Printed in National Cactus and Succulent Journal, 34:28 (1979).
Part 4: HAWORTHIA EMELYAE von Poelln.and a new variety.
M. B. Bayer, National Botanic Gardens of South Africa, Karoo Gardens, Worcester.
Abstract: The retention of the name H. emelyae v. Poelln. over H. picta v. Poelln. and H. correcta v. Poelln. is discussed. There is also a discussion of the relationship of H. emelyae to H. comptoniana Smith. to H. springbokvlakensis Scott and to H. magnifica var. major (Smith) Bayer. A new variety, H. emelyae var. multifolia is described.
Introduction: Haworthia emelyae was described by von Poellnitz in 1937 and named in honour of Mrs Emely Ferguson whom von Poellnitz presumed to have been the collector. Von Poellnitz related his new species to H. willowmorensis v. Poelln., to H. ryderiana v. Poelin. and to H. whitesloaneana V. Poelln.. None of these names is currently upheld (see Haworthia Handbook. Bayer, 1976) and it would be extremely difficult to form a picture of H. emelyae from any vision conjured up from the little tangible evidence associated with them. Similarly the variety H. emelyae var. beukmannii v. Poelln. is known to be a variant of H. mirabilis Haw. (see Bayer, Excelsa 7: 37, 1977) and offers no solution to the problem. There are three photographs supposedly of H. emelyae in the H. G. Fourcade photographic record of Major F. R. Long’s collection. This record was compiled in 1940 and one would have expected less discrepancy at this distance in time than the three photographs actually project. One photograph is of a plant sent in by a collector and recorded…”from the Gamka river, Calitzdorp”. The plant is obviously H. turgida and comparable with the forms of that species from along the lower tributaries of the Gamka River (which actually becomes the Gouritz River after its confluence with the Olifants River in the Rooiberge to the north-east of Vanwyksdorp). The second photograph is of a plant in the collection of W. R. van Ryneveld which is also clearly H. turgida comparable with the Lower Gouritz and Mossel Bay forms. The third photograph is of a plant recorded as from Mrs E. Ferguson herself and this is unmistakably H. reticulata Haw.
Previously published Cact S.Jl 52.1 1980
M. B. Bayer, Karoo Botanic Garden, National Botanic Gardens of South Africa
Haworthia nitidula was described by von Poellnitz in Desert Plant Life (11:192, 1939) from plants collected by Major H. Venter. Venter was very generous with his localities and like most of his others, this no. 15 was also simply cited… “in the environs of Worcester, Swellendam, Caledon and Bredasdorp”. This is an ideal example for demonstrating the problems of identification of haworthias and especially in the section Retusae Haworth. J.R.Brown illustrated H. nitidula in the Cactus and Succulent Journal (18:89, 1946) but apart from this and the original illustration accompanying the description, the plant has not figured in the succulent literature. In G.G.Smith’s records, the species and nine varieties are distinguished. The interesting thing is that these varieties are drawn from three geographically separated species. The object of this article is to present Smith’s photographs and notes of his “varieties” and discuss their actual position as they relate to distribution.
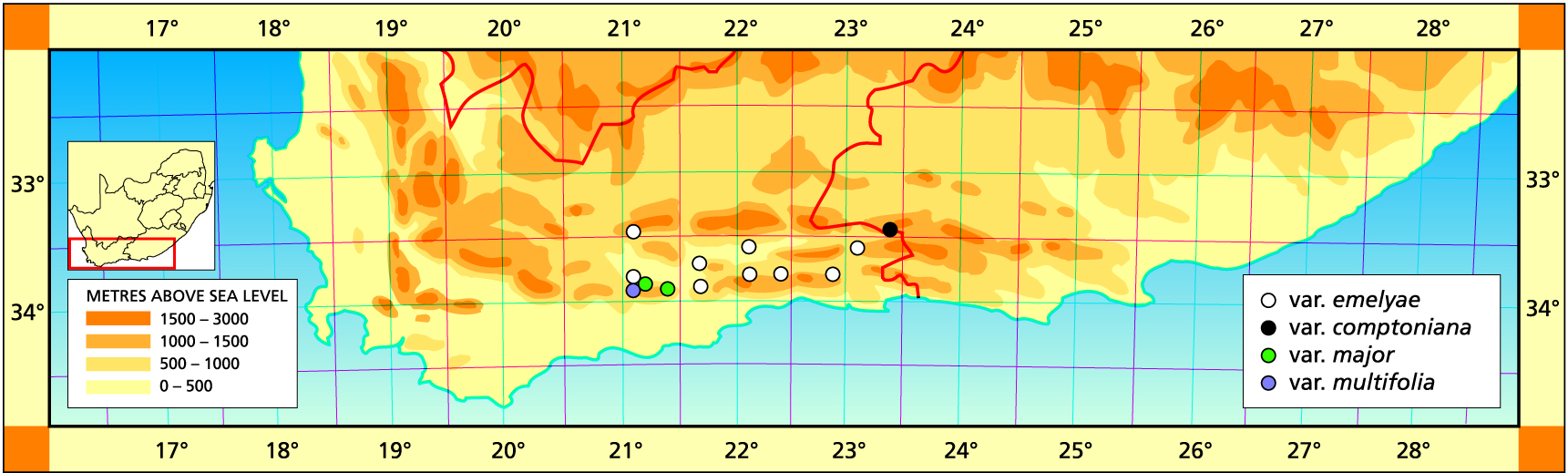
11. Haworthia emelyae V.Poelln., Feddes Repert.Spec.Nov. 42:271(1937). Bayer, Natn.Cact.Succ.J 34:28(1979). Bayer :115(1976). Bayer :38(1982). Type: Cape, locality unknown, Mrs E. Ferguson in Long 322. Not preserved. Lectotype (B&M): icon (B): H. blackburniae V.Poelln., Kakteenk. 9:132(1937). Nom. illegit. non Barker 1937. H. correcta V.Poelln., Feddes Repert.Spec.Nov. 43:103(1938). Type: Cape, Calitzdorp, Mrs E. Blackburn in Triebn. 978. Not preserved. Neotype (designated here): CAPE-3321(Ladismith): Rooiberg, E. Vanwyksdorp(-DA), Mrs Schnettler in KG 335/71 (NBG): H. picta idem. 44:133(1938). V.Poelln., Desert.Pl.Life :126(1939). Type: Cape, Moeras River, Mrs S. Blackburn in Triebn. 1062. Not preserved. Neotype (designated here): icon, Fourcade 72 (NBG).
emelyae: for Emely Ferguson.
Rosette to 10cm φ, seldom proliferous. Leaves 15-20, distinctly retused, pointed, barely translucent, with scattered elongate small flecks, with obscure raised tubercles, lined, reddish-brown hued. Inflorescence simple. Flowers 15-20, white.
1982 – H. emelyae is a widely distributed species occurring from Uniondale through the Little Karoo to west of Ladismith. It occurs in a variety of different geological formations and varies accordingly. The plants have very short recurved leaves with convex end areas usually flecked with pinkish markings. The forms east of Oudtshoorn usually have rounded end areas and are scabrid on the upper leaf faces. Westwards the leaves become pointed and smoother. However, there are clear signs of intergradation with H. magnifica in the Muiskraal area of Riversdale. Doubt has been cast on the origin of Mrs Ferguson’s plants but G.G. Smith’s records clearly indicate that they came from Van Wyksdorp and that they were collected there by a Mrs Le Roux. The var. multifolia has up to 60, more slender, suberect leaves; and was compared in the original description with H. serrata. Other forms of H. emelyae where the few leaves are shortly recurved and round‑tipped can similarly be compared with H. magnifica var. atrofusca. The eastern forms with the flatter, rounded leaf tips can be compared to H. bruynsii and H. springbokvlakensis. H. emelyae is generally non‑proliferous.
1999 – The 1982 discussion is fraught with error as two species are actually involved. The scabrid element referred to above has since been excerpted as H. bayeri and it is fairly obvious that there can be no extension to H. bruynsii which belongs in the Hexangulares. The problem is compounded by the fact that von Poellnitz described three species all of which are synonymous, and yet may have cited H. bayeri among these. The integration with H. magnifica was through the var. major which is now transferred to H. emelyae because of the intermediate populations mentioned above which occur just east of, and in the closer vicinity of, Muiskraal.
a. var. emelyae. The typical variety is rather strongly flecked and the leaves are always retused to horizontal and close to ground level. The neotype cited above is a specimen originating from near Vanwyksdorp and sent by the original collector (Mrs Le Roux) to G.G. Smith.
Distribution: 3321 (Ladismith): 20km W. Ladismith (-BD), NBG144768; Springfontein (-CC), Smith 5787 (NBG); 42km Calitzdorp to Vanwyksdorp (‑DA), Mrs Le Roux in Smith 5437 (NBG); Vanwyksdorp (-DA), Joubert (BOL);Rooiberg, E. Vanwyksdorp (-DA), Mrs Schnettler in KG 335/71 (NBG); SE. Vanwyksdorp (-DC), Bayer in KG257/77 (NBG). 3322 (Oudtshoorn): Mt Hope (-CA), Schnettler in KG582/69 (NBG); Moeras River (-CC), Smith 2447 (NBG); N. Robinson Pass (-CC), Bruyns in KG434/75 (NBG); Zebra (-CD), Smith 2905 (NBG); Erfpacht (-DD), Smith 5808, 6106 (NBG); 3323 (Willowmore): Uniondale to Avontuur (-CA), Smith 2935 (NBG).
Inadequately located: Ladismith, Pillans 858 (BOL); Oudtshoorn, Taylor (BOL).
Haworthia emelyae var, emelyae JDV93/28 north of Herold. Attractively marked end-areas make this a sought-after species.
Haworthia emelyae var, emelyae JDV90/120 south-east of Oudtshoorn. Plants in this population are very comparable with the var. comptoniana with much finer reticulation and a smooth leaf surface.
Haworthia emelyae var, emelyae JDV93/22 south-west of Calitzdorp. The plants get more scabrid and have small white markings in the leaves as does, for example, H. turgida.
Haworthia emelyae var, emelyae JDV84/11 Zebra, also fairly close to the population of H. bayeri.
Haworthia emelyae var, emelyae JDV90/10 south of Oudtshoorn. Usually associated with white quartz outcrops.
Haworthia emelyae var, emelyae JDV93/28 north of Herold. This population has been decimated by road construction and subsequent over collecting.
Haworthia emelyae var, emelyae JDV90/120 south-east of Oudtshoorn. Fairly glabrous plants which strengthen the link to the var. comptoniana.
Haworthia emelyae var, emelyae JDV97/133 south of Vanwyksdorp. One of three new collections which seem to confirm continuity with the var. major.
Haworthia emelyae var, emelyae JDV93/22 south-west of Calitzdorp. Several recent collections seem to confirm the relationship of the varieties.
b. var. comptoniana (Smith) Venter & Hammer Cact.Succ.J(US) 69:77(1997). H. comptoniana Smith, JS.Afr.Bot. 11:76(1945). Bayer 108(1976). Bayer :33(1982). Scott :128(1985). Type: CAPE-3323 (Willowmore): Georgida (-AD), M. Malherbe in Smith 3433 (NBG).
comptoniana: in honour of Prof.R.H. Compton.
1982 – The species with the flattened ends to the leaves are the most sought after, and H. comptoniana is particularly attractive. It occurs in quartz patches and the plants are often well hidden under the stones. The plants are bigger than in H. emelyae (the nearest relative), growing up to 120 mm in diameter in cultivation. The growth form parallels that of H. emelyae exactly and the difference is that it is a bigger and entirely smooth species. The leaf end area is slightly pellucid and reticulated. Where H. emelyae is generally tinted purplish-brown, H. comptoniana is green. Distribution and abundance is not fully known and H. comptoniana is considered very rare in the field. It has proved very easy in cultivation and good seed is easily produced by cross-pollination.
1999 – This variety grows in very close association with H. bayeri but it is not known if other populations exist other than at the type locality. Some forms which are pale coloured and in which there is little contrast between the reticulation and the background colour and translucence of the leaf, are very plain. Other plants are darker coloured or with more conspicuous specks and marked reticulation, and they are are much more attractive.
Distribution: 3323 (Willowmore): Georgida (-AD), M. Malherbe in Smith 3433 (NBG), Smith 3433 (NBG), Bayer in KG114/72 (NBG).
Haworthia emelyae var. comptoniana JDV90/8 Georgida. Recorded only from one population and severely threatened by collectors.
Haworthia emelyae var. comptoniana JDV90/8 Georgida. Also very cryptic in the summer months.
Haworthia emelyae var. comptoniana KG114/72 Georgida. The reticulation and relative length of the leaves are quite variable.
c. var. major (Smith) Bayer comb.nov. H. schuldtiana var. major Smith, JS.Afr.Bot. 12:1(1946). H. maraisii var. major (Smith) Bayer :132(1976). H. magnifica var. major (Smith) Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). Type: CAPE‑3321 (Ladismith): Garcias Pass (‑CC), Smith 5370 (NBG).
major: greater.
1982 – The var. major has an unusually large flower similar to that of H. emelyae and there is evidence of intergradation of these two taxa.
1999 – Although in this variety the leaf tubercles are armed with a spine, the general form of the plants is that of H. emelyae, and populations east of Muiskraal confirm this relationship. The change in relationship forged here has its roots in the new collections of H. magnifica around Riversdale and in the new arrangement regarding H. maraisii. Another consideration is the position of var. paradoxa, mentioned in 1976, and that is its relationship with H. mirabilis. Similarly the following variety and its position need also to be considered.
Distribution: 3321 (Ladismith): Garcia’s Pass (‑CC), Smith 5370 (NBG), Garcia’s Pass (-CC), Dekenah 9 (PRE); Muiskraal (-CC), Smith 3458 (NBG), Bayer in KG118/71 (NBG); Riversdale (-CC), Muir in NBG164/25 (BOL); Sandvlakte (-CD), Bayer in KG138/72 (NBG).
Haworthia emelyae var. major JDV90/19 Muiskraal. A sandstone-dwelling variety with spines on the leaf surfaces.
Haworthia emelyae var. major JDV95/36 Muiskraal. The var. multifolia is now know to occur within a few kilometers of the var. major and these two may not vary continuously.
Haworthia emelyae var. major JDV95/23 Garcia’s Pass. A densely spined seedling grown by P. Bosch and coming from east of the type locality.
Haworthia emelyae var. major JDV90/22 east of Muiskraal. Several quite variable populations associated with shales.
d. var. multifolia Bayer Natn.Cact.Succ.J 34:31(1979). Bayer :39(1982). Type: CAPE‑3321(Ladismith): Springfontein, Riversdale (‑CC), Bayer 1558 (NBG).
multifolia: many leaved.
When this variety was described it was associated and compared with H. serrata. A new population is now known a little further to the west and collections by J. Dekenah suggest that it is possibly a little more widely represented in that general area. The plants have many more upright leaves and the relationship with H. serrata through H. heidelbergensis, which is also now better understood, becomes real. There is a strong suggestion that these links all also involve H. mirabilis and this is perhaps supported by von Poellnitz’ interpretation and citations of varieties of that species.
Distribution: 3321 (Ladismith): Springfontein, Riversdale (‑CC), Bayer 1558 (NBG); W. Springfontein (-CC), Smith 5769 (NBG); E. Springfontein (-CC), Smith 5768 (NBG), Smith 5389 (NBG, PRE), Bayer 1558 (NBG); Muiskraal (-CC).
Haworthia emelyae var. multifolia JDV87/162 Springfontein. Since located at two sites east and west of the type locality.
Haworthia emelyae var. multifolia JDV94/32 Brandrivier. In this population the plants are more robust.
Where are we? Geography plays such a critical role in my perception of the species that it is important to try to understand why and how it touches on the issue. The Langeberg Mountains is a Table Mountain Sandstone about 10km wide and 1500m high running east-west and separating the inland Little Karoo from the Lowland Renosterveld of the southern Cape. There are five main travel routes through those mountains of which two are via river gorges viz. Cogmanskloof, Tradouw, and then there is also the Gouritz River Gorge where there is no road. Haworthia is not generally considered a sandstone and high mountain species because, firstly, there are few records to suggest that and, secondly, because they are averse to the higher moisture levels. However, there are many records in the low and close foothills. H. retusa ‘turgida’ is recorded in the higher areas and in the vegetation associated with the sandstone viz. the Fynbos. Fig. 1 is a view taken looking eastwards from Kleindoorn (Kleindoornrivier). This is about 16km east of Barrydale that sits at the northern end of the Tradouw Pass. The next farm is Brandrivier (B) and beyond that is Springfontein. Muiskraal is marked with an “M” and this is at the northern end of Garcia Pass from Riversdale to Ladismith. The “O” marks Oskop which is beyond Zandkraal and about 15km beyond Muiskraal. Another 10km will take you past Waterval and bring you to Aasvoelvallei at the confluence of the Grootriver with the Gouritz. The last stretch is another 10km over the Cloete’s Pass to Herbertsdale from where one travels back westward 7km to Towerlands.
What is there? I speak of species complexes because the classification and naming of Haworthia seems to defy any agreement. I do not think the nomenclatural system allows it either. South of the Langeberg are the H. retusa and H. mirabilis complexes as well as H. minima and H. pumila, as main role players. North of the mountains is H. arachnoidea and H. mucronata. There are incursions and crossing of the mountains by three of these and my contention that they do not all do so is simply because we do not see beyond the narrow confines of our classification system. North of the mountain there is enough evidence to suggest that H. mucronata and H. arachnoidea is very probably the same one complex despite the few situations where they manifest in near proximity. South of the mountains H. retusa and H. mirabilis have an even more complex relationship. It is difficult to begin to realize that there is a very small step between their manifestations south of the mountain to an integration of them both in H. emelyae that occurs north. I have great sympathy for those who contest the issue. I hope that the figures I present here, as a very small set of images from what may be a very small sample of existing populations, will help bridge the difficulty.
My field experience, however much field experience is a measure of any persons competence to process the product, suggests that there is a predictable coherence in the variation that one observes as one moves from discrete population to discrete population. This progressive variation is also driven by surface geology and habitat. It becomes no problem at all but to realize that as another set of samples, the Aasvoelvallei H. emelyae (see MBB 7850 figs 2a-l) is nothing more than the Towerlands H. retusa ‘turgida’ (MBB7849 figs 3a-b) in another guise. It is the habitats that are so vastly different. Towerlands is south facing mountain slope in sandstone and Fynbos vegetation next to a perennial mountain stream. It was no chance that the rain that caught us there stood in such sharp contrast to the warm sunshine that greeted us an hour later at Aasvoelvallei and H. emelyae a mere 10km away as the crow could fly. That habitat is Bokkeveld shale with a heavy overlay of the very white quartz that veins this formation, and the vegetation is Karoid.
2a. MBB7850 H. emelyae Aasvoelvallei.2b.2c.2d.2e.2f.2g.2h.2i.2j.2k.2l.3a. MBB7849 Haworthia retusa ‘turgida’ Towerlands.3b.
H. mirabilis ‘maraisii’ is present north of the mountains at Montagu and towards Montagu from there to Barrydale. It is also present at Barrydale itself. At Kleindoorn we have an enigma (yet another) where the plants (MBB7847 figs. 4a-s) are very similar to the Barrydale plants and they also seem to flower in summer. But these same plants can be likened to H. emelyae ‘major’ at Muiskraal that flowers in the spring as does H. emelyae (and H. retusa). Figs. 5a-c MBB7844 shows plants at Brandrivier. The dramatic thing is that these are recent photographs and very unlike the plants observed there by me 30 years ago and recognized as H. emelyae ‘multifolia’.
4a. MBB7847 Haworthia emelyae Kleindoornrivier4b.4c.4d.4e.4f.4g.4h.4i.4j.4k.4l.4m.4n.4o.4p.4q.4r.4s.5a. MBB7844 Haworthia emelyae ‘multifolia’ Brandrivier5b.5c.6a. MBB7846 H. emelyae ‘multifolia’ W Springfontain6b. MBB7846 Haworthia emelyae ‘multifolia’ W Springfontain6c.6d.6e.7a. MBB7847 Haworthia emalyae ‘multifolia’ E Springfontein7b.7c.7d.7e.7f.7g.7h.7i.7j.7k.7l.7m.8a. JDV97-44 Haworthia emelyae ‘breueri’ E. Zandkraal8b.8c.8d.8e. JDV97-44 Haworthia emelyae ‘breueri’ E. Zandkraal8f.8g.8h.8i.8j.8k.8l.8m.8n.8o.8p.
Figs 8 are of JDV97-44 (originally GDM429) plants from beyond Muiskraal (where both ‘multifolia’ and ‘major’ occur in disparate habitats) at Zandkraal, where there are three populations to which an appellation ‘H. breueri’ has been fabricated. I downsize that to H. emelyae ‘breueri’ to bring some perspective back to Haworthia nomenclature. I do not include field photographs for Oskop and Waterval, but add two from south of Vanwyksdorp (see figs 9a-c ADH2966) where H. emelyae originated according to the collector (this was a Mrs. le Roux and the plants were sent to Smith via Mrs. Emely Ferguson). The photographs were taken just south of Vanwyksdorp by Adam Harrower and I do hope that readers will realize from this article that it is not at all strange that the von Poellnitz published illustration and type of H. emelyae (figs 10a-b) are so often said to be of H. retusa ‘turgida’. Curiously, H. Fourcade illustrated a plant (his no.144, see fig.11) of H. retusa ‘turgida’ and another (his no. 82, see fig.13) of H. reticulata as H. emelyae. This says a lot for subsequent attempts to interpret and argue vehemently about von Poellnitz’ H. correcta (see fig.12).
9a. ADH2966 Haworthia emelyae S Vanwyksdorp9b.9c.10a. Haworthia emelyae Von Poelln.10b. Haworthia emelyae Von Poelln. Type.11. HGF144 Haworthia retusa ‘turgida’ as Hawortha emelyae12. Haworthia correcta V. Poelln. Type.13. HGF82 Haworthia emelyae ‘turgida’ as Haworthia emelyae
What really struck me on this recent excursion was just how much inviting territory is unexplored. In contrast to that is Kleindoorn where we photographed those plants actually found by Gerhard on a previous visit there with Bob Kent and Kobus Venter many years ago. The plants are on the south side of a sandstone ridge. We could not find them elsewhere on our recent visit and it seems that suitable habitat is just not there for them. There is a most remarkable divide of shale and sandstone on Kleindoorn and while there is/was a form of H. arachnoidea in the sandstone (repeated at Zandkraal), only H. arachnoidea ‘nigricans’ is in the shale. At Brandrivier, E. Springfontein and Muiskraal the ‘multifolia’ is in a sandstone pebble layer that overlays the shale in the manner of the Tertiary terraces south of the mountain. West of Springfontein the plants are again in shale, but without the white quartz of other karoid sites mentioned.
Acknowledgement It is not possible to do this fieldwork without the goodwill of landowners that is threatened by a glut of collectors who often trespass. Therefore I particularly want to thank Bertus le Roux of Kleindoorn, George and Jaco Nel of Brandrivier, Louis Botha of Springfontein, Anton Bredell of Aasvoelvallei, and Gregory Nicholson of Towerlands for access to their property. Gerhard Marx was contributory as was Kobus Venter on a previous excursion. Adam Harrower kindly sent me pictures from his records. ♦
1a.View from above Onverwacht North Westward to Ladismith1b. View looking south west over Muiskraal to Garcia Pass
There used to be a regular bus service between Riversdale and Ladismith and J. Dekenah made use of this for his excursions into the Little Karoo to find plants for G.G. Smith. He thus discovered H. emelyae ‘major’ at the northern mouth of Garcia Pass. He also submitted a single specimen of a plant collected from the karoid veld a little further north. It is that record that suggested to me that ‘major’ was linked to H. emelyae further east and north as well as to ‘multifolia’ to the west. Etwin Aslander found what I regarded as the equivalent of ‘multifolia’ (see figs 2a-i) on a low plateau north of Garcia pass on the farm Muiskraal that is at the foot of the mountains north of Garcia Pass.
Johan Meyer of Barrydale once showed me a cultivated plant that he claimed he collected on the slopes to the north of Etwin’s locality. This was a robust and rather tuberculate plant that was very reminiscent of the forms of H. emelyae at Zandkraal about 12km east. In an earlier chapter I wrote more widely about the problem of land ownership and access and eventually felt that I should look into the situation at Garcia Pass in respect of conservation status of ‘major’, when an opportunity to break the impasse arose.
Fortunately I have quite a long association with Muiskaal and its owners, since I was at the Karoo garden and Mr. Henry Chamberlain, owner of Muiskraal, finally allowed me access. We visited both Etwin Aslander’s population (MBB7858 see figs 2a-i) as well as locating a population on the northern slopes as indicated by Johan Meyer (MBB7859 see figs 3a-i).
3d. MBB7859 Muiskraal3f.3g.3h.3i.1d. Looking eastwards over Muiskraal1e. Looking southwest to Garcia Pass from same spot as above
In addition we also found a population immediately west of the farmstead (MBB7857 see figs. 4a-f). Particularly interesting is the dramatic switches in dominances of shrubby plant species that must be a factor of differences in the very skeletal soils as well as how plants compete with one another (allelopathy). The plants in all three populations are of course similar to the east of Springfontein plants in as much as one generates a mental common denominator for a set of variable plants. This similarity must surely be seen in the similar geologies of the three sites which are on tertiary depositional terraces. We also explored extensively along the foot of the mountain between Muiskraal and Zandkraal without any success.
1f. West Muiskraal
4a. MBB7857 West Muiskraal4b.4c.4d.4e.4f.
However, we also made contact with the owner of the upper areas around Garcia Pass itself and were able to explore both sides of the road at the ‘major’ locality (see MBB7861 see figs 5a-l) as well as the range eastwards.
We did find a new population some 3km east of the farm Onverwacht (see MBB7860 see figs. 6a-i) and Gerhard Marx reports seeing it immediately north of the same farm. We did on a subsequent visit briefly get there, but did not find plants. Some of the plants were very ordinary and it is not surprising that J.R. Brown, in commenting on his collection, felt that he had two species viz. H. ‘major’ and H. maraisii as he might have named them. Definitely the plants I saw could have been mistaken for individuals in several populations in the sandstones south of MacGregor or even nearer Villiersdorp (Moddergat). Unfortunately my camera failed me and I did not get many pictures, but one can see that the plants can have relatively spineless leaf surfaces.
There is something curious about the flowering times. The ‘major’ plants had evidence of recent flower stalks on a few plants while some plants were also beginning to flower. The lower populations at Muiskraal and Zandkraal were already shedding seed. I think it is significant to point out the geological features that I think are the main drivers of ‘difference’. Garcia Pass is a rift through the Langeberg Mountains that are uplifted sand stones. The northern foothills are an eroded remnant of tertiary deposits that overly Bok keveld shale. The Zandkraal populations are associated with quartz outcrops or variants of such as are common throughout the Little Karoo. The vegetation differences between the sandstones, terrace gravels and shales are astounding and illustrated in the fact that botanists, following their original intention that a biome was a major association of plant species driven by massive and extensive landforms and climate, regard the Karoo vegetation and Fynbos vegetation as belonging to two different biomes. The terrace vegetation is primarily Renosterveld, but it has elements of what is now regarded as the Thicket biome. It would not be a serious overstatement of fact to say that no two plant species occur in each vegetation type. The farmer on Onverwacht (Fynbos) is a flower farmer picking from the field, while the present farmer on Muiskraal (generally Karoo) grows olives and grapes. Muiskraal used to be primarily a sheep farm and Onverwach barely.
Despite this observation on compositional differences of the vegetation, or because of it, my contention is that ‘major’ is a variant in the sandstones while ‘multifolia’ is an expression of the same species, H. emelyae, on the tertiary debris. They both go on to varied forms in quartzitic patches in the shales. In my opinion H. emelyae is in a continuum that is formed out of the H. retusa and H. mirabilis complexes south of the Langeberg Mountains, where broadly speaking H. emelyae moves into sandstone as ‘major’ and the other two species occupy that terrain as ‘turgida’.
Acknowledgement: Sincere thanks to Mr. Henry Chamberlain of Muiskraal and Mr. Pierre Vermaak of Onverwacht. Gerhard Marx also motivated with interest and information. ♦
1 December 201116 July 201215 September 201213 December 201213 April 2013August-2013
Here is an interesting series of pictures showing Haworthia leaf replacement over only 16 months. The plant is in an outdoor rockery. Leaves are numbered from oldest to youngest. Leaf loss has not been quite sequential as the 7th leaf outlived the 5th. In the growth condition of September 2012 the leaves show three tiers viz. 1,4,7,10,13 / 2,5,8,11 / 3,6,9,12. The figure for Dec 12 illustrates the “5-tiered” artifact of the spiral sequence with leaves 15/10, 14/9, 13/8, 12/7 and 16/11/6 in each tier.
These pictures demonstrate that the plants grow quite fast. They may not be very long-lived in nature. I estimate a life span of 15-20 years?
The first picture of this plant was taken in Dec 2011 when the leaves were numbered 1 to 10. Here in March 2014 leaves 1 through 12 have died away and there are 10 new leaves. At the same time the stem will be stretching as roots are replaced in a similar sequence. The life of the plant is going to be determined by the way in which the old stem and roots decay and how that decay will affect the ultimate health of the whole plant.
In the field it seems as though the decay of the old leaves and stem is well regulated as a sort of dry composting. In cultivation this can vary from this state to when all the roots and even the base of the stem can rot unhealthily. In the field one does finds moribund old plants that are at the end of their life and with large aged stems. Just how long the plant can maintain good health in either the field or in cultivation is an unknown. A grower just has to depend on experience and feel for his/her plants and this is why some time back I guessed at the normal life of one of these ‘retuse’ haworthia at about 10-15 years. Some things like Tulista pumila and Haworthia coarctata are quite different as the stems can elongate and re-root along the ground. Plant of H. coarctata more than 3 meters in diameter must be very old. Even H. reticulata or H. herbacea do this – look at H. cymbifomis ‘ramosa’. What you can consider is that new leaves and flowers follow a regular process and it is useful to remember that the plants go through a non-growing stage in the year. I tend to think most haworthia grow in the spring and autumn disliking the heat of summer and the cold of winter. But this is just a generalization that points to things that need to be considered. All species are not the same. Good root health is also associated with drainage and adequate air space in the growing medium.
In species like coarctata and reinwardtii – and I doubt that these are two different species – the plants easily lose obvious connection to the mother clone and a single clone can come to occupy a very large area. H. cymbiformis and H. angiustifolia can also spread widely by pieces breaking off to establish somewhere else. H. limifolia does it by stoloniferous expansion as can H. marumiana and H. zantnertiana. But coming back to the solitary non-pupping growth forms it would be interesting to know just where the essential life of the plant resides. Sometimes the stem just seems to get too thick and cutting it back too far also cuts away the support and source of the very young and new centre leaves.
Addendum: 10 March 2023 New leaf formation is an annual event as is new root and stem growth. So the life of the plant is a delicate balance between decay of old and re-placement by new. There are always moribund old plants and there are sometimes seedlings. Seed is not widely and randomly dispersed. They occur less frequently as distance from the parent plant increases. Also, germination and survival in any year is a gamble. Anyone with field familiarity will have observed that no two years are the same. Some events are once-in-forever events. Like the field of red Droasanthemum speciosum one year east of Worcester, or the mass display of pink Drosanthemum micans that is normally yellow there (this yellow variant has the name D. halli). The fields one year of yellow – Moraea gawleri. Another year, rings of pinkish red Ruschia carolii around the termitaria. One year flowering Stapelia thudichumii everywhere in the Tanqua Karoo. And so on. ♦
Incidentally both the soft shale bands within the Sandstone strata as well as the Bokkeveld Shale above, decompose to kaolinic clay.
I took so many pictures that I will just post because I was so fascinated myself. The population is in an area quite new to me and wholly empty as far as haworthia records go – but perfectly predictable if you throw out the chaff. In the meantime, keep thinking.
The plants were all in much the same situations among sparse restioids and grass and very few other succulents. Next I will post better pictures of a few plants and explain a bit more.
What do we have so far? mirabilis, ‘magnifica’, atrofusca, pygmaea, comptoniana, esterhuizenii, Quite good actually – but some serious omissions. Any more offers – these are all excellent.
Just to make sure the name “magnifica” makes sense, here are a few pictures taken at the type locality.
While looking at those magnifica pictures I was struck by these two to show how a retusa influence is present.
A closer look at the new find.
The scenery
Those flat top hills are the remnants of an ancient african plateaus prominent from near Barrydale to beyond Uniondale – north of the Langeberg. – silcrete upper layer like the concreted layers also present topping the inselbergs south of the Langeberg.
So what is it? If H. magnifica makes sense to anyone as a species, that person is lost to me. This new population is in the sequence from Muiskraal eastwards and it is H. emelyae (‘breueri’, ‘wimii’). The identifications put forward are nearly all in the context of H. mirabilis which includes ‘magnifica’. This naturally supports my perceptions that H. emelyae is in fact the karoo extension of H. mirabilis. More widely that H. mirabilis, pygmaea, retusa and emelyae form a single system. If one reads in the Updates the reports of Towerlands/Aaasvoelkrans one will appreciate the connection better. The “magnifica’ as a species myth is simply and easily dispelled by studying the floral data presented in updates. There is no reason why there should be any doubt and confusion that the cabal so enjoys and wallows in. Aloe barbara jeppeae indeed.
The truth of the matter is that there is no taxonomic solution other than recognising this and stop prostituting science in the name of populism. There are hundreds if not thousands of people interested in Haworthia, and millions affected by the formal names of plants. I see some responsibility in respecting this fact and even more so in trying to understand what all this marvelous stuff really means. It comes down to what a species truly is and how it is defined. If I could bring just one person to that realization I might feel comfortable – with no one caring a damn, I am not. :) I have not far or long to go now and I will be posting more on this topic before I am finished.