
27. Haworthia parksiana V.Poelln., Cactus J 5:34(1936). ibid., Feddes Repert.Spec.Nov. 41:205(1937). ibid. 43:104(1938). ibid., Desert.Pl.Life 10:48(1938). Bayer :143 (1976). Bayer :50 (1982). Scott :121 (1985). Type: Cape, Great Brak River, Mrs Helm in PE Parks 636/32. Not preserved. Lectotype (B&M). icon (B).
parksiana: in honour of ‘Mrs Parks’.
Rosette stemless, proliferous, 3-4cm φ. Leaves 25-35, 1,5-3cm long, blackish-green, sharply recurved, minutely tubercled, tip barely pointed. Inflorescence simple, slender, to 20cm. Flowers few, narrow, whitish with dull greenish venation.
1982 – H. parksiana acquired its name in a very odd fashion. Von Poellnitz received the plants from F.R. Long under a Port Elizabeth Parks and Recreation Department collecting number ‘Parks 636/32’. Long had in turn received the plants from Mrs Helm of Great Brak. Von Poellnitz in error attributed the collection to a ‘Mrs Parks’ and hence the name. It is a very distinctive and very small dark species occurring very locally in Mossel Bay and the Great Brak area. Rather than having the flattened retused end area of the typical ‘retused’ species, the leaves are recurved at the ends. The flower buds are also round‑tipped at the end. The most obvious relationship that H. parksiana has with other species is with H. floribunda, although these two species are separated by the Gouritz River valley. H. parksiana, apart from being the smallest species, is also possibly the rarest and occurs only in very small numbers in the wild. It grows completely under the protection of small karroid shrubs, well hidden in the fallen leaf debris or among moss and lichen. The Mossel Bay area receives rain throughout the year but H. parksiana is particular about not having too much water. Offsetting does occur but growth is very slow.
1999 – The species is known to me from 3 localities. One of these is within the township of Great Brak, the other is on a rocky spur jutting out into a district road, and the third is on the very edge of a wheat field. Its continued existence thus seems to be fairly precarious. The relationship is with H. floribunda which is known (in terms of its probable or possible relation to H. chloracantha) at its most eastward, north of Albertinia. That population apparently includes forms which are comparable with H. parksiana. J.D. Venter cites the case where he has grown field collected seed of H. magnifica from south of Riversdale and observed parksiana-like plants among the offspring. This is what should be expected in terms of the reciprocity of forms, and also in terms of a ‘chaos’-driven speciation theory. Otherwise the species is readily recognisable and a relief from the contorted synonymies of other species.
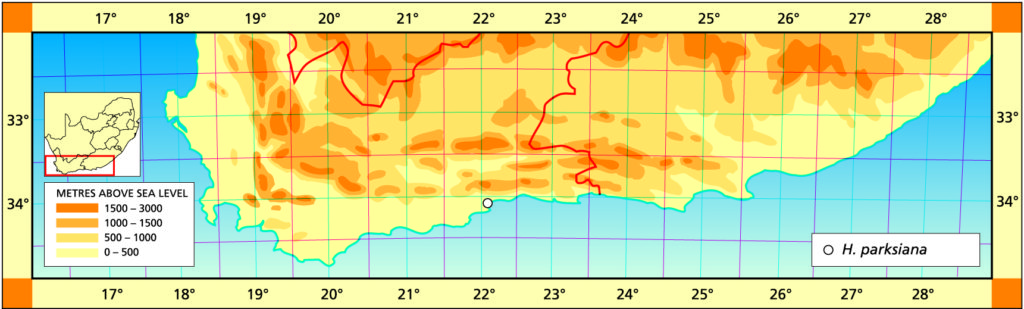
Distribution: 3422 (Mossel Bay): Dumbie Dykes (-AA), Bayer 157 (NBG); Great Brak (-AA), James in NBG8106/45 (BOL), de Wildt (BOL), Smith 2936 (NBG); Botteliersberg (-AA), Bayer (NBG).

Haworthia parksiana JDV94/2 Great Brak. There is not much variation in this species but there is a reason to speculate continuity with H. floribunda. 
Haworthia parksiana JDV96/87 Little Brak. This was a dense population among lichens on a small rock outcrop. 
Haworthia parksiana JDV94/2 Great Brak. Can cluster profusely, yet still be very cryptic under dry conditions.






















































