In trying to close my long history as a writer about Haworthia, I am anxious that my closing thoughts are more understandable than my opening ones were. Update 4 calls for a “paradigm shift”- for me as much as anyone else. My friend Kobus Venter has written … ”I feel that a lot of important descriptive and analytic content gets lost by reducing the species to this level (of superspecies)”. My intention is completely otherwise. My view has altered in the sense that I recognize and admit that all these new names and descriptions of writers like Masa Hayashi or Ingo Breuer are extremely useful and informative and that we are losing content in arguing and disagreeing about them. My contention now is that our problem lies in the fact that we are trying to explain and understand these complex systems things within the context of a restrictive nomenclatural system that is not designed for the purpose. It is confounded by the problem that as a society we do not understand what species are.
The formal nomenclatural system and the use of Latin binomials is too much for us and it does not work for us as a community. One of my critics went so far as to say that she was “turned-off” by my use of haworthia plant names where I omitted to write the prefix “H” in an article which was about nothing else but Haworthia! This in my view is pedantic nit-picking that the code cannot exclude. The same person wrote that the nomenclatural system is very precise and prevents us from getting in a mess. I think we are in a mess because we try to force it to do something that it is not designed to do. We should be more intent on trying to apply the horticultural system that is intended to cope with cultivars, as is now more evident in the literature. The difficulty we face is that the plants are highly variable IN THE FIELD, and we have yet to do more than scratch the surface of that variability.
The code articles that insist on chronological priority are also to my mind problematical (Dr. Paul Forster wrote…”the current framework (ICBN) for decisions is riddled with flaws…” (So there are other problems too?). This is because an earlier name, which may be based on a wholly atypical plant or population, has to be chosen for a species where a later name may be far more appropriate in terms of all other considerations (including historical use and connotation) for all those variants which, are OR NOT YET considered to, comprise a particular species system.
Gordon Rowley’s complaint (penned in the early ‘70’s” should be recalled. This is against my failure to formally recognise names in my Handbook listing such as “ryderiana”, “revendetii”, “cuspidata” etc. that in my opinion were (ARE) horticultural in origin, and therefore did not belong in a formal botanical system. The problem now is that there is a greater mass of variants which are of field origin and the descriptive boundaries of these variants are far tighter and considerably more difficult to record and document in any naming system. Many of those old names that I dropped are being ignored as new ones are being invented.
Dr. Forster has also expressed the opinion that someone who knows the plants has to make the decisions. If I had known at the start what I now know, I might have done a lot better. My contention is that the community leaders need to address the problem as it is now one of general perception and understanding.
I wrote a short note for Haworthiad, to explain a picture of Breuer’s new species H. fusca (MBB7507), and said “… the fact is that it is from a small population just west of Albertinia en route to another of Hayashi’s (?) species H. esterhuyzeniae, and also to Breuer’s H. vincentii. As readers we are being conditioned to accept that there are many kinds of species such as biological species, morphological species, taxonomic species, good species, bad species etc. etc. so a Latin binomial could mean anything (and the word ‘tautology’ has been added to my vocabulary). Botany needs a sensible and practical handle to a squadron of populations from between Albertinia and Great Brak. I would gladly supply this if somehow I could be assured that the act was not seen to be the clown’s contribution to the circus.”
Without any assurance, but with the encouragement of Stirling Baker, I am going to try and produce an explanation.
Put very bluntly and without any apology to a group of people who definitely deserve better, my life experience is that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well. I have already written around the subject a number of times and do not want to repeat what is not necessarily true other than the contribution these thoughts have made to my personal psyche.
In this contribution I am discuss, illustrate and then propose that there are just two species, H. retusa and H. pygmaea in a complex where presently more than nine species and varietal names are being used. I do this in consideration of all the populations of Haworthia known to me in the winter rainfall biome. Thus I recognize the need to rationalize species like H. mirabilis (which will then absorb H. maraisii, H. magnifica and H. heidelbergensis, and H. retusa (which will absorb H. turgida. There is a major problem in that the populations indicate three species in the west, viz. H. mirabilis, H. retusa and H. mutica but these appear to fuse or morph to two in the east. My past treatment of species and varieties like maraisii, magnifica, acuminata, dekenahii, argenteo-maculosa will bear witness to the nature of the (my) problem.
Gist Charles Craib includes a provocative chapter on taxonomy in his marvelous book on The Grass Aloes of the South African Veld (Umdaus Press). Not so curiously, John and Sandra Burrows voice many shared sentiments in a similar grand book on the Ficus species of Southern Africa, Figs of Southern & South-Central Africa (Umdaus Press). These books coincide with my recent experience with Drosanthemum taxonomy to which I have been peripheral since I first tried to collect D. speciosum and its allies. An account of this experience is in manuscript, and like my experience with Oxalis already published, should really be read by people with taxonomic aspirations. Of course they will not, as neither will taxonomists do who seem to already hold very superior individualistic opinions about what species are. There is absolutely no doubt that there is a very commonly held view, that what constitutes a species is based on opinion only.
Perhaps just a single illustration will indicate the kinds of arguments one has to swallow, or refute, if ones perceptions are being threatened or distorted by the decisions which are made and the conclusions which hang on them. In an article which “officially” relegates yet again the single species of Poellnitzia to some other genus (thanks be at least this is within the ambit of the aloe group of genera – the genus Astroloba) some justification is made that … “the resemblance between the flowers of Poellnitzia and the bird-pollinated genus Microloma (Asclepiadacea) is particularly striking”. The author of this really curious statement took me to task when I pilloried it, because there is a publication which conclusively shows that sunbirds do gather the pollinia of at least one Microloma on their tongues. But there are several species of Microloma and I am quite sure that they are not all bird-pollinated and besides, that is not really the point. Here I want to illustrate this time the flowers of Astroloba(Poellnitzia) rubriflora, Microloma sagittata, and M. tenuifolia and that ask readers themselves to decide if the similarity is indeed “particularly striking”. (See figures 1, 2 & 3). The arguments are no different in rationale to those of Charles Craib who treats Chortolirion as a grass Aloe because it meets his particular approach. Whose opinion do we need to follow?
Charles Craib’s decision is based on the usefulness of a classification in respect of his needs. He has kindly sent me a recent manuscript discussing the merits of a new species of grass Aloe that he recognizes from his own knowledge and experience is a product of poor taxonomy. This brings me to the conclusion that taxonomy is not science as I have so often tried to say. Latin names are simply nouns and all that is necessary is that people using names know to what is being referred. A simple example is the fiasco of the name change from Aloe bainesii to Aloe barberae. After 40 years or more where all and sundry have happily used the name bainesii,somebody discovers that another name precedes it by the thickness of a sheet of paper. Aiding or compelling this change in my attitude, I have to admit that my own classification of Haworthia no longer really meets my needs and in the climate of taxonomy that I experience, I am not sure what should be done. Thus in this article I am going to examine what I consider to be a good example of the problems any taxonomist who ever gets a good enough sample will eventually face. The question will have to be asked – are species real?
The area under consideration This is the area east of the Goukou River which bypasses Riversdale, then to south of the high Langeberg Mountain range and reaching Great Brak in the east. The southern limit is virtually the N2 national highway between Riversdale and George, although my sampling drops south of the N2 approaching the Gouritz River from the west, and dropping south to the sea east of the river. The altitude is from sea level to about 250m. The rainfall is moderate and generally falls in winter, although it would be sensible to note that the area is really a westerly extension of the all-year rainfall area forested coastal strip between Port Elizabeth and George. The vegetation is coastal renosterveld although there are areas of Karoid Valley Bushveld especially along the river valleys. The geology is very complex and plays a critical role in plant species distribution and occurrence because the soils are very skeletal. It is often the case that Haworthia is associated with exposed rock where biomass potential and vegetation competition are low. There are five main geological groups represented in the greater Southern Cape area, and the study area is different in that it is dominated by a geologically differentiated formation of the Bokkeveld shales – different to the same formation which dominates west of Riversdale. A subtle factor is that this eastern formation produces, as a weathering product, a range of clays and minerals significantly different to those from the western formation. The eastern part of the study area is dominated by rocks of the more recent Uitenhage formation; while south of the N2 (outside the study area) the formation is the most recent Bredasdorp formation.
I asked Jan Vlok for his views on what may make the area ‘different’. He replied… ”Something I have noted repeatedly is how important river drainage areas are in determining vegetation units and pro parte also those of localised endemics. I have been wondering about the issue why river drainage basins would be so important; and currently think they reflect the complex interaction between several factors – geology, rainfall, landscape of area and in thus the history of the evolution of the area. Sessile organisms – especially plants with limited seed dispersal abilities are obviously strongly affected by these factors.
In your area of interest we have one very important role player – the Gouritz River. It tended to obliterate everything in its way cutting away habitat at a relative fast rate, thus creating many new micro-habitat sites for species to establish from the north and some coming in from the east-or-west. It’s a real “bastard” area with hybrids swarms of e.g. Tritonia and Watsonia species rather common. I guess they are “species in the making”, yes, they look very different from other populations but with no clear cut morphological characteristics (the latter painful from a taxonomic point of view, but great from a functional evolutionary-ecological point of view). Note here that less sessile organisms – especially with wind dispersed seed, e.g. orchids and for that matter asclepiads are less restricted from re-colonising or finding suitable habitat in adjacent river systems. So e.g. Duvalia immaculata has much more freedom to hop from one habitat, over the watershed into the next system, than any of your Haworthia species has. Those have to do the slow march, finding it difficult, but not impossible to cross the river or watershed, and much more slowly. This implies that in the Great Brak to Riversdale area, an east-west dissemination of Haworthia populations has been relatively slow – selecting for numerous micro-habitat specialists, but perhaps each population representing nothing more than parts of one large meta-population.
East of the mighty Gouritz we have the Brakke drainage system (Groot & Klein Brak Rivers). Much smaller & slow cutting into the landscapes, thus greater stability in micro-habitats – added is slightly higher rainfall and certainty of rainfall. Thus no surprise that some populations have evolved to more discrete entities, you call them Haworthia parksiana and H. kingiana.
West of the mighty Gouritz we have the Goukou drainage system. Clearly much more stable as in cutting away the landscape than the Gouritz, but still supplemented by some summer rain. In terms of vegetation mapping I found the area really difficult. Had to look at every corner as I could not predict what will occur where. The closely related sister to the west, the Duiwenhoks has been equally challenging, except that things started panning out better. I guess its related to less reliable summer rain, thus summer drought stress. It is also a less might river, with an odd geology – suddenly conglomerates reappear. So in the Duyvenhoks plant populations start taking on unique characters – as well as do the common and dominant species of the plant communities they occur in.”
Jan Vlok confirms my opinion that the only real factor in the biogeography apart from the complex geology at Great Brak, is the Gouritz Valley. It is mainly the geology which seems to impact on H. parksiana and H. kingiana. The river does not affect either H. minima, or H. turgida. There is some doubt about the interaction and variation of H. chloracantha and H. floribunda across the valley. The populations (splendens, fusca, esterhuizenii, vincentii and argenteo-maculosa are in my opinion a series of consecutive populations which can be regarded as a continuum unbroken by the river valley. There seem to be very few species or any other biogeographical evidence to suggest that Haworthia has been impacted on.
The Haworthia populations Those considered here refer only to the species of the subgenus Haworthia and exclude H. variegata, H. floribunda and H. chloracantha of that subgenus. One must assume that by this stage, most of the relevant populations are known although I cannot claim to have personally explored the full potential of the area. Altogether I know of 29 populations which need to be considered in terms of their probable taxonomic affinities. They are listed according to my interpretation of the present available nomenclature as follows:
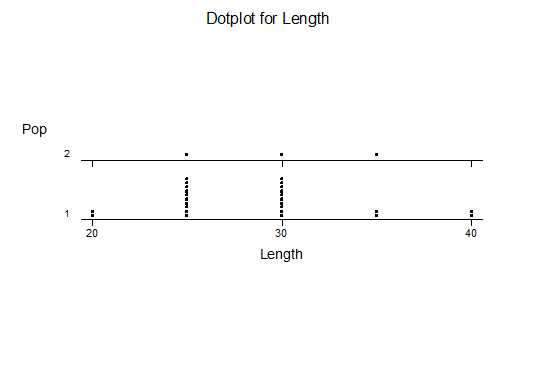
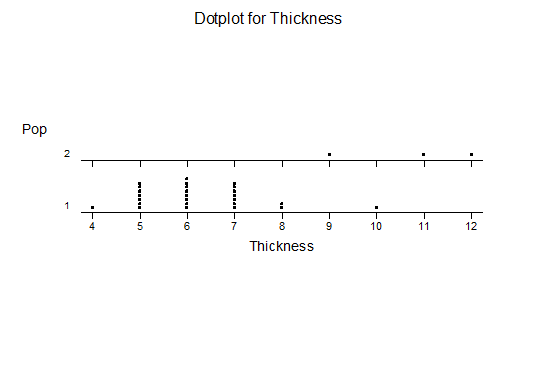
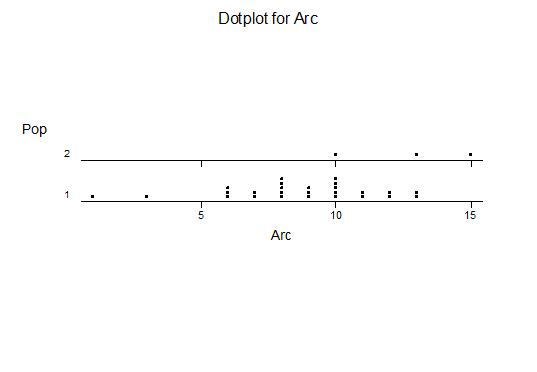
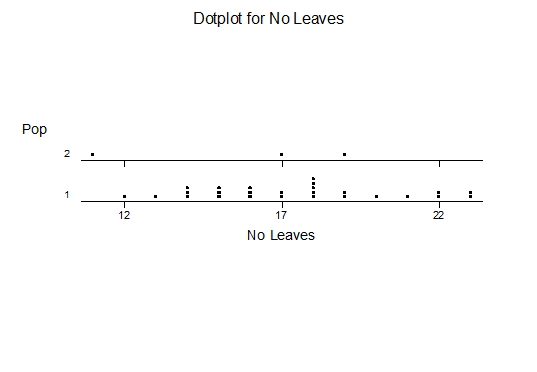
Haworthia turgida (Haworth 1819) Draaihoek, N Draaihoek, Valsch River, Wydersriver, Ouvloere, Die Eiland, Gouritz bridge, S Gouritz Bridge, Brandwacht.
Haworthia pygmaea (von Poellnitz 1929) Herbertsdale, Dumbie Dykes, Mosgas, SW Klipkop, Klipkop, Great Brak, N Great Brak, W Great Brak. (The vars crystallina and major (Pilbeam 1983) are not population based entities).
Haworthia magnifica var acuminata (Bayer 1976) West bank Gouritz at Melkhoutfontein (three separate populations), Vleesbaai.
Haworthia magnifica var dekenahii(Smith 1944) Draaihoek.
Haworthia magnifica var argenteo-maculosa (Smith 1945) Cooper Siding, E Cooper Siding, Humor
Haworthia magnifica var splendens (Venter & Hammer 1998) Soutpanskoppies, Snymanskraal.
Haworthia magnifica var esterhuizenii (Hayashi) Aasvoelberg.
Haworthia magnifica var vincentii (Breuer) Welgevonden.
Haworthiamagnifica var fusca (Breuer) Paulsfontein.
These populations are generally isolated from one another to some degree and it is evident that there are four taxonomic units represented by single populations.
A taxonomic species overview Of course the two words “taxonomic” and “species” used together is tautological as taxonomy is the classification of biological entities of which species is the prime unit. We have to consider what could now be species in terms of some definition, and in terms of all the other populations both inside and outside the study area. In this case there are a great many elements which fall in the same class within the same subgenus of Haworthia. I am not aware of anyone who has really addressed the issue of what comprises a species in the family Aloaceae and certainly not in the genus Haworthia. Breuer did essay a “concept” of Haworthia species, which was in fact a listing of species names for the genus and had nothing to do with questioning the nature of those species. This is in my observation a critical issue in most taxonomic work and it is the subject of many learned publications. I have already made my voice heard on the subject on many occasions and am not going to repeat any of my misgivings other than to say that I want to discuss the above populations in respect of a definition for species which I have fashioned for myself from experience and from a very wide literature.
This definition is that species are dynamic fractal systems of living organisms which are morphologically, genetically and behaviorally continuous in space and time. My attitude is that species have to be considered as phenomena in a creation and that these are spread in space and change with time. When assessing any plant as a species, one has to ask where it comes from, what its attendant companions are, what their variables are, and what their spatial relationship is. One also has to ask, what is all this in relation to geographical considerations including geology, topography, drainage systems, climate, vegetation and other plants of tangibly different families and genera? My experience with many taxonomic works and workers is that it is extremely difficult to approach the subject without extreme caution and also expect a considered response. While I have touted the view that classification is science, it most definitely is not managed in that way. It appears that there are two completely different needs that are met in classification. One is the purported one of botanical science, practiced mostly by herbarium based botanists or in present time, by institutionalized “gel-jocks” trying to lay bare the mysteries of DNA. Both are handicapped by the material available or sampled for their work, AND by their field expertise. The second need is that of the collector who simply needs a name by which to communicate with sources of plants, with other collectors and to have some measure of the difference in appearance of the plant from others he/she knows. Thus both needs will have to be met within the limits of the material known, seen, examined and experienced, and according to the competencies of the scientist, botanist, chemist, collector whoever happens to need an appellation for a plant. Taxonomic accounts and revisions are simply a product of the competence and experience of the writer limited also by the extent to which material, seen and sampled, matches or represents the reality. The term reality also has a special significance and one can rightly ask if this creation and its phenomena have any reality.
The evaluation I do this evaluation in the light of a synopsis of Haworthia which I submitted to the SA Biodiversity Institute for a flora of the Eastern Cape. That synopsis generates the question of how the species of the Western Cape should then be summarized. Quite fortuitously a parallel, although minor, problem seems to be resolved. In about 1976 I suggested that the flora of the Cape would never be understood until it was recognised that the true biome nature of the vegetation and the present-time Cape Flora was that of a Winter Rainfall biome stretching from Southern Namibia to the Eastern Cape. This now appears to be reflected in current botanical thinking where the Succulent Karoo is seen to be intimately linked to the Fynbos. The impact of this is that one really needs to re-think how species elements of the Southern Cape, are linked to those of the Little Karoo. Fortunately this does not really touch the elements I will discuss in this article although there are some peculiarities. For example, there is H. parksiana, H. kingiana, and Duvalia immaculata which are endemic to the Mossel bay area, while H. minima leapfrogs the Langeberg to occur in the Little Karoo.
What is more important is the present classification (of mine) which links most of the elements I am dealing with, to H. magnifica that is in the Riversdale/Heidelberg area. The other principle elements outside the study area bearing on the discussion are H. retusa and H. mutica.
Outside of vegetative physical appearances, there are only two really tangible pointers to relationships. One is population structure and co-existence (do the populations share close habitat or not), and the other is flowering time. Physical appearance is fairly helpful in that there are quite notable differences. There are plants which are generally light coloured and which are either adapted to steep rocky habitats as clumping cliff dwellers or are generally more solitary and adapted to level habitats. These are spring flowering. Then there is a set of populations in which the plants are very seldom in cliff habitats, are dark coloured and flower in late summer.
The first spring flowering group comprises (for the purpose of this article) H. retusa and H. turgida. There is no doubt in my mind that these two are essentially the same species as they never co-exist. The actual situation is very complicated and the recognition of H. mutica as the western representative of H. retusa and which can be said to co-exist with H. turgida, is indicative of that.
The second summer flowering group is even more complex and it is possible to suggest that H. mirabilis as presently constituted, should be enlarged to include H. maraisii, H. heidelbergensis and H. magnifica just west of Riversdale. In the context of biomes I would be rational to suggest that such treatment would need to consider also the incorporation of H. emelyae in view of its possible interaction with H. maraisii.
In respect of the study area it is thus evident that we need to look primarily at flowering time as all the populations are essentially in the light-coloured ambit rather than in the smaller dark green one of H. mirabilis. I do not imply hereby that this is inarguable, but it is evident that we need to understand that in the west we have a triumvirate of possible species with two distinct flowering times, whereas in the study area there seem to be two main elements which are both essentially spring flowering.
How do different people see the situation?
A. Evaluation and observation by Gerhard Marx Both dekenahii (Draaihoek) and fusca flower more or less together with retusa and emelyae (except major) in September-October. Both are in fruit in early November. In cultivation they also flower in March-April but I have never been able to visit them at that time in habitat to see whether they do the same. I would think so.
Splendens (Snymanskraal/Soutpanskoppies) flowers generally (late Dec. to early February). It is the one and only flowering spell for splendens.
Argenteo-maculosa and (its ‘variant’) vincentii also flower together with dekenahii. In Grahamstown argenteo-maculosa flowered for me throughout summer and even into winter. They seem to do the same for Steven Hammer in Vista. But I think in the wild they also have two flowering spells, one in spring and another in fall.
I seem to remember the flowering periods of acuminata are exactly the same as for dekenahii.
Pygmaea from Great Brak and Mossgas flower exactly together with dekenahii , but argenteo-maculosa (Humor and Cooper Siding) flower later. They are still developing flower buds at the moment.
As mentioned above, H. emelyaemajor also flowers later than the ‘picta’ types and ‘breueri’. Major is only starting to flower now, while all other emelyae are just about done releasing seeds.
Thus at present I see the relationships as follows:
H. splendens (Soutpanskoppies/Snymanskraal) is a good species and stands isolated in terms of its flowering time, chunky and glossy leaves and slower growth.
H. dekenahii can perhaps also stand as a seperate species and fusca is definintely a variant, distinguished from the dekenahii by its red-brown colour (meaning no doubt the “rusty”-like difference) greater tendency to offset and geographical separation.
H. argenteo-maculosa – (which I feel it is such an inappropriate name!) – is another separate thing, although it shares superficial characters with splendens on the one hand and acuminata and even turgida on the other. Apart from different flowering time to splendens, it is much ‘softer’ and grows twice as fast and is also smaller with more acuminate leaves that are spotted along the sides. H. vincentii can probably be seen as a variant of argenteo-maculosa as they are so obviously similar, except that vincentii has fewer and longer leaves that are less heavily flecked and it is even more ‘tender’ (softer) than argenteo-maculosa.
H. pygmaea is fine as it stands, with Great Brak/ Dumbie Dykes/ Rooiheuwel/ Mossgas/ Herbertsdale all sharing basically the same characters and flowering time.
Argenteo-maculosa (Cooper Siding and Humor) plants are different in having heavier flecked and chunkier leaves and to me it is a toss of a coin whether they are seen as a variant of dekenahii or pygmaea.
Similarly I think acuminata can just as easily become a variety of turgida or dekenahii rather than of magnifica.
Another option is to keep dekenahii and acuminata variations of magnifica and add fusca, argenteo-maculosa and vincentii also as varieties. But then it is a case of giving them all the same ‘status’ where I feel that fusca and dekenahii are ‘unidentical twins’, while argenteo-maculosa and vincentii are an almost-identical-yet-different duo.
The above decisions are really based upon subtle factors and can be decided by tossing a coin. What I feel is important, is that splendens should be separated as it does not fit in well with the dekenahii–pygmaea bunch or 100% with the magnifica from Riversdale. In terms of flowering time I can see the logic of including it with magnifica and atrofusca, but then dekenahii and acuminata should be removed.
The analysis closes with the words … ”Wait, let is that me sign off before I write myself into a corner.”
A1. Reaction comment by M B Bayer There are two important considerations. One is that Gerhard does not profess to be a botanist or to be too concerned with taxonomy. The other is that he is assessing the situation in terms of his specific life experience in relation to all the variables I mention earlier. He does not see to have considered the many turgida populations and their variation, the Vleesbaai population of acuminata, and another important single population of pygmaea at Herbertsdale, and neither does he relate these to a greatly compounded problem when all the relevant known Western Cape populations are added.
My views are that splendens is significantly different because the two known populations are associated with ochre deposits. Fusca is on a less iron/ochre rich site. Dekenahii is limited to very few plants (and there were ever many) on shale. Can these very small and highly localized elements seriously be regarded as species in terms of my definition? Argenteo-maculosa and vincentii are associated with table mountain sandstone. The argenteo-maculosa populations are also linked to sanstone while the acuminata populations are linked to old river terraces and the Vleesbaai population is linked to sandstone again. The Hebertsdale population is related to Uitenhage conglomerates and all the other pymaea’s to an array of substrates around Mossel Bay, Little and Great Brak.
Turgida occurs on steep rocky river banks (shale cliffs) and in two places can be said to coexist with pygmaea or dekenahii (Dumbie Dykes and Draaihoek). It is not outside the realm of possibility that the differences in vegetative appearances are responses to different habitat and substrate. It should also be noted that the turgida var pallidifolia was described from Draaihoek too. I saw such clones at the site when I visited the area in ca 1972, but on a recent visit no such light coloured clones were apparent. The population further down the Valsch River is also highly patterned as are those at Draaihoek. It should be noted that there used to exist another population to the north of Draaihoek which I would have equated with H. turgida var longibracteata from the Heideberg area or lower Goukou River at Riversdale. The population at Wydersriver is not known to me apart from photographs of G.G. Smith and also seems to fall outside of the ambit of turgida var suberecta as I classified all these eastern populatons of turgida. The one population which I recently examined was on the Wyders River northeast of Ouvloere (Albertinia district) where the plants definitely suggest affinity with vincentii. Examining my photographs, which are reproduced with this article, I thought there was a remarkable similarity between clones of fusca and argenteo-maculosa at Humorum or Humor.
Of course one can adopt a different species definition and even suggest that simple superficial resemblance of a minimal sample is adequate for an opinion forming process. It is certainly difficult to even attempt to examine how these populations became isolated or even to ponder exactly how isolated they really are in relation to pollination mechanisms and seed dispersal.
A2. Response by Gerhard Marx to the re-drafted manuscript I do not know whether to feel flattered or embarrassed that you took my loose comments upon the splendens/pygmaea/turgida group so seriously. Only a few minutes after sending the e-mail away, I wanted to add more questions and comments that partly contradicted the ones I sent.
It is really a complicated puzzle, particularly if one takes flowering time in consideration. When I look from dekenahii to that chunky atrofusca and to fusca and splendens, I do feel that they really neatly fit as varieties under the same species. But then splendens‘ flowering time removes it distinctly from the others and I do not know what to do with that fact.
I feel that then splendens might just as well be thrown in with H. emelyae because of the obvious similarity in appearance. The Moerasrivier/ Saffraanrivier area form of emelyae even has almost the same ‘pimpled’ upper leaves and some glossiness. However, I must say that I do like your proposal to have the whole bunch under H. pygmaea. From a collector and non-scientist view as my own, it feels good and definitely an improvement upon he dispensation as in Haworthia Revisited.
There have been brief moments when I even wondered why they can’t all be retusa. When I first saw H. fusca in the wild I was convinced it is a form of retusa. And after comparing it in cultivation with dekenahii, I became convinced that it is closer to dekenahii. Looking at your Herbertsdale pygmaea photos, I can clearly see shades of acuminata in them, as well as argenteo-maculosa (particularly Cooper Siding) and, of course, Mossgas pygmaea. This supports the idea of having them all as varieties of one ‘superspecies’. But I can find it easier to see esterhuizenii linked to pygmaea than retusa, so that is why I like your idea of having pygmaea as the umbrella species.
To my mind there remains the problem with splendens‘ flowering time that matches up with the mirabilis and atrofusca/magnifica group. And mutica, of course, is again linked to the pygmaea group in terms of flowering time. Particularly if you enlarged our concept of the mirabilis complex with new introductions like ‘shaggy dog’ and plants from ‘Die Kop’ etc.. Maybe splendens could even fit in there.
I really don’t know. That is why I often said that at times I have some sympathy other taxonomists that are inclined to call each differing population (or plant) a separate species ! It is just so much easier.”
A3. Response by M.B.Bayer To this, my response was…”it should not be found to be embarrassing – I think this is perhaps what influences taxonomy and why you do not want to be seen as a taxonomist. You are a taxonomist – we all are. Science and especially nomenclature have put a shroud around the whole process to suggest only those really in the know can make a good decision! What you have not done – and it is not a failing – is to consider the variants of turgida. Flowering time has to be considered with the same reservations as any other character and in the end I think it is an intuitive judgment which has to be made and not a product of scientific reduction and pedantry.
The inclusion of emelyae just adds more problems because there is a whiplash onto heidelbergensis and paradoxa via major and multifolia – and there are no clear answers to that either. The variation just folds back on itself. Hence my references to “fractal” – endless variation never repeating itself! My end point looks as though there is no solution except by consensus in which contributors also recognise intellectual and experiential boundaries?”
B. Comment by anonymous knowledgeable collector “About your ms. I understand that I am looking at it as an under-informed collector, and it makes me realize how completely I have failed to keep up with current writings on haworthias. But like our old friend has demonstrated time and again, ignorance is no obstacle to the collector in offering opinions.
I do agree with you that the populations between Albertinia and Great Brak need – to use your words – a sensible and practical handle. Your ms., however, has not persuaded me that it offers such a handle. From my point of view, to reduce these populations to two species obfuscates rather than clarifies existing population differences. To base your conclusions primarily on flowering time sets aside the differences in physical appearances which have so fascinated us and which have seemed so important in the designation of species. But that must be your objective? The ms., however, does not offer, at least to me, sufficient argumentation that flowering time can be used to solve, or resolve, the Albertinia-Great Brak population differences. And almost as an aside, I must note that, while you write that it appears that there are two completely different needs which are met in classification, you are willing in effect to set aside the collectors’ needs in order to reach a conclusion. The dual naming system which some of us now use because the existing formal system does not meet our needs is bad enough; no need to make it even more prominent by encouraging it.
B1. Response by M.B Bayer My response to this was…”May I incorporate your opinions as I have done with Gerhards?
I think the point which you may miss is that essentially the naming system is for botany, in fact all of biology, and there should be only one goal viz the recognition of living systems. There is absolutely no reason for collectors to insist that Latin binomials are required for their activities. Why on earth cannot collectors use H. pygmaea “splendens“, H. pygmaea “Herbertsdale”, H. pygmaea “vleesbaai”, turgida “pallidifolia“, turgida “sub-erecta” turgida “Wydersriver” etc. What more do they need? It seems fairly obvious to me that there is just a veneer of pretence that the latin names mean something more and which they aspire to? There is no way that anyone else is going to arrive at a classification of these populations (considering all of them) which is more meaningful. What we see in Haworthia is that physical appearances, as variously we see them, are not aiding identification or classification and flowering time does offer a view of a system which reflects over the entire series of populations. But in nortieri I can show that flowering time does NOT do so!
I cannot see how you say I am setting aside collectors needs. I am a collector and I want a system that is true and reflects what I know and experience. This is why I started fiddling with the names in the first place. Why have we so often considered that place names should be used? It is all very well and good for collectors with plants bearing names as they were obtained, to insist that those names have meaning and are immutable as opposed to those who have unnamed plants for which they need a designation.
I spent some time in the field with nortieri. This is a species which is grossly under-represented in collections in relation to field abundance. It must be one of the commonest and most widespread in terms of actual occurrence. We have the names globosiflora, devriesii, albispina, pehlemanniae and agnis as well as nortierimontana and gifbergensis and it is the most under-collected species of the genus. We do not have names for Arizona, Ouberg Pass, Opdie Berg, Moordenaarskaroo, Hex River Pass, Blinkberg, Breekranz, Nuweplaas, Dwasriver, and a few others which could be said to be different. How many more names are we going to generate based on existing approach? This does not worry the collector who is glad to have new names entering the system slowly enough for him to afford to add them.
What I am now saying is that there has to be an agreed system and that it is up to the collectors themselves to come forward with some sensible proposals. As a group they definitely do not have any idea of biological systems or even much grasp of nomenclature, the controls it puts in place and the problems that they also creates. I think I have said elsewhere that I am perceived to be the “trouble-maker” when in fact I seem to have spent my life pointing out that we are basing our hopes on method and system which offers no hope of stability or agreement.”
B2. Collectors reply The collector responded…”I fear that at the present we are far apart both in our individual thoughts and how we understand one another. What to do? There follows several unconnected comments or thoughts of mine on your ms. But first, I don’t mine if you use anything I write, even if it to knock it down, but I would be grateful if you would not use my comments as you have Gerhard’s, with reference to me. To me, what I write are “private” views which may not be supportable and which I may change tomorrow, if not sooner. I still cannot accept in my own mind the idea of using blooming time to formulate species. It is but one factor, and perhaps not even an important one, except where there are two species growing together.
Physical differences still seem the most important to me, per the difference between pygmaea in Great Brak and splendens at Albertinia. Are you not making physical differences important when you exclude certain populations like variegata from your ms? I thought that it was lack of physical differences between the plants at Horn Farm and those near Albertinia which led you to conclude that they were one species – which is going at the issue from a 180 degree direction.
The big “problem”, I guess, is really how to deal with the populations in the area which may or may not be connecting links, and which for one reason or another do not alone deserve species status. I just don’t have an answer. But I don’t think that “blooming time” solves the problem either. Rather, it seems simply to smother the issue.
You write in your ms that there are two completely different needs that are met in classification, yet you seem to give up the one, that of collectors, in your conclusions. If we are talking about taxonomy as the basis for communication, are you not giving up the importance of communication in the combinations that you propose? And yes, there may be two different needs, but why cannot the solution for this contain a system that meets both sets of needs?
I am not wedded to Latin binomial names. We have previously shared an idea of the usefulness of including locality data in a haworthia name. Can that be done without challenging the existing classification system? Every population could be a forma, and then everyone would know in a discussion what plants were being talked about. I recognize the objections and impossibilities, but I think that it is a great idea nonetheless. Meanwhile there are not two collectors in the world who are going to agree on a system for the use of collectors.”
B3. response by M.B. Bayer My response was…”Yes I do regret that our minds do not meet. This is the problem with taxonomy – it is simply not science as I once wrote. I have by no means switched to using flowering time in the way that you suggest. I am recognising that better solution to the problem is needed. Cooper Siding seems to flower with splendens and not with pygmaea nor with Vleesbaai acuminata and nor with Humor. It is simply axiomatic that it is a factor which influences gene flow, but this is disrupted by isolation and what botanist can pretend to understand “island” speciation? Variegata,floribunda and chloracantha present a parallel problem to this pygmaea complex and the question of physical differences to the pygmaea lot is barely relevant. Would any haworthia taxonomist suggest they be treated as one?
My experience with other genera suggests that any two people, who truly look at these things in the field and grow them in the way we do Haworthia, will end up in the madhouse if they are looking for agreement. The very point I have been trying to make is that if classification is to make sense and be useful as a communication medium that it is agreement we have to prioritize. If any one thinks they have valid and strong opinions then they should express them, and they should not lose sight of the fact that Latin names belong to the realm of formal biology. What is the point in dissent which is not balanced by counter argument? In a recent journal an author has at least ten new names to occupy the place of my aristata. This is as if the use of the name aristata itself does not have the potential to generate waves according to the intent of the reader. I wrote at the very outset that my intent was a system of names within the domain of the South African flora as a whole. How will we ever get agreement if we do not even see that need? There is a huge trend to turn everything upside down using DNA technology and now I just have informed comment that standard morphological methods work better!
One should not actually express an opinion without recognising the fact that there is a physical record in the form of the herbarium collections (worldwide) which even take precedence over the plants in the field. Unless ones mind covers all those records how can valid can an opinion be? This is the reason why no two collectors can possibly agree – they are confined to the sample they hold in their minds.
You close with what I think is a fine solution. Refer to the populations by place name within a formal structure and it is that formal structure which I am trying to find.”
We did try to continue this exchange of ideas. My feeling was that it would be endless as the collector is not in fact in a position to evaluate or question any comment in respect of biogeography. Understandably he has to lean on the historic method of dependence on the evidence he has obtained from the material he has seen. Another consideration is that my decision is being influenced by what I think about the western populations where I suggest that H. mirabilis absorbs H. maraisiii, H. magnifica and H. heidelbergsensis. In response to his comments I redrafted the manuscript to explain this.
C. Comment by Steven Hammer “If you say that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well, this sounds contradictory, though one could say that farces and comedies are effective. I suppose the problem is that one is unable to take taxonomy seriously in the way its “strict” pretensions would demand. But if it works in its fashion then all one needs to do is to analyse that fashion and say it’s OK for that purpose.”
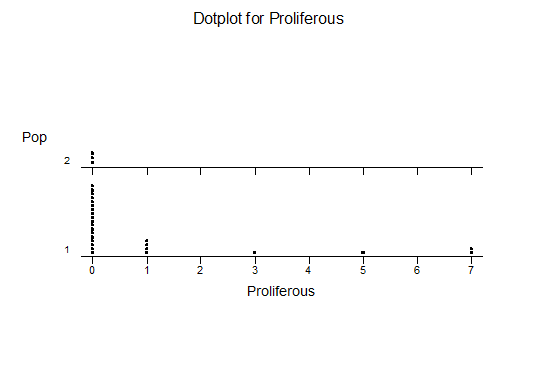
His opinions are that esterhuizenii is far greener than splendens and lacks the capacity to “blush”. Of Dumbie Dykes pygmaea he writes…”it is different – big pale, flabby readily proliferous (lots of turgida traits). Thus it has echoes of vincentii too. But you can see all of those traits in Great Brak pygmaea too.”
In respect of habitat and substrate…”I think it is likely that the difference has its impact – consider the way both western splendens types have changed after a time in cultivation. Soutpans plants look as it they’d been digesting powdered white porcelain! They did not have this look for the first three years. Much more slowly, some of the Snymanskraal plants have acquired it too, while at first they resembled many magnificas, especially those with atrofuscan aspirations.” He adds that it is possible to imagine the Mossel Bay pygmaea transported to Soutpanskoppies and getting bigger and fatter.
In respect of formalities of nomenclature attaching a reality to the entire spectrum of names which they simply do not have, Steven writes…”I love this last sentence. I can see that each of the other elements can be seen as variant of, or a link to, of pygmaea. There are so many cross-links…”
He adds…
”Where orthodox taxonomy gets in trouble is in its hope for absolutes – we are told, for example, that splendens never has spots on its leaf backs. This is not true when one sees a large enough sample. It is true that most esterhuizenii’s have many more spots, much more often, but it is also true that one of the original esters – a Parisi plant – would have passed anyone’s inspection as a western (Soutpans) splendens. If that isn’t a proof of a connection I don’t know what is! It is also true that Parisi’s plants, and their descendents, can be distinguished from the Venter-Marx plants bred from material collected on the same hill – they have more ‘silver’ and are more delicate. That lineages can be distinguished on such a minute (if trivial) level tells us something. Haworthias are very malleable in a pot. If they are that malleable in a pot they must be malleable/shiftable in habitat too.
That brings up continuity. The gene flow may once have been a continuous flow in one direction, but the river’s dried up. So now we have islands in a dried-up stream. Collectors will always want to call splendens a species – it concentrates all the glamour of Haworthia in one shiny package. I don’t know what rank it “deserves” but I suspect it is the least trivial, or the most distant, of pygmaea’s ramifications. If this is pygmaea then it’s the most deeply transformed of the lot. Or it’s got some emelyae links, which would fit, by extension, the maraisii/emelyae link. I do think that co-occurence logic is the strongest logic possible. One can also look at Soutpans and say, hmm, no trace of turgida here!”
C. Response by M.B Bayer I do not consider that if the system works in the fashion that it does, and that it is OK for the purpose, that this helps. It is only working because too little is expected of it. The scientist/botanist believes that the names are providing critical information about species but which they cannot define, and the collector is content with a handle to attach to his plants or communicate about them. My contention is that taxonomy is not providing proper scientific content.
Illustration
1. 7508. Draaihoek. Northwest of Albertinia and the type locality (the only locality) for var. argenteo-maculosa. The substrate is Bokkeveld shale and the vegetation renosterveld. The locality has always been very restricted, with few plants and very unsubstantial. Although there might have been more suitable habitats available in the past, there is very little remaining.
2. Snymanskraal. splendens. The variant was discovered again almost solely due to the persistence of R.W.Kent althouth the next locality may be where Herre originally discovered it. This habitat is quite complex. It is at the convergence of recent sands, ferricrete/silcrete and shale. The ferricrete/silcrete is in a relatively flat-lying formation rather unlike the raised inselbergs that it forms in the west. The vegetation is largely Renosterveld, but with so much sand in the vicinity, there is also Coastal Fynbos.
3. 6751 Soutpan. splendens. Very similar to the preceding habitat and the plants are also very localized in the exposed ferricrete.
4. 7507. Paulsfontein. fusca. This habitat is also on the geological convergence described but here there seems to be high level river gravel which predominates.
5. GM Aasvoelberg. esterhuizenii. Parisi and Dunn seem to have been the discoverers of this population on the low northern slopes of Sandstone hills to the east of Albertinia.
6. 7102 Welgevonden. vincentii. A few kilometers further east, the locality is also Table Mountain sandstone but a different geological series to that at Aasvoelberg. This and the previous four records are in a very simple geographic line and there really is nothing to suggest any biogeographically differences in these habitats other than the detail of substrate. In the general way that Haworthia populations display some degree of observable continuity from one population to the next, so do the plants in these populations. The actual differences may seem quite large but there are parallels at many places for this ecotypification in the genus.
7. JDV86/75 Cooper Siding. argenteo-maculosa. The habitat is again ferricrete/silcrete on an interface with sandstone. Plants west of the siding are actually in Sandstone. It appears that the plants can be found eastwards along the sandstone ridge to near Humor about 10km eastwards. I do not think anyone has really explored the differences in the plants along this ridge. H. floribunda also occurs at the same site and I have seen a hybrid.
8. JDV91/39 Humor. argenteo- maculosa. The plants are on the low slopes of another sandstone ridge which parallels that at Cooper Siding and apparently a different series. I am not aware that anyone has given these plants an individual name, but would be justifiable in terms of all the other names in the series. The plants are quite strongly marked.
9. JDV87/97 Melkhoutfontein, Gouritz Bridge. acuminata. The original locality was destroyed by road-building and these plants survived in cultivation. Populations are also known to the south and north in the bank of Bokkeveld shale which borders the river on the west bank. The plants are odd in that the leaf ends are very acuminate.
10 MBB6747. Vleesbaai. acuminata. The discoverer of this population may be V. DeVries. The substrate is calcareous limestone. It is a similar formation to that at Stilbaai where H. variegata occurs. The plants tend to be quite robust with a curious grey sheen in cultivation.
11. JDV9/135 Herbertsdale. pygmaea. Enst van Jaarsveld found these plants on a steep Uniondale conglomerate slope with H. chloracantha also present. The plants tend to be a large and flabby and not particularly attractive to grow, whereas in habitat they retain their tight soil-level leaf arrangement.
12. EA1200, 6813. Mossgas. pygmaea. This population was found by Paul Schoeman in sandstone just west of Mossel bay. It is important to note that this sandstone is the same formation that runs from south and east of Albertinia in a narrow band sloping down to near sea level at Mossel Bay. Populations 2, 3, 4, 5, 6, 7, 8, and 12 are thus along this band and its contact with younger tertiary formations.
13. ADH275. Northwest of Mosselbay. pygmaea. The geology is exposed ferricrete with river gravels. Adam Harrower found a plant with seed and grew the seedlings here illustrated. Notable is the appearance of plants with the crystalline papillae known in plants from Dumbie Dykes and from Great Brak.
14. EA1218. Klipkop, North Mossel Bay. pygmaea. The substrate is old river terrace. Etwin Aslander’s locality is now a housing estate. The plants are not much different from those in the previous record.
15. EAsn. Dumbie Dykes. pygmaea. The geology becomes very complex from here eastwards as there is igneous extrusion.
16. Great Brak. pygmaea. Various collectors have plants from here. During the early years I observed pygmaea in four different populations in and near Great Brak. They are still present practically within the town, and a short distance to the northwest where they are in gravels. What struck me forcibly at the time was the similarity to H. mutica and in one close comparison of two plants, I could only separate them on the colour of the bracts.
17. MBB7512. Draaihoek. turgida. I will not illustrate more than four populations. Essentially it is significant that we have a situation which parallels that in the Eastern Cape where H. cooperi is is general the plains-dweller and H. cymbiformis the cliff-dweller (in the west there is a morphing and fusion so that cooperi var gordoniana is the plains-dweller and cooperi var picturata and H. transiens the cliff-dweller. In the southern Cape, the separation has been H. turgida the cliff-dweller and H. retusa the plains-dweller. The relationship is less consistent than is the case in the eastern contingent and the very complex continuity of H. mirabilis with elements such as H. heidelbergensis, H. maraisii and H. magnifica (which can be rationally treated as one) suggests that H. retusa should absorb H. turgida. In practical terms H. turgida is the predominant element, and the name retusa is taken because of the requirements of nomenclatural rules. It must be noted that the complex of populations that I have discussed in Haworthia Update Vol. 2 as H. mutica var. nigra represent a part of the interaction of H. turgida and H. retusa and thus in this light would all be H. retusa.
H.. turgida, supporting the contention that it is the cliff-dweller, occurs in the Valsch, Wyders and Gouritz river valleys on very steep sites. It is proliferous, so the plants are often in large shallow rooted clumps. The leaves do not develop the very flat and truncated end-areas as do the plains-dwellers. As far as I am aware it only occurs at two localities east of the Gouritz River. One at Brandwacht north of Mossel Bay and I believe at one east of Little Brak. Only at Draaihoek is it in very close proximity to argenteo-maculosa. It is at this locality where turgida var. pallidifolia was collected. It is both curious and co-incidental that I did observe one or two very pallid plants at the time of my first visit to the site (now apparently no longer present) and formed an impression that the separationof argenteo-maculosa and turgida was actually a question of ecotypic adaptation. A population I did sample was present about 1km north of Daaihoek in which the plants were more comparable with turgida var. longibracteata and these did influence my impression. That population seems to have disappeared as the area is very disturbed (largely due to the dense hyrax population) and I could not find it on a recent visit.
18. 7105 Ouvloere. turgida. The plants here have the same rather obtuse leaves which prompted me to recognize the variety sub-erecta, but I was also struck by the change in the mottling of the leaves and the similarity to this in esterhuizenii and especially vincentii.
19 ADH397 Gouritz Bridge. turgida. This is the well-known form of turgida which once passed under the cultivar name “rodinii” for which Die Hell, up-river in the Swartberg Mountains,was incorrectly given as the origin.
20 EvJ16716 Schoemanshoek, S. Gouritz Bridge. turgida. This is very similar to the former and again from a cliff habitat. There is of course a similarity to acuminata which is only a few kilometers away on the sloping rocky banks of the river. That does vary from the pygmaea complex where I would include it now, as it also tends to be proliferous. I consider that this emphasizes the tight relationship of H. pygmaea and H. retusa (turgida) rather than supporting any reason to fragment them taxonomically as we have done in the past.
Conclusion If I now am required to produce a synopsis which makes broader botanical sense, rather than meeting just the needs of collectors, I would suggest that it is mutually agreed that only two species be recognised. These are H. retusa represented by the populations cited here by the name H. turgida and with the rider that in fact H. turgida should be incorporated under a wider acceptance and recognition of H. retusa; and that ALL the other relevant populations in the area under consideration should be treated as the single species, H. pygmaea. For the collector I would seriously suggest that the formal ranks of subspecies or variety simply be ignored. The species name is used and in inverted commas any other name that is useful to popular discussion and communication. The formalities of nomenclature attach a reality to the entire spectrum of names which they simply do not have. Names are being used to register collection, acquisition, possession and personal achievement, and are recording very little botanical information and understanding of the extraordinary nature of living things.
A prominent taxonomist was very dismissive of my opinions and cursorily explained that…” Yes, species are real, and defined well by their ability to cross freely and produce offspring which again crosses freely. This has been studied and demonstrated since 1750 or so (you may remember that I talked about this in one of our discussions on the subject), but it is naturally not easy to dive into so much work including following up several generations etc. And since related individuals are similar looking, the reciprocal conclusions that similar objects are related has unfortunately been used as being true, which it is not. And yet, species are the only natural unit in the whole of taxonomy, which to determine is the crucial point. Perhaps this sounds rather like the famous Dicta of Bessey, but after having dealt extensively over 38 years by now with species boundaries in teaching, reading, theory and practice, in different vegetation zones and many different groups, I come back to old definition first given by Ray in 1682: “Group of plants derived from common seeds, reproducing their typical features by sowing” and used by most taxonomists since.”
My contention is that this definition is grossly inadequate as it under-estimates the degree of variability that can occur within a species, not only inherently, but especially in relation to spatial distribution and adaptation. ♦
Barry Phipps, in an article reprinted in Haworthiad 20:61, writes that “the term species is a concept”. Donald Levins in The Origin, Expansion and Demise of Plant Species devotes a chapter to “The premise and species concepts”. There is no dearth of literature and the entire subject is indeed, as Levins suggests, a subject of “heated debate”. Levins also quotes from the literature, “… the idea of good species … an artifact of the procedures of taxonomy”, and “… our system of names appears to achieve a reality which it does not possess”. It is comforting for me to read his premise …”that the species is a dynamic entity that undergoes alterations in its gene pool, variation pattern and geographical distribution”, and his advice…”thus it is best to take a pluralistic approach to species’ passages in time, combining genetic and ecological perspectives”.
I myself am not sure if these and many similar statements really mean all that much. The range of living things is surely a reality of the human capacity to observe and rationalize to the limits of their experience and competence. They are phenomena spread in space and changing with time along with the rest of creation. We give them names in order to catalogue, describe and communicate about them. Those people, who make a point of collecting and studying study living objects, give those names and theorise and generalize about their relationships. We do not conceptualise these living things. They obviously are “kinds” and do surely have an objective reality. We may perhaps conceptualise a taxonomic system in which they are studied and classified according to some model or other. This has been an ongoing process since before Linnaeus and it is unfortunate that many writers seem to have become taxonomists with only the International Code of Nomenclature as reference. The result is a plethora of meaningless subjective names or names which can satisfy only the writer and readers who know as much or less. If we do take a pluralistic approach, recognise all the pitfalls, and base our observations on extensive observation, then surely the classification product will be satisfying to ourselves and have some value for others. This is what I have tried to achieve.
In seeking some kind of closure to my own long history as a pseudo-taxonomist and botanist writing about Haworthia, I am going to write about my observations on a number of populations which I attribute to the single species Haworthia nortieri. The newest trend seems to be to term these discrete, isolated, scattered populations as “metapopulations” and theorise about the interchange of genetic material among them and the effect this has on real differences; ultimately reflecting on evolutionary passage. Whatever is attempted or intended, an accurate account of one’s personal experience must surely be of value.
H. nortieri was first noted by Dr. Nortier of Clanwilliam and then collected at the old Doorn River Bridge 45km northwest of Clanwilliam in 1937 and described by G.G. Smith in 1946. Smith also collected a similar plant on the Pakhuis Pass also in 1937, and described this as the var. montana only in 1946. In 1948 he again collected a similar element on the Gifberg Mountain near VanRhynsdorp, which he described in 1950 as var. gifbergensis. In his collecting record, Smith records, also in 1948, the collection of a plant from Doornbosch northeast of Clanwilliam, and described that as H. globosiflora. In the description the collection is attributed to J. Thudichum.
History Thus in 1969 when I started the physical process of arranging the Compton Herbarium records, these four herbarium specimens virtually constituted the total record of the species. My views of the species and its taxonomic record in my 1982 synopsis and 1999 revision were as follows:-
“25. Haworthia nortieri Smith, JS.Afr.Bot. 12:13(1946). Bayer :141(1976). Bayer :49(1982). Scott :88(1985). Type: Cape, Vanrhynsdorp, Smith 1676a (NBG): H. nortieri var. montana idem. 16:6(1950). Type: Cape, Clanwilliam, Smith 1678 (NBG): H. nortieri var. gifbergensis idem. 16:7(1950). Type: Cape, Vanrhynsdorp, Smith 7199 (NBG).
nortieri: for Dr. Nortier.
Rosette stemless, proliferous, 3-5cm φ. Leaves 25-45, soft sub-erect, ovate-lanceolate to obovate, pale to purplish green, with translucent spots on the leaves, small spines on margins and keel. Inflorescence slender, to 30cm. Flowers greyish-white, yellowish in tube.
1982- “H. nortieri occurs in the area between Clanwilliam and Vanrhynsdorp, extending inland to the foot of VanRhyns Pass and on top of the plateau of the northern Cedarberg mountains. The var. globosiflora occurs in the dryer Botterkloof area and is distinguished by the flower having a globose tube while the leaves are shorter and broader. However, the flowers are not always so inflated and plants in the VanRhyns Pass area have normal flowers while being vegetatively similar to the var. globosiflora. This is the basis for rejecting species status for the last-named. H. nortieri occupies the mid‑western geographic locale for the genus. The distribution southwards is not known and there is a big gap between this species at Pakhuis Pass (near Clanwilliam) and a form of H. archeri var. dimorpha in the Elandskloof area southeast of Citrusdal. H. arachnoidea occurs to the north of VanRhynsdorp and also to the north and far west of VanRhyns Pass. It does not, however, seem to appear in the Botterkloof area. H. nortieri is distinguished largely by the opaque leaf surfaces with abrupt, ovoid, pellucid spots. The flowers, and particularly the buds, are greyish in colour but the colour inside the tube is variable.”
1999- “The range of H. nortieri has been extended considerably. The reference to H. archeri var. dimorpha at Elandskloof is quite erroneous and simply arises from the difficulty in relating that collection to the nearest known species. On the other hand, the Elandskloof plants are only known to me from herbarium record and from two living plants collected by Drs Muller-Doblies. These did not seem to unequivocally be H. nortieri. P.V. Bruyns has collected H. nortieri from as far north as the Groenriver, to south as far as Krommeriver in the Cedarberg. The late Harry Hall also collected it in the northeastern Knersvlakte. The most southerly collection is from near Opdieberg, north of Ceres. These plants also resemble the var. globosiflora but the flowers are not globose. At both extremes the plants tend to resemble H. globosiflora vegetatively. The var. nortieri is probably best considered as the more slender-leaved and less robust sandstone ecotype of the species. The plants at Komkans tend to have globose florets, and this is also true of plants at Groenriver where the florets are short and squat. The decision to include H. pehlemanniae within this species is also on account of the flower which is identical to that of var. globosiflora. This is not only in shape, but in colour too. Although the flowers may be the usual white with greenish veins, brownish-green flowers have been observed in both elements. The reference in the original description to quadrantly as opposed to spirally arranged flowers defies plant growth principles. The distribution of the two elements is complementary and gives the species as a whole an extraordinary cosmopolitan character. The colour in the flowers of the typical variety can be remarkable and as if yellow paint had been daubed at the throat of the florets. Habitat ranges from the moist south slopes of the Cedarberg to the dry wastes of Namaqualand and the so-called Moordenaarskaroo.
a. var. nortieri
The typical variety occurs over a very wide area in the Table Mountain sandstones from south-east of Citrusdal to south-west of Nieuwoudtville. In addition it extends out into the Namaqualand lowlands, into very arid conditions. It is thus a very variable taxon in its own right.
Distribution
3017(Hondeklipbaai): Groenriver (-DD), Bruyns 6728 (NBG). 3118(VanRhynsdorp): Komkans (-AA), Bruyns 6146 (NBG); Klipdrif (-BB), Hall 3390 (NBG); Gifberg (-DA), Smith 7199 (BOL,NBG), Thomas in NBG626/69; Steenkampskop (-DB), Bruyns 6167 (NBG); Kobe Pass (-DB), Bruyns 6170 (NBG); Die Kom (-DC), Bayer in KG329/72 (NBG); Doornriver (-DC), Smith 1676 (NBG), Leighton (BOL); W. Doornriver (-DC), Bayer 3637 (NBG); SE. Klawer (-DC), Leipoldt 4146 (BOL), Herre in STE6695 (BOL); E. Doornriver (-DD), Smith 1676a (BOL), Smith 6212 (BOL,NBG), Esterhuysen 6008 (BOL). 3119(Calvinia): Uitkomst (-AC), Barker 10753 (NBG); Vanrhyns Pass (-AC), Smith 6211 (NBG), Hall in NBG656/60 (NBG), Ross-Frames in NBG1200/26 (BOL). 3218(Clanwilliam): S. Clanwilliam (-BD), Van Jaarsveld 8153 (NBG). 3219(Wuppertal): Pakhuis Pass (-AA), Smith 1678 (NBG), ex hort Whitehill NBG68397; Waboomsriver (-AC), Henderson 2212 (NBG); Diamond Drift (-AC), Leipoldt 3107 (BOL); N. Dwarsrivier (-AC), Bruyns in Bayer 6505 (NBG); E. Dwarsriver (-AD), Bruyns in Bayer 6506 (NBG); Heksberge (-CA), Smith 6116 (NBG), Muller-Doblies 79/015 (NBG), E. Elandskloof (-CA), Esterhuysen 3987 (BOL); Sandfontein (-CB), Esterhuysen 27199a (BOL); Cedarberg (-CB), Wagener in NBG11/43 (NBG); Nuwerus (-CB), Bruyns (NBG). 3319(Worcester): NW. Ceres (-AB), Aslander 645 (NBG).
b. var. globosiflora (Smith) Bayer :119(1976). Bayer 49(1982). H. globosiflora Smith, JS.Afr.Bot. 16:11(1950). Scott :87(1985). Type: CAPE‑3119(Calvinia): Doornbosch, N. Doorn River Bridge (‑CD), Smith 7198 (NBG).
globosiflora: rounded flowers.
This variety is not known from only the Doornbosch area south of Botterkloof, which is also relatively unexplored. It has also been collected from as far east as the Ouberg Pass, southwest of Sutherland. The illustration in Scott (:88, 1985) is not of this variety at all and is probably of H. decipiens. Like the var. pehlemanniae, the florets are remarkably globose with a significantly shortened tube, and the two varieties cannot be separated on this score. The flowers of both may be uniformly greyish-green or brownish. Vegetatively, the leaves in the var. globosiflora are distinctively dotted with translucent spots which are absent in var. pehlemanniae.
Distribution
3119(Calvinia): Doornbosch, (‑CD), Smith 7198 (NBG); 50km N. Clanwilliam (-CD), Dyer 3750 (PRE); Botterkloof (-CD), Hall in NBG68414, Villet (BOL); Boontjiesrivier, Kansekraal (-CD), Leipoldt 4119 (BOL). 3220(Sutherland): Ouberg Pass (-AC), Venter 88/40 (NBG).
c. var pehlemanniae (Scott) Bayer comb.nov. H. pehlemanniae Scott, Cact.Succ.J.(U.S.) 54:70(1982). Scott :79(1085). Type: CAPE-3320(Laingsburg): 5km W. of Laingsburg (-BB), Scott 7450 (PRE).
pehlemanniae: for Inge Pehlemann.
Since first collected, this variety has been found at several other localities in the close vicinity of Laingsburg, but also further north in the Moordenaarskaroo and north of Matjesfontein. It differs from the species in the absence of the translucent spots on the leaf, and the vegetative similarity to H. arachnoidea in the same area is deceptive. However, the leaves have a slightly more greyish character than in H. arachnoidea. It does appear to favour shales in relatively exposed situations as opposed to H. arachnoidea which generally prefers cooler south slopes. The two taxa grow in very close association.
Distribution
3221(Merweville): Klipfontein, N. Laingsburg (-CC), Aslander 801 (NBG). 3320(Montagu): 5km SW. Laingsburg (-BB), Scott 7450 (PRE), Bayer 3906 (NBG); N. Laingsburg (-BB), Venter (NBG).
During the last few years several names have been published which in my opinion belong in the context of the above The first of these is H. agnis L. Battista, Alsterworthia International 2(2):8-9, the second H. devriesiiI. Breuer, Avonia21:3 (2003) and the third H. albispina M. Hayashi, Haworthia Study 9:2 (2002). These are all significant records and I do not question the fact that the populations are distinctive and important variants. I simply add that in terms of an objective view of an observer who wishes to understand these plants in the total context of the flora of the region and at another level in the context of the genus Haworthia and its three subgenera, they belong in the one species H. nortieri. For the collector I would suggest that someone make the formal taxonomic statement that these are varieties. The types are deposited who actually knows where, and being outside of the classic regional herbaria they have also not been assessed by the authors of the names, in terms of the total herbarium record. This disregard for rational herbarium practise does, and will continue to, make it difficult to technically formalise a classification.
In referring to the regional flora, I want to explain my viewpoint that botanical classification belongs in the realm of true biology and must be meaningful in terms of all life forms and the conditions which gave rise to their origins and which sustain them. Thus my experience was that the botanical community were defining a Cape Flora based only on the machia-fynbos vegetation peculiar to the predominent sandstone geology of the area. My view was that this was a misinterpretation and that whatever the origins of the fynbos was, there was in present time a Winter Rainfall Biome which included the karoid vegetation of the shales, tillites and igneous rock within and adjacent to the so-called Cape. Species systems, like H. nortieri, would only be understood is this massive impact of geology, skeletal soils and winter rainfall was properly acknowledged.
In a manuscript concerning H. pygmaea and a broader species concept for the genus, I have tried to explain that the reason that I have been drawn into the realm of formal taxonomy, is precisely because there are no reliable and infallible criteria or characters by which the species and the variants van be positively described and identified. The observer is totally dependent on imagery and by the experience garnered in the field and in cultivation. The classification thus can only be assessed in terms of comparable experience and motive.
I have avoided trying to rationalise the morphological differences between H. arachnoidea and H. nortieri because I have found them myself to be a bit obscure. Generally the colour of the leaves in arachnoidea is a darker uniform green as opposed to a pale and often maculate condition in nortieri. Where nortieri does seem to differ is that the mid- to end section of the upper leaf surface tends to be convex. This convexity of the upper leaf surface is reminiscent of the similar characteristic of the species of the southwestern Cape where a retuse truncated end-area often occurs, and may thus account for the truncation in the leaves that characterises some nortieri populations. I have observed this truncation as a response to growing conditions as well, notably in the specimen from the Blinkberg which Ernst van Jaarsveld grew in cultivation as compared with plants observed by myself in the field (see pictures). Similarly plants from Opdieberg collected and grown by Etwin Aslander seemed to have very truncated leaves under his growing conditions. Plants in the field at Arizona north of VanRhynsdorp were very retracted into the soil and the leaves also correspondingly truncated. Alternatively there are populations in which the leaves are attenuate, longer and pointed, bringing me back to an observation I would like to make that such morphological characters and differences are too often used to justify species differences when the variation is in fact an expression of the nature of the species. There are several populations in the greater Touws River area which are doubtful and a good deal of tough fieldwork will be necessary to get a better understanding of these disparate populations which could include both H. maculata as it occurs on and adjacent to the Audensberg at Worcester, as well as H. marumiana var. dimorpha east of Touws River
The variant agnis has been collected twice by Etwin Aslander both east of Nuwerus and at Middelpos near Bitterfontein. It has also been collected by Philip Desmet at Nuwerus. Both collectors observed the presence of H. arachnoidea at Nuwerus and Desmet submitted specimens of a putative hybrid. These plants are indeed small and Dr Ben Zonneveld (private communication) who has researched the mass of total nuclear DNA in many Haworthia specimens maintains that his figures suggest that it is a discrete species. I do not think that his argumentation or his methodology are substantial or convincing and that these kinds of peripheral data sets should be use in taxonomic decision making. In my opinion they could be used to describe variation within a species system more usefully than to support a rickety overburdened classification tree (cladogram). I should add that the Bruyns collection from Komkans, which is west of Nuwerus, is also of small plants and is actually a replication of a collection made by Rolf Rawe in 1969 also at Komkans. That collection was never formally deposited as a herbarium record but I remember it distinctly as there were many plants in the San Merino Nursery at Somerset West closely following the closure of the notable Bernard Carp nursery at Hout Bay.
The variant devriesii is indeed odd as it is from very far east for the species north of Prince Albert. I will show by the illustrations how it relates to H. nortieri by similar ecotypes from the karoid landscapes peripheral to the machia-fynbos of the sandstones where the species is better known. Like albispina, which is from Koup Station, it should simply be regarded as a variant of the existing var. pehlemanniae, although I do not believe that formalizing varieties really is very helpful in the botanical sense. The variation is, in my opinion, largely driven by the nature of skeletal substrate and is not conformable with any classification system that can account for the totality of variation in the species. Both the names as representing formal taxa, will be confounded by complete and true description of local variation within the actual populations the types were sampled from, and further so by samples from populations known to the authors of the names. Thus we have populations in among those cited in my revision as well as new records, particularly from northeast of Nieuwoudtville, and from N VanRhynsdorp, which need to be offset against these “new species”. There are also several new records of my own (Sneeuberg and Katbakkies) and by Ernst van Jaarsveld (Blinkberg, Cedarberg) and by Adam Harrower (Breekkranz, Cedarberg) that add to and extend the already wide variation of the sandstone components of the species. Bruyns (private communication) also reports the species from the Elandsberg in the Tanqua Karoo. Adding new names requires that one cast ones mind back to the situation prior to the revisions by either Col. Scott or myself and question whether these new names require the re-instatement of so many other names put into synonymy in those revisions. At the same time some note must be taken of the degree of exploration and how predictive any new name is likely to be.
Regarding flower characteristics and flowering time, my observation in other genera is that it often appears that variation between species samples is that the observed variation in selected characters, may within a species system exceed that observed between different species systems. I stress the word ‘observed’ because any opinion or judgement can only be made in terms of the sample and information available and utilized by the observer. Thus I concluded that the var. globosiflora belongs in the nortieri system by virtue of populations apparently in the same geographic and floristic range in which the flowers are neither globose nor normal for nortieri. The variant agnis for example has smaller globose flowers, the flowers of pehlemanniae cannot be distinguished from those of globosiflora, while in devriesii the curious conclusion is drawn that the less globose flower suggests an affinity with H. decipiens var cyanea. G.G. Smith was struck by the golden yellow colour in the flower throats of his original nortieri collection, and noted the canary yellow flowers of his var giftbergensis and the yellow-green in the case of var montana. My observation generally is that throat colour is quite variable and cannot be used to rationally separate out lesser taxonomic ranks in the sense of a formal biological and meaningful classification.
Flowering time is often used as in defining species systems. Here again the irony seems to be that variation in flowering time of nortieri is, just as the case of variation in flower morphology, characteristic of the system. Generally flowering time for Haworthia populations covers approximately a minimum six-week window. Different species systems can be found to be in flower virtually throughout the year. In H. cooperi var gordoniana this is late spring to early summer and I have observed that annual climatic conditions may shift this window by as much as six weeks. In the case of the greater H. pygmaea system flowering has been observed to vacillate between an early spring and a late spring flowering. In the Subgenus Robustipedunculares, H. minima populations can differ in a flowering time of late spring or mid-summer. I observe that in nortieri, there are populations in the drier northwest (including agnis) that flower early in spring while in the Cedarberg, flowering maybe as late as summer (December).
Gerhard Marx wrote a good account of albispina and H. arachnoidea var. scabrispina growing together at Koup Station (Aloe 43:6, 2006) in which he notes that the flowering time of the former precedes that of the latter. This is what I also observed in the case of the same two elements growing in close association both north and southwest of Laingsburg. This is also the case at Nuweplaas northeast of Nieuwoudtville where a globosiflora-like element is in close association with H. arachnoidea. It had set and shed seed, while the latter was still flowering.
To be realistic one needs to now consider that we are discussing only about 42 records for an area extending from Groenriver and Bitterfontein in the northwest, down to east of Citrusdal in the south and then eastwards to as far as Prince Albert. This is an enormous area and the actual sample size must be very small in relation to actual occurrence of the species. However, it seems unlikely now that the range is likely to be extended, and that we have a fairly good representation of what the species looks like over its whole range. Further exploration will prove very time consuming especially in the northeastern Cedarberg which is rugged country and largely inaccessible.
Records, collections and illustrations
1. PVB6728 Groenriver (left). Groenriver is in Namaqualandand and inland from Hondeklipbaai. It is northwest of any other known H. nortieri populations. I am not familiar with the area and have not seen the plants. H. arachnoidea is recorded from various places in N Namaqualand but I am also not knowledgeable about its true distribution in that area. There is also a collection PV76745 (right) also from Groenriver that seems to be H. arachnoidea. It would be nice to know more about both these collections.
1. PVB6728 H. nortieri. Groenriver.1b. PVB6745.1 H. nortieri. Groenriver.
2. EA1497 Middelpos, Bitterfontein. Inland from Hondeklipbaai and north of Nuwerus. These are small plants and I also have not seen the population. The flowers are small and tend to globose.
2. EA1497.1 H. nortieri ‘agnis’. Middelpos, Bitterfontein.2. EA1497.2 H. nortieri ‘agnis’. Middelpos, Bitterfontein
3. PVB6146 Komkans. West of Nuwerus. Low lying area and the plants are in among white quartz pebbles. This was probably where Rolf Rawe collected this as a new species which was distributed by San Marino Nursery in 1969.
3. PVB6146.1 H. nortieri ‘agnis’. Komkans.3. PVB6146.2 H.nortieri ‘agnis’. Komkans.3. PVB6146.5 H. nortieri ‘agnis’. Komkans.3. PVB6146.4 H. nortieri ‘agnis’. Komkans.3. PVB6146.3 H. nortieri ‘agnis’ Komkans.
4. EA1442 and EA1443 Kareeberg Nuwerus. This is a low range of hills south of the Khamiesberg. Ewin Aslander also informed me that H. arachnoidea was present with nortieri, and both his collections are of very small plants. JDV95/11 and 95/12 ex EA H. nortieri ‘agnis’. Kareeberg, Nuwerus
4. EA1442.1 H. nortieri ‘agnis’. Kareeberg, Nuwerus.4. EA1443.1 H. arachnoidea. Nuwerus. H. nortieri ‘agnis’. Nuwerus.4. JDV99-12.1 H. nortieri ‘agnis’. Nuwerus.4. JDV99-12.2 H. nortieri ‘agnis’. Numerus.4. JDV95.11.1 ex EA H. nortieri ‘agnis’. Kareeberg, Nuwerus.
5. IDC53 Nuwerus. This collection was sent to me as the putative hybrid H. nortieriXarachnoidea. These were two very small plants under 50mm diam. And differ very slightly from Etwin Aslander’s arachnoidea plants from the same area, except that they seem to be marked with a faint reticulation. I am not sure they are hybrids and in fact do not know if H. nortieri hybridizes with any other species.
5. IDC53.1 H. nortieriXarachnoidea. Nuwerus5. IDC53.2 H. nortieriXarachnoidea. Nuwerus
6. MBB7522. Arizona. This locality is north of VanRhynsdorp on the Knersvlakte on the farm Arizona which now belongs to Western Cape Nature Conservation Board. This is south of Nuwerus and the substrate is shale. The species is also present on the adjacent Kwaggaskop farm belonging to Buys Wiese the well-known nurseryman. The plants are very truncated as the vegetation is sparse and low. The plants were recorded by Annelise leRoux and Steven Hammer who told me of it, and we were taken there by very kind favour of Kobus Kritzinger of Western Cape Nature Conservation Board. *** I have confused the true identity of H. agnis in the above 4 populations and it apparently is not the small plants in the Nuwerus area that I garnered from the place name given in the description, but simply the larger version at Arizona. That is an even stronger contender for inclusion in H. nortieri.
6. 7522.1 H. nortieri. Arizona.6. 7522.2 H. nortieri. Arizona6. 7522.6 H. nortieri. Arizona.6. 7522.9 H. nortieri. Arizona.6. 7522.10 H. nortieri. Arizona.6. 7522.11 H. nortieri. Arizona.6. 7522.12 H. nortieri. Arizona.6. 7522.13 H. nortieri. Arizona.
7. MBB7578. Nuweplaas P.V. Bruyns reported H. nortieri northeast of Nieuwoudtville. We visited the area in early summer and first found H. arachnoidea and then H. nortieri. Nuweplaas is in the extreme southwestern Bushmanland and Aloe dichotoma is virtually at the extreme of its distribution here. The substrate is Dwyka Tillite and the vegetation is Succulent Karoo. There are species like Aloe krapholiana and Euphorbia cylindrical on the farm as well so there is an unusual element in the vegetation. The plants were extremely well-hidden and the plants must be able to tolerate very dry and hot conditions here.
7. 7578 rt, 7577 lt. nortieri cf arachnoidea. Nuweplaas.7. 7578.1 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.4 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.7 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.10 H. nortieri. Nuweplaas, Nieuwoudtville.
8. PVB7167. Steenkampskop. This is the mountain between VanRhynsdorp and VanRhyns’ Pass. The plants are very similar to those at the foot of the pass and generally fairly similar to var. globosiflora, as indeed are those at Arizona and Nuweplaas. The flowers, however, are not globose.
8. PVB6167.1 H.nortieri. Steenkampskop, Vanrhynsdorp.8. PVB6167.2 H. nortieri. Steenkampskop, Vanrhynsdorp.8. PVB6167.3 H. nortieri. Steenkampskop, VanRhynsdorp.
9. MBB7523. Gifberg. This is the first higher altitude record southward from VanRhynsdorp at nearly 650m. The substrate is sandstone on the northwestern corner of this geological formation. The plants are smaller and with more lanceolate leaves than other records. The species has been recorded at Sandfontein a little to the southeast and also at the Koebee Pass to the east.
9. 7523.2 H. nortieri. Gifberg.9. 7523.3 H. nortieri. Gifberg.9. 7523.4 H. nortieri. Gifberg.9. 7523.8 H. nortieri. Gifberg.9. 7523.9 H. nortieri. Gifberg.9. 7523.10 H. nortieri. Gifberg.9. 7523.11 H. nortieri. Gifberg.
10. MBB7524. N. Clanwilliam. The var. montana was described from the Pakhuis Pass and it is not clear exactly where this is on the Pass. The eastern part of the Pass is dry Renosterveld and there is no confirmed record of the species from there to Doringbos at the northern foot of the Pass where var. globosiflora occurs. Derek Tribble described this locality on the western end of the pass, just north of Clanwilliam. At about 300m above sea level it is not really high in respect of the Cedarberg. The plants occur rather sparsely and are fairly solitary. Leaf shape varies from relatively abbreviated to more lanceolate and one plant particularly had leaves with linear translucent markings rather than rounded. It is generally difficult to make these observations under field conditions as the plants are well hidden and often withdrawn into the soil.
10. 7524.1 H. nortieri. N. Clanwilliam.10. 7524.3 H. nortieri. N. Clanwilliam.10. 7524.5 H. nortieri. N. Clanwilliam.10. 7524.6 H. nortieri. N Clanwilliam.10. 7524.7 H. nortieri. N Clanwilliam.10. 7524.8 H. nortieri. N. Clanwilliam.
11. MBB7579. EA1441 (top two). JDV99-11. Trawal Bridge. The species was first described from a collection made near the Bulkshoek Barage between Clawilliam and Klawer. I have seen it there, at the Doorn River Bridge and also on the northern rocky banks of the Olifant’s River near there. At the Trawal Bridge the plants are on a low lying very exposed and hot sandstone ridge. Note the flower photographed and how short the free parts of the petals are.
11. 7579.01 H. nortieri. Trawal Bridge.7579 H. nortieri. Trawal Bridge.11. EAs1441.1 H. nortieri. Trawal.11. EAs1441.2 H. nortieri. Trawal.11. EAs1441.3 H. nortieri. Trawal.11. EAs1441.4 H. nortieri. Trawal.
12. MBB7525. S. Clanwilliam. Whereas the other habitats have been out of or marginal to Fynbos Vegetation, this is true Fynbos at this site. It is west facing and very dry in summer. As the lichen growth shows, the winter conditions will be quite moist. There are several records south of this towards and southeast of Citrusdal which I have not been able to confirm. JDV99-11 H. nortieri. Trawal.
12. JDV99-11 H. nortieri. Trawal.
13. MBB6505 and 7595. N. Dwarsrivier. This is in the Central Cedarberg at about 800m. The plants are on large loose boulders lying in the valley floor. When I first saw the plants I was very impressed with the number of leaves in the rosettes and their attenuation. These impressions can be very unreliable. I still have a large specimen in cultivation but revisited the site recently to establish that the plants are within “the normal range of variation of the species”. I write this rather facetiously because it is actually very difficult to obtain a true impression and my long experience of plant descriptions and identifications suggests to me that taxonomy is characterised by such subjective judgements.
13. 6505.2 H. nortier. N Dawrsrivier, Cedarberg.13. 6505.3 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.4 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.5 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.6 H. nortieri. N Dwarsrivier, Cedarberg.
14. MBB6506. East Dwarsrivier. This collection first came to my notice as a plant said to have been collected by Clive McDowell at the Wolfberg Cracks in the Cedarberg. The plant was H. reticulata and hugely improbable. Fortunately I was able to vist the area. The plants are on a steep south facing riverine rockface and are H. nortieri. They are proliferous and the leaves are shorter and broader than is normally the case.
14. 6506.1 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.2 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.3 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.f1 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.f2 H. nortieri. Dwarsrivier, Cedarberg.
15. ADH2514. Breekkranz. This is a little further south. There is a hiking trail from east of Citrusdal towards Kromrivier. The collector was Adam Harrower and the single plant in cultivation seemed rather robust and thick-leaved. We visited the area and while not reaching the place along the Breekkranz trail indicated by Adam Harrower, we found the plants in the rocks around the Bakkamerfontein ruin. They were abundant and well hidden in rock crevices at about 800m.
15. ADH2514 H. nortieri. Breekkranz, Cedarberg.
16. MBB7580. Bakkamerfontein. The nortieri plants are just at the small distant upper notch on the skyline.
Blinkberg EvJ19550 is taken from just below and looking about 30km to the Sneeuberg (not Sneeukop) in the distance. Only four other populations were recorded in the scanned landscape! The view is just as impressive southwards where three populations would be covered. It is all Table Mountain sandstone – except the Blinkberg itself. Seen in that context, the pygmaea issue is just a small blob in a much more diverse landscape. Photos take at about 10:00am.
16. 7580.1 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.4 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.7 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.11 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.13 H. nortieri. Bakkamerfontein, Krommerivier.
17a. EvJ19550. Blinkberg. Ernst van Jaarsveld has been very helpful in recording interesting populations from inaccessible places. The plant he showed me was also very robust and similar to Adam Harrower’s Beekkranz specimen and I felt it was necessary to visit the site and the plants in habitat.
The nortieri plants are just at the small distant upper notch in the sky-line.EvJ19550.a Blinkberg.
The locality is nearly 30km southeast of Kromriver and moving out of the Cedarberg into a mountaineous area refrred to as the Swartruggens. The geology of the area is fascinating. The north/south road between ceres and Wupperthal runs along the intersection of table Mountain Sandstone to the west and the younger Bokkeveld Shale to the east. The layered rocks is tilted up in the west and down in the east . Because there are harder quartzitic bands in the Bokkeveld formation, the terrain is stepped and one climbs a series on near switchbacks to get to the steep upper more quartzitic Witteberg sandstone which caps the mountain. The Bokkevelf shale here is not rich in succulents and the Haworthia was in a lower band of Dry Mountain Renosterveld at about 850m. The plants were small, well hidden under grass and small bushes and unlike the specimen in cultivation. Again it is impossible to gain an overview of the variability, but I photographed three specimens to show that the leaves can vary from spotted with the translucent dots typical of H. nortieri, or none. The plants here are non-proliferous. The landscape scene among the pictures is a view towards the Central Cedarberg and the pointed peak in the middle distance is Cedarberg Sneeuberg at 2027m. It is equally impressive looking south. It is a rockbound landscape which is difficult to move through and it would require a most remarkable effort to get a better idea of the occurrence and distribution of the plants in that whole area. As it is, we have no records for a huge area extending from Blinkberg in the south to the Pakhuis pass in the northwest, and to the Ouberg Pass in the east near Sutherland. Mrs. J. Marais of Mount Ceder Resort (Blinkberg) reports the plants at a place east of Matjesrivier to the north. H. arachnoidea is not known in the area at all but it does occur as a small densely white spinose form at the SkitteryKloof to the southeast. Curiously the only other record of Haworthia for this area is of a plant like H. venosa var. granulata collected by Elsie Esterhuyzen on top of the Skurweberg Sneekop to the south of Blinkberg. I did climb that peak in 1973 in the hopes of finding the plant, but was unsuccessful.
17. EvJ19550.2 H. nortieri. Blinkberg.17. EvJ19550.3 H. nortieri. Blinkberg.17. EvJ19550.4 H. nortieri. Blinkberg.17. EvJ19550.5 H. nortieri. Blinkberg.EvJ19550 H. nortieri. Blinkberg.17. EvJ19550a H. nortieri. Blinkberg, Cedarberg.17. EvJ19550.7 H. nortieri. Blinkberg.17. EvJ19550.8 H. nortieri. Blinkberg.17. EvJ19550.9. H. nortieri. Blinkberg.EvJ19550.6 H. nortieri. Blinkberg.
17b. MBB7597 & 7599. Kunje, SE Citrusdal. The German team Muller-Doblies showed me a plant from this area. There are other herbarium records from the Waboomsberg further north, but also from the Elandskloof slightly to the west and from the Heksberg to the near south west. The plants are abundant on flat rock sheets where there are small pockets of soil and sparse vegetation. This gets extremely dry in summer while in winter it can be very wet. The plants can be proliferous and there were some very dense localized aggregations of individuals. Size varied quite considerably from small exposed specimens 30mm in diameter, to shade protected plants to 100mm diameter. Flowering seemed to have occurred in late November to December.
17b. Terrain south of Kunje.17b. 7597.1 H. nortieri. Kunje.17b. 7597.2 H. nortieri. Kunje.17b. 7597.3 H. nortieri. Kunje.17b. 7597.4 H. nortieri. Kunje.17b. 7597.5 H. nortieri. Kunje.17b. 7597.6 H. nortieri. Kunje.17b. 7597.7 H. nortieri. Kunje.17b. 7597.8 H. nortieri. Kunje.17b. 7597.9 H. nortieri. Kunje.17b. 7597.10 H. nortieri. Kunje.17b. 7597.11 H. nortieri. Kunje.17b. 7597.12 H. nortieri. Kunje.17b. 7599.1 H. nortieri. S Kunje.17b. 7599.2 H. nortieri. S Kunje.17b. 7599.4 H. nortieri. S Kunje.17b. 7599.5 H. nortieri. S Kunje.17b. 7599.6 H. nortieri. S Kunje.17b. 7599.7 H. nortieri. S Kunje.
18. MBB6158 Skurweberg, Sneeukop. We found these plants in grass in cracks in massive sandstone slabs at the foot of the higher peaks. We also saw them in similar but dryer conditions in the Witteberg formation east of the Katbakkies Pass and west of Skittery Kloof. The whole area seems to provide endless suitable looking habitat and it is surprising that there are not more records, except that people who move around these mountaineous areas are seldom focused on small succulents.
18. 6158.4 H. nortieri. Sneeukop, N Ceres.
19. JDV96-56. EA801. Nevertheless, it is really strange that we have to move nearly 30km south to OpdieBerg to where Etwin Aslander found H. nortieri again. Grown by Etwin, the plants were extraordinarily beautiful being highly colourful and well spotted and very reminiscent of the var. globosiflora except again for the flowers. Again the area is very awesomely rugged and even here one wonders what could have motivated Etwin to have even searched there.
19. JDV96-56.1 H. nortieri. Opdieberg, Ceres.
20. MBB6815 Audensberg and 7526 Die Nekkies (bottom left). I include these two collections in the discussion because they reflect the greater problem in Haworthia, which is to establish the relationship of the different species systems. H. maculata is essentially linked to H. herbacea and H. reticulata, in the Worcester vicinity and well south of OpdieBerg. The two collections illustrated here are from two different biogeographic zones on either side of the karoid zone occupied by H. herbacea. They could thus be as genetically distant from one another as they are from that species. The Die nekkies plant has a browner colouration than the Audensberg plants which can have the purplish colouration of H.nortieri. The flower seems convincing in that the Audensberg plant has the typical fish-tail buds of the southern Haworthia.
20. 6815.1 H. maculata. Audensberg.
21. EA1508 Hex River Pass. This is again a notable collection in an inauspicious looking area. It is in the eastern leg of the Cape fold-mountains in the eastern end of the Hex River valley. The plants have to be considered in the same light as the previous comment. H. arachnoidea is present to the southwest as a very spotted form which could suggest nortieri-like character as well as confounding the situation with regard to H. marumianai var dimorpha. There are two other questionable populations. One is at Sandhills to the west which is a weakly spinose form of H. arachnoidea and then one in the Rabiesberg to the southwest which suggests an affinity with H. herbacea. What this exposes is the paucity of information. There simply are not enough known populations to help any analysis of either the morphology (as non-decisive as this could be) or the biogeography of the species, which is the main ground on which I base my classification. The nearest known H. nortieri population is that at OpdieBerg in the west, Sneeuberg in the north and then var. pehlemanniae at Matjesfontein in the east.
21. EA1508.1 H. nortieri. Hex River Pass.
22. MBB7070. Laingsburg. My few illustrations of H. nortieri var pehlemanniae reflect how little is actually known about the real distribution of many of the species. I have no doubts about the continuity of this element with var. globosiflora in the northwest. It is known at the OubergPass near Sutherland in close proximity to H. arachnoidea. There is the unconfirmed Bruyns’ record from the Elandsberg, Etwin Aslander’s at Klipfontein in the Moordenaarskaroo north of Laingsburg, this collection from just north of Laingsburg, records from just north of Matjesfonein and the original location southwest of Laingsburg on the Agter-Witteberg road.
22. 7070.1 H. nortieri var pehlemanniae. N Laingsburg.
23. MBB7594 Koup. Hayashi must perhaps be commended for recognizing this as different from H. arachnoideai var scabrispina which grows adjacently in very karoid conditions. Both species of course also grow in proximity at Laingsburg. Superficially they could be taken to be the same when in fact the spination and colouration is really different as should be noticable from the illustrations where the origins of the name “albispina” is obvious. Gerhard Marx has written of these populations in Aloe 43:6 (2006) and also illustrates “albispina” from a little further west.
23. MBBsn. H. arachnoidea var scabrispina. Koup.
24. MBB7385. Prince Albert. In the company of Sue and Richard Dean I quite fortuitously found this population discovered by V. DeVries at Prince Albert. It is very far east for the species, but my contention is that species have range and variation. They need to be considered as systems not as single specimens which look a bit different from one another in some sort of directly or indirectly descriptive way. These plants do indeed look a little different, but to make it clear, only one plant is illustrated. The landscape is very similar to that at Arizona north of VanRhynsdorp. The plants have to be retracted into the soil and one cannot form a good opinion without destructive sampling.
24. 7385 H. nortieri var pehlemanniae ‘devriesii’. N Prince Albert.
Conclusion This account is by no means is absolute. I have only illustrated about a half of the known populations and wonder just how many there are which have not been recorded in anyway at all. Of course it can be imagined that in some way more information will better indicate taxonomic entities. My experience persuades me that just the opposite will occur. As my knowledge of the plants in the field has grown, so has my conviction that we are dealing with highly integrated and variable systems. Our taxonomic systems and our understanding of them, especially in the learned ranks where we would expect to find it, are simply inadequate. Our system of classification is based on a belief system that there are simple diagnostic keys which can be derived from simple morphological characters. I think that this is because classification has been built on a slowly growing but still inadequate sampling base. The International Code for Botanical Nomenclature dominates taxonomic thinking which means that nomenclature and the management of names, overrides the knowledge that these names should convey. More attention needs to be paid to explaining different plants as systems that relate to geographic factors and other similar systems.
In terms of the general taxonomic paradigm, it may be reasonable to suppose that the “nortieri” system is composed of discrete taxa. I seriously question that this is so. As the record grows, so does the pattern of increasing variability of otherwise recognizable systems. As regards var pehlemanniae (“albispina”, “devriesii” included) I am quite convinced that even growing these plants in mass from seed cannot produce anything which alters the fact that they belong in this single system which I have discussed in this article. Given the time, the opportunity and the youth, I would try to explore and illustrate these eastern populations in greater depth as well as try and build substantially on the known record. There are huge areas that offer suitable habitats, and similarly huge areas that do not. These areas are largely inaccessible by virtue of the limits of the road network and then the problem of gaining access via land ownership. ♦
Latin names definitely mean different things to different people and my contention is that the real essence of these names should, in addition to their many other usages, be in the relation of plants to their origins, relationships, behaviour and imagined future. A classification can only have the authority that experience and knowledge permit, and be really evaluated and understood by persons with the same evidence before them. In coming to closure I have been exploring some more and with my wife Daphne, made two finds which further convince me that we have to come to a classification by agreement. However, the requirement is that species are seen to be highly complex systems with none of the rigidity and inflexibility that nomenclatural rules imply, nor any of the egocentric authoritarianism that a history, of which I have been a part, suggests.
I have recently written two papers. One deals with the H. notrieri complex, and the other with H.pygmaea. In writing the latter I interwove the comments of observers whose opinions I value, and conclude with the proposal that the elements acuminata, splendens, dekenahii, argenteo-maculosa, fusca and vincentii all be included in the super-species H. pygmaea. Concomitantly I suggest the H. turgida and its variants be absorbed in H. retusa. Behind these two propositions, I was intending to re-enforce my view expressed in Haworthia Update Vol. 3 that H. mirabilis too be re-structured to include magnifica, maraisii, heidelbergensis and any associated varieties. The reason of course been the problem of continuity, however that is understood; and I suggest it is best understood by familiarity with the plants in the field and some recognition of biogeographical factors and the role these may play in driving change, adaptation and evolution.
In Update Vol. 3 I illustrate examples from many of the populations known to me, and also draw attention to populations in the lower Breede River valley which are significant. My most recent excursion has in fact been to try and resolve the dramatic juxtaposition of variants that we found there. We have Adam Harrower’s collection from Sandhoogte and Chris Burger’s collection at Buffelsfontein, both south of the Potberg. Then there are my collections from Stoffelsrivier and one nearer to Infanta. A curious addition I have yet to see in the field is a collection by Ismael Ebrahim of SA Biodiversity Institute from southwest of Vermaaklikheid. That collection resembles my Stoffelsrivier collection and so provides the inevitable link to paradoxa.
At two localities near Melkhoutrivier, between Stoffelsrivier and the Infanta collection, we found plants which have to be related to those mentioned above. The plants are highly variable, as I have now come to expect, and individuals can be likened to badia, acuminata and mutica. The surfaces have a curious sheen and may be incipiently spiny as the Sandhoogte and Buffelsfontein plants also are. The paradox is that we are also driven to the conclusion that plants at Ziekenhuis are the same species.
What re-enforces this seemingly improbable statement, apart from my lengthy dissertation on the matter in Update 3, is a second find we made southeast of the Bromberg. This is the locality for what I suggest is H. rossouwii var. elizeae, and also for several variants of what have, in the past, been classified as maraisii and mirabilis or variants thereof. My new find is a remarkable array of large very dark-green (nearly black), plants which dramatically enforce the close association between mirabilis and maraisii as well as with heidelbergensis. These plants include the elements of both “species” as they might have been understood. What is striking is again the huge variation in the population that I have come to expect in this complex. The plants do lack the opaque dots on the under-leaf surfaces while the upper surfaces may be clearly windowed or opaque. They bring forcibly to my mind a sequence of populations from Heidelberg in the east to Verdwaalskloof near Riviersonderend in the west that I suggested forged the interface of mirabilis and maraisii. Coupled with this is the array of populations both north and south which enforce that continuity and that also lead on to heidelbergensis.
My recommendation is that the nomenclatural code be summarily assigned a secondary role to a dispensation which is more flexibly attuned to the realities of a truly asymmetric species structure with more emphasis on reflecting field relationships. The illusion of reality that the ranks of genus, subspecies and variety provide, should be admitted. Genera should be recognised for their historical value, but for species I would suggest that much more attention be given to the huge asymmetric and asynchronous variability that underlies capacity to change and adapt. With the adoption of H. mirabilis as a superspecies, variants can be indicated by the addition of any other epithet in inverted commas; thus H. mirabilis ‘maraisii‘, H. mirabilis ‘heidelbergensis‘, in the knowledge that there is in fact no clear and infallible distinction. ♦
Statistical analysis of two populations of Haworthia mirabilis (v.Poelln.) M.B.Bayer
M.B.Bayer & L.M.Loucka
Introduction: In discussing H. rossouwii (Aloe 38:31, 2001), Bayer mentions the possible continuity with H. mirabilis var. sublineata. But any comment like this is complicated by the problems of variation, description and circumscription. We want to discuss the variation in the latter element and indicate further where the problems are in the delineating species and varieties. It seems that one of the assumptions of classical plant taxonomy is that of linear dichotomy, black and white, this species or that, and also that there is hierarchical and consistent in-group similarity to some unstipulated degree. Haworthia, and particularly the subgenus Haworthia, presents a problem to those interested in the genus in that the classification is confused and that identifications are difficult. Attempts been made to explain that the classification is confused by the perceptions associated with classical taxonomy, and that the sharp and precise discontinuities suggested by a ‘key’ to the taxa, simply do not occur in the subgenus Haworthia, in fact they do not occur in many other genera, and this simple truth seems to be difficult for some to accept.
Thus this article includes a report of a study done on a batch of seedlings of H. mirabilis var. sublineata. It shows that there is very little probability that one could quantify separation of this from other populations presumed to be the same species.
Outcomes in the biological world are influenced by genetics and environmental factors, but also by chance events. It is because of the probability of chance events that knowledge of statistics is essential to scientific research and our studies of the genus Haworthia.
Plants have parts that can be mechanically measured in terms of spread, height, width, length, thickness, arc; or counted as in the number of leaves or offsets. Other attributes like colour, and surface texture are more subjective. When we observe leaf structures in Haworthia our perception often tries to make sense and draw patterns from many subtle characteristics viz. this plant is species X because it has a pattern that matches our experience. But our brains can be easily fooled when we try to compartmentalize. When we subject our observations to measurement we begin to sort out the characteristics we use to describe one plant, population, or species from another. Viewing this collection in the leisure of the greenhouse, we can see that some leaves were thicker or longer than others, but are they “really”? That is, are they significantly thicker, or longer? When we count the number of leaves, or measure their length can we make some statements to compare and differentiate one population from another? Do the differences we measure translate into differences in overall size and shape of the leaf? Let’s investigate this further by formulating the following question:
Method: Seed of H. mirabilis var. sublineata was field collected at the type locality south of Bredasdorp in April 1997, accessioned as MBB6639. By Sept.2000 these seedlings were at flowering size and 71 plants were available for the study. A second study sample was taken from a population of H. mirabilis var. triebneriana from Fairfield, west of Napier (MBB6643). H. mirabilis is very widespread and this Napier population is apparently no more or less variable than many others known.
Population 1, MBB6639 – 3420CA South of Bredasdorp. April 1997. North facing steep rocky slope, Table Mountain Sandstone with grassy mountain fynbos.
Population 2, MBB6643 – 3419DD Fairfield 15km West of Napier. April 1997. North facing shale ridge, Bokkeveld Shale with grassy renosterveld.
Prior to this study, an attempt was made to reduce the size of the group of MBB6639 seedlings by discarding any single clones which were obviously similar. To do this Bayer tried to arrange the plants according to similarity. He could not find any pattern, and each plant appeared to be fairly unique. Five plants were removed in order to propagate them vegetatively, and for this study a further four clones were removed which were either obviously atypical or retarded. Four smaller apparently under grown clones were retained in the final study batch of 62.
A first attempt was again made to try and identify groups by visual comparison. This proved impossible to do. Attempting to reduce the size of the set by the identification of “identical” pairs also failed. No two plants were sufficiently similar to be regarded as near to identical. The simplest way forward was then to match the plants as closely as possible into pairs, repeating the process five times. After pairing, the pairs were broken and the plants returned to a random array. Pairing was then done again by picking up any one clone and finding the best match. Some re-sorting was done in order to accommodate the last of the clones as the pairing process neared an end. In some cases clones which could not be adequately paired were compared over all the others to find the best match. Although an attempt was made to score the “goodness” of pairs, this proved difficult to do and an inconsistent result was obtained. Some clones simply proved more difficult than others to match.
With five attempts, the total pairings was 310. Sixteen clones were unpaired. Of the pairings, 210 were unique, in 74 cases the same two clones were paired, in 18 cases the same clones were paired three times, in 8 cases four times and in two cases, all 5 times. This shows that there is very little consistent pattern in the group as a whole when trying to match or differentiate based simply on assessing visual cues.
Recognising now that no progress was being made, Bayer considered what ‘characters’ he was using in his visual assessment and then quantified his observations. Some of the characters used in the visual process could include:
a. Size b. Colour c. Leaf number d. Leaf width e. Leaf length f. Leaf thickness g. Leaf recurvature: i. compactness, ii. erect or reflexed h. Leaf marginal spination i. Leaf upper surface: i. translucence, ii. texture, iii. venation, iv. basal opacity, v. basal marking j. Leaf lower surface (as above) k. Proliferation (off-setting)
For statistical analysis it was decided to use the measured spread of the leaves from the tips of the most extended leaves, the height reached by the highest leaf tip measured from the base, the width, length, and thickness of the largest leaf at maximum, and to measure the arching of such leaves by the gap between a line drawn from base to tip and the lower leaf surface (i.e. recuravture). A subjective measurement was made of leaf surface roughness, size and number of spines, and degree of tubercles. The resulting data was analyzed by a Dotplot for each characteristic and a Two-sample T-test used to assess the null hypothesis.
Descriptive Statistics
The MBB6639 plants show the classic bell-shaped normal distribution for most characteristics with the exception of spread and proliferation (off set count). Perhaps the population culling of the runts and natural survival account for the truncated spread data. In time more of the clones may have offsets and thus show a different distribution. For the other characteristics we can see a central tendency and data points spread to the left and right. It’s this central tendency, or average (statistically called the mean) that we’ll study further.
Hypothesis Testing
Question: Are plants from the two populations in this study of the same or different sizes and shape?
Hypotheses: The difference in the average physical characteristics between the two populations is zero; i.e. the null hypothesis (Ho:). If the difference between the means of the samples is among those that would occur rarely by chance when the null hypothesis is true, then the null hypothesis is rejected and the investigator describes the results as statistically significant. That is, the difference between the average of each population characteristic must be great enough to conclude if the populations are the same or different.
Prediction: If the leaves of the two populations are the same size, then the spread, height, width, length, thickness, and arc of the populations should be the same.
Analysis:
The P value we subject our data to is 0.05, or a 5% chance that we would accept that the difference is the means of the two populations is zero when in fact it isn’t.
A null hypothesis is not accepted just because it is not rejected. Data that is not sufficient to show convincingly that a difference between means is not zero do not prove that the difference is zero. Such data may even suggest that the null hypothesis is false but not be strong enough to make a convincing case that the null hypothesis is false. For example, if the probability value P were 0.15, then one would not be ready to present one’s case that the null hypothesis is false to the (properly) skeptical scientific community. More convincing data would be needed to do that. However, there would be no basis to conclude that the null hypothesis is true. It may or may not be true, there just is not strong enough evidence to reject it.
What can we conclude? The two populations appear to be different for leaf width and thickness. For all other characteristics we just don’t have enough data to make any conclusive statement.
We have illustrated the individual clones for the reader to appreciate how different the individual plants may be, and also to consider that the appearances of the plants will vary according to growing medium, exposure to light, season and watering.
Discussion and Conclusion
Our observation is that a statistical approach simply does not penetrate the nuance of difference that visual observation can discern. The data we present is valuable only in that it illustrates the variation within the population MBB6639. The coefficients of variation of the characters used are so high that larger samples would be required to provide more reliable statistics. In this study we have used a second population 6643 which is obviously different. At Bredasdorp there are in fact two other populations. One of these is on a north-facing river bank in among river-eroded sandstone boulders, while across the river about 250M away is a smaller population in Bokkeveld shale. Plants from all three populations are relatively easy to identify. The riverine boulder plants have fewer stumpier and more reflexed leaves, while the shale plants have erect leaves. To what extent these differences are “real” cannot be proven without a replication of the seed collecting and propagating effort done for 6639. In the context of ALL the populations which may comprise the species H. mirabilis it is quite evident that this is impractical. ♦
I was in the field yesterday (March 14th -2007) and then in reflection thought I would relate it to why I write and to what I have written. Someone had been at Sanddrift Drew to look for the Robustipedunculares that grow there, and reported that they could not find H. marginata. This was a bit disturbing to me because that marginata has very slender long leaves and in the vicinity it also hybridizes with H. minima and H. pumila. The particular locality is a fairly prominent flat-topped hill which seems to have been formed from river gravel. Despite being so rocky and fairly steep-sided, the hill has been very severely impacted on by agriculture. The southern Cape soils are very skeletal and agriculture is fortunately concentrated on the lower flatter slopes and to the alluvial flatter areas and eroded shales which can be machined to lands. Rainfall is in winter and distribution is very variable across the landscape. Rainfall patterns have also apparently changed with time, at least as agriculture seems to have developed. There have also been economic and social changes which have altered the fabric of agriculture. There was a time when farming was a way of subsistence. Tractors and fuel was cheap and there was an endless space to tame. The consequence is that huge areas of very marginal land was ploughed and contoured for cropping. This was the fate of most of the Sanddrift hill and H. marginata was thus reduced to a narrow band of Renosterveld vegetation above the very last contour reaching to near the top of the hill. The particular farm seems to have teetered on the verge of failure as both a subsistence farm and a commercial venture for the last forty years. Several very dry periods in that time have driven various owners to financial despair. Presently, however, the farm seems to have fallen in to the hands of what may be a new form of commercial colonialism. It is owned by an English gentleman who has the resources to farm aggressively. Water sees to have been obtained from an expanded and more flexible irrigation scheme and the farm has entered a new phase of development. This is of course happening throughout the country and the threats to the fragments of undisturbed vegetation and rocky outcrops which have given me so much joy are now hugely disturbing.
Haworthia pygmaea now exists (the story is told elsewhere – “The White Widow Reunion” Haworthia Update Vol.2) as a few small seedlings under ageing Pteronia incana in a tiny fragment of very disturbed vegetation near the Sanddrift homesteads and farm buildings. The marginata/pumila habitat along a lower stream is now grossly disturbed by fence and passage way. The marginata/minima habitat was in any case a low road verge cutting fortunately left intact when the road was straightened and moved. Because of the rock and the old road fence which was never removed the habitat has only been subject to grazing pressure. The plants have suffered and the population just survives because I collected and grew seed and then re-planted seedlings. I did unfortunately not do the same for the H. marginata that I thought was relatively safe. Yesterday we found a single plant. We cannot say for certain that it is the only one, so I know that something needs to be done to see if there are plants and propagules somewhere to plant back. There are many other small populations and habitats which are less isolated and more heavily impacted than this one. The future does not look good for them.
It seems almost weird and uncaring to bother much about the classification and names of plants when their very survival is at stake. It is also curious that classification is also seen as an egocentric activity and route to name and fame, rather than as the language and communication process we all use to feel our blind way about creation. What then did I learn from yesterday? What I can add to what I was thinking the day before?
I have been writing about closure. After 45 years of writing about Haworthia have I learned anything? My inclination is to say that I cannot in fact make a final statement and put forward a set of names which will satisfy any universal need and provide an adequate handle to what is out there. Yesterday I was also seeing plants of the genus Acrodon and wondering if it could help me with understanding Haworthia. It seems to me that Acrodon bellidiformis may have the same relation to A. quarcicola that H. mirabilis has to H. maraisii, and probably suggest that other Acrodon species may just as easily be superfluous names which just reflect spatial variation.
This brings me then back to Haworthia names and classification. I have been saying that it is now up to the community to decide what system of names they want to adopt. My contribution is only to have attempted to explain what I have explored in the field and what names or words I can use to do so, against my understanding of what others have done and said. My latest manuscript is entitled “The brutality of reality” in which I expressed the point of view that a formal set of names following the principles of the code for Latin names, should be taken far more lightly. I suggest fewer binomials and abandoning the concept of “typical variety” such as H. mirabilis var mirabilis and any formal designation of ranks such as “subspecies” and “variety” or even “cultivar”. Such names do not have, as I now like to quote, any semblance of reality. They only have meaning in relation to personal knowledge and experience. My application and usage of names in Haworthia has been shaped by my experience of first the literature, then the plants and then personal communications; all changing with the passage of time. My own writing is now available only as a source from which an interested person or group of people can extract and distill names which mean something to them in relation to their particular experience.
The conclusion is that a classification and its names only have meaning in relation to the “truth” (experience, skills, observation, education, intellectual capacity and thus knowledge) of the observer in relation to the “truth” of the user. Conservation therefore should have as its primary motive, the minimization of any human impact on natural habitat which will lead to further losses in the diversity of the different phenomena we observe in our view of creation. Truth itself has no reality within the range of our attachments and interactions within the creation. ♦
In Alsterworthia 7(1):22 Breuer states “No records have been found to indicate that this plant has been discovered before and as it is dictinctive I have decided to name it as a new taxon”. This population is recorded in the old collecting record of G.G. Smith and I searched on the calcretes further to the east as far back as 1969. Unfortunately it never occurred to me then to even look at the remnant of rock in an area largely destroyed by road-building operations. Presently this small ravaged quartzitic outcrop is bisected by a meaningless road which is fenced and I did find the plants there in 2004 – name the place SW Karsriver. Why I looked is because of the mindless destruction of a small valley habitat on the Karsriver about 3km further northeast where a magnificent form of Aloe brevifolia once grew with a population of H. maraisii that has gone with it. I was thus anxious to confirm a maraisii so close to Bredasdorp for reasons best explained elsewhere (Update 3 Chapter 1). Morton Cumming apparently found more than the three plants I saw there across the fence on the north side. I recognized the plants as minima/marginata hybrids and was also a bit nonplussed by the absence of putative parents. Minima was only known at Mierkraal far to the southwest and marginata is known about 10km further to the northeast. I was disturbed by the fact that I could only find the three plants and in February 2005 I visited the site again and collected seed under MBB7453. Cumming seems to have been at the site also early in 2005 and claims to have seen many plants, which surprised me. In the past the site has been grossly disturbed and a constant pain to me is that major road-construction in the late 1960era led to the use of rock outcrops such as this, as gravel sources. The badia-locality at Napier became a major gravel source and could be seen as a huge white scar on the landscape from afar afield as Swellendam. Thus this site at Bredasdorp suffered the same treatment and the land surface has been transformed with the removal of surface rock and gravel. Only the smallest fraction is left and I do hesitate to report the survival of “maraisii” on virtually a single quartz rock remaining on the south side of the road pictured in Alsterworthia. I cannot believe that I would have missed any plants in the area available to be searched. Farming in the area is not mainly devoted to “merino-sheep and grain crops”. Farming in the area has become highly commercialized and water is exported from afar afield as the Theewaterskloof Dam at Villiersdorp. Grain crops are unreliable and with this artificial supply of water, farmers have turned to ostriches and dairy cattle. The result of feed-supplementation has resulted in higher stocking densities and greater trampling and damage to natural vegetation. This has put tremendous pressure on pockets of surviving vegetation that is also exacerbated by a turn to dual purpose Dohne-Merino sheep breeds that graze more aggressively than the original Merino. Additional to this is the destruction of roadside vegetation in what appears to be a deliberate policy of road-engineering to clear verges to the farm fences, and the dreadful application of herbicides for the fear of weed-seeds contaminating crops from those road verges. The possibility that this herbicide application and disturbance of stable natural roadside vegetation will certainly lead to greater weed problems in the future, is left for that dark future.
If there were more plants there at SW Karsriver in 2005 there certainly are not now. When I revisited the site in 2006 one of the three plants had been dislodged presumably by grazing animals, and we replanted it across the fence. At a later visit we found that the plant had sadly died or else, having been in the fenced zone, may be the “small two plant” now gracing a herbarium specimen? In February this year, 2007 I again visited the site in passing and saw that another plant had been broken off. The crown was resprouting and I removed the main body of the plant to grow on in cultivation. Together with this I can report that Sheilam has very successfully germinated the seed I collected and has given me about 20 seedlings for further cultivation and we will return these to the site in due course. Thus 1.5 specimens of H. mortonii represent the species and I hope Sheilam makes a fortune selling this now gravely endangered taxon.!
Breuer in his article makes a reference to MBB6633 as “also this taxon” viz H. mortonii. I already have a problem in that I think professional botanists have reduced taxonomy to a playing field where “the most ignorant and uninformed parade as taxonomists”.
Unfortunately the fragmentation of the literature and the existence of a privately operated journal exacerbate the situation enormously. Both fortunately and unfortunately, it also provides me with a public platform. This MBB6633 is simply H. marginata and what Breuer has observed in cultivation with respect to the two populations and his acquisition of material probably has as much to do with the disappearance of plants as do animals and road-building. Incidentally I periodically visit the marginata at Adoonskop as the northerly population is known. Now in 2007 the plants are very severely grazed down to ground level and the landowner is contemplating turning that non-arable 140ha into a fenced game camp with accompanying ecotourist facilities. Curiously Cumming has reported (private communication) small marginata still further north and I suppose further “research” is going to result in yet another superfluous taxon.
Not over yet! A population of H. mirabilis (var sublineata) used to occur on the south bank of the river course (This river is named Dryriver because like a few “rivers” in the area it only holds water in winter) that runs west to east immediately north of Bredasdorp. In distress at the loss of this population I scoured the wider area to see if by chance it occurred elsewhere. By virtue of a minor miracle we found seven scruffy and bedraggled specimens surviving within a grove of gum trees, covered by a fallen litter of old gum seed capsules, leaves and branches. How they have managed to survive for what must be 50 years or more is very difficult to believe. The site is virtually the same as an historic laundry concretion dating from the 18th century. In searching for the plants we had to scratch and scrape among the litter, raising huge doubts and difficulties in respect of more disturbances and of conservation. Very curious was the additional discovery of a truly depauperate and chlorotic specimen of H. minima from the wreckage. Thus there is no doubt that H. minima was never far from the mortonii locality. It certainly was known a little further east and Breuer is again inaccurate in his reporting that the “coastal area areas from Bredasdorp and further to the south-west are not very well explored for haworthias”. The area is extremely well-known in general botany and has also been explored specifically for haworthias. A proper view from both these perspectives is that this is the southwestern boundary for the genus and it is unlikely that further exploration is going to yield anything new – i.e. based on a rational opinion.
Here I want to point out something. I do not hold a collecting permit from nature Conservation anymore, and yet I have removed the plants from the gum-tree litter at Bredasdorp and also the broken specimen from the DMC10485 site – for which I am fairly confident no permit was issued for so-numbered specimens either. The reason that I do not apply for a permit is manifold. Primarily I suppose it is because I feel I am busy passing my sell-by date (comforted by the fact that some people never were saleable). Secondarily I feel humiliated by the process and the scrutiny of people whom I do not think are knowledgeable or really interested. Thirdly by the challenges of conservation where my activities – however many plants I might remove – are as nothing compared to the mindless destruction of habitat by roadworkers, farmers, other landowners. There seems to be nothing in an environmental impact assessment which lists species by name, that draws anything from those names and bears on the fact that we have living things of different kinds in our living space and we should be very careful indeed that we call it that. The permit system seems to me to be a way of harassing interested people and worded solely to improve the probability of successful prosecution should officials be so lucky as to fall over someone removing so much as a seed from the field without a long list of provenances. There are severe changes taking place in respect of our environment and I think that foolish taxonomy is doing nothing towards helping constructive engagement with conservation issues like this SW Karsriver site presents. Not to speak of proving information which can usefully be added to the knowledge-base we all should gainfully share. ♦
My experience is that Latin names definitely mean different things to different people. I submitted this manuscript as a draft to various people and the response varied from one which was nil, to some sort of general accord. I am, however, no longer confident that botanists either do or will agree with my contention is that the real essence of Latin names should, in addition to their many other usages, be in the relation of plants to their origins, relationships, behavior and imagined future. A classification can only have the authority that experience and knowledge permit, and be really evaluated and understood by persons with the same sort of evidence before them. In coming to closure I have been exploring some more, and with my wife Daphne, made two finds which further convince me that we have to come to a classification by agreement. However, the requirement is that species are seen to be highly complex systems with none of the rigidity and inflexibility that nomenclatural rules imply, nor any of the egocentric authoritarianism that a history, of which I have been a part, suggests.
I have recently written two papers. One deals with the H. nortieri complex, and the other with H. pygmaea. In writing the latter I interwove the comments of observers whose opinions I value, and conclude with the proposal that the elements acuminata, splendens, dekenahii, argenteo-maculosa, fusca and vincentii all be included in the super-species H. pygmaea. Concomitantly I suggest that H. turgida and its variants be absorbed in H. retusa. Behind these two propositions, I was intending to re-enforce my view expressed in Haworthia Update Vol.3, that H. mirabilis too be re-structured to include magnifica, maraisii, heidelbergensis and any associated varieties. The reason of course been the problem of continuity, however that is understood; and I suggest it is best understood by familiarity with the plants in the field and some recognition of biogeographical factors and the role these may play in driving change, adaptation and evolution.
In Update Vol.3 I illustrate examples from many of the populations known to me, and also draw attention to populations in the lower Breede River valley which are significant. Chapter 13 is entitled “Haworthia is confusing” and Kobus Venter kindly used this material to make a presentation to the Succulent Congress at Calitzdorp in 2006. In addition I sent a draft of this recent manuscript to Bob Kent who replied that he was not sure what I meant by agreement. My missive to Bob included two collections made subsequent to Kobus’ presentation and that are a harsh reality check. These need to be added to the material touched on by Kobus. The plants I illustrate come from only a few populations that I know of and I believe that the only logical conclusion that one can draw from that greater body of known populations is that there is one super species, H. mirabilis – among others. My view now is that we need to take ALL the populations known at the very least to me and try to build a rational, consistent and coherent classification which might fit an imagined model of a product for botanical science. Therefore what I mean by “agreement” is virtually that readers have to submit to the uncomfortable situation that I may be right and that there is no alternative but to go along with what I have concluded. The reservation is that new material may necessitate modification and my opinion is that it will probably drive the classification towards a still more conservative position.
My most recent excursion has in fact been to try and resolve the dramatic juxtaposition of variants that we lower down the Breede River. In summation of these, we have Adam Harrower’s collection (Fig.1) from Sandhoogte and Chris Burger’s collection, and (Fig. 2) at Buffelsfontein. These are both south of the Potberg.
Fig. 1. MBB7251 H. mirabilis. Sandhoogte, De Hoop.Fig. 2. MBB7374 H. mirabilis. Buffelsfontein, De Hoop.
Then there are my collections from Stoffelsriver (Fig. 3), one nearer to Infanta (Fig. 4) and then two from Kleinberg (Figs 5 & 6). A curious addition that I have yet to see in the field, is a collection by Ismael Ebrahim (Fig. 7) of SA Biodiversity Institute from southwest of Vermaaklikheid. That collection resembles my Stoffelsriver collection and so provides the inevitable link to paradoxa.
Fig. 3 MBB7520.1 H. mirabilis. Stoffelsrivier.Fig. 4. MBB7248 H. mirabilis. Lower Breede River.Fig. 5. MBB7496.1 H. mirabilis (alias maraisii cf atrofusca. ESE Kleinberg, Matjieskloof.Fig. 6. MBB7516.1 H. mirabilis alias maraisii. Kleinberg.Fig. 7. Ismael Ebrahim sn. H. mirabilis alias paradoxa. Koensrust
At two localities near Melkhoutrivier (Figs 8 & 9), between Stoffelsrivier and the Infanta collection, we found plants which have to be related to those mentioned above. The plants are highly variable as I have now come to expect, and individuals can be likened to badia, acuminata and mutica. The surfaces have a curious sheen and may be incipiently spiny as the Sandhoogte and Buffelsfontein plants also are. The paradox is that we are also driven to the conclusion that the very small plants at Ziekenhuis (Fig. 10) are the same species. Collections, figures, 1 to 10 are a set within an incontestable biogeographic zone where geology is probably the most notable primary variable.
Fig. 8. MBB7608.1 H. mirabilis. Melkhoutrivier W.Fig. 9. MBB7609.1 H. mirabilis. Melkhoutrivier E.Fig. 10. MBB7615. H.mirabilis alias maraisii. Aalwee, Diepkloof.
What re-enforces this seemingly improbable juxtapositioning of such different things, apart from my lengthy dissertation on the matter in Update 3, is a second find we made southeast of the Bromberg (Figs 11 & 12). This is near the locality for what I suggest is H. rossouwii var elizeae, and also for several variants of what have in the past, been classified as maraisii and mirabilis or variants thereof. My new find is a remarkable array of large very dark-green (nearly black), plants which dramatically enforce the close association between mirabilis and maraisi as well as with heidelbergensis. These plants include the elements of both “species” as they might have been understood. What is striking is again the huge variation in the population that I have come to expect in this complex. The plants do lack the opaque dots on the under-leaf surfaces while the upper surfaces may be clearly windowed or opaque. They bring forcibly to my mind a sequence of populations from Heidelberg in the east to Verdwaalskloof near Riviersonderend in the west that I suggested forged the interface of mirabilis and maraisii. Coupled with this is the array of populations both north and south which enforce that continuity and that also lead on to heidelbergensis. So while collection 11 & 12 … may fall outside the compass of a Lower Breede biogeographic zone, it is unlikely that evidence can be found to suggest that it is not in a continuum of many other populations that link it with that zone. We are thus dealing with elements which are grossly different in imagery that constitute one system that can be said to be a “species”. (Note:- the pictures are of single plants and they convey neither the respective sizes of the plants, nor the gross variability within those populations. It is nevertheless true that generally one could probably assign a given specimen to each population – if required to do so).
Fig. 11. MBB7612.3 H. mirabilis. Diamant.Fig. 12. 7627.1 H. mirabilis. Diamant E.
My recommendations are:
1. deficiencies in respect of a species definition be admitted and rectified.
2. the nomenclatural code is summarily assigned a secondary role to a dispensation which is more flexibly attuned to the realities of a truly asymmetric species structure with more emphasis on reflecting field relationships, and to the competence of its users.
3. the illusion of reality that the ranks of genus, subspecies and variety provide should be admitted. Genera should be recognised for their historical and artificial value, but for species I would suggest that much more attention be given to the huge asymmetric and asynchronous variability that underlies capacity to change and adapt.
With the adoption of, say, H. mirabilis as a super species, variants can be indicated by the addition of any other epithet in inverted commas; thus H. mirabilis ”maraisii”, H. mirabilis “heidelbergensis”, H. mirabilis “Melkhoutrivier” in the knowledge that there is in fact no clear and infallible distinction, or pretension that minor ranks have any reality either.
In completing this manuscript and re-reading it, I could not help but keep referring to the book by Felipe Fernadez-Armesto entitled “Truth”. The cover blurb includes the words… ”We need a history of truth – though until now no-one has tried to write one.” There is also a note by Robert Winder which reads … ”A sharp and interesting work bound to enrage specialists in the fields he sprints through.” My own summation is that I have tried to write a truthful account of Haworthia. It will surely also enrage others who try to do the same, as well as others who may perceive the truth in some other fashion. To them I truly apologize. I do not have the answers and find my own writing quite as pompous and irritating as my readers may too. It does appear to me that we have as a society ”… lost faith in the reality of it and lost interest in our search for truth.” ♦
During the time I have worked with plants, I have met many botanists and taxonomists and I particularly had the opportunity to associate closely with one of the most prominent in succulent plant taxonomy. I could never hope to emulate the energy, application, thoroughness and zeal with which that person approached the subject, nor the academic and written achievements. The sharing of ideas was however, a problem and I never felt much more than student. My discomfort with the taxonomic product of this persons work eventually resulted in alienation and eventually I wrote in frustration… ”Taxonomy as a science has to answer the question “Are species real?” starting and ending with proper definition of the word/concept.”
The reply I received was this… ”Yes, species are real, and defined well by their ability to cross freely and produce offspring which again crosses freely. This has been studied and demonstrated since 1750 or so (you may remember that I talked about this in one of our discussions on the subject), but it is naturally not easy to dive into so much work including following up several generations etc. And since related individuals are similar looking, the reciprocal conclusions that similar objects are related has unfortunately been used as being true, which it is not. And yet, species are the only natural unit in the whole of taxonomy, which to determine is the crucial point.
Perhaps this sounds rather like the famous Dicta of Bessey, but after having dealt extensively over 38 years by now with species boundaries in teaching, reading, theory and practice, in different vegetation zones and many different groups, I come back to old definition first given by Ray in 1682:
“Group of plants derived from common seeds, reproducing their typical features by sowing.”
and used by most taxonomists since.
The point I have been trying to make in writing about Haworthia is that consciously or unconsciously, this definition describes the underlying perceptions of most persons. The very problem is the use of this simplistic definition and the fact that it does not work. I do not think I have ever been able to argue this adequately and this response from this paragon of academic botany leads me to this response. The Ray definition of the species flies in the very face of Darwin’s concept of their evolution and adaptation. If species did in fact breed true, there would be no adaptation and no evolution. The nature of creation is change and living things have the capacity in terms of inherent variability to meet this constant change whether it is by slow degree or by cataclysmic event. My contention is that “most taxonomists” have indeed thus been using a faulty definition and wholly underestimate the degree of variability in plants. Their contention may be that they are only providing an approximation of the truth, but this is not the effect achieved nor is it the impression I obtained from long association with this particular taxonomist and others. In Haworthia particularly, variation is pronounced. In some populations where field examination may suggest the plants are all very similar, when grown from seed no two individuals look alike. Vegetative propagation may have contributed to an illusion that plants breed true.
My own definition is that species are a dynamic and fractal group, or groups, of living or past living organisms which are morphologically, genetically and behaviorally continuous in space and time. Quite obviously the discontinuities are not going to be any easier to determine whether one uses Ray’s definition or mine. But what mine does is that it covers the reality that species are spread in geographic space and they have both the variability associated with the range of habitats they occupy as well as the inherent variability which provides them with the flexibility to respond to changes in habitat. Few people have the necessary experience in the field with enough living systems, and with cultivation, to truly encounter the phenomenal variability which underlies capacity to adapt and change. ♦
Recently I have been in communication with three recognized botanists and have their written admissions that…
1. “Taxonomy is in a mess.” 2. That the question of species is “highly controversial”. 3. “The current framework for decisions (for taxonomic decision making) is riddled with flaws, but it is the only one we have. Someone who knows the plants has to make decisions.”
I do presume to have some knowledge of the plants and hence I made a decision to submit a list of names which I think could serve the need of a botanical reality. However, the very botanist who had suggested who should make the decisions then commented … ”It is interesting that all of the ‘new’ discoveries (e.g. H. cummingii to name one) must be forced into the existing classification”. I asked if there was any evidence that force was required to do this. There was no reply.
My conclusion has to be that the botanical nomenclatural and classification system is flawed and that there is actually no way in which Haworthia can be satisfactorily forced into that system. Therefore there is no further contribution that I can make. There is still a huge amount of fieldwork that could and should still be done, but I cannot see that any new records or observations can significantly improve any classification that is connected to the traditional systems and escape the controversy invariably generated when more than one taxonomist becomes involved.
It is now evident in all this that human sensitivities are of far greater consequence than sensibilities. While a classification may be an apparently intellectual and truth-finding process, it may be nothing more than an easily accessible arena where minds can create an illusion of being so occupied. I am deeply sorry that I have thus offended and hurt people by my own activities there. ♦
This note is not strictly after closure because Cameron MacMaster (Cameron knows the plants, especially the bulbs, of the E Cape intimately and was instrumental in the re-location of H. marumiana many years ago.) sent me a picture (Fig.1) of a Haworthia from Glen Avon Falls east of Somerset East some time ago and this has been a lure to me ever since I saw vdW287(PRE). It should be noted that this specimen is cited, I must note a sentiment of considerable reservation which was not conveyed by the rigidity of print, in Haworthia Revisited (p.67) under H. decipiens var. minor… “3225 (Somerset East): in valley behind Bosberg (-DA), van der Westhuizen 287 (PRE).” I have visited the Bosberg in a weak attempt to locate such a plant after a fruitless attempt to determine who and where the collector was and is. The area is intimidating in its vastness as are so many of the hills and mountains of the Cape and with so much still to explore, this area has not been a priority. In fact I have just recognized that while I wrote Revisited in response to pressure, my subsequent exploration has been to seek validation for my own comfort rather than to try and impress anyone. This recent visit to the Bosberg is only because an odd opportunity arose for me to revisit my friends (Ian and Sandi Ritchie) on Kaboega, coupled with interest from a distant botanist acquaintance in Prof. Richard Cowling. Prof. Cowling is one of those rare botanists from whom I have really learned something to think about rather than just to remember. I had contacted him because in my correspondence with Jan Vlok about the vegetation of the Mossel Bay area, he had copied responses to Prof. Cowling. The outcome was that I was introduced to Dr. Syd Ramdhani who is now contracted under Cowling to study the biogeography of Bulbine as a post-doctoral task. Dr. Ramdhani studied Kniphofia and works in the molecular-biology laboratory of Rhodes University managed by Dr. Nigel Barker. Dr. Ramdhani is now also tasked and occupied with a feasibility study of Haworthia as a target group for extended biogeographical research where H. cooperi has been suggested by me as a possible fruitful area of interest. (These botanists have been warned not to be influenced by Bayer!) So I have been aware that the MacMaster plant could signify a replicate of the Kaboega/Helspoort/Plutos Vale/Baviaanskloof/ complexes which suggest that H. cooperi and H. cymbiformis may be one species. My visit to Glen Avon Falls was then added to the familiarization of Dr. Ramdhani with Haworthia on Kaboega.
With the kind assistance of Bill and Allison Brown of Glen Avon farm and then Bill Playdon of Glen Craig, we gained access to the waterfall which is now a National Heritage site. The road cannot be described as suitable for anything but a rugged 4X4 but we took it on in our less suited vehicle to save time (cost = 1 tyre!). We found just 5 plants and photographed three (figs. 2, 3 & 4). The terrain is really rugged and I have no doubt that the plants will occur in greater abundance nearby and also in the greater area of the Bosberg – and that such populations will follow the same pattern now so familiar to me and so seemingly strange to so many haworthiophile writers of my acquaintance. The plants call to mind an article that I wrote in conjunction with John Pilbeam (Cact. Succ. J (US) 46:166, 1974) where we ironically considered H. obtusa to be a variant of H. cymbiformis rather than of H. cooperi. The latter species is everywhere about the Bosberg while H. cymbiformis is to my knowledge far further to the south and east at Alicedale.
Returning to Kaboega, we took a brief look at some of the populations recorded and discussed in Haworthia Update Vol.1. Dr. Ramdhani left us there and we continued our holiday. We had been intrigued by Ian Ritchie’s discovery of a second population of H. sordida (figs. 5, 6 & 7) and while exploring that took a different route back to the vehicle. On the way we found another population of H. cooperi cf cymbiformis (I will be accused of inconsistency here because I have referred to these variants as graciloid, cf viridis, cymbiformoid if not also by some other name. Figs, 8, 9 & 10). I am not going to try and discuss or describe these plants any further than already done in the literature and because the following considerations come to mind.
A botanist acquaintance is most scornful of my opinions and my solutions for Haworthia and wrote to me… ”I believe taxonomy is an explicit, precise discipline governed by certain rules, if they are obeyed, things will fall into place. If they are ignored, you land up in a mess.” This botanist who might well have been raised on Enid Blyton and the famous five, or Bryce Courtenay and Aloe brevifolia in the Barberton hills from Power of One or the BIG tree in The Four Fires, when it comes to secrecy and drama, field botany and field familiarity in particular. The ICBN is no doubt hot-stuff where nomenclature is concerned but pretty damned useless when it comes to what these are used for. If things are going to fall into place it will be because we all make a more concerted effort towards mutual understanding and common purpose.
Secrecy in respect of localities I do sympathize with. The question is if this serves any purpose. My opinion is that, quite apart from the fact that secrecy has no place in science, it does not have any merit. Where secrecy is practiced there can be no organized skepticism and no shared reality or truth. I see it used to obfuscate and obstruct the healthy organization of skepticism by others and possible alternate solutions. There may be a down-side in that unscrupulous individuals may decimate populations or remove valuable genetic material and diversity from the field. The reality is that this is happening all the time at far greater scale than that of a few collectors. If it were not for collectors our knowledge base would have been infinitely smaller and who knows what the negative impact of the dwindling numbers of taxonomists is going to be. My negative view of conservation is that it is a sentiment with its origins in collectors and that it is now functioning to discourage the very interest that gave birth to it.
On our travel home from Kaboega, I kept looking towards the hills and mountains that accompanied us all the way on a 12 hour drive to complete about 1200kilometers. All of those offer suitable habitat and perhaps 2% or a great deal less of the area has been explored with an eye for Haworthia. This brings me finally to a sort of postscript. Russel Scott writes of his encounter with Poellnitzia rubriflora in the Robertson District. What is odd and welcome too no doubt, is that he does not mention the fact that officially the name is Astrolobarubriflora. Over a 6-hour hike he has to suggest that the plants were relatively rare – where he walked of course. His hike could have covered more than 20 miles and perhaps a 20yard wide band – calculated at a total search area of about 2.2% of one sq. mile! It is quite common south of the Breede River between Eilandia and just east of Bonnievale and as Russel mentions seeing H. herbacea I conclude that he was thus within the 600sq mile area I would say is its range. Thus even a 6-hour hike along what must be conceded was a narrow band, has not provided a lot of insight into quite a large area. Very little indeed as to what all there is to be seen. Yet I do not think it is possible to be looking specifically in species and succulent-rich habitat in the said search area without seeing H. mirabilis. Russel has exposed a reality that for someone to really become familiar with any plant group in the field is a dedicated life-time. Only a select few South Africans seem to have achieved that and then only in respect of particular interests. In my life-time of plant interest and field experience, I think I am personally only beginning to have an idea of what it means to really know what field familiarity can be. ♦
Fig.1. Haworthia sp. Glen Avon Falls, Somerset East 22Jan06 101Fig.2a MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.2b MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.2c MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.3a MBB7634.2 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.3b MBB7634.2 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.4a MBB7634.3 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.4b MBB7634.3 H.cooperi cf cymbiformis. Glen Avon Falls.Fig.5 MBB7635 H. sordida. Swartkop, Kaboega.Fig.6 MBB7635 H. sordida. Swartkop, Kaboega.Fig.7 MBB7635 H. sordida. Swartkop, Kaboega.Fig.10 MBB7636 H. cooperi cf cymbiformis. N Swartkop, Kaboega.
There are few succulent collections which do not include haworthias, although these small and insignificantly flowered plants are not good garden subjects. Their size, and shade and shelter requirements make them better suited to intensive cultivation in raised containers under shelter. Very popular with collectors especially prior to World War II, the decline in popularity can be attributed to various factors. Although the genus is credited with some 160 species and more than 250 varieties, it is highly unlikely that more than 90 species and perhaps 10 sub-species would survive a critical revision. Many species have been eliminated in recent years, but there are still many maintained only by the mystery of their origin. Hybrids and ill-defined or inadequate species account for many superfluous names. The result is an artificial pseudo-scientific system of nomenclature in which the classic binomial system is prostituted for a series of horticultural cultivars.
Within Haworthia there are real problems of definition and circumscription of the species. The variability within species is often so great that it is very difficult to circumscribe a species in such a way as to exclude members of other species. The species are best recognised as geographical entities and no species can be described without good reference to locality and distribution. This is the only way in which a sound system of nomenclature directly related to field complexes, and hence a “natural system”, can be derived. Names are the fundamental basis of communication concerning the plants, and the psychology of collecting requires good definition of the “kinds” of things being collected. Nevertheless it is surprising how many persons enthusiastically and vehemently argue the “differentness” of things without seriously considering where the boundaries of difference really lie. The system of nomenclature in Haworthia has been so confused that it has not been possible for collectors to name or acquire plants consistently or confidently.
What are Haworthias? The genus Haworthia is currently experiencing another stormy taxonomic passage and for the present includes the genus Chortolirion Berger. C.A. Parr in the African Succulent Plant Society Bulletin (6: 145, 1971) also included the genera Astroloba Uitew. and Poellnitzia Uitew. However, the writer refuted this in the National Cactus and Succulent Journal 27: 77 (1972). This refutation was not considered by Mrs. A.A. Obermeyer-Mauve in her article in Bothalia (11: 119, 1973) where she tacitly supports Parr’s arrangement.
The genus Astroloba comprises seven species, all of which have leaves arranged in five conspicuous rows. The genus was revised by Mrs. P. Roberts-Reinecke1 in an unpublished thesis submitted for the degree of Master of Science to the Department of Botany at the University of Cape Town. This comprehensive work makes a mockery of the recognised species and demonstrates the remarkable confusion that can arise in taxonomic nomenclature. It is Mrs. Roberts-Reinecke who concludes that there are only seven species with three subspecies, and she also describes the intergeneric hybrid Astroworthia bicarinata (Haw.) Roberts-Reinecke – the hybrid Astroloba rugosa Roberts-Reinecke (nom. nud.) X Haworthia margaritifera (L.) Duval (the latter now H. pumila). Apart from floral characters, there are no caulescent haworthias with five well-defined leaf parastichies and only Poellinitzia rubriflora (Bolus) Uitew. is strictly comparable. This monotypic genus has a unique gasteria-like flower and to lump it either with Haworthia or Astroloba is clearly incorrect. The more recent inclusion of Chortolirion, also monotypic, under Haworthia by Mrs. Mauve is also very questionable. Certainly her argument in which she presents H. graminifolia G.G. Smith as being bulbous and thus comparable with Chortolirion is erroneous. H. graminifolia has no semblance of a bulb or fleshy leaf base, although the broad leaf bases do completely enclose the stem. The real error lies in the unstated contention by both Parr and Mauve, that Haworthia is an integral undifferentiated unit. This is fallacious, as Haworthia is clearly divisible into at least three very well demarcated subgenera (Bayer, 1970).
The first of these is the subgenus Robustipedunculares Uitewaal ex Bayer. When Uitewaal proposed this name he was suggesting it as a subdivision of two sections of the genus. Proper analysis referring to valid natural species shows that there are five species sharing common characteristics.
These are: H. minima Haw. H. marginata (Lam.) Steam. H. margaritif era (L.) Haw. (H. pumila (L)Scott) H. kingiana Poelln. H. poellnitziana Uitew. (now H. minima var. poellnitziana)
They are all acaulescent with robust branching inflorescences; unmistakably similar perianth tubes and arrangements of penanth lobes: rounded seed capsules and black winged seeds. They all occur in the southwestern Cape with H. margaritifera extending into the Little Karoo and H. kingiana occuring in the Mossel Bay area. These are essentially winter growing species which flower in mid-summer. There is some degree of presumed introgression in the case of H. margaritifera and H. marginata in the Ashton area and apparent hybridisation of H. marginata with H. minima around Heidelberg (Cape).
The second subgenus Hexangulares Uitew. ex Bayer was also defined. It comprises both caulescent and acaulescent species, and the flowers are exasperatingly identical throughout. The perianth lobes are practically always similarly presented, but small differences can be detected (also in the case of the perianth tube) in some instances. The peduncle is usually unbranched and the perianth tube is comparatively straight with the veins of the three inner petals positively separating the margins of the outer tepals. There are not more than twenty species in this group and the distribution is quite extensve. H. limifolia Marloth and H. koelmaniorum Oberm. & Hardy occur in the B. Transvaal, Swaziland and N. Natal. The former species is variable so that distinctive geographic types can be recognised. Thus H. koelmaniorum may still best be recognised as a local element within, certainly having no affinity with H.. tuberculata as suggested by the authors of the species. H. tuberculata v. Poelln. is also not allied with the so-called Margaritiferae. It occurs in the Little Karoo, has the same slender branched inflorescence and narrow slender perianth tube with straight upper tepals as H. starkiana v. Poelln. The latter species is fairly localised in the low mountains just north of Oudtshoorn and the hybrid with H. tuberculata probably accounts for “H. smitii” v. Poelln.!
H. tessellata Haw. is very widespread, occurring from south of South West Africa, through the southern Orange Free State to Jamestown (at 6 000 ft.!) and down to the Little Karoo. H. venosa (Lam.) Haw. is probably a synonym of H. recurva Haw., which occurs in the Breede River valley near Swellendam. It is supposedly distinguished mainly by its longer bigger leaves, but some very small forms occur at Napkysmond to the south, while large forms of H. tessellata occur around Oviston and Graaf Reinet. In the Ceres Karoo one finds caulescent subscabrous populations which are very close to the distinctive H. granulata Marl.
H. viscosa (L.) Haw. is a strongly trifarious species occurring widely in the Great and Little Karoo as well as the southern Cape. It varies quite considerably at different localities, and species such as H. beanie G.G. Smith, H. cordifolia Haw. and H. asperiuscula Haw. are probably all superfluous. H. nigra (Haw.) Baker (or H. aspera Haw. if Mrs. Reinecke is correct) is widely spread in the Eastern Cape and Great Karoo. It is a smallish caulescent dark tubercled species which has been recorded in different forms from as far to the northwest as Loeriesfontein and down to Grahamstown in the southeast. H. sordida (Roem. & Schult.) Haw. is a comparatively rare species found from Uitenhage westward to Steytlerville — a very slow growing dark species with the same slender 2-3 branched inflorescence as H. starkiana and H. tuberculata. H. glauca Baker is best known in the glabrous form from the top of the Zuurberg north of Addo, but is undoubtedly synonomous with the slightly tubercled straight leaved “species”, “H. herrei v. Poelln”., “H. jacobseniana v. PoelIn.”, “H. jonesiae v. Poelln.”, and “H. eilvae v. Poelln”., from the area Uitenhage to Willowmore.
H. reinwardtii (Saim-Dyck) Haw. and H. coarctata Haw. are the subject of a recent paper in the National Cactus and Succulent Journal, and many species and varieties can be excised with rational observation of these two species in the field. H. reinwardtii occurs from East London to the Fish River and just beyond, while H. coarctata is found westward to Paterson near Port Elizabeth. These two species are caulescent and are otherwise very close to the acaulescent H. attentuata Haw. and H. fasciata (Willd.) Haw. H. attenuata is best recognised in the area east of Grahamstown and H. fasciata in the fynbos areas around Port Elizabeth and Humansdorp. However, H. attenuata also occurs with the very distinctive finely tuberculate H. radula (Jacq.) Haw. in the lower valley areas of Hankey/Patensie. H. armstrongii v. Poelln. and H. browniana v. Poelln. both consist of small local populations lying very close to each other north of Uitenhage. It is possible that they may have arisen by odd hybridisation of H. glauca and H. fasciata and subsequent vegetative propagation. This is of course highly speculative because no evidence has yet been presented to indicate that hybridisation has led to speciation in Haworthia.
There is no doubt about H. longiana v. Poelln., it has long hard leaves and the offshoots at the stem emerge as strong branchlike out- growths, rather than as softer obviously axillary offsets as in near relatives. H. woolleyi v. Poelln, is most easily allied to H. tessellata although it has the proliferous clumping habit of H. attenuata.
The third subgenus Haworthia is perhaps the most complex of all. These are all acaulescent species (despite “H. ramosa G.G. Smith”) where the peduncle is simple and the outer tepals of the floret have touching margins. The tube is curved and the style upcurved – there is no adhesion between the inner and outer tepals. There are several very decided species with characteristics which evidence a distant relation. H. wittebergensis W.F. Barker with its papery leaves, growing in the mountains south of Laingsburg, cannot really be allied with H. angustifolia of the section Loratae. H. graminifolia G.G. Smith from Schoemanspoort near Oudtshoorn and H. blackburniae W. F. Barker from Van Wyksdorp to west of Ladismith both have fusiform roots and long linear narrow leaves. H. maughanii Poelln. and H. truncata Schonl. have truncated leaves (to use the vernacular, these are the “perdetande”) and truncation occurs in the second leaf of the seedling. This is quite unlike the recurvature of the leaves of the section Retusae, although the functional end purpose appears to be the same. The involved taxonomic confusion presently negates a really rational discussion of the remaining species. The “Retusae” is a section comprising a vast series of intergrading populations extending from Steytlerville to Bredasdorp in the southwestern Cape. This series includes many recognisably distinct taxa such as H. springbokvlakensis C.L. Scott, H. comptoniana G.G. Smith, H. parksiana v. Poelln., H. mutica Haw. and H. mirabilis Haw. etc. where the leaves are strongly reflexed to give an apparently truncated end area. However, several segregates in the same series have normal suberect or incurved leaves e.g. H. reticulata Haw., H. pubescens Bayer. In H. schuldtiana v. Poelln. some of the segregates have “retused” leaves and others lanceolate incurved leaves. The section as it stands at present is an artificial one as some of the principal elements of the series are incorporated in sections such as the Denticulatae Berger and Muticae Berger.
Without good anatomical characters it is virtually a question of guesswork to try and prepare a reasonable diagnosis of the other sections and this may be true of the whole tribe Aloineae. Chortolirion can be described as having a haworthia-like flower if one very crudely regards the flowers of the haworthias as all alike. Actually Chortolinon has the straight tube, tepal arrangement and straight style of the Hexangulares. Whereas there is some difficulty in separating species of that group on floral characters, Chortolirion stands out like a sore thumb. The florets are practically sessile with wide pedicellate attachment while the peduncle is relatively short, fleshy in comparison with Hexangulares and notably glaucescent. Vegetatively Chortolirion has a distinct bulb, the leaves tend to spiral, they have the white markings characteristic of the Leptaloe and the plants are deciduous as is Aloe kniphofioides – the die-back of the leaves is not directly due to fire or frost as suggested by Mrs. Mauve, but a seasonal phenomenon. H. graminifolia in the subgenus Haworthia does not have the semblance of a bulb and it would take more than “eliminating gravity” to get the floret to resemble that of Chortolirion. Mrs. Mauve is also responsible for the inclusion of Chamaealoeafricana in Aloe as A. bowiea Schult. & J.H. Schult. While in essence her treatment may be correct, the facts mobilised in support do not substantiate the change. No one can claim that the generic states in the Aloineae are or were factually correct, but there seems to be little point in merely guessing at a better construction. What is needed is a thorough anatomical study of the entire tribe to include all the genera and aberrant species. Any other basis for creating and recreating categories is unsatisfactory.
The cultivation of haworthia The success of any plant in general cultivation is more certainly a product of inherent adaptability and vegetative or seed fecundity, than dependant on any skills of the gardener. The principal basis of speciation in plants is undoubtedly geographic distribution and habitat selection, so that there are many kinds adapted to specific microhabitats. Any collector attempting to grow all the species of any respectably sized genus must face either fact or disappointment. To be successful the collector must provide “intensive care” facilities for the inevitable difficult plants. Haworthias are no exception to the rule. The gaps in comprehensive collections are not only due to a taxonomic situation loaded with “species” that are not, but also due to species which require luck or skill to cultivate successfully. Of all the variables which affect success in cultivation, soil is perhaps the most complex. Variability in soil seems to play a major role in determining the composition and distribution of plant communities. Haworthias are largely associated with rocky situations with undeveloped soils and it is surprising that most can be grown in the standard soil mix consisting of one part loam: one part sand: one part compost. J.R. Brown, the American doyen, concluded that Haworthias could be grown in almost every type of fairly porous well drained soil. This is mostly true but attention must be paid to shade and watering. Practically all the species occur naturally in shaded conditions and require semi-shade in cultivation. Mature plants are often found in exposed positions in the wild, but these are invariably highly coloured, stunted, withdrawn into the soil, or the leaf tips are necrotic. The best colours and forms are of course obtained with the least possible shade and attenuation or yellow chlorosis of the leaves should not be tolerated.
Watering is a very critical factor and it must be remembered that there are species from the southwestern Cape adapted to winter rain. Conversely there are species averse to winter wetness. There is no fixed principle that winter growers flower to set seed for the start of a new growing season, so flowering is not a good indication of water requirement. H. herbacea and H. reticulata flower in early spring, H. margaritifera in mid-summer, and H. schuldtiana in early winter, yet they are all winter growers. The winter rainfall area is really demarcated by a line drawn from Bredasdorp to the mouth of the Orange River. Rainfall immediately east of this line comes mainly with the autumn and spring solstices, but growth is mainly in winter as a result of adaptation to the intense summer heat and low humidity. East of Ladismith the species appear to be mostly summer growing. In practice the winter growers require a dry spell in late summer. There are some species which have an indeterminate growth cycle, e.g. H. setata Haw. and H. venteri v. Poelln., and injudicious watering can quickly lead to disaster. If in doubt it is best to water when the plants show indications of stress, always bearing in mind that shade may be more desired than water. With some of the really slow growers such as H. sordida and H. woolleyi the temptation to water may be an invitation to the compost heap. They cannot be pushed into vigorous growth and increased shade is again more likely to produce the required results than water.
The summer rainfall soft-leaved Haworthias tend to lose all their roots with winter watering but it is not wise to generalise. H. pilifera cannot be put in the same class as the cliff-hanging (literally) H. cymbiformis–planifolia types. These types are very proliferous and want particularly good drainage below, and a dry winter. H. pilifera will tolerate poor drainage if its water comes sparingly and at the right time. The best specimens are grown in quite intense light conditions forcing retraction into the soil as is their wont in nature. In some variants the leaves may completely truncate with a network of necrotic veins to screen out the sun. In this respect many haworthias resemble the windowed “mesembs” such as Fenestraria, Lithops etc. which reduce surface area (and transpiration) and make provision through surface translucency, for photosynthesis by sunlight in the deeper buried chlorophyll-bearing tissue. Retraction into the soil is a characteristic of the thick-rooted species such as H. truncata and the “retusae”. It is thought to be brought about by contraction of the roots pulling the plant down into the soil. If plants are excavated and allowed to dry out, the roots flex very strongly upwards which would also have the effect of forcing the stem down. However retraction occurs, it is puzzling how the soil is displaced underneath the plant, because H. truncata, among others, may be found in the stoniest of ground. Retraction should thus not be regarded only as a movement per se of the plant downwards, but a state into which the plant grows from the time of germination. If retracted plants are transplanted and well submerged in the soil, the chances of losing them from overwatering are increased. Overwatering can be defined as a combination of giving too much water too often, poor drainage, poor ventilation and incorrect nutrition.
The Hexangulares species such as H. attenuata, H. reinwardtii and H. coarctata are the commonest encountered in gardens. This is because they are very prolific vegetatively, fairly fast growers, and very much more tolerant of exposure to sun. Members of this group seldom have surface translucency and are generally not retracted. H. tessellata grows in very very hot situations but usually in the protection of rock crevices and under stones. H. limifolia is also found where it can get very hot, but survives under the protection of bush or grass canopy. A common species in gardens is a highly fasciated form of H. attenuata which is usually taken to be H. fasciata. The latter species has less attenuate leaves and the upper leaf surfaces are smooth. H. fasciata is not a common species in gardens and this can only be accounted for by soil requirement. The species is found in the acid soils of the Port Elizabeth/Uitenhage area where it grows in the “fynbos” vegetation and exemplifies the problem of suitable soil for cultivation.
Several species are found in the Robertson Karoo in undeveloped soils of the Malmesbury shales. The vegetation is highly differentiated into distinct plant communities over very small distances. Soil pH may vary from 3,9 to 7,8 over a distance of ten paces. However soil pH is not an only consideration. H. margaritifera is found usually in Pteronia paniculata (the composite “ribbokbos”) communities where pH varies between 3,9 and 4,9. It also occurs in Ruschiacarolii (Aizoaceae – vygie or mesem) communities where low pH (4,8) is also recorded and where H. herbacea may be plentiful. It is not unusual for the situation to be reversed with H. herbacea in the Pteronia communities and H. margaritifera in the Ruschia. Yet it is clear that each species has very specific requirements and does not ever appear to co-habit in a single locality. It is often the case that three or four species of Haworthia grow in the same general locality, but clearly confined to their own microhabitats within a stone’s throw of one another. Neither H. margaritifera nor H. herbacea is successful in general cultivation and the reason appears to be a combination of their winter rainfall requirement and their soil needs. H. marginata is similarly rarely successful in cultivation and also (or co-relatedly) rare in habitat.
Propagation of haworthias is more often than not by vegetative offset, as most of the easily cultivated species are also very prolific. Pure seed is not generally available because of the tendency to hybridise, and the plants take three to four years to reach maturity from seed. Seed is easy to obtain by cross-pollination but the parent plants must be isolated from the insects and birds that normally pollinate them. The plants are self-sterile and so the parent plants must be from genetically distinct clones. Pollination is simply effected by inserting a fine hair into the floret of one plant and transferring the adhering pollen to the other. Pollen is released from the youngest flowers and the older flowers bear the receptive stigmas. Pollination is dependent on ideal weather conditions and warm sub- humid mornings are ideal. The seed capsules begin to form immediately after successful pollination and the fertile capsule may ripen in weeks. Failure of pollination. marked by the early abscission of the pedicels. Seed is best sown fresh, in early winter for the winter growers and any time in the summer for the summer growers.
A well-drained loose soil is used, with the seed very lightly covered with the same soil and lightly topped with a thin layer of small pebbles to hold the germinating seeds and tiny seedlings. Where damping- off is a problem, soil sterilisation may be practised or fungicides used. Transplanting follows crowding and must take place with due respect for the growing season and condition of the seedlings. Experience indicates that young plants should first be subject to some drying out and the young seedlings transplanted with intact roots, and then lightly watered. Older mature plants should also be well-rested before moving and if not, all the roots should be stripped off before replanting. Water should also be withheld for a few days after planting and the plants shaded until they start to grow again. Haworthias root best from the stem area closest to the lowest leaves and it is unwise to leave a big section of stem, or plant deeply with the expectation that roots will develop all along the submerged portion. Many species will propagate from leaves but this is a haphazard venture unless properly done. The leaves must be removed as closely as possible to the stem to ensure removal of adventitious growth tissue with the leaf. Gasterias can propagate from portions of leaf, but not Haworthia. Like aloes, non-dividing species can be induced to multiply by damage to the growing point the difficulty in this technique lies in inducing oneself to inflict the necessary damage on a treasured rarity. Excised leaves may be stuck shallowly into a sandy soil-mix and weighted down with small stones if need be. Propagation may also be effected by cutting entire plants into section and treating the cut surface with sulphur or some other fungicide. The writer cannot claim to having achieved any success with rooting hormone but it does seem to produce a good healing of broken surfaces.
Conclusion The published literature on Haworthia prompts one to misquote… “never has so much been written, about which so many know so little!”. Inadequate descriptions, erroneous naming and confused names seem to be the lot of succulent plant groups where the degree of amateur interest has greatly exceeded that of the botanist. E. J. Bullock made some apt remarks concerning the amateur in “Thoughts on the status of Aloe lutescens” in Excelsa No. 3. It is fortunate that there is no such thing as “closed shop” in taxonomic botany but it still behoves the prospective taxonomist to be aware of the levels at which information might now be available. The making of an herbarium specimen and concoction of a Latin description are very elementary barriers to the determined author of a new name. However, the development and maturation of a suitable species philosophy is another matter altogether. Since Darwin and perhaps before, there has been no complete accord on a definition of the “species” although almost everyone has a good idea of what it is. All can agree that the species is the basic unit of classification and it is also fairly easy to accept the classic definition of a species as a grout (or groups) of interbreeding or potentially interbreeding individuals. It is only in the last 30 years that the qualification has been added where all barriers to sexual reproduction are included. However, the difficulty really lies in the fact of change, so that any definition needs to consider variability in a species as it is exhibited in both space and time. There may be groups of plants which are currently not broken into clearly discontinuous groups which can be readily recognised as species. Often the case is that where some elements of a group form very clear subgroups and so species, other elements are highly variable. The former group suggests completeness of adaptation which has been said to be “cause of senescence of the issue”. In other words a well adapted and nonvariable species has probably lost the genetic capacity to change with changing environment and is so threatened. Alternatively, the highly variable “species” or aggregate of elements has the genetic resources to meet changes in the environment. In Haworthia the variable elements are often highly localised and it does not seem possible to evaluate these properly as species or otherwise.
G.W. Reynolds can be considered to have done remarkably good work with Aloe classification for the period in which he worked, and perhaps for much later too. However, no one can contest that problems do not still exist in that genus, and not only at the level of the species. The genus concept in the Aloineae is under pressure as witnessed by the changes implemented by Parr and Mauve. That this pressure has been characterised by a lack of valid new information on which a better grouping can be based, merely serves to emphasise the fact that sound arguments for the older situation never existed. ♦
Written for, and then printed and distributed by Haworthia Study Group, New South Wales – October 1970.
M. B. Bayer, Karoo Garden, Worcester, South Africa.
Much of the confusion in the taxonomy of the genus Haworthia could have been avoided if more attention had been paid to localities and inherent variability of the species. The tendency has been to base species on single specimens and to regard the species as variable concepts subject to personal opinions. According to Stebbins in his book Variation and Evolution in Plants, a species comprises a system of populations separated from each other by complete or sharp discontinuities in their variation patterns, and this must have a genetic basis. Therefore there must be isolating mechanisms to prevent transfer of genes. In Haworthia, most (not all) of the species are found in rock formations and stable situations at moderate altitudes, and hence the populations are well spaced. Haworthia are also insect pollinated, mostly by solitary bees, and as stated by Clausen in The Evolution of Plant Species, it is logical that spatial separation of the plant populations coupled with flight limitations of the pollen vectors, will with time have led to differentiation of highly localised populations. A further problem in Haworthia is that the species do in fact exist at different stages of differentiation. Thus one may be forced for practical reasons to regard a widely ranging series of varying populations as a single species, simply because the degree of intergradation and variability precludes any other alternative. If there is an alternative, it is to recognise varieties in the sense that this concept has been used in the past, i.e., to apply to individual variants and forms.
H. triebneriana, v. Poelln., is a good example, although not an exception, of what one may find. In some cases the variants may have a geographic basis, and it is clear from field observation that H. badia, v. Poelln., H. emelyae var. beukmannii, v. Poelln., and H. triebneriana var sublineata are such variants. The variety rubrodentata may also be, whereas the varieties pulchra, lanceolata and depauperata (all attributed to v. Poelln.) are from the same locality at Stormsvlei. According to the collector, Mr. G.L. Payne, von Poellnitz incorrectly cited the var. lanceolata from Robertson.
A second example is of the two species H. reticulata, Haw. and H. herbacea (M II.) Steam, which have been intensively studied over their distribution range. Here one finds three distinct stages of evolutionary differentiation. In one area one finds populations (not individuals) which are quite anomalous falling neither into one species nor the other. These appear to represent non-differentiated populations which may be remnants of the ancestral forms. In a second area one finds clearly differentiated populations of each species hybridising (introgression) to create an extensive hybrid swarm.
The third instance is within the one species H. reticulate, which varies quite widely over its range with each population intergrading into the next. This is a classical example where the species H. haageana, H. intermedia, H. hurlingii (all v. Poelln.) and H. guttata, Uitew., are really all forms drawn from various points in the range of the parent species, in this case H. reticulata (the correct locality for H. intermedia is stated by Mr. Payne to be at Buitenstekloof, which is within the range of H. reticulata). What has now apparently happened is that migration, probably due to water dispersal, has resulted in two clearly different forms growing en masse near one another. If one ignored the evidence of variability over the whole species range, one would be compelled to regard each as a distinct species.
With these thoughts in mind, we can now examine some variation in the species H. glauca described by Baker in 1880. The locality was given as the Kalahari Region, Orange Free State, which is quite improbable for the species. It is now generally accepted that this species is located in the Zuurberg mountains east of the Sundays River and north-east of Uitenhage. However, the Zuurbergs have western counterparts in the Kleinwinterhoek and Groot River Heights which all together form a chain of mountains running in an east-west direction separating Jansenville and Steytlerville, and extending from near Grahamstown in the east to Willowmore in the west. These form part of the drainage system of the Sundays River which extends northward to include Graaff Reinet.
In 1929 von Poellnitz described H. herrei with locality unknown, but in later publications cited Kendrew, Jansenville, Paardepoort (1937), Kleinwintersfontein and between Jansenville and Glenconnor (1938). The latter locality could well be Paardepoort too, as it is the Mountain pass through the Kleinwinterhoek mountains.
In 1932 von Poellnitz described the variety H. herrei var. depauperata and cited the localities Sundays River between Steytlerville and Port Elizabeth (1932), Paardepoort, Jansenville and Somerset East (1938). The first locality is vague and it is unfortunate that such citations detract so much from von Poellnitz’s work. In 1941 Resende recorded H. herrei var. poellnitzii, but it appears that this is intended to be the var. herrei as Jacobsen (1960) observes.
H. armstrongii and H. eilyae were both described by von Poellnitz in 1937. The former species is cited from Adelaide which is actually the town in which the original collector lived and from which the plants were sent (a common error frequently to be laid at Triebner’s door). From unpublished notes and records of Dr. Fourcade and Major Long, who together compiled a set of photographs and records of named plants, and from Mr. G. G. Smith’s collecting data, it appears certain that the actual locality was Spring Range, some 6 miles north of Uitenhage. H. eilyae was cited from Kleinpoort while the variety H. eilyae var. zantneriana, Resende, was described in 1943, locality unknown.
In 1941 Resende had also described H. carrisoi from the Zuurberg mountains, and the fate of this species is adequately dealt with by Smith (Views on the Naming of Haworthias, 1947) and by Resende (Variability and Taxonomy in the Genus Haworthia). H. carrissoi is undoubtedly a synonym for H. glauca. Two further species were described by von Poellnitz, H. jacobseniana from Miller which is nearer to Willowmore than to Jansenville, and H. jonesiae from Steytlervile (both 1937).
The localities cited for the above species and varieties, considering their similarity, alone tends to belie their validity. If it is accepted that a variety must also satisfy isolation requirements of some degree, in addition to consistent morphological variability, the situation calls for examination.
Evidence available consists of the photographs as published with the descriptions and also at other times, such as by Brown in the American Cactus and Succulent Journal, by van Woerden in Succulenta and by Resende in Succulenta Africana. There are also the records compiled by Fourcade and Long which are of particular value. Major Long was a one-time Director of the Port Elizabeth city parks department and a Haworthia enthusiast. His collection was compiled during the years 1930-40 and he was in close contact with von Poellnitz and with collectors such as Armstrong, Mrs. Emely Ferguson, Mrs. van der Byl, Mrs. Eily Archibald, Mrs. King and Mrs. Jones, to name but a few. In many cases plants photographed by Dr. Fourcade were of plants submitted by the original collectors after whom they were named, such as H. herrei, H. eilyae, H. armstrongii, and H. jonesiae. Only six sets of these photographs were compiled of which four can be accounted for, one is among Long’s papers, one in the Bolus Herbarium, Cape Town, and two among Smith’s papers in the Compton Herbarium, Kirstenbosch. Many photographs were taken by Smith of plants collected by himself or his collaborators and these include of:
H. eilyae – GGS2924 Kleinpoort, GGS3592 Campherspoort, GGS3 644 Campherspoort. H. glauca – GGS12 Zuurberg. H. herrei – GGS6863 7 miles Knoetze to Miller. H. herrei var. depauperata – GGS1184 Somerset East, GGS3658 3 miles Miller to Willowmore. H. jacobseniana – GGS3648 and 3655 Campherspoort, GGS2754 loc.? H. jonesiae – GGS3648a Campherspoort. Unidentified – GGS7048 and 7059 near Campherspoort.
The identifications quoted are those Smith and are not entirely in agreement with the types, but certainly as good as could have been expected under the circumstances. As will be seen inherent variability precludes either accurate identification or segregation of the various forms into the categories imposed by the described taxa. The plants collected by Smith and numbered GGS2754, 3648, 3655, 3644 and 2924 still survive in the collection now at the Karoo Garden. In addition there are his plants GGS7048-50 and 7059 from Campherspoort, 3660 7 miles Miller to Willowmore and 6917 from between Knoetze and Willowmore. Other collections numbered according to a section/species/collection system devised and in use at the Karoo Garden for the Haworthia include 3A/4/ I Zuurberg, 3A/4/2 no locality, 3A/5/1 18 miles NW Willowmore, 3A/5/4-5 Paardepoort, and 3A/6/8-1O Campherspoort. The Karoo Garden collections are multiple plant collections aimed at including the more common variants, and these are all in cultivation, under uniform environment. The collections 3A/6/8-10 are of particular note here as these were collected by the writer and photographs of these accompany this paper. The collections were made along a single transect following the road southward from Klipplaat through the Groot River Heights at Campherspoort and turning west toward Willowmore.
The first photograph (fig.1) is of the stems of four plants collected about 2 miles north of the poort at the locality for H. zantneriana, growing on a sandy plain together with Astroloba foliolosa. Here the plants were seldom more than 20 cm. in diameter or height. The second photograph (fig.2) is of the upper sections of stems of five plants collected within the poort where they were growing en masse, with clumps several feet in diameter with long trailing stems. Within the scraggly karoid scrub, some stems reach a length of 60 cm. and vary in width from 2-5 cm. The third photograph (fig.3) is of stems from plants collected about 3 miles west of the poort and on the north side of the Groot River. Here the plants were slightly more robust than in the case of the other collections but the long straggling stems were not nearly as marked as in the second collection. Colour, leaf tuberculation, plant growth characteristics, leaf-shape, -size and -presentation were all so variable that recognition of specific forms as varieties is impractical. In summation one must conclude that the various elements collected and recorded here, belong to a single species complex. There is no suggestion of geographic variability to suggest infra-specific grouping although it is admitted that for the area east of the Sundays River, evidence is sparse. This may also be true of the area north of Jansenville and also of that around Uitenhage. The writer has seen plants referable to this discussion from this latter area, and from experience in other species complexes is tempted to suggest that H. armstrongii may be a synthetic resulting from introgression of the H. glauca/herrei complex into the H. brownlana/fasciata var. variabilis complex of the Ultenhage area.
In conclusion it is suggested that H. glauca be retained as the species in the section Coarctatae found in the central and lateral drainage area of the Sundays River, possibly differentiated into two varieties H. glauca var. glauca in the Zuurberg mountains in the east, and H. glauca var. herrei in the Kleinwinterberg and Groot Heights in the west.
Further field work is required to substantiate such a proposal and also to elucidate relations of this complex with the Coarctatae of the Grahamstown and the Margaritiferae of the Port Elizabeth/Uitenhage areas. A considerable amount of evidence has accumulated to support the approach to Haworthia taxonomy adopted here and it is hoped that this will lead to a more satisfying classification within the the genus.
Acknowledgement: Acknowledgement is made to the National Botanic Gardens of South Africa where the writer is employed, and under whose aegis the work was therefore done. Gratitude is also expressed to Mr. F.J. Stayner, Curator of the Karoo Garden for unfailing assistance and encouragement. Thanks must also be expressed to Mr. C. Walker of Whaley Bridge, Stockport, whose kind interest prompted this paper.
(Comments by the N.S.W. Haworthia Study Group: This article, by Mr. M.B. Bayer, was received in October, 1970. The gratitude of the Group is extended to Mr. Bayer for his many letters and comments on Haworthia.) ♦
Printed in British Cactus and Succulent Journal 4:45 (1987).
Haworthia is indeed a popular genus which seems to inspire a great deal of controversy and confusion. One would have to be very thick-skinned to be able to ignore past history and not plead for forgiveness for similar transgression. I was just busy trying to clarify, in my own clouded mind, the problem of H. pumila (L.) Duval, when I saw Will Tjaden’s little article on the subject in this journal (3:88, 1985). Gordon Rowley in the same issue reviews the recent books on Haworthia and also mentions the H. pumila versus H. margaritifera debate. Coming so soon on the heels of Fearn versus Cole and Walker versus Bruyns, it would be insensible for Bayer to take up the cudgels against anyone.
In any case I frankly do not know what to do about the problem of the name-game so well expressed by both Rowley and Tjaden, and yet I shamefully have to admit my displeasure at their contribution, or lack of it. In the case of Tjaden, I agree with his comments on name changes and respect this view far more than he suggests. My distress at the recognition of H. pumila (L). Duval is greater than Tjaden has conceived, and all the more because I knew that Col. Scott’s solution offered in 1978 was not correct. Col. Scott was assisted in this instance by Dr L.E. Codd, who is one of our most respected taxonomists. Unfortunately they overlooked Burmann’s Flora Capensis of 1869 and also the fact that another species (H. minima) was involved. While I accepted their decision in the interests of stability and peace, Dr. Onno Wijnands pursued the matter a little more vigorously.
The intention of the Code of Botanical Nomenclature, to bring order and stability to the names of plants, seems fairly obvious. It also seems to me that the Code, being as complex as it is, is either an ass or can often be made an ass of, because its various articles can be used in contradiction to one another. For Mr. Tjaden’s benefit particularly, let me just quote, article 55.2 which is directly relevant to H. pumila… ”On transference of a specific epithet under another generic name, the resulting combination must be retained for the species to which the type of the basionym belongs and attributed to the author who first published it, even though it may have been erroneously applied to a different species.” One of my first experiences with the Code was in the presence of three distinguished taxonomists (one professor and two PhDs) trying to get guidance on the question of application of Linnaeus’s epithet pumila. The problem was so skillfully compounded and evaded that I felt that this simple mortal would have to stick to easier things. I really thought that Dr. Codd had helped solve a complex problem and I am as disconcerted as anyone else that Dr. Wijnands has established otherwise, arguing:—
1. H. pumila (Alt.) Duval is a homotypic synonym of H. herbacea (Mill.) Stear.
2. H. pumila (L.) Duval does not exist, and H. pumila (L.) … cannot be taken up because it would be a later homonym of H. pumila (Ait.) Duval.
3. H. margaritifera (L) Haw. is the correct name for the species concerned.
Gordon Rowley is right in surmising that we have not yet heard the end of the story. I do not yet know the correct position and can only say confidently that it has not yet been arrived at. (A whole lot more straw in the wind followed. The actual outcome was the typification of the Linnaean epithet pumila on as the Commel illustration t10 and this is the valid name in Haworthia). Tjaden is quite wrong in referring to a name-game being played by botanists. Names are part and parcel of the communication process and confusion can just as often be put at the door of gardeners as anywhere else. Duval was a gardener.
Regarding Gordon Rowley’s review, there are a host of major and minor discrepancies between Col. Scott’s perceptions and my own, and I am particularly disappointed in that the review examines none of them nor even reveals them. One of my chief complaints about reaction and response to even my own writing is the undiscriminating attitudes of the audience. Even my respect for Gordon Rowley does not deaden the impression that he has not actually read and comprehended what either Pilbeam, Scott or myself has had to say. Without meaning to be unkind or offensive, I think a much better review could have been written unless Gordon Rowley also finds the audience unselective.
Regarding Rowley’s references to monographs and revisions, I would like to point out that I have used neither term for my work. The cytological data which Rowley refers to as a vast body of published research is an indictment of the subject rather than a source of worthwhile information. Riley and Majumdar (1979), and here I think it unlikely that Rowley read the book, state “all determinations of chromosome numbers should be regarded with suspicion” and this sentence is qualified by a reference to “if they were made from botanical gardens collections of long standing”. It would have been better to have said “from plants of doubtful authenticity”. A common problem with cytological work is that one has to be sure of the material worked on, and although H.P. Riley does cite voucher specimens (which are lodged here at the Compton herbarium), the origin and identification of the material remains doubtful. Even a good dry specimen is not easy to identify. At present the best cytological work, in the sense that Rowley asks for, is that being published by Dr. Canio Vosa at Oxford. At least emphasis is placed on authenticity of material. Dr. Peter Brandham at Kew has also published proper cytological work on Haworthia and all the references available to me are listed in my handbooks. I am sorry if it is not obvious that I had also considered the contents. Brandham’s work is more orientated to chromosomal aberration than to species resolution, and in any case the results of his work on H. coarctata and H. reinwardltii seem to substantiate my mass transfer of varieties.
Rowley for some odd reason cites the basic chromosome counts in H. venosa, H. recurva and H. tessellata. These are all 14 but with polyploids in the latter two. What this bit of information is supposed to convey to anyone evades me completely. Scott mentions these counts too in Cactus and Succ. Journ. (U.S.) 50:77 (1978) with the same degree of breathless reverence. At least Scott points out that there is little possibility of knowing just what Ferguson in 1926 was naming H. recurva. If there is anything in the literature (including the work on chromatography by Riley and IsbeIl, 1963) to suggest any arrangement of species substantially different from mine, I stand helplessly to correction. My handbook was not written to duplicate what I wrote in National Cactus and Succulent Journal 28:80 (1973).
Regarding leaf surfaces, Dr. David Cutler was kind enough to photograph a good number of leaves using electron scanning micrography. I wonder if anyone can read more taxonomic sense into these pictures than I have been able to extract from them. It seems only possible to interpret them against a background of origin and with some prejudgement of taxonomic order. Dr. M. Hayashi of the Tokyo Breeding Research Institute has some very valuable results from tissue culturing and this will to some extent improve our understanding of species relationships. But any method like this which departs from standard herbarium practice of comparing dry barely recognisable specimens, moves onto shaky ground because of the real limits to the material that can be properly examined. At some time or another one has to reach some kind of a decision, and if we are going to wait for clear-cut anatomical, cytological and physiological studies, will we ever get revisions, let alone monographs?
The appearance of Col. Scott’s revision of Haworthia does in fact pose many problems for me and a superficial review is not very helpful to anyone. Who in his right mind wants to enter into a needless and fruitless disputation about who said what when, and about who is right and who is wrong? On the other hand, concerned readers must surely want to know what skeletons are lurking in the cupboard – so let’s open the door and see.
H. herbacea (Mill.) Stear is typified by an illustration in Boerhaave’s Index Alter Plantarum (1711). I apply this name to an element which Scott calls H. arachnoidea, in turn typified by an illustration in Commelin’s Praeludia Botanica (1703). Wijnands points out that this illustration is actually in better agreement with Scott’s H. setata than his H. arachnoidea, and further confounds the issue by stating that H. arachnoidea sensu Bayer is the same element as H. herbacea sensu Bayer, but omitting this relevant citation from his synonymy. Setting this aside, I would concede that my idea of the Boerhaave illustration representing my species causes problems, as I have long nursed the secret apprehension that the illustration more nearly represents H. magnifica V. Poelln. In a way this is a problem that both Scott and myself have been advised to avoid, i.e. selecting neotypes which may later be upended, it would have been difficult for me in 1976 to predict what interpretation Scott would adopt in 1985 and Tjaden may place the blame for confusion where he wishes.
In the case of H. angustifolia Haw. there does not seem to be a problem because nearly all authors imply that Salm Dyck’s illustration (Monogr. 13, t.2 1836) of Aloe stenophylla Schultes represents the species. Fourcade (in Trans. Royal Soc. S. Afr. 21:78, 1932) gives a new name for H. angustifolia sensu Baker, viz. H. monticola. He cites A. stenophylla Schultes and also Salm-Dyck t.2 as synonyms, and states expressly that Schultes’ epithet cannot be taken up in Haworthia because of the priority of H. stenophylla Baker (now Chortolirion). Schultes’ A. stenophylla is a direct transcription of H. angustifolia as described by Haworth. Under Article 63 of the Code, H. monticola is a superfluous name for H. angustifolia Haw. because Fourcade has indisputably included the type of that species. In analysing the discussion of his H. monticola (as I pointed out in Nat. Cact. Succ. J. 37:31, 1982 – one of many references not cited by Scott) it is obvious that Fourcade wholly misinterpreted both the Haworth and Baker descriptions. I do not contest that Fourcade had a different species, but this is only evident from mention of the origin of his specimens. He describes a plant which has translucence in the lower leaf. This is quite nonsensical and there is no comparision between the chlorosis of buried leaves and the translucence of exposed leaves in those species which do exhibit translucence. Scott’s description under H. monticola in this respect also deviates from Haworth, Baker and Fourcade, because he writes of a leaf with a semipellucid upper surface with a number of pellucid spots in the upper third. This is not at all evident in Scott’s illustration of the species and I would go so far as to say that he has two different elements in the picture. The foreground and middle-right plants are in my opinion H. chloracantha var. subglauca (V. Poelln.) Bayer. The other plants resemble specimens I have of H. divergens Bayer from Uniondale which are relatively unspotted. I did not take up Fourcade’s name for the very reasons then suspected and, even if only for that reason, see none to change now. Therefore:
[Syn. H. monticola Fourcade, nom. illegit., sensu Scott, The Genus Haworthia, a Taxonomic Revision, :57 (1976).]
(For the sake of peace and with some doubt about my own nomenclatural expertise, I later just abandoned any argument and reverted to the use of monticola.)
Col. Scott takes up a number of old Kew illustrations as types of Haworth’s species e.g. H. altilinea, H. mucronata and H. aristata. I put my case clearly in both editions of my Handbook and, contrary to Rowley’s supposition, these illustrations have been available to researchers outside Europe and were also seen by both Uitewaal and Von Poellnitz. Scott simply confirms the total confusion in the historical application of these names by illustrating H. habdomadis var. inconfluens (v. Poelln.) Bayer and H. cooperi Baker from Ladismith and Grahamstown respectively, as H. mucronata Haworth. His distribution map implies the inclusion of H. bolusii var. blackbeardiana (v. Poelln.) as well.
Col. Scott’s abandonment of subgenera and the characters which support them is inconsistent with exclusion of Astroloba, Poellnitzia and Chortolirion from Haworthia. If inflorescence characters of this degree are ignored identification must accordingly be complicated. I simply cannot understand Rowley highlighting practically useless cytological evidence and passing this over. The consequences to Scott’s perceptions of Haworthia are immediately disastrous where on page xii of his ‘revision’ the illustration C478 is identified as H. marginata which it cannot possibly be. Jaquin has an identical illustration in Hort. Schoenbr. :421 entitled Aloe pumilio which Scott at least has correctly labelled H. reticulata. On page xiii, C481 is obviously H. marginata and the flower positively excludes H. venosa as Scott suggests. C313 on page xii also compares very unfavourably with H. heidelbergensis on page 131 and I find it very difficult to believe that the species is correctly illustrated anyway. These misidentifications of C478 and C481 cross the boundaries of three major groups of Haworthia first indicated by Uitewaal. Scott finds it difficult to understand why H. bruynsii and H. springbokvlakensis should be separated taxonomically when they are separated… “geographically by only a few kilometres of geologically uniform undulating ground”. I find it equally difficult to accept a classification that for example separates H. scabra Haw. and H. tuberculata v. Poelln even at species level, let alone at sectional level. Scott’s exposition of his species concept does not leave me any the wiser, and the short section on the generic concept speaks for itself.
Without wanting to be unkind, I would have thought that some of the misstatements in Scott’s book should have been apparent to a competent reviewer. Gordon Rowley, having seen H. graminifolia in the field himself, should of all people know that it does sucker. I have avoided giving habitat data like the plague because it is so difficult to make true and usable statements. One has only to read Scott’s discussion of H. pumila to see on what quicksands one treads in this respect. Read first the discussion and then refer to the distribution map. Being right here on my doorstep, I should know this species very well. H. pumila is not found in strictly fynbos vegetation and the statement… “found under fynbos or other karoo scrub” can be valuable information only to the barmy. This species grows here on Malmesbury shales and it is only common in the Pteronia paniculata communities (one of seven such) where pH can be below pH 4 (KCI). These soils are also characterised by Euphorbiamauritanica communities where pH is often above 8, and H. pumila can be at home there too. It also occurs on soils derived from Witteberg quartzites, Dwyka tillite and Bokkeveld shales and I do not believe that this information is going to help any grower in the slightest. H. reticulata is similarly described by Scott as a component of fynbos, but is also said to grow on outcrops of shale. If anybody can be said to deserve credit for making such mutually exclusive statements then truly we can identify this epoch as the age of untruth.
Of what value is a statement for H. minima Haw. which reads… “A constituent of fynbos . . under renosterbos”? Fynbos is one veld-type and renosterbos is a major element of another. H. herbacea (H. arachnoidea sensu Scott) is also on my doorstep, and I am more than surprised to learn that it grows in sandy, well-drained acid soils derived from sandstone. While it has the same distribution range as H. pumila it is fortunately blessed in that it enjoys a higher rainfall maximum. Who will point out that H. angustifolia var. baylissii is represented by a single clone from one of the densest populations of H. angustifolia that I have seen and that in fact has no distribution? Who will point out that Scott’s illustration of H. serrata is definitely not that of my species and that it can best be equated with the form of H. arachnoidea at the northern end of the Tradouw Pass? In the case of H. marumiana sensu Scott, he cites H. archeri Bayer as a synonym and also H. marumiana sensu Bayer. He then goes to great pains in the discussion to exclude the latter and suggest that its “systematic position is not yet clear.” Uitewaal does describe his species as freely proliferous but Scott writes… “H marumiana sensu Uitewaal” is not.
To evaluate this revision of Scott’s we should perhaps look at his section Retusae which he first revised in Aloe 11, 4:17 (1973). The synoptic key (Rowley) consists of 8 steps each with two to three species. One of these steps reads “leaves in 5-tiers” and includes H. retusa and H. heidelbergensis, while another reads leaves in 2-4 tiers” and includes H. mirabilis and H. asperula sensu Scott. Under the description of H. retusa these tiers are described as vertical, while it is wholly unclear what 2-4 tiers means in H. mirabilis or H. asperu!a. The illustration of H. mirabilis shows a plant with 8 near-vertical tiers. In the illustration of H. asperu!a it is possible to resolve the two primary counter spirals, the three secondary spirals, and the five secondary counter spirals. The illustration of H. magnifica facing H. asperula shows beautifully the five secondary counter spirals in near vertical position. (I wrote the article in respect of leaf spirals in Aloe precisely to forestall and nudge both Dr Codd and Col Scott to a rational view.)
In 1973 Scott upheld the name H. nitidula v. Poelln. but the 1985 version does not cite this interpretation under H. longibracteata Smith where it belongs by virtue of its accompanying illustration. H. willowmorensis v. Poelln. sensu Scott 1973 becomes H. correcta v. Poelln. but is cited under the synonomy of H. mirabilis. Similarly H. sublimpidula v. Poelln. of 1973 becomes H. heidelbergensis Smith of 1985, but is not cited as such. The illustration and guide to H. heide!bergensis do not agree with the original description nor with this species as I know it.
The illustration of H. dekenahii is not that element but that of H. turgida var. pallidifolia Smith, which Scott does away with in a new concept of H. retusa. While H. asperula sensu Scott covers everything from H. pygmaea in the east to H. pubescens in the west, four species viz. H. geraldii, H. multilineata, H. fouchei and H. longibracteata are required for a relatively simple situation around Riversdale.
These are only some of many problems which I see in Col. Scott’s book. John Pilbeam’s book is in another class. Except where he enters the taxonomic fray it is the most useful book for the collector. The most serious objection that I have to it is the chapter on geographic distribution. The only nice thing I have to say about that is Pilbeam’s kindness in concealing the origin of most of it. This chapter should have been updated and corrected before publication.
The ‘definitive monograph’ is Gordon Rowley’s pie- in-the-sky. The realities of plant taxonomy are going to prove far more disparaging of hybrids and cultivars by taking them properly out of the rank of genus and species. Haworthia has just been fortunate in having being treated by non-botanists who are sympathetic to the collector. A truly botanical dispensation could reduce the Hexangulares to 11 species, the subgenus Haworthia to 18 and Robustipedunculares to 4. Teasingly I would like to say to Gordon Rowley that he should look at more than just the dust cover when he next reviews a book.
GORDON ROWLEY comments: ”In some books the dust covers are the best parts. But to be able to state that the only reference to chromosomes in all three books is on p.73 of Bayer’s surely indicates some attempt to look between the covers, doesn’t it? Maybe I just wasn’t reading the same volumes that Bruce has. Incidentally, I would love to know what is to become of the luckless hybrids and cultivars if they are to be deprived of the rank of genus and species!
Now, perhaps, could we invite Col. Scott to round off the review with his side of the story?” ♦