68. 2019.7.28 – Coming back to some sort of sensibility after 3 days in the field. Rogan Roth sent me these two pictures of tubeburcled mirabilis from the Swellendam area.
An extraordinary phenomenon. Ask me about it. We can identify 5-6 generations of Aloe ferox. The oldest single mother at a conservative 70 years of age based on the age of plants established at Karoo garden in 1946. The youngest seedlings are current year? I have seen a similar sequence from beyond Uniondale to west of Oudtshoorn.
69. 2019.7.30 – Rogan Roth kindly showed me this population east of Swellendam. It did not strike me as being the same as I had seen in that general area before and I also thought some of the plants could easily be slipped into the Rotterdam “groenewaldii” populations. With less rounding of the lead tips of course. Then it hit me that it appears I have been arguing “groenewaldii” in the context of a supposed discrete “mutica”. This sort of reasoning does not work in Haworthia. “Groenewaldii” is neither mutica, nor retusa, nor mirabilis and neither is it a species, subspecies or even variety of any of them. It is a discrete entity in a vast complex of Haworthias that could all be classed as H. retusa where each population is a discrete and significant part of that true species system. An added problem is that it is not clear where retusa stops. So the Linnaean system based on Darwinian evolution may be quite wrong. Genera and species are just human constructs – they aren’t real things. DNA sequencing may be telling us fairly accurately what the states are, and we are just not presenting or interpreting the product correctly? ♦
70. 2019.8.2, MBB7500 (Die Kop) – This is a really interesting population. Mid-south of area 1 of my map and east of mutica MBB7941 by a few kilometers. It is a “hybrid” population mirabiisXmutica flowering March. It has the name H. hammeri and this itself raises a very interesting phenomenon. Who “discovered” it, who actually collected it and how did material get to a foreign national to describe as a new species? Why is of course something else. Nature Conservation permit regulations are painfully restrictive and heading to where it may become illegal to be in possession of a “species” like this limited to this one locality. Is this another case of the binomial system prostituted for collector interest? A question for the collector (and probably the author of the name too) … “Just what is H. hammeri?”
Some of these look to take on a floribunda like twist and slightly elongated leaf … It seems to be very probable that floribunda is completely integrated into the greater retusa group (species) and this is what I have pointed out in many posts. Floribunda appears in real form a short way east of this – again in a retusoid mirabilid habitat and alone.
These are more pictures from Die Kop. H. ha..ha..mmeri. Flowering time might be a mirabiloid dominant.
The inevitable that I was trying to avoid looms. Stephen Gould wrote his book “Rocks of Ages” providing his view of “non-overlapping magisteria” for science and religion. A very warped one as I wrote in Updates. Science claims dominion over knowledge and dismisses metaphysics as lunacy. Religions claim dominion over spirituality and condemns mysticism as satanic. Both views are plainly nonsensical. Some while ago I posted on metaphysics and indirectly on how the outer world is an illusion of space and time. Reduction and particularization of science leads us out and away from truth. The fact is it is a conscious creation. Knowledge and truth is the same thing and must exist everywhere at once? Species on the physical plane have no reality because all matter is a projection and an illusion. There is humongous literature on all this. I am not telling you this as something new and my discovery. Science, having captured the designation for a process of acquiring knowledge, is suppressive and deliberately dumbs us down to the unreality of physical creation and the truths about it. Matter is created from 5 subtle elements described by countless mystics as earth, water, fire, air and ether. They say, that associated with these are even more subtle elements of sound, taste, touch, and smell. These are energy states (string theory?) and they are represented diagrammatically in the physical form by the 5 platonic solids (they are not solids – they depict energy fields with differing vibratory states). This earth = cube, water = icosahedron, fire = tetrahedron, air = hexagon and ether = dodecahedron. Chaurasi is described as the wheel of 84. 8,400 000 life forms and each life form is a particular combination of the five elements. Plants have only the element of water, invertebrates have fire/air, fire/water, fire/earth, fish are fire/water, reptiles are fire and earth, birds are fire, air and water, mammals and marsupials are fire, air, earth and water. Man alone has all five. All this used to be common knowledge and in Plato’s time, knowledge of the dodecahedron was made secret and divulged to people on the penalty of death! I am no weirdo – I am just telling you how it is.
71. 2019.8.4, MBB7937 – This is a really wonderful locality for mutica that Jakub Jilemicky pointed me to. Some distance E Die Kop and could be the SE limit before mutica is subsumed again in mirabiloids. The quartz is very ferruginous?
That Jakub discovered it is simply indicative of the nature of exploration and “discovery” when there is so much ground to be covered. I will touch on this again.
The group picture of Kobus Venter with Daphne and I was taken here.
72. 2019.8.5, MBB7535 – A little north of the last and on a sparsely quartzy shale substrate. The red leaf colors are probably due to soil factors unknown and I have never grown many muticas. It was a species fairly unknown to me even if one of the first I saw when I was at Karoo Garden. In cultivation I very much doubt you could get such rich color – reds especially seem to be reaction to direct sun in all the retusoids especially. I posted that toothy one before and in the “groenewaldii” series a green many leaved plant similarly toothed. Just another issue of what species descriptions where like with G.G. Smith when single plants were used.
That DK graph makes a point but is also very weak. What we actually have is a two bar graph of confidence and knowledge against time. It is with such a graph that we make assumptions about each others confidence (arrogance) and ignorance (knowledge). The Dunning-Kruger Effect just suggests that there is a tendency in all of us to be more confident in relation to our actual knowledge than we should be. Do we see ourselves as either unassuming and knowledgeable and others as arrogant and ignorant? Is this while we struggle to balance all that against a guess at what it means to be a nice person? I would have to apologize for how I am seen, for the assumptions made as to how I see others, while perhaps others may need to apologize for how they see me and so on and so forth. All this futile wasted effort when we should be objectively trying to seek the truth. Something I do not know is just how all these mutica variants behave in cultivation. I do know I had a mutica and a pygmaea in cultivation that looked identical except for bract colour. ♦
73. 2019.8.6, MBB7944 – It would be truly interesting and instructive to now go southwards and see what happens with variegata, floribunda, and mirabilis, but there are other very significant twists. So lets go to the western boundary of the mutica distributions. A little west and north from Bredasdorp are three populations of H. rossouwii where one would expect to find either mirabilis or mutica. Weird, and it does get weirder still. I wish my staunch opposition :) will consider exploring the significance of the similarity of rossouwii to emelyae multifolia or to some of the mirabiloids. Then a bit further north still are six populations (known to me) of H. mutica and two of mirabilis. But there are many off-road isolated small patches of uncultivated land that will house Haworthia. Just as a matter of vital interest are the two mirabiloids. They are about 200m (MBB7944a) apart but one (MBB7944b) is contiguous with mutica MBB7943). While the mirabiloids are best treated separately, this exercise is about the affinities with the retusoids and muticoids. So let’s just look at 7944a without getting further misled by names such as maraisii, magnifica and a dozen or more others that are the product of intellectual obduracy or laziness, or need for something simpler than a purely botanical solution.
With gentle humour. We can now carry on collecting bits of information like a baboon raiding a mielie (corn) field dropping cobs as fast as they are picked, or try to hold it all in memory and form a greater picture. Please don’t think I am demeaning anyone or animal by the baboon metaphor – I am just expressing how difficult I find it to remember and relate all the populations and their variants to each other while trying to form a reasonably complete picture.
I numbered 7944 as a and b because they are only about 200m apart. 7944b H. mirabilis is contiguous with 7943 H. mutica (to follow), separated in an identical habitat by a shallow eroded depression about 5m wide. What is significant? Do those plants show “hybridism” and floribunda characteristics? Additionally how do you deal with this in the mainstream science arena? Geology would be easy as it is just a straightforward identical Bokkeveld shale series. Vegetation? A detailed Braun Blanquet survey would no doubt reveal difference except that at a Council of Scientific & Industrial Research (CSIR) Congress on Karoo habitats in ca 1984 (Prince Albert) it was agreed that BB was inadequate in arid sparsely vegetated systems on a variable skeletal substrate. If BB was adequate, just think of the time needed to describe and record enough Haworthia habitats to generate a result? By the time the 10th habitat was annotated, the first would be different. Where would you find replication anyway? I am sure you could describe leaf anatomy, shape, form and texture for x number of plants in mind-dazzling language. Would that be better than these pictures? Incidentally, this kind of contiguous occurrence is also evident at two localities east of Riversdale where there is “hybridism” of retusa and floribunda, and again of retusa and mirabilis. Totally different substrates that might be very difficult to identify at a map scale of 1:50 000.
74. 2019.8.7, MBB7943 – All that DK stuff is just a distraction from the actual issue that is of just exploring a classification hypothesis. It is really only my opinion based on my experience and my DK profile. There are some serious issues that I cannot resolve and my conviction is that they are not solvable in the climate of materialism. While there seem to be gross continuities like I have been harping on as well as that of the cooperoids, cymbiformoids and more, there are single population oddities that I cannot fit e.g. elizeae, calcarea, truteri, marxii and also more. There will be another revision sometime in the future by someone else that will bring nomenclature changes based on real problems of typification and ICBN deviations. These will be implemented with a different opinion of the species hopefully based on better sequencing and its interpretation. Wildly hopeful is that it will somehow accord with my definition of “species”.
Note the two plants with flower spikes. It was March and mutica is a spring flowerer – mirabilis late summer. One plant has a healthy peduncle. I have discussed this before and explained that despite this 4-8 month time gap cross-breeding does occur. No one took up the point re aloes and the question of germination and successful reseeding as rare events.
Jakub has done some great exploration and this is an area too that he pointed me to, mid-Nov, same general area. One plant was a monster at 16cm diameter. ♦
75. 2019.8.8, MBB7954 – Skipping 9 populations to get south of Stormsvlei and to the farm Volmoed. It is November. I’ve tried to limit the number of pictures; but there are so many interesting variants. I wish I could show them one by one. They need to be looked at one by one.
Jakub Jilemicky – I remember seeing here a few scabrid clones.
Jakub has made such a valuable contribution. His localities are not the same as mine. He exposes the weakness in all our efforts. Lack of knowledge. Drawing conclusions from inadequate data. Not thinking and not asking questions. If we all honestly decided to sincerely think through this “species” problem and strive to arrive at a common agreed objective, we might be able to reach a solution. We will not do this while classification and revisions are presented and owned by individuals in a climate that does not value truth. Nor while fundamental principles of science i.e. “universality”, “communality”, “no personal gain” and “organised scepticism” are ignored.
There are absolutely mirabiloid genes (if you can call them that) in this population. Just out of sight in one of those little depressions is a mirabilis population that links the maraisoid mirabilis to the badiod and possibly with the muticoid. All this is supporting the hypothesis that there is a single gene pool and one species only? Mirabilis is in this area with the 9 mutica populations I mention – in not a single case do they occur together???? The reason seems to me very “simple”. The difference genetically is so small that there is a subtle element that determines the domination of one over the other in any particular habitat niche?
It is also just a single plant pictured, and as with mutica and retusa, one has to carry images of many plants from many populations to arrive at a valid opinion. I said badioid when I really mean all the mirabiloids in that Napier area. We are led to think in a reductionist manner and this is what enlightened thinkers try to explain to us when they use the word “holistically”. I crudely use the metaphor of the baboon picking cobs because I know what it means to go into the field and see a population of variants and say to what it relates to. Readers tend to forget what has gone before. What is the hypothesis we are looking at? A single gene pool with each population and plant significantly a part of a whole. A principle of classification is that it covers all the parts and not just bits of it. Now we go exploring and anything different is seen as new, instead of asking the question of how does it fit with or help explain what we already know.
A recent treatment of Haworthiopsis is rather depressing as it follows the tired concept of a flat evolutionary tree growing out of a two-dimensional bed of primordial mud. It is only the late Koos Roux as the botanist he became from a history as a horticulturalist and naturalist, who has ever expressed to me the same view I have, i.e. the phylogram that emerges from DNA analysis by Bayesian statistics and its interpretation, is a two dimensional one and cannot express the true reticulate nature of change (is evolution the same thing?) and relationships. Whether the sequencing is past technology or new generation sequencing, this issue remains the same. If Haworthia is to be treated in the same way as Haworthiopsis the result will be comparable to the point at which Smith retired, confounded by the new facts of nomenclatural obligations both real and perceived. A dart board will be used to identify varieties in the continued absence of a species definition that is true. The sad fact is that plant taxonomy is very largely a subjective process. In the Haworthia groups this works for Tulista and for Haworthiopsis (unless one starts imagining subgroups and what purpose do those actually serve?). In Haworthia stricto there is just going to be the usual interpersonal put-down and strife.
76. 2019.8.10, MBB7075 – NE Volmoed between Riviersonderend and Stormsvlei. H. otzenii was described from near here, from a population no longer in existence.
Perhaps one can ask if anything has been proved by these posts to date and since about May? The argument has not just been about Haworthia retusa and its allies, but about classification and science generally. Is it my function to resolve these problems or just to suffer under them?
We have looked at (a guess) about 80 populations. There are possibly 400 more to consider. So we have seen about a fifth or much less of what there is, and I am not counting pictures I do not have from pre-digital years. Is all this seen so far in memory?
Tinus Potgieter asks “If we all honestly decided to sincerely think through this “species” problem and strive to arrive at a common agreed objective, we might be able to reach a solution.” “How to get to this point?” Perhaps by recognition of the problem? When the idea of the national species list first arose, I was asked by PRE “Whose revision should we use, yours or that of Col. Scott?” In present time all hope is attached to “next generation sequencing” while faith lies in existing sequencing results. I have suggested that sequencing is flawed because it ignores the space/time nature of species, and that a definition can and should be derived outside of belief in an apparent objective way of determining them. While collectors and the societies that represent their interests blindly put their faith in science, it is unlikely to happen. As in Haworthiopsis, some half-baked solution will be dressed up in sophisticated nomenclature. The language of DNA sequencing, Bayesian statistics and phylograms will be added to trump and ridicule practical field observation and common sense. True or not will not be a serious consideration. ♦
77. 2019.8.12, MBB7776 west of Riversdale, between Riversdale and Heidelberg – Coming to the end of the first chapter of my story. Lets now try something while we contemplate the reality of the difference between mutica and retusa. What are these next 8 plants? I suggest that they are H. mutica – but it is your opinion that matters now. What do you think?
Let me just add something about this DNA sequencing stuff. The technocrats assume that if they have a sample labelled H. retusa, it comes from an entity that truly exists and can be labelled so conveniently. In the one exercise I was involved in I attempted to get the dudes to replicate. How far it got I do not know beyond the first sequencing run presenting some very uncomfortable results. My comments and questions got me thrown out of the project.
Lawrence Loucka: ‘Replicate’ is a statistics term that means to use multiple samples, make multiple readings, to determine variation within (not between) populations. Because of time and expense botanical DNA studies have generally had insufficient sample sizes to be statistically significant. But costs are coming down and quality is improving.
Stephen Boisvert: Replication just means to repeat the experiment or measurement and get the same result. It can be completely meaningless if you don’t satisfy the requirements for philosophical validity – face validity – construct validity or logical validity – predictive, concurrent, discriminant and convergent. Looks like Bruce’s complaint here is with construct validity, concurrent and discriminant/convergent.
The first issue is a bit more complicated. It’s conceptual at the level of what you think a “species” is. Botany (and zoology) has a strange history on this with one with their notion of Type plants (and animals) where a single instance is held as the defacto species standard and entails a whole host of problematic philosophical assumptions (like a platonic ideal form or aristotelian essentialism) which never quite fit the real physical world or the use of concepts in language and has huge sampling problems (one individual chosen to represent a purported population is extremely problematic to say the least). Still I believe if you clearly articulate your assumptions regardless of your philosophical positions on concepts (platonic idealism, aristotelian essentialism, Wittgensteinian familial resemblance and so on) you should be able to produce useful and communicable knowledge. The big problem is being clear enough both in your own head and in your writing about your assumptions which is difficult because of lot of this is stuff we learn and know at an implicit rather than explicit level. You see this when people can identify and distinguish plants but if you ask them how they differentiate they sometimes can’t tell you. They just know implicitly without explicit rules.
Bruce Bayer: Stephen – I truly appreciate your comments. I have been criticized because I many times cannot identify a Haworthia unless I know where it comes from. Written descriptions are fairly useless with many examples to demonstrate the fact. How do you describe the variation in a population when it is difficult to describe a single plant? No matter how unclear the head, the fact is as you see in these last pictures posted, plants from quite different populations (too often identified as different species) can be identical. What really bothers me is that other peoples heads are not clear enough to grasp this statement. I have been pondering explaining my expressed discomfort with the Haworthiopsis paper in Phytotaxa where my discomfort is seen as a personal attack on the authors.
Ronel Kloppers and Sean Gildenhuys have written an outstanding paper with respect to nomenclature and technicality of taxonomy, but which I regard as weak in both scientific method and “philosophy of concepts”. I am hesitant to deal with the issue because I am not in their intellectual league and not in the least sure where my convictions stem from! I am however extremely concerned that the methodology successful perhaps in Haworthiopsis will be a total disaster if applied to Haworthia stricto sensu.
———-
This next set is just another lot from the same place west of Riversdale. Silly me. I think the first would fit comfortably with H. mutica and these now with H. retusa.
Early on in this series I started by going north and west from the Duiwenhoks River south of Heidelberg, and moved west to H. mutica. Now I skip four very relevant populations and travel eastwards. What is happening is we are going to H. retusa. It will take us across area already covered in detail in the Updates under the cover of mutica var. nigra.1 After that north-east of Riversdale to Kruisriver, further east to the Platkop area, to east Albertinia, to the Gouritz and H. pygmaea. There are three directions northwards to H. emelyae untouched and two southwards that link mirabilis and retusa.
78. 2019.8.14, MBB7794 – SE Heidelberg. Unfortunately I do not have pictures of one population that used to be closer to Heidelberg that would also have been instructive. 7794 is one of 3 populations, one of which is commonly known. I would refer to these as mirabiloid retusa. ♦
79. 2019.8.14 – I think I am going to abandon the topic of “species”. This is H. rossouwii from a place new to me southwest of Heidelberg. Funny that one plant that seemed to me to be a significant variant, was also the target for a browser.
Steven Molteno: Interesting that the one “marginata-form” plant was grazed. Bristles, like tubercles, could be evolved for defensive camouflage reasons, in the dappled light of a nurse bush. Breaking up the appearance, like the stripes on a zebra?
80. 2019.8.15, MBB7732 – From south of the locality for the original and erroneous H. serrata. It does occur further north near the N2 and might still be found at Koppies. Here it is among dense grass and so very vulnerable to fire as what decimated the Koppies population.
81. 2019.8.16, MBB7803 S. Heidelberg – H. rossouwii is a strange species because it has a few populations near Bredsadorp and a few SW Heidleberg. The plants very similar. But there are oddities on the outskirts of the distribution. This is a small form south of Heidelberg in three population, two of which are probably close enough to be considered as one.
Some more MBB7803 from S Heidelberg. There are no doubt two ways of approaching this. Simply giving them a new name or trying to fit them to existing. The latter is what I try to do – so just follow this little story and see what follows. Even if we go to the large hadron collider we still may not find an answer.
82. 2019.8.18 – Unfortunately I do not have pictures for what I eventually called H. rossouwii var. calcarea – an oddity from the De Hoop Reserve, but this is another oddity from the limestones just east of Bredasdorp. I ended up with this as a rossouwii variant as well for lack of any better idea. The east end of the limestones towards Potberg provides its own complications that better involve H. variegata. ♦
83. 2019.8.19 – Another oddity is this elizeae up on the Bromberg near Stormsvlei. A little patch on a lonely sandstone shelf quite unlike H. mirabilis in the area with two plants shown here. The flowering time is November, and for lack of a better fit I think it may belong with H. rossouwii too. Note buds on the accompanying mirabilis plants in September.
84. 2019.8.21 – When I wrote Haworthia Revisited in 1996 H. heidelbergensis was barely known. Since then there have been so many new findings and observations it is almost impossible to detail all the complexities and subtleties associated with it in its later role as an H. mirabilis variant and a newer role as a H. rossouwii one. Or is it really an H. retusa variant? How does one do justice to these things when it is well-nigh impossible to quantify and describe such endless variation. But I was posting about H. rossouwii and I want to take just a few populations to illustrate how tough it gets. Here are three pictures of what was H. heidelbergensis var. minor from JDV85/5.
I add a map and will go through 12 populations to try and show why I suspect that H. rossouwii is involved. This map position has H. rossouwii a few kilometers to the west, and about 35km to the east. Petrophila and calcarea are to the SE and elizeae well away to the NW. There are some other considerations too.
The point of this map is that exploration comes down to significant distance as short as 10 meters. When I started I was more than comfortable with sampling distances a lot greater than 10km.
85. 2019.8.21, MBB8044 – In close vicinity. Pictures taken in February when all these flower and just remember that I suppose February flowering to be a mirabiloid dominant.
86. 2019.8.22, MBB6638 – Several 100m south and is “clearly” mirabiloid. These forms also occur several 100m north of what I referred to originally as H. heidelbergensis var. minor (and lately as H. rossouwii var. minor). They are also present a similar distance to the east. In the next post I will start about 3km west and show populations homing in on this “minor” thing.
By a curious coincidence I saw a ‘banned’ TED talk by Sheldrake. Link His concept of “morphic resonance” seems to offend a large element of the scientific community. But I think he is right and that here is a lot more to the question of energy fields than we might like to think. ♦
87. 2019.8.22, MBB7285 – Starting from the west now is H. mirabilis and again with no two plants quite the same, i.e. Mirabiloids.
88. 2019.8.23, MBB8048 – Skipping out, in a change of mind, 8947, 8052 and 8053, here we are about 800m W of “minor”. It was late February – the mirabiloids (as a general rule) do not like direct sun and are often well hidden and protected in shale cracks and crevices and under sparse or even dense but short plant cover.
Why the impatience? I had some recent interesting correspondence where my correspondent referred to various writers and their respective opinions and followings. Well this is where the problem lies. We ask “who is right?”, instead of “what is true?”. Instead of thinking for ourselves we rely on others to do it for us.
Lawrence Loucka: Have your correspondent read all of Haworthia Updates, then have the conversation again. If your correspondent isn’t willing to make the intellectual investment, then any further dialog would be a waste of time for both of you.
Bruce Bayer: It does not appear to me that any writer has taken my introductions seriously and there does not appear to be anyone who has proposed an alternate species definition either. As Dr Manning wrote, there is just a blind acceptance of a zoological species definition based on inter-species sterility without any consideration of that issue. Usually the thoughts about the constitution for species is dismissed as personal vagary on the part of past writers and excluding the present author. 😄
89. 2019.8.23, MBB8050 – About 400m west of “minor”, skipping 8049 is 8050. Rather surprising to see how much resemblance there is to H. floribunda? But I was reflecting a bit on history and the first journeys into the interior ca 1652 plus. Have we really learned anything? Lawrence sagely posted the introduction to Revisited written in 1966. Even with all the DNA sequencing, has anything since been said that adds anything to our knowledge of Haworthia? I do not think so. The probability that all these things I suggest are a species “H. retusa“, may indeed be a step closer to reality without actually getting there.
90. 2019.8.25, MBB7821 – Now about 150m west, and the transition to greener, more, and more erect leaves is stronger. These are all just subtle effects on a small scale that occur among all these populations that I suggest are probably the same species. No amount of sequencing, quantification, acrobatics and verbal gymnastics to please collectors and irritate alternate experts, is going to remove the element of doubt and consequent confusion.
91. 2019.8.26, MBB8045 – About 15m away from the original mini-habitat of minor. This multi-leaved habit is not characteristic of the mirabiloids but then just what is?
92. 2019.8.27, KG36/70 – Finally back to the original locality for “minor” using the original collecting number more than 30 years later. What does one make of this? I ended up placing it, rather desperately as a variant of H. rossouwii. There is no better reason than the curious resemblances one finds in various mirabiloid populations to that species. I know H. rossouwii from 9 populations and nowhere is it in the company of any other species. What I have shown is how intimately involved H. floribunda is with this greater retusoid raft of populations. H. rossouwii much less so, but one cannot doubt that there are cross connections. Does it get subsumed into H. mirabilis also despite different flowering time? Depending on DNA sequencing to provide answers when the sampling, analysis and interpretation is done without good insight into the nature of the problem, is not going to provide a satisfactory answer. Especially so when the DNA results so far available suggest so little difference between species.
Jakub Jilemicky: I have once seen rossouwii and mirabilis together – on Oudekraalkop farm.
Bruce Bayer: Thank you Jakub – that is really interesting. ♦
Conclusion: There is a serious problem of personal perception. I am now 84yrs old with all the accompanying ailments of the inevitability of decay. I have had a long break from Haworthia and have a wish to see, from my perspective, those things resolved that are not. The very nature of the journey and the experience, has involved conflict and personal differences. I can’t apologise because how can I be sure that I am either right or wrong? My motive has been, as far as is possible, purely to grasp the truth and writing about the issue has been central to the process. It all involves a very serious journey into philosophy and science, and even into the meaning of those two words viz. love of knowledge and “to know”. Do you know anything if what you know is not true?”
These posts are not intended to follow what has gone before. They are just a platform for me to reach out and meet people that may be asking the same kind of questions I have asked and who just might want to see things through another lens. Polish my own perceptions and fix distortions. In my journey with Haworthia I have met countless people from both ends of the intellectual and academic spectrum and do not want to pretend about where I might fit as it could easily be what is perceived as the wrong end and justifiably so. There is no wrong end for anybody else. It is about motive and those with the right motive hopefully will gravitate here of their own volition.
Haworthia are not poisonous plants and have given me a vast amount of pleasure. I treasure all the people encountered in my personal journey. In these posts too.
This has been a small excursion into the amazing world of variation and exploring the problem of names to fit. It does seem to me that science offers more promises than solutions. Personally I feel that science has disappointed and tricked me into trying to find answers where it has hidden the truth. Darwinian selection and survival seems to be a dead horse. A species model just does not fit no matter how one tries to define the word species. Botany with its taxonomy and nomenclature are a breeding ground for argument and disagreement. A collective approach was attempted in the genus Aloe, but in my opinion that was a disaster as it never addressed the core problem. My suggestion is that this should be tried again but recognizing that the myth and reality of species needs to be admitted and seen for what it is. This will have to be a process and it should start with next generation sequencing? No – it needs to start with us.
From the practical point of view, and to give credit where credit is due, I think an account should be given of the eastern flank of the greater retusoid puzzle. The person who holds the key, the understanding, insight and the pictures for this is Kobus Venter for whom far more credit should have been given for the publication of Haworthia Revisited. Much the same could have been said for Steven Hammer. When I undertook to write revisited, I said I could not do so without the help of these two stalwarts. That I failed to acknowledge them adequately, is because I did not think the book was complete enough to be of such credit to anyone. But I have also come a very long way since that book with the publication of the Updates (print and digital) and the involvement of Harry Mays and Lawrence Loucka. A final account should include the case of Brandrivier H. emelyae var. multifolia to indicate or prove, that classification is just a moment in time and we may never arrive at an answer that is true.
93. 2019.9.1, MBB7719 – East of Stormsvlei – when you doubt if mirabilis can still present itself as any different, it manages to do so. No retusoids in the near vicinity either.
94. 2019.9.2, MBB7513 – Yes this is also H. mirabilis, and it is not surprising that Haworthia classification is so controversial. Variation within and between populations is just incredible. These particular plants were on very large lumps of manganese concreted rock. ♦
95. 2019.9.19 – During the course of these posts about the retusoid complex, I received a comment about Haworthiopsis that has relevance. This was that H. tessellatata, H. granulata and H. venosa (group A) presented the same problem as the relationship of H. coarctata, H. reinwardtii and H. fasciata (group B). It simply is not true and just illustrates the misconceptions that befuddle communication and decision making. In group A, I know of no evidence whatsoever of any interaction between the three “species”. Not that this diminishes the problem. They are geographically discrete and only between H. granulata and H. tessellata is there some doubt about continuity along the border of the Roggeveld Plateaux near Middelpos. In the case of group B, it is quite different. The evidence is that H. coarctata and H. reinwardtii are confounded in the coastal area between East London and Alexandria, and that they are essentially different on the basis of polyploidy as tetraploids vs sterile triploids. This has not been clarified. Also the populations often appear to be vegetatively propagated single clones reflected in the many G.G. Smith names. Evidence was posted in this group to remind and suggest that H. coarctata is confounded with H. fasciata in the Port Elizabeth area. This is also reflected in the von Poellnitz name H. fasciata forma variabilis. It is also evident that populations in the Lower Gamtoos (see pictures) are difficult to distinguish as either H. fasciata and H. reinwardtii (a situation touched by coastal inundation in the distant past). Furthermore there is the interaction of H. fasciata with H. glauca north east of Uuitenhage (“H. armstrongii” and “H. browniana”). Most difficult is the problem on the Zuurberg north of Kirkwood where H. glauca and H. coarctata become indistinguishable. While Haworthiopsis being such a relatively “easy” taxonomic group, there are still serious problems that explode when it comes to recognizing internal infraspecific groups such as subspecies, varieties, and forms. All this is secondary to the vexing problem of species definition. The genus Haworthia is far more complex and bewildering. It is frustrating to finally recognize that while South African Botany accepts the new generic dispensation for the Asphodelaceae, the International Organisation for Succulent (IOS) plants does not (yet). ♦
I struggle to really get my mind around principles and practices that get so shrouded in a language of their own. This cognitive dissonance business is a case in point. A person adjusts their belief system according to what they are psychologically more comfortable with and can easily depart from the truth in the process? I cannot believe that Haworthia is unique in the problems we experience in trying to arrive at answers. The only difference in the extent or degree in which we are able to collect together and directly experience for ourselves the vast diversity they present. By working in a botanic garden I was just unfortunate enough to have had this experience with many different families and genera. It is not a unique experience and others have had the same opportunities. Why has not each of us who have had this kind of experience come to an agreement about the difficulties of classification? Cognitive dissonance? Perhaps it does not suit the needs and realities of our daily lives and how we relate to others who without that experience, but with greater intellect and authority, know considerable more and tell a different story. This happens against a vast background of history. Why I say this is because I am presently trying to find names for a very rich local flora. So Watsonia is a case in point. There is a revision by a very distinguished botanist but it is only with cognitive dissonance that I can believe this revision will give me the tools to correctly identify the 4-6 species that occur here. But it is not only Watsonia that I struggle with. I can probably very conservatively list about 50 or more others as examples that mirror the same issues that the H. retusa affiliates do. But it is almost impossible to even mention these difficulties with the myriad of amateur enthusiasts and even botanists who engage at all with plant identification and are convinced the authority exists in the books and people they consult with.
Us so-called Haworthia experts need to have a good look at the Watsonia monograph for purposes of a reality check. I have every admiration and respect for a remarkably able, competent and leading taxonomist. But if the argument and data put forward for the support of many of his taxonomic decisions are anything to go by, the classification of Watsonia is a disaster area (at least for the winter rainfall area) that no amateur (me included) dares venture into. It is totally dependent on a cognitive dissonance that the author must know what he did because no one else could, without a very comprehensive study of all the herbarium material allied with several (multiple) seasons spent in the field making own observations.
Dr Peter Goldblatt, The genus Watsonia. Annals of the Kirstenbosch Botanic Gardens, Vol 19, 1989 ISBN 0 620 12517 9
Read critically W. borbonica and W. rodgersii. See dubia and strictiflora. For the SW Cape species it is just an endless list of problems riddled with subjective statement like “clearly” and “I prefer”. There is also the reference to “hybrid swarms”. Just as in Haworthia, the term seems to have been used to obscure what the variation obscures. Simply put, hybridization flaunts the reliance on a zoological species definition. The existence of hybrids is blamed for the inability to provide a “useful” key! Look at dubia and strictifolia, and then amabilis. Humilis and laccata? Then try to sort out meriana and all its possible affiliations and in turn the affiliations of its affiliates. Look at the distributions and correspondences. In effect the author says “Trust me” confident that no one will ever be able to track through all those specimens and all that field work again. When time and change will also have decimated more of the evidence. At the same time I doubt if anyone except another polymath, will ever be able to track the types, their names, and the nomenclature as competently or in the same way. It is a tribute to Peter Goldblatt that this makes for such an interesting adventure. You could derive a lot of pleasure and genuine knowledge in the process.
Why Watsonia? Well – why not Oxalis, or Asparagus, or Conophytum, or Ceropegia, or Drosanthemum, or Crassula, or so many other genera I have been confronted with…
I find it rather difficult to deal with some of the complaints I have heard about my preoccupation with “species”, but there is truly a problem that needs to be addressed. Science seems to have been reduced to a social construct that all knowledge is confined to the material creation. Within this there is a species concept that is also just a construct. This is that living organisms can be sorted into discrete gene pools called species, and currently that these are identifiable by DNA sequencing. Also concurrent is that it does not work adequately and that a new next generation sequencing will iron out the problems. Funny all this, because evolutionary theory is that this has all come about by random mutation and selection. There is some doubt about this and the concept (upon concept) of “intelligent design” suggests that if this was true, the vast majority of organisms would either be trying to get into such groups or trying to get out. So there is also a concept that there are no such things as species? How does poor average citizen deal with this? By referring to Sheldrake’s “morphic resonance” of course? Because we have heard of names like Adam, Moses, Noah, Aristotle, Plato, Hippocrates, Cleopatra, Napoleon, Genghis Khan, Freud, Kinsey, Harvey Weinstein, Piglet, Pluto, Bacon, Newton, Einstein, Marilyn Munroe, Watson, Crick, James Thurber, Walter Mitty, Bob Hope, Charlie Chaplin, Aldous Huxley, Noel Coward, Mickey Mouse, Donald Duck and more; we believe all the wisdom enshrined in these multitudinous characters is embedded in our own individual consciousness. So we entertain ourselves by arguing amongst us about everything and anything on the basis of a great common wisdom or that of entirely our own, which in reality is grossly deficient. By limiting our science to physical creation and this misguided social construct, we miss the reality that it also tells us that physical creation it is possibly just an illusion of time and space, and that true science expands to a far greater reality. This is in the metaphysical realm that is just another ridiculous social construct depicted as unavailable, out of reach and out of bounds. Is it then satanic and spirituality just a myth? Among many other things, the existence of metaphysical realms suggests that physical creation is a small part of a greater illusion and that there is an ultimate reality. It suggests that there are discrete life forms in creation and that therefore it is worth arguing about what they are, until such time as we recognise their significance as conscious entities that serve a purpose. I can’t wait.
November 3, 2019 – Lawrence asked for Watsonia pictures. This is a smallish red-flowered plant from an unusual remnant of vegetation here in Fisherhaven. It is a truly fascinating problem examining the issue of how do we know that what we know is true. It is an absolute act if faith to imagine that being presented with a monograph that you will consequently be able to identify the species it deals with – and no offence whatever to the author. ♦
96. 2019.10.29 – These are pictures of H. bayeri that Gerhard Marx sent me and he knows the species better than I do.
Here are the only other pictures I have of H. bayeri. I wish I did know more about that group of retuse-leaved species from the eastern Little Karoo e.g. springbokvlakensis, bruynsii and comptoniana. I have memories of odd individual plants from S Oudtshoorn to the western end Baviaanskloof as well as several reports of finds that need explanation.
Reviewing my thoughts on springbokvlakensis and bayeri in respect of my perception of the greater retusa group, I think someone with an open mind, should really consider the possibility that this is also a single gene pool? I am busy with a short discussion of the perception we have about species and how we may just be locked in an acceptance of a mindless species concept. Do you remember what a botanist said about using a species definition based on non-interbreeding elements and morphology simply being used as a surrogate? ♦
A review of the plants photographed recently around Albertinia … I only know of 23 populations in this area that relate in different ways to the names Haworthia turgida, H. retusa, H. mirabilis, H.emelyae, H. pygmaea, H. splendens, H. vincentii, H. acuminata, H. fusca, H. dekenahii, H. argenteo-maculosa, H. pygmaea, and H. esterhuizenii. I still need to get pictures of H. splendens, H. vincentii, and H. turgida on the Wydersriver.
In essence there are two variants – the clumping cliff dwellers (H. turgida) and the mostly solitary flats dwellers (H. retusa). The flat dwellers are far more variable than the cliff dwellers. If all the variation is considered against the differences in each of the habitats, it seems very unreasonable to suggest that these populations are anything but a single species (H. retusa) expressing itself differently according to local dictates and demands. This is same issue of variability occurs in these plants all the way westwards.
97. 2020.03.04 – Esterhuizenii – Aasvoelberg. I was concerned that I am not sure anymore of flowering time because essentially retusa and mirabilis are separated by a late summer vs a spring flowering. None of the populations I saw now in late February showed any sign of buds or flower. Only the Humor plants had just two old inflorescence of about two months age. The fact it is a SYSTEM. The retusa elements are lighter coloured signified by having an element “pallidifolia”, while the mirabilis element has the element “nigra”. Recognising species as systems, variation and differences can be seen in a more realistic way. This does help to overcome the pressures that ignorance, egocentrism, emotion, commercialism and intellectualism bring to bear on classification. H. retusa ‘esterhuizenii’?
The map is useful although it does raise the question of how confidential should we be about locality. Part of this quest was to examine how populations have fared in the years since “discovery” and also in the 50 years since I have been directly involved. My observation is that the main threat is excessive animal pressure – and/or combined with drought. The Aasvoelberg population seems to be very small and a secondary threat is alien weed infestation. But I really do not know the actual size and health of the population. I did see plants further west long ago, but it will be a real mission to explore there properly. An important point about conservation is that it is really only as systems that species can cope with natural and even man-imposed change. It is only with a proper and true definition of the term “species” that we can even discuss them sensibly. “Adaptive” must surely be part of that?
98. 2020.03.05 – This is H. retusa (turgida) at Buisplaas on the Gouritz River between the Gouritz Bridge and Herbertsdale. This form was informally known as H. rodinii and is known from the Gouritz Bridge and also south of that. As a cliff dweller it is normally clumping. The first time I saw such a plant it was a specimen reputed to have come from Die Hell much further north in the Swartberg Mts. VERY unlikely, but H. retusa should be sought in the Gouritspoort where the river passes through the Langeberg Mts. There are many populations of these cliff dwellers and countless variants. I do not and cannot, dispute the fact that names present a problem for suppliers and collectors, but that is not a problem that can be settled by science. Association of a botanical binomial and place is the only solution that makes sense to me. Taking out the geographical detail diminishes the sense of any name. But this series is to explain and expand on this issue.
These are also Buisplaas but in a totally different habitat several hundred meters from the cliff dweller. One picture shows a very proliferous clone. This same juxtaposition of clumping and non-clumping occurs elsewhere and is a pronounced feature of the H. cooperi and H. cymbiformis problem in the Eastern Cape. These Buisplaas populations validate my conviction that the Western cape retusoids are in fact a single species system. There has been a predictive element in all my exploration since the revision of 1999 Haworthia Revisited and this population simply fits a pattern. Comparing all the pictures of this series I am now showing will demonstrate this too. H. retusa ‘pygmaea’. I write the name like this because in the absence of a species definition, formal lower ranks have to be meaningless. Furthermore the nature of local variation is such that it is generally impossible to circumscribe ranks. A very good example of this is the persistent reference to H. mirabilis var. magnifica which can absolutely not be supported on the basis of a type specimen representing an original group of such similar or like plants.
99. 2020.03.07 – Despite the drought and animal activity (and human predation presumably) – the Cooper Siding plants are in great health. I am posting all my pictures (of Haworthia – there are many other plant species of significance present) because the reality of diversity within populations gets zero attention in citizen science and little more in academia. But do not get me wrong. I write “citizen science” not citizen cognition. Also I do not ignore the fact that “citizen science” also features in academia. These plants (a plant?) was described as H. dekenahii var. argenteo-maculosa. In my Handbooks 1976, 1981), I placed the two under H. retusa. Was this wrong? No – because moving them to H. pygmaea as I did in my revision (1999) highlights the reality that this is a single system and not just spatially isolated populations that can be treated as different.
In writing about Albertinia and the conservation state of these Haworthia populations (that really bothers me), some very serious thoughts come to mind. Do we really want to know the problem and the answer(s)? The problem is environmental deterioration – climate change. This is coupled with the decline in moral values across the counter. But do we want to acknowledge this and really want to know what is true. That all of existence is actually a grand illusion that we may choose to be locked in to. I should not have suggested that climate change is the main factor – although Albertinia has been experiencing the worst drought in living memory. Livestock pressure also definitely impacts negatively on vegetation, and our dietary addictions prevent us recognizing this fact.
100. 2020 03.08 – Dekenahii is at Draaihoek on the Valse River northeast of Albertinia. Juxtaposed again on a nearby rocky cliff is “turgida pallidifolia”. A floribunda / chlorocantha variant is in close proximity in its own preferred space. A different form of the turgida retusoid used to be on north facing cliffs a little further north at Weltevrede. Keep in mind that I am considering a bigger picture in which differences within populations negate differences between them, and especially when habitat is considered. I also juggled dekenahii between retusa and pygmaea and whatever you do with it, the best explanation I can offer as a binomial is H. retusa ‘pygmaea’. Better still is to add the unwritten meaning that it is actually Helgaard Oosthuizen’s discovery of H. retusa as a group of plants at Draaihoek circa 1943 given to Smith by Dekenahii. Names standing alone may be good for business but the interest lies in the information attached to them and which they point to.
This is the “turgida” form of H. retusa at Draaihoek. These plants have more pointed leaves and are more colourful than the “pallidifolia” that I remember 59 years back. But the precise locality may have been different. I remember plants more like those at Tweekuile further east along the river that I will still post pictures of. At Weltevrede further upstream the plants had longer leaves. Steven Molteno coincidentally reminded me of H. marumiana and the exact same conundrum I am trying to explain here, but magnified manifold over the vastness of the Karoo and our ignorance of both marumiana and the Karoo.
101. 2020.03.10 – Tweekuile is north of Albertinia and just a few km east of Draaihoek on the Valse River. This is what H. retusa ‘turgida’ looks like there. It occurs to me that sensible taxonomy, like prevention and cure of disease, is not a sustainable business model 😅
Without any simple and practical measure, how does one deal with an endless array of slightly different looking things affected by age, exposure to direct sun and an also seemingly endless lot of agencies affecting appearance. Compare these two Cooper Siding and Humor pygmeas with the Tweekuile turgidas.
102. 2020.03.11 – Have a good look at the rest of Humor. I suppose it is still fashionable to dismiss variation like this as “hybrid swarm”. Problem here is what parents where and when. To be strictly correct, is there really any option but to say H. retusa ‘pygmaea’? ♦
103. 2020.03.11 – Melkhoutfontein. The weird history of “acuminata” is not very flattering. It started with me when I described this as a variety of H. retusa. But I did say some interesting things in 1981 that lead to my present point of view. One is that I said that acuminata may be eventually shown to link H. retusa and H. pygmaea. This is exactly what the present thread is about without any conscious effort to support what I might have predicted. I think I may have changed acuminata to a variety of H. mirabilis in Haworthia Revisited? Weird too, but I would not be ashamed. That would reflect the reality – it is one system and countless opinions can be woven around the evidence that supports the idea. The original population of acuminata was destroyed and although I did find plants nearby I did not think such a strong population as this still existed so close by. This population is at Melkhoutfontein on the Gouritz River southeast of Albertinia. The habitat is old river terrace. There were several smaller populations to the north. Coming are populations to the east that inspire further support for the kind of continuities that occur between practically all Haworthia populations.
104. 2020.03.16 – Johnson’s Post plants are also something else. Large clumps on clay pedestals on an ancient river bank? An amazing sight.
105. 2020.03.17 – These plants are at Vleesbaai and I have no doubt that a species name has been coined for them without condemning the need for a name. It is the formal species part that bothers me. From my perspective these are H. retusa ‘pygmaea’. Steven Molteno can give a good habitat description. The conservation aspect is interesting because it is a small rocky area in an expanse of cultivated and heavily stocked lands. The real threat to the plants is the severe stocking pressure and the irony is that grazing is also necessary to keep vegetation under control. I will, I’m sure, come back to the conservation issue.
Steven Molteno – If these are the plants I’m thinking of (they look like it) then they’re all growing on a lone outcrop of quartzitic sandstone of the Skurweberg formation (Ss), Table Mountain Grp. This exposure is a rather eroded ridge very near Vleesbaai, surrounded by a lot of calcrete and alluvium I think.
There’s another (much larger) exposure of this same formation (Ss), not far away to the north. It’s that line of tall hills that runs parallel to the N2, south of the road, and east of Albertinia. Interesting that this larger Skurweberg formation ridge is also home to some unusual mirabiloid-retusoid confluences (the ‘vincentii’ & co. towards the western end of that ridge of hills; pygmaea ‘argenteo-maculosa’ towards its eastern end).
I think this correlation with a specific geological formation is very suggestive of broader connections across this whole Gouritz region.
Soils are very rudimentary on these outcrops. Lithic leptosols, plants growing very close to (or in) bedrock. Eroded grit from quarzitic Skurweberg formation is probably rather acidic, especially with the contribution of broken-down organic material that’s pretty much its only supplement!
Up near Johnson’s Post it’s a completely different story though.
There, the geology is very complex, with lots of funny “slivers” of different formations, all parallel to each other but zig-zagging across the flats.
At Johnson’s Post though, the river’s carved out a series of little kloofs in the terrain. The central valley floor is quite wide and filled with alluvium. The haworthias grow along +-north-facing slopes of Kirkwood formation (Kk). It’s often reddish, but here it’s a light-brown (almost a little ‘greenish’?) siltstone with greyish shales and bit of sandstone sometimes. This is what was called the “variegated marls”, and it erodes into a heavy, fine-grained clayey loam. Very muddy if wet. The same formation (Kk) is the main home for H.pygmaea in the Mossel bay and Brakke regions to the east. There is a bit up near Herbertsdale, and a long strip of it going east-west through Riversdale and Heidelberg. Another patch is just east of Swellendam, and I think there’s probably an exposure at Klaasvoogds but its very eroded. Everywhere except Klaasvoogds, it’s inhabited by the floribunda/chloracantha complex. Sometimes also these funny mirabilis-retusa integrades like near Heidelberg.
To return to Johnson’s Post, above the Kirkwood formation you get this pebbly calcarenite (De Hoopvlei fm I think), but it’s on the flatter area on top and I don’t know of Haworthia there. It’s also mostly ploughed though.
Landtypes at Johnson’s Post are also interesting. Eastern part is Fc15. Fc15-39 are lime-rich river valleys, networking across most of the Overberg, that are always inhabited by retusa ‘turgida’ and by one of the floribundoids. Usually not a mirabilis though.
Western part of Johnson’s Post is landtype Ae133 (obviously these aren’t strict boundaries though, they grade). The south-eastern extremity of the same landtype (Ae133) is where those Haworthia at Vleesbaai live – i.e. Ae133 connects these two populations.
There’s a similar landtype, Ae113, just north of the Breede River mouth (!)
Vegtype at Johnson’s Post is what Jan Vlok calls: Southern Cape Valley Thicket. It’s a thin belt that winds along the Gouritz and its tributaries, with a strip along the lower Goukou too. It nearly always has some sort of toothed-leaved floribundoid/chloracanthoid (along the lower Goukou its H. variegata) as well as H. retusa turgida. The correlation in distribution is quite strong. These haworthias almost seem to trace out this vegtype on the map. I think the correlation with a specific geological formation is very suggestive of broader connections across the whole Gouritz region. These sandstones are very much older than the surrounding sediments.
106. 2020.03.18 – This is Paulsfontein just west of Albertinia. A small population in an intensively grazed and weed encroached area. A reminder to keep in mind within and between population variation. This has the Latin epithet ‘fusca‘ and I personally fuddle over whether to address it as pygmaea or mirablis. So I suggest H. retusa because these populations simply mirror what I was showing for populations to the west where names like mutica, groenewaldii, badia, joleeniae, bobii, hammeri etc. are collector useful. Conservation is poking me in the ribs but I am not finding the right ambit for that discussion.
A very powerful supporting argument for my thesis of H. retusa as a seemingly massive conglomerate, is the close association with H. floribunda. H. chloracantha, H. parksiana and H. variegata; as well as in the continuities one sees among those ‘species’.
Essie Esterhuizen wrote (31 July 2021) … Populations in that area confuse me. I am not sure whether I deal with the western end of Haworthia pygmaea or eastern end of Haworthia retusa.
Bruce Bayer replied … Delighted to hear Essie’s voice. Essie I do not know what you have followed of mine and I have no idea of where your thoughts have taken you. My conclusions are that we have all got things wrong about botanical classification and that Haworthia is just showing us. So I think species are vast fractal systems. At the least H. retusa (incl. turgida) mirabilis, mutica, pygmaea and emelyae are a single system?
I am so glad Essie has commented because I have just felt so in need of expressing my present feelings about the aloids generally. There was a congress on Aloe classification about 10 years ago that was hailed as a breakthrough for the subject. I saw it as a total farce because the essential nature of species was ignored. My experience and slight knowledge of the DNA methodology applied in the latest solution for the genera (and species), is that it is supremely cringeworthy. All it gets right is the three Haworthioid genera where sequencing was not needed to reach that conclusion. Part of my fears for the worst are based on the fact that two of the major sequencing studies were initiated to explore the phylogeny of just Haworthia species. That the results were written up to totally avoid discussion of the initial hypothesis will never cease to confound me. My confidence in aloid classification is at zeropoint.
107. 2020.03.19 – I wish I had pictures from my visit to this site about 20 years ago. The place? Lodewykstenk east of Albertinia and east of Aasvoelberg – esterhuizenii. There were many plants. This time I really struggled to find only these 3. Why? Certainly not collectors. Drought and severe animal pressure. The plants have the name H. vincentii. But really. Can you name plants in casual fashion like this without a knowledge of what the name is directed at? A basic life form. I consider that it fits comfortably with all these plants I have been dealing with here.
108. 2020.03.19 – Flat or cliff? What the thesis is that these plants belong in a single gene pool and the enormous variability is due to the demands of habitat as much as anything else. Cliff is vertical rock. A level habitat may have the same rock to soil ratio. Whatever the vast range of non-steep rock sites, most significant is that animals (livestock) cannot graze or trample a vertical rock wall? This is then the cliff hanging form of H. retusa east of Tweekuile, north of Albertinia, but a bit off the Valse River. A fortuitous new record to demonstrate just how much exploration is still needed. ♦
109. 2020.03.22 – Soutpankoppies. What is there to do? The Albertinia area is really absorbing. Westwards the gene pool (retusa/mirabilis/turgida/mutica) is confused as it tries to express itself in so many different ways, whereas at Albertinia it simplifies to two and, in my opinion, then one. At the same time there is a second gene pool comprising floribunda/chloracantha/variegata/parksiana. The area is not well explored and there are clearly small islands of vegetation that may still hold some secrets. The photographs here are of ‘splendens’ at Soutpankoppies that I perceive to be the most eastern expression of ‘mirabilis’. Retusa in its typical form ends a little to the west. Further exploration in the Albertinia area will surely expose the facts about these two hypothesized gene pools.
“Splendens”. Not for nothing is it specially protected in an enclosure. But this underlines the real problem of conservation – a huge subject. This particular enclosure is of interest for several reasons. When I first heard of it I assumed it was where the plants were seen in ca 1996. But it is not. So what has happened to that small population? There was also a third ‘population’ in the immediate vicinity of an old small farmstead. Local disturbance and alien weed invasion seem to have annihilated that one. The splendens enclosure is hugely successful in respect of the short term health of the specific population and even satellite imagery demonstrates that. The impact of livestock activity and weed invasion outside of the enclosure are obvious. But the total effect is confounded by the presence of surface rock (ferricrete?). The splendens population would have been as safe in the absence of any fence. The other two populations were far more vulnerable. It is simply not practical to enclose vulnerable ‘species’ and this was a hot topic in professional conservation as far back as 1970 if not before. We cannot ignore the problem of taxonomy, because the essence of conservation is surely the preservation of diversity. At what level now does one do this? The current trend in conservation is the identification of “red-data” species. I think this is complete nonsense because it amounts to plum-picking in which the end-product is the maintenance some kind of museum record of what was?
110. 2020.03.23 – An enthusiast commented that Haworthia magnifica made sense to him. Well here are pictures of plants at the type locality (Riversdale Commonage or now known as the Werner Frehse Reserve) as was indicated to me by the collector of the original material i.e. J. Dekenah. If you want confirmation that the name, as a species name, makes sense it is worth using google to locate the type and see the images that come up. What would be useful is for that enthusiast to set out to demonstrate why this particular name makes sense to him against all the other names that may do the same.
This is probably what H. magnifica is imagined to be, based on my experience with the name, the trade and the literature. But the fact is the population at the Frehse Reserve is substantially fragmented and the plants are highly variable. Not only that, anyone who has seen a retusa/mirabilis hybrid will recognise that influence in the pictures I’ve posted.
111. 2020.03.25 – Here is a set of pictures of plants propagated from seed from a H. mirabilis population (MBB6651) 2km further south from the Frehse Reserve and what was purported to be H. magnifica. The habitat is a bit different because here the plants are in white clay (kaolinite) whereas at Frehse they are in shale. What is common to these plants that makes them different to an impossible to define commonality of H. magnifica???? It is a problem throughout ALL the mirabiloids and retusoids of the Southern cape and extending to the emelyoids of the Karoo. Examine the last picture and you will see the ultimate trickster. The floribundoids (variegata, chloracantha and parksiana) have a show-in too.
The coloration in this plant is what we see in the old ‘mutica var. nigra‘ population. When one talks of magnifca one cannot dismiss a series of mirabilis populations that occur from Frehse Reserve at Riversdale westwards to Riviersonderend. ♦
112. 2020.03.26 – The trickster – H. floribunda. This is the Draaihoek Albertinia site where I was not sure (ca 1970) if it was floribunda or chloracantha. I went back to check ca 2001. The place was grazed and trampled to a frazzle – sheep and/or ostriches. I found a few specimens right at the far end. They pass for floribunda and especially if you recognise that floribunda has a good many faces. The last picture is from west looking east – so this is the top rocky edge of a south facing drop. The water is the Valse River. Dekenahii is across the way.
So in these very distressing times, lets see where we go next.
113. 2020.03.26 – This is H. chloracantha from east of Tweekuile along the Valse River north Albertinia, so also further east from Draaihoek. A small dense patch of plants under bush at the upper end of a south facing slope – escaping full exposure to sunlight. A few leaves show the round leaf tip of floribunda but the plants are also much more proliferous than I have ever seen in that ‘species’. It is very useful to consider all the variants in this complex to see how and why I arrive at my opinions on their classification. This is not a glamour complex but as systems they work in exactly the same way. I really regret not having good pictures and especially since I first saw this species at Great Brak in 1969.
114. 2020.03.27 – Ouvloere is east of the previous as that is from Tweekuile but the plants are really odd. Unlike the previous these plants are in and under low ground hugging plants at the top of a slope. This must be about 8km west of the Gouritz River.
How would you know this is H. floribunda?
The answer to … How would you know this is H. floribunda is because I say so? So please forgive my apparent arrogance. I would dearly love to show you all the plants and populations that lead to this conclusion because you would surely be able to then make that decision for yourself. These two pictures are also floribunda from very near to ‘splendens’ at Dekriet (Snymanskraal) west of Albertinia. A strange habitat where the rocky ferricrete is exposed but with a collapsed eroded depression in which the plants grow around the edges. When this time of tribulation is over, I hope to go back and get habitat pictures. Reminiscent of parksiana?
115. 2020.03.30 – The Valse Rivier joins the Gourits at Die Eiland where there is now H. chloracantha and eastwards. It supplants floribunda entirely – or is it truly just the same species?
116. 2020.03.30 – The last post was of plants from N Die Eiland (Buisplaas). From the top edge of a west facing shale cliff. These are also Buisplaas but from a low north facing slope of alluvium. I never imagined chloracantha could be so abundant and so resistant to the impact of habitat. But it becomes one of the most remarkable of the species on closer scrutiny.
The great awakening will be when one can truthfully and openly write about, discuss, the reality of basic life forms i.e. species. When we are free of the prejudices, beliefs and misconceptions of science as practiced and religions as preached. The great awakening will be when one can truthfully and openly write about, discuss, the reality of basic life forms i.e. species. When we are free of the prejudices, beliefs and misconceptions of science as practiced and religions as preached. ♦
117. 2020.03.31 – This is just north of Herbertsdale under renosterbos on shale. A serious lack is pictures of a population nearby on a very steep, north-facing crumbling clay ‘cliff’ with karoid bushveld plants growing wherever they can fit. Here the plants make enormous clumps and themselves are individually large to leaves 60-70mm. Also missing are pictures from a short way to the south-west on an east-facing cliff of conglomerate. There the plants are small, solitary and dark-colour red 😊. These demonstrate the impact of habitat. Of course also seriously lacking is a species definition and agreement on the issue. Systems, systems, systems.
118. 2020.03.31 – This is chloracantha further south at Johnson’s Post with ‘pygmaea’. Large dense clumps seemingly holding clay pedastals together. These are associations of systems and in my opinion very significant in recognising where species stop and start. Not that this simplifies the problem. Incidentally chloracantha in a small green form also occurs in the Gouritz River gorge through the Langeberg.
119. 2020.04.01 – Chloracantha occupies an incredible range of habitats and varies accordingly. These are it at Cooper Siding in feldspathic sandstone. Not far away it is also in transitional zone to shale and the plants also looking a bit different. In both cases it is with “pygmaea” and I have see hybrids despite the season difference in flowering times. Curiously I was once asked … “how do you know it is a hybrid?” Well I do not know how to answer that. It is like asking, “How do you know you which is your left foot?”
Lawrence Loucka – Please say some more about hybrids. ‘I know it when I see it.’ may be true but how to confirm? What do you see – parent proximity, morphology, flowering overlap, habitat? If not hybrid, then what?
Bruce Bayer – I think you are asking me to identify my left foot. What can it possibly be if it is not derived from the plants before your eyes? I forget now the name of the hybridity test based on mensuration and statistical analysis. I tried to use it on what were such obvious hybrids between herbacea and reticulata – and as you well know in H. mirabilis var sublineata, getting significance requires an inordinate sample size. So of course it is parent proximity. whereabouts of the “hybrid”. And what is difference in flowering time where there is 4-5 months between peak flowering time of each putative parent and a flowering time that can be acted on the the seasonal rainfall – and individual plants that simply do their own thing. With DNA we cannot even separate the species so how do we use that? It comes down to nitpicking and finding a way to discount the obvious. How do you confirm that all the plants before your eyes are two species when you have no definition of what the word means and cannot specify the actual criteria that are common to the respective entities? That is why it is possible to just come out with names like hammeri, joleeniae, vincentii etc. and find an innocent accepting audience? 😄 Best way of looking at the issue.
Jakub Jilemicky – Aren’t these floribunda v. dentata?
Bruce Bayer – Jakub, I would like you to detail the differences between floribunda and chloracantha. There seems to be a serious misconstrual of classification here. First you have to arrive at the species name and then at the next level down. In Haworthia I have virtually abandoned “nextleveldown” because the species level is already confounded. You cannot get a variant of one species in a population of another where the populations and species are arranged and derived as they are in Haworthia. Can someone explain it better? Prof. Cronquist once asked of his taxonomic proteges and fellow experts…”Do we know what we are doing?” By and large haworthia taxonomists do not even seem to know what taxonomists even should be doing. 😄
Jakub Jilemicky – True, this group of plants are very challenging. I’m using your books as a source of taxonomy. I have seen quite a few population from Hartenbos in the East to Riversdale in the West. And I’m lucky to grow quite a big collection from many sources of this plants, so I can compare them both in cultivation and in nature. H. floribunda v. dentata seems for me to continue into chloracantha v. denticulifera. Around Gouritz river it is almost impossible for me to distinguish them. Towards Herbetsdale the same happens with chloracantha. There are a few characteristic true chloracantha populations north of town, others are resembling floribunda. Yes, there can be selected true TL plants, which are characteristic, but all between seems for me to be one big continuity🤷♂️
Bruce Bayer – Yes Jakub, it is not just this group. I think I actually generated the name floribunda var. dentata. In retrospect it was a stupid thing to do because there is no such a taxonomic entity IF you realize that “variety” should mean a population entity and not “variant” meaning individual variation – in which case “form” would be better? My posts are doing what you describe – showing that there is 1/2/3? big continuity/ies.
Cooper Station – Jakub Jilemicky
Bruce Bayer – Thanks Jakub – I also very recently posted toothed chloracantha at Copper Siding. I say chloracantha only because there are none of the stronger floribunda features i.e. smoother surfaces, rounded leaf-tips, keel absent for large part of leaf end, and greyish green. The outward turning leaves are more floribundoid than chloracanthoid and we cannot forget parksiana. If we could just get over this hurdle of talking sensibly about these obvious mismatches and evidences of continuity, we might begin to properly look at real problems like the Hex valley plants. the Prince Albert problems, grasp what Kaboega populations tell us and many more.
120. 2020.04.01 – Stay calm, be patient, all things will pass. I found I had pictures of ‘pygmaea vincentii‘ from Welgevonden = Lodewykskraal east Albertinia. Was there with Kobus and there were countless plants then – August 2008. They were not well drawn into the soil and perhaps very vulnerable to close grazing? In this crisis it is worth considering that nature does not need us and can smile at our puny efforts, destructive ways and thoughtless actions. Also bless us for the few things we may be doing right. It is all under control.
Frantisek Vesely – Hello Bruce, all these plants were growing on the same spot?
Bruce Bayer – Hello Frantisek. It is a north facing rocky slope about 1km wide and I remember many plants all along the slope in about a 50m wide band. That was 2008. This March we could only find 3 plants very shrunken into rock cracks. It is possible that others are simply sand and debris covered in a very desiccated state and rain may bring them out.
Jakub Jilemicky – I was there in 2011 and after few hrs search we found only 5-6 plants in rock crevices…
Frantisek Vesely – Thank you. I would say there could be some interaction of var. esterhuizenii with var. turgida/suberecta. That would explain why the plants which are considered to be H. vincentii just look so odd but on the other hand they are somewhat intermediate between these two.
Bruce Bayer – Thank you Jakub – useful info
Bruce Bayer – Frantizek good observation. I wish I could express or, firstly, understand it better. It is not necessarily “interaction” but rather a question of distribution of genetic material on the basis that there is a single gene pool interconnected in some incomprehensible way and expressing itself on the basis of localised habitats. It is difficult to believe that at one time there were a great deal more populations and more gene exchange between them than there is now. Perhaps that was the case. The nearest “turgida” to vincentii is “rodinii” but we have the problem that here in the east the total gene pool is reducing from a mirabilis/retusoid (incl turgida) to retusoid (incl turgida)/ pygmaeoid (incl retusa/emelyaoid). Vincentii would represent the fade out of mirabilis via splendens and esterhuizenii. Dekenahii and fusca the fade out of retusa and fade in of pygmaea. It is something that needs exploration both literally and figuratively in the mind.
121. 2020.04.02 – This is a population from east of Herbertsdale. Here they grow in kaolinite and are buried up to their leaftips. Very much like those decaying white ‘pressure burst’ common in the Riversdale area and of course westwards. In the first picture of three clones, the one on the left is a plant from S Herbertsdale. How can you now name this population? There is something very wrong in the way we use and apply names? What do we mean by the word “species”. To say this population 3km E Herbertsdale has elements of parksiana and floribunda implies that they truly are different species. How would we know? ♦
122. 2020.04.07 – Like with the present pandemic we are coming to something difficult to comprehend. I went to Botlierskop, Rooiheuwel farm, north of Klein Brakrivier hoping to see a slender long-leaved form of Haworthia chloracantha (a JDV record). Instead I came across this population of rather stubby plants. Keep this population in view – two close populations to follow and then an issue of proximities!
Also on a rock slab about 70 meters away. Is this still chloracantha or is it parksiana?
Frantisek Vesely – We saw the similar situation – on slope there was chloracantha (+ hybrids), on flat a few dozens meters away there was parksiana and its hybrids with chloracantha – not difficult to recognize what happened there as they were so close and mixed up but on one spot most of them were chloracantha-like and the other spot were more parksiana-like plants.
600M away there is a population of uniform parksiana. But how can we say hybrids for the previous? How long have these populations been in existence? Is transfer of pollen a problem and over what distance? There seem to me to be a host of unanswerable questions in general about hybridization as an explanation for variability. It is linked to the more general question of co-occurrence – of mixing or non-mixing of species. Ring clines? Also something we should really try to document – i.e. the proximity of the various species to one another. Add to that the occurrence of hybrids.
123. 2020.04.09 – This population, east of Riversdale, is an aspect that really calls for attention – proximity of kinds (species) and hybridization. At this locality there is H. retusa and H. floribunda in contiguous/adjunct/closely neighbouring populations. Among the floribundas there is a single putative hybrid? How often does this happen and why are kinds (species) so seldom growing together? When is an entire population comprised of hybrids? What comes in between? Let me just add that I see very little in the taxonomic literature dealing with the problems of variability that exist in many genera other than Haworthia. It is simply obscured.
I am quite aware that my comments are often cryptic and often deliberately so. As a society we are very misled and never more so than in respect of the present pandemic. I do not see how we can put that aside and think that we can at the same time depend on our collective intelligence to solve and understand anything else e.g. taxonomy of Haworthia. It is a conscious creation and that consciousness has to be lifted to a level that we see things for what they really are. Hybridity is at the root of species definition and as such a critical step to recognising species. At the same time geographical location is also critical. To to think anything can be explained while there is a paranoia about revealing it, is wishful thinking.
2020.04.12 – There is a problem with classification where priority creates difficulty. The name Haworthia retusa has nomenclatural priority over the name turgida, when in actual fact the turgida form is the main one and retusa is a variant of that. It is much easier to then understand, describe and explain the idea that retusa becomes mutica in the west, retusa and mirabilis become pygmaea in the east, while turgida and mirabilis become emelyae in the Karoo. That might be where I should now leave things. There are a hoard of complexities that just get lost in the needs and activities of the majority who have no need of much more than a lot of desiderata. ♦
Lawrence suggested I finish what I had to say about the retusa combobulation. Actually I find myself just plain exhausted trying to explain why I name things the way I do when so many otherwise knowledgeable enthusiasts are in real ignorance about the fundamental problems of Haworthia classification and identification. I was just writing about populations of plants in a very wide area between Steytlerville and Oudtshoorn, explaining that it was nigh impossible to decide whether they are decipiens, mucronata, bolusii or cooperi once the easy conclusion was made that they were not arachnoidea. This all reduces argument in the retusoids to absurdity. Someone tell me what these are and why?
I felt the need to say something about how, at the end of my useful life, what my feelings are about (my) classification of Haworthia. It so happens that I was extraordinarily fortunate to have had the confidences of some very distinguished people as they closed their lives down. People like Prof. R H. Compton, Dr R.A. Dyer, Dr Alan Codd, Mrs A. Obermeyer, Dr Courtney-Latimer, Dr O. Hilyard and others in botany. also in other sciences like Dr H.K. Munro, Dr B.K. Petty, Dr B. Smit from entomology. Col. C.L. Scott came to the same place. The common factor was an admission that really they did not know very much. I thought a word that would cover the state of mind would be “pragmatism”. That is until I checked the definition of the word, “Perceptual inattention to the specific functions comprising inquiry led realists and idealists alike to formulate accounts of knowledge that project the products of extensive abstraction back onto experience.” What about the product of my experience “experience” – the knowledge that you acquire by the time you do not need it anymore) projecting abstraction??? The funny thing is that the rules of nomenclature confound the purpose of classification. The purpose of classification is to identify all the basic life forms and to organise them in the system and order of their historic origins. Just as pragmatism can be left unexplained and confounded in many words and pages of incomprehensible text, so is my classification of Haworthia. A point is “groenewaldii”, where I suggest it really should be H. mutica var. groenewaldii. This is a conceptual trap! Combobulation cannot be avoided. All the evidence I see is that there is a single system that what we call turgida is the strongest possible contender for origin. The problem is that nomenclatural rule is that if retusa has its origin in turgida, the name retusa must be upheld for the system. Do you follow that??? In my explanation of the “species” mutica, I explain it as emanating from the interaction of mirabilis and retusa. So it is quite wrong to use the combination of mutica var. groenwaldii. It should be retusa var. groenewaldii and still more correctly H. turgida var. groenewaldii. I am presently really just enjoying sharing my thoughts about some “species” that will surely trouble anyone wanting certainty in identification and names. ♦
In a series of postings in the summer of 2019, Bruce Bayer discussed the taxonomy of Haworthia retusa and related H. turgida, H. pygmaea, H. mirabilis, H. magnifica, H. emelyae, H. mutica, H. groenwaldii, H. floribunda. Here are supplement references. ~ Lawrence Loucka
H. retusa (L.) Duval in Pl. Succ. Hort. Alenc.: 7 (1809): Aloeretusa L. (1753). Lectotype, designated by Scott (1985): Illustration in Commelin, Horti Med. Amstelod. 2: t. 6 (1701). Epitype, designated by Breuer & Metzing (1997): Blikbonnie, E Riversdale, Dekenah s.n. NBG144772 (NBG).
retusa: with leaf-tips bent back thumb-like.
Rosette stemless, slowly or seldom proliferous, to 12cm φ. Leaves 10-15, turgid, rigid, with pronounced retused end-area, pointed tips, variously lined and windowed. Without surface spination and usually without spination on margins and keel. Colours brownish or green and seldom purpling. Inflorescence simple, robust, to 30cm. Flowers compacted on inflorescence, white with greenish-brown veins.
Haworthia retusa (L.) Duval J. Commelin, Horti medici Amstelodamensis rariorum tam Orientalis, vol. 2: t. 6 (1701) http://plantillustrations.org/illustration.php?id_illustration=122188Haworthia retusa (L.) Duval [as Aloe retusa L.] Botanical Magazine (Curtis), t. 433-468, vol. 13: t. 455 (1799) [S.T. Edwards] http://plantillustrations.org/illustration.php?id_illustration=7534Haworthia retusa (L.) Duval [as Aloe retusa L.] A.P. de Candolle, P.J. Redouté, Plantarum Historia Succulentarum (Plantes grasses), vol. 1: t. 45 (1799-1837) [P.J. Redouté] http://plantillustrations.org/illustration.php?id_illustration=48020Haworthia retusa (L.) Duval [as Aloe retusa L.] J.S. von Salm-Reifferscheidt-Dyck [Salm-Dyck], Monographia generum Aloes et Mesembryanthemi, Aloe, vol. 1: , fig. 9.3 (1849-1863) http://plantillustrations.org/illustration.php?id_illustration=184034Haworthia retusa (L.) Duval [as Aloe retusa L.] Collection des vélins du Muséum national d’histoire naturelle, vol. 8: t. 63 () [C. Aubriet] http://bibliotheques.mnhn.fr/medias/detailstatic.aspx?INSTANCE=exploitation&RSC_BASE=IFD&RSC_DOCID=MNHN_VEL_PORTEFEUILLE008_FOL063Haworthia retusa (L.) Duval J. Moninckx, Moninckx atlas, vol. 3: t. 7 (1682-1709) http://plantillustrations.org/illustration.php?id_illustration=138251
H. retusa var. turgida (Haw) Bayer (2012)H. turgida Haw. in Suppl. Pl. Succ.: 52 (1819). Neotype, designated by Breuer & Metzing (1997): Swellendam, Breede River Bridge, Bayer 2420 (NBG).
turgida: swollen.
Rosette partially stemless, proliferous, 5-10cm φ. Leaves 20-40, turgid often as thick as broad, recurved or slightly retused, generally mottled, yellow-green to pink in sun, margins and keel lightly spined. Inflorescence simple, 15-20cm. Flowers slender, brownish-white with darker venation.
H. pygmaea V.Poelln. in Repert. Spec. Nov. Regni Veg. 27: 132 (1930). Neotype, designated by Breuer & Metzing (1997): hills E Great Brak, Fourcade 4759 (BOL).
pygmaea: dwarf.
Rosette stemless, slowly proliferous, 6-10cm φ. Leaves 12-15, retused, round-tipped, surface pellucid with obscure raised tubercles, sometimes intensely papillose. Inflorescence simple, robust, to 30cm. Flowers white with greenish veins.
H. mirabilis (Haw.) Haw. in Syn. Pl. Succ.: 95 (1812): Aloemirabilis Haw. (1804). Neotype, designated by Bayer (1977): Illustration in Curtis’ Bot. Mag.: t. 1354 (1811). Epitype, designated by Breuer & Metzing (1997): Skuitsberg, between Caledon and Greyton, Bayer 2453 (NBG).
mirabilis: wonderful.
Rosette stemless, proliferous, to 7cm φ. Leaves 10-15, retused, 3-4cm X 1,5cm, markedly retused, acute above, face translucent and lined, dark green, with marginal spines turning reddish in the sun. Inflorescence slender. Flowers narrow, elongate, biarcuate bud, upper lobes pinched at tips.
H. mirabilis var. magnifica (V.Poelln.) Bayer (2012): Haworthiamagnifica V.Poelln. in Repert. Spec. Nov. Regni Veg. 33: 240 (1933): H. maraisii var. magnifica (V.Poelln.) Bayer : 131 (1976). Lectotype, designated by Breuer & Metzing (1997): Riversdale, Ferguson.
magnifica: magnificent.
Rosette stemless, slowly proliferous, to 8 cm φ. Leaves spreading, retused to ground level, dark green to purplish, scabrid to finely spined margins, end-area slightly translucent between the veins, surfaces with small slightly raised tubercles. Inflorescence slender, to 40cm. Flowers brownish veined, few open, upper lobes pinched at tips.
H. emelyae V.Poelln. in Repert. Spec. Nov. Regni. Veg. 42: 271 (1937). Lectotype, designated by Breuer & Metzing (1997): [unpublished image] (B).
emelyae: for Emely Ferguson.
Rosette to 10cm φ, seldom proliferous. Leaves 15-20, distinctly retused, pointed, barely translucent, with scattered elongate small flecks, with obscure raised tubercles, lined, reddish-brown hued. Inflorescence simple. Flowers 15-20, white.
H. mutica Haw. in Revis.:55 (1821). Lectotype, designated by Bayer (1978): [image] (K; later published in Excelsa 8: 50 (1978). Epitype, designated by Breuer & Metzing (1997): NE Soetrivier Bridge, Bayer KG623/69 (NBG).
mutica: without a point.
Rosette stemless. non-proliferous, 6-8cm φ. Leaves 12-15, retused, blunt-tipped, brownish-green, in habitat developing purplish cloudiness, barely pellucid with several longitudinal lines. Inflorescence simple, to 20cm. Flowers white with brownish veins.
Description: Always growing singly, but in a couple of cases growing in groups of plants of different ages. Rosette: acaulescent, 40-50 mm Ø (exceptionally up to 100 mm). Leaves: 5-7(8), fleshy, spreading, end-area in most cases very rounded (resembling H. springbokvlakensis), 20-25 mm long, 13-18 mm broad, 11-13 mm thick. Leaf surface of back and leaf-base, smooth, opaque; colour dull dark green. Margins: opaque, smooth. Back without keel. End-areas: 13-18 mm long, face translucent, with 3-5 whitish parallel lines, the middle one the longest (sometimes parallel lines are ± partly of greenish colour). Surface of the end area, rough with small papillae. End area of some specimens are with ± whitish, ± raised dots, in a few cases the dots are blended into cloudy flecks. Leaf-tip: very rounded, without teeth. Inflorescence: 300-600 mm long, 1.2-2 mm diam., Peduncle: 200-300 mm long. Sterile Bracts: 15-20, 5-7 mm long. Raceme: 100-300 mm long. 8-18 flowers, 2-5 flowers open at one time. Fertile Bracts: 3-4 mm long. Pedicels: 2-4 mm long, 1-1.2 mm diam. Flowers: 14-16 mm long. Perianth: 3 -4 mm diam. Flower-face: 9-12 mm long. Upper part: 6-7 mm long, 3-5 mm broad. Lower part: 5-7 mm long, 5-9 mm broad. Fruit: 11-18 mm long, 2.5-4 mm thick. Flowering time February–March. Type Information Collector: J. Groenewald s.n. (= MBB7801) Locality: 3420 (-BA), Buffeljagsrivier, Western Cape, SA.
Haworthia groenewaldii ‘typical’ specimen with whitish flecks and parallel lines with some green colour.
Haworthia groenewaldii in Habitat. Specimen without whitish flecks, growing in between Merxmuellera disticha, a hard and unpalatable grass.
Another specimen of Haworthia groenwaldii with little whitish flecksA specimen of Haworthia groenewaldii with inflorescence among a group of smaller plants.
H. floribunda V.Poelln. in Repert. Spec. Nov. Regni Veg. 40: 149 (1936). Neotype, designated by Bayer (1982): [unpublished image] (B). Epitype, designated by Breuer & Metzing (1997): Blackdown, NE Heidelberg, Bayer 158 (NBG).
floribunda: many flowered.
Rosette stemless, up to 3cm φ, slowly proliferous. Leaves 20-30 dark green opaque, upto ovate-lanceolate, spreading, twisted with flattened, rounded tip, margins scabrid to dentate. Inflorescence simple, to 250mm. Flowers 10-15, greenish-white, few open together.
Journal of the Linnean Society vol 18 – 1881, J. G. Baker ‘On Aloinea and Yuccoidea’ – 3. Haworthia starting on 209 (page 197). H. retusa 220 (208). H. turgida on the next page.














































































































































































































































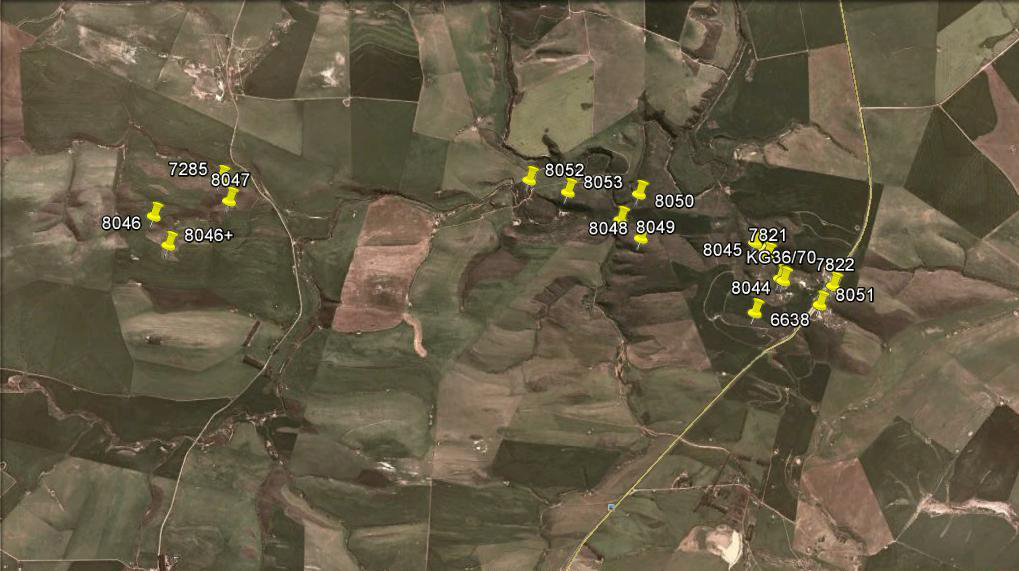










































































































































































































































































































































































































































































































![Haworthia retusa (L.) Duval [as Aloe retusa L.]
Botanical Magazine (Curtis), t. 433-468, vol. 13: t. 455 (1799) [S.T. Edwards]
http://plantillustrations.org/illustration.php?id_illustration=7534](https://haworthiaupdates.org/wp-content/uploads/2019/07/H-retusa-2.jpg)
![Haworthia retusa (L.) Duval [as Aloe retusa L.]
A.P. de Candolle, P.J. Redouté, Plantarum Historia Succulentarum (Plantes grasses), vol. 1: t. 45 (1799-1837) [P.J. Redouté]
http://plantillustrations.org/illustration.php?id_illustration=48020](https://haworthiaupdates.org/wp-content/uploads/2019/07/H-retusa-3.jpg)
![Haworthia retusa (L.) Duval [as Aloe retusa L.]
J.S. von Salm-Reifferscheidt-Dyck [Salm-Dyck], Monographia generum Aloes et Mesembryanthemi, Aloe, vol. 1: , fig. 9.3 (1849-1863)
http://plantillustrations.org/illustration.php?id_illustration=184034](https://haworthiaupdates.org/wp-content/uploads/2019/07/H-retusa-4.jpg)
![Haworthia retusa (L.) Duval [as Aloe retusa L.]
Collection des vélins du Muséum national d’histoire naturelle, vol. 8: t. 63 () [C. Aubriet]
http://bibliotheques.mnhn.fr/medias/detailstatic.aspx?INSTANCE=exploitation&RSC_BASE=IFD&RSC_DOCID=MNHN_VEL_PORTEFEUILLE008_FOL063](https://haworthiaupdates.org/wp-content/uploads/2019/07/Aloe-retusa.jpg)