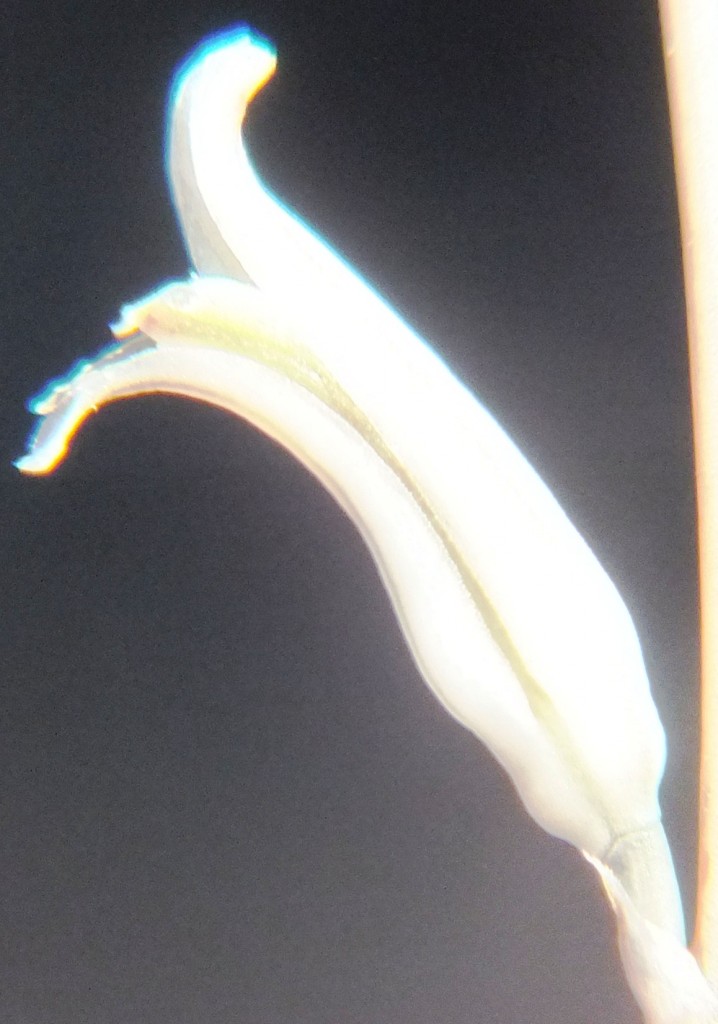
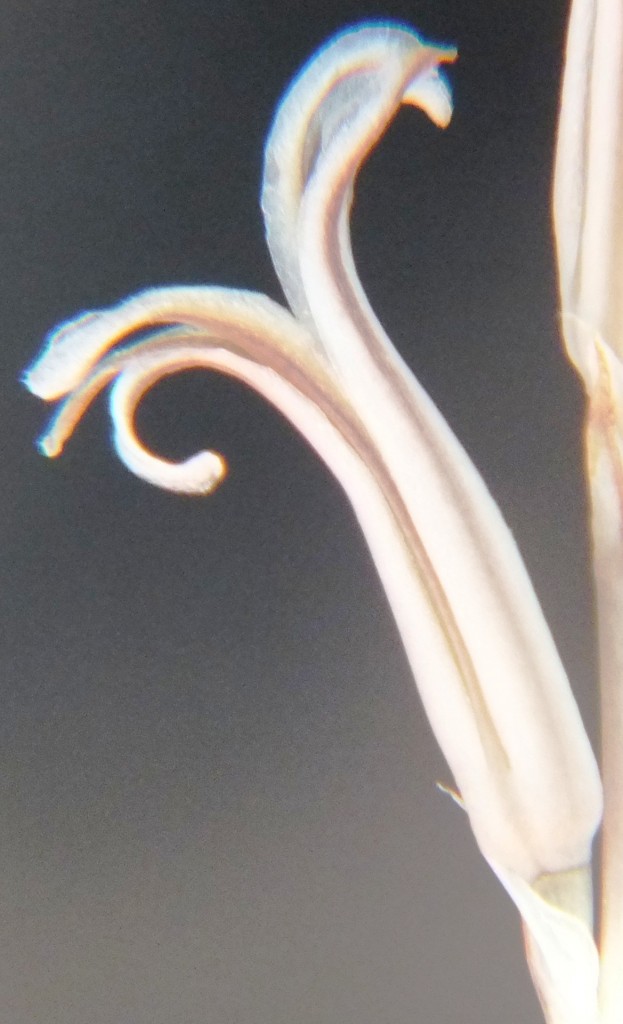
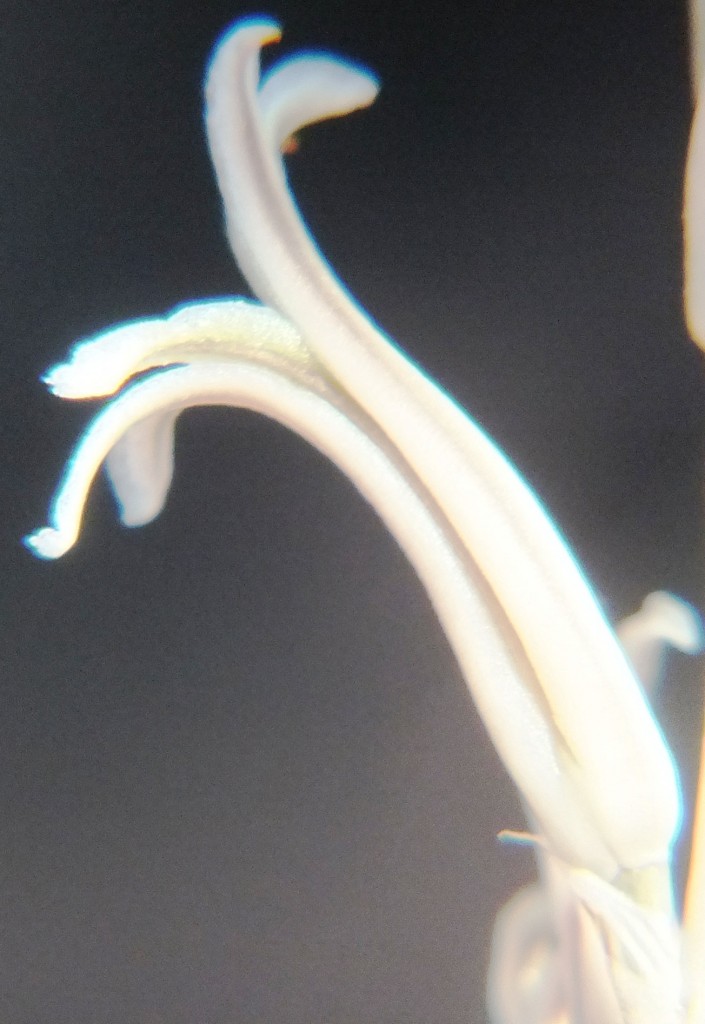
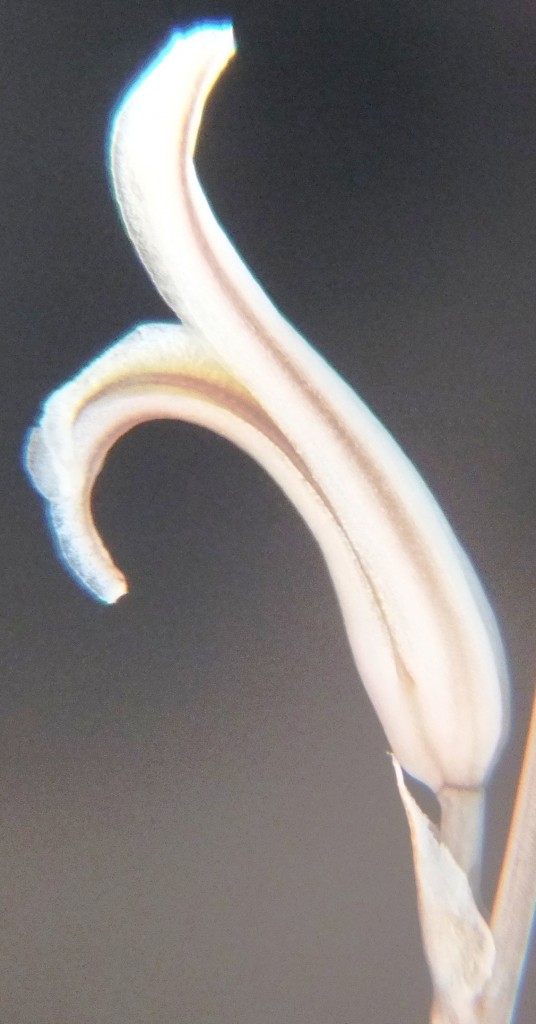
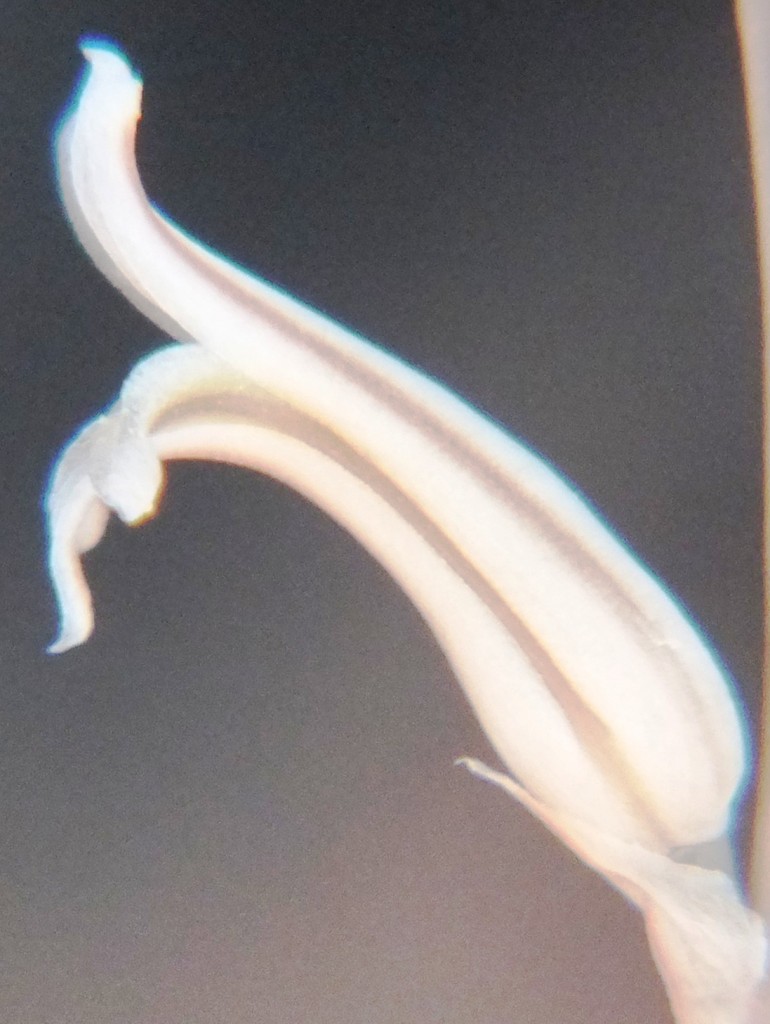
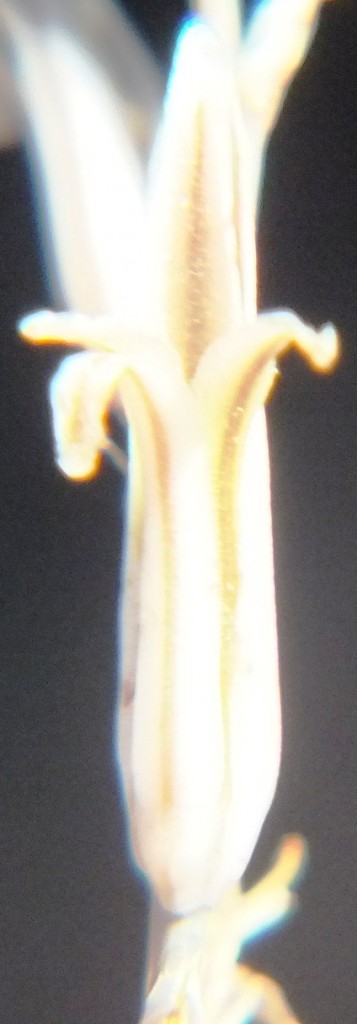
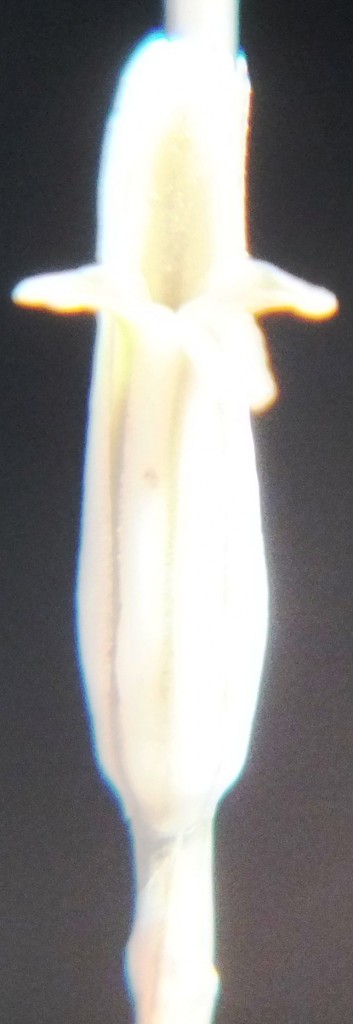
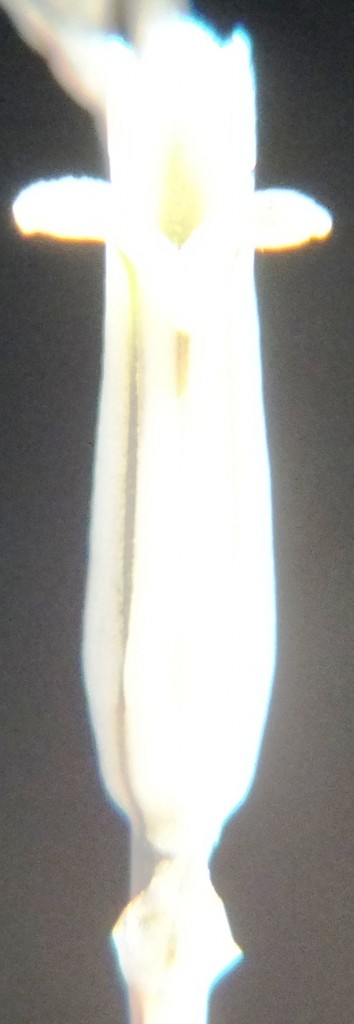
Two further records and data for Haworthia mirabilis.
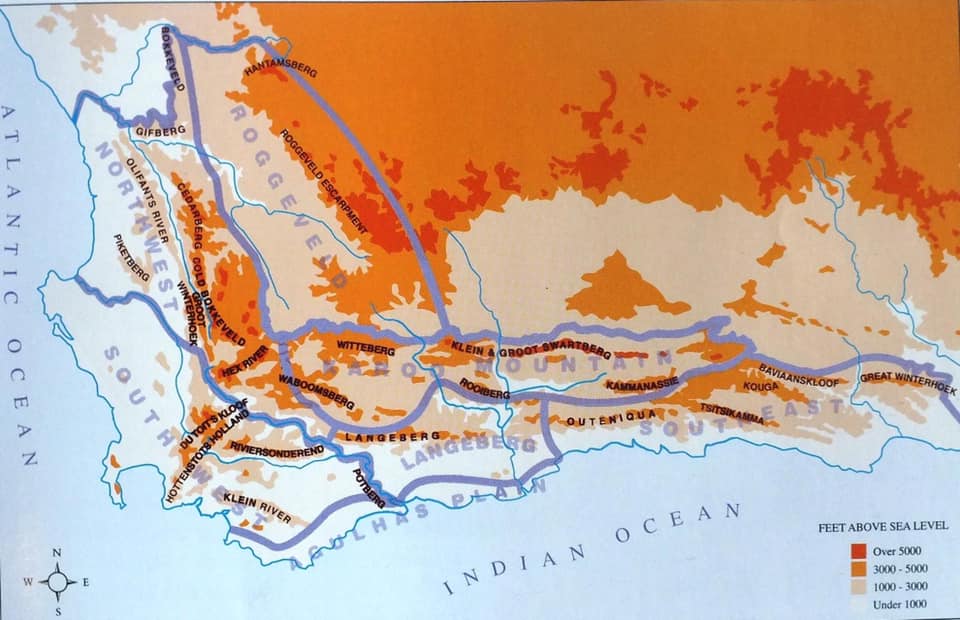
Kobus Venter drew my attention to a second population of H. mirabilis on the eastern boundary of the farm Schuitsberg, which is the origin of the var. beukmannii of von Poellnitz. But the real motive for exploration in that area was a photograph sent to me by Messrs. Daryl and Priscilla Hackland of a plant on the Zigzag Path just east of the northern end of the town of Greyton. It was their son Andy Hackland who observed the plants and thought I might be interested – indeed. The bright green colour of the plant struck me as most unusual for that area where most of the populations known are south of the river in shale and they are of the brown and red hues. This is not strictly true because records of an expedition by Messrs. Beukman, Otzen and others mentions a small “black” species just before the entrance to Greyton. I have never found that.
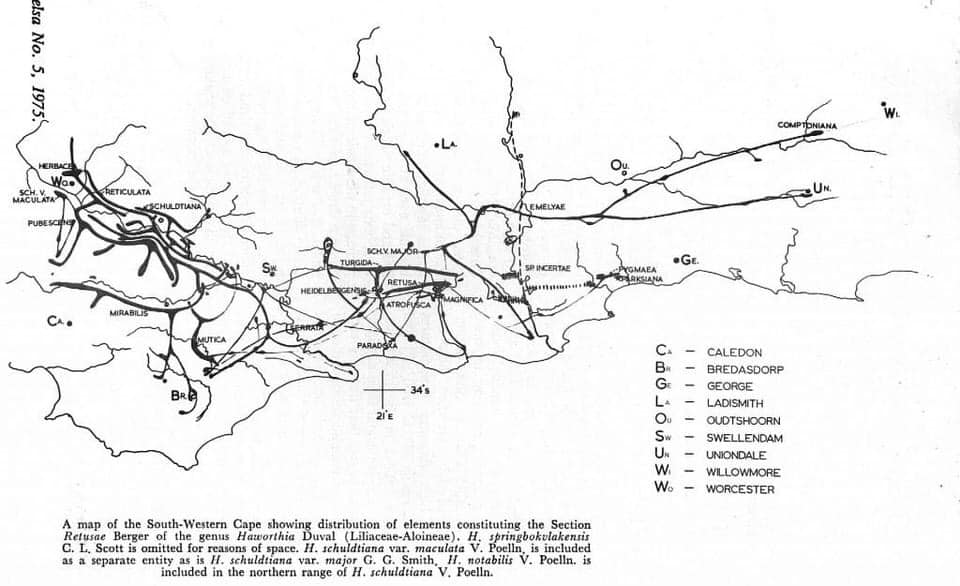
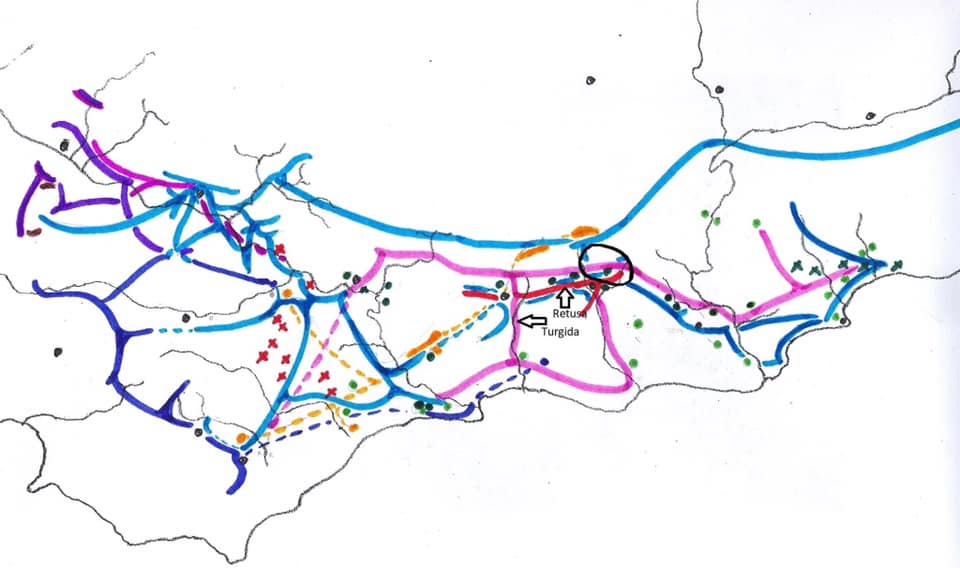
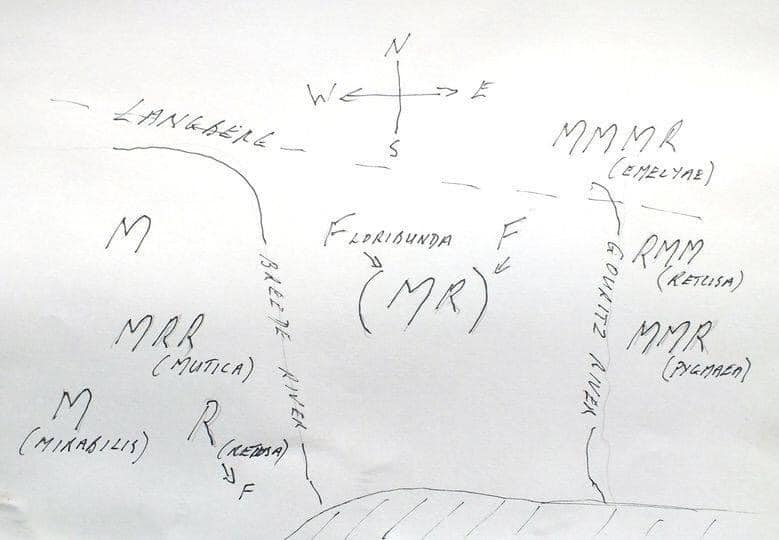
However, it is well known that Haworthia mirabilis occurs along the course of the Boesmansrivier on the way to MacGregor. I once observed it there myself hiking part way from the northern end of the walking trail down the river to Greyton. Kobus Venter has also seen it while walking from the southern end. I included this variant under the varietal name ‘consanguinea’ that is actually based on populations on the north side of the mountain in the Dwarswaterkloof west of MacGregor. Ms. Dawn Schwegmann once showed me plants from that general area but I never did establish exact origin. What did strike me about the Boesmansrivier plants is the very close resemblance to Haworthia retusa ‘turgida’ when I still considered populations that occur in Table Mountain Sandstone in the Tradouw Pass and north of Riversdale as a discrete species. The only difference to me was the summer flowering time and the resemblance of the flowers to those of H. mirabilis from the Bredasdorp/Napier area. This observed similarity was largely based on a very small sample, colour, and the fact that the flowers were available simultaneously. I must have assumed that this similarity was unlikely to include ‘turgida’. Unfortunately photography was expensive and almost unmanageable for this sort of detail in that era and I had to observe and record from memory.
While I now have only the memory of the turgida-like element in the pass and a myriad of varying populations of H. mirabilis at the northern end and around MacGregor, I have had no evidence of transition to the red-brown plants (eg ‘rubrodentata’) south of the Riviersonderend River from east of Genadendal to at least Lindashof east of Riviersonderend town. This is as the species occurs in putatively Bokkeveld strata.
The Hacklands saw the plants at the top end of the Zig Zag path and the geology here will need a better description than I can give without a geology map. The vegetation is transitional to Fynbos and the stratum an argillaceous one (clayey) not obviously Sandstone nor shale. But the plants are so different. They are smaller than in populations in the area that I know south of the river, fairly bright green and becoming rather yellow in summer stress dress. A physical description of the plants defies description and I would be a loss to refer to the plants as anything but a variable lot. In the amazing context of Latin names in Haworthia I would like to refer to the plants as Haworthia mirabilis ‘hacklandii’ – if it can be forgotten and forgiven (it unfortunately cannot be) that I question the merit and motives for using personal names for taxa. In any case, the serious limitation is to know how to descriptively and sensible circumscribe such a population. The alternative reference is H. mirabilis MBB8040 or H. mirabilis NE Greyton, or H. mirabilis Zig Zag Path. Whatever name is used, this population is in my opinion quite an awesome demonstration of the morphological continuity that I have come to expect in Haworthia. A wide range of comparisons is possible if one takes single clone after clone. I thought ‘sublineata’, ‘mundula’, ‘ turgida’ (from a range of populations) as well as ‘notabilis’ and even ‘elizeae’ and ‘mutica’). The departure from the wider dark green colour of the mirabiloids is the most unexpected feature although this is just what occurs with ‘consanguinea’. This latter is the ground truth and I fully expect that further populations occur to complete a transition to the even more obviously ‘turgidoid’ variant known from deeper in to the Boesmansriver valley. Later driving eastwards and a little further south towards Schuitsberg, we observed very similar habitats 2-3km further north of the road in foothills of the Riviersonderend Mountains.
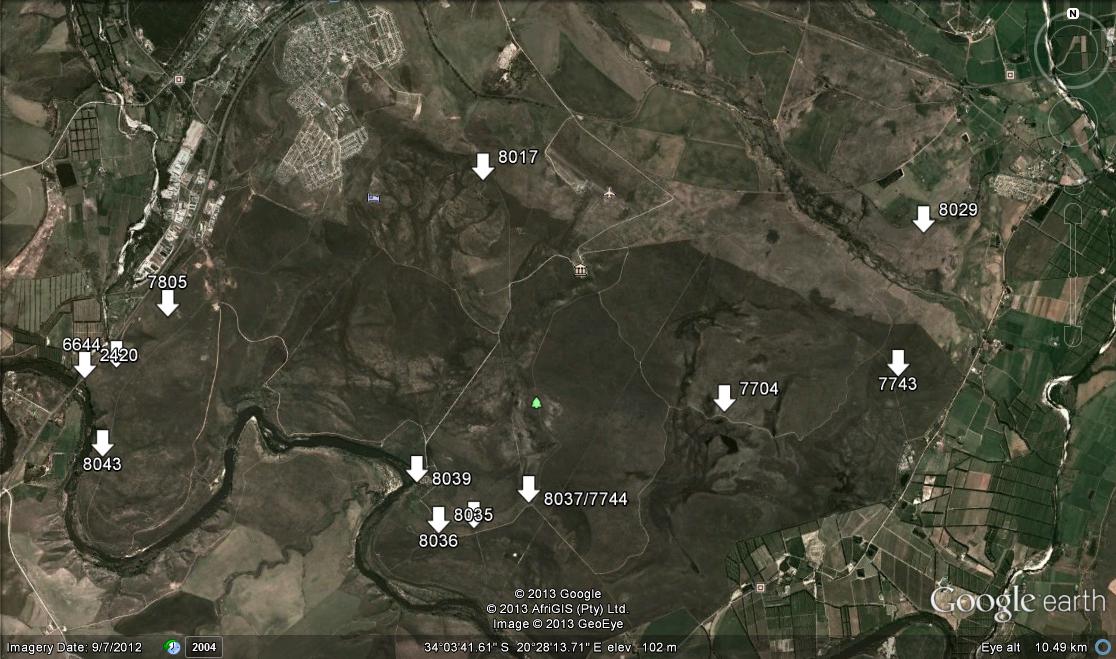
Among the pictures of the plants is a view from the top of the Zigzag Path looking north into the Boesmansriver valley (or gorge). The hill on the right front seems to be similar to that of the Path and similar low rocky hillsides occur at several places. Significantly there is one immediately north of Riviersonderend where the river passes by on the north side rather than on the south away from the higher hills away from the main range. Kobus has persistently observed that there must be plants there. The evidence of the Zigzag Path occurrence definitely suggests that this must be so and that many more populations exist unrecorded simply because of lack of exploration specifically for Haworthia. While Andy observed the plants at the top of the Path, they are widespread all the way down to the bottom. It is quite amazing that the presence of the plants has not come in to my ken sooner. It is just an observation of mine that Haworthia distribution may be far wider than we suspect simply because Haworthiophiles have not ventured into unlikely areas.


























 that runs towards MacGregor 25km to the NE - and to H. mirabilis 'consanguinea'. The weathered cardboard in the foreground is a very sad but loving tribute by children to their grandfather - messages of love and longing painted on two little stones, on the remains of a shrivelled dried wreath.")













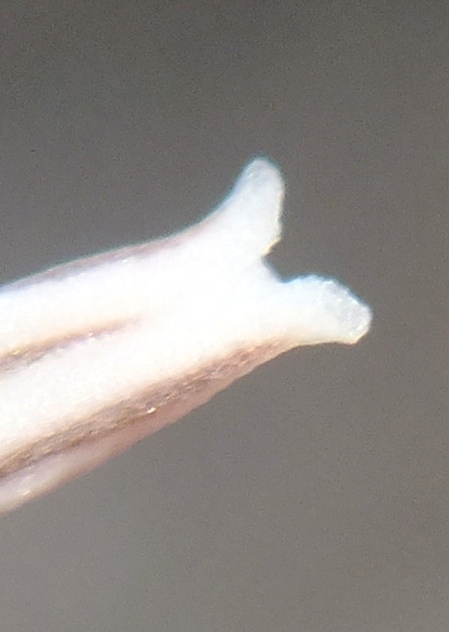
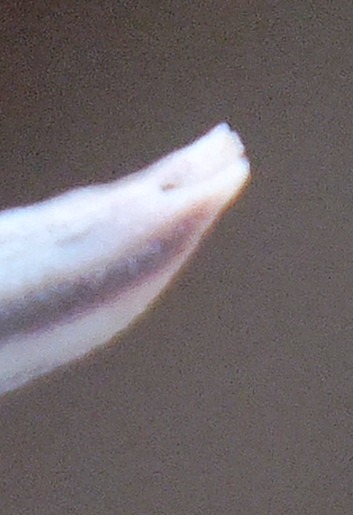
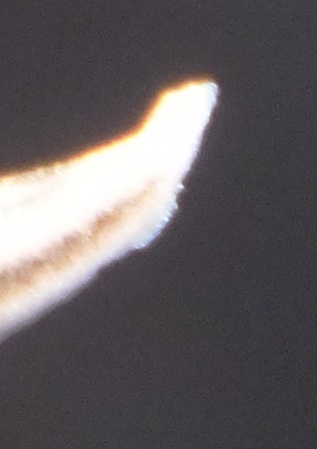
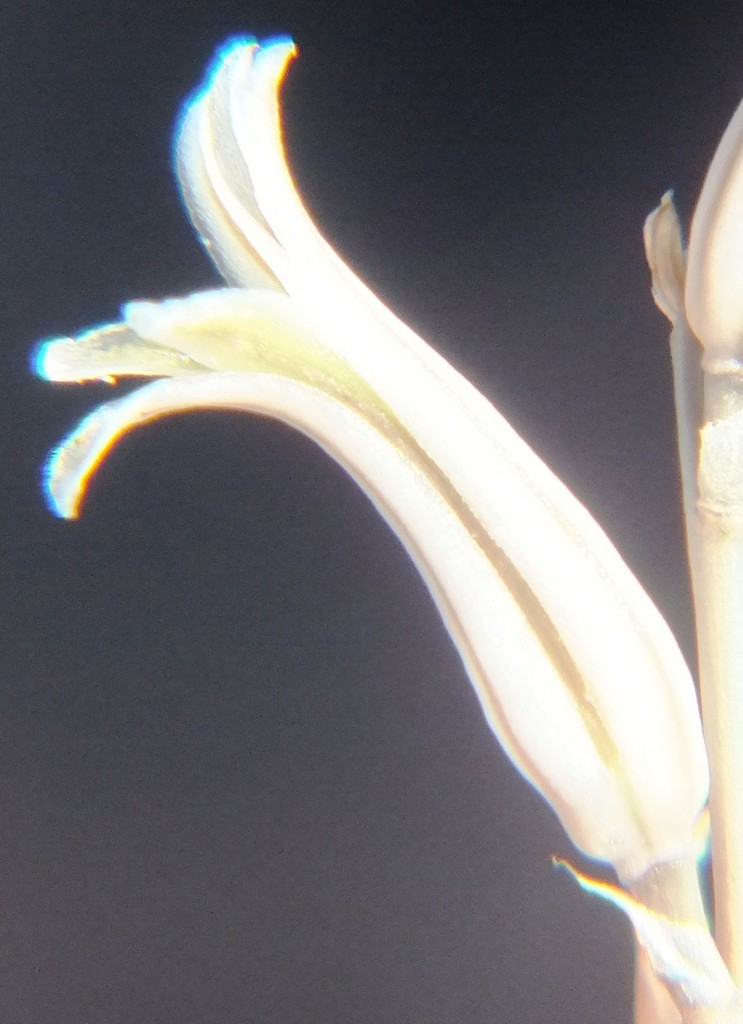
Flower profiles, faces, and buds
































































From Greyton we went to Schuitsberg driving further east than previously. This population seems to have led to the speculation that the type of H. mirabilis as illustrated in Botanical Magazine t1354 (1811) could have come from here. In a recent article in Alsterworthia, Gerhard Marx attempts to ridicule my typification of the name ‘mirabilis’. I frankly think this is much more a point scoring effort than a genuine attempt to arrive a taxonomic truth to which Marx claims to aspire. Apart from speculated similarities of some or other clone to that picture, is the fact that this is an old transport route dating to that era. There is a plaque at that site dated 1988 commemorating a 300year history. The problem for me is the variation among the plants that renders the nomenclatural system and the typological approach to naming dismally doubtful. There is just no Haworthia mirabilis var. mirabilis.
Schuitsberg is the origin of H. emelyae var. beukmannii von Poellnitz .I associate that name and description with an exceptional robust, compact and retused version of H. mirabilis (a version with much longer erect leaves with spined margins viz var. rubrodentata {red teeth} was named from further west along the river). I visited Schuitsberg and adjacent farm Nethercourt in the early 1970s. The plants I saw were indeed robust. However I visited there again 2012 and found the place previously visited destroyed and no plants among the many in the adjoining rocky slopes that resembled what I perceived to be ‘beukmanii’. This is repeated in my experience of this second locality on Schuitsberg east (MBB8041) of where I knew the plants. One can refer to the many variants illustrated with this article. Are these all individually or collectively closer to any early illustration on which the name ‘mirabilis’ can be based? It was only a single large very robust plant with leaves nearly 30mm wide and 20mm think that I would have said also agrees with the original acceptance of the “element”. So what can we say here of all these variants? This is a fact of the retusoid species and close relatives. They are simply highly variable with respect to leaf shape, arrangement, markings, armature, and colour, as well as flower characters. The consequence of these is gross aberration of the formal nomenclatural system and whole host of Latin names that are very useful to collectors, commercial nurseries and any other activity where a formal impressive name adds value. What we have at a place like Schuitsberg is that we have plants that can be labelled ‘beukmani’ because they might look ostensibly like the original described plant or its image, or just because it happens to come from Schuitsberg.






































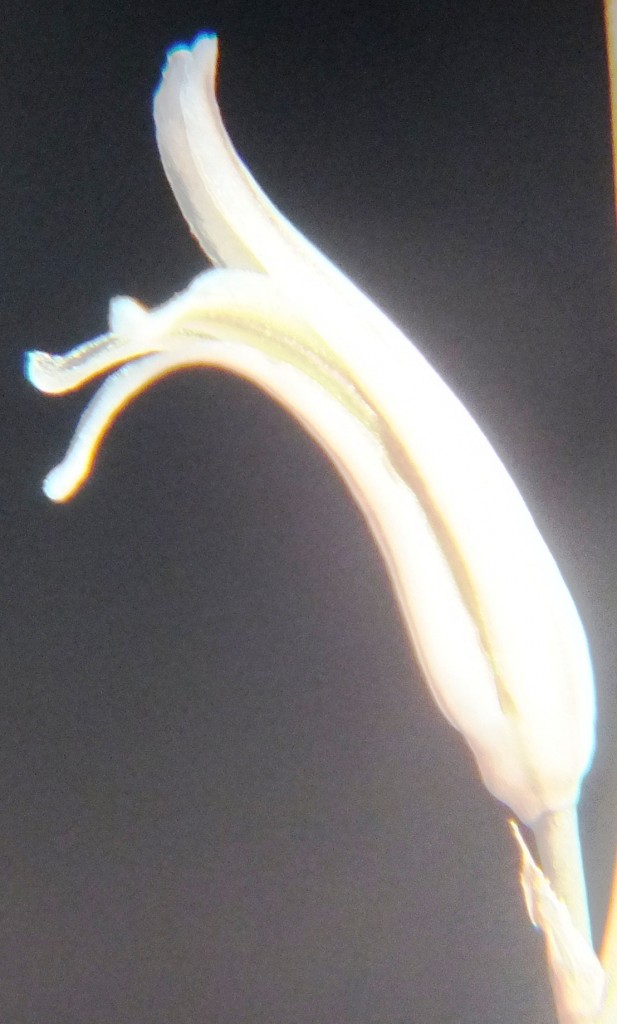
Flower profiles, Faces, and Buds.



















































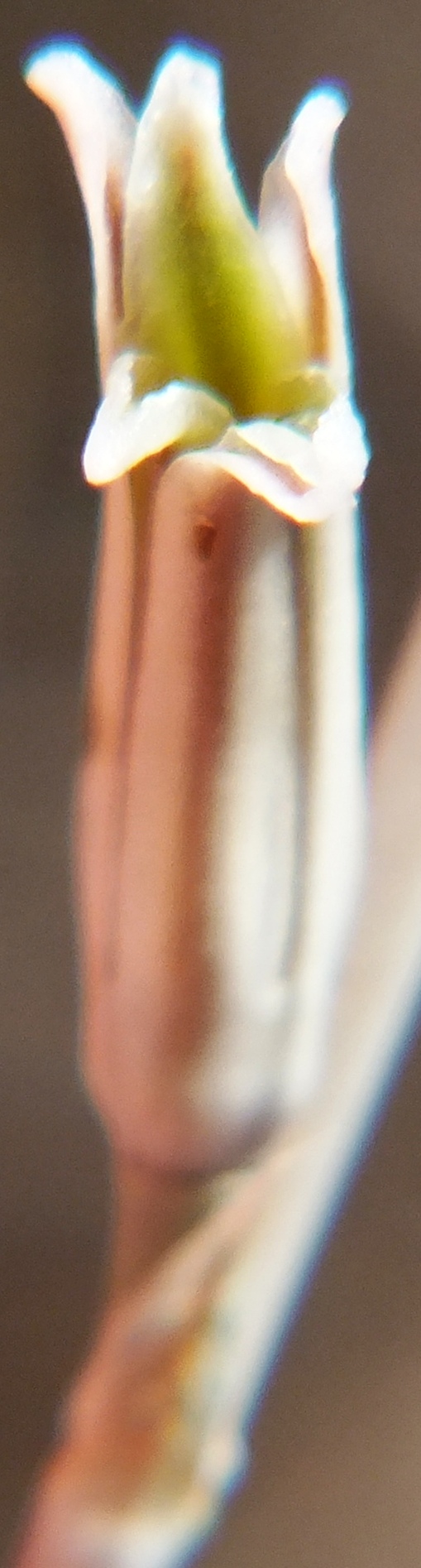




Regarding the flowers of both localities that I illustrate here, there is again the high variability that I discuss and illustrate in all the preceding parts of Haworthia Update 8. What we now need is a computer wizard who can perhaps explore the possibility of computerised pattern recognition. In this way to look at trying to arrive at a composite single image that captures the identification of a population. It boggles my mind that someone might think, say ‘beukmannii’, is an array of plants scattered through many populations to so constitute a species.
I have tried to approach this whole problematic, provocative, disputatious subject from a rational objective view based on my own education, experience and acquired knowledge to place it in an environment acceptable to formal botany. That environment has not been all that helpful and conducive to a solution either.
Acknowledgement
I would like to thank Dr. Daryl and Mrs. Priscilla Hackland, and Andy Hackland for information and assistance in visiting the Greyton Zigzag Path population. Mr. P.G. Viljoen not only kindly allowed us access to Schuitsberg but took the trouble to drive us there to show us a wider view of the localities and also the commemorative plaque that mark the old travel route. ♦