These are a bit west of the Slangriver and taken in July during the wet season. So the plants are green, turgid and about as much translucency as they can show. It occurs to me now that I always thought of words rather than pictures, but in this digital age this is about how many pictures needed and of better quality.
Essie Esterhuizen commented “It was in this area where I realised in the early 90’s that I don’t have the ability to sensibly distinguish between the then maraisii, magnifica, mirabilis and heidelbergensis.”
To which Bruce Bayer replied. “Well Essie, its not a question of your ability. It simply cannot be done. Those names just existed because of the history of the subject and even now there is surely some doubt that you and I are stating the fact that a true classification does not meet the perceptions or needs of enthusiasts and collectors. Their need are real and it is time amateur journals started assuming some responsibility for a way of using names that is useful, while being accurate.”
This is also west of the Slangriver and not just making a mockery of any attempt to subdivide H. mirabilis into heidelbergensis, atrofusca, jakubii, magnifica, maraisii, shuldteniana and many more, you can clearly see that floribunda is involved too. I do not know of any floribunda southwest of Heidelberg except the evidence that appears in these small mirabiloids that this is where it has gone. Soumen Aditya is a good observer and it will be nice to know how his opinions of the names that are developing.
Soumen Aditya replies “Sir, true…one population and its species concepts no value at this moment. Horrible variations each single species and definitely link each others. Floribunda is a simple variation under many mirabiloid included maraisii. Thank you.”
This is a few km downstream on the Slangriver from toonensis and a great opportunity for someone to explore the prediction that more populations will exhibit the similarities that follow geographical order. There are some unique plants in this post and the Slangriver is no stranger to oddity. It is not easily accessible. It follows a strange path – first southward and then westwards to flow into the Breede River between Malgas and Infanta where there are many mirabilis and some turgida variants.
Some way to the northwest and the plants seem to have lost translucency while the leaves are shorter stouter and perhaps darker green. But there are MANY populations of this ilk varying in one direction or another, supporting the truth of Essie’s comment that categorizing them by any of the more relevant names is just not possible.
While considering what to post next on Mirabilis Travels I realized that I have to break mirabilis up and organize my pictures better. There are practically speaking 12 geographical zones and I only have pictures for about 200 populations of at least 400 I have visited that fall into the category I recognize as H. mirabilis. I cannot believe that anyone can dispute the statement that these populations and the internal variation form a continuum and any taxonomic subdivision will be purely artificial. In fact I go further and state that the species H. turgida, retusa, pygmaea, mutica and emelyae are more probably than not, included. When the involvement of H. floribunda with it’s associations, and then the probability that H. emelyae has further connections are considered as well, the enormity and complexity of the situation can be imagined. But here are pictures from two further populations to go with the previous SW Heidelberg.
Each of those mirabilis zones has something going on pointing to the complexity I refer to, but right now I am posting about the heidelbergensis mirage. This is the Swellendam zone and the population one from S Buffeljagsrivier. The home of groenewaldii, that is a variant that for me puts the whole retusa/mirabilis complexity into perspective.
This, further south from Buffeljagsriver. The plants were not in good shape and this is just one of the sad facts of life. It raises the issue of how to decide what it is. But the situation is that there is no clear idea of just what constitutes heidelbergensis. West of Buffeljagsrivier there are 6 known populations of mirabilis that I cannot state why exactly they are the same. There is an obvious similarity in respect of size, colour and flower but confoundment with the floribunda/variegata variant as well. Can it be understood and can it be explained? So lets rather continue southwards down the Breede River for the moment.
Further south along the Breede River is a “farm” where we found 8 populations of mirabilis and I will just show plants from 3 localities to show how they vary. See the floribunda influence too – floribunda as a recognizable population is absent south of the Bontebok Park but reappears as such down along the western end of the Potberg. But remember we are actually on a heidelbergensis mission to show that it is fiction just as magnifica and maraisii (and others) are.
Eureka – have I got it? For years I have been pondering the problem of how and why the form of rossouwii appears in mirabilis ’heidelbergensis’, mirabilis ’sublineata’, and emelyae ‘multifolia’. Essie has independently indicated that he observes this too. I should actually be travelling, in my series of posts about heidelbergensis, from west to east to show this. Sadly I do not have pictures of the heidelbergensis I saw further east of Heidelberg, nor any of Essie’s for the eastern Hessequas area. DNA does not create form. It only provides the materials to do so. Form depends on the creative power of the metaphysical element (causal mind – consciousness) of each life form? I will put a question mark there again, because this is a serious issue we need to contemplate. The photos I add are multifolia variants from 4 populations from Little Karoo. The last one touches on Warwick asking why I fingered a single variant from S of “heidelbergesis var. toonensis” along the Slangriver.
So we are still south of Buffeljags along the Breede River. Looking for heidelbergensis evidence. Not much in this lot but I did also interject with a post referring to rossouwii. Very interesting because now one has to consider all populations, related species AND their distribution. Because this is a whole and species cannot be summarily treated like cherries or plums and picked by whim and fancy. Haworthia rossouwii is very odd because it occurs at two centers about 30-40km apart and then appears to reoccur in a smaller form just south of Heidelberg.
Bit of a dilemma. Rather than go south I will for the moment go west from Swellendam towards Stormsvlei. The end will shock as it really demonstrates the impossibility of arriving at a solution on the mechanistic platform.
The predictability of a working hypothesis? Essie sent me this lovely picture of a plant from the mountains between Heidelberg and the Little Karoo. But like me Essie has only just awaken to the fact that you cannot get people to believe what you say unless you can give them a greater visual experience than you had yourself? Essie has a LOT to tell us. This is a link between the turgidoides (i.e. retusa and mirabilis together therein) and H. emelyae. It will eventually be recognized to also include rossouwii?
A little further west in the Luiperdsberg area west of Swellendam.
Now between Luiperdsberg and Bromberg on the way to Stormsvlei. There is not a smooth transition and a huge number of populations nevertheless supports the contention that this is a single system viz H. mirabilis.
I don’t have pictures of mirabilis on the south side of the Bromberg so these are from north. Also I am going to skip going still further north into the Worcerster Robertson Karoo and beyond the Riviersonderend River. I wrote all about mirabilis in Updates about 2006 and don’t have pictures for the complexity in that area and westwards, which would help to ease the oddity of transformation.
Tony Brook asked if mirabilis is the oldest name. Bruce answered “A very uncomfortable question. I have no doubt that it is atrovirens – but it is a tricky path and I am no fan of the nomenclatural code or the fancy footwork that goes with it. Goes back to Stearn and typification of old names.”
These Bromberg variants will surely excite the splitters but really when one sees so many of the populations from McGregor to Swellendam as well as to the north; and even the ecological semi-desert to the south, you have no option but to abandon an impossible point of view. Unfortunately the population of ‘heidelbergensis v. scabra’ (i.e. before my even conservative view was proved hopeless) at Leeurivier east of Drew, was ploughed out despite the non-arable nature of the shale. But there are pictures in Revisited and also in early volume of Updates re: H. mirabilis.
Authoritarianism is a real problem in present society and this comes up in plant classification too. So it is not compulsory to see these plants as H. mirabilis too. It is just my personal opinion. The plants on the north slopes of the Bromberg are in sandstone and these now are along the Riviersonderend River in manganic tertiary deposit. This is very unusual habitat. Taken late February as they flower in the heat of summer.
These were taken 3 years earlier and In January when cooler wetter conditions prevail. There are four microhabitats in a very small area.
Now this is a real freak for which I have no explanation. A single small population in a weird habitat on the south side of the Bromberg. Sandstone slabs and very wet. About 200m away is shale and also what one expect of H. mirabilis. But this flowers in November and has in-turning leaves that one finds in, say herbacea. Although there are individuals, as you can see, in which the leaves are erect-spreading. The flowers are nothing unusual except the buds are flattened top to bottom and the ends also turned down slightly like-in mirabiloids. So it throws everything out of the window or into disarray. It does not alone meet the requirements of a species and there is zero indication of what its real affinities may be. My deeper feelings are that it mirrors a dark side of (at least Haworthia) classification and that the real species is the entire “genus”. For the present I have speculated, as I have suggested in this series, that floribunda/variegata/chlorocantha are intimately connected to the turgidoids, and suspect that rossouwii is as well. I have treated this H. eliseae as a variant of rossouwii, as I have done with ‘calcarea’, ‘petrophila’ and my own ‘heidelbergensis var. minor”. Let me add something. Classification is not some thing confined to part of the group so that you can cherry-pick where and to what you want to apply a name. A classification has to be ALL inclusive and the specimens remaining in the incertae folders are only there because they are too poor to permit any decision.
The myth of heidelbergensis has dissipated to the west and north and morphed into others, so let us go back down the Breede River and see what mirabilis holds for us in that direction. There’s a gap that I haven’t explored, the 20km stretch of the Breede River as it twists and turns south of Swellendam. The next two populations are south of Malgas, east of the river.
Here are more from the east bank of the Breede River south of Malgas. Note that the populations on the west bank all along the northern lower reaches of the Potberg are incredibly complex with names like joleneae, bobii confounded with atrofusca, floribunda, variegata, turgida, and mutica.
As an after thought … did anyone see this? Some how or other these two forms of H. eliseae came about, and I suspect it may have something to do with Rupert Sheldrake’s suggestion of morphic fields. Is this an overlap of the Worcester/Robertson herbaceoids and the more southerly mirabioids? I settled for a rossouwii/mirabilis interaction that is not entirely convincing. These two pictures were taken at the same time and place in one small population
I am not sure how to deal with the west bank populations. Essentially I am talking about H. mirabilis and have been exploring the heidelbergensis phenomenon. On the lower west bank of the Breede River we have a long conjunction of shale and sandstone mixed with tertiary deposits. The vegetation is either fynbos or renosterveld and there are vast differences over short distances. H. turgida in its best form is present (along the river itself) and so is the floribunda/variegata element. The result is two contrasting versions of mirabiloids. I will post the 6 or so turgidoid influenced populations first (fynbos) and then the more dark mirabiloids (Renosterveld) after. Note that there are no absolutes. On the decaying shales where there is kaolinite, we could be back west of Riversdale. I will start with what deviously and/or distastefully, acquired the name ‘bobii’.
This is just a few km west of bobii and again a link in a chain that makes a mockery of populations as species. I don’t think the Potberg is well explored at all. While this is on the eastern and north facing flank, there are two other populations known on the southern slopes travelling westwards.
On the southern face of the Potberg a few km further west and well off the beaten track …
Again travelling west along the south side of the Potberg is a similar population of plants. Unfortunately I only have two pictures of plants in cultivation that are not very useful except to show how growing conditions affect appearances. We arrived at site late afternoon in rainy weather. It was a long hike through thick brush and the light was also too poor for film photography.
Leaving the Potberg, returning to bobii at Infanta and then searching along the lower slopes between the Potberg and the Breede River. Just in what respect and by what margin do the next three populations differ from the previous 4? This was March 2006.
It is a bit of overdose because these are pictures of the same population as previously but taken a year later in September.
by J.M. “Essie” Esterhuizen and M. Bruce Bayer. While Bruce Bayer is the writer of this piece is by convention prime author, Essie Esterhuizen is cited as such for the following reason; it honours an astonishing amount of field work and profound insights that many taxonomic botanists never acquire. It is instigated by Essie’s more recent exploration and comments he has made that reflect the writer’s own conclusions. Thus this article is intended simply to mirror the thoughts of open minds looking for answers in a domain dominated by academics.
Introduction and general preamble The two of us have been long interested in nature and particularly Haworthia. We have a combined experience of at least 120 years of exploring the subject that we would like to share. Neither of us makes any claims to some or other authority or qualification that gives us more authority than anyone else. We have simply had an enormous privilege to have spent so much time in the field learning about these fascinating plants. Just like anyone interested in anything, we have needed a vocabulary and words or names to understand, organize and communicate our thoughts and words. This is either as internal private mental processes or as external communication with other people. The most important words we need are designations and names for the very objects of our interest. Perhaps for our own purposes we do not need words at all, but for external communication we certainly do. Then it becomes problematic because agreement, definitions and all kinds of conventions and guidelines exist that may not correlate with our individual capabilities, character, belief or value systems. In fact all this may clash with what our deepest individual understanding tells us.
The facts are that we have an organized system of information gathering and exploration we term ‘science’. Within that we have various disciplines of which botany exists for the plant kingdom. Within that we have a sub-discipline (taxonomy) of description, identification and classification. To guide this process international rules of nomenclature are agreed upon to provide a powerful framework of names that Essie and I now try to evaluate as a framework for our observations in that group of plants we know as Haworthia. Already there is a problem because Haworthia have been known in history as Aloe at the birth of the Linnaean system before being separated as Haworthia. At present there is one faction that maintains this arrangement and another that expounds Haworthiasenso stricto, Haworthiopsis, and Tulista as a formalization of three distinct sub-groups (subgenera) that both of us have no problems with.
This all seems very good and well until one faces the harsh reality that science as a discipline is concerned with physical creation. Rather than serving the purpose of information gathering across every single aspect of sentient life, it is in fact restricted to what has been described as mechanistic materialism. This is where explanations are sought for anything and everything by processes of experimentation, measurement, analysis, reason and logic. Plant taxonomy, at its base, rests on the principle that there are basic life forms (species) as in the animal world and that there are basic discrete life forms kept apart by virtue of their closed reproductive systems. This is derived or adapted from zoology and tacitly transferred to botany when there is enough evidence to demonstrate that fertility can extend across generic barriers. There is in fact no species definition. There is also no recognition of metaphysical realities or even the possibilities of their existence. Curiously a formidable intellectual who has edited and largely authored a massive six volume encyclopedia of mystic terms by John Davidson M.A. (Cantab) concluded that the definition of the species concept as a basic life form was not the same for science and mysticism. This is patently absurd.
Endless debate can ensue, but Essie and I, in our quest to understand Haworthia, have simply wanted to know what these basic forms are in practical terms so that we can organize and communicate our interests and thoughts with each other or with anyone else that may have common interest. In our quest we have simply used the hundreds and hundreds of names that have been coined for Haworthia species to come to know what these tell us and how useful they are to us in understanding them. Rationally each name is based on the tacit perception that it applies to a group of plants that are, or potentially are, interbreeding. This is blatantly untrue and no good definition actually exists to our knowledge that in any way regulates what is formally described as a species. An example could be cited where a single plant specimen has been selected to represent a hypothetical, fictitious species. No name in mentioned to avoid hurt. So we (the authors) have labored under this handicap where no one knows what a name even means, in trying to use and apply the many names sensibly. Both of us have written extensively about our field experience and what we will do now is collaborate in this article to explain how we have used what is available to explain the relationship of various populations. This is with the recognition and realization that the plants occur in highly localized limited localities, are highly variable and yet somehow seem to be integrated and related according to a geographic modality.
It is simply not rational that we may only confidently apply one or other of the many names if we know where a plant comes from. Lawrence Loucka has done an incredibly good job in creating a compendium of Bayer’s writing in a website Haworthia Updates (https://haworthiaupdates.org/). This culminates in a set of what were Facebook postings (in Haworthia Shucked ) under the title “Mysteries”). In these, particularly “part 36”, it is posited that named species such as retusa, turgida, mirabilis, mutica, pygmaea and emelyae are in fact one and the same species. All Bayer’s writing since the publication of his Haworthia Revisited in 1999 is in pursuit of the goal that Essie independently undertook circa 1980 viz. Just what constitutes Haworthia in respect of different life forms that intrigue all of us even it we do not cognitively know this. The section “Mysteries” in Haworthia Updates deals specifically with Bayer’s exploration in the Albertinia area while Esterhuizen has evidence from an additional six localities that provide the very evidence that Bayer was expecting and looking for.
Together we thus consider that the six species are in fact one single system according to a definition of plant species that we agree on and repeat here … A species is a complex dynamic conscious fractal system of living organisms that are metaphysically, and physically (genetically, morphologically, physiologically, behaviorally) continuous in space and time. It seems logical that these systems must be open as it the same one consciousness that pervades all of creation, and surely that is incontestable. Furthermore there is constant change and not necessarily at all in an interminably slow Darwinian selective way. Both of us have observed dramatic changes within our time spans of observation.
The evidence The evidence for our conclusions is in fact already out there in Haworthia Updates and also in Essie’s more recent Facebook Page Haworthia Veld Memories (https://www.facebook.com/Haworthia-Veld-Memories-110859041540965/). Here we simply post a selected set of 54 pictures of plants from 27 populations from between Riversdale and Albertinia to as far east as Great Brak, east of Mossel Bay. The published names, primarily of species (sic!) rank our present discussion covers are the six already named viz. retusa, turgida, mirabilis, mutica, pygmaea and emelyae, and then fusca, splendens, dekenahii, argenteomaculosa, acuminata, rodinii, pallidifolia, suberecta, esterhuizenii and vincentii. There are no doubt many more names that sprout from a need that should not be met by resort to formal botanical taxonomy or nomenclature. No further discussion of those names is deemed necessary because that is what the literature on the subject already does.
Very important in consideration of these pictures, is the question of randomness and bias. The pictures we have selected were based on selecting what is reasonable from those that are available. Unfortunately the range of pictures available is generally very limited for most populations. Secondly it is difficult to avoid any kind of bias when photographing. Populations can be extremely variable and one can select pictures to either prove similarity or difference. This needs then to be adequately weighted to geographical relationships and habitat. As an example of the latter point is that it the contention of the authors that turgida is really a cliff hanging variant of a flats dwelling retusa. Just how does one formulate a single image from a mélange of different ones? This is probably where the difficulty really lies in reaching agreement because we may simply be doing so differently.
Essie’ perspective. (I have adapted Essie’s direct written contribution to this third person version to try and get a better fit …) Understanding Haworthia is like putting together a giant jig-saw of many thousands of pieces. Only nature knows the full picture, and we will only see that if we can complete the puzzle.
The difficulty we truly struggle with is this. If we try and apply the many names made available to us for plant populations we do know, to further new populations we come across in my field jaunts, we end up nonplussed and confused. In fact sometimes a name applied to some or other population does not seem appropriate even for many of the plants in those same populations.
Essie grew up in Heidelberg so it was just logical that H. heidelbergensis would be the first species he came to contend with. It was described as coming from under one bush just west and practically within Heidelberg itself. What he found there did not match what Smith or anyone else wrote or said. Later he found a second population nearby in which there was nearer agreement at least with the original description. But he also looked at about 10 other populations in the general area and could not imagine how one could apply the criteria of the description to all the variants he was seeing. As originally selected and described, H. heidelbergensis is not even representative of the population from which it came. He had a similar experience with H. turgida that occurs abundantly in and around Heidelberg. It just seemed irrational to formally recognize all the variations. His exploration further east of Heidelberg gradually lead him into a swamp of variables and names that included amongst many more H. heidelbergensis, H. magnifica, H. retusa and H. atrofusca. He could not see or decide where one started or stopped or how the descriptions could possibly explain the situation.
Widening his exploration still further to the east and then west, south and north simply added to his dilemma. Just where did a “species” stop or start? H. dekenahii was a particular problem for him because he saw it change status from a discrete species to a variety of either H. mirabilis, H. pygmaea or H. retusa. He ended up in a situation that he could not give a name to a plant unless he knew where it had originated. This is not something any of us would like to admit to because there are four reasons for the unfortunate truth of the issue. Firstly there is the extraordinary intrinsic genetic variation of the plants themselves, secondly the variation coupled to age and growing conditions (in the field or in cultivation), thirdly the extraordinarily idiosyncratic way in which names are generated and imagined to have meaning, and lastly what we all do have to admit to viz. personal perceptions.
Concluding remarks No formal taxonomic alterations are suggested. The communities and persons involved and interested in these plants need names and we still use names just as if they never disappeared from a formal revision and new classification. Bayer suggested in his very first Handbook (1976) that it is quite unnecessary to consider names as lost just because a taxon is no longer regard as formally valid. It seems foolish to us to expect formal taxonomy to submit to a hobbyist interest, even though it should definitely underlie whatever names are used there.
More important is the idea put forward here that H. retusa and H. mirabilis are a single system that cannot be subdivided into any set of names with equal rank. It does not offer any solution in respect of better delimitation of other species. We have not presented any evidence regarding H. emelyae for example, or how species like H. herbacea and H. reticulata relate to the system we have covered here. The article does not touch on a definite and incredible role that H. floribunda plays as part of the puzzle. Nor on the intricate close relationship this species has with H. chlorocantha, H.variegata and H. parksiana. Furthermore we are both acutely aware that the same complexity is evident among other acknowledged species. So even having a species definition is not resolving the issue of names.
Dr. Davidson has suggested that perhaps the metaphysical element needs to be explored and understood. What this means is that we cannot to continue believing, as “science” intends us to do, that consciousness has evolved in creation. Instead it is creation that has evolved from consciousness. Rather than cite any references to this concept we suggest searching the internet on the topic. The conclusion really seems to us that the systems, organizations and individuals (tasked or self-anointed) to describe and name plant species and their variants, are simply not up to the task. This is largely because of a paradigm of science that is self-limiting.
End note While drafting this article I learned that Davidson has in fact authored a book ”Natural Creation or Natural Selection”. I would have asked “Intentional Creation or Accidental Creation”. Unfortunately too few of us are intellectually up to arguing the subject and it seems to me that our whole society has been dumbed down and fooled into believing that we human beings are the central and most important entities in creation viz. anthropocentrism. There is another belief paradigm that all forms of life have intrinsic value viz. biocentrism. Personally I favour this point of view, even going so far as to say that it underlies this curious preoccupation and brotherhood I feel with Haworthia! There are other authors such as Capra, Sheldrake and Meyer that emphasize a creation that stems from consciousness rather than the converse.
Acknowledgement Both of us have an enormous debt to the incredible number of people who we have touched on our journey and the best we can do is just adopt an attitude of great gratitude for all the interactions great or small, our last hurrah.
Suggested literature Capra, F. The Web of Life, Anchor Books. 1996 Davidson, J. Natural Creation or Natural Selection, Vega Books. 2012 Roads, Michael. Talking with Nature, Journey into Nature, Six Degrees Publishing Group. 2018 Sheldrake, Rupert. A New Science of Life, J.P.Tarcher. 1982 Meyer, Steven. Darwin’s Doubt, Harper One. 2010
Sitting back with time to think and contemplate! Society needs recalibration and while I honestly feel it is actually happening there is still a thick coating of sheer ignorance that holds us back. Here is a set of photographs from what is a very small area with 4 major habitat conditions. Sheet rock under partial shade. Huge pocketed conglomerate boulders shrouded in lichens. A broken small bolder verge and slope. And a gravelly shale area with small low scrub. Look how the plants vary. In my opinion this is all H. mirabilis MBB7513 and a small representation of a vast collage of individuals and populations that comprise that system.
The flower pictures of H. mirabilis MBB7513 show how variable those can be too – despite the familiar “the flowers are all the same”.
This article follows Last Hurrah, an article by the same authors, that is requisite prequel that presents the argument for a single Haworthia retusa mirabiloid system.
Introduction The intention is simply to relate the field experience of two observers who have struggled to use and apply names as they have appeared in the literature, and deal with the extraordinary opinions and views on the classification of these plants. While Bayer has written extensively about the entire S Cape Complex, Essie’s extensive field work has yet to bear fruit and be documented. So in this article the aim is to deal with those relevant names that both Essie and me (as scribe) have applied, or seen others apply, to the retusoids and mirabiloids of the specific area South of Riversdale to Still Bay on the coast. Of course it is problematic that it would be more realistic to regard H. retusa as a name lesser to turgida in respect of field fact when nomenclatural rules require the name retusa be used based on chronological priority. We then expect readers to understand that retusoid and turgidoid are the same, where the suffix “-oid “means resembling or likeness: and also not quite the same. This latter because we see H. turgida as a cliff-dwelling, generally clump forming element, whereas H. retusa is a level-ground version and generally non-clumping.
Several additional names are used in the context we want to write about. Magnifica is one of them. This is a species name applied by Von Poellnitz to a plant collected in the Freshe Reserve at Riversdale by Dekenah. The populations of mirabiloid in that area are incredibly variable and it is simply not sensible to use it in the sense of a species name as even Bayer did in 1976. But we will use it to refer to the likeness as we illustrate the variants. Asperula is a Haworth name and its likeness appears in a Salm-Dyck illustration and cannot be linked to any particular population. Our observation is that turgida and retusa both transition to mirabiloid represented by darker green, rough-surfaced and marginally spinose plants. Vernalis and enigma and are names similarly used for transitional elements north-east of Riversdale. Jakubi is a name applied to a mirabiloid southernmost down the Goukou River. Turgida itself is a very common all the way down the Goukou where at Stilbay Smith described it as longibracteata. Certainly its presence as an apparently discrete element all the way along the Goukou may appear contradictory to our claim. But we are not basing our opinions on frivolous limited local situations like this.
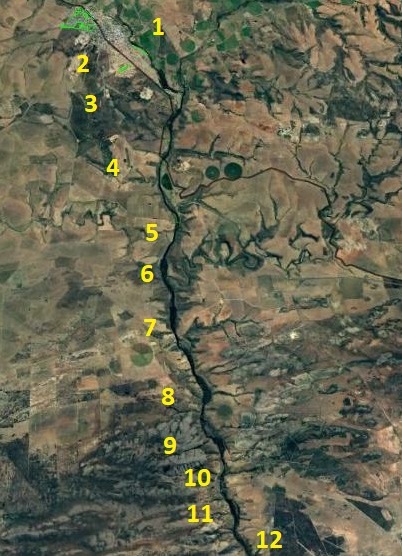
Observations We refer to the accompanying map on which 12 numbers show the approximate location of mirabiloids populations from immediately east of Riversdale down the Goukou River valley for about 20km, with another 16km to before the mouth of the river. We limit the number of pictures because it simply is not practical to present all the illustrations that might tell a story.
We start with group 1 as on the map, which is Komserante immediately east of Riversdale. A mirabiloid occurs there nearly contiguously with a very proliferous large retusoid that was described as H. geraldii. That retusoid was part on a minor continuum that extended southward where I also obtained another name viz. H. fouchei. Both those names can simply be subsumes under H. retusa. The group 1 mirabiloid have been masquerading, thanks to many pretentious taxonomists, as H. asperula.
It essentially refers to a dirty green, rather roughish retusoid with some degree of spination. (Essie’s view was that somehow this common phenomenon in retusoids is due to an influence from H. floribunda, but this is discussed in the prerequisite reading of Last Hurrah). Bayer dismissed the name based on the amazing, wholly confused application of the name by an array of authors. To re-institute that name for the incredible milieu of variants in this Komserante population borders on the irrational – it is irrational. Our conclusion is that this population is unquestionably directly connected to the widespread interaction and interconnectedness of the retusoids and the mirabiloids, as is also the case at Kruisriver where the names enigma and vernalis seem to have been applied. These are names superfluous to science however great they are for commerce and the enjoyment of growing and collecting things. To illustrate locality 1 we have selected pictures to show the fit to the Lower Goukou.
The localities 2 and 3 are within the Freshe Reserve at Riversdale that is where the species H. magnifica originated from. I am partly to blame for the perpetuation of this name as I used it before I had come to grips with the nature of a greater mirabiloid reality. I had also used the name maraisii. My reference number for the Reserve is MBB7163 but there are several localized populations within the reserve all with the incredible variation that is such a feature of Haworthia populations. Our previous article referencing MBB6651 H. mirabilis from SW Riversdale illustrates this.
2. JME 12. JME 22. JME 33. JME 13. JME 23. JME 3
Locality 4 is a population from further south, while localities 5, 6 and 7 are close and along the lower Goukou. Locality 7 corresponds latitudinally with my own observations at Windsor in MBB7818. While two of the pictures are from the different locality indicated by Essie, we show several from the Windsor site. The Windsor plants are not specifically relevant except in a deeper and wider context that is not called for here.
Locality 12 is where H. jakubi originated from. My reference is MBB7820 and I have included some of my own photos here because there is also a second population for this site a little further north.
Here is a further excerpt from Essie’s outline … “The author of ‘jakubii’ does end by saying that the taxon is intermediate between Haworthia mirabilis ‘paradoxa’ and Haworthia mirabilis ‘magnifica’, listing shape, surface, coloration and solitary growth as reasons. This may seem reasonable on the surface of things. But it is totally unreasonable if one considers that one could take representatives from the same and different populations of these, and use the same argument to justify many more names within those two elements as variable components of a much greater cohort of variants.” (The quote is taken from Bayer 2009). It is difficult to know if the author of jakubi had any real authority to make such sweeping statements, but certainly we agree.
The magnifica aspect is the one that captured Essie’s interest and this is what he set out to explore. Also he felt that plants are so variable true and the characters being used to separate them so flexible that had the author of the name really understood that, he would never have described yetanother new species. There is of course a real problem in involving paradoxa and then bobii, from Vermaaklikheid and Infanta westwards as discrete, as the author did. In our view those two elements are mirabiloid and part and parcel of one species system. Vermaaklikheid is 25km west of Locality 12 and Infanta is again 25km further west. Here the geology and general topography play a role and there is nothing that really provides home to mirabiloids as do the inland areas. So any conclusion is really speculative based on intimate knowledge of population relationships inland.
Locality12 is still 16km from the mouth of the Goukou and while H. turgida is superabundant from at least locality 5 all the way to the coast, H. mirabilis seems to halt at point 12. H. variegata is present east of the river in the limestone formations yet it does not seem to reach to the Gouritz river eastwards. That it is absent west of the Goukou and then re-appears west of the Breede River, is just remarkable. We think this supports the mirabiloid/retusoid concept especially in view of our view of a second system made up of floribundoid/chlorocanthoid/parksianoid/variegatoid.
Discussion and conclusion Essie asks just what characters can be effectively used to make authoritative decisions about species if they are so variable. In one of my very first publications I explained how the extreme variability made quantification a statistical nightmare. Another publication explains the problem of examining surface structure and texture using electron-scanning microscopy. It proves the probable flaws of drawing on characters like hairs or spines or different degrees of tuberculation for classification.
So what Essie has done, is spent a vast amount of time and energy in the filed with the very purpose of understanding the names bandied about by people with vested and commercial interest in plants. In this case Essie was motivated by the description of jakubii and comments on its affinity primarily with a so-called magnifica. His determination to arrive at some sort of truth and finding these other populations down the Goukou is nothing short of remarkable. The area is not easily accessible and has involved lonely hikes of 10 or more kilometers where there are no roads or formal trails.
Essie and I wholly agree that the concept of H. mirabilis is about as ethereal as things can get, so is that of H. mutica, H. pygmaea and H. emelyae. What we suggest is trying to get to basics. We are not contesting any of these names in respect of interesting, fascinating and collectable variants. The issue is the nature of the names and their validity in a true system that science is expected to provide. It does not do this. Just consider the earlier definition of “species” that has ghosted on forever and underpins plant classification by default. It is actually derived from animal classification and reads … ”a group or groups of organisms that are interbreeding or potentially interbreeding”. Unexpressed is the vast detail of what interbreeding and subsequent survival of progeny entails.
The evidence from these populations is surely that there is some kind of a continuum from Riversdale itself all the way down to the jakubii locality. Additionally, the Komserante and Freshe Reserve plants suggest that a retusoid element is also present within at least those populations.
We stand by the comments we have made in Last Hurrah. These are … “The difficulty we truly struggle with is this. If we try and apply the many names made available to us for plant populations we do know, to further new populations we come across in my field jaunts, we end up nonplussed and confused. In fact, sometimes a name applied to some or other population does not seem appropriate even for many of the plants in those same populations.”, and finally this … ”More important is the idea put forward here that H. retusa and H. mirabilis are a single system that cannot be subdivided into any set of names with equal rank.”
Reference Bayer, M. B. (2009). Haworthia jakubii – another new species? Haworthia Updates – Essays on Haworthia Volume 5, Chapter 14, pp167-169 [link] ♦
Introduction. Apart from the vexing question of species delimitation, there is a truly curious phenomenon in how the species interact spatially and also behaviorally. The authors here believe in a systems approach as the only rational approach to Haworthia. With H. floribunda as part of one system (viz. with H. variegata, H. chlorocantha and H. parksiana), H. retusa is another (viz. with H. turgida, H. mirabilis, H. pygmaea, H. mutica and probably H. emelyae). Within these two systems there is secondary interaction and we are trying to unravel the role of the two prime elements with H. mirabilis (as a secondary in the H. retusa system). Bayer has discussed floribunda in HaworthiaUpdates.org and it is not necessary to repeat that here. In this article we are only dealing with single populations of H. floribunda and H. retusa that occur contiguously at a locality SW of Riversdale where they were located by Essie. H. mirabilis does occur 600m to the south-east as a discrete population in a kaolinite exposure site as opposed to the rocky more stable nature of the H. floribunda/H retusa site. It is important to note two things:
This locality is southwest of multiple discrete populations of H. mirabilis as well as H. retusa, while …
It is the only known occurrence for H. floribunda south of the N2 highway from west of Heidelberg to beyond Albertinia.
Preamble. This is neither a taxonomic and nomenclatural debate nor any attempt to explain how these things came about. It is simply the synthesis of the experience of two field enthusiasts and explorers trying to rationalize what they observe in the field. Trying to make sense of many latin names. It is Essie’s suggestion that H. floribunda and H.retusa interact at a deep level and that H. retusa becomes rougher and darker where the two are in close proximity. They do flower at different seasons with a general and approximate 4 month time barrier. Bayer on the other hand suggests that the two species definitely do produce hybrids despite difference in flowering time (observed in several places as isolated individual hybrids). The apparent roughening and darkening observed in H. retusa is more likely to be consequent to a deeper direct connection with H. mirabilis that may have nothing to do with hybridization. As observed elsewhere, hybridization seems to be a convenient excuse to explain phenomenon that have no other obvious origin. What then happens when H. retusa and H. floribunda virtually share habitat here SW Riversdale.
To understand the situation better we have to look at that H. mirabilis population mentioned to the south (MBB6651). I personally have no photographs because I saw the population before I had a digital camera. But I grew them from seed and the accompanying pictures show the incredible variation that suggests an interaction of the retusoid, the mirabiloid and the floribundoid. Essie made a special excursion to obtain 3 field photographs.
The plants. Multiple photographs of the two species accompany this discussion. H. floribunda can be seen to be H. floribunda with perhaps one notable exception that could be dismissed as a hybrid with the greener coloured H. retusa. The H. retusa plants on the other hand do show some variation. The plants are smaller and generally greener than is the case with other nearby populations.There are indeed some plants on the much darker side and this is where an important distinction may be made between the retusoid and the mirabiloid factions (also spring vs late summer flowering) of a single system. The retusoid system is characterized by light coloration with yellows greens and pinks, while the mirabiloid faction is darker green to blackish green with rusty reds and browns. Two plants are shown from an essentially retusoid population about 33m north.
Conclusion. We do not draw any conclusion from this single location that simply adds fuel to our belief in system and that far less formal names are needed to explain the incredible complexity and high degree of variation. (Note to writer … “Keep it simple, stupid”.) We are further suggesting that all these varied populations are elements of just two complex species systems, and not the fragmented cherry-picked elements that the many Latin names suggest.
Footnote by MBB. It is necessary to write something about Essie Esterhuuizen because whether he knows it or not, he has made an incredible contribution to Haworthia by field exploration that is beyond extraordinary and exceptional. I asked him of he could explain his dedication and commitment to Haworthia and this is what he wrote …
“I was fortunate to have grown up in Heidelberg in the Western Cape. As children we spent most of our time playing in the veld around the town. From a very early stage I was thus exposed to Haworthia and needless to say it formed part of the plants on our rockery. Because of their small size I kept 8 different plants in my so called “collection” which I regarded as a collection of notice. Around 1982 I visited the Karoo Botanical Gardens to see if I could find names for the “small aloes” on the rockery when Bruce at that time the Curator of the garden showed me the garden’s collection. Suddenly I realized that if I want to continue with my hobby – the challenge is humongous.”
Now Essie is Afrikaans speaking and Afrikaans is a language very dear to me because it is what I refer to as a heartfelt language – a language of real people and not like English that is used and abused by many cultures and castes. So I asked Essie to rather write in his home language and leave it to me to try to extract the really personal essence of his interest and opinions. So translated by me, this is what he says…
“I am not a scientist, a collector, or a nurseryman – just a plain field observer. In my first field excursion involving the name heidelbergensis, I was shocked to find that what was known and written about these plants was just a drop in a bucket. Just judging and identifying plants on the basis of what is written, or in collections, does not reflect what the situation is in the field. On the other hand, to explain what one observes in the field verbally or in writing is extremely difficult. Field observation becomes like a treasure hunt. When you think you have found some sort of answer, you find another clue to lead you on.”
The crux of the matter is that it is extremely difficult to articulate what is meant in this last sentence. By putting our heads together we agree that there is pattern in this very complex system, as we have just explained in the article “Last Hurrah”. We have planned to collaborate in presenting the evidence for the continuity we say exists within and between two species systems. Essie’s final sentence reads …”For me my observations will never come to an end … the challenges are too great, continuous and wide ranging”.
Fig. 1 H. mirabilis 6651JME SW RiversdaleFig. 2 H. mirabilis 6651JME SW RiversdaleFig. 3 H. mirabilis 6651JME SW RiversdaleFig. 4 H. mirabilis 6651JME SW RiversdaleFig. 5 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 6 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 7 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 8 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 9 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 10 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 11 H. mirabilis 6651 (cult. exSW Riversdale)Fig. 12 H. mirabilis 6651 (cult. exSW Riversdale)
A note on a H. floribunda and H. retusa interaction was written by Bruce Bayer based on the opinions, observations, and exploration of Essie Esterhuizen.