It is a matter of considerable concern to me that the lack of “adequatio” for the understanding of Haworthia is so conspicuous by its absence. Three reviews appeared for 5 small essays published as a supplement to Haworthiad and also as a portion of Haworthia Update Vol. 5. To suggest that these reviews are anything but some sort of myopia is an understatement and I find it difficult to try and explain why. My reason is simply that many criticisms simply arise out of the fact that the reviewers are not familiar with my writing and have no basis for assuming that they are up to the task of reviewing what I have written. Nothing is being learnt from history where the same old arguments about opinions hardly differ from those voiced 60 years ago.
The real issue is that there is confusion in the minds of collectors as the user public, about the uses and application of Latin names in the pursuit of their interests. Professional botany offers no assistance and any numbers of poorly equipped authors of necessity, invent and re-invent types, the interpretation of names and then their application. This does nothing to assist the collector. The reviews of that small segment of my writing do not help either and certainly do not generate a climate that encourages me to write any further. One review calls for a revision when the message is that a formal revision might just be intellectual vanity when a practical list of names and explanation is presented to add to an existing Revision. A second review seems to tout a mistaken belief that self fertility and polyploidy may be explanations for the intractable problems that we have in the self-sterile non-polyploid haworthias. A third review is seemingly facetious comment that may obscure my opinions as those of the reviewer and acceptable as such, but not if seen to be mine. None of these reviews will be helpful to anyone who may need to decide what set of names makes more sense than another. There is no common voice and any reader is left to make up his own mind in the absence of any good authority. Perhaps this is something I should be personally grateful for because there are noises emanating from various sources that my opinions also threaten understanding rather than enhancing it. I have to accept that it may be difficult for anyone to agree with me who has not had the same exposure to the intractability of biological systems that seems to have been my lot in life.
I regret that I did not, prior to publication, see the forward to Vol. 2 that I hoped would generate “common voice”. It, unfortunately, is just a bit of personalized chitchat about the writer. It can hardly generate any confidence in what he may have written about and does not attempt to do so.
My revision in 1996 (published 1999) admits the problems of typification and my attempt to stay within the historically recognized interpretations and applications of names. A revision is fundamentally a reconciliation of new information and new “collections” and the Update Volumes have been an ongoing reporting of new finds and observations. More than one reviewer seems to think that a change of mind is indicative of weak argument rather than a product of new and better information.
Update Vol. 1 (2001), published by Umdaus Press includes one essay regarding Haworthia mucronata and five others dealing almost wholly with Haworthia cooperi and its variants.
Subsequent volumes were published by Alsterworthia. Update Vol. 2 appeared in 2006 in two parts comprising a total of 17 chapters and a list of Bayer accessions. Update Vol. 3 was published in 2007, also in two parts and comprising 15 essays. Update Vol. 4 was also published in 2007 in one part comprising 10 essays of which two were intended to close the saga. However, a surge of inquiry and energy made me decide to have a last serious effort to try an answer some outstanding questions in my own mind. So Update Vol. 5 was published in 2009 and comprises 16 essays. Perhaps, if nothing else, these essays just demonstrate how much there is out there in the field that has escaped attention.
Update Vol. 6 is simply a small set of essays that address some loose ends of which there are still many. One essay in particular will remain unwritten because, while I have some information, it is not enough to base any conclusions on. This concerns the area between Oudtshoorn and Uniondale. It is also not the only gap in my experience and knowledge. The chapters in Update Vol. 2 should demonstrate just how intensively the field has to be explored to get to grips with the possible dynamics of the plants. This view should be re-enforced by the subsequent volumes. While I have tried to hold to a belief in “species” and names relevant to some sort of system, it seems to me that as a society we are denied the freedom to really know what the meaning and purpose of creation is. Science itself seems to insist that there is none and botanists largely view the observance of nomenclatural rules as the primary criterion in the application of names.
My experience and observation now suggest to me that creation is purposeful and meaningful. Life is manifested in various distinctive forms and these forms are manifested according to their DNA. This DNA may be the fundamental stuff of consciousness in living things too; flowing from and responsive to the energy fields of physical bodies such as the galaxy and the solar system. Life on earth is driven by catastrophic events of varying degree and varying intervals, so that species simply represent those sets of living things as they come, change and go with successive events. We do not only need to consider what species might be and mean, but also when. Who knows? ♦
There is a very curious parallel in the problems and aspects of classification and identification of haworthias and chameleons that I personally find a bit mystifying. Classification of succulent plants and especially Haworthia is often done outside of mainstream science, which means that it is not done by trained professional botanists, who can stand in the intellectual arena with academics and degreed intellectuals. It has been pointed out to me that this is the problem in Haworthia and also why professionals do not want to attempt to resolve the issue because the nomenclature has been so confused and complicated by all the bungling that has taken place. Having been on the fringes, and indeed even failed attempting to cross the bridge into academia, I find it very difficult indeed to reconcile my life experience in the amateur arena with what I encounter from the professional one. This is because I have observed an ever widening gap between the perceptions of the non-science individual compared to that of the professional. My interest in chameleons is simply another aspect of my interest and passion for living things and I am distressed that the knowledge and understanding of these fascinating animals, as with Haworthia, is so clouded by ignorance and confusion.
It is a bit problematic for me to explain why I think classification and plant names is such a big issue and why it is important to me, but fundamental to me is trying to understand and know something about plants and animals as an extension of my life experiences.
Recently a very welcome and valuable booklet, Chameleons of Southern Africa has been authored by Krystal Tolley and Marius Burger and published by Struik Publishers (Cape Town). What makes it so interesting for me beyond my interest in the animals themselves, is the remarks the authors make in the preface in respect of writing the species accounts. The problem of finding identifying characters was a key issue and there is constant reference to confusion. There is of course no statement about what a chameleon species actually is and there is this curious statement, “There are few consistencies of identification characteristics, and we admit that we sometimes initially identify a chameleon by means of ‘gut feeling’.” What makes this so curious is that there is nothing in the booklet to indicate there is any more to the mystery of identification and how a final identification could be made to possibly help the even more than average individual to arrive at own conclusion.
In Haworthia (as indeed is the case in many plant genera) the description of species has been based on a basis of simple visual (morphological) difference, with barely any consideration of variation and in the total absence of a definition of a species beyond the assumption that if they look different they are different (species).
One has to ask what is it that is referred to as ‘gut-feeling’? I have often heard and read that classification and taxonomic statement is a matter of opinion and felt compelled to respond by quoting a writer who made the distinction between mere opinion and informed opinion. “Gut-feel” may vary from the instant response of the ignorant to a fleeting image, to the deeply considered intuitively driven reservation of another who is highly informed and who has a vast bank of experience, images and literature to draw from.
The significant thing is that any decision-making in the chameleon booklet has been facilitated by results from DNA analysis – *please see the footnote. Now I must make it clear that I am not attacking the book or the authors. I want to draw attention to the problems inherent in a methodology which amounts to nothing more than saying “trust me”, and the very serious problem that own judgment is by implication quite useless. As a very knowledgeable observer wrote to me “Our own eyes and brains are seemingly no longer good enough to serve the world – well that is what the RNA-DNA lab technicians are trying to tell us.” There have been several attempts to evaluate the classification of Haworthia by DNA sequencing that do little more than ring alarm bells. Without any pretence at understanding all the convolutions of genetics and the repository of DNA in all its forms, I would have to ask again, what does one do to organize one’s scepticism about the results that are thrust upon one? If the total facies that an individual organism presents to the eye and senses is not a reasonable representation of the information encoded in the DNA, then we are absolutely helpless. Could one say that we have been put into this creation without the sensory capabilities to distinguish and discriminate anything?
Recently I submitted photographs of two chameleons to an authority for identification stating that I thought they were not the Karoo Dwarf. The reply I received was “It looks like the Karoo Dwarf. What makes you think it is not?” Apart from the fact that I had already given reasons, the truth is that my correspondent could not possibly have said why he thought it did look like the Karoo Dwarf, thereby shifting the responsibility back on to me. Are we stuck with the sad and unavoidable fact that unless one can get a DNA result, one cannot know what you are looking at?
What has now happened with respect to Haworthia DNA where we have the same dreadful problem of identification and understanding? I have elsewhere (see Haworthia Update 3 (Ch 3: p92) discussed the papers by Messrs. Treutlein et al and I summarily dismiss what I think may be unpublished results by another source I must politely keep to myself. There was a rather immodest statement by Messrs. Wallace and Noll published in Haworthiad that promised to unfold the problems of Haworthia classification by DNA analysis. The results have never been disclosed and there is a disconcerting failure by the authors to respond to any inquiry. More recently Dr. Syd Rhamdhani has also been busy sequencing the DNA of a range of Haworthia and species of related genera. The work was initiated in an attempt to relate variability to habitat diversity, but to preclude a total negative result it was deemed advisable to widen the scope to ensure that the methodology was applicable at a broader scale. The results demonstrated, at least from my personal perspective, a complete failure to relate the DNA sequences with the realities of the classification of at least the semi-rational mind of an experienced and informed observer.
Group1A
Group 1Aa
Code
Collector Number
Taxon name
Subgenus
Locality
Source
54
MBB 7128
angustifolia var. baylissii
Haworthia
Wellsgate, E. of Kirkwood
Field
47
MBB 6843
marumiana var. reddii
Haworthia
Inverbolo, Stutterheim
Sheilam
58
MBB sn
truncata v. maughanii
Haworthia
Sheilam
Group 1Ab
Code
Collector Number
Taxon name
Subgenus
Locality
Source
35
JDV 90/57
monticola var. asema
Haworthia
S. of Kruis River
Field
40
JDV 97/128
mucronata var. mucronata
Haworthia
SE of Barrydale
Sheilam
6
SR&TDsn 4
angustifolia
Haworthia
Mountain Drive (Gtown)
Sheilam
Group 1Ac
Code
Collector Number
Taxon name
Subgenus
Locality
Source
32
EvJ 17548
cymbiformis var. transiens
Haworthia
Horee
Field
52
MBB 7032
mutica var. mutica
Haworthia
Rietfontein
Sheilam
Group1B
Group 1Ba
Code
Collector Number
Taxon name
Subgenus
Locality
Source
30
PVB 7047
zantneriana
Haworthia
N. of Redcliffe, Willowmore
Sheilam
62
MBB sn
reticulata
Haworthia
Bosfontein
Sheilam
48
MBB 6875
maraisii
Haworthia
Cogmanskloof, Ashton
Shadowlands
51
MBB 6985
rossouwii var. calcarea
Haworthia
De Hoop
Group 1Bb
Code
Collector Number
Taxon name
Subgenus
Locality
Source
33
GM 623
marxii
Haworthia
Rooi Nek Pass S. of Laingsberg
36
JDV 91/91
cymbiformis var. setulifera
Haworthia
Rainbow Valley
Sheilam
4
SR & TDsn 2
cooperi var. gracilis
Haworthia
Mayfair (Gtown)
Field
18
SR & BBsn Site 5- 27/6
cooperi var. viridis
Haworthia
Kaboega, Dassiekop NE of farmhouse
15
SR & BBsn Site 3- 26/6
cymbiformis var. obtusa
Haworthia
Glen Avon falls
Field
16
SR & BBsn Site 2- 27/6
cooperi var. ‘puberula’
Haworthia
Kaboega, W of Wilgefontein
Field
1
SR&TD 719
cymbiformis var. incurvula
Haworthia
Platosvale
Field
21
SR & BBsn Site 3- 18/10
cooperi var. viridis
Haworthia
Kaboega, De Plaat
Field
24
SR & BBsn Site 1- 19/10
cooperi var. spec.1
Haworthia
Kaboega, Kaboegapoort
Field
23
SR & BBsn Site 1- 19/10
cooperi var. spec.3
Haworthia
Kaboega, Kaboegapoort
Field
25
SR & BBsn Site 1- 19/10
cooperi spec. 4
Haworthia
Kaboega, Kaboegapoort
Field
22
SR & BBsn Site 4- 18/10
cooperi var. viridis
Grassland form
Haworthia
Kaboega, W of Wilgefontein
Field
38
JDV 93/45
cymbiformis var. obtusa
Haworthia
Kagasmond
Sheilam
27
SR & BBsn Site 1- 19/10
cooperi spec.2
Haworthia
Kaboega, Kaboegapoort
Field
39
JDV 94/98
decipens var. xiphiophylla
Haworthia
W. of Coega
Sheilam
64
BBsn
aristata
Haworthia
E of Hopewell farm
Field
Group 1Bc
Code
Collector Number
Taxon name
Subgenus
Locality
Source
13
SR & BBsn Site 1- 18/10
cooperi typical
Haworthia
Kaboega, Gertpiets area
Field
26
SR & BBsn Site 2- 19/10
cooperi var. viridis
Haworthia
Kaboega, back of the Dam (Spekboomberg)
Field
44
MBB 6791/4
cooperi var. picturata
Haworthia
Andrieskraal
Sheilam
3
SR & TDsn 1
cooperi var. pilifera
Haworthia
Mayfair (Gtown)
Field
Group 1Bd
Code
Collector Number
Taxon name
Subgenus
Locality
Source
63
MBB sn
parksiana
Haworthia
Mossel bay
Sheilam
50
MBB 6983
rossouwii var. rossouwii
Haworthia
Soutkloof, Napier
Sheilam
67
MBB sn
pulchella var. pulchella
Haworthia
Constable Station
Sheilam
43
MBB 6762
puchella var. globifera
Haworthia
Touwsberg, Little Karoo
Sheilam
Group2A
Group 2Aa
Code
Collector Number
Taxon name
Subgenus
Locality
Source
46
MBB 6829
Poellnitzia rubiflora
NA
Rooikloof
Shielam
71
AJ290302
Poellnitzia rubiflora
NA
Genbank
Group 2Ab
Code
Collector Number
Taxon name
Subgenus
Locality
Source
73
MBB sn
Aloe bowiae
NA
???
Sheilam
9
SR&TD S3-29/8
Aloe speciosa
NA
Plutosvale
Field
10
SR&TD S6-29/8
Aloe africana
NA
Bothas Hill (Grahamstown)
Field
11
SR&TD S7-29/9
Aloe lineata
NA
Stones Hill (Grahamstown)
Field
72
AJ290289
Aloe vera
NA
Genbank
Group 2Ac
Code
Collector Number
Taxon name
Subgenus
Locality
Source
7
SR & TDsn 6
attenuata
Hexang.
Plutosvale
Field
34
JDV 85/145
scabra var. morrisiae
Hexang.
Schoemanspoort
Sheilam
31
EvJ 16840
attenuata var. glabrata
Hexang.
Collywobbles
Sheilam
56
MBB 7179
viscosa
Hexang.
Constantia, W. of Willowmore
Sheilam
65
MBB sn
venosa var. venosa
Hexang.
Breede Riv., Swellendam
Sheilam
41
JDV 1942
attenuata var. attenuata
Hexang.
Soutkloof, Addo
Sheilam
28
Clark & Pienaar 120
venosa var. tessellata
Hexang.
Farm Hughdale 161, Kikvorsberge (Noupoort Region, N. Cape)
Group 2Ad
Code
Collector Number
Taxon name
Subgenus
Locality
Source
66
MBB sn
reinwardtii
Hexang.
Wesley
Sheilam
68
AJ290299
kingiana
Robusti.
Genbank
5
SR & TDsn 3
coarctata
Hexang.
Ecca Reserve (Gtown)
Field
42
MBB 6380
pungens
Hexang.
Braam River
Sheilam
17
SR & BBsn Site 3- 27/6
sordida
Hexang.
Kaboega, E of Olifantskop
Field
Group 2Ae
Code
Collector Number
Taxon name
Subgenus
Locality
Source
19
SR & BBsn Site 2- 28/6
nigra
Hexang.
DePlaat, Kaboega
Field
61
MBB sn
venosa var. woolleyi
Hexang.
Springbokvlakte
Sheilam
74
AJ290298
Gasteria liliputiana
Hexang.
Genbank
45
MBB 6809
fasciata
Hexang.
Gamtoos
Selecta Succ.
57
MBB 7695
glauca
Hexang.
15 km S of Kaboega Farm house, Darlington Dam to Kirkwood
Field
20
SR & BBsn Site 1- 27/6
glauca
Hexang.
Kaboega, W of Wilgefontein
Field
Group 2Af
Code
Collector Number
Taxon name
Subgenus
Locality
Source
59
MBBsn
glauca
Hexang.
????
12
SR sn 30/6/08
glauca
Hexang.
Gertpiet se Kraal
Field
Group 2Ag
Code
Collector Number
Taxon name
Subgenus
Locality
Source
60
MBB sn
Hybrid
arachnoidea X blackburniae v. graminifolia
Haworthia
Schoemanspoort
Sheilam
55
MBB 7145
limifolia var. glaucophylla
Hexang.
Three Sisters, Barberton
Sheilam
Group2B
Group 2Ba
Code
Collector Number
Taxon name
Subgenus
Locality
Source
2
SR843
Astroloba foliolosa
Kaboega, De Plaat
Field
69
MBB 6756
Astroloba bullata
??
Sheilam
49
MBB 6952
marginata
Robusti.
N. of Ashton
Sheilam
70
AJ290292
Astroloba foliolosa
Genbank
Group 2Bb
Code
Collector Number
Taxon name
Subgenus
Locality
Source
53
MBB 7096
pumila
Robusti.
Konings River
Sheilam
37
JDV 92/51
blackburniae
Haworthia
Voorbaat
Sheilam
29
EA 1211
kingiana
Robusti.
Great Brak
Selecta Succ.
Group 2Bc
Code
Collector Number
Taxon name
Subgenus
Locality
Source
8
SR&TD S2-29/8
Aloe tenuior
Committees Drift Junction
Field
Dr. Rhamdani has so far generated results from DNA sequencing using the nuclear ITS and chloroplast trnL-F markers. The former, on the evidence of the “species” considered, seems to give a good view of the generic possibilities, with not enough information with respect to the species. The trnL-F spacer results contain several spurious associations if my existing classification (Bayer 1999) and geographic distance are valid considerations. A poster presentation of the results has been published in Alsterworthia International 9(3)13-17.) Dr. Rhamdani has been kind enough to send me the product of an alternative manipulation of his results that I tend to equate with an attempt to display the relationships of samples as they would be in geographic space. One can see from the groupings (see tables below) that there are gross anomalies that cannot possibly be taken to have classification truth. Group 1Aa for example has H. angustifolia, H. marumiana ‘reddii’ and H. truncata ‘maughanii’ nested together. Group 1Bd includes H. parksiana and H. rossouwii. The presence of H. kingiana in two different groups viz 2Ad and 2Bb has to be dismissed as an identification error where I would stake my reputation on EA1211 rather than AJ290298. Each group can be tediously discussed, but one needs to consider that the actual focus of the exercise was to explore the geographic relationship of DNA sequences below species level. Far too much was expected of a still poorly understood technology.
Coming back to chameleons, I have the problem that, being interested in the creatures, I want to know what they are. Having familiarized ourselves (my wife and I) with the Cape Dwarf (Bradypodium pumilum) we came into contact with the (then) Robertson Dwarf (B. gutturale now Little Karoo dwarf). The distribution given by W R. Branch and the descriptive detail did not seem to match what we were experiencing in the field and we concluded that the Robertson Dwarf and the Karoo Dwarf (B. karroicum) were surely the same species. We then encountered what we thought was the Western Dwarf (B. occidentale) and were derailed by the opinion given us from a DNA analysis of the “population” observed, that it was a variant of the Cape Dwarf viz. a “renosterveld form”. What we were told did not satisfy our “gut-feel” however profound and true it may have been or is. Then the booklet was published that confirmed our conviction that the Robertson and Karoo Dwarves were being confused (The true Karoo dwarf is now said to equate the E cape dwarf B. ventrale!), but not throwing any more light on the question of the “Renosterveld form” of the Cape Dwarf.
We then learned of a chameleon in the Gouritz River valley. On seeing these we concluded that on morphological grounds they were not the Little Karoo Dwarf and had to fall within the ambit of the Swartberg and Eastern Cape Dwarves as accepted by Tolley and Burger. These are the chameleons “identified” by an authority as the Karoo Dwarf despite our submission of evidence why it was not. This led us back to the published DNA work and a paper by Tolley et al that discussed specifically the Eastern Cape Dwarves and a number of variants suggested being new species. My experience in Haworthia generated the strong “gut-feel” that this may be quite wrong as wide field experience suggests that species are highly complex systems that defy the conventional taxonomic approach. Further sampling would quite probably generate a host of still other variants and require explanation. It is quite obvious that the chameleons cannot be identified with certainty using morphological characters and this is equally true for Haworthia. My experience is that it is true for many plant genera. Nevertheless, I think that this is because morphological criteria are so heavily relied on with not enough reference to geographic distribution and a rational species definition, as opposed to an open-ended species concept. In another paper by Tolley et al, the two species B. melanocephalum and B. thamnobates are shown to be genetically identical, despite the fact that they are known to be morphologically different and geographically separated. Thus it was a pleasant surprise to find another article (Miller & Alexander (2009) that probes this question and speculates that it is possible that the DNA sequences used in the analysis may not be representative of the full genome. The available results of DNA sequencing for Haworthia suggest to me that those sequences are also not adequately or accurately representing a true or reasonable phylogeny. A problem I then also have is that a phylogram is a simple two-dimensional tree that is being drawn to illustrate the relationship of elements that are spread in at least two-dimensional space derived from change with time. In my opinion far too much is thus inferred from complex statistical procedures that generate a wholly theoretical diagram wholly beyond penetration by ordinary skepticism. I would suggest that the so-named bootstrap and posterior probability values may be subject to the same apparently many vagaries of multivariate statistical procedures that are not perhaps fully understood or even recognized by those who use them.
Dr. Rhamdhani suggested that I mention some of the limitations of the sequencing process that he lists:
1. mostly extant samples are used as these are only available for analysis (except in some cases fossil can be sequenced).
2. small sample size.
3. use of a few selective markers.
4. hybridization and incomplete lineage sorting distorts and complicates patterns.
5. appropriate analytical tools have not been formulated to handle the data and the above issue.
6. manipulation, misuse etc. of species concepts to justify desired results – this can be related to the “gut-feel” issue that this article raises.
I did complain about the problem of probing results published in complex technical papers when the results conflict with my “gut-feel”. The response was that “our eyes and our intuition are frequently tricked by the complexity of the real world…” This is flatly not true for Haworthia and I strongly suspect that it is not true for chameleons either. In Haworthia we are tricked by a very simplistic approach and the failure of taxonomic botany to serve interested parties. My kind authority I referred chameleons to also wrote “I know the genetic information may seem very confusing, but it is essential to remember that the genes tell us about genealogy and not morphological or behavioural similarity.” I have said before that the traditional approach of taxonomy was always to relate morphology, and whatever other information one could acquire, to phylogeny. It is an unlucky fabrication foisted on us by DNA sequencing that this is a new direction for classification. To totally believe that, say, less than a thousand base pairs on a tiny section of mitochondrial DNA is giving what must be seen as a final answer that defies the doubt of an observer is perhaps too much to ask.
An end point in my chameleon inquiry came when Dr. Tolley conceded (as most DNA experts now seem to be doing) “DNA is obviously not a final answer, and everyone that understands well how evolution works, already agrees on that. However, when used wisely, in connection with morphology (which is subject to phenotypic plasticity, strong selection, and local adaptation) it obviously can be useful in understanding the evolutionary history of organisms. We can use DNA to give us clues as to the evolutionary lineages, which sometimes can be the same as what we humans like to call “species”. For me, “species” is partly an artificial construct because evolutionary processes do not suddenly jump, and divide one thing into two species, there are gradual changes, and sometimes (more often than we care to admit), they do not fit in the box we have labelled “species”. She exposes the fundamental problem that no one probably understands properly how evolution works nor do any of us have the wisdom to use DNA information to best advantage.
References
Ashadee, K.M. & Alexander, G. L. 2009. Do dwarf chameleons (Bradypodium) show developmental plasticity. African Zoology 44:45-54.
Tolley, K.A et al. 2004. Phylogenetics of the southern Africa dwarf chameleons, Bradypodium (Squamata: Chameleionidae). Molecular Genetics and Evolution 30:354-365
Tolley, K.A. & Burger, M. 2007. Chameleons of Southern Africa. Struik, Cape Town.
Treutlein, J. et al. 2003a. Phylogenetic relationships in Asphodelaceae (subfamily Alooideae) inferred from chloroplast DNA sequences (rbcL, matK) and from genomic fingerprinting (ISSR). Taxon 52:193 207.
Treutlein, J. et al. 2003b. Evidence for the polyphyly of Haworthia (Asphodelaceae Subfamily Alooideae: Asparagales) inferred from nucleotide sequences of rbcL, matK. ITS1 and genomic fingerprinting with ISSR-PCR. Plant Biology 5:513-521.
Note. The Chamelion DNA study rests on the sequencing of about 40 individual specimens as opposed to the 1000 plus used for the former morphological revision. ♦
In Haworthia Update 4 I wrote an essay about the haworthias east of Albertinia in which I discussed their relation to H. retusa and H. mirabilis, while generally lumping them largely in H. pygmaea. There are of course real ‘turgida’ populations as far east as near Mossel Bay, so I argued the case for an interplay of the two former species that over the whole distribution range generated two ‘species’ in the east viz. H. pygmaea and H. retusa (to include ‘turgida’), and three ‘species’ in the west adding H. mutica to H. mirabilis and H. retusa.
Recently I had the pleasure of meeting Gregory Nicholson who is studying botany at University, Cape Town. He surprised me by telling me that there was a Haworthia on his parent’s property west of Herbertsdale. It was not in fact so much surprising as confirmation of the belief I formed on a trip a short while before that there must be haworthias in the very suitable terrain of the Jakkals River valley 6km west of Herberstdale. The surprise came when Greg indicated the position of the plants much deeper into the mountains.
We visited the site while exploring H. emelyae north of the Langeberg Mountains and I will report that in an additional short article. We over-nighted east of Barrydale and a puncture made us detour via Riversdale rather than take the northern dirt road. It was raining heavily at Albertinia, but we could see sunshine on the Langeberg in the direction of Herbertsdale. We traveled the dirt road through the upper Gouritz, because we also wanted to gauge the condition of the gorge through the mountain, as Greg reports the presence of H. retusa ‘turgida’ some distance in. Towerlands is some 8km on and it was still just sunny when we arrived. We walked immediately the several 100m to Greg’s site along a good path among true Fynbos vegetation and found the plants very easily in a ferruginous sandstone ridge sloping into the river. Then the rain came and made picture taking a bit difficult (see MBB7489 figs. 1-16).
The plants are quite small and there was not much clustering. Unfortunately the weather restricted us so we could not look for better habitat around the corner where Greg had actually pointed. Greg’s photographs had made me lean to H. pygmaea and indeed I do not think one can deny the reality of this if one also considers the odd plants at Herbertsdale itself (JDV97/135) that I assign to H. pygmaea. However, the similarity to ‘turgida’ is just as striking and Daphne and I concluded that we could recognize similarities to ‘dekenahii’ and even ‘mutica’. To make it even more confusing, I felt, and still do, that the plants had similarities to H. emelyae. This latter was confirmed in my mind when we continued our trip via Cloetes Pass back to the Little Karoo to just west of the Gouritz gorge and a population of H. emelyae known to us from Gerhard Marx. I will submit that in a subsequent short note.
I include a habitat scene (fig. 1 – the arrow marks the locality) and note that looking south east one can see Albertinia and Aasvoelberg in the distance. We know that ‘turgida’ is present in the sandstones of the Langeberg, but this find suggests it is far more abundant and widespread than present records suggest. We need to now what the plants in the upper Gouritz look like.
Acknowledgements Gerhard Marx has been extremely helpful and informative and I am most grateful to Gregory Nicholson for his contribution. ♦
Kaboega is a set of farms on the northeast of the Zuurberg Mountain range, north of Kirkwood and off the Addo National Park. I wrote about the haworthias that occur there in Haworthia Update Vol.1. There is also an article in Aloe 40:10 (2003) in which there is a discussion of the variation of those haworthias as related to geology and topography. My wife and I frequently visit Kaboega to renew relationships with Ian and Sandy Ritchie who live there. Each time we go we try to explore some different area. We generally end-up with something that is notably new.
Part 1.
There is a real problem in trying to reconcile the populations we see with the names that are available and the way in which I have tried to formalize them myself. The problem is that Kaboega seems to occupy some sort of central and neutral position and it is by no means easy to arrive at any clean rational classification. Three of my species are involved, and I have to say they are “mine” because other authors are in strong disagreement. The three species I see are H. cymbiformis, H. cooperi, and H. aristata. It is firstly necessary to explain that I interpret the name H. aristata in Haworthia Revisited quite differently from what I might have done earlier; and quite differently from other authors who have simply taken the easy route and associated the name with Little Karoo elements for which I use the name H. mucronata. My interpretation of the name will be quite evident from my writings and from the pictures submitted with this article. The use of the name H. cymbiformis with respect to Kaboega is a major problem for someone like myself who is firmly convinced that geographical relationships are foremost in the recognition of species as living systems. On Kaboega, plants that look like H. cymbiformis seem to proceed out of a complex that is surely H. cooperi. If one properly considers all the populations that I ascribe to H. aristata one is seriously confronted with the reality that it is also a geographic variant of H. cooperi.
During June 2008 we were again at Kaboega and found two further populations of ostensible H. aristata. One of these viz. MBB7698 (see figs 1a-d) is only about 1km east of Buffelsnek on the western boundary where I recorded MBB7012. We did also see the same plants on a ridge still further east and north of Wilgerfontein where the very green and proliferous cymbiformis-like plants seem to reach their western limit. MBB7698 is effectively on the top of the Spekboomberg ridge, whereas the cymbiformoids are a little to the west and on the south slopes. It would be really interesting to know what occurs further westwards towards Darlington Dam.
We found H. aristata again practically central to the farm and about 1,5km south east of the Weir (MBB7703, see figs 2a-c, and these plants may have been illustrated before elsewhere incorrectly as MBB7697, a Gasteria). This is still about 1km north of the east-facing riverine cliff that houses the very green cymbiformoid plants (MBB7636, see figs 3a-c) of which other populations have already been reported to the east at De Plaat (3 populations) and also to the west at Koks Dam and Spekboomberg.
I have probably argued for recognition of H. aristata as separate from H. cooperi. Such an entity can probably be recognized as occupying the lowlands to the north of the Zuurberg. Both H. cooperi and H. bolusii var. pringlei occur to the north at Ripon and it appears that the latter may be represented by a population northeast of Stonefountain. There is a problem with proper, as opposed to the formal way either Breuer or myself may have done typification of the name H. gracilis, where we have both apparently just guessed that the name applies to the Helspoort variants of H. cooperi. In fact von Poellnitz’ name H. gracilis rest on the citation of a really mixed bag from not just Hellspoort, but from five quite disparate locations across almost the entire Cape. I use the name ‘gracilis’ only in reference to Hellspoort and it is evident to me that H. aristata is in the H. cooperi continuum. Look at the pictures and see what you think. I have added three pictures (MBB7701, see Figs 4a-c) taken of plants above Klipfontein at the eastern end of Kaboega that I refer to as H. cooperi ‘puberula’ and related to another record MBB6908 at Wilgerfontein.
1a. MBB7698 H. aristata E Buffelsnek, Kaboega1b. MBB7698 H. aristata E Buffelsnek, Kaboega1c.1d.2a. MBB7703 H. aristata N Swartkop, Kaboega2b. MBB7703 H. aristata N Swartkop, Kaboega2c. MBB7703 H. aristata S Swartkop, Kaboega3a. MBB7636 H. cooperi cf. cymbiformis N Swartkop, Kaboega3b. MBB7636 H. cooperi cf. cymbiformis N Swartkop, Kaboega3c. MBB7636 H. cooperi cf. cymbiformis N Swartkop4a. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega4b. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega4c. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega
Part 2.
Variants of H. cooperi on Kaboega
The various populations of H. cooperi are discussed or referred to in several of my essays published in the Haworthia Updates and elsewhere. I have also been privileged to go back to Kaboega several times and further explore there. Each visit generates something new and different and evokes thoughts of what is still unseen, unreported and unknown.
On this occasion Daphne and I were actually hoping to find and photograph perhaps a chameleon, as workers had reported seeing one abut six months previously in the vicinity of the Klipfontein guest house on Kaboega. This is in the northeastern end of the set of farms that now constitute Kaboega. We thought we should explore the upper slopes where there was more sunlight and basking opportunity for the chameleons in the cold winter sun.
We climbed up the mountain via a very overgrown route that served as a passage for stock to access the higher mountain slopes, but also serve as a donkey pack route for the transport of supplies over the mountain in earlier days. This is of course peripheral to plant exploration. Nevertheless it is so fascinating to dwell on the fact that the enlarged game farm is actually comprised of about seven smaller farms that once were home to farmers and family. All abandoned as circumstances have changed over the years. It is as though man invaded this rich treasure house of natural beauty, sucked it dry and abandoned it again leaving nature to now heal the wounds. This is very unlikely because of the game fences and the absence of predators. Rainfall is marginal and the Kudu population is huge. Nature will have to respond in some dramatic and drastic way if she has anything in mind, and she is perfectly capable of that.
We moved east of our original visit to the mountain top where we had seen and photographed a few scattered plants before. We know that if we find one plant, to ask “Now where are they?” This confinement to such discrete, small habitats is so characteristic that we laugh off the notion that to explain the vicarious distributions over great distance, the plants were wider spread at some distant time in the past. Something else is at work and these populations maintain pattern and coherence beyond material means.
Eventually Daphne, whose eyes are now better than mine, did find the first plant and the body of the population we had only touched before. So these are the pictures. The first three pictures are of MBB6901 H. aristata (See figs 5a-c) on the lower country north of the mountain about 3km away on a neighboring farm Doornkloof, the next one would be a cultivated plant of MBB7701 H. cooperi ‘puberula’ nom. nud. (See fig. 6) in sandstone and Fynbos vegetation in the mountain south of Klipfontein. The rest are H. cooperi (no appellation – MBB6940, See figs 7a-p) just east of my previously reported site north of Klipfontein, also in sandstone and transitional Fynbos. This term is used to simply imply one of these strange anomalies where karoid and Fynbos elements mingle. Many more pictures are really needed to indicate that each plant looks different and we spent a long and enjoyable time calling back and forth to come and observe one more gem.
Some of the clones would correspond to clones in the parent population of H. cooperi ‘viridis’ at Perdepoort about 20km west while others to H. cooperi ‘gracilis’ from Helspoort north of Grahamstown, 50km east. Thus to my mind there is no satisfactory solution to the naming of these plants in our conventional understanding of Latin names.
The last photograph is a view across the Klipfontein valley and over the old greater farmyard to a mountain ridge and over that to the Zuurberg Mountain itself. Between is the Vyeboomfontein farm. The black arrows mark a point on the near ridge where there is a Bushman painting site and a large quartz rock that is home to plants like MBB7101. That mountain ridge extends westward 1-2km to the abandoned DePlaat farmhouse where there are again populations of H. cooperi on northwest and south aspects, at different altitudes and with their own variants. More conventional H. cooperi ‘pilifera’ can be found on the far mountain while Kaboegapoort is a gorge through the mountain with still another set of populations and variants.
It is all a wondrous never ending drama.
5a. MBB6901 H. aristata Doornkloof5c. MBB6901 H. aristata Doornkloof5b. MBB6901 H. aristata Doornkloof6. MBB7701 H. cooperi ‘puberula’ S Klipfontein in cultivation.7a. MBB6940 H. cooperi N Klipfontein7b.7c.7d.7e.7f.7g.7h.7i.7j.7k.7l.7m.7o.7p.7q.
Part 3.
General comments on the plants on Kaboega.
The southern end of Africa is home to one of the six or seven plant kingdoms of the world. This is the so-called Cape Floral Kingdom. Botanists in the past seem to have misinterpreted this flora and not realized that it maintains its integrity for two reasons. The first is that it is on the table Mountain Sandstone formation and the second is that it is in a winter rainfall zone. Its origins are no doubt ancient, but in present day terms it is part of a present day winter rainfall area which includes the Succulent Karoo. Kaboega sits on the eastern verge of this biome and is therefore in a huge tension zone between inland, upland, summer and winter rainfall (and the stress of dryness) vegetation groupings.
Apart from annual rainfall stresses, broken topography, skeletal soils and an enormous range of exposed geological formations and rock series, contribute to an extraordinary range of varied habitats. Thus South Africa and especially the Cape, offer plant species huge opportunities to adapt and change according to ecological differences and stresses. Kaboega is a wonderful example of the way in which plant species have adapted to these different habitats.
Kaboega has plant species from all of the biomes in the Cape, and the vegetation ranges from upland grassy fynbos, renosterveld, succulent karoo, valley bushveld and countless variation of these. The main rocks are quartzitic, dwyka tillite (of glacial origin) and shales. In addition there is a small amount of alluvial terrace.
What is so dramatic is that the different species occupy sometimes very limited habitats and despite this tight habitat choice (requirement) it is very difficult to specify what those requirements are for any group of species.
The genus Haworthia is represented by two of its subgenera. The Subg. Hexangulares is represented by three species:-
a. H. glauca, which is present mostly on the upper quartzitic ridges where Euphorbia polygona and the asclepiad Huernia brevirostris occur. This form of H. glauca includes some evidence of apparent interaction with H. coarctata which comes from further east.
b. H. sordida is an isolated population south of the Zuurberg and then westwards to Steytlerville. It was known on Kaboega from only one small hill where it grew under renoster bush. It was only while researching this visit that a second group was found.
c. H. nigra, which is very widespread in the greater karoo, but rare this far south and so far it is only known from one small population on the river terrace gravels.
The second subgenus Haworthia is only represented by many varieties of but one species viz. H. cooperi and this recognition of one species is only arrived at by discarding the traditional view of plant taxonomy that has no definite and decisive definition for the word species. The Kaboega plants are known from about 30 populations. These vary dramatically, but continuously, according to habitat and this has to be considered in terms of continuing similarities from Kaboega outwards into the Karoo, the Western Cape and the Eastern Cape. This suggests that there are very complex species systems and species cannot be as easily identified and described by visible characters, as a loose definition has in the past allowed. The Kaboega populations are very different from each other, and could be said to be from four or more species if fancy rules. In fact they are probably adapted by habitat and are evidence of the continuity of them all in one species complex.
It is population structure which is so significant and if one considers all the plant species that occur on Kaboega, one comes to realize that they occupy often unique habitats. There is one level of generalists such as Acacia karoo, Olea africana, Aloe ferox, etc which seem to be ubiquitous. But actually most of the species can be seen to have a “home”. There are about seven species of Aloe on Kaboega and seldom do more than two share habitat. As one goes through a general list of names, one finds that one has to visit very specific places to find the plants. One can also find places like this one spot at Klipfontein that can be described as a “hotspot” for Kaboega. These are small species rich areas, which host species which do not occur generally and may in fact be very rare for Kaboega. By also occurring on Kaboega like this, they represent a genetic entity and resource which may be quite unique
Acknowledgement. I am most grateful to Ian and Sandra Ritchie for their generous hospitality and interest for the many times and hours we have spent fossicking in the wilderness of Kaboega. ♦
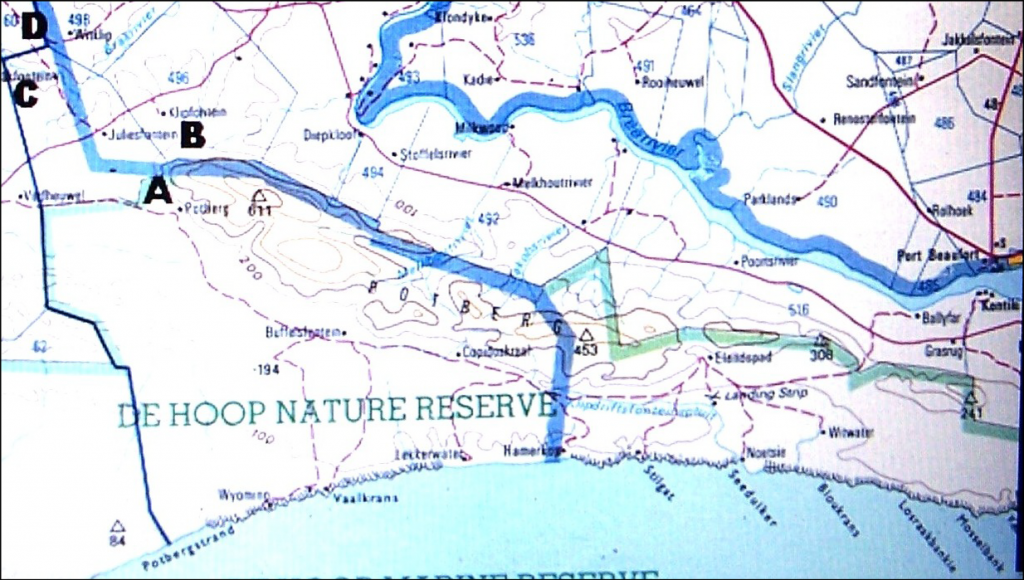
Where are we? Geography plays such a critical role in my perception of the species that it is important to try to understand why and how it touches on the issue. The Langeberg Mountains is a Table Mountain Sandstone about 10km wide and 1500m high running east-west and separating the inland Little Karoo from the Lowland Renosterveld of the southern Cape. There are five main travel routes through those mountains of which two are via river gorges viz. Cogmanskloof, Tradouw, and then there is also the Gouritz River Gorge where there is no road. Haworthia is not generally considered a sandstone and high mountain species because, firstly, there are few records to suggest that and, secondly, because they are averse to the higher moisture levels. However, there are many records in the low and close foothills. H. retusa ‘turgida’ is recorded in the higher areas and in the vegetation associated with the sandstone viz. the Fynbos. Fig. 1 is a view taken looking eastwards from Kleindoorn (Kleindoornrivier). This is about 16km east of Barrydale that sits at the northern end of the Tradouw Pass. The next farm is Brandrivier (B) and beyond that is Springfontein. Muiskraal is marked with an “M” and this is at the northern end of Garcia Pass from Riversdale to Ladismith. The “O” marks Oskop which is beyond Zandkraal and about 15km beyond Muiskraal. Another 10km will take you past Waterval and bring you to Aasvoelvallei at the confluence of the Grootriver with the Gouritz. The last stretch is another 10km over the Cloete’s Pass to Herbertsdale from where one travels back westward 7km to Towerlands.
What is there? I speak of species complexes because the classification and naming of Haworthia seems to defy any agreement. I do not think the nomenclatural system allows it either. South of the Langeberg are the H. retusa and H. mirabilis complexes as well as H. minima and H. pumila, as main role players. North of the mountains is H. arachnoidea and H. mucronata. There are incursions and crossing of the mountains by three of these and my contention that they do not all do so is simply because we do not see beyond the narrow confines of our classification system. North of the mountain there is enough evidence to suggest that H. mucronata and H. arachnoidea is very probably the same one complex despite the few situations where they manifest in near proximity. South of the mountains H. retusa and H. mirabilis have an even more complex relationship. It is difficult to begin to realize that there is a very small step between their manifestations south of the mountain to an integration of them both in H. emelyae that occurs north. I have great sympathy for those who contest the issue. I hope that the figures I present here, as a very small set of images from what may be a very small sample of existing populations, will help bridge the difficulty.
My field experience, however much field experience is a measure of any persons competence to process the product, suggests that there is a predictable coherence in the variation that one observes as one moves from discrete population to discrete population. This progressive variation is also driven by surface geology and habitat. It becomes no problem at all but to realize that as another set of samples, the Aasvoelvallei H. emelyae (see MBB 7850 figs 2a-l) is nothing more than the Towerlands H. retusa ‘turgida’ (MBB7849 figs 3a-b) in another guise. It is the habitats that are so vastly different. Towerlands is south facing mountain slope in sandstone and Fynbos vegetation next to a perennial mountain stream. It was no chance that the rain that caught us there stood in such sharp contrast to the warm sunshine that greeted us an hour later at Aasvoelvallei and H. emelyae a mere 10km away as the crow could fly. That habitat is Bokkeveld shale with a heavy overlay of the very white quartz that veins this formation, and the vegetation is Karoid.
2a. MBB7850 H. emelyae Aasvoelvallei.2b.2c.2d.2e.2f.2g.2h.2i.2j.2k.2l.3a. MBB7849 Haworthia retusa ‘turgida’ Towerlands.3b.
H. mirabilis ‘maraisii’ is present north of the mountains at Montagu and towards Montagu from there to Barrydale. It is also present at Barrydale itself. At Kleindoorn we have an enigma (yet another) where the plants (MBB7847 figs. 4a-s) are very similar to the Barrydale plants and they also seem to flower in summer. But these same plants can be likened to H. emelyae ‘major’ at Muiskraal that flowers in the spring as does H. emelyae (and H. retusa). Figs. 5a-c MBB7844 shows plants at Brandrivier. The dramatic thing is that these are recent photographs and very unlike the plants observed there by me 30 years ago and recognized as H. emelyae ‘multifolia’.
4a. MBB7847 Haworthia emelyae Kleindoornrivier4b.4c.4d.4e.4f.4g.4h.4i.4j.4k.4l.4m.4n.4o.4p.4q.4r.4s.5a. MBB7844 Haworthia emelyae ‘multifolia’ Brandrivier5b.5c.6a. MBB7846 H. emelyae ‘multifolia’ W Springfontain6b. MBB7846 Haworthia emelyae ‘multifolia’ W Springfontain6c.6d.6e.7a. MBB7847 Haworthia emalyae ‘multifolia’ E Springfontein7b.7c.7d.7e.7f.7g.7h.7i.7j.7k.7l.7m.8a. JDV97-44 Haworthia emelyae ‘breueri’ E. Zandkraal8b.8c.8d.8e. JDV97-44 Haworthia emelyae ‘breueri’ E. Zandkraal8f.8g.8h.8i.8j.8k.8l.8m.8n.8o.8p.
Figs 8 are of JDV97-44 (originally GDM429) plants from beyond Muiskraal (where both ‘multifolia’ and ‘major’ occur in disparate habitats) at Zandkraal, where there are three populations to which an appellation ‘H. breueri’ has been fabricated. I downsize that to H. emelyae ‘breueri’ to bring some perspective back to Haworthia nomenclature. I do not include field photographs for Oskop and Waterval, but add two from south of Vanwyksdorp (see figs 9a-c ADH2966) where H. emelyae originated according to the collector (this was a Mrs. le Roux and the plants were sent to Smith via Mrs. Emely Ferguson). The photographs were taken just south of Vanwyksdorp by Adam Harrower and I do hope that readers will realize from this article that it is not at all strange that the von Poellnitz published illustration and type of H. emelyae (figs 10a-b) are so often said to be of H. retusa ‘turgida’. Curiously, H. Fourcade illustrated a plant (his no.144, see fig.11) of H. retusa ‘turgida’ and another (his no. 82, see fig.13) of H. reticulata as H. emelyae. This says a lot for subsequent attempts to interpret and argue vehemently about von Poellnitz’ H. correcta (see fig.12).
9a. ADH2966 Haworthia emelyae S Vanwyksdorp9b.9c.10a. Haworthia emelyae Von Poelln.10b. Haworthia emelyae Von Poelln. Type.11. HGF144 Haworthia retusa ‘turgida’ as Hawortha emelyae12. Haworthia correcta V. Poelln. Type.13. HGF82 Haworthia emelyae ‘turgida’ as Haworthia emelyae
What really struck me on this recent excursion was just how much inviting territory is unexplored. In contrast to that is Kleindoorn where we photographed those plants actually found by Gerhard on a previous visit there with Bob Kent and Kobus Venter many years ago. The plants are on the south side of a sandstone ridge. We could not find them elsewhere on our recent visit and it seems that suitable habitat is just not there for them. There is a most remarkable divide of shale and sandstone on Kleindoorn and while there is/was a form of H. arachnoidea in the sandstone (repeated at Zandkraal), only H. arachnoidea ‘nigricans’ is in the shale. At Brandrivier, E. Springfontein and Muiskraal the ‘multifolia’ is in a sandstone pebble layer that overlays the shale in the manner of the Tertiary terraces south of the mountain. West of Springfontein the plants are again in shale, but without the white quartz of other karoid sites mentioned.
Acknowledgement It is not possible to do this fieldwork without the goodwill of landowners that is threatened by a glut of collectors who often trespass. Therefore I particularly want to thank Bertus le Roux of Kleindoorn, George and Jaco Nel of Brandrivier, Louis Botha of Springfontein, Anton Bredell of Aasvoelvallei, and Gregory Nicholson of Towerlands for access to their property. Gerhard Marx was contributory as was Kobus Venter on a previous excursion. Adam Harrower kindly sent me pictures from his records. ♦
1a.View from above Onverwacht North Westward to Ladismith1b. View looking south west over Muiskraal to Garcia Pass
There used to be a regular bus service between Riversdale and Ladismith and J. Dekenah made use of this for his excursions into the Little Karoo to find plants for G.G. Smith. He thus discovered H. emelyae ‘major’ at the northern mouth of Garcia Pass. He also submitted a single specimen of a plant collected from the karoid veld a little further north. It is that record that suggested to me that ‘major’ was linked to H. emelyae further east and north as well as to ‘multifolia’ to the west. Etwin Aslander found what I regarded as the equivalent of ‘multifolia’ (see figs 2a-i) on a low plateau north of Garcia pass on the farm Muiskraal that is at the foot of the mountains north of Garcia Pass.
Johan Meyer of Barrydale once showed me a cultivated plant that he claimed he collected on the slopes to the north of Etwin’s locality. This was a robust and rather tuberculate plant that was very reminiscent of the forms of H. emelyae at Zandkraal about 12km east. In an earlier chapter I wrote more widely about the problem of land ownership and access and eventually felt that I should look into the situation at Garcia Pass in respect of conservation status of ‘major’, when an opportunity to break the impasse arose.
Fortunately I have quite a long association with Muiskaal and its owners, since I was at the Karoo garden and Mr. Henry Chamberlain, owner of Muiskraal, finally allowed me access. We visited both Etwin Aslander’s population (MBB7858 see figs 2a-i) as well as locating a population on the northern slopes as indicated by Johan Meyer (MBB7859 see figs 3a-i).
3d. MBB7859 Muiskraal3f.3g.3h.3i.1d. Looking eastwards over Muiskraal1e. Looking southwest to Garcia Pass from same spot as above
In addition we also found a population immediately west of the farmstead (MBB7857 see figs. 4a-f). Particularly interesting is the dramatic switches in dominances of shrubby plant species that must be a factor of differences in the very skeletal soils as well as how plants compete with one another (allelopathy). The plants in all three populations are of course similar to the east of Springfontein plants in as much as one generates a mental common denominator for a set of variable plants. This similarity must surely be seen in the similar geologies of the three sites which are on tertiary depositional terraces. We also explored extensively along the foot of the mountain between Muiskraal and Zandkraal without any success.
1f. West Muiskraal
4a. MBB7857 West Muiskraal4b.4c.4d.4e.4f.
However, we also made contact with the owner of the upper areas around Garcia Pass itself and were able to explore both sides of the road at the ‘major’ locality (see MBB7861 see figs 5a-l) as well as the range eastwards.
We did find a new population some 3km east of the farm Onverwacht (see MBB7860 see figs. 6a-i) and Gerhard Marx reports seeing it immediately north of the same farm. We did on a subsequent visit briefly get there, but did not find plants. Some of the plants were very ordinary and it is not surprising that J.R. Brown, in commenting on his collection, felt that he had two species viz. H. ‘major’ and H. maraisii as he might have named them. Definitely the plants I saw could have been mistaken for individuals in several populations in the sandstones south of MacGregor or even nearer Villiersdorp (Moddergat). Unfortunately my camera failed me and I did not get many pictures, but one can see that the plants can have relatively spineless leaf surfaces.
There is something curious about the flowering times. The ‘major’ plants had evidence of recent flower stalks on a few plants while some plants were also beginning to flower. The lower populations at Muiskraal and Zandkraal were already shedding seed. I think it is significant to point out the geological features that I think are the main drivers of ‘difference’. Garcia Pass is a rift through the Langeberg Mountains that are uplifted sand stones. The northern foothills are an eroded remnant of tertiary deposits that overly Bok keveld shale. The Zandkraal populations are associated with quartz outcrops or variants of such as are common throughout the Little Karoo. The vegetation differences between the sandstones, terrace gravels and shales are astounding and illustrated in the fact that botanists, following their original intention that a biome was a major association of plant species driven by massive and extensive landforms and climate, regard the Karoo vegetation and Fynbos vegetation as belonging to two different biomes. The terrace vegetation is primarily Renosterveld, but it has elements of what is now regarded as the Thicket biome. It would not be a serious overstatement of fact to say that no two plant species occur in each vegetation type. The farmer on Onverwacht (Fynbos) is a flower farmer picking from the field, while the present farmer on Muiskraal (generally Karoo) grows olives and grapes. Muiskraal used to be primarily a sheep farm and Onverwach barely.
Despite this observation on compositional differences of the vegetation, or because of it, my contention is that ‘major’ is a variant in the sandstones while ‘multifolia’ is an expression of the same species, H. emelyae, on the tertiary debris. They both go on to varied forms in quartzitic patches in the shales. In my opinion H. emelyae is in a continuum that is formed out of the H. retusa and H. mirabilis complexes south of the Langeberg Mountains, where broadly speaking H. emelyae moves into sandstone as ‘major’ and the other two species occupy that terrain as ‘turgida’.
Acknowledgement: Sincere thanks to Mr. Henry Chamberlain of Muiskraal and Mr. Pierre Vermaak of Onverwacht. Gerhard Marx also motivated with interest and information. ♦
It is always assumed that botanists have a good grip of their subject, as one supposes for the scientists in other disciplines. Any science is the advance of knowledge by observation, hypothesis and testing by systematic enumeration and experiment. This is furthered by replication and review by other scientists. Botany is far behind the exact sciences, because of the very nature and complexity of living things and systems, and also behind because zoology where animal life is obviously more organized than is the case with plants and where more attention is focused.
1. Looking down the north face Die Nekkies east to west2. Same spot looking east northeast
It has always been recognized that South Africa has a unique flora and traditionally the difference from other floras was attributable to the extraordinary richness, diversity and endemism of the so-called Fynbos that came to be regarded as the Cape Flora. Just what the Cape Flora comprised was extraordinarily vague and the early limits were stated in terms of latitude and longitude. In the mid-20century it was recognized that South Africa should be divided into biomes, as major components of the vegetation, and these were defined as units with wide and expansive boundaries. So while the Fynbos biome was recognized among others, no connection seemed to exist between its boundaries and those of the major geological systems. The essential problem is that the boundaries of this biome are by no means wide and expansive, because one can literally step from Fynbos vegetation into Renosterveld, Sandveld and even Karoo. This is no more dramatically demonstrated than at Die Nekkies where it is obvious that the vegetation differences are primarily a result of geological substrate fortuitously linked to rainfall gradients determined by continental land-form, latitude and topography.
3. Same as figure 1, but from further east4. Still further east looking west northwest5. Looking north with M marking the location of Haworthia maculata, K – Karoo Garden6. Near east end with P marking the site of Haworthia pubescens, R indicating Rooiberg to the east7. View south to Villiersdorp
The significance of Die Nekkies is that it is formed from a narrow band of the Witteberg rock formation distorted in two transverse planes of the folded Cape Sandstone systems. Die Nekkies hills are directionally east to west and tilted down from the south. Northwards they are lost under the flood plain of the Breede River. Eastwards and southwards there is a wedge of Witteberg and Karoo formations that drives against the Cape Sandstones all the way south west to Villiersdorp. The consequence is a dramatic vegetation barrier related to the very different soil chemistry of Witteberg quartzites as opposed to Cape Sandstone, which is as dramatic where Bokkeveld Shale and Dwyka Tillite meet.
What is in effect the case, and apparently not really conceded by botanists, is that there are primarily two floras in a single winter rainfall biome. Some botanists insist that vegetation differences are simply fire driven. Here it is again difficult to separate the consequences of different fuel loads due to the substrate characteristics and the plants adapted to each substrate from the effects purely due to the fire, but it is nevertheless quite obvious that the major differences are substrate.
This is all a very broad view of a complex phenomenon that emerges when one begins to look at the individual species components of the vegetation. Vegetation classification is really based on dominant species and seldom has any attempt been made to understand species distributions that do not coincide with broad general pattern. When this has been attempt-ed it has been very seriously flawed by the problem of identification. This is a matter that botanists seriously underestimate and consistently fail to address.
It is simply assumed by general botanists that taxonomists in herbaria are competent and able, and that revisionary work and primary taxonomy have kept pace with technological progress and research in all other fields. This is not the case. Taxonomy has for many years been a forgotten science and less and less resources are allocated to either revisionary research or identification. It is simply not possible to collect, mount, store and record the variation of even a fragment of the species flora within any herbarium structure. A related fact is that there is no definition of what a species actually is. Very often it is just assumed that a definition is in place that is satisfied when some, usually morphological, criterion suggests that a specimen and sometimes more than one specimen does not agree with some ill-defined norm dependent on the taxonomist and the herbarium in which he/she is stationed.
It is self-evident that in the nature of evolutionary processes that are presumed to take place, that variation must be intrinsic to any species. This means that species need to be considered as systems that respond and change according to events that also change substrate and climate. It also means that species systems cannot be the same and a model (definition) needs to be in place that recognizes that each species (system) may be in an individual relation to time and space.
Two examples from Haworthia can be given for the significance of Die Nekkies in terms of species diversity and distribution and the difficulties that poor taxonomy introduces to obscure pattern. The first is for Haworthia maculata. Where there is already considerable doubt as to a proper classification of the Alooideae in the Asphodelaceae, Haworthia is persistently seen to be a single natural group. While there is no definition in place for the species category, the genus category is assumed to have some reality. It in fact does not. It is quite evident that some species do have very close affinities that would satisfy the perception that these form good genera, there are many species that elude such simple categorization. In Haworthia, as it is traditionally constituted, there are three distinct sets of species based on a range of characters both vegetative and floral. It is an extraordinary reflection on botanical classification that this has never been properly accepted. The genus Aloe is widely distributed in South Africa and more than 150 species are recognized. However, only two species can be seen as strictly Cape Sandstone and hence Fynbos “biome” species, although several are closely associated. There is only one species of Haworthia subgenus Haworthia that can truly be said to be a Fynbos species and that is H. nortieri. This subgenus is poorly represented north and west of the Fynbos “biome”, but it is represented by a vast array of variants in the Eastern, Southern Cape and Little Karoo that are difficult to aggregate into species. The variants seem to be ecotypes and it is such ecotypes that do adapt to sandstone habitats. There are none such variants in the sandstone mountains south and west of Die Nekkies where only one “species” viz. H. maculata occurs in localized population at several points along the range. This “species’ is very closely related to H. herbacea that is very common on all the other skeletal soils to the east, both north and south, but very rarely penetrating sandstone habitats. Where H. maculata occurs on the eastern limits of Die Nekkies, there is a subtle change of rock and shallow soil to a more Karoid habitat, and the plants tend to be more clump forming and spinose, suggesting a shift to H. herbacea. This similarity shift is replicated in H. herbacea where it occurs east of the brickworks near the Brandvlei Dam wall. Ecotypes similar to, and classified as, H. maculata occur on the Witteberg formation hills southwards to Villiersdorp, but never in close association with H. herbacea. Curiously, a very similar variant to H. maculata occurs again high in the sandstones north of Worcester on the Audensberg peaks and also lower down in the Sandhill area. Thus H. maculata is a significant marker in the relation of the Asphodelaceae and Haworthia to the distinction of “Succulent biome” and “Fynbos” biome.
8. MBB 7526 Haworthia maculata Die Nekkies, Brandvlei Dam9. MBB 7526 Haworthia maculata Die Nekkies, Brandvlei Dam
Haworthia subgenus Hexangulares is represented by only one species in the south western Cape. This is Haworthia venosa that occurs only in rock faces flanking the lower Breede River and Gouritz River. Its related species H. tessellata is a Karoid element. Haworthia subgenus Robustipedunculares is tightly allied to a winter rainfall “biome”, but consists of only four species. Three of them are endemic to the Southern Cape and three of these do not extend further west than Ashton and Bonnievale. H. pumila is abundant in the Worcester/Robertson Karoo and extends as far afield as Laingsburg, Anysberg and Barrydale, but south of the Langeberg it is not nearer to Swellendam than at Drew Station. South of the Riviersonderend it is only at Stormsvlei, near Protem and at Vaandrigsdrift. It does not occur on the Witteberg formations south of Die Nekkies, but it does occur on the eastern lower slopes of Die Nekkies itself. So here again Die Nekkies is the evident limit of this species too – the second example.
Possibly the very dramatic case of Drosanthemum halli and D. bellum more clearly suggests the significance of local diversifying factors. D. halli is yellow flowered and abundant whereas D. bellum is pink and localized. They do not co-occur. It can be dramatically shown that D. hallii is nothing but a geographic variant of the bi-coloured D. micans that occurs north and west of Worcester, and that D. bellum can only be different because of very site specific factors.
H. maculata at the western end of Die Nekkies appears to be a solitary plant but, as it occurs more eastwards, it becomes proliferously clumping as the vegetation also becomes increasingly Karoid. Why this should be so is not clear.
The many accompanying illustrations (MBB7526 figs. 8-38) show the scenery and topography and the range of variation in H. maculata. One can see that it is at home in both the Restiods of the Fynbos and under Pteronia paniculata of the Karoid.
All the following photos are MBB 7526 Haworthia maculata. Die Nekkies, west end.
Acknowledgement I would like to thank Anso Le Roux of Pokkraal, Rawsonville for drawing my attention to H. maculata much further west along Die Nekkies than previously recorded and for inspiring a walk along the full eastern limb. ♦
It is very difficult to write about particular plants when one has to contend with the fact that one is not sure what Latin names may mean to the reader. I wrote a piece with closure in mind and thought I would send it to a non-taxonomist botanist whom I think is a tribute to the profession. Extracts from his response are “I am an ardent supporter of your species concept and couldn’t agree more strongly with your statement that without variation, there would be no evolution… I do agree that the pervasive species concepts force us to ignore the most interesting and productive research avenue: documenting and understanding variation in the field.”
It is worth considering what he has said about “pervasive species concepts” and just what he might mean when he says they force us to ignore documentation and understanding of variation in the field. It seems to me that the converse is the truth. Failure to properly understand and document variation has contributed to the pervasiveness of false concepts which fly in the face of science. I keep harping away at this question of lack of definition, because it is not apparent to me that any reader appreciates my point of view. To my mind a key issue is made of nomenclature and the rules that govern it, and very little attention at all is given to whether these Latin names help our understanding at all. I feel that my contribution contributes mostly to documentation of variation and that I cannot do much in respect of understanding. This short article should explain why.
I have tried to explain what I have observed in the three species Haworthia cooperi, H. cymbiformis and H. transiens as I understand the various populations that might be seen to constitute three discrete systems. My problem is that they do not necessarily constitute such discrete systems at all. Even considered together as one system, it is not entirely sure in my mind that they are truly separated from any other species belonging in the same sub-genus Haworthia. It is not improbable that they diffuse into H. decipiens and into H. mucronata that are in turn diffused into other elements. If one considers that a species definition based on breeding true has been “… used by most taxonomists since 1682.” (I quote this from a prominent taxonomist), one must recognize that taxonomists have then been ignoring the innate variability which drives change, adaptation and evolution. I am not sure that I have not been making the same mistake although I have consciously tried to recognize that discontinuities of any kind do not necessarily suggest discrete species.
Someone once wrote that “specialization leads to extinction”. If species become too specialized, they lose the capacity to adapt and evolve according to an ever-changing environment. At the other extreme, species that can maximize variation within their individuals and populations will benefit most in those changing circumstances. If taxonomy has been guilty of paying insufficient attention to variation as a concomitant of evolution in present time, it may mean that we may generally be recognizing species at the wrong level. We may collectively be attaching far too much attention to morphological and even genetic differences without due account of distribution and potential for change both internal to the species and external from the environment.
Therefore, I am deeply skeptical that my classification of Haworthia is a true reflection of the situation in respect of actual species. I have been influenced by a paradigm in taxonomy that genera and higher taxa were largely artificial and determined by arbitrary choices that they should be approximately equal in size and number. Without taking this too seriously, there is a huge problem when it comes down to species and to consider that some in a genus may be tightly grouped in terms of limited variation and distribution, while others form vast complexes.
Jan Vlok kindly sent me a picture of a Haworthia from the Little Langkloof. This is the upper reaches of the Keurboomriver Valley which runs inland from Plettenberg Bay, turning west north of the Knysna area and cutting across the Prince Alfred’s Pass road between Knysna and Uniondale. The only Haworthia species in the area are H. transiens and H. scabra and they are abundant there. Jan said the plant was in Karoid vegetation and was generally solitary. I suggested that it was probably an ecotype of H. transiens and that otherwise it may have been H. cooperi var. gordoniana. I have written an account of the interaction of those elements in the Baviaanskloof to the north and west, and it is quite evident to me that there is a close relationship. It should be noted that there is also a major variant of H. scabra closer to the Prince Alfred’s Pass road, so that there is a precedent in adaptive change. I was recently able to visit the area myself and confirm that the plants are in this very same category of interaction between H. cooperi and H. transiens. Thus, in my opinion, they parallel the kind of relationship other variants of H. cooperi has with H. cymbiformis, and in a changed dispensation for taxonomy it may be agreed to regard these two species and H. transiens as one species. It is possible that continuity may still be found with H. mucronata further west. While I write “changed dispensation”, I regard it as essential that it first be understood that there is still a vast amount of exploration and recording to do and that any final solution will only be achieved by collaborative effort and agreement. Professional taxonomists will have to make the decisions, but they will have to first prove themselves competent to do so.
Kliprivier plants in cultivation.
The above was the article as printed in Haworthiad. It deals with my collection MBB7586 (see figs. 1a-e) from east of Die Vlug in the Prince Alfred’s Pass south of Uniondale. My conclusion was that H. transiens was but another expression of H. cooperi at the western end of its distribution zone and possibly transition to H. mucronata.
I had collected seed that I asked Etwin Aslander to grow, which he did very successfully. The seedlings were initially extraordinarily variable and there were some striking plants that gave indications of reticulate marking that would have shamed H. cooperi ‘picturata’. After Etwin transplanted them the plants looked much more ordinary and I selected a few to grow myself. So this short note is just to present pictures of these and demonstrate the badly beaten horse of variability.
Figs. 1 are a series of pictures of plants in the field that can be said to be representative without leaving out too much. Figs. 2a and b are of my permit limited collection grown in my outside rockery in relatively full sun. Figs 2c. to 2h are the selected clones that I grew on to show readers what happens between field, nursery and greenhouse table. Any wonder that you can justifiably complain when you do not get what you think you ordered?
I originally omitted names and would now use the name H. cooperi ‘transiens’.
Figures 2a and b are of my permit limited collection MBB 7586 of Haworthia cooperi ‘transiens’. Kliprivier grown in my outside rockery in relatively full sun.
Figs. 2c to 2h. Selected clones of plants grown from habitat seed in cultivation, grown on to show readers what happens between field, nursery and greenhouse table.
Acknowledgement Jan Vlok very kindly informed me of the existence of this locality east of Die Vlug. ♦
How did this start? Is it possible to see anything with such a quick peek? Somewhere in my memory bank is my stated opinion that understanding H. arachnoidea would assist any botanist towards a better understanding of classification. This was long before I came to see that this is a lot more necessary than I thought then. It is not just botanists who seem dumbed down to the reality of a diversity that is necessary for response to change and largely denied by the nomenclatural code and how it is practiced. We have not understood change or what changes can occur. During perhaps only the last 15 to 20 years has it become apparent that changes are cataclysmic and frequent. I will skip the fact that the violence of catastrophe, or the drivers of catastrophe, may also induce change at even a cellular level.
I usually go into the field with a specific goal while also carrying a host of peripheral questions in my mind. While exploring the problem of H. schoemanii, I was thus also thinking of Conophytum. After photographing a species seen at Laingsburg, I mentioned to Steven Hammer that I had also seen it in the southeastern Tanqua Karoo (Bakoven). This was news to him and so in my wish to re-establish the reality of H. venosa subsp. granulata recorded at Patasriver (actually Patatsriver road and that is another story that will be told) as well as the Conophytum for Steven, I undertook an expedition to the Tanqua.
It was co-incidence and luck that brought us into contact with Dr. De Ville Wickens, who is an expert on the geology of the Gondwanaland connection to the Tanqua mountains and who now owns Bizaansgat to the northwest of the farms Droogekloof and Bakoven. Dr. Wickens facilitated our expedition by offering us accommodation at Bizaansgat and we were thus able to spend more time there than otherwise.
1. ADH3140 Haworthia nortieri Skitterykloof2.
Derek Tribble did alert me to a small Haworthia in the Skitterykloof about 20km to the west of Bizaansgat. I had myself seen the plant and concluded that it was H. arachnoidea. Fairly recently Adam Harrower gave me a small plant from the vicinity of Skitterykloof and I confess that I was a bit confounded because it did implicate H. nortieri (See ADH3140, figs 1 & 2) particularly because the leaves were so spotted. Why I say this is because the combined presences of the look-alikes H. arachnoidea and H. nortieri ‘Albispina’ at Koup and elsewhere, as well as their often superficial resemblances, now make me a little wary of snap decisions. I later concluded from the flower that Adam’s plant was in fact H. arachnoidea, and not because I am convinced I could separate the two on the basis of the flower only. So this report covertly deals with the reality of a massive variation within H. nortieri and also H. arachnoidea and hope that readers will generate their own open mindedness to realize that I am drawing conclusions here that could or should be better substantiated. I also hope that this does not mean that a whole new set of Latin names is required to try and do so.
Because my initial field work was to obtain an overview of the genus, I soon stopped paying much attention to H. arachnoidea, because it seemed to be so ubiquitous. A big mistake! Unfortunately digital cameras were then not available and space in the herbarium (contrary to what my learned friend has to say) was and still is at a premium. The consequence is that I do not have a proper physical or mental record of all the plants and populations that I might have seen. In fact I cannot even fully remember how or why I linked H. arachnoidea to H. decipiens the way I did in my Handbook.
What happened on this expedition is that at Droogekloof we were surprised to find a very small Haworthia that I take to be H. arachnoidea (see MBB7644a, figs 3a–j). It is important to note the differences in colour, spination and leaf-spotting and/or reticulation. These plants were single and scattered over a fairly wide area. They varied in size from as little as 20mm to an average of about 30mm diam and become extremely cryptic in the dry summer.
3a. MBB 7644a H. arachnoidea Droogekloof3b.3c.3d.3e. MBB 7644a H. arachnoidea Droogekloof3f.3h.3i.3j.3g.
We continued to Bakoven and found it again (see MBB7644, figs 4a-e). Fig. 4c. is a view from a mountain top at Bakoven looking NNW towards Droogekloof and then Bizaansgat in the distance. Fig. 4b illustrates the weak inflourescence with few flowers generally true for many of the populations we observed. We the retraced our steps and saw similar plants approximately midway back to the Droogekloof population (MBBsn see fig. 5). The next day at Bizaansgat we saw exactly the same thing at our first three stops and then came across an absolute field of them at a fourth. Again at several more stops on Bizaansgat (MBB7646 see figs 6a-h) and yet again on another farm 5km to the east.
4a. MBB7644 H. arachnoidea Bakoven4b.4c. View northwards from Bakoven4d. MBB7644 H. arachnoidea Bakoven4e.5. MBBsn H. arachnoidea Droogekloof6a. MBB7646 H. arachnoidea Bizaansgat6b.6c.6d.6e.6f.6g.6h.
The next week took us on the road, the Patatsriver road, from the southern Tanqua to Matjesfontein east of Laingsburg. We stopped on the advice of Gerhard Marx at a well known spot at Perdekraal where Lithops, Didymaotus and Tanquana occur. We saw the same small H. arachnoidea there (MBB7658 see figs 7a-c).
7a. MBB H. arachnoidea Perdekraal7b.7c.
From there we turned north to Bantamsfontein for H. venosa subsp. granulata and we were surprised to find again the same H. arachnoidea at every place we stopped on the farm (MBB 7660 see figs 8a-e).
8a. MBB7660 H. arachnoidea Bantamsfontein8b.8c.8d.8e
We travelled on in the direction of Matjiesfontein to Patatsriver and again found it ubiquitous (MBB7666 see fig 9).
9. MBB7666 H. arachnoidea Patatsrivier
We went on to Keurkloof just northwest of Matjesfontein, and found it on the two hillsides we searched (MBB7670 fig.10).
10. MBB7670 H. arachnoidea Keurkloof
From there we went to Bulhouer, which is north Matjesfontein, and found it at four places (MBB7671 & 7672 see figs. 11a-d and 12a-e).
11a. MBB7671 H. arachnoidea Bulhouer S11b.11c.11d.
It is very important to note that Emil Heunis recorded the form ‘scabrispina’ on this farm Bulhouer and this is what I was expecting to find. However, my four samples were observed at regular intervals of about 1km from south to north. The southern populations were indistinguishable from any we had seen from Perdekraal in the west, but the last population, (MBB7672 figs 12a-e), were as close to as must be possible. My contention is that we thus observed continuity from the west to the east.
12a. MBB7672 H. arachnoidea Bulhouer N12b. MBB7672 H. arachnoidea Bulhouer N12c.12d.12e.
On northwards to Dwarsindieweg some 15km further and we found the small forms (MBB7677 see figs 13a-b) en route as well as commonly at Josephkraal another 16km east (MBB7678 see fig. 14). We found it again the next day north of Dwarsindieweg (MBB7686 see figs 15a-d) on the high upper slopes of Klein Tafelkop (MBB7688 see 16 fig 54). Heading for home we stopped on a backroad on the way to the Patatsriver road at Witrantjies and found it yet again (see figs 17a-c). We had been very cautious all the time to be sure we were dealing with one thing and not two. On this last stop we were confronted with the first drama when it seemed fairly certain that we had also found H. nortieri and this turned out to be false. We did return later to climb a high mountain at Volstruisfontein west of Dwarsindieweg hoping to find H. marumiana. Instead we again found small H. arachnoidea (MBB7802).
13. A view from south Dwarsindieweg.13b. MBB7677 H. arachnoidea Dwarsindieweg S14. MBB7678 H. arachnoidea Josephkraal15a. MBB7686 H. arachnoidea Dwarsindieweg15b.15c.15d.16. MBB7688 H. arachnoidea Klein Tafelkop17a. MBB7693 H. arachnoidea Volstruisfontein18a. KG16/72 H. arachnoidea Ladismith18a. KG16/72 H. arachnoidea Ladismith18b.18c.
It was not flowering time. The pictures that should have accompanied this article will show that the arachnoidea we found were often still accompanied by an odd old inflorescence. A weak spindly thing with evidence of seldom more than three old capsules.
Generally the plants were very small and we very often came across seemingly mature plants that were down to 20mm in diameter, and seldom exceeding 35mm. Invariably we could find the odd bigger plant in any single population array. The spination was fairly variable in respect of size, spacing and arrangement. The leaves could be short and squat or narrow and elongate. Colour was usually on the bright green side and I do not know what the summer colour would be. Leaf markings were interesting because often there was a slight reticulate pattern to the leaves and translucent dots and stripes and margins could be observed. This seemed to diminish in a cline from west to east. We did conclude, because this is what we also wanted to observe, that there was some evidence for continuity with H. arachnoidea var. scabrispina. There was the one population, however, north of Bulhouer where the plants were indeed bigger (50-60mm diam) and there was a definite tendency to form overarching brown spination so characteristic of scabrispina. There were of course the odd variants elsewhere and one group of four plants east of Dwarsindieweg could have come from the very confounded interface of H. arachnoidea and H. mucronata that is covered in the Latin H. arachnoidea var. nigricans. Readers should recognize that there is no way in which the nomenclatural code can be used to account for this diversity and variation, and it is time that more space and recognition is given to this very common phenomenon in the general context of writing, speech and communication.
The pictures will include one of ‘candida’, an epithet conveniently supplied by M. Hayashi for a variant of H. arachnoidea that occurs very close to Ladismith (KG16/72 see figs 18a-c). The spines in these plants often are black at the tips and the same phenomenon can be observed in massive plants that occur near Worcester in the “typical” variety arachnoidea. But I would like to break away from this usage. The epithet arachnoidea is applied to every single plant of that species wherever it occurs. Varietal names of any kind may be attached and I especially recommend the form and the formalization that Gordon Rowley advocates too; this is the format example…H. arachnoidea ‘scabrispina’. For more precision a whole host of abbreviated prefixes could adorn the label e.g. cf ‘scabrispina’ meaning “compare with” or v ‘Scabrispina’ as a cultivar or var. scabrispina as a generalisation.
There are a set of varietal names in H. arachnoidea already, and the proper address for these small Tanqua plants discussed in this article is in terms of my own Revision H. arachnoidea v. namaquensis. I do not think that my dispensation is adequate or good enough for the scale of our present knowledge, but neither do I think formalization is going to do much more than arouse an egocentric mass of still more Latin epithets. So I strongly suggest that we as a community simply accept something like H. arachnoidea ‘tanqua’ or ‘minuta’ and drop any pretension that Latinization adds any lustre or glamour to plants that are already fascinating and attractive. One of my objections to what I find a really irksome process of formalization (typification, full citation of basionym and all that stuff), almost senseless, because unless a microchip is also implanted it is unlikely that there will be any chance of associating any plant out of habitat back to its origins.
I am not going to write any further about H. nortieri cf ‘pehlemanniae’ here because I need to know more as well as confirm something about the flowers and flowering times of both sets discussed here. There were the occasional dark coloured plants in the arachnoidea (and excuse me dropping the ‘H’ – it really is not necessary in this informal environment), but I doubt very much that anything further will support the suspicion that arachnoidea will now always be confusable with nortieri. It turns out that what I though might be H. nortieri because of the very grey colour tone of the leaves was in fact also H. arachnoidea. A specimen flowering in cultivation produced non-globose flowers.
Acknowledgement: Dr. De Ville Wickens, Bizaansgat; Mr. P Schietekat, Stellenbosch; Mr. M. Jan duToit, Remhoogte Boerdery; Mr. Pieter de Graaf, Patatsriver; Mr. C.K. Francois, Bulhouer; Mr. Wynand Theron, Dwarsindieweg; Mr. Johan Kriel, Citruspoort (Volstruisfontein); Messrs. Stephen and Gail Louw, Keurkloof. Hospitality and access to properties is deeply appreciated and valued. ♦
I have become increasingly concerned about the poor relations that exist between collectors and the authority of Nature Conservation. The argument that collectors threaten and despoil natural populations is very real and I do not dispute at all that Conservation authorities have a very valid complaint. They have a function to perform. On the other hand there is an interaction between human beings and nature in all its forms that should be fostered to the benefit of both sides.
Nurseries, traders and collectors are as much of the picture as are conservationists, institutions, researchers and landowners. It is unfortunate that there is no non-government party that lobbies for the rights and activities of the former group, but it is not my intention nor within my competence to argue all the aspects of the case.
I strongly believe that people have the right of access to nature in all its forms and the issue is one of individual responsibility and proper consideration of consequences. An appreciation of and sensitivity to nature should be reflected in whatever we do in our lives. My own collecting impulses led me to institutional employment where I could exercise my interest to what I thought were efforts more worthy than my personal interests. From that position I also did try to share and extend privileges to a wider circle. It is in this way that I became involved with Sheilam Nursery. It was not my wish or intention that my collection should have come to be housed there. However, Sheilam has succeeded over a period of nearly 40 years to maintain a fairly true record of my collections obtained as propagated material from the Karoo Garden at Worcester. My offer of permitted collections dating from my revision of Haworthia in 1966 to the Karoo Garden was rejected and for a while resided with Etwin Aslander at Brackenfell. It has since passed to Garth Schwegman at Sheilam who has taken a particular interest in the maintenance and propagation of that collection.
Hand pollination of haworthia has become the norm and Sheilam has made a massive effort to generate plants from this. Of course growing plants from seed is a lengthy process, but is richly rewarded with an astonishing array of variants and beautiful plants.
Need I say how important this activity can be for an understanding of the plants and the way this is expressed in a classification? A good example is of the hybridization found between H. lockwoodii and H. arachnoidea ‘scabrispina’ at the Floriskraal Dam east of Laingsburg (see figs 1-3). The hybrid is very reminiscent of plants that I have seen from between there and Prince Albert; and also southeast of the Rooinek Pass through the Witteberg, and again northeast of Calitzdorp. Roy Mottram underscored the issue of hybridization in Haworthia classification. I have not ignored the issue, but rather viewed it as an imponderable in the continuum between populations that seems to have pattern.
Figures 4 and 5 are seedlings of a collection of mine from north east of Nieuwoudtville of H. nortieri. They could be related to ‘globosiflora’ and it will be interesting to see the flowers. However, it may be foolish to draw any conclusion from whatever flower shape emerges in the broader nature of the variation already observed although not adequately documented, both in H. nortieri and in other species complexes. Plants with these stubby deeply mottled leaves can be the consequence of growing conditions just as much as of genetic variation intrinsic to a ’species’.
Figs 6-12 are seedlings of another collection of mine (MBB7608) from Melkhoutrivier south of the lower Breede River and north of the Potberg Mountain. This collection is reported in Chapter 7 of Update 4, I think very aptly entitled “The brutality of reality in Haworthia”. It is one of 12-15 populations in the greater area that support my suggestion of H. mirabilis as including H. maraisii, H. magnifica and H. heidelbergensis. I suspect that H. mutica is also involved to some degree or other, while it is not difficult to extend comparisons to as far afield as Riversdale and further to H. emelyae in the Little Karoo.
Collectors will no doubt always have a problem with plants that do not match the perceptions they attach to names. Kobus Venter has always expressed anathema for anything but random selection when collecting specimen plants. This is in some degree correct because the specimens are supposed to represent a population. However, the opinion on a population is largely based on field observation. Subsequent cultivation and observation do not greatly influence that, while oddities and unusual forms may help mould opinion. While Nature Conservation may object to the introduction of permitted collections eventually into the collecting arena, I see no ethical grounds for this. I insist that it is an undeniable right that people have right of access to a natural resource with the proviso that there is no pressure on sustainability. ♦
When I was at the Karoo Garden I became a bit befuddled by the way botanists referred to the Cape Floral Kingdom. It seemed to me that they used the term for the vegetation that was on the Table Mountain sandstones and conveniently excluded that which was not. Thus the “Fynbos” vegetation, characterized by its Ericaceae, Proteaceae and Restionacea, was synonymous with this floral kingdom. An official document was published at the time which purported to classify the Southern African vegetation into biomes as major floral assemblages with very broad boundaries. It did not make sense to me because my observations were that the “fynbos”, however different in terms of historical origin, was essentially a flora of the sandstones, and that there was rather a winter rainfall biome which included karoid (Succulent Karoo mainly) flora. The role of geological substrate and skeletal soils seemed to me to be pivotal as there are places where one can virtually take a single step from one vegetation assemblage into another.
My impression is that it is the dramatic range of geological formations and highly eroded and eroding (in geological time) surfaces that is responsible for the extraordinary diversity and variation of the Cape Flora. My experience was also that taxonomists were not really aware that this might be significant in respect of plant classification. Botanising in the north and east of the country is not quite the same as botanising in the Cape and certainly it appears to me that European botanists familiar with the plants of a non-eroding land mass do not, or did not, give enough attention to the relation of substrate to plant variation. Unfortunately I am not a geologist, but I want nevertheless to illustrate a few plant populations which demonstrate the complexity of the problem where dramatic differences in soils over very small distances are reflected in variation in vegetation and in the plant species themselves.
In the Compton herbarium, there is a specimen of a Haworthia collected by Prof. Compton. It is labeled as from “Potberg sandy flats”. Now I digress because there is a problem of semantics. The word “retuse” is currently defined in botany and in the Collin’s Dictionary as “end rounded with a central depression”. In Smith’s “Dictionary of Plant Names”, the name “retusus” is said to mean “with a rounded slightly notched tip”. But this is not how it is understood nor used in Haworthia. Haworth, and other writers of the time, used the term(s) “retuso-deltoid” to describe leaves that are “bent back like the end of the thumb”. Thus “retuso-“ meaning “bent back” and “deltoid” referring to the triangular end-area. It definitely does not refer to the end notching of a flat leaf face. When we speak of the “Retuse” haworthias, we mean plants with bent back leaves and a flat triangular end-area. Prof. Compton’s specimen was of such plants where in the dried herbarium specimens the margins of the leaf ends seem to have been cartilaginous.
No other collections of Haworthia were known to G.G. Smith from the Potberg area other than perhaps H. minima. The Potberg area (see map) is also quite extensive as it is about 35 kilometers long and about 10km wide. My records at the time of writing Haworthia Revisited were virtually limited to reports of Luckhoff, Otzen and Herre of a small black species (maraisii!) and two collections of such a plant by both Frank Stayner and Peter Bruyns. I had twice been down to Infanta to be shown H. variegata by Chris Burgers on one occasion, and when I collected H. turgida south of Malgas on another. However, Chris Burgers had also reported two different plants, and one of these was from Buffelsfontein, south of the Potberg inside the De Hoop Nature Reserve. It was not possible to form any opinion about Burgers’ specimens without seeing live plants and I assumed that they were probably in the “mirabilis” complex. Adam Harrower recently came to light with a collection from Sandhoogte. This is also south of the Potberg, but about 15km east of Buffelsfontein. I was fortunate enough to see this population and was of the opinion that it was close to H. mirabilis. I was not convinced that this was what Prof Compton had recorded, but the phrase “sandy flats” also stuck in my craw. The Sandhoogte and Buffelsfontein populations (figs 1 to 5) are in very rocky streamside slopes and sandy flats are not the place to look for any Haworthia. The leaves in these plants are sub-erect and this seems to be related to habitat where slope and cliff dwelling plants differ from those on the flats. I regret that the pictures are not strictly comparable. We had spent the day trying to relate the De Hoop Oxalis records and scrappy specimens to correct names, and the final search for Burger’s Haworthia was more difficult than we had anticipated. By the time we found the plants, the light was too poor for photographing them. The photographs of the two cultivated plants obscure the close similarity of the plants in the field populations.
While partly motivated by trying to locate Trichodiadema and Acrodon species for Dr. Heidi Hartmann, my wife and I discovered extraordinarily pilose “retuse” plants along the lower Breede River (figs 6-9) This is about 4km over the Potberg from Sandhoogte and I surmised it to be the same, as both those and the Buffelsfontein plants evidence incipient spination. All these collections flower in late summer and I consider them to belong in the greater “mirabilis” group as it is becoming increasingly clear that fewer species is more probable than more.
On our most recent visit to the area, we discovered an almost smooth version of the plants being discussed, at a point about 4km over the mountain from Buffelsfontein (figs 10-12). The habitat is extremely rocky and gravelly, as at the lower Breede River, and I am sure that this is now related to Prof. Compton’s collection. The fact that the leaves are retuse in these latter two populations is related to the levelness of the area. I include all four populations in an informal category I refer to as H. mirabilis “pilosa”, despite the fact that in one population only, the plants are startlingly hairy.
Now I come back to my starting point. The geology of the area is most complex. The Potberg is mostly Table Mountain Sandstone except for the eastern most tip which is Cape Limestone. At the foot of the western end there is Bokkeveld shale which is home to a tiny remnant of renosterveld and all of this vegetation type that is included in the De Hoop Reserve. South of the Potberg it is mostly sandy and the vegetation is “fynbos”. North of the Potberg is a different story all together. While the sandstones meet Bokkeveld shale, the situation is confounded by the extensive and complex tertiary deposits characterized by ferricretes, silcretes, terrace and fluvial gravels and other. This is reflected in the vegetation of the area and close alongside Fynbos genera one can find dense patches of Succulent Karoo species such as Gibbaeum austricola and Acrodon. If one can step from one vegetation type into another like this, it is axiomatic that the complex exposure of this mother material is going to have a dramatic influence on the species that comprise the vegetation.
While I think that the collections I record here are related to what Prof. Compton collected, the habitats can definitely not be described as “sandy flats”. However, I can by no means be said to have explored the area thoroughly. The plants are very cryptic and can be confined to very small and localized habitats so they are very easy to miss. ♦
In an earlier article I described how Haworthia floribunda (at B on map) transmutes eastwards to H. variegata (at A on map) between the localities Klipfontein and Kleinberg, which are north of the Potberg Mountain. This is despite the fact that they both occur in close proximity at the northwestern end of the mountain. Difficulties now arise in the immediate vicinity to the west (at point W) and this extends northwestwards (to points C and D on the map). While we can confidently ascribe names to floribunda and variegata at those particular sites, the plants to the west and northwest are confounding. They fall into a no-man’s-land of these two species with H. mirabilis, H. maraisii, H. heidelbergensis and even H. mutica thrown in.
Localities and their names can also be confusing, so they are listed as follows:
A – the north and western point of the Potberg mountain on the De Hoop Nature Reserve and adjoining both Klipfontein and Juliesfontein farms (Juliesfontein farm has been renamed Poteberg Farm)
B – Byeneskop, a small hill on the western boundary of the farm Klipfontein
C – the farm Brakfontein
D – a hilltop north of Brakfontein named Witklip Kop
W – the two farms at Die Kop, Wydgelee, at the entrance to the De Hoop reserve
Figs 1 to 4 are from immediately west of A and show characteristics of floribunda in respect of the leaf tips. Figs 5 & 6 are of plants also from close by and the influence of H. maraisii is more evident. Generally the leaves are much shorter and stouter than one would expect in H. floribunda and the small size and numerous leaves suggest H. heidelbergensis. Both pointed leaves and leaves with the flattened and rounded tips may occur on the same plant (see figs 7 & 8).
Figs 9 to 13 are of two collections on the same ridge to the south of the Brakfontein homestead and are generally similar to the plants in figs. 1 to 8.
9. MBB6886 H. heidelbergensis – maraisii Brakfontein10. MBB6887 H. heidelbergensis – maraisii Brakfontein11. MBB6887 H. heidelbergensis – maraisii Brakfontein12. MBB6887 H. heidelbergensis – maraisii Brakfontein13. MBB6887 H. heidelbergensis – maraisii Brakfontein
Figs 14 to 22 are plants from a small hill to the north of Brakfontein. There were few plants there as the hill is severely grazed by sheep. There is quite a strong resemblance to H. mirabilis in that the leaves are firmly sub-erect and slightly more robust. The hill is very typical of the ferricrete/silcrete/gravel inselbergs which occur throughout the southern Cape. They are characterized as hard, conglomerated, solid rock that caps the hills, and these are underlain by Bokkeveld shale that has decomposed to kaolin. Exposed Bokkeveld shale is the dominant parent material for the soils of the area and these are arable, whereas the rocky inselbergs are not. The consequence is that habitats suitable for Haworthia are the relatively un-weathered skeletal soils which can only support sparse vegetation. Thus distribution of the plants is island-like and no doubt this plays a significant role in the way the plants relate to each other. ♦
14. MBB6890 H. heidelbergensis – maraisii N Brakfontein15. MBB6890 H. heidelbergensis – marasii N Brakfontein16. MBB6890 H. heidelbergensis – marasii N Brakfontein17. MBB6890 floribundoid leaf18. MBB6890 floribundoid leaf19. MBB6890 more pointed leaf20. MBB6890 more pointed leaf21. MBB6890 more pointed leaf22. MBB6890 more pointed leaf
Aloe haworthioides was described by Baker in 1887, moved to a unispecies genus Aloinella by Lemée in 1939 and then into Lemeea by Heath in 1993. This was wisely all undone by G. F Smith et al in 1995. This taxonomic dance is one of those events which probably bring taxonomy into disrepute in the minds of people otherwise respectful of scientific process.
A. haworthioides is rather symbolic of the deeper problem in the genus Aloe and related genera. This problem is that the genera are not firmly embedded in divine dispensation and there is much room for doubt about how they could be classified. Heath’s play with the generic status of the nine small Madagascan aloes rather demonstrates what can pass for well-reasoned, logical argumentation, just as surely as the same arguments were dismembered by Smith et al.
In pondering Haworthia, I have had to consider flower structure in relation to plants which are not Haworthia. So here I present a number of illustrations which show just how different A. haworthioides actually is. To answer “From what?” is a lot more difficult because of all the options available. A. haworthioides is a very small plant and seldom seems to reach a height of more than 80mm. It has many small slender leaves which are quite spinose to the point of “arachnoid’ hairiness and hence its visual similarity to Haworthia (fig.1 above).
Fig. 2Fig. 3Fig. 4Fig. 5Fig. 6
The flower is something else and I suppose it could be described as “aloid”. The inflorescence is sub-capitate to cylindric with the flowers congested at the end of the peduncle (fig. 2). The tube of the floret is quite short and the lobes are free to the base (fig. 3). To what extent fusion of florets in the genus is indicative of relationship is a question I wish I could answer. It is nearly as problematic as the question of zygomorphy and actinomorphy, which is exacerbated by the fact that observers will insist there is some sort of twilight zone where the exclusivity of these states dissipates. In bulbs, writers describe the petals as “united into a tube” rather than plump for a word like “fused”. The conditions exist where there is a true tube which has no visible differentiation into tepals, where the tepals are fully fused and yet visible, tightly adnate and almost inseparable, and fully free. All four conditions may occur in the same flower with increasing distance from the base along the “tube”. The use of the word tepal is also prone to some doubt because it is used when the inner and outer petals of a flower are “similar” and it is not certain what constitutes “dissimilar”.
It is instructive to consider that Reynolds abandoned any system of formal sections for his second volume of Aloes of tropical Africa. Where in his first volume, he went to great pains to formally maintain hierarchy. In the second volume, he grouped 13 very small Madagascan species together and wrote “They do not comprise a homogenous group. Species with widely divergent characters are included only on account of their small size to facilitate identification”.
What is quite dramatic about A. haworthioides is the nature of the stamens. The filaments are exserted, but they are very broad so that they form an apparent extension of the floral tube. It is again interesting to turn to Reynolds. He writes “Perianth white to pale pink, slightly campanulate…Filaments dilated, remarkably thick and fleshy, the 3-inner narrower and lengthening before the three outer, with their anthers in turn exserted.” I am inclined to grasp these words as justification for the poetic license or plain hermetics of my own writing. In the plant I have, the flower is hardly describable as campanulate (bell-shaped), the filaments are certainly subequal in length but very similar in width, while anthers are barely exserted beyond the length of the filaments. Reynolds poetic license seems to extend to the variety aurantiaca where “the whole spike, including the axis, bracts and perianths of a beautiful bright orange-red, with the stamens yellow”. Again in the plant at my disposal it seems as though it is only the filaments which are orange-red and the anthers only are yellow. The style is relatively short and is not exserted like the filaments.
I am sure that many other Aloe species could be analysed in critical fashion and probably more effectively than this, to show that the genus is possibly a very artificial construct. I say this because observers have tended in the past to take the same broad uncritical view of Haworthia, as Dr. L. A. Codd once did, when he wrote to me to say that “we are ignoring the small differences”. This was in response to my complaint that ALL Haworthia flowers were said to be small and the same when his staff lumped Chortolirion into Haworthia.
It seems to me that we are facing an onslaught on classification driven by the novelty and promise of molecular studies where DNA is seen as the ultimate solution to problems of classification. I have my very grave doubts. Aloe haworthioides is an oddity and while DNA studies will surely support this and enable better explanation of its relationships, identification and recognition by name may be better left as it is. Molecular biologists will lead us the way of Reynolds, which is the appreciation of the fact that hierarchical structure in classification is often misconstruction. We also do not all perceive names and their meaning in the same way. If we simply altered our way of thinking by an educational process perhaps we would be better off. The binomial system has worked remarkably well to date, and can continue to do so if we meddle with it with more circumspection. ♦