsvlei. The species was originally described from where the plants are quite large for the species.
Haworthia maraisii var. maraisii JDV87/118 Rondeheuwel. A population where the species seems to hybridize with H. mirabilis.
Haworthia maraisii var. maraisii JDV86/7 Rooivlei. Like the Napier plants, very dark and with more abrupt leaf-tips
Haworthia maraisii var. maraisii JDV94/18 southeast of Worcester. The first collection from here was by Roy Littlewood. The plants were very dark with erect pointed leaves, but that population appears to have vanished.
b. var. meiringii Bayer :134(1976). H. magnifica var. meiringii Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :45(1982). Type: CAPE‑3320 (Montagu): E. of Bonnievale (‑DC), Bayer in KG 224/70 (NBG).
meiringii: for P.L. Meiring.
1982 – The var. meiringii appears vegetatively very like a smaller, darker green version of H. herbacea, until it flowers. Also to the west it intergrades into the more characteristic retuse‑leaved var. maraisii.
1999 – This variety has the growth form of H. herbacea but is a smaller darker green species with the same flowers and flowering time as H. maraisii. It occurs east of Bonnievale, and westwards transposes to a more rigid-leaved form with erect thin scabrid leaves which practically co-occurs with the typical form of that species. Immediately west of Bonnievale it appears to intermingle with H. heidelbergensis in dense populations.
Distribution: 3319 (Worcester): W. Bonnievale (-DD), Smith 3822 (NBG). 3320 (Montagu): Bonnievale (-CC), Marloth 11855 (PRE); E. Bonnievale (‑CC), Smith 3948 (NBG), Bayer in KG 224/70 (NBG); W. Bonnievale (-CC), Bayer 1214, 1217 (NBG), Bayer in KG2/71 (NBG), Bayer 1218, in KG7/71, in KG9/71 (NBG).
Inadequately located: Bonnievale, Malherbe in Smith 3428 (NBG), Smith 5060 (NBG), van der Merwe 95 (BOL); ex hort, Hurling & Neil (BOL); Drew, Hurling & Neil (BOL).
Haworthia maraisii var meiringii JDV96/15 west of Bonnievale. The density of this population is surprising.
Haworthia maraisii var meiringii JDV90/94 east of Bonnievale. The shape and colouration of the plants is very reminiscent of H. herbacea.
Haworthia maraisii var meiringii JDV87/121 west of Bonnievale. Becomes very variable proceeding westward, and at this locality the plants are very spotted.
c. var. notabilis (V.Poelln.) Bayer :141(1976). H. notabilis V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938): H. magnifica var. notabilis Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :45(1982). Scott :146(1985). Type: Cape, Wolfkloof, G.J. Payne in Triebn. 1103. Not preserved. Lectotype (B&M): icon (B): H. schuldtiana var. erecta Triebn. et V.Poelln. ibid. 49:25(1940). Type: Cape, Bonnievale, Stellenbosch. Not preserved: H. nitidula var. opaca V.Poelln., Desert.Pl.Life 20:4(1948). Type: Cape, Klaasvoogds. Not preserved:
notabilis: noteworthy.
1982 – The variety notabilis (then of H. magnifica) also has erect leaves which are darker green and more turgid than in the case of H. herbacea.
1999 – There is difficulty in comparing this variety with H. maraisii. The decision to place it here was taken on account of the variation at the type locality, the similarity of the flowers and flowering times, and also because of the forms originally seen at Klaasvoogds, which were darker and more compact than the more turgid of the Wolfkloof forms. At Wolfkloof, a turgid, lighter green form grows on the east of the valley where it is on shales. On the west side is a granite formation and the forms are very toothed and have longer more slender leaves. There also appears to be a third form a little to the southwest which has the firmer textured leaves of H. reticulata which co-occurs there. More than one population is now known at Klaasvoogds and the plants appear to vary substantially. There is yet another population at Agtervink which is comparable to the Klaasvoogds and eastern Wolfkloof plants. Finally there is H. maculata var. intermedia at Buitenstekloof which has a different flowering time. It is these odd indistinctly related populations which appear to be influenced by ecotypic (largely geological) factors. All the populations discussed here are on rocky sites associated with the great Worcester fault line and the granite and dolomite formations which are exposed there.
The original decision to include this element with var. maraisii was based on quite an extensive study of the flowers and flowering times of a wide range of populations in the Worcester/Robertson Karoo. Variation within those populations which are more obviously of the var. maraisii, seemed to exceed that between them and var. notabilis. Flowering time was also originally thought to have been a very strong character but it seems to break down here as it may in H. magnifica.
Distribution: 3319 (Worcester): Wolfkloof, Robertson (‑DD), Smith 3984 (NBG, PRE), Scott 2204 (PRE), Bayer 1208, 1209 (NBG), Fourcade 166 (NBG), Fourcade 192 (NBG), Hurling & Neil in NBG 2115/37 (NBG); Robertson (-DD), Payne in PRE39466; Vinkrivier (-DD), Bayer 121 (NBG); Klaasvoogds (-DD), Stayner & Bayer in KG 638/69 (NBG).
Haworthia maraisii var. notabilis JDV87/197 northwest of Robertson. This variety occurs as three different ecotypes in close proximity.
Haworthia marasii var. notabilis JDV87/197 Woolfkloof northwest of Robertson. The granite form is darker coloured and more spinescent.
20. Haworthia marumiana Uitew., Cact.Vetp. 6:33(1940), Nat.Cact.Succ.J 9:20(1947). Bayer :133(1976). Bayer :23(1982). non Scott :78(1985). Type (see B&M): Cape, Ladismith and Mossel Bay, Stellenbosch 6610 and 7773 (AMD).
marumiana: in honour of Dr M. van Marum.
Rosette stemless, very proliferous, to 7cm φ. Leaves erect, incurved, softish, margins and keel with spines, purplish-green in colour, opaque, with reticulate patterning. Inflorescence simple, to 200mm. Flowers smallish, white.
1982 – The distribution of this species is most interesting. Uitewaal gave the locality as Ladismith and also a second locality at Mossel Bay, both attributed to H. Herre. A collection in the Kirstenbosch garden attributed to C. Luckhoff also appears to have been this species, and this was recorded as from Heidelberg, Cape. Smith did not apply the name to any of his field collections. The recorded localities are not borne out by any other records or collections and thus the origin remained something of a mystery. In 1972 Mr C. McMaster informed the writer of a species growing in the mountains south of Tarkastad, and this proved to be the missing species. It is a small very proliferous species and the only tetraploid so far recorded in the subgenus Haworthia. The distribution is rather surprising as it occurs north of Tarkastad itself, at Cradock, Graaff-Reinet, New Bethesda, Murraysburg, Nelspoort and Beaufort West. It is very possible that H. pearsonii may also have been this species as specimens of H. marumiana do have brown‑striped flowers. However, this is not certain and H. pearsonii is regarded still as insufficiently known. It is possible that H. batesiana is a glabrous form of H. marumiana. H. archeri may be a southwestern variant of H. marumiana also, although the distance between known localities of these two species is rather great. On the other hand the known localities of H. marumiana are also very disjunct.
1999 – Col. Scott again has a fairly valid point that H. archeri is actually the original H. marumiana particularly in respect of the localities associated with the latter name. However, my contention expressed in several of my early writings, is that interpretation of species based on illustrations or only descriptions, with no solid herbarium record, are the foundations for contention. If I could extend this argument I would add that proper weight must also be attached to the concept of a species which has distribution range and variation. Secondly I also based my opinion on the plants received from several different sources as well as on plants in Smith’s collection ostensibly of the type, which I would have assigned to H. marumiana as I recognised it, and not to this southwestern counterpart. Smith’s plants of this species were from various sources including Uitewaal. Thirdly, it is evident from the herbarium record that the Stellenbosch Garden collection of Herre had included plants from the Nuweveld mountains at Beaufort West. I could not locate the representative specimen which Scott cites (Scott 6354) in the PRE herbarium. This species must also be considered together with H. cymbiformis var reddii in the Cathcart/Queenstown area. H. marumiana is now regarded as a very variable and widely distributed species of the Central and Southern Karoo.
a.var. marumiana. The range of this species has been extended considerably since even the New Handbook. Several collections have been made far north of Queenstown and these are as different from the typical variety as var. archeri. There seem to be two different elements. One is from Sterkstroom and the Penhoek Pass in which the plants are quite large and with fine patterning on the leaves. Other collection from north and northwest are of plants with more succulent turgid leaves, and somewhat more translucent in the leaf reticulation.
Inadequately located: ex hort, Smith 6940 (NBG); Cradock, Archer 873 (BOL).
b.var. archeri comb.nov. H. archeri Barker ex Bayer, JS.Afr.Bot. 47:791(1981). Bayer :29(1982). H. marumiana sensu Scott :78(1985). Type: CAPE-3320 (Montagu): Whitehill (-BA), Archer in NBG 68145 (NBG).
archeri: for Mr J. Archer.
1982 – The specimen of H. archeri in the Compton Herbarium was suspected of originating anywhere but in the Whitehill area until collected there by Peter V. Bruyns in 1977. It is a very small compact brownish green species reaching to 60mm in diameter. The relationship to other species is very unclear and the other species in the same subgenus occurring in that area are H. arachnoidea, H. pehlemanniae, H. lockwoodii and H. wittebergensis. An affinity with H. magnifica is highly unlikely because of the geographic barrier and floral differences. It is very conceivable that it is related to H. marumiana which is, however, also not known nearer than in the Nuweveld mountains at Beaufort West. Vegetatively the plants are almost identical to H. marumiana from as far afield as Tarkastad and Graaff-Reinet, and it is primarily floral differences and geographical distribution which justify its recognition. It grows on Dwyka sandstones among karroid vegetation but on the cooler south slopes. The Whitehill area west of Laingsburg is north of the Witteberg mountain range and technically lies on the border of the winter and summer rainfall areas. However, it is most probable that H. archeri prefers a dry summer and what little moisture it can get, in the winter. H. archeri is not proliferous, as is H. marumiana, and is so far unknown in cultivation.
1999 -The manuscript name by Miss Barker was intended for Mr Archer who initially curated the Karoo Botanic Garden when it was still situated at Whitehill. This element has proved decidedly more common than first supposed and also proven to be very proliferous. It has been since also found at several places in the greater Sutherland, Merweville, and Frazerburg areas, and this seems to confirm the connection with H. marumiana. The two varieties come very close together geographically at Prince Albert too, where some populations are still within recognisable range as var. marumiana (albeit with more distinctive patterning on the leaves). The reticulate patterning on the leaves in the var. archeri appears to be generally finer than is the case with var. marumiana and the leaves are less flaccid., but the var. archeri has less marking on the leaves than is generally the case. The leaves also tend to curve outward and are a little more scabrid. The var. archeri thus is fairly common on the higher-lying areas between the Rooiberg Pass (S. Laingsburg) and to well north of Sutherland. There is a high degree of variation across that range, and one collection from near the Floriskraal Dam is glabrous and very like a small glabrous form of H. arachnoidea var. nigricans. There is also a population in the Gamkapoort area which has broader leaves and less spination. The flat elevation of the upper petals seems to be distinctive and there is an odd occurrence of the petal presentation in some populations where the plants would otherwise be regarded as simply H. arachnoidea.
c.var. batesiana (Uitew.) Bayer comb.nov. H. batesiana Uitew., Nat.Cact.Succ.J 3:101(1948). Bayer :101(1976). Bayer :30(1982). Scott :100(1985). Type (see B&M): Cape, Graaff-Reinet, Ferguson (AMD).
batesiana: in honour of G. Bates.
1982 – As it is presently known, H. batesiana is an unusual and distinctive small species of up to 5cm in diameter recorded from the Valley of Desolation near Graaff-Reinet. It is smooth, bright green with a light reticulate pattern on the leaves. It also offsets very freely to form large clumps. A recent collection by Prof. D. Muller-Doblies from the Valley of Desolation consisted of still smaller darker green plants (up to only 25 mm in diameter) in a tighter rosette of spined leaves. These clearly belong to H. marumiana and also fit into the distribution pattern for that species. The writer has not succeeded in finding H. batesiana in the field, but a collection from Klipplaat northwest of Cathcart is clearly comparable. However, the plants there are too robust to be regarded as H. batesiana and it appears that there is a tendency towards H. cymbiformis. The possibility of H. batesiana being only a glabrous variant of H. marumiana cannot be ruled out; this is also suggested by a collection of the latter species from south of New Bethesda.
1999 – Several collections of this variety are now known. These are by J. Bouwer in the Valley of Desolation, a collection by P.V. Bruyns from the Kamdeboo mountain, and another collection also by P.V. Bruyns from Tandjiesberg to the east of Graaff-Reinet. In both these latter collections the plants vary quite considerably and are all glabrous. The Klipplaat (Waterdown Dam) population is assigned to H. cymbiformis var. reddii.
Distribution: 3224 (Graaff-Reinet): Valley of Desolation (-AB), Hurling and Neil in NBG68978, Smith 6955 (NBG), Bouwer (NBG), Scott 985 (PRE); Kamdebooberg (-AC), Bruyns 2978 (NBG); Uitkomst (-AD), Bruyns 2994 (NBG); Tandjiesberg (-BC), Bruyns 3245 (NBG,BOL).
d.var. dimorpha comb. nov. H. archeri var. dimorpha Bayer, JS.Afr.Bot. 47:793(1981). Bayer :29(1982). Type: CAPE-3320 (Montagu): Constable Station, W. Laingsburg (-AD), H. Hall in Smith 7418 (NBG).
dimorpha: two shapes.
1982 – The variety dimorpha (of H. archeri) has much fewer leaves, which in cultivation both grow much larger (up to 120 mm diam.) and flex outward. This variety grows west of the typical variety and occurs in fynbos vegetation on Table Mountain Sandstone.”
1999 – Not much more light has been thrown on this variant. There has been no evidence of a connection to H. nortieri to the west. There have been some indications that a hard spinescent form resembling H arachnoidea occurs which has the same floral characters. Attention should perhaps be paid to the block patterned reticulation that occurs in the seed capsules of this species and also in H. pulchella. Interestingly, the same white tubercles which characterise this variety, also occur in the population of var. archeri cited above from Riethuisies in the Frazerburg area.
Distribution: 3320 (Montagu): Constable Station, W. Laingsburg (-AD), H. Hall in Smith 7418 (NBG).
e.var. viridis var.nov. Type: CAPE-3322 (Oudtshoorn): S. Prince Albert, Bayer 3620 (NBG, Holo.).
virida: green.
Differs from the typical in being light-green with narrower, more-erect leaves. (A var. marumiana foliis erectis angustioribus subviridibus differt).
Plants collected by P.V. Bruyns at Vrischgewaagd west of Prince Albert, are brighter green than normal and the rosettes are fairly tight and reminiscent of H. pulchella, which is a much more scabrid species. The reticulate patterning on the leaves is still apparent.
Distribution: 3321 (Ladismith): Vrischgewaagd (-BD), Bruyns 6252 (BOL).SW. Kliphuisvlei (-BD), Thompson 2166 (PRE). 3322 (Oudtshoorn): S. Prince Albert (-AC), Krige (BOL), Bayer 3620 (NBG), Bruyns 2601 (NBG).
21. Haworthia mirabilis Haw., Syn.Pl Succ. :95(1812). Bayer :136(75). Bayer :47(1982). Bayer, Excelsa 7:37(1977). pp. Scott :116(1985). Aloe mirabilis Haw., Trans.Linn.Soc. 7:9(1804). Ker-G., Curtis’ Bot.Mag. t1354(1811). Type: Cape, Masson. Not preserved. Neotype (designated here): Icon t1354, Curtis’ Bot.Mag.: H. mundula Smith, JS.Afr.Bot. 12:8(1946). H. mirabilis ssp. mundula (Smith) Bayer :139(1976). Bayer :47(1982). Type: CAPE‑3419 (Caledon): Mierkraal, Bredasdorp (‑DB), M. Otzen 10 in Smith 5479 (NBG).
mirabilis: wonderful.
Rosette stemless, proliferous, to 7cm φ. Leaves 10-15, retused, 3-4cm X 1,5cm, markedly retused, acute above, face translucent and lined, dark green, with marginal spines turning reddish in the sun. Inflorescence slender. Flowers narrow, elongate, biarcuate bud, upper lobes pinched at tips.
1982 – A full account of H. mirabilis is given in Excelsa (Bayer, 1977). It is intimately associated with H. magnifica but the nature of the relationship is not fully understood. The species grow near together at Stormsvlei and an intermediate population occurs a few miles to the south. The species also approach one another near the Potberg at the mouth of the Breede River, at Bredasdorp and, unconfirmed, in the area east of Drew. The initial distinction was that H. mirabilis (as H. triebnriana) was smooth on the leaf faces while H. magnifica was not. H. mirabilis is generally a larger plant, and with more translucence in the leaves than H. magnifica. The flower is a little larger and generally brown‑veined. Several names such as H. willowmorensis, H. triebneriana, and H. rossouwii are placed under H. mirabilis on rather speculative grounds. The synonymy thus indicates a very variable species and this is in fact so. There are several places, notably at Bredasdorp and Napier, where ecological differences have resulted in forms very different from one another. At Napier the differences are of such an order that the subspecies badia is recognised. At Greyton, a form high in the mountains, with the typical flower of H. mirabilis, occurs. For the present it is regarded as a variant of H. mirabilis. There are also variants in the limestone formations of the coastal belt. The subspecies mundula is a very proliferous variant south‑west of Bredasdorp. It is possible that H. magnifica var. paradoxa belongs with H. mirabilis (see H. magnifica) but the coastal area has not been fully explored so that a good decision is not yet possible.
1999 – The typification of this species is obvious although Haworth in 1812 does not cite the Botanical Magazine illustration. He does cite the preceding t.1353, Aloe recurva under Haworthia recurva, so this is quite an extraordinary oversight. There is an illustration in the Kew library apparently labelled H. mirabilis which Scott regards as illustrating this species. The illustration is almost identical to t.1353 and is of that species viz. H. recurva Haw. rather than of H. mirabilis. The illustration is of a trifarious plant and, despite the annotation ‘mirabilis, ‘there can be no question of any confusion between this and the description of H. mirabilis which reads ‘quinquefarius’. The other points are that 1. Ker-Gawler obtained his plants from Haworth and acknowledges this source; 2. Haworth refers to its singularity and ‘productions of art’; 3. Haworth places it between H. margaritifera and H. translucens(?).
This has been an old problem in Haworthia – the proliferation of names without the first establishment of the application of the old. The recognition of the correct application of the species name requires some additional modification to the structure of the species. While citing the wrongly labelled Kew illustration, Scott also omits mention of the Ker-Gawler illustration. In addition to this he appears to have identified several collection of H. maraisii as H. mirabilis which is also apparent from his distribution map. These two species do not co-occur and yet they seem to be discrete. Two populations are known which appear to be interactions of the two elements.
a.var. mirabilis. It is clear that H. mirabilis had to have some application and this has previously been found in terms of a host of varieties. In retrospect it is apparent that the only variety that satisfies the requirements of the species as originally described is H. mundula of Smith, transferred only to H. mirabilis as a subspecies in 1976. It is a problem of the nomenclatural system that the species may be described from a non-representative element, as has happened here where only one deviant population is concerned. The specimen nominated as the epitype by Breuer and Metzing is again unfortunate because this is the variant beukmannii.
Distribution: CAPE‑3419 (Caledon): Mierkraal, Bredasdorp (‑DB), M. Otzen 10 in Smith 5479 (NBG), Smith 3951, 5364, 5612 (NBG).
Inadequately located: ex hort, Malherbe in NBG303/60.
Haworthia mirabilis var mirabilis JDV96/33 southwest of Bredasdorp. G.G.Smith had made no attempt to relate his collections to Haworth’s original species. In fact very few authors have made the attempt to relate names to a concept of specie as natural phenomena.
Haworthia mirabilis var mirabilis JDV96/33 southwest of Bredasdorp. Very proliferous and mat-forming at the single locality known.
Haworthia mirabilis var mirabilis JDV92/84 southwest of Bredasdorp. The plants colour deeply in the dry summer, usually reddish.
b.var. badia (V.Poelln.) Bayer comb.nov. H. badia V.Poelln., Kakteenkunde 7:76(1938). Haworthia mirabilis subsp. badia (V.Poelln.) Bayer :101(1976). Bayer, Excelsa 7:42(1977). Bayer :47(1982). Type: Cape, Napier, G.J. Payne in Triebn. 1058. Not preserved. Lectotype (B&M): icon, Kakteenk. en Kakteenfr. :76(1938).
badia: reddish brown.
This is a robust, slowly proliferous variety growing in sandstone derived depauperate clay, in among pebbles and low-growing fynbos. The leaves are quite attenuate and develop a very deep shiny brown colour in the sun. Unfortunately most of its rather unique habitat has been destroyed by quarrying activity and invasion by alien vegetation.
Distribution: 3419 (Napier): Napier (‑BD), I. Williams 616 (NBG), Payne in PRE 34870, Smith 3239, 3269, 5207, 5480 (NBG), Bayer in KG628/69, Rossouw in NBG2095/37 (BOL); 3km Napier to Caledon (-BD), Scott 2207 (PRE).
Haworthia mirabilis var. badia JDV88/66 west of Napier. Usually a chestnut brown in the field.
Haworthia mirabilis var. badia JDV88/66 west of Napier. Often protected under low shrublets. Can be surprisingly difficult to see.
Haworthia mirabilis var. badia JDV88/66 west of Napier. The shiny end area of the leaf is conspicuous and characteristic.
c.var. beukmannii (V.Poelln.) Bayer comb.nov. H. emelyae var. beukmannii V.Poelln., Feddes Repert.Spec.Nov. 49:29(1940). Type: Cape, Caledon, C. Beukman. Not preserved. Lectotype (B&M): icon (B). Epitype (designated here): CAPE‑3419 (Caledon): Skuitsberg (‑BA), Smith 3969 (NBG).
beukmannii: for C. Beukman.
This is a very robust form with strongly retused leaves and spined margins.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. The plants are large despite growing in restricted shale cracks.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. The leaf tips are generally retused.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. Caledon. Occasionally proliferous.
Haworthia mirabilis var. beukmannii JDV88/70 Skuitsberg. It has been suggested that, historically, this was the possible origin of the species.
d.var. calcarea var.nov. Type: CAPE-3420 (Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG, Holo.).
calcarea: pertaining to lime.
Differs from the species in having short erect leaves, with a short retused end-area. It is proliferous and the rosettes, at least in cultivation, tend to be raised as opposed to remaining flattish to the ground. (A var. mirabilis foliis brevioribus erectis viridibus sordidus differt).
First collected by C. Burgers in the De Hoop Nature reserve where it grows on low-lying limestone rocks. There are two other collections from the Potberg area which may have some connection with this variety, or with the species. One is by Prof. Compton in which the plants appear to have been in sand and very obscure. At first sight they appear to be of H. mutica, but they have very pointed leaves. The other collection is also by C. Burgers and appear to be of plants with more slender and longer leaves from the lower slopes of the Potberg. There is also a collection from the Potberg once made by A. Mitchell of a very small plant, like the variety consanguinea described below, but which is H. variegata var. modesta. H. heidelbergensis is also known from this area.
Distribution: 3420 (Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG); NNE. Buffelsfontein (-BC), Burgers 2018 (NBG).
Haworthia mirabilis var. calcarea JDV84/65 De Hoop. An unusual occurance for the species in limestone.
A small proliferous, relatively soft-leaved form comparable with the small proliferous mountain form of H. turgida. (A var. mirabilis rosulis parvioribus prolificantibus facile et foliis virellis differt).
It is difficult to deal with these montane forms which seem to occur in the sandstone ranges from the Potberg, the Riviersonderend, the Langeberg, the Swartberg and the Cedarberg mountains. This particular element seems to be associated with the lower-lying varieties in the shales, in the same way that H. turgida transposes to retusa-like forms. Although looking very similar to the sandstone forms of H. turgida, it has the very narrow elongate buds of H. mirabilis, and also the leaves have the brownish-red coloration associated with H. mirabilis. The P.V. Bruyns collection cited below is significantly different from the two others as the leaves are quite slender.
Inadequately located: Near McGregor, Esterhuysen 5218, 5219 (BOL).
Haworthia mirabilis var. consanguinea JDV88/63 Dwarswaterkloof southwest of McGregor. This form is also in sandstone and the leaves are slender and slightly cylindrical.
Haworthia mirabilis var. consanguinea JDV84/76 Die Galg northeast of Greyton. This form is very similar to H. turgida except that it has the brownish-red tints of H. mirabilis, and also a similar flower.
f.var. paradoxa (V.Poelln.) Bayer comb.nov. H. paradoxa V.Poelln., Feddes Repert.Spec.Nov. 33:240(1933). idem. Desert.Pl.Life 99:90(1937). H. maraisii var. paradoxa (V.Poelln.) Bayer :143(1976). H. magnifica var. paradoxa (V.Poelln.) Bayer, Nat.Cact.Succ.J 32:18(1977). Bayer :45(1982). pp. H. asperula Haw. sensu Scott :119(1985). Type: Cape, Vermaaklikheid, Mrs Ferguson in Stellenbosch 6692. Not preserved. Neotype (B&M): Riversdale, Ferguson (BOL).
paradoxa: paradoxical.
Von Poellnitz did not explain his epithet and simply related it to his H. schuldtiana (H. maraisii). He was struck by the spines on the leaf face which are unusual. A greater paradox is the relation of this element to H. emelyae var. major which also has a very spined leaf surface. It does not follow that such spines are a dichotomously allocated character. Their occurrence in H. magnifica does not necessarily suggest that var. paradoxa is better placed there on these and geographical grounds, than with H. mirabilis. The var. paradoxa also seems to be associated with limestones and there is no other known link with H. mirabilis east of the Breede River. The solution which I offer is the somewhat dubious connection of the species through H. heidelbergensis, and possibly also through the two populations ascribed to var. calcarea.
Distribution: 3421 (Riversdale): Vermaaklikheid (‑AC), J. Dekenah 8 (NBG), Kramer 433 (PRE), Fourcade 277 (NBG), Smith 3272, 5388, 6109 (NBG).
Inadequately located: Ferguson 1 (BOL).
Haworthia mirabilis var. paradoxa JDV94/89 south of Vermaaaklikheid. Proliferous and growing on coastal limestones.
Haworthia mirabilis var. paradoxa JDV94/89 south of Vermaaaklikheid. Very similar both vegetatively and florally to H. emelyae var. major.
g.var. sublineata (V.Poelln.) Bayer comb.nov. H. triebneriana var. sublineata V.Poelln. in Feddes Repert.Spec.Nov. 44:135(1938). Type: Cape, Bredasdorp, G.J. Payne in Triebn. 1106. Not preserved. Neotype (designated here): CAPE-3420 (Bredasdorp): S. Bredasdorp (-CA), Smith 3966 (NBG).
sublineata: almost lined.
This is also a sandstone variant from south of Bredasdorp. The leaves are relatively long and slender. A similar, well-lined but more robust form used to be abundant immediately north of the town too along the river bank. It should be expected to occur further to the west.
Distribution: 3420 (Bredasdorp): S. Bredasdorp (-CA), Smith 3966 (NBG), Stayner in KG209/60; N. Bredasdorp (-CA), Smith 3252, 3266, 3830, 3976 (NBG).
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. A more robust form used to occur to the north of the town as well.
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. A lighter green colour and with long attenuate leaves.
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. Very similar forms also occur in the sandstones north of Riviersonderend mountains.
h.var. triebneriana (V.Poelln.) Bayer comb.nov. H. triebneriana V.Poelln., Cactus J 5:33(1936). idem. Feddes Repert.Spec.Nov. 41:214(1937). idem. Cactus J 6:36(1937). idem. Desert.Pl.Life 99:101(1937). idem. Feddes Repert.Spec.Nov. 47:8(1939): H. mirabilis Haw. pp. Bayer :136(1976). pp. Bayer :47(1982). pp. Bayer, Excelsa 7:42(1977). pp. Scott :69(1985). Type: Cape, Strydomsvlei, Mrs Helm in Triebn. 841. Not preserved. Lectotype (Bayer, 1976): icon. (B): H. willowmorensis V.Poelln., Feddes Repert Spec.Nov. 41:216(1937). non Scott, Aloe 11:42(1973). Type: Cape, Willowmore, Mrs Helm in Triebn. 840. Not preserved. Lectotype (B&M): icon (B): H. triebneriana var. depauperata idid. 43:94(1938). ibid. Desert.Pl.Life 9:101(1937). Type: Cape, Stormsvlei near Robertson, Payne in Triebn. 990. Not preserved. Lectotype (Bayer, 1976): icon. (B): H. triebneriana var. multituberculata idem. Feddes Repert Spec.Nov. 44:135. idem. 47:10(1939). Type: Cape, NW. Napier, G.J. Payne in Triebn. 1111. Not preserved. Lectotype (Bayer, 1976): icon, (B): H. triebneriana var. rubrodentata Triebn. et V.Poelln. ibid. 47:10(1939). Type: Cape, between Villiersdorp and Greyton, G.J. Payne in Triebn. 1143. Not preserved. Lecotype (B&M): icon (B). Epitype (designated here): CAPE-3419(Caledon): near Genadendal (-BA), Bayer in KG692/69 (NBG): H. triebneriana var. napierensis ibid. Type: Cape, Napier, G.J. Payne in Triebn. 1145. Not preserved. Neotype (designated here): CAPE-3419(Caledon): Skietpad (-BD), Bayer 4642 (NBG): H. triebneriana var. turgida Triebn. ibid. 47:11(1939). Type: Cape, N. of Napier, G.J. Payne in Triebn. 1107. Not preserved: H. triebneriana var. subtuberculata V.Poelln. idem. 47:10(1939). Type: Cape, S. Caledon, G.J. Payne in Triebn. 1114. Not preserved: H. triebneriana var. pulchra V.Poelln. ibid. 49:29(1940). Type: Cape, N. Stormsvlei, Stellenbosch 19. Not preserved. Neotype (designated here): CAPE-3420(Bredasdorp): Stormsvlei Pass, Beukmann in Smith 5609 (NBG): H. rossouwii V.Poelln., Kakteenkunde 7:75(1938). Type: Cape, Napier, Rossouw in Triebn. 1059. Not preserved. Lecotype (B&M): icon (B). H. nitidula V.Poelln., Desert.Pl.Life 11:192(1939), Bayer, Cact.Succ.J(U.S.) 52:10(1980). Type: Cape, Worcester, Swellendam etc., Venter 15. Not preserved. Lectotype (B&M): icon (B). Epitype (designated here): CAPE-3419(Caledon): near Greyton, Bayer in KG31/70 (NBG).
triebneriana; in honour of W. Triebner.
In recognising the typical variety as the previously named ssp. mundula, it becomes necessary to find a name for the main body of the species, and this is now taken from the first name that can be fairly definitely associated with the species. This is despite the improbable locality cited for H. triebneriana and considering the very poor records associated with Triebner’s contributions. Where the typical variety of the species is restricted to the single locality southwest of Bredasdorp, this generalised variety is widespread in the area from Caledon, east to Swellendam, to Napier and back to Caledon. At Swellendam there is an additional population to the one south of Stormsvlei where this species is confounded with H. maraisii.
Distribution: 3419 (Caledon): Uitkyk (-AB), Smith 5562 (NBG), Bayer in KG682/69 (NBG); N. Uitkyk (-AB), Bayer in KG28/70 (NBG); Dagbreek (-BA), Bayer 2453 (NBG); near Greyton (-BA), Smith 3245, 3260, 3265, 3419, 3811, 3904, 3905, 3968, 5481, 5482, 5643 (NBG), Bayer in KG 30/70, in KG31/70 (NBG); Near Genadendal (-BA), Bayer in KG692/69 (NBG); Skietpad (-BD), Bayer 4642 in KG510/70 (NBG); Mierkraal, Napier (-BD), Bayer in KG681/69 (NBG). 342O(Bredasdorp): Stormsvlei (-AA), Smith 3254, 3457, 3971, 5609, Bayer in KG26/70 (NBG), De Kok 296 (NBG); Near Breede River (-AB), Tomlinson 13680 (PRE).
Inadequately located: ex Triebner, Smith 4947 (NBG); Venter in NBG6290/39; Caledon, Venter 13, 15 (BOL).
Haworthia mirabilis var. treberiana JDV96/47 southeast of Greyton. Large plants which are often subjected to fire.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad, Napier. The typical variety is the residue of plants of the species not named as varieties. May be robust or relatively small.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad. This was the locality for the ‘var. napierensis’ V.Poelin.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad. Some plants are somewhat tuberculate.
Haworthia mirabilis var. treberiana JDV90/34 west of Genadendal. Occasionally the marginal spines may be conspicuously reddish.
22. Haworthia monticola Fourc., Trans.Roy.Soc.S.Afr. 21:78(1937). Scott :57(1985). Type: Cape, George and Uniondale districts, Fourcade 2498 (K): H. divergens Bayer :113(1976). Bayer :38(1982). Type: CAPE‑3322 (Oudtshoorn): Molen River (‑DD), Bayer 175 (NBG).
monticola: mountain dwelling.
Rosette stemless, proliferous, 2-4cm φ. Leaves 30-40, elongate, lanceolate, 2-6cm long, tips incurving, margins and keel with small spines, upper surfaces often with pellucid spots. Inflorescence slender, to 30cm. Flowers sparse, white.
1982 – The name of this species [i.e. divergens] is derived from its probable relationship with H. variegata and H. angustifolia. It occurs in the Outeniqua mountains between Oudtshoorn and Willowmore and is separated from H. variegata, which it closely resembles, by the Langeberg mountains, the Gouritz River Valley and the intercession of H. chloracantha. It is separated from H. angustifolia by a distance of about 160km. The leaf tips tend to curve inwards and the marginal spines are more conspicuous than in related species. Towards Uniondale and Willowmore the species is still strongly variegated but becomes smaller. Beyond Willowmore there is a suggestion that it may intergrade with H. zantneriana.
1999 – Fourcade’s argument that Baker in Flora Capensis of 1880, was describing this species in mistake for Haworth’s H. angustifolia, is weak in the extreme. His reference to translucence in the lower leaf area is nonsensical and is simply a consequence of submergence which occurs in most species when so growing. There is no doubt from his specimens that he was dealing with a different species, and why he should have decided to draw Baker’s, or even Schultes earlier description into doubt is odd. They were both based on Haworth’s species and probably on Salm-Dyck’s illustration. This is why I applied a new name viz. H. divergens. However, if it is argued that Fourcade’s species has no connection to Baker, and is based on and typified by different material (Fourcade’s own specimens), then I presume his name should stand.
Affinities between this species and H. angustifolia are often cited. However, there is a problem of distribution and geographical co-herence. It is far more probable that the connection of the two species is through H. zantneriana and the problem can only be fully understood through a good knowledge of the mountain ranges of the southern Cape. There are very few collections of H. monticola east of Willowmore. The furthest is at Sandvlakte in the Baviaanskloof where the plants have fairly turgid leaves with some translucence. This is however, in the southern mountains of the Baviaanskloof and thus cannot connect to H. zantneriana. If it did prove continuous with that species it would have to be east of the Perdepoort to the north of Willowmore. There is a collection of P.V. Bruyns (6316) from that area which may prove to be this missing link. In fact the more recent collection also by Bruyns (7071) cited under H. zantneriana var. minor, is from the specified area and seems to confirm the continuity of the two taxa. A collection by J.D. Venter from the Witberg east of Willowmore, may do the same. Here the plants are less denticulate, very spotted and rather softer than usual for the species. There does not seem to be any likelihood that there is a direct connection with H. angustifolia across the barrier of the Groot River Valley. The Oudekraal collections of that species (its var. baylissii) do allow suggestion of such an affinity.
a. var.monticola . The typical variety is very similar in appearance to H. variegata with mottling of the leaf surfaces. Around Uniondale these markings are less conspicuous and the leaves are blackish. Southeast of Willowmore the leaves are almost spineless and the plants are very mottled.
Distribution: 3322 (Oudtshoorn): 13km N. Klaarstroom (-BC), Bruyns 2255 (NBG); Molen River (‑DD), Bayer 175 (NBG), Smith 2895 (NBG); Erfpacht (-DD), Smith 5809 (NBG). 3323 (Willowmore): Leeukloof (-AB), Stayner in KG117/62 (NBG), Venter 91/139 (NBG); Boesmanspoortberg (-AB), Bruyns 6316 (BOL); Perdepoort (-AB), Branch 461 (NBG); Georgida (-AD), Bayer 3363 (NBG); Witberg (-BC), Venter 92/102 (NBG); S. Willowmore (-BC), Stayner in KG644/61 (NBG); Uniondale (-CA), Stayner in KG242/70 (NBG); Langkloof (-CA), Taute in NBG1282/26 (NBG); 9km W. Uniondale (-CA), Fourcade 3606 (PRE); E. Uniondale (-CA), Bayer 3384 (NBG); NE. Uniondale (-CA), Smith 3664, 3665 (NBG); Nuwekloof (-DA), Bruyns 1655 (NBG). 3324(Steytlerville): Bosrugpad, Sandvlakte (-CA), Boycott in NBG144868; Dam se Drif (-CA), Bruyns 1844 (NBG).
Inadequately located: Uniondale, Herre 5686 (BOL); Oudtshoorn, Taylor 31 (BOL).
Haworthia monticola var. monticola JDV94/64 southeast of Willowmore. A smallish dark form.
Haworthia monticola var. monticola JDV86/90 Uniondale. Only slightly variegated.
Haworthia monticola var. monticola JDV86/79 northwest of Uniondale. The leaves are more incurved at the tips, otherwise there is a resemblance, even in the flowers, to H. angustifolia.
b. var. asema var.nov. Type: CAPE-3321 (Ladismith): Calitzdorp, Besemkop (-DA), J.D. Venter 12 (NBG, Holo.).
asema: without distinguishing marks.
Leaves smoother, more turgid, generally shorter, more uniformly grey-green, flowering very much earlier. (A var. monticola radicibus crassioribus et foliis brevioribus obesioribus griseo-viridibus differt).
This variety is widely separated from the main body of the species and is known at two localities northeast of Calitzdorp. It does not have clear relationships with H. monticola nor with any other species. The root system is less branching, the roots much more succulent, it flowers very much earlier and the off-sets are very quickly free of the mother plant. It may be the species noted by Von Poellnitz as H. angustifolia from Calitzdorp (also commented on under H. pulchella). It is not given species status simply because it is inadequately known and there is no better information relating it to other species.
23. Haworthia mucronata Haw., Haworth, Suppl .. Pl. Succ.: 50 (1819). Type: Cape, exhort. Kew. Not preserved. Lectotype: icon (K): H. helmiae V.Poelln., Feddes Repert. Spec. Nov. 41: 201 (1937). Scott: 99 (1985). H. unicolorvar. helmiae (V.Poelln.) M.B.Bayer: 121 (1976), Bayer: 59(1982): Type: Cape, Heidelberg, Smithers in Triebn. 901, Gt Brak River, Mrs Helm in Triebn. 901. Not preserved . Lectotype (B&M): icon, (B). Epitype (designated here): CAPE-3322 (Oudtshoorn): Schoemanspoort (-AC), Bayer 171 (NBG): H. V. Poelln., Feddes Repert. Spec. Nov. 44: 233 (1938), Scott: 98 (1985). Type: Cape, Little Karoo, Mrs E. Ferguson. Not preserved. Neotype (designated here): CAPE-3321 (Calitzdorp): On the Gamka East road (-DA), Scott 1050 (PRE): H unicolorV.Poelln., Kakteenkunde 10: 133 (icon), 154 (1937). Bayer: 165 (1976). H. unicolor var. unicolor (V.Poelln .) M.B.Bayer: 59 (1982): Type: Cape, Barrydale, A.J. Joubert in STE5230. Not preserved. Lectotype (B&M): icon in Kakteenk. & Kakteenfr.: 133 (1937): Epitype (designated here): CAPE-3320 (Montagu): Barrydale (-DC), J. Dekenah 234 (NBG): H. mclarenii V.Poelln., Des. Pl. Life 11: 107 (1939). Type: Cape, near Barrydale, G. McLaren in Triebn. 1160. Not preserved. Lectotype (B&M): icon (B): H. aristata Haw. sensu. V.Poelln., Feddes Repert. Spec. Nov. 43: 92 (1938). Ibid. 49: 27 (1940). Sensu Jacobsen 2: 537 (1954). Sensu CL.Scott, Nat. Cact. Succ. J. 35: 11 (1980). Ibid.: 110 (1937): H. serrata M.B.Bayer sensu CL.Scott: 62 (1985).
mucronata: pointed.
Rosette stemless, proliferous, 6-12cm φ. Leaves 30-45, soft, incurved, broadly ovate-lanceolate, slightly pellucid, with translucent margins and keel, both often spined. Inflorescence simple, robust. Flowers many, compact on peduncle, broad across upper tube, white with green venation.
NOTE: Considerable confusion surrounds the above and following synonymies. The species H. unicolor and H. rycroftiana are now included here, and one variety of the former, var. venteri, is transferred to H. arachnoidea. Haworthia habdomadis and its varieties are also subsumed in this new arrangement, under H. mucronata.
1982 – The original specimen of H. unicolor was from the town of Barrydale and pale green plants of this kind can still be found in that immediate area. However, von Poellnitz also included citations from Montagu and Ockertskraal. In a later publication (1938), he included the species in synonymy under H. aristata on the basis of plants received from Long from the Cango Caves. These plants included forms which von Poellnitz felt represented both his H. unicolor and H. aristata. However, in Feddes Repert. Spec. Nov. 43:92(1940), he continues the discussion and cites specimens, one later to be described as H. mclarenii. It seems clear that these citations by vn Poellnitz refer to the better known and highly variable H. unicolor var. venteri. The locality of H. mclarenii cited by Von Poellnitz as Tulbagh was patently erroneous. G.G. Smith records that W.E. Armstrong wrote “I had sent away a batch of plants, some from Mr McLaren and others from Miss de Klerk of Tulbagh and of the batch L693, Arm. 180 and Trieb.1160 actually collected by Miss de Klerk of Uitzicht, P.O. Tulbagh, at Barrydale, was erroneously named H. mclarenii”. There is thus no doubt that this species is synonymous with H. unicolor. A photograph by Fourcade (no.224) of H. mclarenii also confirms this view. The type locality for H. venteri was recorded by Smith as ” 3/4m. Barrydale‑Lemoenshoek” although Major H. Venter’s localities are very broadly stated in publication and practically valueless. Only Smith’s notes give any better clue to these but in this case there is still no certainty. However, there is a very common species extending from Barrydale to Oudtshoorn from which this variety is clearly drawn. At Barrydale itself the plants are rather pale green with elongated sparsely setate leaves ‑ this is regarded here as the species H. unicolor. Eastward the plants are more robust and often glabrous, usually distinguished by particularly dark‑purplish coloration towards the leaf tips. The further eastward one progresses the more setate the plants become until at Oudtshoorn, south of the Cango Caves, the plants are comparatively small and more softly setate (H. unicolor var.helmiae). It is not in the least clear what the relationship is between H. unicolor and H. arachnoidea. They do not appear to cohabit which suggests that they may be conspecific and only differentiated on an ecotypic basis. This is borne out by the pale arachnoidea‑like population at the northern entrance to the Tradouw Pass, south of Barrydale. However at Ladismith H. arachnoidea is present as a very distinct heavily setate form, and similarly south of the Cango Caves it occurs near to H. unicolor var. helmiae as a softer haired but also highly setate form. Cognisance must also be taken of H. integra from apparently west of Ladismith. The species H. unicolor is therefore not well‑known or properly understood and this solution must be regarded as suspect. Von Poellnitz’s citations of localities for the var. helmiae are highly confusing and in Feddes Repert. Spec. Nov. 44:223(1938) are totally disparate. The type is cited in 1938 as “Great Brak River, Mrs Helm” and three Triebner numbers are added to this. Mrs Helm’s strong personal recollection (private communication, and acknowledgement to Col. C.L. Scott) is that the original plants were collected at a specific site south of the Cango Caves, Oudtshoorn. The photograph extant in the Botanical Museum Dahlem (which must serve as the type) is of a very poor specimen, but it can be reasonably allied with plants from the site given by Mrs Helm. It is concluded that it is an extension of the complex to which it is here referred. J.R. Brown’s illustrations in Cactus Succul.J(U.S.) 18:39(1946) are not of this variety.
1999 – The confusion here is not only due to human error and perversity. The problem lies in trying to explain the difficulties of the situation itself, and then in the light of all the different perceptions about the species which may or may not be involved. Col. Scott distributes elements of the single typical variety into four species (including his perception of H serrata Bayer) in three different sections, and this exemplifies how difficult it is to arrive at a consensus. To understand the changes effected here, one should read the synonymy thoroughly. The changes are consistent with the explanations given in 1982 and it is felt that var. venteri should in fact go under H. arachnoidea under an older name nigricans. Similarly var. helmiae is subsumed under H. arachnoidea as well. The remnant, H. unicolor, correctly belongs to H. mucronata Haw. but not in the sense used by Scott where elements of H. bolusii, H. cooperi, H. aristata, H. arachnoidea and possibly two other species are also included.
Three factors are important in ringing these changes:-
A collection from northeast of Montagu (cited under the var. inconfluens) in which the plants are highly variable and which could be assigned to var. habdomadis, var. mucronata, var. integra and possibly to H. arachnoidea var. nigricans.
The relationship of H. arachnoidea to this species via its varieties.
The relationship of H. rycroftiana to H. integra V.Poelln. as interpreted by Scott (the name integra is now upheld over the former).
Apart from these factors, the characteristic of the species is the translucence along the margins and keels, which separates it from H. arachnoidea. Unfortunately the confusion within the ambit of the species itself confounds interpretation of relations with other species. Hayashi (unpublished) repeats and emphasizes the probable relationship of the species with H. cymbiformis through its var. transiens, as conjectured in 1982. One cannot exclude a similar relationship with H. cooperi as also conjectured under the var. habdomadis. The inclusion of all those diverse elements in H. mucronata sensu Scott, does not hint at the real relationship of the species with H. decipiens and H. lockwoodii north of Seweweekspoort. There is considerable difficulty in unequivocally designating specimens.
a. var. mucronata. The typical variety occurs around Barrydale and it is clear that there are many variants including the transformation to arachnoidea-like plants in Tradouw Pass. It was these variants which initially suggested to me that the variation to H. arachnoidea var. nigricans was continuous. However, the glabrous forms of the latter should always be distinguished by the characteristic of non-tranlucent keel or margins to the leaf. The leaves of this variety (the argument can be extended to the varieties integra and nigricans) may become very spiny. The loss of marginal translucence, particularly in the northwestern Little Karoo makes it very difficult to distinguish it from H. arachnoidea.
Distribution: 3320(Montagu): Barrydale (‑DC), J. Dekenah 234 (NBG), Smith 3911, 3992, 6572 (NBG), Smithers (BOL), Bolus in NBG697/32 (BOL), Hurling & Neil (BOL), Jordaan (BOL), Villet in KG180/61 (NBG), Stayner in KG119/71 (NBG); 18km W. Barrydale (-DC), Scott 6200 (PRE); Hills S. Barrydale (-DC), Scott 5089 (PRE). 3321(Ladismith): Algerynkraal, Brakkloof (-AD), Curator PRE Bot. Garden 3628 (PRE).
Inadequately located: Montagu, Rogers in BOL15478.
Haworthia mucronata var. mucronata JDV92/53 north of Barrydale. This typical variety has a thinner, less turgid leaf than the more widespread var. inconfluens.
Haworthia mucronata var. mucronata JDV92/55 south of Barrydale. At the original locality the plants are mostly without marginal spines to the leaves.
Haworthia mucronata var. mucronata JDV92/50 Perdekop. Here, as at most localities other than the type, the plants have spines on leaf margins and keels.
b. var. habdomadis (V.Poelln.) Bayer comb.nov. H. habdomadis V.Poelln., Desert.Pl.Life 11:88(1938). Scott :106(1985). H. inconfluens var. habdomadis (V.Poelln.) Bayer :121 (1976). H. habdomadis var. habdomadis (V.Poelln) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :41(1982). Type: Cape, Seweweekspoort, A.J. Joubert in Stellenbosch 7692. Not preserved. Neotype (B&M): Seweweekspoort , Barker & Lewis in NBG2764/32 (BOL).
habdomadis: referring to Sevenweekspoort.
1982 – Haworthia habdomadis is a species of the Little Karoo and it extends from Anysberg in the west to as far as Uniondale in the east. Three varieties are recognised as in H. unicolor and these two species have a puzzling relationship. Apart from a locality south of Vanwyksdorp, they are not known to occur together. H. habdomadis var. habdomadis is an element occurring in the sandstone mountains between Sevenweekspoort and just west of Ladismith. The var. inconfluens is more opaque and is usually spineless. This variety occurs in a form at Anysberg barely distinguishable from H. cymbiformis var. transiens, in that it has the same rounded leaf tips. The var. morrisiae is generally a brilliant green colour and occurs in the Oudtshoorn/Volmoed areas. However, it also occurs at Prince Albert and at Van Wyksdorp. It has both pointed and unpointed leaf forms and the distribution does not seem to strictly complement that of the var. inconfluens. Furthermore the form at Van Wyksdorp has the purple tinges associated with H. unicolor var. venteri. The form at Van Wyksdorp has the characteristic leaf shape of H. unicolor var. venteri with the same well‑defined keel and terminal bristle, and at this locality it grows practically in conjunction with that species. The two species flower simultaneously and there do not seem to be significant differences in the flowers. The presence of H. habdomadis var. inconfluens north of Uniondale needs confirmation of identification.
1999 – The circumscription here narrows this variety down to the sandstone forms with a more compact translucent rosette. Although also known from just west of Ladismith, very little further information regarding its distribution is available. Spined forms of this nature are known north of Seweweekspoort and also east of De Rust and presumed to be the var. inconfluens. The recurrence of such soft, short leaved forms north of Montagu is surprising and perhaps re-enforces the perception of ecological adaptation, in this case to low-nutrient sandstone. If this is the case, similar forms should occur on the south facing rocks of the Touwsberg, Anysberg and Warmwaterberg.
Distribution: 3321 (Ladismith): Dwarsrivier (-AC), Bayer 1618 (NBG); Swartbergkloof (-AC), Joubert 182 (BOL); Seweweekspoort (‑AD), H. Hall in NBG69371, Hall 2199 (PRE); Smith 7524 (NBG), Malherbe in NBG654/41, Compton & Lamb in NBG2340/27 (BOL), Barker & Lewis in NBG2764/32 (BOL); Lower Seweweekspoort (-AD), Smith 6991 (NBG).
Haworthia mucronata var. habdomadis JDV97/120 Seweweekspoort. The leaves are shorter and broader than in the typical variety or the var. inconfluens.
Haworthia mucronata var. habdomadis JDV97/120 Seweweekspoort. There are several sandstone mountain ridges in the Little Karoo, and where H. mucronata or H. arachnoidea occur on them, they occur as smaller, softer variants.
c. var. inconfluens (V.Poelln.) Bayer comb.nov. H. altilinea var. limpida fa inconfluens V.Poelln., Feddes Repert.Spec.Nov. 45:169(1938). H. mucronata var. limpida fa inconfluens idem. 49:29(1940). H. inconfluens (V.Poelln) Bayer :121(1976). H. habdomadis var. inconfluens (V.Poelln) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :41(1982). Type: Cape, Ladismith waterfall, H. Herre in STE7695. Not preserved. Lectotype (B&M): icon, Triebner 1031 (B): H. bijliana var. joubertii V.Poelln., Cactus J 5:36(1936). idem., Feddes Repert.Spec.Nov. 41:196(1937). H. setata var. bijliana sv joubertii idem. 44:224(1938). H. setata var. joubertii (V.Poelln.) Jacobsen, Hand.Succ.Pl. 2:593(1960). Type: Cape, Ladismith, A.J. Joubert in Triebn. 878. Not preserved. Neotype (designated here): CAPE-3321 (Ladismith): N. Ladismith (-AD), Smith 5734 (NBG).
inconfluens: coming together.
It is not known to what von Poellnitz alluded when he named this form. Presumable he was referring to the lines of the leaves joining towards the leaf-tips, or he may have been alluding to the coming-together of different geographic forms. The var. joubertii was collected north of the town of Ladismith where spined forms of this large variety grow. Just south of the town there is a massive population of spineless plants. This variety is not well-represented in the herbarium although it is known as far afield as Hoekvandieberg and Bellair Dam. Eastward it is known to the Huisriver Pass and again northeast of Calitzdorp. From there it is apparently replaced by the green var. morrisiae. A pale grey-green counterpart is found again from Uniondale eastward and this is probably the route to either H. cymbiformis or to H. cooperi. There is another transformation north of Seweweekspoort, already referred to, which involves H. decipiens and H. lockwoodii. Where previous authors have simply confounded H. bolusii var. blackbeardiana with H. mucronata, these populations would surely have driven them finally to a more realistic appreciation of the actual problems involved.
Distribution: 3320 (Montagu): Montagu (-CA), Dymond in NBG2096/32 (NBG); Knipes Bath (-CA), Smith 3990 (NBG); Dammetjies (-CB), Stayner in KG47/67 (NBG); Jakkalsfontein (-DA), Bayer 4438 (NBG); Anysberg Pass (-DA), Bayer 1986 (NBG), Martin 57 (NBG); Warmwaterberg (-DA), Smith 7310 (NBG), 3321(Ladismith): 8km W. Ladismith (-AC), (NBG); Dwarsrivier (-AC), Bayer in KG567/71 (NBG); N. Ladismith (-AD), Smith 5734 (NBG); Ladismith (-AD), Pole-Evans in PRE34908, Malherbe in NBG587/40; Die Berg (-BC), Bayer 159 (NBG); Huis River Pass (-BC), Hardy 320 (PRE), Smith 6887 (NBG); Boerbonefontein (-CA), Laidler 361 (NBG); Noukloof (-CA), Laidler 84 (NBG); 4,5 km south of Ladismith(‑CA), Smith 5503, 5730, 5731 (NBG), Bayer 1628, in KG 590/71 (NBG); 9km SW. Ladismith (-CA). 3322 (Oudtshoorn): Schoemanspoort (-AD), Taylor (BOL).
Haworthia mucronata var. inconfluens JDV91/97 south of Langsburg. The plants are normally a paler green with more turgid leaves than the typical variety, and without spines too.
Haworthia mucronata var. inconfluens JDV89/56 Prinsberg, western Little Karoo. The similarity of the varieties stresses the difficulty of circumscribing them even geographically.
Haworthia mucronata var. inconfluens JDV86/102 south of the Ladismith. In some populations the plants are more spinose and the historic confusion with H. bolusii var. blackbeardiana is quite justified.
Haworthia mucronata var. inconfluens JDV91/74 east of Rooinek Pass in the Witteberg. Here the plants may have spinescent leaf margins and somewhat resemble H. decipiens.
d. var. morrisiae V.Poelln., Feddes Repert.Spec.Nov. 49:29(1940). Scott :83(1982). H. altilinea var. morrisiae V.Poelln., Feddes Repert.Spec.Nov. 45:168(1938). H inconfluens var. morrisiae (V.Poelln.) Bayer :121(1976). H. habdomadis var. morrisiae (V.Poelln.) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :41(1982). Type: Cape, Oudtshoorn and Calitzdorp, Mrs Morris in Long 484. Not preserved. Lectotype (B&M): icon (B).
morrisiae: for Mrs G. Morris.
This is a bright, almost emerald-green, variety from around Oudtshoorn and Calitzdorp. It is known westward to south of Vanwyksdorp. Greenish forms of the species also occur further west at Anysberg Pass and from southwest of the Anysberg.
Distribution: 3320 (Montagu): Touwsfontein (-CB), Bayer 5296 (NBG). 3321 (Ladismith): W. Vanwyksdorp (-CB), Bayer in KG568/71 (NBG); Bergplaas (-BC), Bayer in KG323/70 (NBG); Warmbron (-DB), Bayer 1621 (NBG). 3322 (Oudtshoorn): between Oudtshoorn and Mossel Bay (‑CA), J. Dekenah 197 (NBG); Rooikrans (-CA), Van Niekerk 521 (BOL); Volmoed (-CA), Bayer in KG109/74 (NBG); SW. Oudtshoorn (-CA), Smith 5782 (NBG), Taylor 11776 (PRE); Rooikoppies (-CB), Schnettler in KG334/71 (NBG); Hazenjacht (-CB), Bayer in KG120/71 (NBG); Kamanassie Dam (-CB), Bayer in KG151/72 (NBG); 8km S. Oudtshoorn (-CA), Bayer 2099 (NBG).
Inadequately located: Oudtshoorn, Taylor in NBG981/28 (BOL), Vlakteplaas, Frey (BOL).
Haworthia mucronata var. morrisiae JDV94/44 southeast of Vanwyksdorp. In the area it grows in close association with H. arachnoidea var. nigricans.
Haworthia mucronata var. morrisiae KG90/60 west of Oudtshoorn. The chestnut brown leaf tips are characteristic.
e. var. rycroftiana Bayer Bayer, JS.Afr. Bot. 47:795(1981). Bayer :54(1975). Type: CAPE‑3321 (Ladismith): Gouritz River between Van Wyksdorp and Herbertsdale (‑DC), Bayer 1701 (NBG).
1982 – The relationships of this species ( ie. H. rycroftiana) are enigmatic. The locality in the Gouritz Valley supports the view that it is related to the H. turgida complex, especially in view of the reported occurrence of the latter species northwest of Calitzdorp. However, the general facies of the plant and its occurrence north of the Langeberg mountains suggest a more probable affinity with H. unicolor or H. habdomadis. H. unicolor is known at its nearest 10 km south of Van Wyksdorp where is grows with H. habdomadis var morrisiae. H. rycroftiana is distinguished from both of these by its more ovate and turgid leaves. It occurs at its only known locality on a steep slope where it is clump‑forming ‑ also reminiscent of H. turgida. The locality and appearance of the plants suggests that this species is a link between the species of the South‑western Cape and the Little Karoo. This particular area between Van Wyksdorp and Zebra, south of Oudtshoorn is entirely unknown as far as Haworthia is concerned and the middle Gouritz valley is also largely unexplored. (P.V. Bruyns has since collected from two populations in this latter area, which are patently the same species).
1999 – Cognizance must be taken of H. integra V. Poelln. as discussed and explored by Scott. This also helps to explain a population which has been distributed as a glabrous variant of var. habdomadis which occurs within the Gamkapoort. References to H. turgida in the 1982 seem to be quite erroneous because that species does not have the translucent margins and keels to the leaves. It also is not known north of the Langeberg mountains as suspected from records. Also around Calitzdorp, var. integra may include spined forms. There is, and will be, probable difficulty in separating these spined forms from H. arachnoidea var. nigricans and more attention to this problem is required.
Distribution: 3321 (Ladismith): 23km E. Ladismith (-AC), Bayer in KG77/71 (NBG); Kruisrivier (-BD), Stayner in KG857/60 (NBG); Vensterkrans, Algerynskraal (-CA), Laidler 376 (NBG); Muiskraal (-CC), Chisholm in NBG774/38 (NBG); Gamka East (-DA), Blackburn (BOL); On Gamka East road (-DA), Scott 1050 (PRE); Danielskraal (-DA), Bruyns 2219 (NBG); Waterkloof (-DA), Bruyns 2231 (NBG); Badspoort (-DB), Bruyns 3724 (BOL); Gouritz River between Van Wyksdorp and Herbertsdale (‑DC), Bayer 1701 (NBG). 3322 (Oudtshoorn): N. Oudtshoorn (-AC), Smith 5089, 5783 (NBG); Schoemanspoort (-AD), Smith 5085 (NBG), Bayer 171 (NBG).
Haworthia mucronata var. rycroftiana JDV87/98 Waterkloof, Rietvlei. The leaves are smooth and thicker than in the typical variety. The acuminate leaf tip is also shorter.
Haworthia mucronata var. rycroftiana JDV90/62 Gouritz River Gorge, type locality for this variety.
Haworthia mucronata var. rycroftiana JDV91/6 Gamkakloof. A smaller form with shortis leaves and resembling a glabrous form of var. habdomadis.
Haworthia mucronata var. rycroftiana JDV92/57 Gamka East. This is one of the several populations where H. mucronata and H. arachnoidea are confounded.



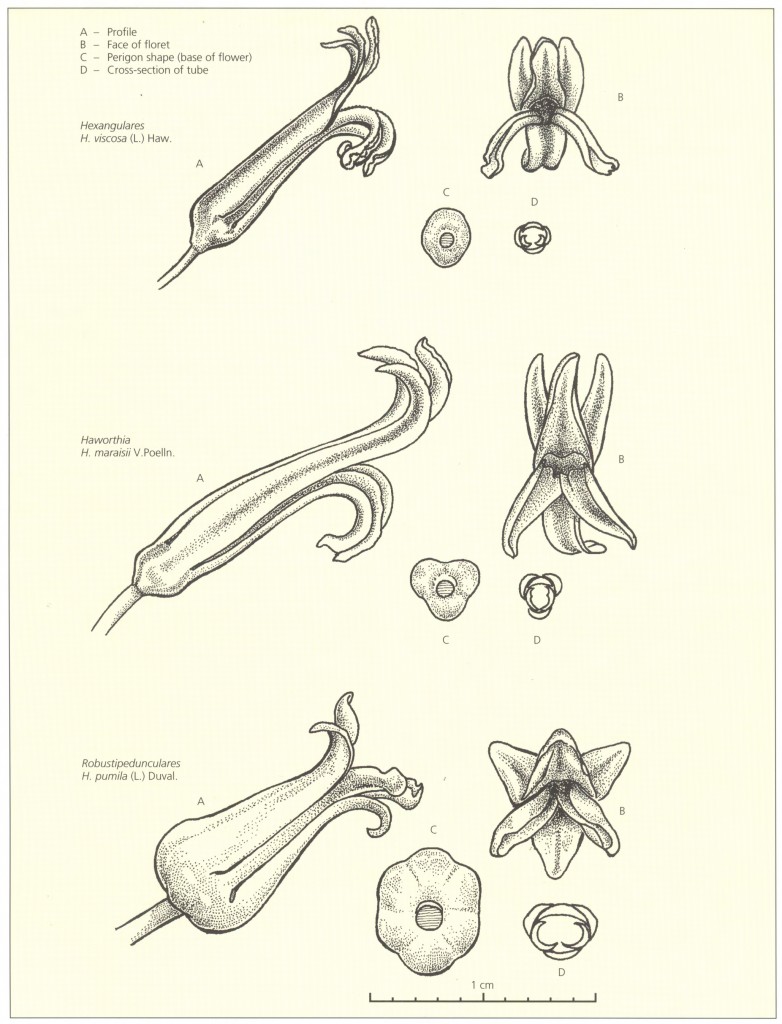

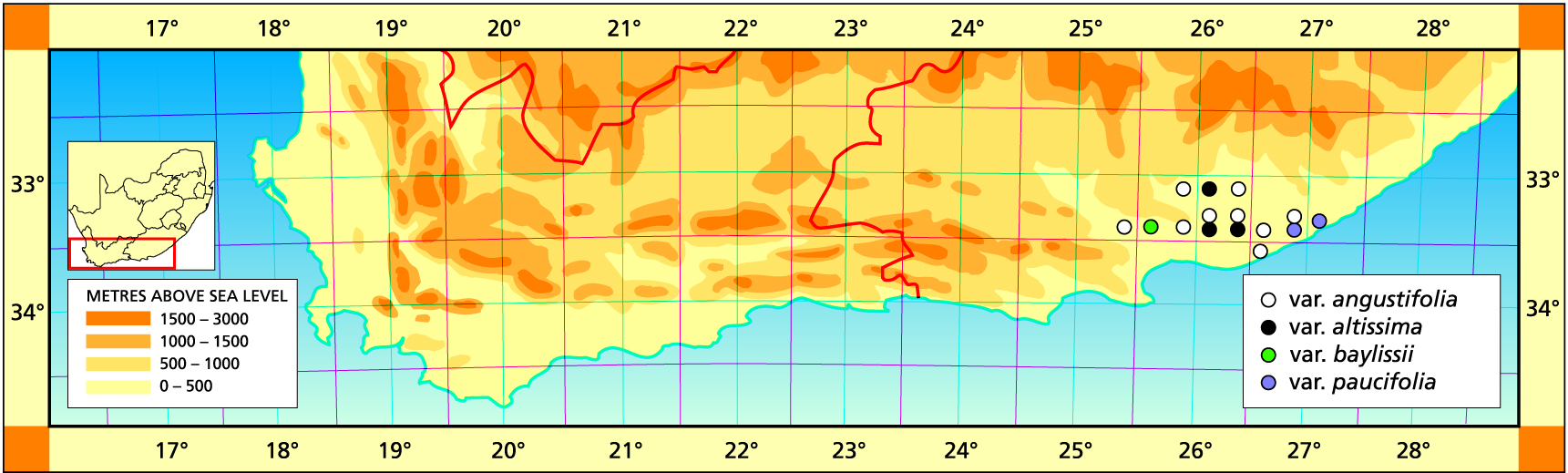







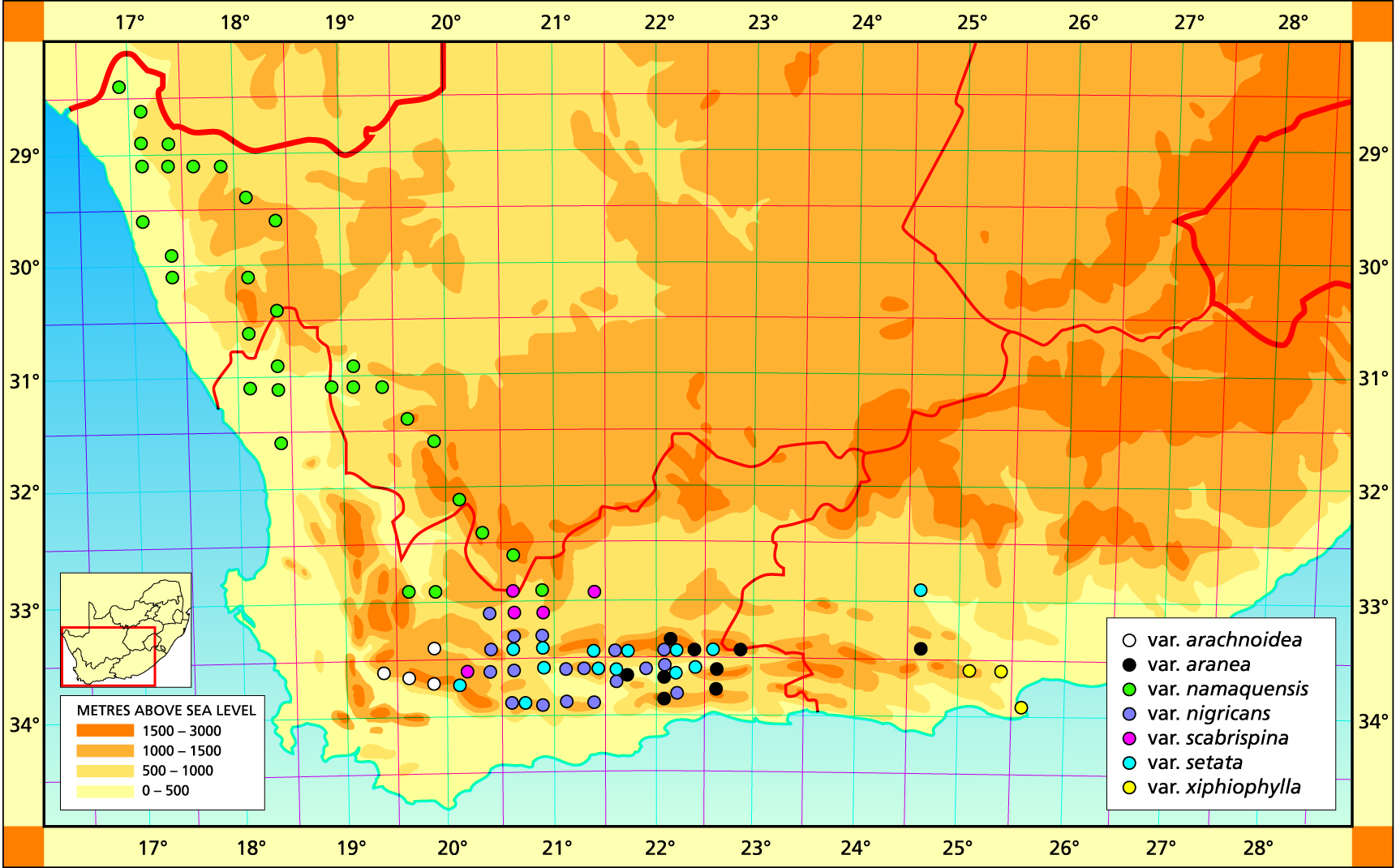























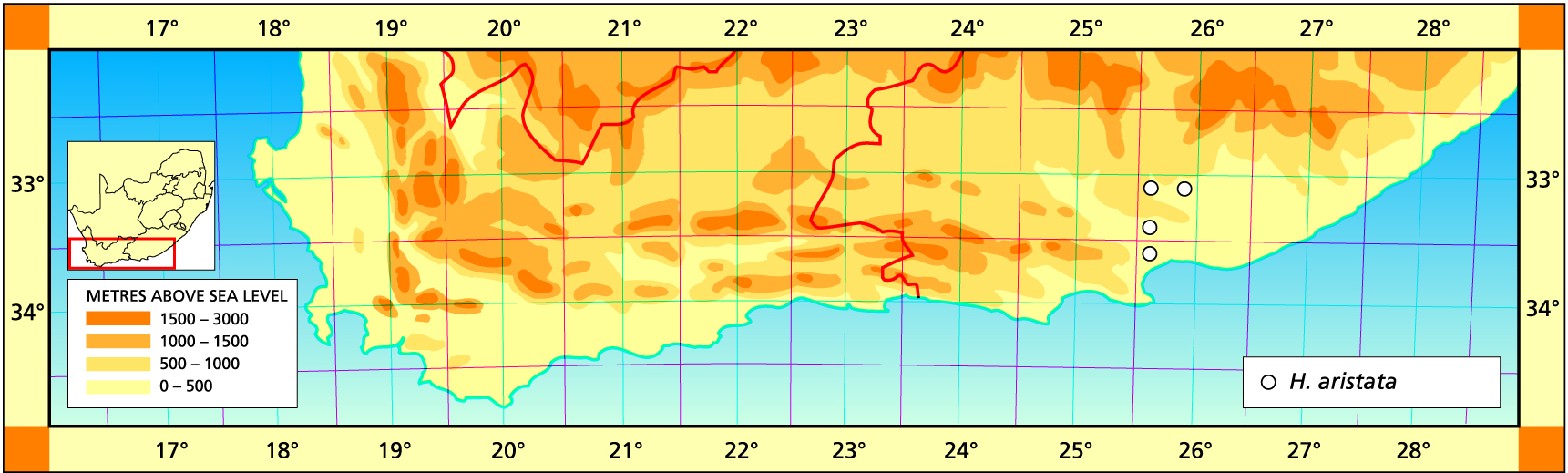


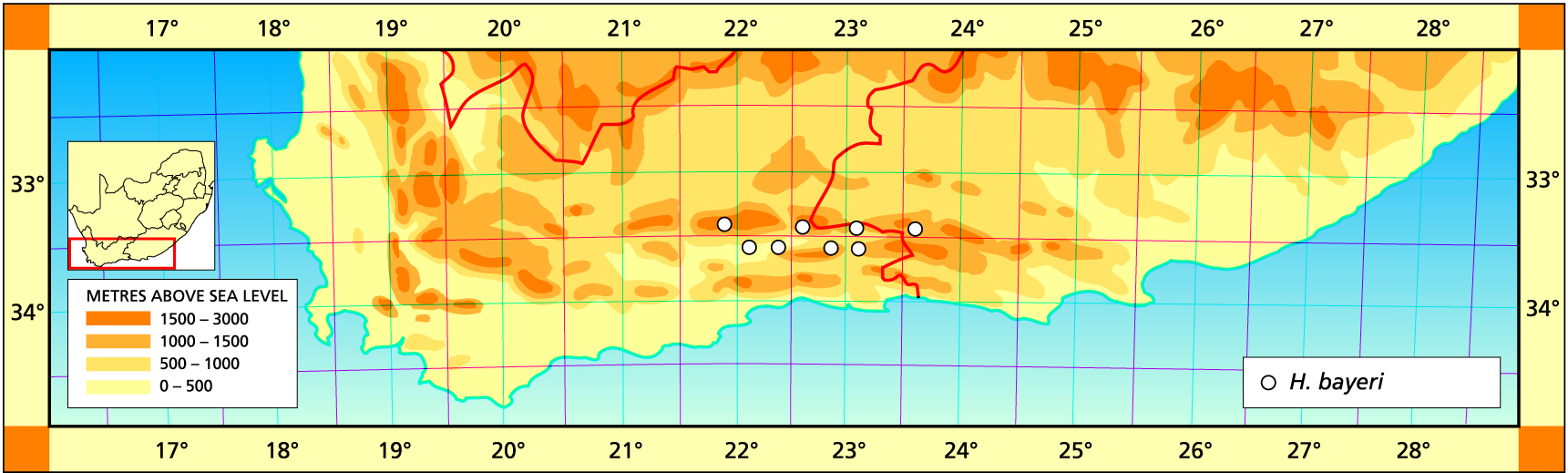




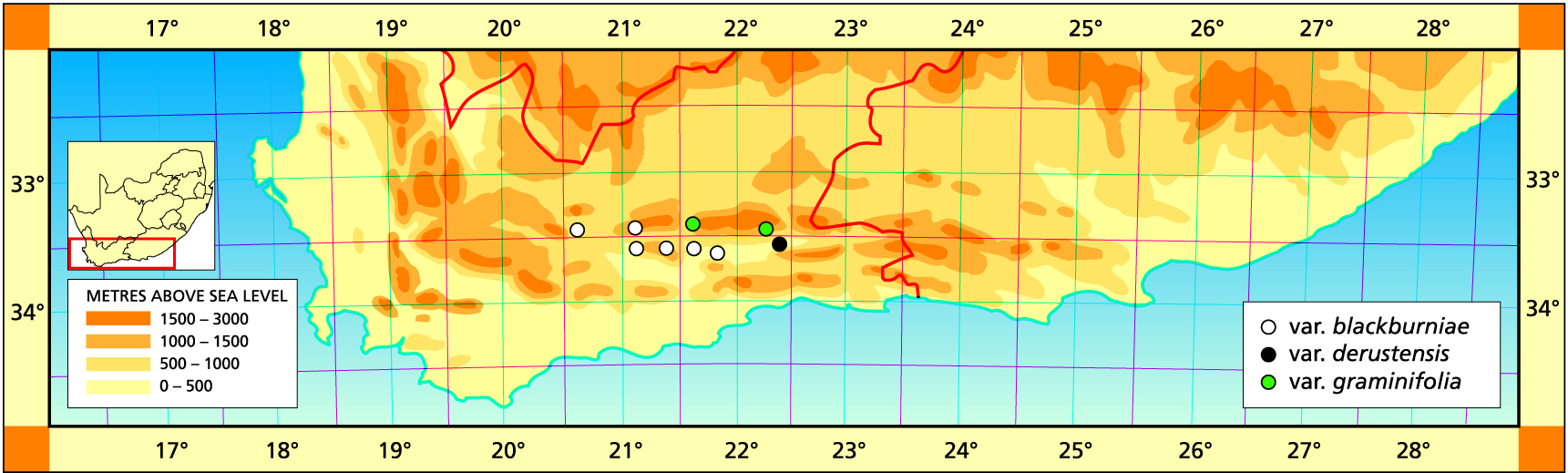







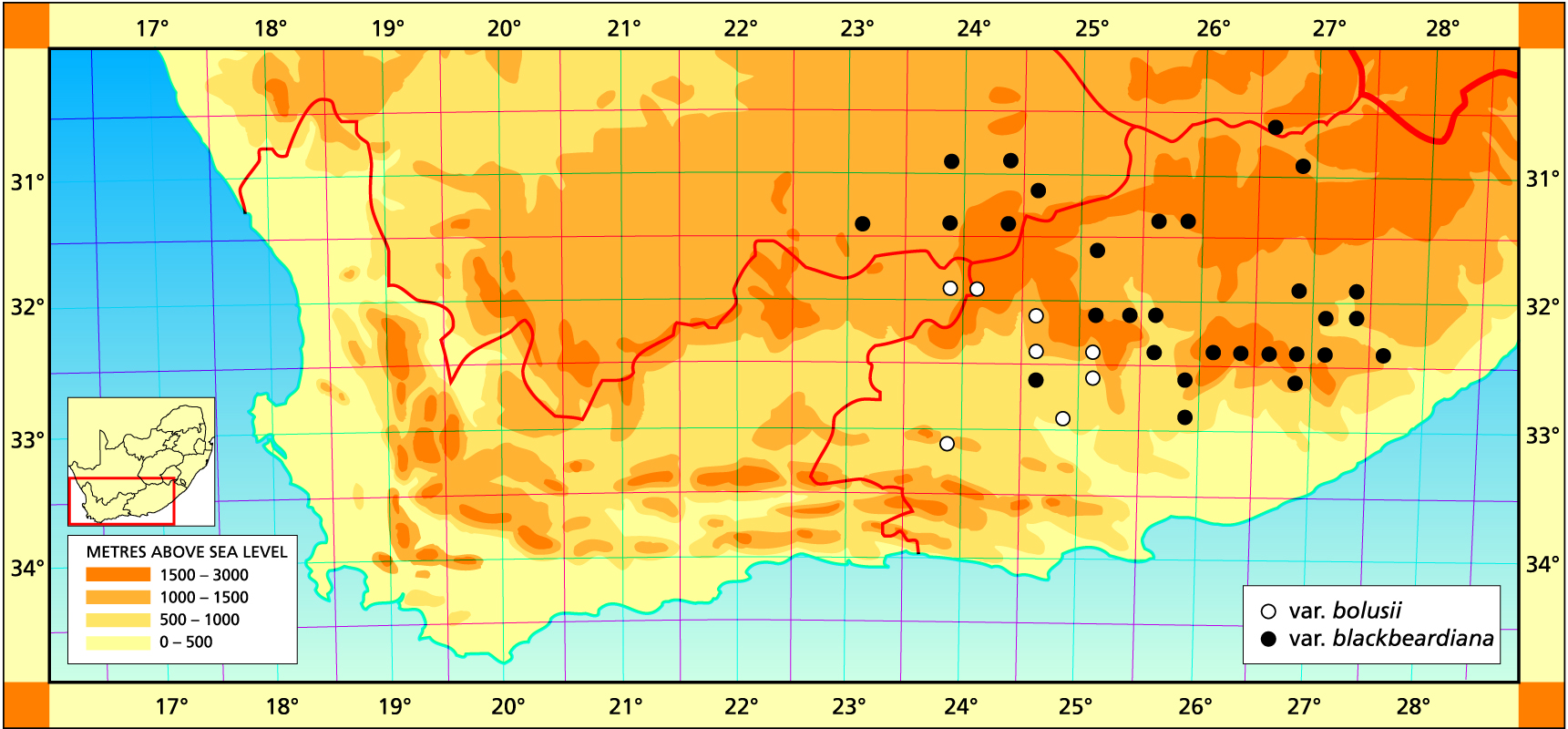





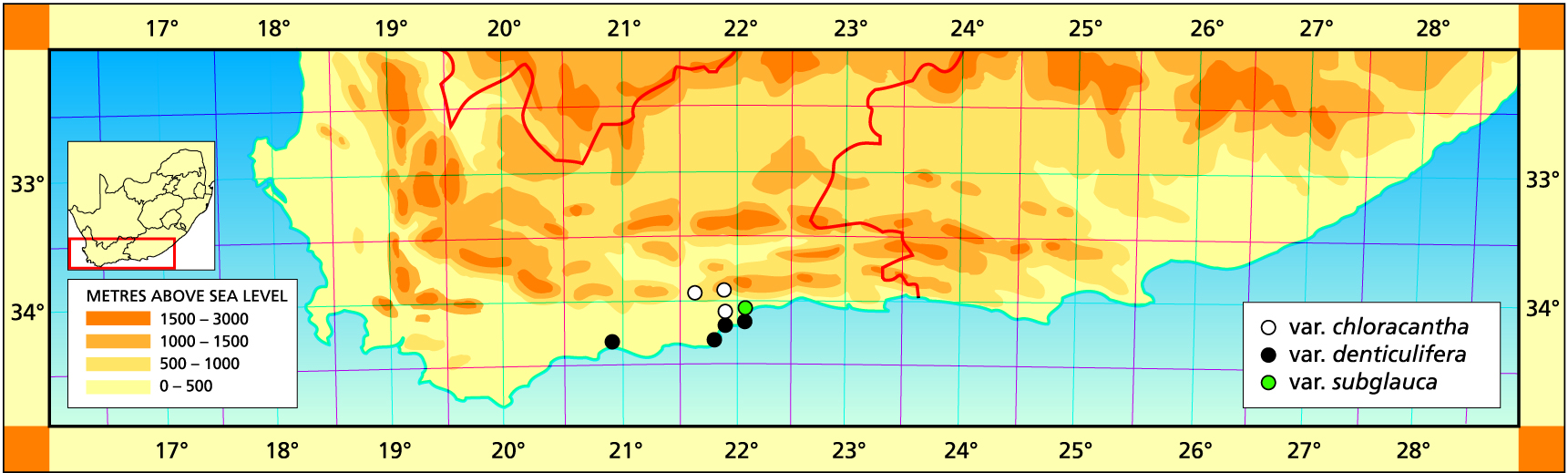







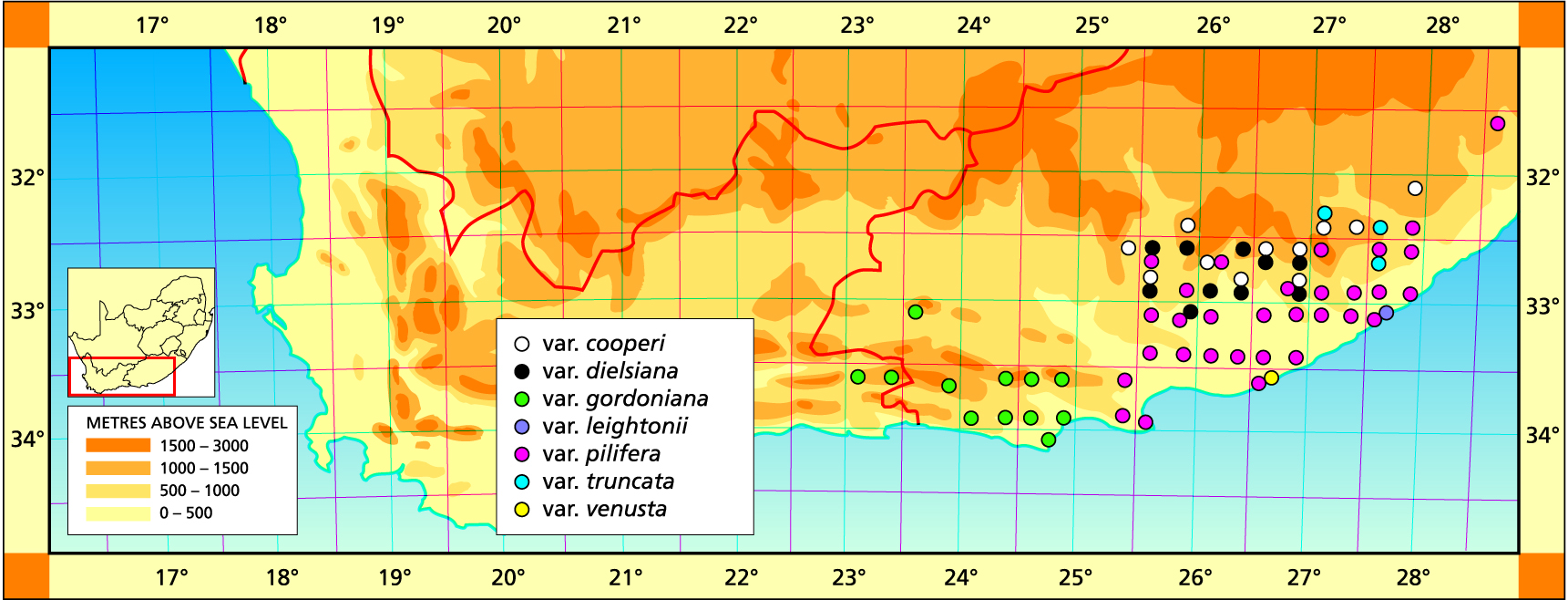



















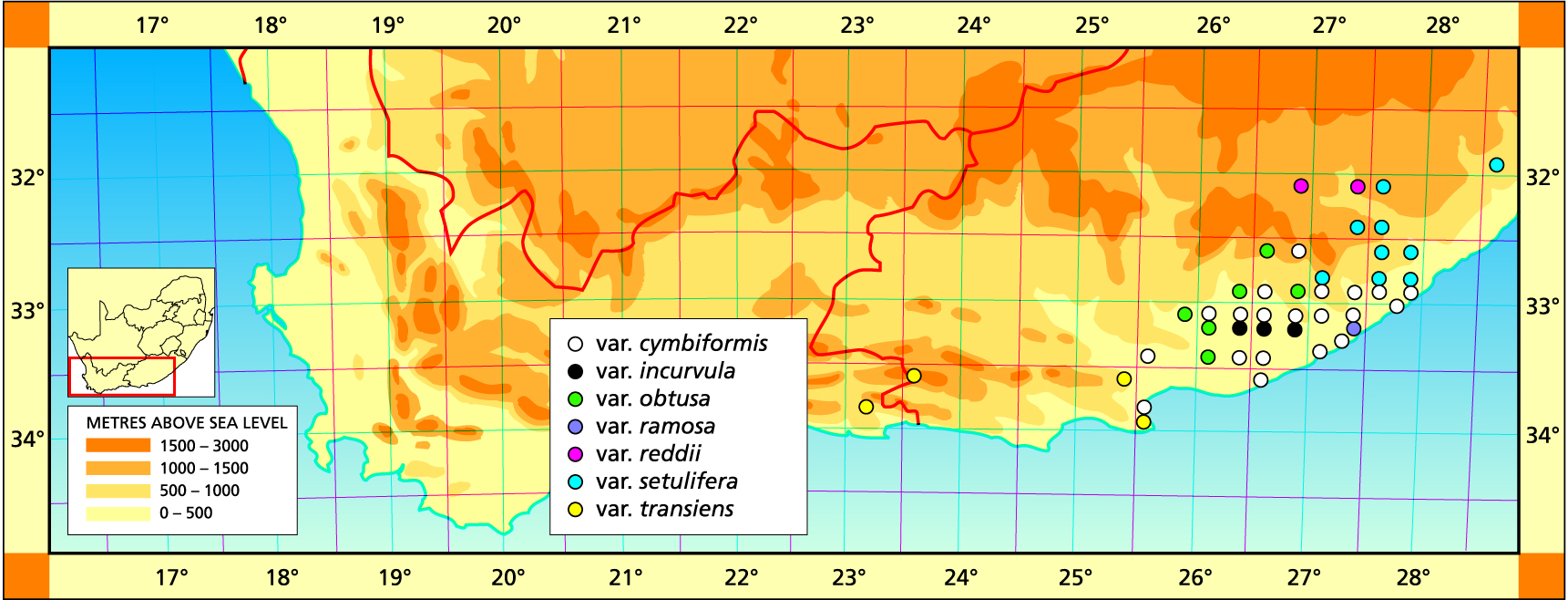












![H. cymbiformis - Curtis’s Botanical Magazine, vol. 21 t. 802 (1805) [S.T. Edwards]8067](https://haworthiaupdates.org/wp-content/uploads/1999/01/H.-cymbiformis-Curtis’s-Botanical-Magazine-vol.-21-t.-802-1805-S.T.-Edwards8067.jpg)
![H. cuspida - Addisonia, vol. 23 t. 741 (1954-1959) [M.E. Eaton] 162903](https://haworthiaupdates.org/wp-content/uploads/1999/01/H.-cuspida-Addisonia-vol.-23-t.-741-1954-1959-M.E.-Eaton-162903.jpg)