(Haworthiamaculata var. livida (Bayer) Bayer, comb.nov. H. pubescens var. livida Bayer in Haworthia Revisited, p.134, 1999, Umdaus) Type: Cape-3319 (Worcester): S Lemoenpoort (-CD), Bayer 1128 (NBG, Holo.).
I described Haworthiapubescens var. livida in Haworthia Revisited (Umdaus, 1999), in the full knowledge that it was in a twilight zone of inadequate information. It is a good example of how Latin names give plants a false reality. The system forces decision making without any slack being cut for doubt. This is thus a good opportunity to demonstrate what inexperience and ignorance add to the process of classification. In the small area along the Breede River north of the Brandvlei Dam near Worcester, the species H. herbacea, H. maculata and H. pubescens grow in close proximity. H. herbacea is ubiquitous throughout the Worcester/Robertson Karoo, while H. maculata has a curious distribution in that area. It occurs at widely separated localities on the western fringe of H. herbacea and I have wondered about its relationship to that species because of the similar flowers and flowering time. H. pubescens is only known from a small set of low ridges east of the Brandvlei Dam where it grows in close proximity to H. herbacea. It also has similar flowers but it flowers a little later in late spring as opposed to early spring.
When I first discovered plants at Lemoenpoort (Hammansberg) about 20km further south I found very few plants and the only thing I could do with them was regard them as in the same context of the Breede River species and hence their description as a variant of that H. pubescens.H. herbacea is very abundant in this area too but almost exclusively in Bokkeveld Shales and Dwyka Tillite. The doubtful plants at Lemoenpoort occur in Witteberg Sandstones. A population of plants was also known from the top of the Moddergat Mountain (Ouhoekberg) to the west that created some problems for me as it seemed to relate to the absence of H. mirabilis from both the Hammansberg and Ouhoekberg. Not only that but it also looked very similar to H. maculata. H. mirabilis is known at its closest east of these two mountains at several localities along the eastern ridges of the Ouhangsberg and Droogerivierberg mountains that are in turn east of Hammansberg. H. maculata is very abundant along the long ridge of low hills north of the Brandvlei Dam. Very similar plants occur high in the Audensberg peaks north of Worcester and also lower down in the Kanetvlei (Sandhills) area west of Heatlie Peak. These hills are very difficult to explore and I speculate a connection to a population at Buitenstekloof to the east. There is a population here that obfuscates the interpretation of the name H. intermedia VPoelln. I lumped this population under H. mirabilis var. notabilis. I have to rely on memory here because having set flower morphology and flowering time aside (without completely ignoring them), I did not have the methodology in place to record detail. The flower was rather similar to H. herbacea and H. maculata and it also flowered late spring. I was not, and am not, sure that there is not a connection between the Buitenstekloof plants and the Kanetvlei plants.
It is unfortunate that Gerhard Marx has expressed his opinion that I have ignored flowers in my classification of Haworthia. While there is no doubt that this is partially true, there are very good reasons why I did this. The main reason is the historic one. It was understood and accepted that the flowers in the species as they were then recognized, were very similar. Even the subgeneric differences were not seen to be significant. My own early attempts at classification also indicated that the flowers were not going to be of assistance at the level at which differences between populations and species were being recognized. In fact I had observed similarities of the flower in populations that were clearly different taxonomic entities. Conversely I observed big floral differences in populations that I imputed to be the same “species”. Because of the added seasonality of flowers, I thus put them aside to focus on exploration and explanation on the basis of the vegetative structures and geographic distribution. Marx’ observation therefore comes at a time when that work is substantial and flowers can now be used to clarify and verify relationships. My conviction is that those relationships are best understood in relation to distribution and we are only now in a position to assess what value the flowers may add to those interpretations.
This article thus reports on the exploration of the Ouhangsberg, Hammansberg and Moddergat (Ouhoekberg) mountains as well as some work done in the Brandvlei Dam area. Again I put flower and flowering time aside – for the moment. This is in the same way I am inclined to the considered view that geographic information and field observation are the essential ingredients whereby ANY method has to provide explanation that satisfies physical experience. The alternative is that what we see and experience physically and mentally, is just the illusion that metaphysicists ascribe to creation.
Populations recorded
The following populations (see map) were found and observed:-
1. KG669/69 N Brandvlei Dam – set 1 2. MBB164 N Brandvlei Dam – set 1 3. MBB1119 (and 6815) Audensberg – set 2 4. MBB1120 (and 7994) Sandhills, Kanetvlei – set 2 5. MBB1128 (and 7066) W Lemoenpoort (type locality var. livida) – set 3 6. MBB1145 Ouhoekberg, Moddergat – set 5 7. MBB2591 NE Brandvlei Dam – set 1 8. MBB4461 (and 6514) Buitenstekloof – set 6 9. MBB7270 Ouhoekberg W – set 5 10. MBB7266 E Lemoenpoort – set 4 11. MBB7271 Cilmor Winery, Dewetsberg – set 7 12. MBB7526 Die Nekkies W. Brandvlei Dam – set 1 13. MBB7991a Ouhoekberg E – set 5 14. MBB7991b west Ouhoekberg E – set 5 15. MBB7992 Hammansberg midpoint – set 8 16. MBB7993 Hammansberg W – set 8 17. MBB7997 Sandberg H. pubescens, type locality – set 9 are for MBB7997 cf H. pubescens N of point 11 18. MBB7995 and 7996 Brickfield H. herbacea ‘submaculata’ – set 10 19. MBB7984, 7985 and 7988 Droogerivierberg, H. mirabilis – set 11 20. Unknowns – set 12
Map references for Haworthia maculata.
There are 4 records for the Brandvlei Dam but in Update 6, I report on H. maculata from the pump station east of the Resort, very abundant to the eastern point of Die Nekkies. Three of the records are for west of the Resort while I have not confirmed Etwin Aslander’s observation that he had seen it at the extreme western end of Die Nekkies. G.J. Payne in 1970 communicated to me that he had seen a Haworthia south of the Brandvlei Dam above and west of the Brandvlei Hot Spring. I was unable to confirm that in a single brief visit there ca. 1976. I have not been able to relocate plants at Sandhills but have no doubt that it will be found there if more thorough and extensive fieldwork is done there. In fact a very important consideration is that there is still enormous scope for exploration.
On each visit to the Ouhoekberg I have found plants in different local populations and none of the last four records is the same as seen in my first visit. At the original Lemoenpoort I always struggled to find plants at all, and the same applies to the locality to the southeast. At my last visits to these two places there have been many plants but relatively difficult to find.
H. herbacea is abundant in the Hammansberg and Ouhangsberg area as well as east of Cilmore. It has not been observed near the Ouhoekberg. It occurs between Cilmor and Die Nekkies in a larger form that acquired the name H. submaculata V.Poelln.; and I did suspect that there was a continuity with H. maculata that does not now seem to be the case. I exclude H. pubescens from this report. It occurs with H. herbacea east of Cilmor, but it also occurs on the northeastern slope of Dewetsberg north of the H. maculata recorded habitat. There are/were plants of H. mirabilis vegetatively very similar to H. pubescens at the Breede River Bridge southwest of Roberson. Their flowers and flowering times (March) correspond with H. mirabilis in that area. H. pubescens flowers in November (later than H. maculata at Die Nekkies or of H. herbacea – Sept/Oct) and the flower is similar to that of H. herbacea. It seems fairly obvious that H. pubescens fills the gap for H. mirabilis and the flower and flowering time are problematic.
The plants illustrated:
Set 1. Map points 1, 2, 7, 12. Die Nekkies Figs. 1.1 to 1.5 7526 H. maculata. Die Nekkies. Figs 1.6 to 1.10, sn* H. maculata, Die Nekkies – East.
A chapter in Update 6 deals with H.maculata – Die Nekkies Biomes, and Haworthia maculata These few images give scant insight into the variability in respect of rosette and leaf form, colour and marking, and proliferation.
Set 2. Map points 3, 4. Audensberg and Sandhills Figs. 2.1 to 2.5 MBB6185 H. maculata. Audensberg. Figs. 2.6 to 2.36 H. maculata, Vreesniet, Kanetvlei.
Unfortunately I found non-digital photography to be costly, cumbersome and unmanageable. So I have no field images for either population on the high Audensberg or at my original Sandhills site. However, I revisited the area and found the plants quite abundant a little further north at Vreesniet that is also Kanetvlei/Sandhills. The plants were almost confined to narrow rock cracks and their presence suggests to me that there is a very probable connection now eastwards to the Unknowns and even further on to the Osplaas H. arachnoidea, Etwin Aslander’s Hex Pass plants, and then still further to the H. marumana ‘dimorpha’ questionable.
Figs. 2.1 to 2.5 MBB6815 H. maculata. Audensberg.
Figs. 2.6 to 2.36 H. maculata, Vreesniet, Kanetvlei.
Set 3. Map point 5. Figs. 3.1 to 3.34 MBB7066 H. maculata, Lemoenpoort.
This is the type locality (i.e. MBB1128, W. Lemoenpoort) for H. maculata var. livida that I think is largely untenable or unnecessary fragmentation. Lemoenpoort is the valley that separates Hammansberg from Ouhangsberg. The first few plants seen were in exposed situations and had the purplish or bluish-grey colour that prompted the Latin name. This was maintained in cultivation. I linked it at the time to H. pubescens that seemed more probable at the time than to H. maculata and then simply because of the perceived demands of a relatively inflexible nomenclatural system. There are now seven new populations that contribute and improve understanding of H. maculata as a species. All these localities are in Witteberg Sandstone and the type locality is only slightly different in that the stratum of rock is less feldspathic (i.e. mineralized).
Set 4. Map point 10. SE Lemoenpoort Figs. 4.1 to 4.28 MBB7266 H. maculata SE Lemoenpoort.
This is a more west facing slope and also relatively low down on the mountain side. The plants were in rocks bordering an eroded softer shale band. They probably also occur higher on the slope in less mineralized sandstone. Generally the plants were very cryptic and tended to be solitary.
MBB7266 SE Lemoenpoort, view to southwestMBB7266 SE Lemoenpoort, looking up at locality.
Set 5. Map points 6, 9, 13, and 14. Ouhoekberg. Figs. 5.1 to 5.24 MBB7991a H. maculata, Ouhoekberg E. Figs. 5.25 to 5.28 Panoramic views. Figs. 5.29 to 5.33 MBB7991b H. maculata, Ouhoekberg.
These populations were first observed as one on a higher eastern point of the Ouhoekberg above Moddergat in about 1975. George Lombard accompanied Kobus Venter and me there in 1996 and we found them on the western high point. I located them nearby in 2004. We found them again recently as two more populations on the eastern heights near where I must have observed them first. Note the reticulation in the dried fruit capsule. It can be very much more evident in species like H. pulchella but I seriously doubt its diagnostic value. I have included views to… (a) north to show the water of the Brandvlei Dam in the far distance. The area between has not been adequately explored. What is interesting is the geology. The mountains on the left are the Table Mountain Sandstones, nearer is the valley where the soft Bokkeveld Shale has been eroded away, and then comes the Witteberg Sandstone with a neck of soft shale, and then low down is Dwyka tillite. H. herbacea is present on the Dwyka outcrop barely visible in the middle right. (b) The second view is looking east at first the Hammansberg and beyond that Ouhangsberg with H. mirabilis on its eastern flanks. The view looking southeast is over low Bokkeveld shale ridges with an abundance of H. herbacea. The view south is to Villiersdorp where Wolfkloof (not the Robertson Wolfkloof) is a deep valley behind the Table Mountain Sandstone left of the gap through the Rooihoogte Pass. Here is where H. herbacea ‘lupula’ occurs, unusually in sandstone. Altitude and skeletal soils of different origins contribute hugely to the genetic mosaic. Arable depositional soils exclude Haworthia and obviously inhibit contact between populations.
Ouhoekberg view to north.Ouhoekberg view to east.Ouhoekberg view to southeast.Ouhoekberg view to south.
Set 6. Map point 8. MBB4461 and 6514 H. mirabilis ‘notabilis’. Buitenstekloof
I am sorry that I have probably contributed to the confusion here. It started with von Poellnitz with a species he described as H. intermedia from this locality where both H. mirabilis ‘notabilis’ and H. reticulata occur, but he included E. Cape H. cooperi in the synonymy so it is not quite clear what he meant. While H. reticulata can be easily recognized for what it is at Buitenstekloof, the other population I ascribe to H. mirabilis ‘notabilis’. But I am not certain because I recall it flowering in November and with a flower type nearer to H. maculata than to H. mirabilis. It appears to me that further exploration may expose continuity westwards to the Sandhills/Audensberg populations of H. maculata and even northwards to unknowns.
H. mirabilis ‘notabilis’ (with or without the inverted commas) occurs from Klassvoogds between Ashton and Robertson, westwards through Wolfkloof, Robertson to Agtervink and further westwards still to Buitenstekloof. This all along the great Worcester fault line and intrusive igneous rock. The dolomite is apparently difficult to explain geologically and is associated with the granite-like extrusions in the area.
Habitat at BuitenstekloofView further into Buitenstekloof
Set 7. Map point 11. Cilmor. Figs. 7.1 to 7.3 MBB7271
An interesting point here that H. herbacea (map point 18) occurs between this point and all the Die Nekkies populations. I am not sure that the area between can be fully explored although I have been into it.
Set 8. Map points 15 and 16. Hammansberg. Figs. 8.1 to 8.21 MBB7992 H. maculata, Hammansberg.
The first locality 7992 midway between the eastern and western ends of the mountain was very exposed and very sparsely vegetated. The sandstone was very mineralised and fractured. The second locality to the west was well vegetated. In the view northward, the low hill in the near middle is the Draaivlei mountain that is Dwyka Tillite and hosts an abundance of H. herbacea as does the Dwyka strata on the northern side of the Hammansberg. The south side is Witteberg Sandstone with Bokkeveld shale on the lower south-facing slope. The view from the third locality southeastwards, shows massive valleys running into the Table Mountain Sandstone of the Riviersonderend mountains on the right. If one takes in mind that this is a short way east of the Wolfkloof Valley where H. herbacea ‘lupula’ occurs, then this area begs exploration. I do have a recall of H. herbacea in low altitude sandstone either near there or at a similar site at MacGregor to the east. The interesting view is the one looking eastwards that really gives the scale of exploration needed. The furthest mountains are the Langeberg Table Mountain Sandstsones and Buitenstekloof is nested in foothills there and in intrusive igneous rock. The coming closer is the Rooiberg, then Gemsbokberg, then Gannaberg, then Ouhangsberg and Hammansberg itself. Rooiberg is fairly well explored although the significance of H. herabcea ‘flaccida’ is not explained. I doubt if a anything new will crop up on the Hammansberg. But the intervening three definitely hold key populations marginal to these covered in this posting. The Langeberg? I only have the set 12 unknowns and cannot see myself resolving those.
view from Hammansberg eastwardsview from Hammansberg northwardsview Hammnsberg to the southeast
Set 9. Map point 17. H. pubescens. Sandberg. Figs. 9.1 to 9.20 MBB7997 H. pubescens, Dewetsberg.
I long ago observed H. pubescens southwest of Sandberg and south of the Breede River on the Dewetsberg. This is not much further than 500m north of a population of H. maculata at Cilmor Cellar. Visiting the site again I observe that the plants are slightly different to the Sandberg plants. While they are equally small, cryptic and dark coloured, they have less surface spinuliferation that gives rise to the species name. Also some plants are distinctly spotted in the way that H. maculata are and these spots may give way to translucens when the plants grow in less light. See the images showing leaf surface detail of H. pubescens compared to the two collections of the Brickfields plants (images 21-27) of H. herbacea in Set 10, map point 18.
Set 10. Map point 18. H. herbacea ‘submaculata’. Brickfield, Brandvlei Dam. Figs. 10.1 to 10.20 MBB7995 H. herbacea, S Brandvlei Brickfield. Figs. 10.21 to 10.53 MBB7996 H. herbacea, E Brandvlei Brickfield.
I have associated this locality with von Poellnitz H. submaculata and treated it as a synonym of H. herbacea. However, here I first illustrate a population about 600m south of that where the plants are in the usual size range for the species ie.30-40mm diam. At the locality east of the Brickfield and next to the Breede River, the plants are 1/3 to 1/2 as large again and can form huge clumps. North-west from this is a population of H. maculata at the extreme end of Die Nekkies and only about 300m distant, and this population I have always regarded as somewhat intermediate. What is interesting to note is the huge variation in leaf shape and armature within each population. In both cases the plants are in Witteberg Sandstones and at the first site this is both in a very shale-like stratum as well as in a highly quartzitic one. This is unusual for H. herbacea. Both populations wedge in geographically between H. maculata populations and in no known case do both occur.
7995 Habitat looking northwards
Set 11. Map point 19. H. mirabilis. Droogerivierberg. Figs. 11.1 to 11.11 MBB7984 H. mriabilis, Droogerivierberg. Figs. 11.12 to 11.24 MBB7985 H. mirabilis, Sandberg S. Figs. 11.25 to 11.31 MBB7988 H. mirabilis, Trappieskraalkloof.
In this area H. herbacea is common in the Dwyka Tillite but not in the Witteberg Sandstone. But instead of H. maculata, H. mirabilis is present there and commonly so. Where at one time I considered that H. pubescens was very probably an extension of H. mirabilis to a western and northern limit, this does not seem to be the case as it is so closely linked to H. maculata and H. herbacea. On the other hand I have wondered about the similarity of the Moddergat H. maculata to H. mirabilis. H. mirabilis is in the eastern areas of the Ouhangsberg and now we know that H. maculata occurs not far away on the western slopes. The centre area remains unexplored and this needs to be done as it may also help understand the idea of two principle role players viz. H. retusa and H. mirabilis as precursors of a very complex Southern Cape assemblage.
Set 12. Map points 20. Unknowns. Figs. 12.1 to 12.7 MBB7865 H. cf. arachnoidea. Keurkloof, SE Dedoorns. Figs. 12.9 to 12.15 EA1441 unknown, Hex Pass.
I have only two sets of pictures for this set. These are…(a) MBB7865 that Ernst van Jaarsveld drew to my attention at Keurkloof, south east of De Doorns in the Hex River Valley. There is a continuum of suitable habitat between there and Sandhills/Kanetvlei; (b) EA1441 and this number may be incorrect, but it is at the near-base of the Hex River Pass still further east than Keurkloof. In between is Osplaas Station where there are dramatic forms of H. arachnoidea with very spotted leaves, not to forget the nortieri-like form of H. arachnoidea at Kanetvlei about 200m north of MBB7994 H. maculata.
H. cf. arachnoidea. Keurkloof, SE Dedooorns 0087865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 0067865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 005
Conclusion The nature of the populations of H. maculata on Die Nekkies varies quite considerably in that there is an area east of the Resort where the plants form huge clumps. Individual rosettes can be quite large. But there are also points where the plants tend to be solitary, hidden in rock cracks or even truncated into the soil. The plants in the Southern populations are smaller and tend to be solitary. There may be differences in flowers and flowering time but this will really be significant in relation to H. mirabilis rather than between the populations recorded here or to H. herbacea. Attention must be paid to the way in which H. herbacea is geographically wedged in between populations of H. maculata. Equally significant is the character of H. pubescens southwest of Sandberg (Dewetsberg) where the plants have less spinuliferous leaf surfaces and also spotting (maculation). This is one of the best pieces of evidence I have for the geographic continuities that exist throughout the genus both in respect of species of how I think species can be recognized and of how difficult decision making is. In essence this report only dismisses the idea that the Lemoenpoort population can be assigned to H. pubescens. I doubt if it is rational to even formally recognize it as different to H. maculata as “variety” itself is a mythical status. Fragmentation by names is a rather archaic approach to classification that serves hardly anything but a semi-commercial need.
Acknowledgement Many people contributed in one way or another to this report … Mr Hentie deWet of Moddergat, Messrs Poffie and Hettie Conradie of Sandberg (South), Mr Pieter Naude of Vreesnicht, Mr Johan and Marie Fourie of Buitenstekloof, Mr Nico Marais of Worcester (Brandvlei) Brickfield, Mr A. Groenewald of Cilmor Cellar and Stefan Hugo of River Farm. Messrs PD and Anso leRoux provided much logistic support, interest and company as did Kobus Venter, Etwin Aslander and Werner Voigt. Lawrence Loucka has managed and facilitated the archiving of publications and pictures.
There seems to be so much harping about my departure from the International Code of Botanical Nomenclature (ICBN) that I obviously need to try and explain myself better. The real issue is that we are dealing with a group of plants that is largely appreciated for its vegetative characters and not for its small and unexceptional flowers. Because the plants are small and so commonly grown by collectors the numbers of plants in cultivation and close observation is large. The plants also do vary in respect of leaf morphology, arrangement, and surfaces to a greater extent than in many other genera. Furthermore, the variation is also exaggerated by growing conditions. The fact that flowers are not used in the classification process beyond the level of sub-genera means that there is an almost total reliance on vegetative characters for classification. The nomenclatural system in botany tends to be a typological one, which means that reliance is placed on descriptions very often derived from single specimens. This is particularly so when the nomenclatural types are simply old illustrations that have been used to arrive at identifications and names by consecutive authors for decades. Thus use of those identifications and names, and their continuing re-interpretation causes a great deal of either grief or great personal satisfaction depending on just who is being affected by the process. The fact that the names should indicate “species” is lost from sight and totally obscured by the additional absence of any good universally accepted explanation for what a species is or might be.
Explanation in and around all this has been a large part of my writing since I started this in 1962. Thus I will not enlarge on the subject but rather try to explain again this way using a species name generated by Col. C.L. Scott with whom I had some considerable altercation. It must be understood that Col. Scott was not a biologist and it is just a simple fact that the problems that the above brief remarks expose, trouble many professional taxonomists to this day. I do not condemn Col. Scott and express my respect and admiration for him and am very grateful to him for the friendship he later extended to me before he died.
The example I will take is that of his species Haworthia geraldii. It comes from a small hillside east of Riversdale running south to north, named Komserante. This is an Afrikaans term meaning the ridge around a small geographic basin. In the way that haworthias are assembled in small habitat defined and localised populations, there are three recognizable populations of plants (of the subgenus Haworthia) along this one ridge about 1km long. The southern population used to be complemented by still another that occurred where the stream bed left the basin at the southern end. A plant from that southern population was named as H. foucheii by K. von Poellnitz. The northern population is a bit problematic and I initially included it in my then concept of H. magnifica. Since then however, it has become clear to me that my idea of species was too conservative and that H. magnifica as described by von Poellnitz from south east of Riversdale (now the Frehse Reserve) is part and parcel of a huge complex that I regard as H. mirabilis. Thus this northern population is seen by me as H. mirabilis but complicated by the fact that it is largely influenced by hybridization with the next population or populations south. A plant (or perhaps, and doubtfully, plants) have been given the name H. vernalis by the Japanese writer M Hayashi. The name H. geraldii is attached to the second population southwards. The plants are very proliferous, form large clumps and the leaves are usually quite strongly retused (“bent back like a thumb”). The name H. foucheii is simply attached to the third because the plants there tend to be solitary and the leaves are also fairly erect and spreading as described and pictured for the original from the fourth population that was off the ridge.
The problem is now that we have descriptions and plants in each population that do not match the descriptions. While I assign the northern population and all the plants in it to H. mirabilis, other writers use the names H. vernalis, H. magnifica and even the confused name H. asperula. What they mean is by no means clear and in actuality a lot more names and descriptions would then be required to name each of the countless variants there for what might be an original pure species and variants or for first second or third generation hybrids with H. geraldii or H. foucheii as those names were applied.
But the problem is far more extensive than just these four populations. After years and years of field exploration it has become evident to me that like H. mirabilis, H. retusa is also highly diverse and I see it to include all the variants of H. turgida. In fact I suggested a long time ago that H. retusa is simply the solitary large form of H. turgida in low-lying level areas as opposed to the more common and extensive clumping cliff dwelling forms of that species. This is also why I object to the requirements of the ICBN that require the name H. retusa to take precedence for the species for historical reasons when it would be far more realistic to take it as a variant of H. turgida for biological reasons.
It has always been simply evident to me that H. geraldii and H. foucheii are variants of H. retusa. The problem now is that the plants in the two populations vary so much that, while the names are indeed useful for commercial and collector use, there can be problems that the use and application of the names will be confused.
Look at it like this. In respect of H. geraldii; it came from one population at Komserante, Riversdale and not all the plants are the same. I think this one population and all those plants in it belong to the species H. retusa, so I called it H. retusa var. geraldii. (var. = variety). Because not all the plants look the same that means that only some are truly geraldii and only the plants that meet Scott’s description are actually var. geraldii. Because the plants multiply vegetatively and Scott only took a piece, the original plant may still be there. This means that we should recognize it as H. retusa var. geraldii forma geraldii and in cultivation as H. retusa ‘Geraldii’ or just as H. ‘Geraldii’. What do we do with the other 100 plants or more in the population that are not H. retusa ‘Geraldii’? I do not think they can all get names so I take away the capital letter, leave out “var.” and use ‘and’ (inverted commas) to show that the identification is, and will, be a bit uncertain unless you know from a label that the plant comes from that particular population at Komserante. The same applies to H. retusa var. foucheii. In this case the original population is gone and I am not sure if H. ‘Foucheii’ is still in cultivation. So I use the name H. retusa ‘foucheii’ for a second population on Komserante. Some of the plants in this population look like some of those in the H. retusa‘geraldii’ population but not quite like H. ‘Geraldii’ itself. So for me the answer is MBB7780 H. retusa ‘geraldii’, Komserante and MBB7781 H retusa ‘foucheii’, Komserante.
It can be seen that a complication comes in when more than one population is involved. The problem then is that you can put them in a line so that some plants in the first population look like plants in the second, some in the second look like plants in the third, some of those look like those in the fourth until at the end none of the plants in the last population look like any in the first. The trouble in the field is that the line is not straight and it can also go off in different directions. Some of those directions may end up where they started. One simply cannot be truly confident about many of those names that are given to plants. Serious and proper consideration must be given for how different they may be even within the populations. Not to say of the shared similarities and still added variation from geographically adjacent populations. It may happen that I may write H. ‘retusa’ (and add number and place name) for a population that may better (not necessarily correctly) be identified as H. retusaXmirabilis. Other writers who are not biologists and have other considerations in mind may want to give a new name altogether. That new name and description may be again really only for one plant, or maybe a few more in a population, that have real or imagined novelty value and attraction. But they take no account of all of the less attractive variants or other populations that comprise the biological whole by close proximity if for nothing else.
The above discussion is integral to the rationalized list of names that Dr John Manning helped me produce. There are some species names there that I have deliberately presented as synonyms or variants of the species I recognize, because the authors of those names fail to convince me that they have any understanding of the situation at all. The word “species” is apparently simply a convenient naming system for oddities and novelties and not any scientific construct to explain the natural phenomena of living systems and their parts.
To summarize; I have written a report on flower characters in which I refer to the Komserante populations and I have used the following names and my own convention as follows:-
Please note that the way I have omitted the word variety from the following names and also used inverted commas …
7779 H. mirabilis, Komserante (the northern population – I could add ‘vernalis’ but it carries all the baggage of doubt about actual status). 7780 H. retusa ‘geraldii’, Komserante. 7781 H. retusa ‘foucheii’, Komserante. 7920 H. retusa ‘nigra’, Van Reenens Crest.
The omission of the word variety is for two reasons…
1. Economy
2. To convey the idea that the actual indication of status is not certain as I have used the name to indicate a population or populations rather than a single described plant. The prime and overriding uncertainty is that we cannot know what a species is that I have described for Haworthia. Thus how can we possibly rank populations at levels below?
The use of inverted commas reinforces what I want to convey. This is that the individual plants in the populations are variable and it may not be easy to always identify the plants (individual or population) according to a more formal classification.
Any departure from the ICBN or the way the names are treated in formal botany conveys the difficulty that I personally find in trying to reconcile formal nomenclature with names that are so often tied to single plants.
Conclusions. The nature of the populations of H. maculata on Die Nekkies varies quite considerably in that there is an area east of the Resort where the plants form huge clumps. Individual rosettes can be quite large. But there are also points where the plants tend to be solitary, hidden in rock cracks or even truncated into the soil. The plants in the southern populations are smaller and tend to be solitary. There may be differences in flowers and flowering time but this will really be significant in relation to H. mirabilis rather than between the populations recorded here or to H. herbacea. Attention must be paid to the way in which H. herbacea is geographically wedged in between populations of H. maculata. Equally significant is the character of H. pubescens southwest of Sandberg (Dewtsberg) where the plants have less spinuliferous leaf surfaces and also spotting (maculation). This is one of the best pieces of evidence I have for the geographic continuities that exist throughout the genus both in respect of species of how I think species can be recognized and of how difficult decision making is. In essence this report only dismisses the idea that the Lemoenpoort population can be assigned to H. pubescens. I doubt if it is rational to even formally recognize it as different from H. maculata as “variety” itself is a mythical status. Fragmentation by names is a rather archaic approach to classification that serves hardly anything but a semi-commercial need.
Acknowledgement. Many people contributed in one way or another to this report … Mr. Hentie deWet of Moddergat, Messrs. Poffie and Hettie Conradie of Sandberg (South), Mr. Pieter Naude of Vreesnicht, Mr. Johan and Marie Fourie of Buitenstekloof, Mr. Nico Marais of Worcester (Brandvlei) Brickfield, Mr. A. Groenewald of Cilmor Cellar and Stefan Hugo of River Farm. Messrs. PD and Anso leRoux provided much logistic support, interest and company as did Kobus Venter, Etwin Aslander and Werner Voigt. Lawrence Loucka has managed and facilitated the archiving of publications and pictures.
MBB1128 (MBB7066 W. Lemoenpoort) is the type locality for H. maculata var. livida. Plant pics are available in Haworthia Update Vol. 9. A myth corrected to, population 5, set 3. Here are flower photos from 8 December 2012.
MBB4461 (and (MBB6514) H. mirabilis ‘notabilis’ Buitenstekloof
In Haworthia Update Vol. 9 and a recent posting on Haworthia nortieri Bruce Bayer makes reference to this Haworthia maculata population, MBB7994 from Kanetvlei.
MBB7991 Moddergat above Ouhoekberg E, ex hort. single clone
From Updates Volume 9 Set 5 Bayer writes, “These populations were first observed as one on a higher eastern point of the Ouhoekberg above Moddergat in about 1975. George Lombard accompanied Kobus Venter and me there in 1996 and we found them on the western high point.”
These photos were taken 20 December 2012 from a single ex hort. clone and hence consistency, but still illustrative.
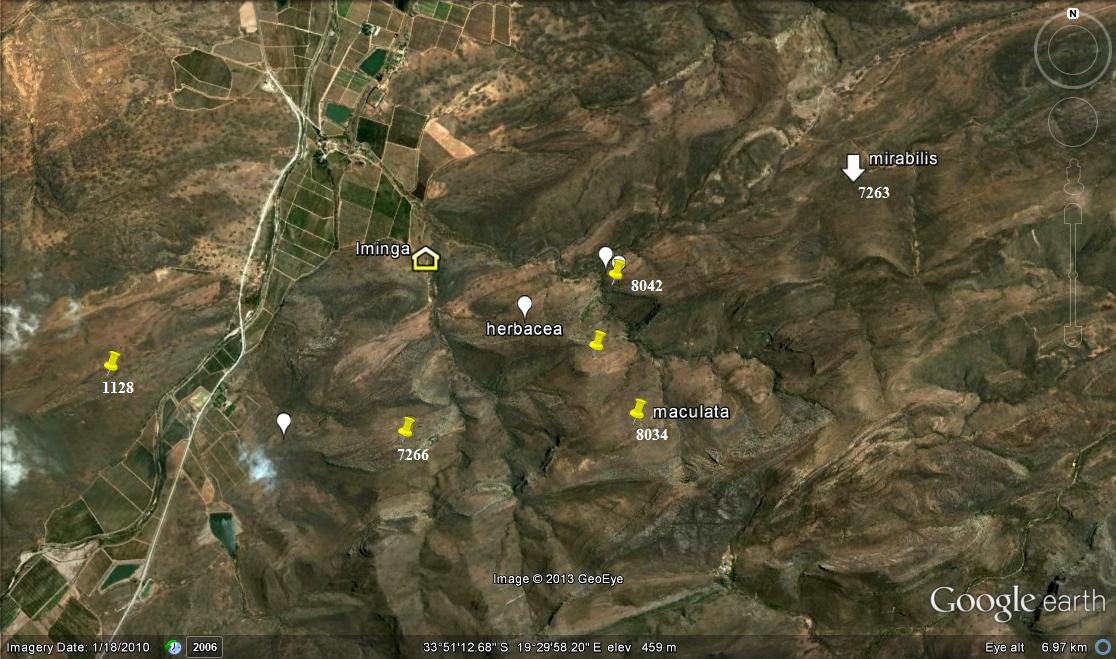
8034 Haworthia maculata ‘livida’, Ouhangsberg.
These plants occur in Pteronia communities on very dry and sparsely vegetated north slopes. The geology is Witteberg Sandstone but the particular strata the plants prefer seems to be a softer more erodible and less quartzitic stratum than they occupy at Lemoenpoort to the west. The plants appear to have flowered early in December coinciding with the flowering at Lemoenpoort and southeast Lemoenpoort. The two localities where we observed the plants are approximately midway between H. mirabils at Droogerivierberg to the east and Lemoenpoort to the west. There is no suggestion that they form any continuum and the flowering times are different.
The name of the farm is Iminga Mountain Reserve, a large tract of quite non-arable countryside that is appreciated by the owner as her garden.
Acknowledgement – I deeply appreciate the freedom to explore Ouhangsberg granted by Ms Suzi Broomberg.
Following on from the 8034 record, we returned to explore still further east in a direct line from the 8034 localities to the H. mirabilis population on Droogerivierberg (see map). We found plants at the same altitude as we had observed H. herbacea on the previous visit. The plants were again very cryptic and hard to find. Flowering was also long past and the few capsules present contained very little seed. The situation with respect to habitat was a little different and these plants were just off a truly sandstone stratum and in a narrow band of renosterveld. H. herbacea was within 50m of the nearest H. maculata in a more karoid vegetation similar to that where we had seen H. maculata on the previous visit. This is difficult to understand. Southeast of the Brandvlei Dam, H. herbacea occupies habitat where one would expect to find H. maculata. But it does not do so in the Ouhoekberg nor Hammansberg. Here north of the Ouhangsberg there does not seem to be a visible difference in the choice of habitats for the two species but they cannot be said to truly co-occur i.e. grow together in such close proximity that they intermingle.
The spatial relationships of the various populations are now quite significant. The spread from localities east to west on the map is 5.23km. The distance between known H. mirabilis and nearest known H. maculata is nearly 1.5km. There is no indication of transition and present evidence is that there is no continuity between these two species. There is however still that space between to explore as well as a massive valley south between the two populations in question.
Fig. 1 Image of Iminga Mountain Reserve.Fig. 2 Westwards towards homestead.Fig. 3 Below locality at fig 2, southward.























































































































































































































































































































































































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}