The ‘retuse’ haworthias are among the most fascinating of the genus and therefore Colonel Scott’s recent revision is of particular interest. This work…‘A revision of the genus Haworthia, Section Retusae” was published in ALOE, Dec.1973 and forms a new taxonomic record of the species involved. The object of the present paper is to examine the validity of this record particularly in the light of past history of Haworthia. A considerable literature on Haworthia is now available much of which points to the problems likely to be encountered in the group. Much of it also points to the consequences of inadequate species concepts for the group.
37. Haworthia turgida Haw., Suppl.Pl.Succ. :52(1819). Salm-Dyck, Monogr. 9:t5(1837). Bayer :163(1976). Bayer :57(1982). H. retusa sensu Scott :113(1985). Type: Cape, ex hort Kew. Not preserved. Neotype (B&M): icon, Bowie (K): H. laetivirens Haw., Suppl.Pl.Succ. :53(1819). Salm-Dyck, Monogr. 10:t3(1837). Type: Cape. Not preserved. Neotype (designated here): icon t.3, Salm-Dyck: H. caespitosa V.Poelln., Cactus J 5:33(1936). V.Poelln., Feddes Repert.Spec.Nov. 43:103(1938). Type: Cape, near McGregor, G.J. Payne in Triebn. 586. Not preserved. Lectotype (B&M): icon (B): H. caespitosa fa subplana V.Poelln. ibid. 44:232(1938). Type: Cape, Uniondale, G. Helm in Triebn. 874. Not preserved: H. caespitosa fa subproliferans V.Poelln. ibid. Type: Cape, Calvinia. Not preserved.
turgida: swollen.
Rosette partially stemless, proliferous, 5-10cm φ. Leaves 20-40, turgid often as thick as broad, recurved or slightly retused, generally mottled, yellow-green to pink in sun, margins and keel lightly spined. Inflorescence simple, 15-20cm. Flowers slender, brownish-white with darker venation.
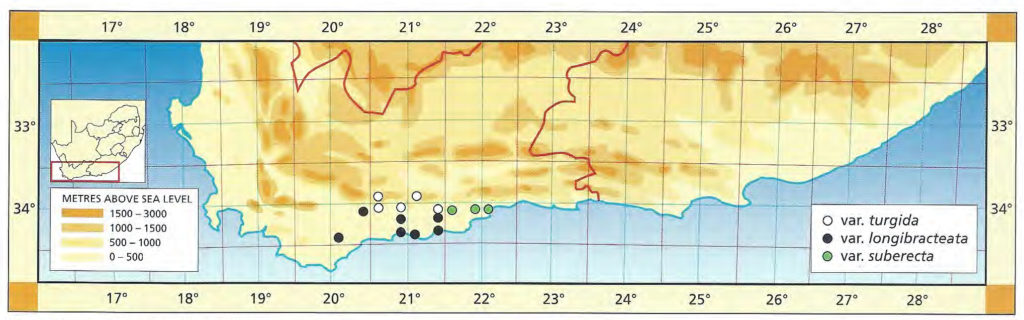
1982 – H. turgida is the most widely distributed of the Southwestern Cape species and also the most variable. It occurs in both the higher mountains in sandstone, and also on the lower lying shales. It is very subject to ecotypic variation and the best evidence of this is north of Bredasdorp eastward apparently through the coastal limestones, northward to Swellendam where there is possibly intergradation with H. reticulata, then eastwards again in the Langeberg Mountains and down all the main river valleys including the lower Breede River valley. The eastern limit is near Little Brak. The northern limit is of course the Langeberg mountain range and it is unlikely that H. turgida occurs inland along the Gouritz river ‑ despite an unconfirmed report of a collection northwest of Calitzdorp. Some of Von Poellnitz’s forms and varieties were recorded from as far afield as Calvinia and Steytlerville. Although the possibility of such collections cannot be completely ruled out, it is unlikely that these could have been relatives of H. turgida. There are forms in the Potberg mountains as well as in the Riviersonderend mountains which may intergrade with other local species (e.g. at Greyton with H. mirabilis) in the same way that H. turgida is related to H. retusa in the Heidelberg and Riversdale areas, or at Albertinia. It is a fascinating problem as it is obvious that affinities at one locality may not be the same at another. Thus it is quite conceivable that H. turgida may have an affinity with H. magnifica var. notabilis at Robertson, with H. maculata in the Hex River valley and with H. archeri through the inland mountains. H. turgida is generally a small species up to about 30mm diameter in the sandstones, however, in shales it may grow up to 80mm diameter. It is always very proliferous and grows on steep rocky slopes as opposed to H. retusa which is less proliferous and on level areas. H. turgida is also not withdrawn into the ground. This contrast of proliferation and withdrawal into the soil as opposed to clump formation also occurs in the related species H. reticulata and H. herbacea, and is evident in less strongly related species such as H. cooperi and H. cymbiformis.
1999 – The early illustrations by Salm-Dyck hardly allow room for doubt about this species, and yet the localities that von Poellnitz cites are grossly off the mark for the species and varieties which he described and cited. The problem that Col Scott had with this species and with H. retusa should also be explained here. It arises out of a misconception about leaf tiers and the statement ‘quinquefarious’ in the early literature. This term refers to vertical leaf tiers and it is apparent from a manuscript in the Grahamstown (Albany) Museum (and also from the revision in Aloe 11, 1973) that Scott concluded this to mean horizontal leaf layers. Also Scott’s treatment, where he applies the name H. retusa to this element, as well as to a number of von Poellnitz’ species and varieties H. laetivirens, is baffling. Breuer and Metzing have nominated a neotype which is not a good representation of the typical variety from the sandstones of the Langeberg mountains.
Of the southern Cape species, H. turgida is unquestionably the main role player. It occurs in recognisable form from Bredasdorp to east of Mossel Bay. It forms a continuum with H. reticulata and H. herbacea which are in the Worcester/Robertson area, and ranges from the Langeberg high peaks to the valleys running seaward and southward. It is unfortunate that apart from Smith’s names, few others have any geographical credibility. The affinities suggested in the 1982 discussion are in retrospect improbable. If there is a further association which needs exploration, it may be the link with the inland species through the high mountain forms. It does not seem sound to speculate on the chronological origins of the elements at altitude as opposed to those at lower levels.
a.var. turgida The typical variety based on Salm-Dyck’s renditions, rather than on the Kew illustration cited by Scott, which I have not seen, is considered to be the one in the Tradouw Pass east of Swellendam. The leaves are almost recurved, highly mottled and moderately spined. The higher mountain forms in the sandstones are generally smaller and note has to be taken of similarities between these smaller softer elements of apparently disparate species e.g. H. mirabilis var. consanguinea, H. maculata, and H. vlokii, all at high altitude in sandstones. At Heidelberg H. turgida expresses its full potential in terms of ecotypic variation where it passes from sandstones, to shales and on to the clays of the Witteberg series. It also appears to hybridise with both H. heidelbergensis and with H. floribunda in that area.
Distribution: 3320 (Montagu): Tradouw Pass (-BC), Read (BOL), Smith 3247, 5160, 6784, 7517 (NBG); S. Barrydale (-DC), Smith 3902 (NBG). 3321(Ladismtih): 16km N. Riversdale (-CC), Smith 7197 (NBG); NE. Riversdale (-CC), Smith 5385 (NBG). 3420(Bredasdorp): Buffeljachts (-BA), Smith 4941 (NBG); Heidelberg (-BB), Smith 5044 (NBG), Kramer in NBG841/60; N. Heidelberg (-BB), Smith 6203 (NBG); Bayer in KG240/72, in KG 241/72 (NBG); Blackdown (-BB), Smith 5546 (NBG). 3421(Riversdale): Glen (-AB), Muir 3006 (BOL); Klein Kruisriver (-AB), Smith 6075, 6080 (NBG).
Inadequately located: ex hort. Whitehill, NBG68371, NBG68056, ex hort. Ross-Frames, NBG81/44, NBG2937/32, NBG68057, NBG1827/28, Smith 3240, 3429), 3498, 3904, 3950 (NBG); Swellendam, Ross-Frames (BOL); Riversdale, Muir in NBG1827/28 (BOL); Albertinia, Muir 1300 (BOL).
Haworthia turgida var. turgida JDV89/20 north of Heidelberg. This population seems to have been influenced by H. heidelbergensis.
Haworthia turgida var. turgida JDV89/23 north of Heidelberg. A little further north the plants are transforming to the smaller typical appearance.
Haworthia turgida var. turgida JDV85/75 Tradouw Pass. It is characteristically found in the sandstones and can occur in dense masses on steep rock faces.
b.var. longebracteata (Smith) Bayer comb.nov. H. longebracteata Smith, JS.Afr.Bot. 11:75(1945). Scott :127(1985). Type: CAPE-3421(Riversdale): near Stilbaai, Dekenah 18 in Smith 5378 (NBG).
longebracteata: long bracts.
This variety is fairly large and represents the lower-lying larger forms which occur from Bredasdorp across to the Kafferkuils River east of Riversdale. The typical form is from the Stilbaai area where the leaves tend to be suberect with a fairly pronounced end-area. To the west this end-area is less pronounced and both the Bredasdorp and the Swellendam (southwest) forms are reminiscent of the flatter non-retused leaves of H. reticulata.
Distribution: 3420 (Bredasdorp): 5km SW. Swellendam (-AB), Bayer 2420 (NBG), van der Merwe in NBG1184/28, Malherbe in NBG204/41; Napky (-AB) Bruyns in KG37/77 (NBG); 20km N. Bredasdorp (-AC), Venter in NBG868/38, Malherbe in NBG470/42, Smith 3949, 5484 (NBG), Bayer in KG34/70 (NBG); Kransriviermond (-BB), Smith 5752 (NBG); Diepkloof, S. Malgas (-BC), Bayer 2533 (PRE); 3km W. Vermaaklikheid (-BD), Kramer 434 (PRE). 3421(Riversdale): Kafferkuils Bridge (-AB), Smith 6795 (NBG), Bayer 4479 (PRE); S. Riversdale (-AB), Smith 7181, 7204 (NBG); Duiwenhoksriver (-AC), Bayer 2672 (NBG); Brakfontein (-AC), Smith 6107 (NBG), Bayer in KG94/71 (NBG); Botterkloof (-AD), Smith 5378 (BOL,PRE); Kransfontein Farm (-AD), Bohnen 7735 (NBG,PRE); Stilbaai (-AD), Fouche 46 (PRE); Dekenah 18 in Smith 5378 (NBG), Smith 2242, 2811, 5382, 6794 (NBG).
Inadequately located: Heidelberg, Ferguson (BOL).
Haworthia turgida var. longibracteata JDV89/24 north of Stilbaai. Always on rocky steep slopes and in dense clusters.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Very like H. reticulata except for the larger, more turgid and spreading leaves. The flower is much less conspicuous.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Becoming very colourful under stress.
Haworthia turgida var. longibracteata JDV86/6 north of Bredasdorp. A very isolated population.
c.suberecta V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, George district, Mrs Helm in Triebn. 844. Not preserved: Neotype: CAPE-3422(Mossel Bay): Brandwacht (-AA), Bayer in KG631/69 (NBG): H. turgida var. subtuberculata V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, Mossel Bay, Lategan. Not preserved: H. turgida var. pallidifolia Smith, JS.Afr.Bot. 12:10(1946). Type: CAPE-3421(Riversdale): Draaihoek, Albertinia, J. Dekenah 146 in Smith 5714 (NBG).
suberecta: leaves almost erect.
Here the variety is extended to include all these more strongly retused, clump-forming populations which extend all the way from Brandwacht, northeast of Mossel Bay, to Albertinia. This variety is strongly mottled and the leaf ends are slightly truncated and rounded. Smith based his var pallidifolia on a pale-coloured variant which is actually not very common. The Gouritz River form is the most truncated and as, in the typical variety, is highly mottled with semi-translucent dots.
Distribution: 3421 (Riversdale): Draaihoek, Albertinia (-BA), J. Dekenah 146 in Smith 5714 (NBG), Smith 5045, 5490 (NBG), Bayer 4476 (PRE); Weltevrede (-BA), Smith 5537, 7202, 7203 (NBG), Bayer 4471 (NBG); Wydersriver (-BA), Smith 2890 (NBG); Droogerug (-BA), Muir 3006 (BOL); Nuweberg (-BA), Bohnen 9024 (NBG); E. Valsch River Bridge (-BA), Smith 5714, 6912 (NBG), Bayer 4477 (NBG); Gouritz River (-BB), Fouche 48 (PRE), Smith 3857, 3961 (NBG), Bayer 4478 (NBG). 3422 (Mossel Bay): Brandwacht (-AA), Herre in STE6374 (BOL), Bayer in KG631/69 (NBG)
Inadequately located: Mossel Bay, Smith 4952, 5078 (NBG); ex hort Smith 647 (NBG), Naude in NBG1
Haworthia turgida var. suberecta JDV84/74 northwest of Mossel Bay. Leaves abbreviated and rounded at the tips.
Haworthia turgida var. suberecta JDV87/108 Grouritz River. If not so proliferous this would probably be a much more popular plant.
Haworthia turgida var. suberecta JDV94/101 Brandwacht north of Mossel Bay. On stony clay slopes.
Haworthia turgida var. suberecta JDV94/88 northwest of Albertinia. The origin of Smith’s var. pallidifolia in which the leaves are very pale.
I wrote a short note for Haworthiad, to explain a picture of Breuer’s new species H. fusca (MBB7507), and said “… the fact is that it is from a small population just west of Albertinia en route to another of Hayashi’s (?) species H. esterhuyzeniae, and also to Breuer’s H. vincentii. As readers we are being conditioned to accept that there are many kinds of species such as biological species, morphological species, taxonomic species, good species, bad species etc. etc. so a Latin binomial could mean anything (and the word ‘tautology’ has been added to my vocabulary). Botany needs a sensible and practical handle to a squadron of populations from between Albertinia and Great Brak. I would gladly supply this if somehow I could be assured that the act was not seen to be the clown’s contribution to the circus.”
Without any assurance, but with the encouragement of Stirling Baker, I am going to try and produce an explanation.
Put very bluntly and without any apology to a group of people who definitely deserve better, my life experience is that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well. I have already written around the subject a number of times and do not want to repeat what is not necessarily true other than the contribution these thoughts have made to my personal psyche.
In this contribution I am discuss, illustrate and then propose that there are just two species, H. retusa and H. pygmaea in a complex where presently more than nine species and varietal names are being used. I do this in consideration of all the populations of Haworthia known to me in the winter rainfall biome. Thus I recognize the need to rationalize species like H. mirabilis (which will then absorb H. maraisii, H. magnifica and H. heidelbergensis, and H. retusa (which will absorb H. turgida. There is a major problem in that the populations indicate three species in the west, viz. H. mirabilis, H. retusa and H. mutica but these appear to fuse or morph to two in the east. My past treatment of species and varieties like maraisii, magnifica, acuminata, dekenahii, argenteo-maculosa will bear witness to the nature of the (my) problem.
Gist Charles Craib includes a provocative chapter on taxonomy in his marvelous book on The Grass Aloes of the South African Veld (Umdaus Press). Not so curiously, John and Sandra Burrows voice many shared sentiments in a similar grand book on the Ficus species of Southern Africa, Figs of Southern & South-Central Africa (Umdaus Press). These books coincide with my recent experience with Drosanthemum taxonomy to which I have been peripheral since I first tried to collect D. speciosum and its allies. An account of this experience is in manuscript, and like my experience with Oxalis already published, should really be read by people with taxonomic aspirations. Of course they will not, as neither will taxonomists do who seem to already hold very superior individualistic opinions about what species are. There is absolutely no doubt that there is a very commonly held view, that what constitutes a species is based on opinion only.
Perhaps just a single illustration will indicate the kinds of arguments one has to swallow, or refute, if ones perceptions are being threatened or distorted by the decisions which are made and the conclusions which hang on them. In an article which “officially” relegates yet again the single species of Poellnitzia to some other genus (thanks be at least this is within the ambit of the aloe group of genera – the genus Astroloba) some justification is made that … “the resemblance between the flowers of Poellnitzia and the bird-pollinated genus Microloma (Asclepiadacea) is particularly striking”. The author of this really curious statement took me to task when I pilloried it, because there is a publication which conclusively shows that sunbirds do gather the pollinia of at least one Microloma on their tongues. But there are several species of Microloma and I am quite sure that they are not all bird-pollinated and besides, that is not really the point. Here I want to illustrate this time the flowers of Astroloba(Poellnitzia) rubriflora, Microloma sagittata, and M. tenuifolia and that ask readers themselves to decide if the similarity is indeed “particularly striking”. (See figures 1, 2 & 3). The arguments are no different in rationale to those of Charles Craib who treats Chortolirion as a grass Aloe because it meets his particular approach. Whose opinion do we need to follow?
Charles Craib’s decision is based on the usefulness of a classification in respect of his needs. He has kindly sent me a recent manuscript discussing the merits of a new species of grass Aloe that he recognizes from his own knowledge and experience is a product of poor taxonomy. This brings me to the conclusion that taxonomy is not science as I have so often tried to say. Latin names are simply nouns and all that is necessary is that people using names know to what is being referred. A simple example is the fiasco of the name change from Aloe bainesii to Aloe barberae. After 40 years or more where all and sundry have happily used the name bainesii,somebody discovers that another name precedes it by the thickness of a sheet of paper. Aiding or compelling this change in my attitude, I have to admit that my own classification of Haworthia no longer really meets my needs and in the climate of taxonomy that I experience, I am not sure what should be done. Thus in this article I am going to examine what I consider to be a good example of the problems any taxonomist who ever gets a good enough sample will eventually face. The question will have to be asked – are species real?
The area under consideration This is the area east of the Goukou River which bypasses Riversdale, then to south of the high Langeberg Mountain range and reaching Great Brak in the east. The southern limit is virtually the N2 national highway between Riversdale and George, although my sampling drops south of the N2 approaching the Gouritz River from the west, and dropping south to the sea east of the river. The altitude is from sea level to about 250m. The rainfall is moderate and generally falls in winter, although it would be sensible to note that the area is really a westerly extension of the all-year rainfall area forested coastal strip between Port Elizabeth and George. The vegetation is coastal renosterveld although there are areas of Karoid Valley Bushveld especially along the river valleys. The geology is very complex and plays a critical role in plant species distribution and occurrence because the soils are very skeletal. It is often the case that Haworthia is associated with exposed rock where biomass potential and vegetation competition are low. There are five main geological groups represented in the greater Southern Cape area, and the study area is different in that it is dominated by a geologically differentiated formation of the Bokkeveld shales – different to the same formation which dominates west of Riversdale. A subtle factor is that this eastern formation produces, as a weathering product, a range of clays and minerals significantly different to those from the western formation. The eastern part of the study area is dominated by rocks of the more recent Uitenhage formation; while south of the N2 (outside the study area) the formation is the most recent Bredasdorp formation.
I asked Jan Vlok for his views on what may make the area ‘different’. He replied… ”Something I have noted repeatedly is how important river drainage areas are in determining vegetation units and pro parte also those of localised endemics. I have been wondering about the issue why river drainage basins would be so important; and currently think they reflect the complex interaction between several factors – geology, rainfall, landscape of area and in thus the history of the evolution of the area. Sessile organisms – especially plants with limited seed dispersal abilities are obviously strongly affected by these factors.
In your area of interest we have one very important role player – the Gouritz River. It tended to obliterate everything in its way cutting away habitat at a relative fast rate, thus creating many new micro-habitat sites for species to establish from the north and some coming in from the east-or-west. It’s a real “bastard” area with hybrids swarms of e.g. Tritonia and Watsonia species rather common. I guess they are “species in the making”, yes, they look very different from other populations but with no clear cut morphological characteristics (the latter painful from a taxonomic point of view, but great from a functional evolutionary-ecological point of view). Note here that less sessile organisms – especially with wind dispersed seed, e.g. orchids and for that matter asclepiads are less restricted from re-colonising or finding suitable habitat in adjacent river systems. So e.g. Duvalia immaculata has much more freedom to hop from one habitat, over the watershed into the next system, than any of your Haworthia species has. Those have to do the slow march, finding it difficult, but not impossible to cross the river or watershed, and much more slowly. This implies that in the Great Brak to Riversdale area, an east-west dissemination of Haworthia populations has been relatively slow – selecting for numerous micro-habitat specialists, but perhaps each population representing nothing more than parts of one large meta-population.
East of the mighty Gouritz we have the Brakke drainage system (Groot & Klein Brak Rivers). Much smaller & slow cutting into the landscapes, thus greater stability in micro-habitats – added is slightly higher rainfall and certainty of rainfall. Thus no surprise that some populations have evolved to more discrete entities, you call them Haworthia parksiana and H. kingiana.
West of the mighty Gouritz we have the Goukou drainage system. Clearly much more stable as in cutting away the landscape than the Gouritz, but still supplemented by some summer rain. In terms of vegetation mapping I found the area really difficult. Had to look at every corner as I could not predict what will occur where. The closely related sister to the west, the Duiwenhoks has been equally challenging, except that things started panning out better. I guess its related to less reliable summer rain, thus summer drought stress. It is also a less might river, with an odd geology – suddenly conglomerates reappear. So in the Duyvenhoks plant populations start taking on unique characters – as well as do the common and dominant species of the plant communities they occur in.”
Jan Vlok confirms my opinion that the only real factor in the biogeography apart from the complex geology at Great Brak, is the Gouritz Valley. It is mainly the geology which seems to impact on H. parksiana and H. kingiana. The river does not affect either H. minima, or H. turgida. There is some doubt about the interaction and variation of H. chloracantha and H. floribunda across the valley. The populations (splendens, fusca, esterhuizenii, vincentii and argenteo-maculosa are in my opinion a series of consecutive populations which can be regarded as a continuum unbroken by the river valley. There seem to be very few species or any other biogeographical evidence to suggest that Haworthia has been impacted on.
The Haworthia populations Those considered here refer only to the species of the subgenus Haworthia and exclude H. variegata, H. floribunda and H. chloracantha of that subgenus. One must assume that by this stage, most of the relevant populations are known although I cannot claim to have personally explored the full potential of the area. Altogether I know of 29 populations which need to be considered in terms of their probable taxonomic affinities. They are listed according to my interpretation of the present available nomenclature as follows:
Haworthia turgida (Haworth 1819) Draaihoek, N Draaihoek, Valsch River, Wydersriver, Ouvloere, Die Eiland, Gouritz bridge, S Gouritz Bridge, Brandwacht.
Haworthia pygmaea (von Poellnitz 1929) Herbertsdale, Dumbie Dykes, Mosgas, SW Klipkop, Klipkop, Great Brak, N Great Brak, W Great Brak. (The vars crystallina and major (Pilbeam 1983) are not population based entities).
Haworthia magnifica var acuminata (Bayer 1976) West bank Gouritz at Melkhoutfontein (three separate populations), Vleesbaai.
Haworthia magnifica var dekenahii(Smith 1944) Draaihoek.
Haworthia magnifica var argenteo-maculosa (Smith 1945) Cooper Siding, E Cooper Siding, Humor
Haworthia magnifica var splendens (Venter & Hammer 1998) Soutpanskoppies, Snymanskraal.
Haworthia magnifica var esterhuizenii (Hayashi) Aasvoelberg.
Haworthia magnifica var vincentii (Breuer) Welgevonden.
Haworthiamagnifica var fusca (Breuer) Paulsfontein.
These populations are generally isolated from one another to some degree and it is evident that there are four taxonomic units represented by single populations.
A taxonomic species overview Of course the two words “taxonomic” and “species” used together is tautological as taxonomy is the classification of biological entities of which species is the prime unit. We have to consider what could now be species in terms of some definition, and in terms of all the other populations both inside and outside the study area. In this case there are a great many elements which fall in the same class within the same subgenus of Haworthia. I am not aware of anyone who has really addressed the issue of what comprises a species in the family Aloaceae and certainly not in the genus Haworthia. Breuer did essay a “concept” of Haworthia species, which was in fact a listing of species names for the genus and had nothing to do with questioning the nature of those species. This is in my observation a critical issue in most taxonomic work and it is the subject of many learned publications. I have already made my voice heard on the subject on many occasions and am not going to repeat any of my misgivings other than to say that I want to discuss the above populations in respect of a definition for species which I have fashioned for myself from experience and from a very wide literature.
This definition is that species are dynamic fractal systems of living organisms which are morphologically, genetically and behaviorally continuous in space and time. My attitude is that species have to be considered as phenomena in a creation and that these are spread in space and change with time. When assessing any plant as a species, one has to ask where it comes from, what its attendant companions are, what their variables are, and what their spatial relationship is. One also has to ask, what is all this in relation to geographical considerations including geology, topography, drainage systems, climate, vegetation and other plants of tangibly different families and genera? My experience with many taxonomic works and workers is that it is extremely difficult to approach the subject without extreme caution and also expect a considered response. While I have touted the view that classification is science, it most definitely is not managed in that way. It appears that there are two completely different needs that are met in classification. One is the purported one of botanical science, practiced mostly by herbarium based botanists or in present time, by institutionalized “gel-jocks” trying to lay bare the mysteries of DNA. Both are handicapped by the material available or sampled for their work, AND by their field expertise. The second need is that of the collector who simply needs a name by which to communicate with sources of plants, with other collectors and to have some measure of the difference in appearance of the plant from others he/she knows. Thus both needs will have to be met within the limits of the material known, seen, examined and experienced, and according to the competencies of the scientist, botanist, chemist, collector whoever happens to need an appellation for a plant. Taxonomic accounts and revisions are simply a product of the competence and experience of the writer limited also by the extent to which material, seen and sampled, matches or represents the reality. The term reality also has a special significance and one can rightly ask if this creation and its phenomena have any reality.
The evaluation I do this evaluation in the light of a synopsis of Haworthia which I submitted to the SA Biodiversity Institute for a flora of the Eastern Cape. That synopsis generates the question of how the species of the Western Cape should then be summarized. Quite fortuitously a parallel, although minor, problem seems to be resolved. In about 1976 I suggested that the flora of the Cape would never be understood until it was recognised that the true biome nature of the vegetation and the present-time Cape Flora was that of a Winter Rainfall biome stretching from Southern Namibia to the Eastern Cape. This now appears to be reflected in current botanical thinking where the Succulent Karoo is seen to be intimately linked to the Fynbos. The impact of this is that one really needs to re-think how species elements of the Southern Cape, are linked to those of the Little Karoo. Fortunately this does not really touch the elements I will discuss in this article although there are some peculiarities. For example, there is H. parksiana, H. kingiana, and Duvalia immaculata which are endemic to the Mossel bay area, while H. minima leapfrogs the Langeberg to occur in the Little Karoo.
What is more important is the present classification (of mine) which links most of the elements I am dealing with, to H. magnifica that is in the Riversdale/Heidelberg area. The other principle elements outside the study area bearing on the discussion are H. retusa and H. mutica.
Outside of vegetative physical appearances, there are only two really tangible pointers to relationships. One is population structure and co-existence (do the populations share close habitat or not), and the other is flowering time. Physical appearance is fairly helpful in that there are quite notable differences. There are plants which are generally light coloured and which are either adapted to steep rocky habitats as clumping cliff dwellers or are generally more solitary and adapted to level habitats. These are spring flowering. Then there is a set of populations in which the plants are very seldom in cliff habitats, are dark coloured and flower in late summer.
The first spring flowering group comprises (for the purpose of this article) H. retusa and H. turgida. There is no doubt in my mind that these two are essentially the same species as they never co-exist. The actual situation is very complicated and the recognition of H. mutica as the western representative of H. retusa and which can be said to co-exist with H. turgida, is indicative of that.
The second summer flowering group is even more complex and it is possible to suggest that H. mirabilis as presently constituted, should be enlarged to include H. maraisii, H. heidelbergensis and H. magnifica just west of Riversdale. In the context of biomes I would be rational to suggest that such treatment would need to consider also the incorporation of H. emelyae in view of its possible interaction with H. maraisii.
In respect of the study area it is thus evident that we need to look primarily at flowering time as all the populations are essentially in the light-coloured ambit rather than in the smaller dark green one of H. mirabilis. I do not imply hereby that this is inarguable, but it is evident that we need to understand that in the west we have a triumvirate of possible species with two distinct flowering times, whereas in the study area there seem to be two main elements which are both essentially spring flowering.
How do different people see the situation?
A. Evaluation and observation by Gerhard Marx Both dekenahii (Draaihoek) and fusca flower more or less together with retusa and emelyae (except major) in September-October. Both are in fruit in early November. In cultivation they also flower in March-April but I have never been able to visit them at that time in habitat to see whether they do the same. I would think so.
Splendens (Snymanskraal/Soutpanskoppies) flowers generally (late Dec. to early February). It is the one and only flowering spell for splendens.
Argenteo-maculosa and (its ‘variant’) vincentii also flower together with dekenahii. In Grahamstown argenteo-maculosa flowered for me throughout summer and even into winter. They seem to do the same for Steven Hammer in Vista. But I think in the wild they also have two flowering spells, one in spring and another in fall.
I seem to remember the flowering periods of acuminata are exactly the same as for dekenahii.
Pygmaea from Great Brak and Mossgas flower exactly together with dekenahii , but argenteo-maculosa (Humor and Cooper Siding) flower later. They are still developing flower buds at the moment.
As mentioned above, H. emelyaemajor also flowers later than the ‘picta’ types and ‘breueri’. Major is only starting to flower now, while all other emelyae are just about done releasing seeds.
Thus at present I see the relationships as follows:
H. splendens (Soutpanskoppies/Snymanskraal) is a good species and stands isolated in terms of its flowering time, chunky and glossy leaves and slower growth.
H. dekenahii can perhaps also stand as a seperate species and fusca is definintely a variant, distinguished from the dekenahii by its red-brown colour (meaning no doubt the “rusty”-like difference) greater tendency to offset and geographical separation.
H. argenteo-maculosa – (which I feel it is such an inappropriate name!) – is another separate thing, although it shares superficial characters with splendens on the one hand and acuminata and even turgida on the other. Apart from different flowering time to splendens, it is much ‘softer’ and grows twice as fast and is also smaller with more acuminate leaves that are spotted along the sides. H. vincentii can probably be seen as a variant of argenteo-maculosa as they are so obviously similar, except that vincentii has fewer and longer leaves that are less heavily flecked and it is even more ‘tender’ (softer) than argenteo-maculosa.
H. pygmaea is fine as it stands, with Great Brak/ Dumbie Dykes/ Rooiheuwel/ Mossgas/ Herbertsdale all sharing basically the same characters and flowering time.
Argenteo-maculosa (Cooper Siding and Humor) plants are different in having heavier flecked and chunkier leaves and to me it is a toss of a coin whether they are seen as a variant of dekenahii or pygmaea.
Similarly I think acuminata can just as easily become a variety of turgida or dekenahii rather than of magnifica.
Another option is to keep dekenahii and acuminata variations of magnifica and add fusca, argenteo-maculosa and vincentii also as varieties. But then it is a case of giving them all the same ‘status’ where I feel that fusca and dekenahii are ‘unidentical twins’, while argenteo-maculosa and vincentii are an almost-identical-yet-different duo.
The above decisions are really based upon subtle factors and can be decided by tossing a coin. What I feel is important, is that splendens should be separated as it does not fit in well with the dekenahii–pygmaea bunch or 100% with the magnifica from Riversdale. In terms of flowering time I can see the logic of including it with magnifica and atrofusca, but then dekenahii and acuminata should be removed.
The analysis closes with the words … ”Wait, let is that me sign off before I write myself into a corner.”
A1. Reaction comment by M B Bayer There are two important considerations. One is that Gerhard does not profess to be a botanist or to be too concerned with taxonomy. The other is that he is assessing the situation in terms of his specific life experience in relation to all the variables I mention earlier. He does not see to have considered the many turgida populations and their variation, the Vleesbaai population of acuminata, and another important single population of pygmaea at Herbertsdale, and neither does he relate these to a greatly compounded problem when all the relevant known Western Cape populations are added.
My views are that splendens is significantly different because the two known populations are associated with ochre deposits. Fusca is on a less iron/ochre rich site. Dekenahii is limited to very few plants (and there were ever many) on shale. Can these very small and highly localized elements seriously be regarded as species in terms of my definition? Argenteo-maculosa and vincentii are associated with table mountain sandstone. The argenteo-maculosa populations are also linked to sanstone while the acuminata populations are linked to old river terraces and the Vleesbaai population is linked to sandstone again. The Hebertsdale population is related to Uitenhage conglomerates and all the other pymaea’s to an array of substrates around Mossel Bay, Little and Great Brak.
Turgida occurs on steep rocky river banks (shale cliffs) and in two places can be said to coexist with pygmaea or dekenahii (Dumbie Dykes and Draaihoek). It is not outside the realm of possibility that the differences in vegetative appearances are responses to different habitat and substrate. It should also be noted that the turgida var pallidifolia was described from Draaihoek too. I saw such clones at the site when I visited the area in ca 1972, but on a recent visit no such light coloured clones were apparent. The population further down the Valsch River is also highly patterned as are those at Draaihoek. It should be noted that there used to exist another population to the north of Draaihoek which I would have equated with H. turgida var longibracteata from the Heideberg area or lower Goukou River at Riversdale. The population at Wydersriver is not known to me apart from photographs of G.G. Smith and also seems to fall outside of the ambit of turgida var suberecta as I classified all these eastern populatons of turgida. The one population which I recently examined was on the Wyders River northeast of Ouvloere (Albertinia district) where the plants definitely suggest affinity with vincentii. Examining my photographs, which are reproduced with this article, I thought there was a remarkable similarity between clones of fusca and argenteo-maculosa at Humorum or Humor.
Of course one can adopt a different species definition and even suggest that simple superficial resemblance of a minimal sample is adequate for an opinion forming process. It is certainly difficult to even attempt to examine how these populations became isolated or even to ponder exactly how isolated they really are in relation to pollination mechanisms and seed dispersal.
A2. Response by Gerhard Marx to the re-drafted manuscript I do not know whether to feel flattered or embarrassed that you took my loose comments upon the splendens/pygmaea/turgida group so seriously. Only a few minutes after sending the e-mail away, I wanted to add more questions and comments that partly contradicted the ones I sent.
It is really a complicated puzzle, particularly if one takes flowering time in consideration. When I look from dekenahii to that chunky atrofusca and to fusca and splendens, I do feel that they really neatly fit as varieties under the same species. But then splendens‘ flowering time removes it distinctly from the others and I do not know what to do with that fact.
I feel that then splendens might just as well be thrown in with H. emelyae because of the obvious similarity in appearance. The Moerasrivier/ Saffraanrivier area form of emelyae even has almost the same ‘pimpled’ upper leaves and some glossiness. However, I must say that I do like your proposal to have the whole bunch under H. pygmaea. From a collector and non-scientist view as my own, it feels good and definitely an improvement upon he dispensation as in Haworthia Revisited.
There have been brief moments when I even wondered why they can’t all be retusa. When I first saw H. fusca in the wild I was convinced it is a form of retusa. And after comparing it in cultivation with dekenahii, I became convinced that it is closer to dekenahii. Looking at your Herbertsdale pygmaea photos, I can clearly see shades of acuminata in them, as well as argenteo-maculosa (particularly Cooper Siding) and, of course, Mossgas pygmaea. This supports the idea of having them all as varieties of one ‘superspecies’. But I can find it easier to see esterhuizenii linked to pygmaea than retusa, so that is why I like your idea of having pygmaea as the umbrella species.
To my mind there remains the problem with splendens‘ flowering time that matches up with the mirabilis and atrofusca/magnifica group. And mutica, of course, is again linked to the pygmaea group in terms of flowering time. Particularly if you enlarged our concept of the mirabilis complex with new introductions like ‘shaggy dog’ and plants from ‘Die Kop’ etc.. Maybe splendens could even fit in there.
I really don’t know. That is why I often said that at times I have some sympathy other taxonomists that are inclined to call each differing population (or plant) a separate species ! It is just so much easier.”
A3. Response by M.B.Bayer To this, my response was…”it should not be found to be embarrassing – I think this is perhaps what influences taxonomy and why you do not want to be seen as a taxonomist. You are a taxonomist – we all are. Science and especially nomenclature have put a shroud around the whole process to suggest only those really in the know can make a good decision! What you have not done – and it is not a failing – is to consider the variants of turgida. Flowering time has to be considered with the same reservations as any other character and in the end I think it is an intuitive judgment which has to be made and not a product of scientific reduction and pedantry.
The inclusion of emelyae just adds more problems because there is a whiplash onto heidelbergensis and paradoxa via major and multifolia – and there are no clear answers to that either. The variation just folds back on itself. Hence my references to “fractal” – endless variation never repeating itself! My end point looks as though there is no solution except by consensus in which contributors also recognise intellectual and experiential boundaries?”
B. Comment by anonymous knowledgeable collector “About your ms. I understand that I am looking at it as an under-informed collector, and it makes me realize how completely I have failed to keep up with current writings on haworthias. But like our old friend has demonstrated time and again, ignorance is no obstacle to the collector in offering opinions.
I do agree with you that the populations between Albertinia and Great Brak need – to use your words – a sensible and practical handle. Your ms., however, has not persuaded me that it offers such a handle. From my point of view, to reduce these populations to two species obfuscates rather than clarifies existing population differences. To base your conclusions primarily on flowering time sets aside the differences in physical appearances which have so fascinated us and which have seemed so important in the designation of species. But that must be your objective? The ms., however, does not offer, at least to me, sufficient argumentation that flowering time can be used to solve, or resolve, the Albertinia-Great Brak population differences. And almost as an aside, I must note that, while you write that it appears that there are two completely different needs which are met in classification, you are willing in effect to set aside the collectors’ needs in order to reach a conclusion. The dual naming system which some of us now use because the existing formal system does not meet our needs is bad enough; no need to make it even more prominent by encouraging it.
B1. Response by M.B Bayer My response to this was…”May I incorporate your opinions as I have done with Gerhards?
I think the point which you may miss is that essentially the naming system is for botany, in fact all of biology, and there should be only one goal viz the recognition of living systems. There is absolutely no reason for collectors to insist that Latin binomials are required for their activities. Why on earth cannot collectors use H. pygmaea “splendens“, H. pygmaea “Herbertsdale”, H. pygmaea “vleesbaai”, turgida “pallidifolia“, turgida “sub-erecta” turgida “Wydersriver” etc. What more do they need? It seems fairly obvious to me that there is just a veneer of pretence that the latin names mean something more and which they aspire to? There is no way that anyone else is going to arrive at a classification of these populations (considering all of them) which is more meaningful. What we see in Haworthia is that physical appearances, as variously we see them, are not aiding identification or classification and flowering time does offer a view of a system which reflects over the entire series of populations. But in nortieri I can show that flowering time does NOT do so!
I cannot see how you say I am setting aside collectors needs. I am a collector and I want a system that is true and reflects what I know and experience. This is why I started fiddling with the names in the first place. Why have we so often considered that place names should be used? It is all very well and good for collectors with plants bearing names as they were obtained, to insist that those names have meaning and are immutable as opposed to those who have unnamed plants for which they need a designation.
I spent some time in the field with nortieri. This is a species which is grossly under-represented in collections in relation to field abundance. It must be one of the commonest and most widespread in terms of actual occurrence. We have the names globosiflora, devriesii, albispina, pehlemanniae and agnis as well as nortierimontana and gifbergensis and it is the most under-collected species of the genus. We do not have names for Arizona, Ouberg Pass, Opdie Berg, Moordenaarskaroo, Hex River Pass, Blinkberg, Breekranz, Nuweplaas, Dwasriver, and a few others which could be said to be different. How many more names are we going to generate based on existing approach? This does not worry the collector who is glad to have new names entering the system slowly enough for him to afford to add them.
What I am now saying is that there has to be an agreed system and that it is up to the collectors themselves to come forward with some sensible proposals. As a group they definitely do not have any idea of biological systems or even much grasp of nomenclature, the controls it puts in place and the problems that they also creates. I think I have said elsewhere that I am perceived to be the “trouble-maker” when in fact I seem to have spent my life pointing out that we are basing our hopes on method and system which offers no hope of stability or agreement.”
B2. Collectors reply The collector responded…”I fear that at the present we are far apart both in our individual thoughts and how we understand one another. What to do? There follows several unconnected comments or thoughts of mine on your ms. But first, I don’t mine if you use anything I write, even if it to knock it down, but I would be grateful if you would not use my comments as you have Gerhard’s, with reference to me. To me, what I write are “private” views which may not be supportable and which I may change tomorrow, if not sooner. I still cannot accept in my own mind the idea of using blooming time to formulate species. It is but one factor, and perhaps not even an important one, except where there are two species growing together.
Physical differences still seem the most important to me, per the difference between pygmaea in Great Brak and splendens at Albertinia. Are you not making physical differences important when you exclude certain populations like variegata from your ms? I thought that it was lack of physical differences between the plants at Horn Farm and those near Albertinia which led you to conclude that they were one species – which is going at the issue from a 180 degree direction.
The big “problem”, I guess, is really how to deal with the populations in the area which may or may not be connecting links, and which for one reason or another do not alone deserve species status. I just don’t have an answer. But I don’t think that “blooming time” solves the problem either. Rather, it seems simply to smother the issue.
You write in your ms that there are two completely different needs that are met in classification, yet you seem to give up the one, that of collectors, in your conclusions. If we are talking about taxonomy as the basis for communication, are you not giving up the importance of communication in the combinations that you propose? And yes, there may be two different needs, but why cannot the solution for this contain a system that meets both sets of needs?
I am not wedded to Latin binomial names. We have previously shared an idea of the usefulness of including locality data in a haworthia name. Can that be done without challenging the existing classification system? Every population could be a forma, and then everyone would know in a discussion what plants were being talked about. I recognize the objections and impossibilities, but I think that it is a great idea nonetheless. Meanwhile there are not two collectors in the world who are going to agree on a system for the use of collectors.”
B3. response by M.B. Bayer My response was…”Yes I do regret that our minds do not meet. This is the problem with taxonomy – it is simply not science as I once wrote. I have by no means switched to using flowering time in the way that you suggest. I am recognising that better solution to the problem is needed. Cooper Siding seems to flower with splendens and not with pygmaea nor with Vleesbaai acuminata and nor with Humor. It is simply axiomatic that it is a factor which influences gene flow, but this is disrupted by isolation and what botanist can pretend to understand “island” speciation? Variegata,floribunda and chloracantha present a parallel problem to this pygmaea complex and the question of physical differences to the pygmaea lot is barely relevant. Would any haworthia taxonomist suggest they be treated as one?
My experience with other genera suggests that any two people, who truly look at these things in the field and grow them in the way we do Haworthia, will end up in the madhouse if they are looking for agreement. The very point I have been trying to make is that if classification is to make sense and be useful as a communication medium that it is agreement we have to prioritize. If any one thinks they have valid and strong opinions then they should express them, and they should not lose sight of the fact that Latin names belong to the realm of formal biology. What is the point in dissent which is not balanced by counter argument? In a recent journal an author has at least ten new names to occupy the place of my aristata. This is as if the use of the name aristata itself does not have the potential to generate waves according to the intent of the reader. I wrote at the very outset that my intent was a system of names within the domain of the South African flora as a whole. How will we ever get agreement if we do not even see that need? There is a huge trend to turn everything upside down using DNA technology and now I just have informed comment that standard morphological methods work better!
One should not actually express an opinion without recognising the fact that there is a physical record in the form of the herbarium collections (worldwide) which even take precedence over the plants in the field. Unless ones mind covers all those records how can valid can an opinion be? This is the reason why no two collectors can possibly agree – they are confined to the sample they hold in their minds.
You close with what I think is a fine solution. Refer to the populations by place name within a formal structure and it is that formal structure which I am trying to find.”
We did try to continue this exchange of ideas. My feeling was that it would be endless as the collector is not in fact in a position to evaluate or question any comment in respect of biogeography. Understandably he has to lean on the historic method of dependence on the evidence he has obtained from the material he has seen. Another consideration is that my decision is being influenced by what I think about the western populations where I suggest that H. mirabilis absorbs H. maraisiii, H. magnifica and H. heidelbergsensis. In response to his comments I redrafted the manuscript to explain this.
C. Comment by Steven Hammer “If you say that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well, this sounds contradictory, though one could say that farces and comedies are effective. I suppose the problem is that one is unable to take taxonomy seriously in the way its “strict” pretensions would demand. But if it works in its fashion then all one needs to do is to analyse that fashion and say it’s OK for that purpose.”
His opinions are that esterhuizenii is far greener than splendens and lacks the capacity to “blush”. Of Dumbie Dykes pygmaea he writes…”it is different – big pale, flabby readily proliferous (lots of turgida traits). Thus it has echoes of vincentii too. But you can see all of those traits in Great Brak pygmaea too.”
In respect of habitat and substrate…”I think it is likely that the difference has its impact – consider the way both western splendens types have changed after a time in cultivation. Soutpans plants look as it they’d been digesting powdered white porcelain! They did not have this look for the first three years. Much more slowly, some of the Snymanskraal plants have acquired it too, while at first they resembled many magnificas, especially those with atrofuscan aspirations.” He adds that it is possible to imagine the Mossel Bay pygmaea transported to Soutpanskoppies and getting bigger and fatter.
In respect of formalities of nomenclature attaching a reality to the entire spectrum of names which they simply do not have, Steven writes…”I love this last sentence. I can see that each of the other elements can be seen as variant of, or a link to, of pygmaea. There are so many cross-links…”
He adds…
”Where orthodox taxonomy gets in trouble is in its hope for absolutes – we are told, for example, that splendens never has spots on its leaf backs. This is not true when one sees a large enough sample. It is true that most esterhuizenii’s have many more spots, much more often, but it is also true that one of the original esters – a Parisi plant – would have passed anyone’s inspection as a western (Soutpans) splendens. If that isn’t a proof of a connection I don’t know what is! It is also true that Parisi’s plants, and their descendents, can be distinguished from the Venter-Marx plants bred from material collected on the same hill – they have more ‘silver’ and are more delicate. That lineages can be distinguished on such a minute (if trivial) level tells us something. Haworthias are very malleable in a pot. If they are that malleable in a pot they must be malleable/shiftable in habitat too.
That brings up continuity. The gene flow may once have been a continuous flow in one direction, but the river’s dried up. So now we have islands in a dried-up stream. Collectors will always want to call splendens a species – it concentrates all the glamour of Haworthia in one shiny package. I don’t know what rank it “deserves” but I suspect it is the least trivial, or the most distant, of pygmaea’s ramifications. If this is pygmaea then it’s the most deeply transformed of the lot. Or it’s got some emelyae links, which would fit, by extension, the maraisii/emelyae link. I do think that co-occurence logic is the strongest logic possible. One can also look at Soutpans and say, hmm, no trace of turgida here!”
C. Response by M.B Bayer I do not consider that if the system works in the fashion that it does, and that it is OK for the purpose, that this helps. It is only working because too little is expected of it. The scientist/botanist believes that the names are providing critical information about species but which they cannot define, and the collector is content with a handle to attach to his plants or communicate about them. My contention is that taxonomy is not providing proper scientific content.
Illustration
1. 7508. Draaihoek. Northwest of Albertinia and the type locality (the only locality) for var. argenteo-maculosa. The substrate is Bokkeveld shale and the vegetation renosterveld. The locality has always been very restricted, with few plants and very unsubstantial. Although there might have been more suitable habitats available in the past, there is very little remaining.
2. Snymanskraal. splendens. The variant was discovered again almost solely due to the persistence of R.W.Kent althouth the next locality may be where Herre originally discovered it. This habitat is quite complex. It is at the convergence of recent sands, ferricrete/silcrete and shale. The ferricrete/silcrete is in a relatively flat-lying formation rather unlike the raised inselbergs that it forms in the west. The vegetation is largely Renosterveld, but with so much sand in the vicinity, there is also Coastal Fynbos.
3. 6751 Soutpan. splendens. Very similar to the preceding habitat and the plants are also very localized in the exposed ferricrete.
4. 7507. Paulsfontein. fusca. This habitat is also on the geological convergence described but here there seems to be high level river gravel which predominates.
5. GM Aasvoelberg. esterhuizenii. Parisi and Dunn seem to have been the discoverers of this population on the low northern slopes of Sandstone hills to the east of Albertinia.
6. 7102 Welgevonden. vincentii. A few kilometers further east, the locality is also Table Mountain sandstone but a different geological series to that at Aasvoelberg. This and the previous four records are in a very simple geographic line and there really is nothing to suggest any biogeographically differences in these habitats other than the detail of substrate. In the general way that Haworthia populations display some degree of observable continuity from one population to the next, so do the plants in these populations. The actual differences may seem quite large but there are parallels at many places for this ecotypification in the genus.
7. JDV86/75 Cooper Siding. argenteo-maculosa. The habitat is again ferricrete/silcrete on an interface with sandstone. Plants west of the siding are actually in Sandstone. It appears that the plants can be found eastwards along the sandstone ridge to near Humor about 10km eastwards. I do not think anyone has really explored the differences in the plants along this ridge. H. floribunda also occurs at the same site and I have seen a hybrid.
8. JDV91/39 Humor. argenteo- maculosa. The plants are on the low slopes of another sandstone ridge which parallels that at Cooper Siding and apparently a different series. I am not aware that anyone has given these plants an individual name, but would be justifiable in terms of all the other names in the series. The plants are quite strongly marked.
9. JDV87/97 Melkhoutfontein, Gouritz Bridge. acuminata. The original locality was destroyed by road-building and these plants survived in cultivation. Populations are also known to the south and north in the bank of Bokkeveld shale which borders the river on the west bank. The plants are odd in that the leaf ends are very acuminate.
10 MBB6747. Vleesbaai. acuminata. The discoverer of this population may be V. DeVries. The substrate is calcareous limestone. It is a similar formation to that at Stilbaai where H. variegata occurs. The plants tend to be quite robust with a curious grey sheen in cultivation.
11. JDV9/135 Herbertsdale. pygmaea. Enst van Jaarsveld found these plants on a steep Uniondale conglomerate slope with H. chloracantha also present. The plants tend to be a large and flabby and not particularly attractive to grow, whereas in habitat they retain their tight soil-level leaf arrangement.
12. EA1200, 6813. Mossgas. pygmaea. This population was found by Paul Schoeman in sandstone just west of Mossel bay. It is important to note that this sandstone is the same formation that runs from south and east of Albertinia in a narrow band sloping down to near sea level at Mossel Bay. Populations 2, 3, 4, 5, 6, 7, 8, and 12 are thus along this band and its contact with younger tertiary formations.
13. ADH275. Northwest of Mosselbay. pygmaea. The geology is exposed ferricrete with river gravels. Adam Harrower found a plant with seed and grew the seedlings here illustrated. Notable is the appearance of plants with the crystalline papillae known in plants from Dumbie Dykes and from Great Brak.
14. EA1218. Klipkop, North Mossel Bay. pygmaea. The substrate is old river terrace. Etwin Aslander’s locality is now a housing estate. The plants are not much different from those in the previous record.
15. EAsn. Dumbie Dykes. pygmaea. The geology becomes very complex from here eastwards as there is igneous extrusion.
16. Great Brak. pygmaea. Various collectors have plants from here. During the early years I observed pygmaea in four different populations in and near Great Brak. They are still present practically within the town, and a short distance to the northwest where they are in gravels. What struck me forcibly at the time was the similarity to H. mutica and in one close comparison of two plants, I could only separate them on the colour of the bracts.
17. MBB7512. Draaihoek. turgida. I will not illustrate more than four populations. Essentially it is significant that we have a situation which parallels that in the Eastern Cape where H. cooperi is is general the plains-dweller and H. cymbiformis the cliff-dweller (in the west there is a morphing and fusion so that cooperi var gordoniana is the plains-dweller and cooperi var picturata and H. transiens the cliff-dweller. In the southern Cape, the separation has been H. turgida the cliff-dweller and H. retusa the plains-dweller. The relationship is less consistent than is the case in the eastern contingent and the very complex continuity of H. mirabilis with elements such as H. heidelbergensis, H. maraisii and H. magnifica (which can be rationally treated as one) suggests that H. retusa should absorb H. turgida. In practical terms H. turgida is the predominant element, and the name retusa is taken because of the requirements of nomenclatural rules. It must be noted that the complex of populations that I have discussed in Haworthia Update Vol. 2 as H. mutica var. nigra represent a part of the interaction of H. turgida and H. retusa and thus in this light would all be H. retusa.
H.. turgida, supporting the contention that it is the cliff-dweller, occurs in the Valsch, Wyders and Gouritz river valleys on very steep sites. It is proliferous, so the plants are often in large shallow rooted clumps. The leaves do not develop the very flat and truncated end-areas as do the plains-dwellers. As far as I am aware it only occurs at two localities east of the Gouritz River. One at Brandwacht north of Mossel Bay and I believe at one east of Little Brak. Only at Draaihoek is it in very close proximity to argenteo-maculosa. It is at this locality where turgida var. pallidifolia was collected. It is both curious and co-incidental that I did observe one or two very pallid plants at the time of my first visit to the site (now apparently no longer present) and formed an impression that the separationof argenteo-maculosa and turgida was actually a question of ecotypic adaptation. A population I did sample was present about 1km north of Daaihoek in which the plants were more comparable with turgida var. longibracteata and these did influence my impression. That population seems to have disappeared as the area is very disturbed (largely due to the dense hyrax population) and I could not find it on a recent visit.
18. 7105 Ouvloere. turgida. The plants here have the same rather obtuse leaves which prompted me to recognize the variety sub-erecta, but I was also struck by the change in the mottling of the leaves and the similarity to this in esterhuizenii and especially vincentii.
19 ADH397 Gouritz Bridge. turgida. This is the well-known form of turgida which once passed under the cultivar name “rodinii” for which Die Hell, up-river in the Swartberg Mountains,was incorrectly given as the origin.
20 EvJ16716 Schoemanshoek, S. Gouritz Bridge. turgida. This is very similar to the former and again from a cliff habitat. There is of course a similarity to acuminata which is only a few kilometers away on the sloping rocky banks of the river. That does vary from the pygmaea complex where I would include it now, as it also tends to be proliferous. I consider that this emphasizes the tight relationship of H. pygmaea and H. retusa (turgida) rather than supporting any reason to fragment them taxonomically as we have done in the past.
Conclusion If I now am required to produce a synopsis which makes broader botanical sense, rather than meeting just the needs of collectors, I would suggest that it is mutually agreed that only two species be recognised. These are H. retusa represented by the populations cited here by the name H. turgida and with the rider that in fact H. turgida should be incorporated under a wider acceptance and recognition of H. retusa; and that ALL the other relevant populations in the area under consideration should be treated as the single species, H. pygmaea. For the collector I would seriously suggest that the formal ranks of subspecies or variety simply be ignored. The species name is used and in inverted commas any other name that is useful to popular discussion and communication. The formalities of nomenclature attach a reality to the entire spectrum of names which they simply do not have. Names are being used to register collection, acquisition, possession and personal achievement, and are recording very little botanical information and understanding of the extraordinary nature of living things.
A prominent taxonomist was very dismissive of my opinions and cursorily explained that…” Yes, species are real, and defined well by their ability to cross freely and produce offspring which again crosses freely. This has been studied and demonstrated since 1750 or so (you may remember that I talked about this in one of our discussions on the subject), but it is naturally not easy to dive into so much work including following up several generations etc. And since related individuals are similar looking, the reciprocal conclusions that similar objects are related has unfortunately been used as being true, which it is not. And yet, species are the only natural unit in the whole of taxonomy, which to determine is the crucial point. Perhaps this sounds rather like the famous Dicta of Bessey, but after having dealt extensively over 38 years by now with species boundaries in teaching, reading, theory and practice, in different vegetation zones and many different groups, I come back to old definition first given by Ray in 1682: “Group of plants derived from common seeds, reproducing their typical features by sowing” and used by most taxonomists since.”
My contention is that this definition is grossly inadequate as it under-estimates the degree of variability that can occur within a species, not only inherently, but especially in relation to spatial distribution and adaptation. ♦
This chapter is based on recent field exploration and embroiders around many aspects of Haworthia species discussed in earlier chapters. What should be striking is that new populations follow the very predictable geographic pattern that all my earlier exploration has exposed and in my estimation confirm in every way what I consider a sound and satisfactory taxonomic solution and help explain its limitations.
1. Haworthia minima.
Two new populations of this widespread species are recorded:
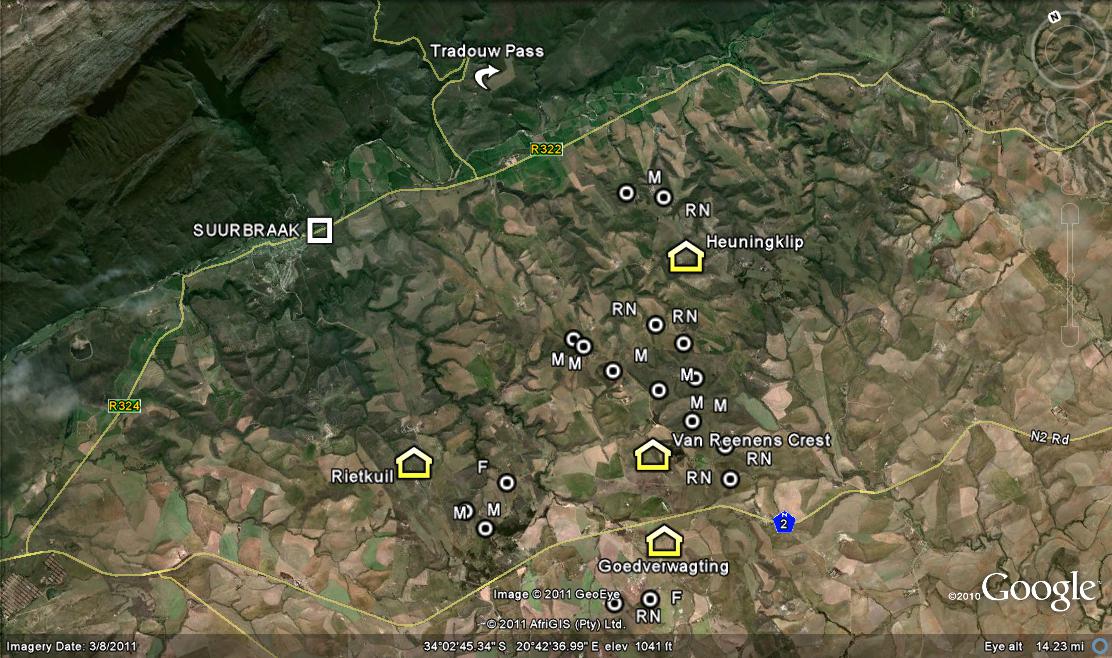
MBB7807 Swartrug, SE Heidelberg. This is a steepish east facing slope. The plants are small, the leaf length barely exceeding 50mm and with a tendency to form clusters in the shallow clay banks that rift the area. There are many plants in quite a small area and it curious to see them within very large clumps of Euphorbia tridentata. This Euphorbia itself has a strange geographic distribution being known at Riversdale, Calitzdorp and then at Cradock in the Eastern Cape. (see Figs 1)
MBB7813 NW Skeiding, E Tradouw Pass. This is a high rainfall area and we were expecting to find H. mirabilis in the lower pressure bursts and white clay that can be seen from the farmhouse far away. But instead we came across H. minima at the highest point of the hill among sparse grass. They are rather green in colour and it is interesting to note in the one plant pictured that it has non-tuberculate upper leaf surfaces and white cartilaginous margins and keel of the leaves. (see Figs 2)
2. Haworthia rossouwii.
MBB7803b Morning Star, S Heidelberg. Morning Star was known to J. Dekenah but the area has been very developed since those years. It was very surprising to find this small form in abundance at the top of a high hill in ferricrete pebbles and virtually no vegetation ground cover. However on this subsequent visit we crossed through an internal farm fence and found the plants in abundance about 150m further away and over quite a large area. We later looked at a lower promising hillside and small area nearer the Morning Star homestead and found the plants again. (see Figs 3)
Looking from Morning Star to Kransrivermond 097
3. Haworthia mirabilis.
This item reports on 6 populations, three of which are new. They confirm the continuity down the Duiwenhoks River and also southwards and westwards from east of Riversdale through to west of Robertson and Greyton and Napier to the south of that. Attention is drawn to south of Heidelberg where the ‘enigma’tic relationship of this species with H. retusa is suspected.
MBB7808 Swartrug, SE Heidelberg. This farm is well known as the source of ‘Haworthia chromatica’ that I dismissed as an element in the complex surrounding the e H. retusa (mutica) ‘nigra’. The names Droekloof and Doringkloof have been used while the map reference gives Diepkloof for a large area extending to west of the Duiwenhoks River. The plants are on an east facing boulder terrace in a stony low bush site with some grass. The plants are small and show the expected wide range of variation. They are fairly similar to the plants in the lichen patch just east of Heidelberg that I have always mistakenly taken to be the origins of H. heidelbergensis, but here are subtle differences that mark a population difference, but I will come back to this. (see Figs 4)
Looking from Swartrug towards Riversdale.
MBB7809 Koeisekop, SE Heidelberg. This is on the same farm and a similar ferricrete hillside southwards. There are several hilltops by the same name in the general area and I cannot even speculate on the origins of the name (“cowshead”). The plants are larger than in the previous population and very abundant. The similarity to populations in the Swellendam and Riviersonderend area is in my opinion striking. Odd clones are miniature images of H.mirabilis from the southwestern populations west of Napier. (see Figs 5)
MBB7220 Morning Star, S Heidelberg. I have discussed this population before and I repeat it here because I did not have many field photographs. It is only 300m from the Morning Star MBB7221 H. retusa ‘nigra’’ population and they flower and seed virtually in unison. Therefore they probably also reflect on the strange interaction between the two major systems. What I have surmised is that H. floribunda gets absorbed into H. mirabilis at the southern limits of its distribution and in these pictures there is clear evidence of the rounded and flattened leaf tips of that species. Therefore we may actually here have a three way species interaction. Reinforced by the presence of three different things at Kransriviermond, covered later under 4. H.retusa ‘turgida’. (see Figs 6a and b)
MBB7221 Morning Star
MBB7811Kransriviermond, S Heidelberg. Populations further down the Duiwenhoks river have been reported on and this one fall within that mould and the same one as the previous Swartrug and Koeisekop populations. The population is smaller and the plants too are quite small. (see Figs 7)
MBB7814 Skeiding, W Heidelberg. This area is also reported on but digital photography makes it so much easier to record the enormous variability that such populations exhibit. Some of the plants are typically ‘atrofusca’ while others have leaves ended in an awn. Leaf surface may be smooth and there is even an indication of smooth leaves. What is notable is the absence of H. floribunda from what is the sort of habitat it can be associated with and can one can speculate its absorption into H. mirabilis with leaf shapes that demonstrate this. H. floribunda does appear again about 15km to the southwest in association with H. retusa and in the absence of H. mirabilis. (see Figs 8)
The Skeiding homestead
MBB7233 Die Plotte, Heidelberg. “H. heidelbergensis” correctly has its origin in this near vicinity and these small summer flowering elements are certainly part of a large assemblage of populations in the central area of what I now come to see as all H. mirabilis. Here discussion must pass to the following. (see Figs 9)
4. Haworthia retusa‘turgida’.
A difficulty that I experience is in the need to dismiss the impression and general idea we have that there are tiered ranks. H. retusa has many populations and past treatment has placed most of the variants in H. turgida. To suggest that these are really two separate systems is not realistic and confounds any explanation for a few anomalies that exist.
MBB7810 Tierkloof, SE Heidelberg. Tierkloof is quite a deep valley that pushes out of the Duiwenhoks to the east. We found this huge population of large clump forming plants and several such populations are known just north of Heidelberg itself. I have applied the epithet ‘longebracteata’ to these and this is no more satisfactory than generating names for each of a very wide range of populations. There are populations along the Duiwenhoks that are far more inclined to the ‘turgida’ element and Essie Esterhuizen wrote a very good account of the host of variable populations that he encountered. (see Figs 10)
H. retusa ‘turgida’ at Tierkloof, Swartrug
MBB7812 Kransriviermond, S Heidelberg. We only saw three plants at this locality and these plant mirror the preceding, if a bit smaller. The problem here is that we also have the element I myself described and named as H. retusa ‘mutica’ and I originally linked this “variety” with H.. mutica following what G.G. Smith had done and for predictive reasons. The fact that ‘turgida’ now occurs with it is a huge hurdle to cross. Think carefully. (see Figs 11)
MBB7804 Kransriviermond.I have written about his population in the chapter “Haworthia enigma”. This explains that the plants are not all leaden-black as perceived. They are spring flowering – as ‘turgida’ is. However, the range of forms and colour virtually replicates the plants in MBB7721 at Morning Star (pictures in the “enigma” chapter) where they are spring flowering. Across the river from both these populations is a weak population of small plants of H. mirabilis (7811 above) summer flowering. We know that this major difference in flowering season is not a total barrier to hybridization. (see Figs 12)
MBB7234 Die Plotte, Heidelberg. This is a population of large clump forming plants on a north slope about 300m south of MBB7233 H. mirabilis ‘heidelbergensis’. It is spring flowering. When I first saw these plants I dismissed (actually I never dismiss anything – I maintain reservations as I do now) these plants as large variants of ‘heidelbergensis’, and actually collected one magnificent specimen that looked to me just like H. mirabilis, say, from west Napier. Looking at the range of variation one is compelled to concede that here again is an infusion of H. mirabilis into a population that is primarily H. retusa in that vast shadowland it casts with ‘turgida’. (see Figs 13)
Conservation I add these comments for several reasons surrounding the poor image that collectors have in the eyes of conservationists, and the very rickety fence I therefore occupy. There is a note reprinted in a recent issue of the newsletter To the Point. This taken from Times magazine and seems to be a rather hyped up condemnation of widespread destruction based on a view of internet websites. These are also monitored by RSA law enforcement that is rightfully disturbed by the high number of locality and collector citations that are indicative of illegal collecting. I personally no longer apply for a “collecting” permit for the prime reason that I am concentrating on photographic record, as a herbarium record no longer serves any better practical short term purpose. The herbaria are overburdened and I prefer living plants to dead ones. After 70 years of field activity I feel treated like a pariah and reduced to subservience under less-informed inexperienced people operating under a rigid and oppressive legislation aimed at acquiring a prosecution rather than wisely trying to direct activity in a positive direction. This activity of collection is, and I have no doubt about this either, a human right imbedded in the very reason for creation and our place in it. Conservation has its roots in the passion that people have for nature and it is nurtured by experience, contact and knowledge of its varied components. It is just ridiculous that one can go to any animal or plant centre here in South Africa and see plants and animals from anywhere else in the world and none indigenous. It is Ok to, say, keep Madagascan Chameleons in captivity where it is someone else’ responsibility to secure them. But keeping local ones is taboo? It is OK to freely sell cacti that do not occur here but are now being dug out in tons in (if that is really true). Recently it has become illegal to so much as possess Encephalartos latifrons and even the National Botanic gardens can no longer propagate and sell this plant. While we talk “plant indigenous” on all public fronts, the fact is that the acquisition, cultivation and sale of indigenous plants is viewed in the most negative light – a viewpoint encouraged by conservators.
This is control by law, a large degree of pure ignorance and by improper application of power derived from legislation. This is not the way to go. It is quite evident from the Times magazine article that legislation has the negative effect of creating a false value and glamour to discovery, Latin names, acquisition and possession. I believe that knowledge and understanding is true power, not legislation and enforcement.
The collecting numbers, localities and information provided here is to promote understanding and knowledge of plants, and thereby to promote their continued existence against all the vagaries of nature that “threaten” them, least of it which may be actually human greed.
Acknowledgement I am glad to acknowledge the company, comments and interest of Gerhard Marx and whose organization sceptisism I also value. Landowners always impress me with their generosity and graciousness in allowing us access. On this occasion they include Mr. and Mrs. Nelie and Wimpie Jacobs of Diepkloof, Mr. Cloete and Belinda Fourie of Melkboom (B&B), Mr. and Mrs. Coetzee and Sarita Uys of Morning Star, Messrs. Christo and Pieter van Deventer of Doornvlei, Mr. and Mrs. Neels and Anne-Lise Uys of Skeiding (B&B) and then Ms. Jennifer Steggie of Heidelberg. In view of the comments on conservation noted above, I note that there are several farms that now offer accommodation and an outdoors experience and the opportunity to view these plants in their extraordinary natural surroundings. ♦
My experience with Haworthia dates back to my childhood and on to nearly 70 years of observation. However, my interest was only able to properly manifest when I began work at the Karoo Botanic Garden in 1969 and it has since been through many phases. I wrote a formal taxonomic revision of the genus in 1999 and have spent a good bit of the last nine years adding to and verifying what I wrote. Haworthia has always been regarded as a problem child of botany to be avoided by professional taxonomists for various reasons including an apparent phobia of the many amateur collectors peering over the shoulder while at work. This has puzzled me because it seemed to me that if the need for good classification and identification was so strong there was an obligation on botany to provide the service. So my involvement has been largely by default. I was trained in an agricultural and entomological tradition with a totally different and unsophisticated approach to things like taxonomy, systematics and nomenclature. In the infant science that agriculture then was in South Africa, I can barely claim that my MSc is much more than an indication that I tried to learn something beyond normal schooling. While trained as an agricultural entomologist, my leaning was to plants and I eventually came to the Karoo garden to do what I liked best viz. exploring plants. Unfortunately the route is via identification and names and so I have walked a long road through the minefield that this is. Was this only in respect of Haworthia? No! This is a persistent misconception. Haworthia is only different because it has attracted such close and sustained amateur interest by so many for so long. I experienced failing classification in many other genera. To be fair I think the real reason is the lack of importance attached to the whole function of plant classification. It even seems as if many modern botanists pursue the study of plant relationship under the guise of systematics that is not committed to providing formal names and identifications.
The problem in the genus Haworthia begins with weak botany from the moment Linnaeus decided that there were four small plants that belonged together as one species in the genus Aloe. This alone generated a nomenclatural problem the dust of which is still to settle. A worse and obscured fact is that because of their small flowers these plants were and are still treated as one genus – one set. This is not true. The allied genera of the Alooideae of the Asphodelaceae are not comfortably classified and the three sets of plants that constitute the genus Haworthia are not only florally distinctive, they are also phytogeographically and behaviorally different. Thus while there was or is this huge flaw at this classification level, it is actually bizarre that sense can ever be thought to have reigned at the species level. The proliferation and confusion of names coupled with the associated argumentation about names has even led me to doubt my sanity, or even lose it as my critics will happily claim. My impression now as a non-botanist is that taxonomic botany somehow lost sight of the requirement that a biological classification should mirror and reflect the evolutionary processes or phylogeny that underlies our view of species and their origins. Worse, is the actual absence of a clear and unsullied definition of what a species is and hence what a Latin binomial as a supposedly formal scientific term of reference actually means. Cladistic methodology in plant taxonomy is in my opinion nothing more than a belated and even misguided attempt to arrive at or return to some semblance of scientific truth. Molecular biology is now being desperately seen as a holy grail that will summarily correct reluctantly conceded failures of the past.
There is no doubt that the mess in the classification of Haworthia is the product of non-botanists, but it is absolutely incorrect to exculpate the contributory role of taxonomic botany and professional botanists who are or were taxonomists. My interest has by no means been confined to Haworthia and the need for names has largely been driven by a perpetual curiosity about many different plants in the field environment. I am, by the route my life has taken me, unusually and almost uniquely experienced in the relation between botany, botanists and interest of the layman in plants, as it is reflected in the names we use for them and how we respectively do so. Consequently I think that I am now qualified to express some opinions on what Haworthia can teach us quite apart from what the hostilities that wage across the whole taxonomic front suggest.
I pointed out rather timorously that in Oxalis, morphological characters were not as indicative of species as revisions and identification keys generally seem to imply. The dramatic changes in our classification systems that the results of molecular biology now seem to threaten, is a manifestation of this simple fact rather than any great new understanding of what really goes on in the plant kingdom. My fear is that molecular biology is likely to fudge most of its promise because the basic tenets of classification, the mistakes of the past and the reality of the field are still not properly appreciated. This is what I think Haworthia has to tell us. I made use of Oxalis to test my observations that we can look at Haworthia as composed of elements that reflect the very nature of the habitats and environments where they are found. I did this because Haworthia plants simply do not have any array of characters by which species can be arrived at whereas Oxalis has a plethora of such apparent (taxonomically bewitching, tempting, and misleading) means. While the few facts that there are in respect of differentiating Haworthia into three groups (subgenera as presently constituted) has been lost and confounded in the broader classification of the Alooideae there is not much else to help verbally to define the species. It is not to say that modern technology has not been resorted to, to help clarify the issue. Both cladistic and molecular studies have been done but the results flawed by weak hypothesis and perhaps just weak science too.
How then do I think sense can be made of Haworthia? It is firstly by defining the species as dynamic fractal systems that illustrate the responses of plants to skeletal habitats and geological substrate in the wider context of landscape and topography across the total distribution area. Furthermore, there is an obvious correlation to be made with vegetation and other biological systems both plant and animal. I cannot pretend to know a great deal about any of these things, but also suggest that nothing more is needed other than awareness that this is what biological diversity and species are essentially about anyway.
It has taken me nearly 40 years to arrive at the big picture of Haworthia. It is as a non-taxonomist that I say categorically that the three main elements in the present genus need to be seen and registered as “different”. Then we can focus on the subgenus Haworthia where the greater difficulty lies and which I believe offers the greatest insight into what plant species are and what the difficulties are in recognizing and defining them. Also into what Latin binomials should or might mean.
For this I am going to take just two species systems and ignore some of the peripheral facts that might confuse the issue rather than illustrate the profound nature of species interactions. I take H. retusa (Figures 1 to 4) and H. mirabilis (Figures 5 to 10). In order to do this I have to bypass even my own conservative classification by considering H. turgida and its many variants to be the riverine and cliff dwelling version of H. retusa. H. mirabilis is more complex and I have to include three other species viz H. maraisii, H. magnifica and H. heidelbergensis for which spatial factors and major geological substrate are the prime differentiators. Both my two selected species are highly variable both within and between populations. It is in fact the intra-population variability that I underestimated. Growing plants from field collected seed can result in a hundred or more individual plants that are all different in appearance.
The two species share the same distribution range, but with an extension of H. mirabilis into the Worcester/Robertson Karoo, while H. retusa extends further east from Albertinia to Great Brak. H. mirabilis flowers in late summer and H. retusa flowers in spring. They never grow in direct association! The populations of each are highly localized and when in proximity are usually separated by hundreds of meters, and difficult to ascribe to habitat conditions given the ranges of habitat that respective populations may occupy. I have already noted and warned of the reality that there is evidence of interaction between these two clearly defined systems. In addition, somehow or other, the species H. pygmaea emerges as discrete in the eastern Mossel bay area, while H. mutica sits in an island between Swellendam and Riviersonderend in the west.
What is most striking is the variability within each system and this is where the crunch comes and what my pictures should illustrate. They are but a small sample of many hundreds and I have adopted an informal method of captioning the pictures as the formalities of nomenclature would need to be followed to depart as I have from the existing status. Not only are my pictures supposed to illustrate the problem of variability within species, figures 2 and 3 show variability within a population and figures 3 and 10 show two very similar plants from two quite different species systems. Note that the plant in figure 3 is in flower and that the similarity to figure 10 is rather exaggerated by photograph scale as in H. mirabilis ‘toonensis’ the plants are smaller.
Taken individually from the morphological extremes between and even often within populations, it will stretch the credibility of the most ardent taxonomic lumper to claim that any two of these are representatives of the same species. It is an unpalatable truth that things that look different are the same just as the converse is that things that look the same are different. When this reality is properly conceded by the professional taxonomist, perhaps we will succeed in demystifying and deglamorising the Latin binomial. Botanists who are not taxonomists will arrive at a better understanding of their plant world and consequently we who look to them for understanding will be better served. We need to be more realistic about plant description and identification and also correspondingly more conscious of the real nature of diversity. Conservation is not to be seen as maintaining a list of names for which there is a use. It is to be seen as maintaining the options for change and the conservation of a far greater diversity than any list of names can hold. It is botanists (quoted in Levin, 2000) who wrote “our system of names appears to achieve a reality which it does not in fact actually possess”. I think this implies that even the Latin binomials we so modestly, reluctantly or even portentously use do not convey what they should; but rather the product of our individual limited experience, knowledge and perception. My feeling is that species do have a reality and for Latin names to have any sense, they need to be more circumspectly applied. There is little merit in having a highly regulatory nomenclatural code and little value in what the names actually refer to. ♦
In Haworthia Update 4 I wrote an essay about the haworthias east of Albertinia in which I discussed their relation to H. retusa and H. mirabilis, while generally lumping them largely in H. pygmaea. There are of course real ‘turgida’ populations as far east as near Mossel Bay, so I argued the case for an interplay of the two former species that over the whole distribution range generated two ‘species’ in the east viz. H. pygmaea and H. retusa (to include ‘turgida’), and three ‘species’ in the west adding H. mutica to H. mirabilis and H. retusa.
Recently I had the pleasure of meeting Gregory Nicholson who is studying botany at University, Cape Town. He surprised me by telling me that there was a Haworthia on his parent’s property west of Herbertsdale. It was not in fact so much surprising as confirmation of the belief I formed on a trip a short while before that there must be haworthias in the very suitable terrain of the Jakkals River valley 6km west of Herberstdale. The surprise came when Greg indicated the position of the plants much deeper into the mountains.
We visited the site while exploring H. emelyae north of the Langeberg Mountains and I will report that in an additional short article. We over-nighted east of Barrydale and a puncture made us detour via Riversdale rather than take the northern dirt road. It was raining heavily at Albertinia, but we could see sunshine on the Langeberg in the direction of Herbertsdale. We traveled the dirt road through the upper Gouritz, because we also wanted to gauge the condition of the gorge through the mountain, as Greg reports the presence of H. retusa ‘turgida’ some distance in. Towerlands is some 8km on and it was still just sunny when we arrived. We walked immediately the several 100m to Greg’s site along a good path among true Fynbos vegetation and found the plants very easily in a ferruginous sandstone ridge sloping into the river. Then the rain came and made picture taking a bit difficult (see MBB7489 figs. 1-16).
The plants are quite small and there was not much clustering. Unfortunately the weather restricted us so we could not look for better habitat around the corner where Greg had actually pointed. Greg’s photographs had made me lean to H. pygmaea and indeed I do not think one can deny the reality of this if one also considers the odd plants at Herbertsdale itself (JDV97/135) that I assign to H. pygmaea. However, the similarity to ‘turgida’ is just as striking and Daphne and I concluded that we could recognize similarities to ‘dekenahii’ and even ‘mutica’. To make it even more confusing, I felt, and still do, that the plants had similarities to H. emelyae. This latter was confirmed in my mind when we continued our trip via Cloetes Pass back to the Little Karoo to just west of the Gouritz gorge and a population of H. emelyae known to us from Gerhard Marx. I will submit that in a subsequent short note.
I include a habitat scene (fig. 1 – the arrow marks the locality) and note that looking south east one can see Albertinia and Aasvoelberg in the distance. We know that ‘turgida’ is present in the sandstones of the Langeberg, but this find suggests it is far more abundant and widespread than present records suggest. We need to now what the plants in the upper Gouritz look like.
Acknowledgements Gerhard Marx has been extremely helpful and informative and I am most grateful to Gregory Nicholson for his contribution. ♦
The writing of my grand finale was inspired by several things. One of these was another item of a mind-numbing foray into the classification of Haworthia. So I asked that deep thinker and observer, Gerhard Marx, for a devil’s advocate (abbrev. DA) point of view which he has done with the same competence he has as an artist. I have many times in my writing addressed the issue of a species definition and produced one too. Not surprisingly the first thing the DA does is dismiss my definition without producing one of his own. Simply being able to say that an indeterminate number of plants from some population are sufficiently different in respect of a character or two from other haworthia, is motivation enough for the generation of a new name?
The case of H. groenewaldii Breuer, described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald is the case in point. It presents the description of this supposed new species from Buffeljags east of Swellendam. The article is written in the first person (Breuer) who quotes extensively from Gerhard’s e-mails, and includes a piece by Jannie Groenewald under the heading “Description of the Vegetation type and distribution”. The overall impression is of an article that conforms to the style of a forgotten era and it is not possible or sensible to attempt a rational dismissal. Who is actually responsible for the article and how does one correct misleading statements without giving offence?
There is probably only one main issue that can be made level-headed sense of. This is the statement in the article…”another (it is not clear what the other was) striking feature was the fine but rough leaf texture…” Then quoted again…”the typical H. mutica grows only 20km to the west at Dankbaar”. But the Dankbaar plants also have the same shiny leaf surface. The word “typical” is generally overused in the article and I would counter claim that the Dankbaar plants are typically H. mutica and reinforce my argument for the similarity to the Buffeljags plants. But this is not the main issue, nor is it the least of many rectifications the article almost demands. In the new-age freedom of communication there are virtually no secrets anymore and this adds to my discomfort because the article only touches on some of the issues clouding Haworthia classification. The DA says I should meet this following argument in support of an H. groenewaldi’…” it is consistently smaller than H. mutica and reminds superficially actually of H. bruynsii or springbokvlakensis with its rounded leaves. Kobus Venter’s feeling was that it is closest to H. magnifica var atrofusca and I agree 100%.” These are subjective statements and I cannot agree about the size as I have seen many H. mutica populations that I do not think supported the opinion. The roundness of the leaves is also subjective. The element H. mirabilis ’atrofusca’ in any case has plants with variably rounded leaves, so which do you mean?. The name ‘mutica’ means leaves without a point and rounded and it is no co-incidence that Col. Scott initially identified H. springbokvlakensis as H. mutica until I pointed out to him that I thought H. mutica was actually the plants (note plural) in the Bredasdorp area. Scott plagiarized my words describing them alongside old wagon tracks that suggested their discovery in earlier times.
DA lists several more indeterminates…”all the unique morphological features like small size, rounded rough-textured and uniquely flecked leaves”. Of course there are not several more as I have already dismissed two and this leaves the third viz. the “uniquely flecked leaves”. Firstly not all the plants have flecked leaves and I have shown that these occur in H. mutica at Klipport too. The main issue is flowering time and I agree this is problematic. But this is actually so in respect of the whole Southern Cape Haworthia complex where it is quite evident that there are two flowering periods, summer and winter and that despite this difference there is hybridization. What happened in the past?
DA writes…”MBB 7801 flowers…with the typical thin tender peduncles and delicate flowers identical to that of H. maraisii and some H. magnifica and H. mirabilis. Normal H. mutica flowers in spring (Sept-Oct) and has more robust peduncles and flowers.” This word “typical” again, does not convey the truth. There are no floral characters by which one can separate the three “species” the sentence suggests. I have also seen small H. mutica (Crodini) with very small weak inflorescences and comment on this as a general variable. See fig. 149, a recent picture of peduncles of H. mutica from Grootvlakte, Riviersonderend. There is a population of H. mirabilis (I presume Breuer and Marx would use the name H. magnifica var. magnifica) from south Riversdale MBB6651. The plants are very small and cryptic and when I collected seed it was from small slender inflorescences with 3-4 seed capsules. In cultivation I raised a large number of plants that were all different and some were huge. The inflorescences were robust as well with 15 -20 capsules. So this reference to floral characters is simply a loose statement and I do not believe that a proper overview of the flowers in H. retusa and H. mirabilis will reveal any dramatic differences. I have taken good note of flowers and in particular the strange fish-tail bud-tip that characterizes the larger set of populations in the southern Cape.
DA states that I… ”made several blatant errors by ignoring the importance of flower features in his studies”. It this one supposed error or several? I have not ignored flowers at all and it is a blatant error to say so. I simply did not and still do not find them useful. It is fraudulent to suggest otherwise. That fish-tail bud is highly significant and I made a great effort to quantify the differences between H. reticulata and H. herbacea. That the attempt failed was only because the sample required to get to statistical difference became too large. Nevertheless I then tackled a good number of H. mirabilis populations that also have the same bud tip. I concluded that there was a difference between the populations south of the Langeberg and those north and then had to discard that as erroneous because the same problem of variability occurred as with the rosette of leaves. Even the general flowering times I gave for the area was quite wrong – flowering time ranged from November to late April. It is odd that the fish-tail bud tip does actually influence the tips of the upper tepals in that the margins tend to be flattened together. But there is a huge difference in that in H. herbacea and H. reticulata the upper tepals are held far out from the mid-axis (hence ‘subregularis’ or ‘cowboy-hat’) while in H. mirabilis the tepals may meet above the middle upper tepal.
More by the DA… ”So, only careful observation and comparison of diagnostic features, starting with flowers, is the solution.” This is a weak sop because it is something that I started to do and found quite unhelpful. The history is that the small aloid genera were virtually recognized by the fact that the flowers were small. The differences between the subgenera were dismissed by Dr L.A. Codd with the statement that “small differences” were being ignored. DA overlooks a single major binding factor between H. retusa and H. mirabilis viz. bud similarity. I have also discussed in detail the whole question of diagnostic features and went to the genus Oxalis to substantiate my observations that small detail is not helpful. It comes back to the issue of what is a species? I maintain that they are not these artificial elements circumscribed by morphological detail. They are systems set in a geographical framework. The species, whatever they are, have surely not arisen from independent sources. My field work is very extensive and I have shown over and over again that the populations are linked so that when one finds a single population like at Buffeljags, one asks how it fits. It is not any misconstrual by G.G. Smith that a sample from a population south of Heidelberg was likened to H. mutica. In his time it was not known where H. mutica even occurred and hence he made a mistake in describing H. otzenii. DA is defending a methodology that failed miserably and is not going to provide any solutions in the future.
Well, well, well one might say. The DA maintains that Bayer has lost his way, to support an argument that he had stopped “doing taxonomy”. DA is not strong on logic but it is intended that way and neither are the DA’s observations necessarily correct. DA writes this … “Bruce’s dilemma is that he spent a lifetime and hundreds of thousands of Rands doing his research and while he did indeed do an invaluable contribution, he is now faced with the fact that he can’t do that full and neat integration of it all because 1) there is more variety in the wild than what his dispensation recognizes and he feels overwhelmed by the idea of having to try and do a real truth-reflecting revision at this ripe age and 2) he made several blatant errors by ignoring the importance of flower features in his studies.” Point two I have already dismissed, but DA cannot be serious when he suggests that there is some kind of dilemma involving money and time spent. This is an issue that I will have to address when I meet the Angel of Death.
I do recognize that there is great ‘variety in the wild” quite contrary to DA’s claim that here is more variety than what my dispensation recognizes. The main gist of my writing has been this extraordinary variation. I am not overwhelmed by it at all and I maintain that my dispensation is the only way to rationally express it. To say it confuses me is not an overstatement because it is clarification that I have been seeking. Now Steven Hammer puts it very well… ”it’s almost as if you are being blamed for nature’s complexities” and I said the same thing in different words long ago. The Latin binomial system fails and I have argued and shown evidence for the need of a new and different model What is more I have proposed such a model too. Anyone who contemplates at any age, the “truth reflecting revision” the DA demands will be faced with a daunting task of citing the synonymy of all the species described or recognized by Breuer and Hayashi (let alone cite all the collecting records). That alone could occupy a lifetime. Not only that, but to also cover the usages and references by the many authors who have written about the plants would be a humungous task. I suspect that DA is implying that I have become senile with a “ripe old-age” comment. My response is that at least my senility was preceded by some degree of clarity in contrast to what I am forced to conclude about contemporary Haworthia taxonomists! I see no reason whatsoever to depart from a logical argument about species definition and the significance of geographical distribution from my species list of February 2009. Effectively this is a revision and it just does not have literature and specimen citations that meet the needs of “a full and neat integration”. Let me state categorically that there is not the slightest hope of such a pie-in-sky solution.
So we ask Lawrence Loucka’s question… ”How best to separate facts and knowledge from personalities and ego?” First present the facts. I want to add to the information that I presented in my article on “Haworthiaretusa ‘nigra’ – A grand finale”. There I discuss and illustrate plants from the wider general area of Buffeljags and especially from the Tradouw area to the northeast. Covered are four populations from the farm Heuningklip, and three from the farm Rietkuil. Here I want to discuss and illustrate eight more populations from the farm Van Reenens Crest that lies between. The gist of my argument is that classification of Haworthia is far beyond this nonsensical and simplistic generation of new names in exactly the same way that a miasma of names was generated in the years before 1948.
It is not just chance that the area I have recently been exploring has been so neglected for so long. It is a high-lying and fairly featureless area and very grassy in comparison to the Karoid Broken Veld, Karoid Valley Bushveld remnants, and the rocky hills, streams and river beds of the areas north east and west. The vegetation is a grassy Renosterveld and the species diversity seems extraordinarily low. There is not much there to attract a succulent plant enthusiast and explorer compared to far more enticing and rewarding countryside elsewhere. My exploration there is a direct result of my having exhausted most other options available to me. An added consideration is that of prediction when long experience has proven that connections between populations exist and need to be found.
So it is not surprising that we found 8 new populations, 6 in the context of H. mirabilis as I circumscribe it, and 2 for H. retusa ‘nigra’ in the same vein of personal circumscription. These populations were all new to me and very exciting. Nevertheless I would not dream of seeing them as new species anymore than I did for H. mutica ‘Buffeljags’. The Van Reenen’s Crest populations fit a pattern and need to be considered in the context of the bracketing populations at Heuningklip and Rietkuil discussed in the preceding chapter. The collections and illustrations are mapped in an accompanying map and are identified as follows:-
The populations.
Set 1 – H. mirabilis. The plants in the following six populations are collectively different from those at either Rietkuil or Heuningklip. Using the erroneous or absent logic that characterizes the description of “H. groenewaldii”, there is no doubt that at least one new Latin binomial should be generated. Far from being confused by the huge variation I have seen in Haworthia and guided by a systems view of species, I see these as being representative of one species. It would be useful if proponents of some other dispensation take these pictures individually and arrange them according to the characters they so diligently use to denote their many different species. The question of flower character I laugh off because I have tried to use it and know it will fail. I will come to flowering time again. Plant size is barely significant because in such clay soils the plants do not grow to the size that they will in cultivation. Generally the plants in these populations fall in the category 35mm to 800mm diam. But it is difficult to gauge age and the direct effects of niche favourability that will favour growth and size.
1. MBB7914 H. mirabilis, Van Reenens Crest See figs 1-19. This was a small population as can be judged from fig.1 where the pressure burst and exposed white clay area barely exceeds 100sq m. The plants were only on the perimeter of the area. This is what makes it quite difficult to assess what the population structure may have been in the era before farming and cultivation. Very seldom indeed have I found plants anywhere in pristine arable or even semi-arable landscapes. Fig. 2 is a view looking across to Heuningklip and the locality of MBB7896 H. retusa ‘nigra’.
Fig. 1 View of locality MBB7914.Fig. 2 View from locality MBB7914 to Heuningklip.Fig. 3 7914 H. mirabilis
2. MBB7915 H. mirabilis, Van Reenens Crest See figs 20-27. Fig. 20 is a view looking south to a very large area that we did not explore because we chose instead a similar valley immediately beyond the one pictured. The MBB7915 locality was rather odd. In fact the whole exercise was in the respect that we kept being reminded how similar the situation was to Kiewietsvlakte where H. mirabilis and H. retusa play out a similar show. In this case there was no actual pressure burst. In this case there was less white clay and there were the characteristic platform-like ridges as if made by grazing animals. There were few plants and while rather scattered, were still confined to what I can only describe as low-biomass bearing area i.e. shallow gravelly soil that does not support dense vegetative cover.
Fig. 20 View from locality MBB7915.Fig. 21 7915 H. mirabilis.
3. MBB7916 H. mirabilis, Van Reenens Crest See figs 28-48. Fig. 28 is a view of the white clay in an area very like that pictured in Fig. 20. Very heavily utilized by grazing animals, it is surprising that there were so many plants. The variability is quite evident from the pictures and some plants stand out as very different.
Fig. 28 The clay soils in which the plants often grow. Locality MBB7916.Fig. 29 7916 H. mirabilis
4. MBB7917 H. mirabilis, Van Reenens Crest See figs 49-70. This was a dramatic population because the plants were not quite where we expected them to be. Fig. 49 shows a more typical habitat in the distance. Here the plants were rather in a very brown gravelly situation and absent a short distance away where there was more white clay. As for all these populations there was a wide range of surface textures and also leaf shapes and endings. Often the leaves had a very distinct mucro (end-awn or point). Equally often the leaves were without any point and rounded and this was not true for all the leaves on any single plant. It is very problematic to make statements about the plants because when we were looking at Rietkuil plants we were reminded of a host of more easterly populations and plants. In this case even variants of H. emelyae came to mind.
Fig. 49 View from locality MBB7917 to that of MBB 7919.Fig. 50 7917 H. mirabilis.
5. MBB7918 H. mirabilis, Van Reenens Crest See figs 71-87. This population was unusual in that the plants were generally small and proliferous. Sometimes there were as many as 20 rosettes in a clump.
Fig. 71 7918 H. mirabilis.
6. MBB7919 H. mirabilis, Van Reenens Crest See figs 88-116. One needs to refer to the map to see how the populations now relate to one another. These were again very variable as I have come to expect in the whole H. retusa/H. mirabilis collective – to coin yet another adjective to do what I do not think the formal system allows. Note carefully fig.100 in which a young bud is clearly visible.
Fig. 88 7919 H. mirabilis.Fig. 100
Set 2 – H. retusa ‘nigra’ We found just these two populations.
7. MBB7920 H. retusa ‘nigra’, Van Reenens Crest See figs 117-137. These plants were in full flower and setting seed as one expects for the summer-flowering H. retusa elements. In fig. 128 a plant is shown with well-developed end-awn and fig. 134 a plant with rounded leaf tips. Fig. 137 is simply a view of the flower that does not even depict the more normal fish-tail bud tip – see next!
Fig. 117 7920 H. retusa ‘nigra’.Fig. 128Fig. 134Fig. 137
8. MBB7921 H. retusa ‘nigra’, Van Reenens Crest See figs 138-148. Fig. 138 is a plant that could have come from an H. retusa ‘retusa’ population from around Riversdale, while Fig. 146 could have come from H. retusa ‘turgida’ from somewhere around Heidelberg. Fig. 147 clearly shows what I mean by the fish-tail buds. Fig. 148 shows that there has been quite extensive exploitation of the white clay (kaolin). Fortunately (?) unlike the bentonite that is mined on large scale, the kaolin pockets are often small and restricted and apparently not commercially viable.
Fig. 138 7921 H. retusa ‘nigra’.
Conclusion The plants in the above populations cannot be categorized without reference to the totality of populations and I will try and summarize my point of view. These populations occupy a position very central to the distribution of both H. retusa and H. mirabilis. I need to quote DA again who wrote … ”His (Bayer’s) emphasis is upon phylogeny and understanding the evolutionary processes at work in Haworthia and he loves getting involved with very complicated arguments and theories which (frustratingly or delightedly) to him nobody quite follows or is really interested to follow.”
DA is most fortunate to be able to dismiss the basis of classification and what all biologists involved in classification try to do. It is quite wounding to say that I enjoy this theorizing. I would be far happier if there was no contradiction and I could wander out in to the veld and be assured that there were smart neat boxes and simple ways of filling them from a manageable list of names and unequivocal identifying characters. My observation is that all this so-called “taxonomy” practiced by too many writers is so flawed because the very basis of biological variation is wholly ignored and forgotten. It does not take rocket science to imagine that science was supposed to set us free from superstition and blind belief and allow us to individually question the nature of creation.
Here we have a real problem that is overridden by a basic problem in formal classification. If I could start now afresh with a clean slate and knowing what I now do, I would still be handicapped by the restrictions of the nomenclatural code. This is that I do not believe that the name H. mirabilis as typified by a plant that comes from an outlying population truly represents the species. Perhaps this is a point not understood by the collector community, and in fact even by botanists in other disciplines. The “type” is only there to serve as a reference point for the NAME and may not be remotely typical of the species that this name is applied to. What is unfortunately true as with the crop of taxonomists who have muddied the classification of Haworthia in more ways than one, is that more often than not the application of names is not to species at all but to an odd assortment of plants that meet no criterion for a basic unit of life forms. In the case of the name and type of H. retusa there has been curiously no problem and the vast variation there is forgotten simply because of familiarity. The interesting thing is that the name H. mirabilis was not even in use before I resurrected it in about 1976, so anything that remotely departs from an unfamiliar norm must be “new”? For H. retusa ‘nigra’, I must have got the name ‘nigra’ from the old literature and reference to the Kransriviermond collection that Smith informally listed as H. mutica. The point is that in my acknowledged ignorance I attached the name to H. mutica because of the absence of an end-awn and a relatively rounded leaf tip to the plants seen. (Looking back to all my pictures I see that in some plants there is definitely an end-awn.) The dark-colour has resulted in a name that is not appropriate for the populations that can be considered to constitute the taxon. Neither is the population at a geographic end-point and as a probable hybrid entity representative. This statement also only scratches at the surface of a real problem of origins and changing faces of the plants with time.
Reflecting about relationship as a taxonomist is required to do, and with due respect to DA and the weird views of my preoccupation with relationships the invitation to him required; it seems to me that the unseen and unknown element, now only evident (H. retusa ‘nigra’) as a widespread and common element in this central area, is important. It may be the precursor to the real way in which H. retusa and H. mirabilis have developed in close synchrony and association in both easterly and westerly directions.
The really strange occurrences are evident from the map herewith (fig. 150). The two species do not grow in direct close association in the area under consideration. There are places where they do e.g. Skeiding about 20km eastwards; KomseRante, Riversdale; Soetmelksrivier, E Riversdale. Why is this? There is no observable difference in habitat or associated vegetation, and still there is pattern in the distribution. It appears to me that they are separated by flowering time (that does not preclude hybridization as field hybrids show) and in character by colour and a tendency to greater surface roughness of various kind and degree in H. mirabilis. The mystery deepens when H. floribunda appears in two places but again as sole occupant of habitat seemingly favourable to either H. retusa or H. mirabilis. Pattern is broken here because the evidence seems to be absorption into H. mirabilis in other places where the two species do closely meet. The conclusion has to be that I have come no nearer to a neat solution. The H. retusa ‘nigra’ populations cited above will attract no takers, but the H.mirabilis populations will. I create no new name because my personal view of Haworthia does not call for one.
Fig. 149 Peduncles of H. mutica, E RiviersonderendMap of Tradouw South
Acknowledgement I particularly want to thank Gerhard Marx for his considered points of view he has crafted as my Devils Advocate. He is an extraordinary grower and observer, and an artist of the highest order. He had driven me to more carefully and thoroughly explore and document what I see and think. I am extremely grateful to him. Lawrence Loucka has been a great help as an advisor, and so has Steven who brings something special to the whole field in the way he “communicates” with plants and about them. Kobus Venter is ever supportive. Access to Van Reenens Crest was through kindness of Trevenan Barry unsoured by the pressures of harvest time. ♦
A review of the plants photographed recently around Albertinia … I only know of 23 populations in this area that relate in different ways to the names Haworthia turgida, H. retusa, H. mirabilis, H.emelyae, H. pygmaea, H. splendens, H. vincentii, H. acuminata, H. fusca, H. dekenahii, H. argenteo-maculosa, H. pygmaea, and H. esterhuizenii. I still need to get pictures of H. splendens, H. vincentii, and H. turgida on the Wydersriver.
In essence there are two variants – the clumping cliff dwellers (H. turgida) and the mostly solitary flats dwellers (H. retusa). The flat dwellers are far more variable than the cliff dwellers. If all the variation is considered against the differences in each of the habitats, it seems very unreasonable to suggest that these populations are anything but a single species (H. retusa) expressing itself differently according to local dictates and demands. This is same issue of variability occurs in these plants all the way westwards.
97. 2020.03.04 – Esterhuizenii – Aasvoelberg. I was concerned that I am not sure anymore of flowering time because essentially retusa and mirabilis are separated by a late summer vs a spring flowering. None of the populations I saw now in late February showed any sign of buds or flower. Only the Humor plants had just two old inflorescence of about two months age. The fact it is a SYSTEM. The retusa elements are lighter coloured signified by having an element “pallidifolia”, while the mirabilis element has the element “nigra”. Recognising species as systems, variation and differences can be seen in a more realistic way. This does help to overcome the pressures that ignorance, egocentrism, emotion, commercialism and intellectualism bring to bear on classification. H. retusa ‘esterhuizenii’?
The map is useful although it does raise the question of how confidential should we be about locality. Part of this quest was to examine how populations have fared in the years since “discovery” and also in the 50 years since I have been directly involved. My observation is that the main threat is excessive animal pressure – and/or combined with drought. The Aasvoelberg population seems to be very small and a secondary threat is alien weed infestation. But I really do not know the actual size and health of the population. I did see plants further west long ago, but it will be a real mission to explore there properly. An important point about conservation is that it is really only as systems that species can cope with natural and even man-imposed change. It is only with a proper and true definition of the term “species” that we can even discuss them sensibly. “Adaptive” must surely be part of that?
98. 2020.03.05 – This is H. retusa (turgida) at Buisplaas on the Gouritz River between the Gouritz Bridge and Herbertsdale. This form was informally known as H. rodinii and is known from the Gouritz Bridge and also south of that. As a cliff dweller it is normally clumping. The first time I saw such a plant it was a specimen reputed to have come from Die Hell much further north in the Swartberg Mts. VERY unlikely, but H. retusa should be sought in the Gouritspoort where the river passes through the Langeberg Mts. There are many populations of these cliff dwellers and countless variants. I do not and cannot, dispute the fact that names present a problem for suppliers and collectors, but that is not a problem that can be settled by science. Association of a botanical binomial and place is the only solution that makes sense to me. Taking out the geographical detail diminishes the sense of any name. But this series is to explain and expand on this issue.
These are also Buisplaas but in a totally different habitat several hundred meters from the cliff dweller. One picture shows a very proliferous clone. This same juxtaposition of clumping and non-clumping occurs elsewhere and is a pronounced feature of the H. cooperi and H. cymbiformis problem in the Eastern Cape. These Buisplaas populations validate my conviction that the Western cape retusoids are in fact a single species system. There has been a predictive element in all my exploration since the revision of 1999 Haworthia Revisited and this population simply fits a pattern. Comparing all the pictures of this series I am now showing will demonstrate this too. H. retusa ‘pygmaea’. I write the name like this because in the absence of a species definition, formal lower ranks have to be meaningless. Furthermore the nature of local variation is such that it is generally impossible to circumscribe ranks. A very good example of this is the persistent reference to H. mirabilis var. magnifica which can absolutely not be supported on the basis of a type specimen representing an original group of such similar or like plants.
99. 2020.03.07 – Despite the drought and animal activity (and human predation presumably) – the Cooper Siding plants are in great health. I am posting all my pictures (of Haworthia – there are many other plant species of significance present) because the reality of diversity within populations gets zero attention in citizen science and little more in academia. But do not get me wrong. I write “citizen science” not citizen cognition. Also I do not ignore the fact that “citizen science” also features in academia. These plants (a plant?) was described as H. dekenahii var. argenteo-maculosa. In my Handbooks 1976, 1981), I placed the two under H. retusa. Was this wrong? No – because moving them to H. pygmaea as I did in my revision (1999) highlights the reality that this is a single system and not just spatially isolated populations that can be treated as different.
In writing about Albertinia and the conservation state of these Haworthia populations (that really bothers me), some very serious thoughts come to mind. Do we really want to know the problem and the answer(s)? The problem is environmental deterioration – climate change. This is coupled with the decline in moral values across the counter. But do we want to acknowledge this and really want to know what is true. That all of existence is actually a grand illusion that we may choose to be locked in to. I should not have suggested that climate change is the main factor – although Albertinia has been experiencing the worst drought in living memory. Livestock pressure also definitely impacts negatively on vegetation, and our dietary addictions prevent us recognizing this fact.
100. 2020 03.08 – Dekenahii is at Draaihoek on the Valse River northeast of Albertinia. Juxtaposed again on a nearby rocky cliff is “turgida pallidifolia”. A floribunda / chlorocantha variant is in close proximity in its own preferred space. A different form of the turgida retusoid used to be on north facing cliffs a little further north at Weltevrede. Keep in mind that I am considering a bigger picture in which differences within populations negate differences between them, and especially when habitat is considered. I also juggled dekenahii between retusa and pygmaea and whatever you do with it, the best explanation I can offer as a binomial is H. retusa ‘pygmaea’. Better still is to add the unwritten meaning that it is actually Helgaard Oosthuizen’s discovery of H. retusa as a group of plants at Draaihoek circa 1943 given to Smith by Dekenahii. Names standing alone may be good for business but the interest lies in the information attached to them and which they point to.
This is the “turgida” form of H. retusa at Draaihoek. These plants have more pointed leaves and are more colourful than the “pallidifolia” that I remember 59 years back. But the precise locality may have been different. I remember plants more like those at Tweekuile further east along the river that I will still post pictures of. At Weltevrede further upstream the plants had longer leaves. Steven Molteno coincidentally reminded me of H. marumiana and the exact same conundrum I am trying to explain here, but magnified manifold over the vastness of the Karoo and our ignorance of both marumiana and the Karoo.
101. 2020.03.10 – Tweekuile is north of Albertinia and just a few km east of Draaihoek on the Valse River. This is what H. retusa ‘turgida’ looks like there. It occurs to me that sensible taxonomy, like prevention and cure of disease, is not a sustainable business model 😅
Without any simple and practical measure, how does one deal with an endless array of slightly different looking things affected by age, exposure to direct sun and an also seemingly endless lot of agencies affecting appearance. Compare these two Cooper Siding and Humor pygmeas with the Tweekuile turgidas.
102. 2020.03.11 – Have a good look at the rest of Humor. I suppose it is still fashionable to dismiss variation like this as “hybrid swarm”. Problem here is what parents where and when. To be strictly correct, is there really any option but to say H. retusa ‘pygmaea’? ♦
Also southeast of Heidelberg is this population of turgida just to demonstrate how variable it also is. Here it has erect narrow leaves and shows the colouration of the retusoids. Haworthia turgida occupies about 75% of the area that mirabilis does but they never actually share habitat. I remind you that I do not like the nomenclatural system and where it occurs that the name retusa would be required to be preferred over turgida, when in fact retusa is a lesser variant of turgida.