18. Haworthia magnifica V.Poelln., Feddes Repert.Spec.Nov. 33:239(1933). Bayer, Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). H. maraisii var. magnifica (V.Poelln.) Bayer :131(1976). Type: Cape, Riversdale commonage, Mrs E. Ferguson. Not preserved. Lectotype (B&M): Riversdale, Ferguson (BOL).
magnifica: magnificent.
Rosette stemless, slowly proliferous, to 8 cm φ. Leaves spreading, retused to ground level, dark green to purplish, scabrid to finely spined margins, end-area slightly translucent between the veins, surfaces with small slightly raised tubercles. Inflorescence slender, to 40cm. Flowers brownish veined, few open, upper lobes pinched at tips.
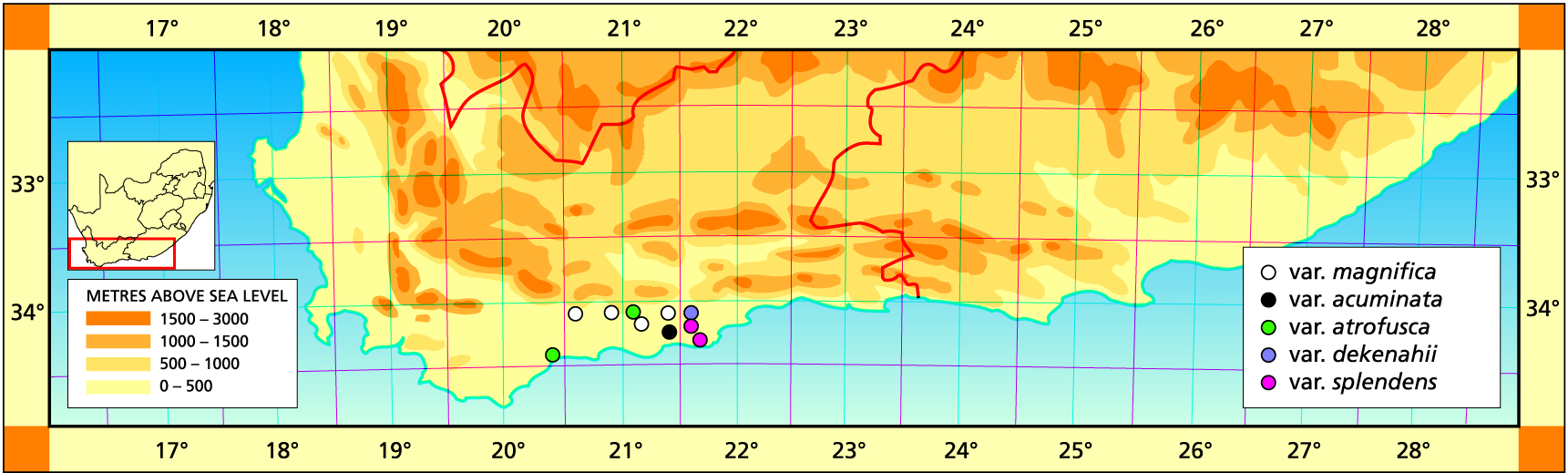
1982 – Haworthia magnifica is an incredibly variable species and it will not be possible to gain any insight into probable affinities with H. turgida, H. retusa, H. emelyae and H. mirabilis without in‑depth study. It is characterized by its dark‑green colour, small size, short green‑lined buds and flowers, and ‘fish‑tail’ bud tips. It flowers in March ‑ April although this varies and only serves as a character to distinguish the species from H. mirabilis. Only the more notable and widely spread forms are recognized as varieties here. There are several other populations which could justly be named as varieties. The variety major has an unusually large flower similar to that of H. emelyae and there is evidence of intergradation of these two taxa. The var. paradoxa may constitute a south‑eastern link between H. magnifica and H. mirabilis, except that truer forms of H. magnifica var. maraisii intercede along the Breede River and at Bredasdorp. The var. maraisii at its type locality at Stormsvlei is quite a robust form which adopts many guises in the Robertson/Worcester area. South of Stormsvlei there is an aberrant population which appears intermediate between the var maraisii and H. mirabilis. However, just north of Stormsvlei, H. mirabilis occurs in a small, many‑leaved form. The var. meiringii appears vegetatively very like a smaller, darker green version of H. herbacea, until it flowers. Also to the west it intergrades into the more characteristic retuse‑leaved var. maraisii. The var. notabilis also has erect leaves which are darker green and more turgid than in the case of H. herbacea. The var. atrofusca was originally collected from west of Riversdale and is characterized by its rounded leaf tips. The var. magnifica has unusually long acuminate leaf tips and is restricted to a small area southeast of Riversdale, recurring again just west of Heidelberg. H. magnifica has been recorded north of both Montagu and Barrydale and as far south as Cape Infanta and westwards to Bredasdorp. Consideration of the variability of this species and distribution of variants is very helpful towards understanding variability in the genus as a whole.
1996 – The comment ‘in-depth study’ is quite inappropriate in the Haworthia literature which generally suffers from any formal objective study. What has happened since 1982 is much further collecting which has brought some new perceptions. The problems with this species are exemplified by Scott’s circumscription and synonymy of H. asperula where it is barely possible to separate all the diverse elements included in it. Esterhuizen wrote in ALOE (1996) that it was easier for him to separate the var. atrofusca from var. maraisii, than from magnifica. This perspicacious observation is all the more so for the new collections from east of Riversdale, and also for the changed view of H. heidelbergensis. It has been decided to separate the elements maraisii and magnifica because it appears that they relate to each other as H. heidelbergensis does to H. mirabilis. It is also more realistic that some of the varieties that were with magnifica are rather variants of H. maraisii. Thus H. magnifica is applied to the eastern populations which are rather more robust and in which the end-area of the leaf is longer and more pointed. If the end-area is shortened it is also rounded. H. magnifica is usually more greenish than the nearly black H. maraisii. Where previously considering H. retusa as a major element has led to problems, the perception that H. turgida and H. magnifica are the main role players makes it easier to understand what the different populations may represent. This should be apparent from the varieties recognised here, and in the corresponding discussion.
a. var. magnifica. Originally from only south of Riversdale. The name is now also applied to the population from south of the Tradouw Pass, and to that just east of Riversdale. These are quite robust plants with rather scabrid, sub-tuberculate leaves with denticulation of the margins at least. The surfaces may also be slightly scabrid with the tubercles bearing small spines. The plants are lighter green in colour than H. maraisii.
Distribution: 3420 (Bredasdorp): Tradouw Pass (-BA), Smith 6788 (NBG). 3421 (Riversdale): S. Riversdale at beacon (-AA), Smith 5372 (NBG), Bayer in KG83/71 (NBG); Reserve (‑AB), J. Dekenah 16 (NBG); E. Riversdale (-AB), Smith 5376, 5376a, 5748 (NBG), Bayer in KG92/71 (NBG); Riversdale (-AB), Dekenah 6a (PRE); (-AB), Smith 5372 (PRE); Riversdale (-AB), Muir 3553 (PRE).
Inadequately located: Riversdale, Smith 3900, 5057 (NBG).
Haworthia magnifica var. magnifica JDV93/35 Tradouw Pass. The plants here are very variable and usually much greener than typical.
Haworthia magnifica var. magnifica JDV93/13 south-east of Riverdale. Characteristic of these plants are the very dark colour and acuminate, retused leaf-tips.
Haworthia magnifica var. magnifica JDV97/131 Also north-east of Riverdale. Two adjacent populations which show translation to H. retusa.
Haworthia magnifica var. magnifica JDV94/6 north-east of Riversdale is very similar to plants at Tradouws Pass.
Haworthia magnifica var. magnifica JDV97/39 Barrydale. North of the Langeberg, these plants are more like H. maraisii. However, it seems probable that they will be shown to link up with H. emelyae var. major along the northern slopes of this range.
b. var. acuminata comb.nov. H. retusa forma acuminata Bayer :94(1976). H. retusa var. acuminata Bayer :53(1982). Type: CAPE‑3421 (Riversdale): N. of Gouritzmond (‑BD), Bayer in KG 311/7 (NBG).
acuminata: sharp pointed.
Previously under H. retusa, this variety has been transferred here because of the now restricted view of that species, and the new concept of H. magnifica. It is only known from the one locality as the original nearby locality appears to have been destroyed.
Distribution: 3421(Riversdale): N. of Gouritzmond (‑BD), Bayer in KG 311/7 (NBG), Bayer 2423 (NBG); S. Gouritz Bridge (-BD), Smith 5047 (NBG); 9.5km Gouritz to Albertskraal (-BD), Smith 3946 (NBG).
Haworthia magnifica var. acuminata JDV94/94 north of Gouritzmond. The leaf-tips are much more acuminate than in H. retusa, and the plants are darker green.
Haworthia magnifica var. acuminata JDV91/39 north of Gouritzmond. Plants are proliferous.
c.var. atrofusca (Smith) Bayer Natn.Cact.Succ.J 32:18(1977). Bayer :44(1982). H. atrofusca Smith, JS.Afr.Bot. 14:41(1948). Bayer :100(1976). Scott :130(1985). Type: CAPE‑3421 (Riversdale): (‑AA), J. Dekenah 225 in Smith 6169 (NBG).
atrofusca: very dark brown.
This variety was represented by a single small population to the west of Riversdale and characterised by the blunt rounded leaf-tips. This is in effect a single character which occurs in other populations and other species. The scope of the variety is widened to include the large element to the north and west of Riversdale collected mostly by C. Craib (unpublished). These are large brownish-green to blackish plants which are densely and finely tubercled against a scarcely translucent background. The original variety and populations to the west include forms with sharply pointed leaves. There is a very interesting population in the Potberg area to the west of the Breede River. The habitat is identical to that of the type locality. It is very unusual to encounter such a vicariant distribution record in the genus where isolated plants so closely resemble the type.
Distribution: 3420 (Bredasdorp): NW. Kathoek (-AD), Bayer & Bruyns 6549 (NBG). 3421(Riversdale): (‑AA), J. Dekenah 225 in Smith 6169 (NBG), Bayer in KG202/70 (NBG); Droerivier (-AA), Bayer 2665 (NBG).
Haworthia magnifica var. acuminata JDV90/91 west of Riversdale. Leaf-tips rounded.
Haworthia magnifica var. acuminata JDV97/146 west of Riversdale. Leaf-tips rounded. The ‘aubergine’ locality of C. Craib.
Haworthia magnifica var. atrofusca JDV90/91 west of Riversdale. The dark coloured species seem to need more protection from direct sunlight.
Haworthia magnifica var. acuminata JDV92/97 north-west of Riversdale. More robust plants with even more rounded leaf-tips.
d. var. dekenahii (Smith) Bayer comb.nov. H. dekenahii Smith, JS.Afr.Bot. 10:140(1944). H. retusa var. dekenahii (Smith) Bayer :53(1982). Type: Cape, on farm Draaihoek (-BA), J. Dekenah 86 in Smith 5489 (NBG).
dekenahii: for Japie Dekenah, a born naturalist.
This element as a species was completely discarded in the 1976 handbook and resurrected as a variety of H. retusa in 1982. Col. Scott upholds it as a species but illustrates H. turgida var. pallidifolia which co-occurs with it. The significant things about this variety are the raised tubercles on the leaves, the silver flecks, and the blunt rounded leaf-tips.
Distribution: 3421 (Riversdale): Draaihoek (-BA), J. Dekenah 86 in Smith 5489 (BOL,NBG,PRE).
Haworthia magnifica var. dekenahii JDV87/146 north-west of Albertinia. Plants are fairly scabrid, darkish-green and the leaf-tips are rounded.
Haworthia magnifica var. dekenahii JDV87/146 north-west of Albertinia. Plants do not proliferate and can be very cryptic.
e. var. splendens Hammer and Venter Cact.Succ.J(U.S.) in ms. Type: W. Albertinia (-BA), Venter (NBG).
splendens: splendid.
The name is indeed apt. I first saw this plant in about 1970 in a visit to Dr Hans Herre. I simply assumed the unlabelled pinkish-red plant with the shiny black raised tubercles was an unusual specimen of H. emelyae which itself was practically unknown to me at that time. The full story of this variety is better told by the authors whose persistence and tenacity led to its rediscovery at what is presumed to be a second locality – the first apparently having been destroyed. Strangely enough a further population was discovered fatefully, and perhaps fortuitously, by Mary Parisi and Ed Dunne to the east of Albertinia. The word fatefully are used with some deliberation because these two people deliberately avoided the contamination of plans aforethought in wanting to do their own exploration and discovery. This eastern population flowers earlier together with the var. acuminata and also with H. emelyae. There is obviously some significance to this which implies some cross-mountain connection and throws some doubt on the relation of the H. emelyae varieties.
Distribution: 3421 (Riversdale): E. Albertinia (-BA), Marx sn. (NBG); W. Albertinia (-BA), Venter (NBG).
Haworthia magnifica var. splendens JDV97/127 Albertinia. Spectacular glazed pieces of pottery.
Haworthia magnifica var. splendens JDV93/59 west of Albertinia. The tubercles on the leaves can be very dark coloured.
Haworthia magnifica var. splendens JDV97/127 Albertinia. Possibly this is the spot where Herre first collected this variety.
Haworthia magnifica var. splendens Kent sn. Originally collected by Dr. Hans Herre from west of Albertinia.
Haworthia magnifica var. splendens JDV93/59 west of Albertinia. Rediscovered by Venter, Kent and Marx. Flowers here in late summer.
Haworthia magnifica var. splendens JDV94/108 east of Albertinia. The leaf surface in this population are very mottled. Flowering time is in the spring although it seems as though the plants may also flower in late summer in cultivation.
Haworthia magnifica var. splendens JDV94/108 east of Albertinia. Plants from here have more acuminate leaves than in plants from west of Albertinia; also the leaf-tips are densely white-spotted.
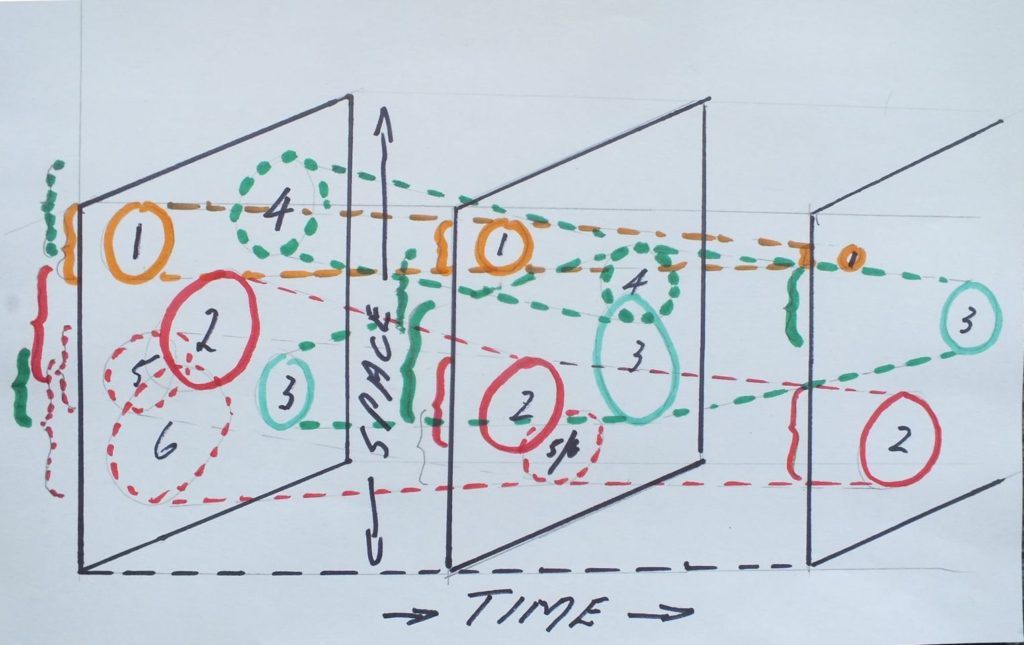
Introduction After writing Haworthia Revisited in 1996, I became aware of just how inadequate readers seem to be to the task of assimilating all the available literature on Haworthia, in the botanical and intellectual climate in which we live. It seems as though the more information we have the more confused we become. In order to generate the material needed to disprove or fortify my classification hypothesis, I have spent a further considerable amount of time in the field and in cultivating plants from seed. Unfortunately the editorial support and speed of publication has not kept pace with my own effort and much of my writing and my evidence is still in manuscript form. This short essay was therefore to put forward only a little more evidence to show just how complex plant species are – not necessarily only in Haworthia.
In my first Handbook (1976), I anticipated H. maraisii and H. magnifica to be separate species – the former west of, and the latter east of Heidelberg, Cape. In the second book (1982), I felt that I was dealing with a single species and referred to a population just west of Heidelberg as H. magnifica var. magnifica (instead of as H. maraisii). I did this transposition quite deliberately in order to suggest that the distinction between the two species was very arbitrary. In my later revision (1999) I separated H. magnifica and H. maraisii again. The rationalisation is given there and I just need to explain that I thought this was a better way of communicating the nature of the variable populations attached to each of two elements. It was then evident that populations relevant to magnifica and maraisii were proving to be more radically different than a single species hypothesis could comfortably accommodate. Also new evidence was accumulating for the nature of:
1. H. heidelbergensis, which was not even included in the previous debates. 2. H. floribunda, also proving to be more variable than predicted. 3. H. mirabilis.
The overall relationship of these three species is far closer to both H. magnifica and H. maraisii than even my original conservative approach suggests.
In the essay which deals with H. floribunda I make two points particularly relevant to this article:
1. The geographic facts, viz..”Westwards (from Gouritz River), the Swellendam area seems to provide a vegetation interval which H. turgida seems to bridge with difficulty, and which also marks some kind of a break for the H. magnifica/H. maraisii complex. This divide may be a function of inadequate collecting or the lack of adequate habitat”.
2. The populations known to me. To quote from the floribunda manuscript “There are three more collections to be considered:- by Denis DeKok near Swellendam (Figs.25a, b & c MBB6644), about 10km west of Swellendam (MBB6861 not illustrated), and from the farm Rondeheuwel south of Stormsvlei (Figs.26a, b & c MBB6882, (Bayer in KG326/71). This latter collection has previously been reported (Haworthia Handbook 1976) under one of the populations intermediate with H. mirabilis, and is cited in Haworthia Revisited under H. maraisii. These populations, as well as that southwest of Heidelberg (Figs27a, b & c MBB6663, Bayer in KG107/74 – cited under H. magnifica, Bayer, 1999) confound the issue enormously. Thus we actually have four populations which cannot with confidence be allied with either H. magnifica, H. maraisii or with H. mirabilis. The reality is that neither H. heidelbergensis nor H. floribunda can be excluded from the consideration of these populations. The case for each “species” needs to be dealt with separately.”
Primarily illustrated here are plants from the population MBB6644 which occurs west of Swellendam. The population is not remarkable for the variation observable there among the individual plants, and I have taken it to demonstrate that there was, and is, predictive value in my classification hypotheses. Much of my recent writing has been directed at the very weak concepts and perceptions that appear in our general understanding of “species” and their variability. This has an impact on individual credibility, plausibility and the truth with which we examine our human condition and pursue our hobbies.
Results The photographs depicted are all taken at virtually the same distance so that the size of the plants is relatively correct; with the largest plants being near 90mm diam. (The pots are 90mm square.)
The first illustration depicts a conventional form of H. maraisii from a population from north of McGregor Fig. 1 MBB6646 H. maraisii var. maraisii). The species was first described from Stormsvlei where the plants are a little larger and more tuberculate than these pictured. Actually it has become difficult to say just where this species stops and starts, and this will become obvious. There is a degree of translucence in the leaves and there is a conspicuous vein down the centre of the leaf. This is by no means a ‘character’ for the species, and an article could be written about the variation in venation as well as about this single population (MBB6646) too.
The Swellendam plants have been named as H. maraisii=mirabilis in the preceding chapter and are illustrated as follows:
Fig.2, 3 and 4 are the clones 3, 8 and 17 of MBB6644. The surfaces are a little less tuberculate than the McGregor plant shown, but the only significant difference is actually that they are slightly bigger plants, the spination is more obvious and the leaves tend to be fewer and more erect. These are also by no means diagnostic characters. There is little translucence and the venation is inconspicuous. The identification of the population as H. maraisii thus seems obvious.
However, clones 2 and 4 illustrate a narrowing of the leaf (Figs.5 & 6 MBB6644). Clone 15 is a small plant also with smaller narrower leaves (Fig. 7 MBB6644). Were its leaves more erect instead of so recurved, it could be perhaps be taken to be H. heidelbergensis var. scabra.
Clones 1 and 3 (Figs.8 & 9) of MBB6663 are from west of Heidelberg and would easily be lost among the Swellendam plants if unlabeled. In the preceding chapter the population is identified as H. magnifica var magnifica. Clone 2 of this population (Fig.10 MBB6663) is a plant with more erect leaves – but …
Clones 7, 10, 11, 14 and 16 of MBB6644 (Figs.11 to 15) have a similar narrowing and erect bearing of the leaves, not to say that the plants are otherwise identical. This condition of the more erect leaves seems to be the more general one in both populations, MBB6644 and MBB6663. This is almost the crux of the problem in the classification of Haworthia, and in my experience it is not a problem unique to Haworthia. In all of biology there is this variability that requires statistical method to establish what “average” is. When one is dealing with different growing condition, and the nuances of texture, colour, shape, leaf recurvature and number, it is in fact not possible to generate such a “mean”.
Clones 9 and 13 of MBB6644 (Figs.16 & 17) are unusual in either H. maraisii or H. magnifica. The leaves tend to narrow quite considerably, but the leaves can also be unusually narrow and erect in populations of H. maraisii as in a population west of Robertson (Fig.18 MBB6647.8).
I include an illustration of a plant from a population of H. floribunda var. dentata from south of Swellendam (Fig. 19 MBB6881), – a population that tends to link floribunda with either maraisii or heidelbergensis to expose the reality that the link extends to H. mirabilis. (Apparently in a Dutch journal a writer has suggested that H. floribunda var. dentata is a form between H. floribunda and H. chlorocantha var. subglauca! This would be a complete misrepresentation of my classification hypothesis which has been fully explained and there is no reason for such a mistake. The var. dentata is represented by many populations and Chapter 3 details the possible relationship of the species floribunda and chlorocantha without any frivolity about the varieties and forms.)
Fig. 19. MBB6881.6 H. floribunda var dentata. Oudekraal, Heidelberg.
Also illustrated is a plant from a population very close (NW Kweekkraal), both geographically and in appearance, to H. magnifica var. atrofusca west of Riversdale (Fig.20 MBB6817). I have included it here because general colouration and texture are the same and it evidences the typical round-tipped leaves of floribunda which occurs so frequently in maraisii and magnifica (see older leaves in Fig.2 or Fig.8).
Fig. 20. MBB6817.1 H. magnifica var atrofusca. NW Kweekkraal, Riversdale.
Clones 1, 5 and 18 of MBB6644 (Figs.21, 22 & 23) are small rather nondescript specimens, while is clone 64 of MBB6639 (Fig.24) is a plant of H. mirabilis var. sublineata from south of Bredasdorp. These last four illustrations depict a series of smaller plants which bear a resemblance to H. heidelbergensis. The difference in the latter is the lighter green colouration, slightly more translucence and more conspicuous spination. These are significant in the context of these collections and these photographs. However, put into the context of all the populations of H. heidelbergensis, H. floribunda, H. mirabilis, H. maraisii, H. magnifica, also H. emelyae and the total variability among these, it is impossible to stipulate difference.
The four problem populations, MBB6644, MBB6861, MBB6882 and MBB6663 are not as distinctive as suggested by the way in which I have specified them. They have also to be seen in the light of the variability of the individuals in those populations, and the many other populations which abut geographically onto them. Thus where I suggest they confound a clear difference between the two species H. maraisii and H. magnifica, this is not only how it is to be understood. The essay concerning H. floribunda will illustrate how extensive the problem actually is. These four populations can be discussed in terms of each of the species named in the previous paragraph.
One has to come back to biological variation and consider how one is to circumscribe a species in such a way as to facilitate identification. Here we have a single population in which the basic features such as colour, size, texture, spination, venation and translucence, are so variable that it is impossible to establish what the mean or average plant is. The problem is compounded by the fact that the variables can also not be broken down to discrete quantities. I have shown only the whole plant and thus the vegetative features. The flowers are less variable both within and between populations and ‘species’. It is already clear from the literature that species in the sub-genera of Haworthia can barely be separated on the basis of their flowers. To expect that the flower can then provide a ‘signature’ or character by which population variability can be understood, is fallacious. Floral character in fact must persuade one that there are less species than we wish to find from the vegetative characters which attract us to the plants in the first place. Flowering time is useful but it is also not diagnostic. Generally flowering time in the species mentioned in this article lies between September and May. It is no more useful than the geographic location of the population. H. maraisii tends to flower from March to May and H. mirabilis is generally earlier. However, to establish the actual flowering times for each population in terms of different seasons and years, and as a mean for the individuals in each population, must be a herculean task.
What I have done is to take geographic distribution as a key element in the classification. My experience is that distribution reflects the relationship between populations and between species. This is just a logical extension of the processes we should expect from speciation processes associated with genetic interbreeding, and isolation which obviates it. “Genetic drift” should reflect the probabilities associated with pollination across distance and with the nature of the pollinating agent. Similarities between populations should reflect this, as well as the probabilities of distribution by whatever agent such as wind or water. Naturally these will not be absolute, and the classification hypothesis will only be as good as the information allows, with the proviso that it is more information we are going to require if a better hypothesis is to be formulated.
What is striking in all my formal and informal communication with interested parties is the failure to appreciate what judgmental factors may underlie the vexing thing we regard as a “species”. I need to make it quite clear that I take a species to be a system of individuals judged on a very wide variety of factors – including geographical relationships and the influence these seem to have on other similar systems belonging to other genera.
In Haworthia, I do not deceive myself that these systems are as discrete as my classification may suggest. That is why I wrote that I was recognising nodes in a complex interconnected array. Someone put forward their own solution to one such array by suggesting that the intermediate element be recognised as a species. This is particularly unacceptable when one considers the full implication. All the intermediates could be recognised as species and the present species then as intermediates! The position is that my classification was undertaken as a proper revision when a considerable amount of new data was available; sufficient to consider it really representative and adequate on which to base an overview of the genus. For persons now to suggest an alternative classification without a proper and thorough insight into all that old material, and without adding adequately to the known record, will border on the irresponsible and inconsiderate of the purpose of classification.
[1] Breckenridge (Haworthiad 19:4, 2005) for an example among others, admits to being wholly lost in respect of a remark I made (Haworthiad 18:44, 2004) in respect of varietal names. I wrote “… (variety) is a useful rank only if there is no typical variety, and if it is recognized that the species name is used for all the variants both formally named AND unnamed.” I was referring to the practice in nomenclature where the description of an inferior rank (variety in this case) automatically creates a “typical” rank. It is obvious that if a species is described from limited material, it is improbable that the description will include all the variants. Thus any new material of that species may differ from the described. Formally giving this new material a name should in fact broaden the initial description. If the ramifications of this fact are not apparent to the reader, then no amount of writing and explanation will lift the fog of misunderstanding. ♦
In a very interesting book by Stephen Gould entitled “Rock of Ages”, in which he propounds his principle of NOMA – non-overlapping magisteria. This states that science and religion should not be confused nor mixed.
So this is not a confession of confusion – you do not confess to what is obvious. It is an admission, and an admission can be construed as an apology. But, as a rhetorical question, how can one apologize and expect forgiveness when one continues to walk the errant path?
I started to write about Haworthia to dispel confusion, and yet more than 40 years on, this confusion has not become any less. The conclusion I have come to (and I wish it was a closure) is that the prime source of confusion is simply the human condition. In mystic philosophy one can read… “Born in ignorance, we live in ignorance and we die in ignorance.”
I think that my interest in Haworthia stems from my conscious effort to dispel this primal confusion and find some of the order in my view of creation. The classification of plants suggested just one small piece of my world which was available to me, and Haworthia as one group which no one else could explain to me. What have I now learned and what contribution does this make to dispel confusion?
My courage to now say something more directly arises from a recent request by SANBI to write a synopsis of Haworthia for an E. Cape Flora. I feel that I have done that fairly successfully. The problem is now to produce a similar product for the SW Cape and this is considerably more difficult.
To explain Haworthia one has to first cross the hurdle of the present classification of the Asphodelaceae and the perceptions that Aloe, Haworthia and other genera are clear discrete groups that the classification suggests. Ignore completely the issue of whether the so-called species are clearly separable. Haworthia consists of three quite discrete groups which have no closer relation to each other than do any other of the extant Asphodeloid genera. They each have their own problems and idiosyncrasies but it is the subgenus Haworthia which concerns me most.
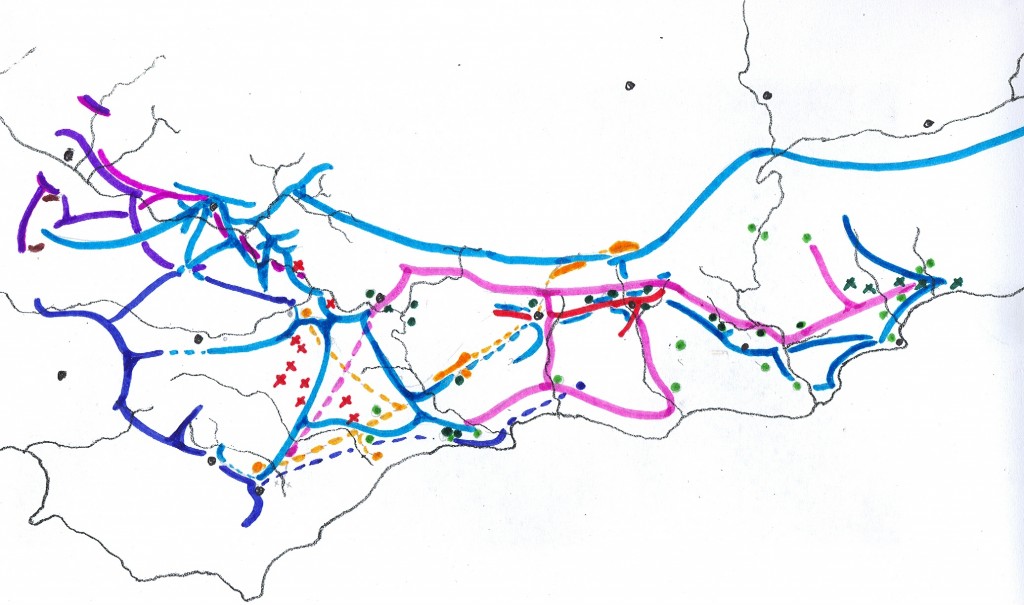
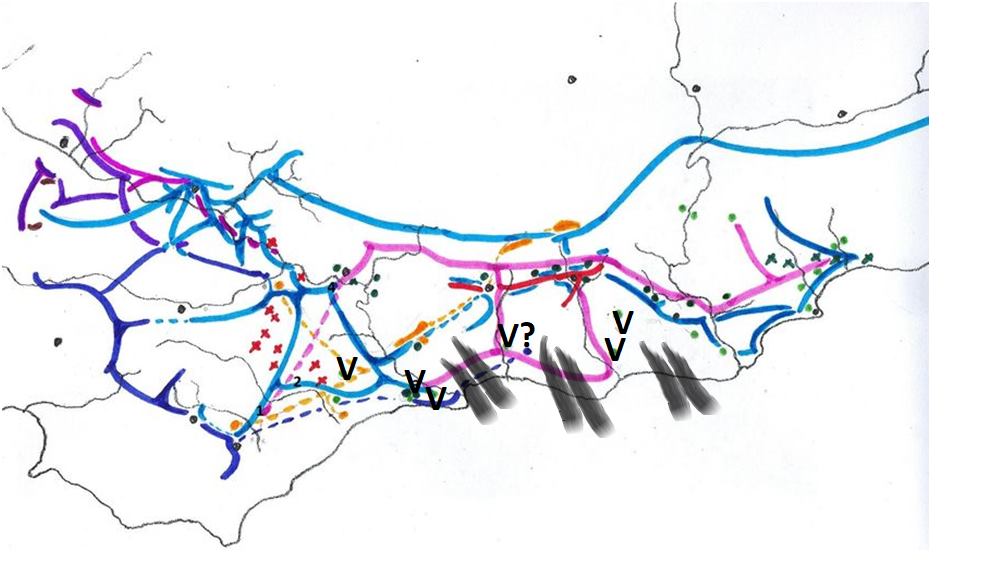
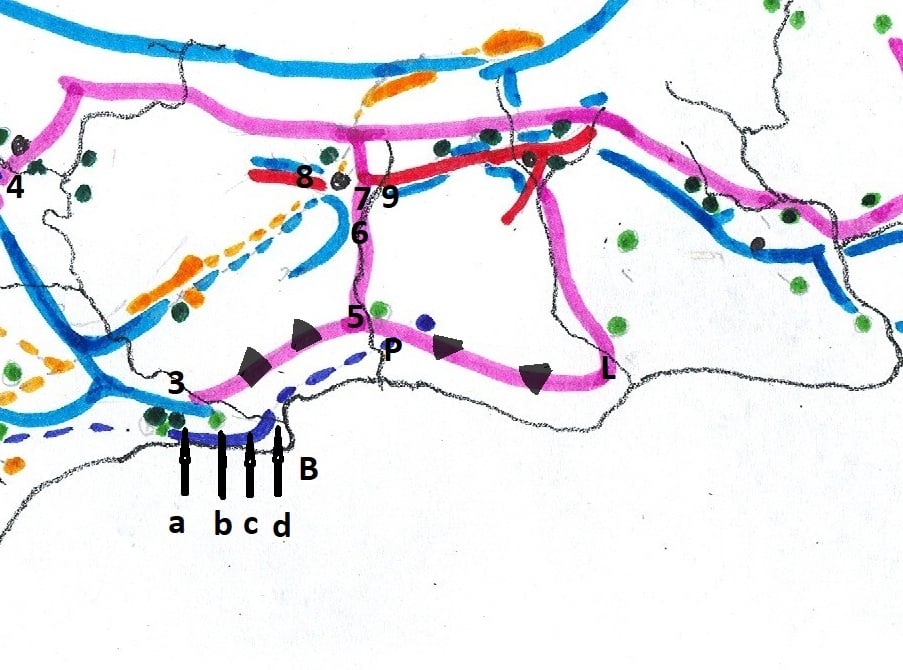
In 1975 I prepared a map to illustrate the relationship of the SW Cape species (primarily south of the Langeberg Mts.) and this was published in Excelsa (5, 1975). In this map I tried to illustrate the problem of continuity which simply makes it impossible to recognize clear closed groups of plants which we can with any truthfulness say are species. It is the reality for Haworthia, and my conviction is that the model which underlies the Latin binomial system is fundamentally flawed. It works because our ignorance clouds its shortcomings.
Map 1 Distribution of SW Cape species of Haworthia
Updating my map may show why (Map.1). It is perhaps possible to identify three main elements that are nevertheless extremely difficult to map. Very seldom do we find all three growing in close proximity and never do we find any of the components I suggest doing so either. We often struggle to suggest which of the three we are being faced with.
The names we can use for these three are:-
H. retusa – to include or cover mutica, turgida and pygmaea
H. mirabilis – to include or cover maraisii, magnifica, emelyae and heidelbergensis
H. floribunda – to include or cover parksiana, chloracantha, floribunda and variegata.
There are other elements viz:-
H. herbacea – to include or cover reticulata, maculata and pubescens
H. rossouwii –
But this is a simplification and not a solution.
What simply has to be recognized as a fact of life that this problem is not of my creation and I do not believe that the use of any technical sophistry, trick of classification or nomenclature will possibly dispel it. These “species” are as interconnected as the strands of a complex web as intricate as any a confused spider could construct.
Essentially one has a mid- to late-summer flowering group and a spring flowering group, but they are linked. H. pygmaea, in the retusa complex, actually springs from the mirabilis complex; while its counterpart mutica in the west, seems to spring from the retusa complex. Emelyae, in the mirabilis complex, seems to be linked back to rossouwii via multifolia. These are the kinds of realities one has to deal with.
The relationship of the three main elements I have mentioned differs from point to point on the map, and to describe these relationships means that one has to virtually deal with populations and groups of populations one-by-one. I have done that, particularly in the book Haworthia Update Vol. 1n and again in Vol. 2. But since the drafting of the last manuscript, I have been to the field again and found more material, which I will show now.
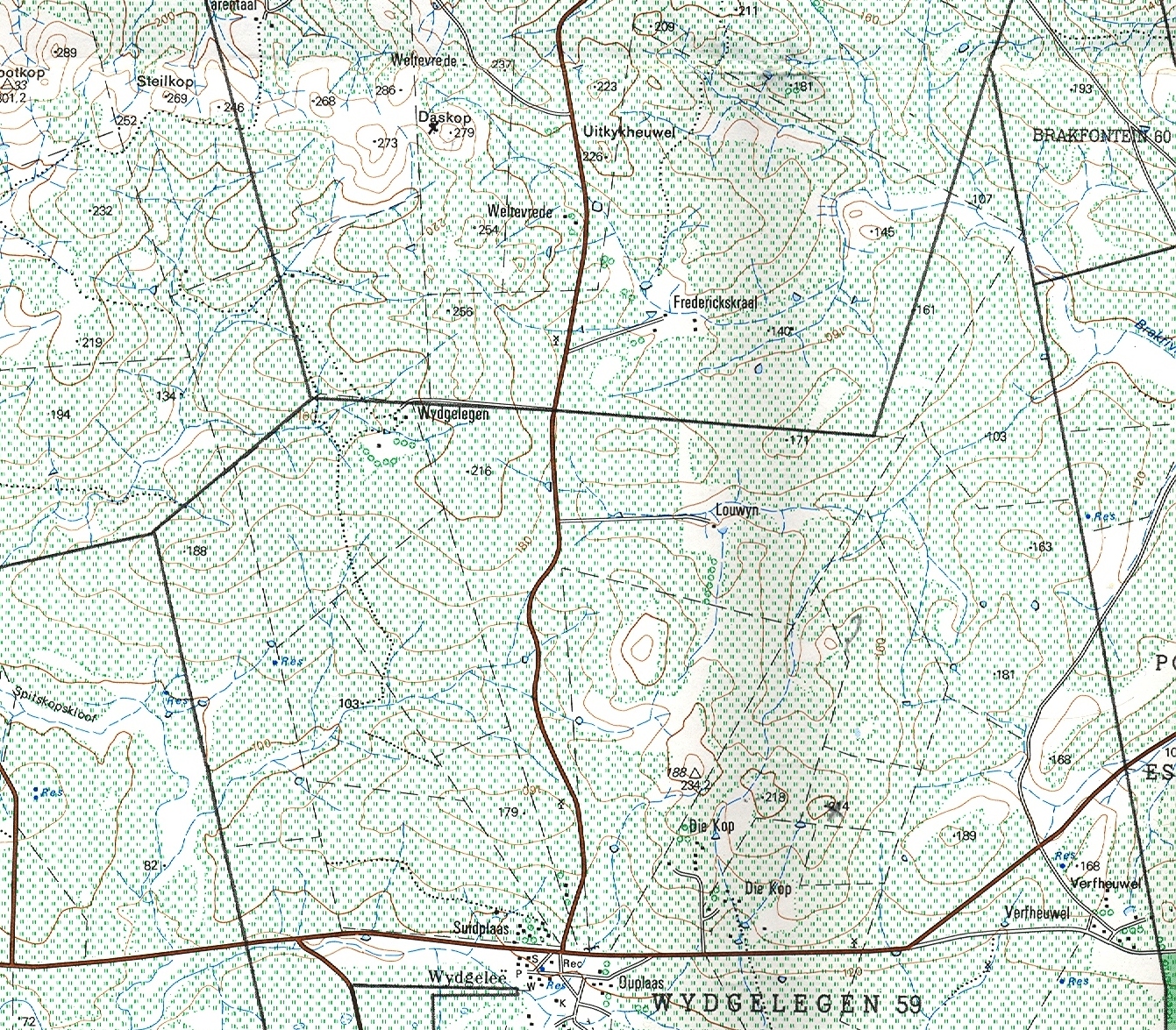
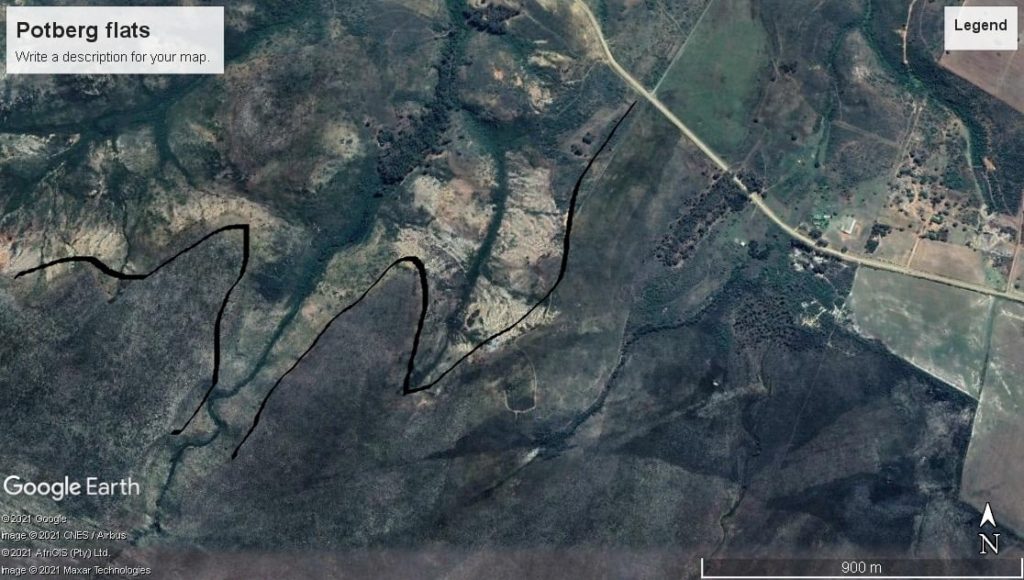
Essentially there are three areas which I will cover:- a. the southern Potberg and where the Breede River enters the sea south of Swellendam (Map 2); b. secondly Klipfontein at the northwest of the Potberg (Map 3); c. thirdly an area just west of that which I also call Die Kop, Wydgelee (Map 4). The localities are not indicated precisely on these maps but the detail is in archived and available information.
Two of three relevant populations are discussed in Chapter 15. These are ADH594 Sandhoogte, and MBB7248 Lower Breede River H. mirabilis (pilosa). I refer to CB2018 and I can now report and illustrate it myself. This is MBB7251 (Figs 2a & b) from Buffelsfontein within the DeHoop Nature reserve. In the field the plants are not distinguishable from ADH 594 and grow in exactly the same Fynbos habitat viz a very rocky and grassy streamside slope with a dry northern aspect. The illustrations are of plants cultivated and hence far greener and softer than in habitat. What is striking is the tendency to surface spination which is such a feature of MBB7248.
b. Northwestern Potberg.
Some of these populations are also covered in Chapter 15. These are MBB6890, MBB6886, MBB6882, MBB6889 and MBB6544 (figs A18…22). My comment that H. variegata occurs here too should be borne in mind. This is on the upper slopes of the extreme western end of the Potberg. There is another record of C Burger’s from further east and south of Diepkloof for which I coined the name H. variegata var hemicrypta. Some doubt will now be thrown on the wisdom of that decision. The mirabilis populations referred to above are on the farm Juliesfontein and my new records are now east of that towards Diepkloof along the Breede River.
1. MBB7487 H. floridunda. Byeneskop (figs 3a to g). No good opinion is ventured in Chapter 15 concerning the populations noted above. Those populations seem to fall in the context of an interaction between H. maraisii/heidelbergensis/floribunda and I warn of infusion of H. variegata. The Byeneskop is on the very eastern border of the farm Juliesfontein and the plants are surprisingly and confidently comparable with the typical form of H. floribunda. The leaves are generally fairly strap like and flattened to the characteristic twisted ends as evidenced in floribunda. A few plants (see fig.3f) do have shorter and stubbier suberect leaves which characterize so many collections that confound the recognition of the relevant species. This particular habitat can be described as Renosterveld on silcrete with dominance of white clay and much grass and other succulents. Similar plants were also seen about a kilometer eastwards on the verge of vegetation transitional to Fynbos.
2. MBB7492 Klipfontein East (figs 4a to d). This is again a typically H. floribunda-like population. Whereas the preceding tends to be slightly tuberculate with the rougher greyer texture and colour of H. maraisii which would suggest floribunda var dentata, these plants tend to have a smoother leaf surface. Pictures of the flower are included although of very little diagnostic significance. The habitat is a steep rocky sandstone hillside vegetated with low Fynbos species.
3. MBB7494 Klipfontein East (figs 5a to d). The plants suggest H. floribunda except that the colour of the leaves is greener and there is a tendency to translucence, or less opacity of the leaves. The white flecks in the leaves are quite conspicuous and are more often encountered in H. turgida. Their significance taxonomically is probably nil as these markings/inclusions may appear in plants as remote as H. angustifolia var baylissii on the Zuurberg, E. Cape. The habitat is again short grassy Fynbos but unusual because of the presence of finely textured white quartz rock.
4. MBB7495 Klipfontein East (figs 6a to c). While we have been looking at plants on a series of buttresses of the Potberg, these plants are now closely associated with large white quartz boulders. They are small and although very similar to the preceding, the leaves seem to have lost the characteristic rounded ends and the margins are relatively strongly toothed.
5. MBB7496 Kleinberg (figs.7a to g). This population is only about 1km east of the preceding but there is a transition to Renosterveld. The habitat is again silcretious with the typical white clay substrate. This seems to be a very erodable surface and plant cover is quite low. The plants are surprising because they depart quite radically from the floribunda-like mode of the preceding populations. They are now distinctly magnifica-like and particularly reminiscent of var atrofusca except much smaller. Curiously a population across a small valley southwards in sandstone and Fynbos is in keeping with the floribunda-like finds westwards.
6. MBB7499 Juliesfontein (figs 8a & b). We returned to Juliesfontein to look at a promising rocky slope between Byeneskop and our previous closest collection on Juliesfontein. These are almost directly north of H. variegata and as can be seen from the pictures they revert to the forms described in Chapter 15 for that area viz. there is a similarity to heidelbergensis. The more leaves are more frequently pointed rather than rounded and translucence and venation is more conspicuous.
c. Die Kop, Wydgelee.
The nearest plants dealt with in other chapters are MBB7539 at Tarentaal that I liken to magnifica var atrofusca and MBB6030 at Napky which is maraisoid with floribunda-like character (type locality for H. maraisii var simplicior with rotate leaf-tips). It is important to note that the Tarentaal plants are not uniformly larger that my identification suggests and also that H. mutica is also present at its most southeasterly locality known south of Napky. Tarentaal only just appears on the map and the mutica locality is just off the map to the upper right (northeast) corner. It should be noted that populations assigned to H. heidelbergensis occur to the west at Kathoek, Beyersdal, N. Windheuwel and terminating in the contentious var minor at Rooivlei north of Bredasdorp. I think if an overview was now made of all these populations I have assigned to heidelbergensis in different chapters and in different publications, it will have to be concluded that it may represent the pivotal element in the whole classification process. Where it was the least represented species in the period preceding publication of Haworthia Revisited it is now far better known and any hypothesis prepared to test relationships in these southwestern Cape species Haworthia will need to question this centrality. Die Kop is a set of 5-6 silcrete and ferricrete hilltops of varying size and height but otherwise not significantly different from a large number of such habitats extending from Stormsvlei in the northwest and terminating against the Potberg and Breede River in the southeast. Collections from several of these have already been reported on in Chapter 15 with the general observation that their identification is in the uncertain linking maraisii and mirabilis. I hope, in the minds of readers, that I have not excluded heidelbergensis from this milieu.
1. MBB7500 Die Kop East (figs 9a to v). I doubt if any collection has generated such reservations in my mind as this one has and many illustrations are used to express the problem. The plants are large and up to 80-100mm in diameter. They are closely spaced in a small grassy, non-rocky area which seems to be geomorphically stable. They are extremely variable and resemblances extend from badia to retusa to mutica. The leaves may be short and rounded or fairly long and attenuate, closely adpressed to the ground (recurved) or sub-erect. One notable feature is that the plants have a milky colouration to the leaves that Seven Hammer once remarked on as a feature of H. mutica. While variability is known to be great, it is seldom that it is as dramatic as this in a population of plants as they occur in the field.
2. MBB7502 Die Kop West (figs 10a to c). The plants here are smaller and more widely distributed and scattered over a larger area. The habitat is skeletal and rocky and the vegetation more herbaceous than at the eastern site. The similarity of the plants to those of 7500 is close but the variation is less extreme. There is less evidence of a mutica influence and a return to more or less the maraisii/floribunda/heidelbergensis archetype.
Fig. 9 7502 Die Kop W
3. MBB7503 Die Kop Mid (figs. 11a to i). The first impression I had was that the plants included more mirabiloid influence. Not as big as 7500 and also scattered on a skeletal slope. At one point there was a gravel patch with Gibbaeum austricola which is commonly found on such gravel patches associated with ferricrete inselbergs.
4. MBB7504 Die Kop North (figs 12a to k). The plants are smaller and there is a more evident floribunda influence. I did not think that these plants were significantly different from the many populations discussed in Chapter 15 between Tarentaal and Stormsvlei.
Conclusions: It should be very evident from all the information provided so far that more information is not producing a better picture of discrete elements. To the contrary and this reinforces the conclusion already mooted, that movement to a new view of “species” in the subgenus Haworthia is necessary, but is it possible?
Acknowledgement: Mr. Keith Spencer, DeHoop Nature Reserve. Messrs. Christine & Malcolm Wallace, Ziekenhuis. Mr. John Douglas-Hamilton, Ballyfar. Mr. Johan Groenewald, Klipfontein, Potberg. Messrs. Saartjie and Neil Neetling, Juliesfontein. Messrs. Adele & Vlooi DuToit, Die Kop & Frederickskraal. Messrs. Pet & Johannes Uys, Die Kop. Messrs. Sofia & Jurie Vermeulen, Tarentaal
I wrote a short note for Haworthiad, to explain a picture of Breuer’s new species H. fusca (MBB7507), and said “… the fact is that it is from a small population just west of Albertinia en route to another of Hayashi’s (?) species H. esterhuyzeniae, and also to Breuer’s H. vincentii. As readers we are being conditioned to accept that there are many kinds of species such as biological species, morphological species, taxonomic species, good species, bad species etc. etc. so a Latin binomial could mean anything (and the word ‘tautology’ has been added to my vocabulary). Botany needs a sensible and practical handle to a squadron of populations from between Albertinia and Great Brak. I would gladly supply this if somehow I could be assured that the act was not seen to be the clown’s contribution to the circus.”
Without any assurance, but with the encouragement of Stirling Baker, I am going to try and produce an explanation.
Put very bluntly and without any apology to a group of people who definitely deserve better, my life experience is that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well. I have already written around the subject a number of times and do not want to repeat what is not necessarily true other than the contribution these thoughts have made to my personal psyche.
In this contribution I am discuss, illustrate and then propose that there are just two species, H. retusa and H. pygmaea in a complex where presently more than nine species and varietal names are being used. I do this in consideration of all the populations of Haworthia known to me in the winter rainfall biome. Thus I recognize the need to rationalize species like H. mirabilis (which will then absorb H. maraisii, H. magnifica and H. heidelbergensis, and H. retusa (which will absorb H. turgida. There is a major problem in that the populations indicate three species in the west, viz. H. mirabilis, H. retusa and H. mutica but these appear to fuse or morph to two in the east. My past treatment of species and varieties like maraisii, magnifica, acuminata, dekenahii, argenteo-maculosa will bear witness to the nature of the (my) problem.
Gist Charles Craib includes a provocative chapter on taxonomy in his marvelous book on The Grass Aloes of the South African Veld (Umdaus Press). Not so curiously, John and Sandra Burrows voice many shared sentiments in a similar grand book on the Ficus species of Southern Africa, Figs of Southern & South-Central Africa (Umdaus Press). These books coincide with my recent experience with Drosanthemum taxonomy to which I have been peripheral since I first tried to collect D. speciosum and its allies. An account of this experience is in manuscript, and like my experience with Oxalis already published, should really be read by people with taxonomic aspirations. Of course they will not, as neither will taxonomists do who seem to already hold very superior individualistic opinions about what species are. There is absolutely no doubt that there is a very commonly held view, that what constitutes a species is based on opinion only.
Perhaps just a single illustration will indicate the kinds of arguments one has to swallow, or refute, if ones perceptions are being threatened or distorted by the decisions which are made and the conclusions which hang on them. In an article which “officially” relegates yet again the single species of Poellnitzia to some other genus (thanks be at least this is within the ambit of the aloe group of genera – the genus Astroloba) some justification is made that … “the resemblance between the flowers of Poellnitzia and the bird-pollinated genus Microloma (Asclepiadacea) is particularly striking”. The author of this really curious statement took me to task when I pilloried it, because there is a publication which conclusively shows that sunbirds do gather the pollinia of at least one Microloma on their tongues. But there are several species of Microloma and I am quite sure that they are not all bird-pollinated and besides, that is not really the point. Here I want to illustrate this time the flowers of Astroloba(Poellnitzia) rubriflora, Microloma sagittata, and M. tenuifolia and that ask readers themselves to decide if the similarity is indeed “particularly striking”. (See figures 1, 2 & 3). The arguments are no different in rationale to those of Charles Craib who treats Chortolirion as a grass Aloe because it meets his particular approach. Whose opinion do we need to follow?
Charles Craib’s decision is based on the usefulness of a classification in respect of his needs. He has kindly sent me a recent manuscript discussing the merits of a new species of grass Aloe that he recognizes from his own knowledge and experience is a product of poor taxonomy. This brings me to the conclusion that taxonomy is not science as I have so often tried to say. Latin names are simply nouns and all that is necessary is that people using names know to what is being referred. A simple example is the fiasco of the name change from Aloe bainesii to Aloe barberae. After 40 years or more where all and sundry have happily used the name bainesii,somebody discovers that another name precedes it by the thickness of a sheet of paper. Aiding or compelling this change in my attitude, I have to admit that my own classification of Haworthia no longer really meets my needs and in the climate of taxonomy that I experience, I am not sure what should be done. Thus in this article I am going to examine what I consider to be a good example of the problems any taxonomist who ever gets a good enough sample will eventually face. The question will have to be asked – are species real?
The area under consideration This is the area east of the Goukou River which bypasses Riversdale, then to south of the high Langeberg Mountain range and reaching Great Brak in the east. The southern limit is virtually the N2 national highway between Riversdale and George, although my sampling drops south of the N2 approaching the Gouritz River from the west, and dropping south to the sea east of the river. The altitude is from sea level to about 250m. The rainfall is moderate and generally falls in winter, although it would be sensible to note that the area is really a westerly extension of the all-year rainfall area forested coastal strip between Port Elizabeth and George. The vegetation is coastal renosterveld although there are areas of Karoid Valley Bushveld especially along the river valleys. The geology is very complex and plays a critical role in plant species distribution and occurrence because the soils are very skeletal. It is often the case that Haworthia is associated with exposed rock where biomass potential and vegetation competition are low. There are five main geological groups represented in the greater Southern Cape area, and the study area is different in that it is dominated by a geologically differentiated formation of the Bokkeveld shales – different to the same formation which dominates west of Riversdale. A subtle factor is that this eastern formation produces, as a weathering product, a range of clays and minerals significantly different to those from the western formation. The eastern part of the study area is dominated by rocks of the more recent Uitenhage formation; while south of the N2 (outside the study area) the formation is the most recent Bredasdorp formation.
I asked Jan Vlok for his views on what may make the area ‘different’. He replied… ”Something I have noted repeatedly is how important river drainage areas are in determining vegetation units and pro parte also those of localised endemics. I have been wondering about the issue why river drainage basins would be so important; and currently think they reflect the complex interaction between several factors – geology, rainfall, landscape of area and in thus the history of the evolution of the area. Sessile organisms – especially plants with limited seed dispersal abilities are obviously strongly affected by these factors.
In your area of interest we have one very important role player – the Gouritz River. It tended to obliterate everything in its way cutting away habitat at a relative fast rate, thus creating many new micro-habitat sites for species to establish from the north and some coming in from the east-or-west. It’s a real “bastard” area with hybrids swarms of e.g. Tritonia and Watsonia species rather common. I guess they are “species in the making”, yes, they look very different from other populations but with no clear cut morphological characteristics (the latter painful from a taxonomic point of view, but great from a functional evolutionary-ecological point of view). Note here that less sessile organisms – especially with wind dispersed seed, e.g. orchids and for that matter asclepiads are less restricted from re-colonising or finding suitable habitat in adjacent river systems. So e.g. Duvalia immaculata has much more freedom to hop from one habitat, over the watershed into the next system, than any of your Haworthia species has. Those have to do the slow march, finding it difficult, but not impossible to cross the river or watershed, and much more slowly. This implies that in the Great Brak to Riversdale area, an east-west dissemination of Haworthia populations has been relatively slow – selecting for numerous micro-habitat specialists, but perhaps each population representing nothing more than parts of one large meta-population.
East of the mighty Gouritz we have the Brakke drainage system (Groot & Klein Brak Rivers). Much smaller & slow cutting into the landscapes, thus greater stability in micro-habitats – added is slightly higher rainfall and certainty of rainfall. Thus no surprise that some populations have evolved to more discrete entities, you call them Haworthia parksiana and H. kingiana.
West of the mighty Gouritz we have the Goukou drainage system. Clearly much more stable as in cutting away the landscape than the Gouritz, but still supplemented by some summer rain. In terms of vegetation mapping I found the area really difficult. Had to look at every corner as I could not predict what will occur where. The closely related sister to the west, the Duiwenhoks has been equally challenging, except that things started panning out better. I guess its related to less reliable summer rain, thus summer drought stress. It is also a less might river, with an odd geology – suddenly conglomerates reappear. So in the Duyvenhoks plant populations start taking on unique characters – as well as do the common and dominant species of the plant communities they occur in.”
Jan Vlok confirms my opinion that the only real factor in the biogeography apart from the complex geology at Great Brak, is the Gouritz Valley. It is mainly the geology which seems to impact on H. parksiana and H. kingiana. The river does not affect either H. minima, or H. turgida. There is some doubt about the interaction and variation of H. chloracantha and H. floribunda across the valley. The populations (splendens, fusca, esterhuizenii, vincentii and argenteo-maculosa are in my opinion a series of consecutive populations which can be regarded as a continuum unbroken by the river valley. There seem to be very few species or any other biogeographical evidence to suggest that Haworthia has been impacted on.
The Haworthia populations Those considered here refer only to the species of the subgenus Haworthia and exclude H. variegata, H. floribunda and H. chloracantha of that subgenus. One must assume that by this stage, most of the relevant populations are known although I cannot claim to have personally explored the full potential of the area. Altogether I know of 29 populations which need to be considered in terms of their probable taxonomic affinities. They are listed according to my interpretation of the present available nomenclature as follows:
Haworthia turgida (Haworth 1819) Draaihoek, N Draaihoek, Valsch River, Wydersriver, Ouvloere, Die Eiland, Gouritz bridge, S Gouritz Bridge, Brandwacht.
Haworthia pygmaea (von Poellnitz 1929) Herbertsdale, Dumbie Dykes, Mosgas, SW Klipkop, Klipkop, Great Brak, N Great Brak, W Great Brak. (The vars crystallina and major (Pilbeam 1983) are not population based entities).
Haworthia magnifica var acuminata (Bayer 1976) West bank Gouritz at Melkhoutfontein (three separate populations), Vleesbaai.
Haworthia magnifica var dekenahii(Smith 1944) Draaihoek.
Haworthia magnifica var argenteo-maculosa (Smith 1945) Cooper Siding, E Cooper Siding, Humor
Haworthia magnifica var splendens (Venter & Hammer 1998) Soutpanskoppies, Snymanskraal.
Haworthia magnifica var esterhuizenii (Hayashi) Aasvoelberg.
Haworthia magnifica var vincentii (Breuer) Welgevonden.
Haworthiamagnifica var fusca (Breuer) Paulsfontein.
These populations are generally isolated from one another to some degree and it is evident that there are four taxonomic units represented by single populations.
A taxonomic species overview Of course the two words “taxonomic” and “species” used together is tautological as taxonomy is the classification of biological entities of which species is the prime unit. We have to consider what could now be species in terms of some definition, and in terms of all the other populations both inside and outside the study area. In this case there are a great many elements which fall in the same class within the same subgenus of Haworthia. I am not aware of anyone who has really addressed the issue of what comprises a species in the family Aloaceae and certainly not in the genus Haworthia. Breuer did essay a “concept” of Haworthia species, which was in fact a listing of species names for the genus and had nothing to do with questioning the nature of those species. This is in my observation a critical issue in most taxonomic work and it is the subject of many learned publications. I have already made my voice heard on the subject on many occasions and am not going to repeat any of my misgivings other than to say that I want to discuss the above populations in respect of a definition for species which I have fashioned for myself from experience and from a very wide literature.
This definition is that species are dynamic fractal systems of living organisms which are morphologically, genetically and behaviorally continuous in space and time. My attitude is that species have to be considered as phenomena in a creation and that these are spread in space and change with time. When assessing any plant as a species, one has to ask where it comes from, what its attendant companions are, what their variables are, and what their spatial relationship is. One also has to ask, what is all this in relation to geographical considerations including geology, topography, drainage systems, climate, vegetation and other plants of tangibly different families and genera? My experience with many taxonomic works and workers is that it is extremely difficult to approach the subject without extreme caution and also expect a considered response. While I have touted the view that classification is science, it most definitely is not managed in that way. It appears that there are two completely different needs that are met in classification. One is the purported one of botanical science, practiced mostly by herbarium based botanists or in present time, by institutionalized “gel-jocks” trying to lay bare the mysteries of DNA. Both are handicapped by the material available or sampled for their work, AND by their field expertise. The second need is that of the collector who simply needs a name by which to communicate with sources of plants, with other collectors and to have some measure of the difference in appearance of the plant from others he/she knows. Thus both needs will have to be met within the limits of the material known, seen, examined and experienced, and according to the competencies of the scientist, botanist, chemist, collector whoever happens to need an appellation for a plant. Taxonomic accounts and revisions are simply a product of the competence and experience of the writer limited also by the extent to which material, seen and sampled, matches or represents the reality. The term reality also has a special significance and one can rightly ask if this creation and its phenomena have any reality.
The evaluation I do this evaluation in the light of a synopsis of Haworthia which I submitted to the SA Biodiversity Institute for a flora of the Eastern Cape. That synopsis generates the question of how the species of the Western Cape should then be summarized. Quite fortuitously a parallel, although minor, problem seems to be resolved. In about 1976 I suggested that the flora of the Cape would never be understood until it was recognised that the true biome nature of the vegetation and the present-time Cape Flora was that of a Winter Rainfall biome stretching from Southern Namibia to the Eastern Cape. This now appears to be reflected in current botanical thinking where the Succulent Karoo is seen to be intimately linked to the Fynbos. The impact of this is that one really needs to re-think how species elements of the Southern Cape, are linked to those of the Little Karoo. Fortunately this does not really touch the elements I will discuss in this article although there are some peculiarities. For example, there is H. parksiana, H. kingiana, and Duvalia immaculata which are endemic to the Mossel bay area, while H. minima leapfrogs the Langeberg to occur in the Little Karoo.
What is more important is the present classification (of mine) which links most of the elements I am dealing with, to H. magnifica that is in the Riversdale/Heidelberg area. The other principle elements outside the study area bearing on the discussion are H. retusa and H. mutica.
Outside of vegetative physical appearances, there are only two really tangible pointers to relationships. One is population structure and co-existence (do the populations share close habitat or not), and the other is flowering time. Physical appearance is fairly helpful in that there are quite notable differences. There are plants which are generally light coloured and which are either adapted to steep rocky habitats as clumping cliff dwellers or are generally more solitary and adapted to level habitats. These are spring flowering. Then there is a set of populations in which the plants are very seldom in cliff habitats, are dark coloured and flower in late summer.
The first spring flowering group comprises (for the purpose of this article) H. retusa and H. turgida. There is no doubt in my mind that these two are essentially the same species as they never co-exist. The actual situation is very complicated and the recognition of H. mutica as the western representative of H. retusa and which can be said to co-exist with H. turgida, is indicative of that.
The second summer flowering group is even more complex and it is possible to suggest that H. mirabilis as presently constituted, should be enlarged to include H. maraisii, H. heidelbergensis and H. magnifica just west of Riversdale. In the context of biomes I would be rational to suggest that such treatment would need to consider also the incorporation of H. emelyae in view of its possible interaction with H. maraisii.
In respect of the study area it is thus evident that we need to look primarily at flowering time as all the populations are essentially in the light-coloured ambit rather than in the smaller dark green one of H. mirabilis. I do not imply hereby that this is inarguable, but it is evident that we need to understand that in the west we have a triumvirate of possible species with two distinct flowering times, whereas in the study area there seem to be two main elements which are both essentially spring flowering.
How do different people see the situation?
A. Evaluation and observation by Gerhard Marx Both dekenahii (Draaihoek) and fusca flower more or less together with retusa and emelyae (except major) in September-October. Both are in fruit in early November. In cultivation they also flower in March-April but I have never been able to visit them at that time in habitat to see whether they do the same. I would think so.
Splendens (Snymanskraal/Soutpanskoppies) flowers generally (late Dec. to early February). It is the one and only flowering spell for splendens.
Argenteo-maculosa and (its ‘variant’) vincentii also flower together with dekenahii. In Grahamstown argenteo-maculosa flowered for me throughout summer and even into winter. They seem to do the same for Steven Hammer in Vista. But I think in the wild they also have two flowering spells, one in spring and another in fall.
I seem to remember the flowering periods of acuminata are exactly the same as for dekenahii.
Pygmaea from Great Brak and Mossgas flower exactly together with dekenahii , but argenteo-maculosa (Humor and Cooper Siding) flower later. They are still developing flower buds at the moment.
As mentioned above, H. emelyaemajor also flowers later than the ‘picta’ types and ‘breueri’. Major is only starting to flower now, while all other emelyae are just about done releasing seeds.
Thus at present I see the relationships as follows:
H. splendens (Soutpanskoppies/Snymanskraal) is a good species and stands isolated in terms of its flowering time, chunky and glossy leaves and slower growth.
H. dekenahii can perhaps also stand as a seperate species and fusca is definintely a variant, distinguished from the dekenahii by its red-brown colour (meaning no doubt the “rusty”-like difference) greater tendency to offset and geographical separation.
H. argenteo-maculosa – (which I feel it is such an inappropriate name!) – is another separate thing, although it shares superficial characters with splendens on the one hand and acuminata and even turgida on the other. Apart from different flowering time to splendens, it is much ‘softer’ and grows twice as fast and is also smaller with more acuminate leaves that are spotted along the sides. H. vincentii can probably be seen as a variant of argenteo-maculosa as they are so obviously similar, except that vincentii has fewer and longer leaves that are less heavily flecked and it is even more ‘tender’ (softer) than argenteo-maculosa.
H. pygmaea is fine as it stands, with Great Brak/ Dumbie Dykes/ Rooiheuwel/ Mossgas/ Herbertsdale all sharing basically the same characters and flowering time.
Argenteo-maculosa (Cooper Siding and Humor) plants are different in having heavier flecked and chunkier leaves and to me it is a toss of a coin whether they are seen as a variant of dekenahii or pygmaea.
Similarly I think acuminata can just as easily become a variety of turgida or dekenahii rather than of magnifica.
Another option is to keep dekenahii and acuminata variations of magnifica and add fusca, argenteo-maculosa and vincentii also as varieties. But then it is a case of giving them all the same ‘status’ where I feel that fusca and dekenahii are ‘unidentical twins’, while argenteo-maculosa and vincentii are an almost-identical-yet-different duo.
The above decisions are really based upon subtle factors and can be decided by tossing a coin. What I feel is important, is that splendens should be separated as it does not fit in well with the dekenahii–pygmaea bunch or 100% with the magnifica from Riversdale. In terms of flowering time I can see the logic of including it with magnifica and atrofusca, but then dekenahii and acuminata should be removed.
The analysis closes with the words … ”Wait, let is that me sign off before I write myself into a corner.”
A1. Reaction comment by M B Bayer There are two important considerations. One is that Gerhard does not profess to be a botanist or to be too concerned with taxonomy. The other is that he is assessing the situation in terms of his specific life experience in relation to all the variables I mention earlier. He does not see to have considered the many turgida populations and their variation, the Vleesbaai population of acuminata, and another important single population of pygmaea at Herbertsdale, and neither does he relate these to a greatly compounded problem when all the relevant known Western Cape populations are added.
My views are that splendens is significantly different because the two known populations are associated with ochre deposits. Fusca is on a less iron/ochre rich site. Dekenahii is limited to very few plants (and there were ever many) on shale. Can these very small and highly localized elements seriously be regarded as species in terms of my definition? Argenteo-maculosa and vincentii are associated with table mountain sandstone. The argenteo-maculosa populations are also linked to sanstone while the acuminata populations are linked to old river terraces and the Vleesbaai population is linked to sandstone again. The Hebertsdale population is related to Uitenhage conglomerates and all the other pymaea’s to an array of substrates around Mossel Bay, Little and Great Brak.
Turgida occurs on steep rocky river banks (shale cliffs) and in two places can be said to coexist with pygmaea or dekenahii (Dumbie Dykes and Draaihoek). It is not outside the realm of possibility that the differences in vegetative appearances are responses to different habitat and substrate. It should also be noted that the turgida var pallidifolia was described from Draaihoek too. I saw such clones at the site when I visited the area in ca 1972, but on a recent visit no such light coloured clones were apparent. The population further down the Valsch River is also highly patterned as are those at Draaihoek. It should be noted that there used to exist another population to the north of Draaihoek which I would have equated with H. turgida var longibracteata from the Heideberg area or lower Goukou River at Riversdale. The population at Wydersriver is not known to me apart from photographs of G.G. Smith and also seems to fall outside of the ambit of turgida var suberecta as I classified all these eastern populatons of turgida. The one population which I recently examined was on the Wyders River northeast of Ouvloere (Albertinia district) where the plants definitely suggest affinity with vincentii. Examining my photographs, which are reproduced with this article, I thought there was a remarkable similarity between clones of fusca and argenteo-maculosa at Humorum or Humor.
Of course one can adopt a different species definition and even suggest that simple superficial resemblance of a minimal sample is adequate for an opinion forming process. It is certainly difficult to even attempt to examine how these populations became isolated or even to ponder exactly how isolated they really are in relation to pollination mechanisms and seed dispersal.
A2. Response by Gerhard Marx to the re-drafted manuscript I do not know whether to feel flattered or embarrassed that you took my loose comments upon the splendens/pygmaea/turgida group so seriously. Only a few minutes after sending the e-mail away, I wanted to add more questions and comments that partly contradicted the ones I sent.
It is really a complicated puzzle, particularly if one takes flowering time in consideration. When I look from dekenahii to that chunky atrofusca and to fusca and splendens, I do feel that they really neatly fit as varieties under the same species. But then splendens‘ flowering time removes it distinctly from the others and I do not know what to do with that fact.
I feel that then splendens might just as well be thrown in with H. emelyae because of the obvious similarity in appearance. The Moerasrivier/ Saffraanrivier area form of emelyae even has almost the same ‘pimpled’ upper leaves and some glossiness. However, I must say that I do like your proposal to have the whole bunch under H. pygmaea. From a collector and non-scientist view as my own, it feels good and definitely an improvement upon he dispensation as in Haworthia Revisited.
There have been brief moments when I even wondered why they can’t all be retusa. When I first saw H. fusca in the wild I was convinced it is a form of retusa. And after comparing it in cultivation with dekenahii, I became convinced that it is closer to dekenahii. Looking at your Herbertsdale pygmaea photos, I can clearly see shades of acuminata in them, as well as argenteo-maculosa (particularly Cooper Siding) and, of course, Mossgas pygmaea. This supports the idea of having them all as varieties of one ‘superspecies’. But I can find it easier to see esterhuizenii linked to pygmaea than retusa, so that is why I like your idea of having pygmaea as the umbrella species.
To my mind there remains the problem with splendens‘ flowering time that matches up with the mirabilis and atrofusca/magnifica group. And mutica, of course, is again linked to the pygmaea group in terms of flowering time. Particularly if you enlarged our concept of the mirabilis complex with new introductions like ‘shaggy dog’ and plants from ‘Die Kop’ etc.. Maybe splendens could even fit in there.
I really don’t know. That is why I often said that at times I have some sympathy other taxonomists that are inclined to call each differing population (or plant) a separate species ! It is just so much easier.”
A3. Response by M.B.Bayer To this, my response was…”it should not be found to be embarrassing – I think this is perhaps what influences taxonomy and why you do not want to be seen as a taxonomist. You are a taxonomist – we all are. Science and especially nomenclature have put a shroud around the whole process to suggest only those really in the know can make a good decision! What you have not done – and it is not a failing – is to consider the variants of turgida. Flowering time has to be considered with the same reservations as any other character and in the end I think it is an intuitive judgment which has to be made and not a product of scientific reduction and pedantry.
The inclusion of emelyae just adds more problems because there is a whiplash onto heidelbergensis and paradoxa via major and multifolia – and there are no clear answers to that either. The variation just folds back on itself. Hence my references to “fractal” – endless variation never repeating itself! My end point looks as though there is no solution except by consensus in which contributors also recognise intellectual and experiential boundaries?”
B. Comment by anonymous knowledgeable collector “About your ms. I understand that I am looking at it as an under-informed collector, and it makes me realize how completely I have failed to keep up with current writings on haworthias. But like our old friend has demonstrated time and again, ignorance is no obstacle to the collector in offering opinions.
I do agree with you that the populations between Albertinia and Great Brak need – to use your words – a sensible and practical handle. Your ms., however, has not persuaded me that it offers such a handle. From my point of view, to reduce these populations to two species obfuscates rather than clarifies existing population differences. To base your conclusions primarily on flowering time sets aside the differences in physical appearances which have so fascinated us and which have seemed so important in the designation of species. But that must be your objective? The ms., however, does not offer, at least to me, sufficient argumentation that flowering time can be used to solve, or resolve, the Albertinia-Great Brak population differences. And almost as an aside, I must note that, while you write that it appears that there are two completely different needs which are met in classification, you are willing in effect to set aside the collectors’ needs in order to reach a conclusion. The dual naming system which some of us now use because the existing formal system does not meet our needs is bad enough; no need to make it even more prominent by encouraging it.
B1. Response by M.B Bayer My response to this was…”May I incorporate your opinions as I have done with Gerhards?
I think the point which you may miss is that essentially the naming system is for botany, in fact all of biology, and there should be only one goal viz the recognition of living systems. There is absolutely no reason for collectors to insist that Latin binomials are required for their activities. Why on earth cannot collectors use H. pygmaea “splendens“, H. pygmaea “Herbertsdale”, H. pygmaea “vleesbaai”, turgida “pallidifolia“, turgida “sub-erecta” turgida “Wydersriver” etc. What more do they need? It seems fairly obvious to me that there is just a veneer of pretence that the latin names mean something more and which they aspire to? There is no way that anyone else is going to arrive at a classification of these populations (considering all of them) which is more meaningful. What we see in Haworthia is that physical appearances, as variously we see them, are not aiding identification or classification and flowering time does offer a view of a system which reflects over the entire series of populations. But in nortieri I can show that flowering time does NOT do so!
I cannot see how you say I am setting aside collectors needs. I am a collector and I want a system that is true and reflects what I know and experience. This is why I started fiddling with the names in the first place. Why have we so often considered that place names should be used? It is all very well and good for collectors with plants bearing names as they were obtained, to insist that those names have meaning and are immutable as opposed to those who have unnamed plants for which they need a designation.
I spent some time in the field with nortieri. This is a species which is grossly under-represented in collections in relation to field abundance. It must be one of the commonest and most widespread in terms of actual occurrence. We have the names globosiflora, devriesii, albispina, pehlemanniae and agnis as well as nortierimontana and gifbergensis and it is the most under-collected species of the genus. We do not have names for Arizona, Ouberg Pass, Opdie Berg, Moordenaarskaroo, Hex River Pass, Blinkberg, Breekranz, Nuweplaas, Dwasriver, and a few others which could be said to be different. How many more names are we going to generate based on existing approach? This does not worry the collector who is glad to have new names entering the system slowly enough for him to afford to add them.
What I am now saying is that there has to be an agreed system and that it is up to the collectors themselves to come forward with some sensible proposals. As a group they definitely do not have any idea of biological systems or even much grasp of nomenclature, the controls it puts in place and the problems that they also creates. I think I have said elsewhere that I am perceived to be the “trouble-maker” when in fact I seem to have spent my life pointing out that we are basing our hopes on method and system which offers no hope of stability or agreement.”
B2. Collectors reply The collector responded…”I fear that at the present we are far apart both in our individual thoughts and how we understand one another. What to do? There follows several unconnected comments or thoughts of mine on your ms. But first, I don’t mine if you use anything I write, even if it to knock it down, but I would be grateful if you would not use my comments as you have Gerhard’s, with reference to me. To me, what I write are “private” views which may not be supportable and which I may change tomorrow, if not sooner. I still cannot accept in my own mind the idea of using blooming time to formulate species. It is but one factor, and perhaps not even an important one, except where there are two species growing together.
Physical differences still seem the most important to me, per the difference between pygmaea in Great Brak and splendens at Albertinia. Are you not making physical differences important when you exclude certain populations like variegata from your ms? I thought that it was lack of physical differences between the plants at Horn Farm and those near Albertinia which led you to conclude that they were one species – which is going at the issue from a 180 degree direction.
The big “problem”, I guess, is really how to deal with the populations in the area which may or may not be connecting links, and which for one reason or another do not alone deserve species status. I just don’t have an answer. But I don’t think that “blooming time” solves the problem either. Rather, it seems simply to smother the issue.
You write in your ms that there are two completely different needs that are met in classification, yet you seem to give up the one, that of collectors, in your conclusions. If we are talking about taxonomy as the basis for communication, are you not giving up the importance of communication in the combinations that you propose? And yes, there may be two different needs, but why cannot the solution for this contain a system that meets both sets of needs?
I am not wedded to Latin binomial names. We have previously shared an idea of the usefulness of including locality data in a haworthia name. Can that be done without challenging the existing classification system? Every population could be a forma, and then everyone would know in a discussion what plants were being talked about. I recognize the objections and impossibilities, but I think that it is a great idea nonetheless. Meanwhile there are not two collectors in the world who are going to agree on a system for the use of collectors.”
B3. response by M.B. Bayer My response was…”Yes I do regret that our minds do not meet. This is the problem with taxonomy – it is simply not science as I once wrote. I have by no means switched to using flowering time in the way that you suggest. I am recognising that better solution to the problem is needed. Cooper Siding seems to flower with splendens and not with pygmaea nor with Vleesbaai acuminata and nor with Humor. It is simply axiomatic that it is a factor which influences gene flow, but this is disrupted by isolation and what botanist can pretend to understand “island” speciation? Variegata,floribunda and chloracantha present a parallel problem to this pygmaea complex and the question of physical differences to the pygmaea lot is barely relevant. Would any haworthia taxonomist suggest they be treated as one?
My experience with other genera suggests that any two people, who truly look at these things in the field and grow them in the way we do Haworthia, will end up in the madhouse if they are looking for agreement. The very point I have been trying to make is that if classification is to make sense and be useful as a communication medium that it is agreement we have to prioritize. If any one thinks they have valid and strong opinions then they should express them, and they should not lose sight of the fact that Latin names belong to the realm of formal biology. What is the point in dissent which is not balanced by counter argument? In a recent journal an author has at least ten new names to occupy the place of my aristata. This is as if the use of the name aristata itself does not have the potential to generate waves according to the intent of the reader. I wrote at the very outset that my intent was a system of names within the domain of the South African flora as a whole. How will we ever get agreement if we do not even see that need? There is a huge trend to turn everything upside down using DNA technology and now I just have informed comment that standard morphological methods work better!
One should not actually express an opinion without recognising the fact that there is a physical record in the form of the herbarium collections (worldwide) which even take precedence over the plants in the field. Unless ones mind covers all those records how can valid can an opinion be? This is the reason why no two collectors can possibly agree – they are confined to the sample they hold in their minds.
You close with what I think is a fine solution. Refer to the populations by place name within a formal structure and it is that formal structure which I am trying to find.”
We did try to continue this exchange of ideas. My feeling was that it would be endless as the collector is not in fact in a position to evaluate or question any comment in respect of biogeography. Understandably he has to lean on the historic method of dependence on the evidence he has obtained from the material he has seen. Another consideration is that my decision is being influenced by what I think about the western populations where I suggest that H. mirabilis absorbs H. maraisiii, H. magnifica and H. heidelbergsensis. In response to his comments I redrafted the manuscript to explain this.
C. Comment by Steven Hammer “If you say that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well, this sounds contradictory, though one could say that farces and comedies are effective. I suppose the problem is that one is unable to take taxonomy seriously in the way its “strict” pretensions would demand. But if it works in its fashion then all one needs to do is to analyse that fashion and say it’s OK for that purpose.”
His opinions are that esterhuizenii is far greener than splendens and lacks the capacity to “blush”. Of Dumbie Dykes pygmaea he writes…”it is different – big pale, flabby readily proliferous (lots of turgida traits). Thus it has echoes of vincentii too. But you can see all of those traits in Great Brak pygmaea too.”
In respect of habitat and substrate…”I think it is likely that the difference has its impact – consider the way both western splendens types have changed after a time in cultivation. Soutpans plants look as it they’d been digesting powdered white porcelain! They did not have this look for the first three years. Much more slowly, some of the Snymanskraal plants have acquired it too, while at first they resembled many magnificas, especially those with atrofuscan aspirations.” He adds that it is possible to imagine the Mossel Bay pygmaea transported to Soutpanskoppies and getting bigger and fatter.
In respect of formalities of nomenclature attaching a reality to the entire spectrum of names which they simply do not have, Steven writes…”I love this last sentence. I can see that each of the other elements can be seen as variant of, or a link to, of pygmaea. There are so many cross-links…”
He adds…
”Where orthodox taxonomy gets in trouble is in its hope for absolutes – we are told, for example, that splendens never has spots on its leaf backs. This is not true when one sees a large enough sample. It is true that most esterhuizenii’s have many more spots, much more often, but it is also true that one of the original esters – a Parisi plant – would have passed anyone’s inspection as a western (Soutpans) splendens. If that isn’t a proof of a connection I don’t know what is! It is also true that Parisi’s plants, and their descendents, can be distinguished from the Venter-Marx plants bred from material collected on the same hill – they have more ‘silver’ and are more delicate. That lineages can be distinguished on such a minute (if trivial) level tells us something. Haworthias are very malleable in a pot. If they are that malleable in a pot they must be malleable/shiftable in habitat too.
That brings up continuity. The gene flow may once have been a continuous flow in one direction, but the river’s dried up. So now we have islands in a dried-up stream. Collectors will always want to call splendens a species – it concentrates all the glamour of Haworthia in one shiny package. I don’t know what rank it “deserves” but I suspect it is the least trivial, or the most distant, of pygmaea’s ramifications. If this is pygmaea then it’s the most deeply transformed of the lot. Or it’s got some emelyae links, which would fit, by extension, the maraisii/emelyae link. I do think that co-occurence logic is the strongest logic possible. One can also look at Soutpans and say, hmm, no trace of turgida here!”
C. Response by M.B Bayer I do not consider that if the system works in the fashion that it does, and that it is OK for the purpose, that this helps. It is only working because too little is expected of it. The scientist/botanist believes that the names are providing critical information about species but which they cannot define, and the collector is content with a handle to attach to his plants or communicate about them. My contention is that taxonomy is not providing proper scientific content.
Illustration
1. 7508. Draaihoek. Northwest of Albertinia and the type locality (the only locality) for var. argenteo-maculosa. The substrate is Bokkeveld shale and the vegetation renosterveld. The locality has always been very restricted, with few plants and very unsubstantial. Although there might have been more suitable habitats available in the past, there is very little remaining.
2. Snymanskraal. splendens. The variant was discovered again almost solely due to the persistence of R.W.Kent althouth the next locality may be where Herre originally discovered it. This habitat is quite complex. It is at the convergence of recent sands, ferricrete/silcrete and shale. The ferricrete/silcrete is in a relatively flat-lying formation rather unlike the raised inselbergs that it forms in the west. The vegetation is largely Renosterveld, but with so much sand in the vicinity, there is also Coastal Fynbos.
3. 6751 Soutpan. splendens. Very similar to the preceding habitat and the plants are also very localized in the exposed ferricrete.
4. 7507. Paulsfontein. fusca. This habitat is also on the geological convergence described but here there seems to be high level river gravel which predominates.
5. GM Aasvoelberg. esterhuizenii. Parisi and Dunn seem to have been the discoverers of this population on the low northern slopes of Sandstone hills to the east of Albertinia.
6. 7102 Welgevonden. vincentii. A few kilometers further east, the locality is also Table Mountain sandstone but a different geological series to that at Aasvoelberg. This and the previous four records are in a very simple geographic line and there really is nothing to suggest any biogeographically differences in these habitats other than the detail of substrate. In the general way that Haworthia populations display some degree of observable continuity from one population to the next, so do the plants in these populations. The actual differences may seem quite large but there are parallels at many places for this ecotypification in the genus.
7. JDV86/75 Cooper Siding. argenteo-maculosa. The habitat is again ferricrete/silcrete on an interface with sandstone. Plants west of the siding are actually in Sandstone. It appears that the plants can be found eastwards along the sandstone ridge to near Humor about 10km eastwards. I do not think anyone has really explored the differences in the plants along this ridge. H. floribunda also occurs at the same site and I have seen a hybrid.
8. JDV91/39 Humor. argenteo- maculosa. The plants are on the low slopes of another sandstone ridge which parallels that at Cooper Siding and apparently a different series. I am not aware that anyone has given these plants an individual name, but would be justifiable in terms of all the other names in the series. The plants are quite strongly marked.
9. JDV87/97 Melkhoutfontein, Gouritz Bridge. acuminata. The original locality was destroyed by road-building and these plants survived in cultivation. Populations are also known to the south and north in the bank of Bokkeveld shale which borders the river on the west bank. The plants are odd in that the leaf ends are very acuminate.
10 MBB6747. Vleesbaai. acuminata. The discoverer of this population may be V. DeVries. The substrate is calcareous limestone. It is a similar formation to that at Stilbaai where H. variegata occurs. The plants tend to be quite robust with a curious grey sheen in cultivation.
11. JDV9/135 Herbertsdale. pygmaea. Enst van Jaarsveld found these plants on a steep Uniondale conglomerate slope with H. chloracantha also present. The plants tend to be a large and flabby and not particularly attractive to grow, whereas in habitat they retain their tight soil-level leaf arrangement.
12. EA1200, 6813. Mossgas. pygmaea. This population was found by Paul Schoeman in sandstone just west of Mossel bay. It is important to note that this sandstone is the same formation that runs from south and east of Albertinia in a narrow band sloping down to near sea level at Mossel Bay. Populations 2, 3, 4, 5, 6, 7, 8, and 12 are thus along this band and its contact with younger tertiary formations.
13. ADH275. Northwest of Mosselbay. pygmaea. The geology is exposed ferricrete with river gravels. Adam Harrower found a plant with seed and grew the seedlings here illustrated. Notable is the appearance of plants with the crystalline papillae known in plants from Dumbie Dykes and from Great Brak.
14. EA1218. Klipkop, North Mossel Bay. pygmaea. The substrate is old river terrace. Etwin Aslander’s locality is now a housing estate. The plants are not much different from those in the previous record.
15. EAsn. Dumbie Dykes. pygmaea. The geology becomes very complex from here eastwards as there is igneous extrusion.
16. Great Brak. pygmaea. Various collectors have plants from here. During the early years I observed pygmaea in four different populations in and near Great Brak. They are still present practically within the town, and a short distance to the northwest where they are in gravels. What struck me forcibly at the time was the similarity to H. mutica and in one close comparison of two plants, I could only separate them on the colour of the bracts.
17. MBB7512. Draaihoek. turgida. I will not illustrate more than four populations. Essentially it is significant that we have a situation which parallels that in the Eastern Cape where H. cooperi is is general the plains-dweller and H. cymbiformis the cliff-dweller (in the west there is a morphing and fusion so that cooperi var gordoniana is the plains-dweller and cooperi var picturata and H. transiens the cliff-dweller. In the southern Cape, the separation has been H. turgida the cliff-dweller and H. retusa the plains-dweller. The relationship is less consistent than is the case in the eastern contingent and the very complex continuity of H. mirabilis with elements such as H. heidelbergensis, H. maraisii and H. magnifica (which can be rationally treated as one) suggests that H. retusa should absorb H. turgida. In practical terms H. turgida is the predominant element, and the name retusa is taken because of the requirements of nomenclatural rules. It must be noted that the complex of populations that I have discussed in Haworthia Update Vol. 2 as H. mutica var. nigra represent a part of the interaction of H. turgida and H. retusa and thus in this light would all be H. retusa.
H.. turgida, supporting the contention that it is the cliff-dweller, occurs in the Valsch, Wyders and Gouritz river valleys on very steep sites. It is proliferous, so the plants are often in large shallow rooted clumps. The leaves do not develop the very flat and truncated end-areas as do the plains-dwellers. As far as I am aware it only occurs at two localities east of the Gouritz River. One at Brandwacht north of Mossel Bay and I believe at one east of Little Brak. Only at Draaihoek is it in very close proximity to argenteo-maculosa. It is at this locality where turgida var. pallidifolia was collected. It is both curious and co-incidental that I did observe one or two very pallid plants at the time of my first visit to the site (now apparently no longer present) and formed an impression that the separationof argenteo-maculosa and turgida was actually a question of ecotypic adaptation. A population I did sample was present about 1km north of Daaihoek in which the plants were more comparable with turgida var. longibracteata and these did influence my impression. That population seems to have disappeared as the area is very disturbed (largely due to the dense hyrax population) and I could not find it on a recent visit.
18. 7105 Ouvloere. turgida. The plants here have the same rather obtuse leaves which prompted me to recognize the variety sub-erecta, but I was also struck by the change in the mottling of the leaves and the similarity to this in esterhuizenii and especially vincentii.
19 ADH397 Gouritz Bridge. turgida. This is the well-known form of turgida which once passed under the cultivar name “rodinii” for which Die Hell, up-river in the Swartberg Mountains,was incorrectly given as the origin.
20 EvJ16716 Schoemanshoek, S. Gouritz Bridge. turgida. This is very similar to the former and again from a cliff habitat. There is of course a similarity to acuminata which is only a few kilometers away on the sloping rocky banks of the river. That does vary from the pygmaea complex where I would include it now, as it also tends to be proliferous. I consider that this emphasizes the tight relationship of H. pygmaea and H. retusa (turgida) rather than supporting any reason to fragment them taxonomically as we have done in the past.
Conclusion If I now am required to produce a synopsis which makes broader botanical sense, rather than meeting just the needs of collectors, I would suggest that it is mutually agreed that only two species be recognised. These are H. retusa represented by the populations cited here by the name H. turgida and with the rider that in fact H. turgida should be incorporated under a wider acceptance and recognition of H. retusa; and that ALL the other relevant populations in the area under consideration should be treated as the single species, H. pygmaea. For the collector I would seriously suggest that the formal ranks of subspecies or variety simply be ignored. The species name is used and in inverted commas any other name that is useful to popular discussion and communication. The formalities of nomenclature attach a reality to the entire spectrum of names which they simply do not have. Names are being used to register collection, acquisition, possession and personal achievement, and are recording very little botanical information and understanding of the extraordinary nature of living things.
A prominent taxonomist was very dismissive of my opinions and cursorily explained that…” Yes, species are real, and defined well by their ability to cross freely and produce offspring which again crosses freely. This has been studied and demonstrated since 1750 or so (you may remember that I talked about this in one of our discussions on the subject), but it is naturally not easy to dive into so much work including following up several generations etc. And since related individuals are similar looking, the reciprocal conclusions that similar objects are related has unfortunately been used as being true, which it is not. And yet, species are the only natural unit in the whole of taxonomy, which to determine is the crucial point. Perhaps this sounds rather like the famous Dicta of Bessey, but after having dealt extensively over 38 years by now with species boundaries in teaching, reading, theory and practice, in different vegetation zones and many different groups, I come back to old definition first given by Ray in 1682: “Group of plants derived from common seeds, reproducing their typical features by sowing” and used by most taxonomists since.”
My contention is that this definition is grossly inadequate as it under-estimates the degree of variability that can occur within a species, not only inherently, but especially in relation to spatial distribution and adaptation. ♦
Steven Hammer, in his inimitable style, put a very fresh face on Haworthia in Cact. Succ. J. (Los Angeles) 80:140 (2008). He drew attention to the wonder of the plants in cultivation for the collector, contrasted to a reality of unglamorous scruffiness in the field as per the lens and pen of Bayer. It has fallen to my lot as a very unwilling taxonomist to reduce the fascination these plants have for me, and for so many others, to the mundane vortex of labels, their proliferation and continual amendment. The fact is whether on a label or on the tongue, a name is a part of any language we use to talk to each other; but we are not learning anything from a well-documented history and in Haworthia seem to remain lost in a maze.
The unhappy truth for Haworthia is that by the time von Poellnitz in Germany, G.G. Smith in South Africa, F. Resende in Portugal, A.J.A. Uitewaal in Holland, W. Triebner in Namibia, J.W. Dodson and J.R. Brown in USA had either left or abandoned the scene, there were any number of names that meant very little more than the Latin they were written in. J.R. Brown presented a talk, A brief review of the Genus Haworthia, to the Los Angeles Cactus and Succulent Society that was published in the Cactus and Succulent Journal of America 29:125-135 (1957). He noted the number of species and varietal names at 160 and 370 (!) respectively, arranged in 20 sections.
While J.R. Brown was winding down (his last note on Haworthia was published in 1970), I was busy trying to make sense of a two large files that seemed to form the body of a manuscript by G.G. Smith for which Mrs. M. Courtenay-Latimer had drafted a title… “A monograph of the genus Haworthia.” This manuscript comprised a collection of all current species descriptions arranged in the purported twenty sections of Berger and accompanied by many illustrations from the original publications, as well as by many of Smith’s own photographs and those of H.G. Fourcade. We know that Smith retired in a huff, but was there really good reason for his exit?
It is quite evident from the legacy of notes that the manuscript could not have been anywhere near complete. I drew attention to this in an article…”Reflections on G.G. Smith’s Haworthia collection” that was published in Excelsa 8:46-52 (1978). While stuck in the paradigm of 20 sections, he envisaged many more species and varieties and I cannot imagine how he could ever have reconciled this with the evidence of his compiled manuscript. I suspect he realized this too, hence the (self-directed?) huff.
It took me several years to absorb the specimens in the Compton Herbarium while at the same time taking Smith’s meticulous (in the sense of the literature from which it was extracted) collation of material to pieces. Putting it all back together as best I could, I eventually produced Haworthia Handbook (1976) published by National Botanic Gardens of South Africa. It was not a great work and neither was it very well-received at the time (though it has had a curiously perky afterlife once it went out of print). Charles Glass wrote to me that he was a bit disappointed (by its elliptic compressions?). Gordon Rowley was kind enough to point out many nomenclatural errors and a page of errata had to be inserted.
However, I was actually committed to helping Col. Scott write a formal revision as I had no aspirations to do so myself. He had been refused access to Smith’s collection, as the Curator of the Compton Herbarium had concurred with Mrs. Courtenay-Latimer (and perhaps Smith himself) that Col Scott would not do credit to a formal revision. It became quite problematic for me when I realized that this estimate of Scott’s botanical capability was uncomfortably close to true. This led me to re-write my handbook as The New Haworthia Handbook (1981), also published by National Botanic Gardens, hoping that Col. Scott would follow the ideas expressed in my optimistic foreword.
This did not happen and Scott’s…The Genus Haworthia – A Taxonomic Revision, appeared in 1983 published by Aloe Books, Johannesburg, as a closely and clumsily kept secret from me.
My honest opinion is that this book was a total disaster, taxonomically though not photographically, and it was most disheartening and confounding to consider that the flattering introduction was by Dr. L. E. Codd, then head of the national Division of Botany and of the Pretoria Herbarium. Not only that, Dr. Beth Gibbs Russell later telephoned me to ask whose version of Haworthia the herbarium and the institute should follow! I do not know what my response was other than thinking how unfair it was to place the onus on me to judge. Whatever I might have said, that did not stop the institute from publishing an extraordinary mish-mash of names for Haworthia in a national species list taken from Scott, myself and other sources. It was simultaneously no comfort to welcome the grand old man of Haworthia, J.W. Dodson, to the Karoo Garden. Bewildered by what he saw in the collection there, he could do no more than express his conviction that Col. Scott had produced a very workable revision!
As far as I was concerned, Haworthia was all emptied back into Smith’s “monograph,” and readers may imagine why I found this so disturbing. Around 1993-4 news of a new succulent lexicon to be compiled by Urs Eggli to replace the famous Jacobsen set was circulated. At the same time, by virtue of my contact with the Compton Herbarium, I was asked to write a synopsis of Haworthia for a list of plant species for the Cape Flora. These were the defining factors that led me to writing a formal revision. I cannot bring myself to detail the communications with other aspirants to taxonomic fame then and since, as these border on the bizarre and do nothing more than add to the disrepute of taxonomy in the minds of plant collectors who care about and are compelled to use Latin names.
When I eventually wrote my revision I had no illusion that it would be the last word on the subject. Indeed it seems in retrospect that it may well have been but the first! Not only has there been an explosion of new names and opinions, I have also been driven to examine and re-examine those areas of my understanding built not only on available field knowledge and exploration, but also on Smith’s vast legacy. It is necessary to note that the contributions of people like Jay Dodson and J.R. Brown played no small part either. The little book written by Brown, viz., Succulents for the Amateur, played such a significant part in cultivating my interest in plants that I have been very sensitive to the resonance of the devotion and interest Brown showed for Haworthia. This respect for others like F.R. Long and H.G. Fourcade, whom I never met, extended to people like Frank Stayner, G.J. Payne, Gordon King and so many others I met solely as a consequence of these plants. That the human relations aspect of their classification and identification took such a downturn in the wake of Smith’s criticism of, primarily, Resende in 1947 is a sad record in history. Relations never recovered and my involvement in and distress at the later half-repetitions of history is at the heart of this article.
Since 1995 my accession records have climbed from 6400 to 7700 and these numbers represent almost entirely new population records for Haworthia and I trust that I have not used my own numbers where a very few contributing collectors have helped me. I have written innumerable articles for Haworthia journals while Umdaus published Haworthia Update Vol 1 in 2002. This seems to have been a commercial failure but my belief is that editors and publishers have never understood the issues. I could not help because despite my excessive attempts to explain the problems causing strife and confusion in the literature, I really never exposed the heart of the matter. Fortunately for me Harry Mays of Alsterworthia kindly published Haworthia Updates Vols 2, 3 and 4 between 2003 and 2007.
In these volumes I tried to address the realities of variation and the confoundment that exists in my own and the communal mind, about what species are or might be as related to a perceived botanical and scientific process for naming plants. But if 40 delegates could be brought together as happened recently in an effort to create a united front just for the single genus Aloe, it does seem fairly senseless to be functioning so alone in Haworthia where a greater need exists. In my “unsparing” view of things I find it extraordinary that the symposium did not seem to address the wider problem of genera in the Alooideae. I am quite sure that if this were done first, it would facilitate the common understanding of lower ranks. In Haworthia there has been absolutely no common will to reach any kind of consensus and no reason has been found for that. My conviction has grown that Latin names, and the way the nomenclatural system is used, do nothing more than strain good will in that they implies a reality in a staged relationships of plants that is simply not true. If I were a more dedicated taxonomist I would have simply worked towards a functional generic arrangement: Haworthia for the glamour queens, Jayarbia (honoring J.R. Brown) for the Hexangulares, and Catevala for the Robustipedunculares.
I recently submitted a short list of names that I think should serve Haworthia to the Haworthia Society in England. As Gordon Rowley pointed out in a letter to me… “The problem is that Bruce Bayer changes his mind.” Well, I do want to change my mind about that list again. I would advise incorporating H. heidelbergensis, H. magnifica and H. maraisii and their variants all under H. mirabilis. Furthermore I would suggest abandoning the formal lower ranks of subspecies, variety or forma for appended third epithets in inverted commas. The list of species is thus reduced to a realistic 54.
But there is still a wealth of undiscovered things out there and the many existing names mean very little to me in understanding what I have seen and continue to find. So I give a brief overview of the product of very recent fieldwork where in less than 10 days my wife and I found 24 previously unrecorded Haworthia populations. These included two populations of H. floribunda, one of H. retusa ‘nigra’ (implementing a name change and a new way of expressing difference), five of H. rossouwii, one of H. minima, one of H. marginata, two of H. mirabilis ‘paradoxa’ and 12 of H. mirabilis. While each of these has a story attached, I am only going to touch on the latter.
I have written in detail about H. mirabilis in Haworthia Update 3:87 in a chapter that deals primarily with the populations in the western area of the distribution. I also wrote in detail of associated plants in Update Vol. 2:51-79 dealing with populations east of Heidelberg in the eastern area. So these new populations are relevant to the central zone and I will add several others that were found earlier but subsequent to the Update 3 publication, plus another by Marc Mougin (brought to my attention by Diederik Van den Abbeele). They all deal with an area extending eastwards from Stormsvlei and the Bromberg Mountain to the Karringmelks River southwest of Heidelberg and southwards to Koppies and Stuurmanskraal, where there are populations already covered in Updates.
Five of the populations can be unequivocally assigned to H. mirabilis ‘atrofusca’ and yet with a fair degree of variation. These are all just west of Heidelberg. The other seven that do nothing more than confound understanding of anything that can be construed as H. maraisii, H. magnifica, H. heidelbergensis or H. mirabilis, occur southwest of Heidelberg and extend westward to Diamant (east of the Bromberg). The only significant thing about the two groups is that where ‘atrofusca’ is evident in the populations, H. floribunda may be in the vicinity. Where H. floribunda is not present the seedlings resemble that species as an odd indication that perpetuated juvenile characters underlie the species floribunda. The variation within the populations is quite dramatic and it is indeed difficult to verbally report and sensibly convey this to a reader. The plants vary in size, texture, markings, leaf armature, color and translucence. So I submit pictures with the manuscript and offer the following comment as best I can:-
Fig . 1 Map of greater Swellendam area showing population points
A map is provided as Fig.1. Greater Swellendam area to show positions of Haworthia populations noted.
1. MBB7254 Bromberg. (Fig. 2). This really belongs with a large set of populations extending westwards and north along the Riviersonderend Mountains and into the Robertson Karoo. I start with it because it serves as a virtual ‘outgroup’ — and this is because the southern slopes of the Bromberg are different to the northern slopes. They are quartzite (silicon oxides) sandstones and the northern slopes felspathic (other silica salts). This difference in mother material manifests itself in the Haworthia populations and can be used to explain why H. maraisii v.Poelln. in the shales and siltstones at Stormsvlei are so different to H. mirabilis ‘depauperata’ VPoelln. in the quartzitic sandstones less than 1km north. The plants are relatively small and have the reddish color often associated with H. mirabilis south of the Riviersonderend Mountains.
Fig. 2 7254 H. mirabilis. Bromberg.
2. MBB7612 Diamant W (Figs 3 & 4). These are very big plants up to 110mm in diameter with a similarity to ‘maraisii’ by virtue of their blackish-green color and more retused (bent back) leaves. The classic difference of ‘maraisii’ and mirabilis as being separated by translucence absent in the leaves or present, is again flouted in this population as in so many others; and similarly leaf surface texture is variable. The significant thing is that the mountain they are on is an extension of the Bromberg (Riviersonderend Mountain tail) but it is the northern slope and the sandstones are felspathic.
Fig. 3 7612.3 H. mirabilis. Diamant W.Fig. 4 7612.4 H. mirabilis. Diamant W.
3. MBB7626 Diamant Mid (Figs 5 & 6). This population is less than a kilometer east of the receding on a more eastern and thus cooler aspect. The plants are a little smaller, the leaves generally more erect and are a little more reminiscent of mirabilis to the southwest in that spines are better developed on the margins and surfaces than in the case of classic ‘maraisii’ (I am trying to show that ultimately there is no infallible distinction).
Fig. 5 7626.1 H. mirabilis. Diamant Mid.Fig. 6 7626.2 H. mirabilis. Diamant Mid.
4. MBB7718 Diamant E (Figs 7, 8, 9 & 10). This is a huge population on a lower elevation than the preceding and the skeletal habitat is significantly different. It is on the interface of the Felspathic sandstone and shales and siltstones of an older formation while a strong vein of quartzite transects it. In addition there is evidence of an eroded away, and later, ferricrete deposit. The plants are also extremely variable, reflecting the range of divergent surfaces although generally much smaller than preceding. Leaf surfaces are mostly roughish.
Fig. 7 7718 H. mirabilis. Diamant E.Fig. 8 7718 H. mirabilis. Diamant E.Fig. 9 7718 H. mirabilis. Diamant E.Fig. 10 7718 H. mirabilis. Diamant E.
5. MBB7633 Luiperdsberg (Figs 11 & 12). A small population quite high up on a slope of felspathic sandstone. These are small plants and because of quite a grassy habitat the leaves are a little elongated generating the range of forms that I have generally assigned to a little understood or known range of plants and populations in H. ‘heidelbergensis’ Smith.
Fig. 11 7633 H. mirabilis. Luiperdskop.Fig. 12 7633 H. mirabilis. Luiperdskop.
6. MBB7719 Dagbreek (Figs 13, 14, 15 & 16). An established population in felspathic sandstone but marginal to shale and siltstone. Very grassy but with occasional rocky ridges so that the plants are concealed in grass tufts or wedged in crevices. The plants are moderately sized to 60mm diameter and differ in that many have a rather smooth surface with corresponding finer and more closely spaced marginal spines. Some plants develop a shiny opacity about the leaf tips that suggest surfaces perhaps comparable with the smoothness of the H. retusa complex.
Fig. 13 7719 H. mirabilis. Dagbreek.Fig. 14 7719 H. mirabilis. Dagbreek.Fig. 15 7719 H. mirabilis. Dagbreek.Fig. 16 7719 H. mirabilis. Dagbreek.
7. MBB7704 Bontebok Park (Figs.17, 18, 19, & 20). There is an earlier collection (6644) from the ferricrete/shale interface alongside the N2 highway near the northeastern corner of the park, which I considered belonged in a series of five populations that seemed to be neither H. ‘maraisii’ nor generally accepted H. mirabilis. It is on ferricretes but there is some problem how to relate this apparent river terrace gravel with the solid caps of the inselbergs to the east and west. There is nothing common in the general suite of species that could be described as Fynbos on the former and grassy Renosterveld on the latter. The plants are moderate in size; surfaces can be quite rough with a tendency to spination, and the marginal spines quite coarse. Leaves erect or spreading.
Fig. 17 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 18 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 19 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 20 7704 H. mirabilis ‘maraisii’. Bontebok Nat.Pk.
8. MBB7705, 7706, 7713, 7714, 7715 Uitvlugt (Figs 21, 22, 23, 24, 25, & 26). These are five quite independent populations on the one farm that is unusual in that it seems to be on predominantly an earlier variant of Cenozoic ferricretes. The plants are generally related to H. ‘atrofusca’ with rounded leaftips and roughish surfaces. The actual habitats are all different in respect of surface detail and associated vegetation. The plants vary in size between the different populations from small (30mm diam) to large (70mm diam). In the one site where the soil is whitish bentonite, the plants are small and also have a curious similarity of color to that of the clay and the associated plant Gibbaeum haaglenii (austricola).
Fig. 21 7705 H. mirabilis ‘atrofusca’. Uitvlugt NW.Fig. 22 7706 H. mirabilis ‘atrofusca’. Uitvlugt.Fig. 23 7713 H. mirabilis ‘atrofusca’. Uitvlugt W.Fig. 24 7713 H. mirabilis cf ‘magnifica’. Uitvlugt W.Fig. 25 7714 H. mirabilis ‘atrofusca’. Uitvlugt E.Fig. 26 7715 H. mirabilis ‘atrofusca’. Uitvlugt N.
9. VA-RW07-27 (Figs 27, 28, 29 & 30). This population located by M Mougin, was reported to me by D. Van den Abbeele. It is much closer to Heidelberg and obviously in a ferricrete related habitat. The plants are all very ‘atrofusca‘-like with some evidence of the pointed leaf-tips that would have been ‘magnifica’. The surfaces have the slightly riffled texture that one associates with ‘atrofusca‘ rather than the tendency to roughish surfaces or almost spinescent tubercles associated with, but not peculiar only to ‘magnifica’.
Fig. 27 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 28 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 29 VA-RW07-27 H. mirabilis cf ‘magnifica”. Basryhoogte. M.Mougin.Fig. 30 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. D. Van den Abbeele.
10. MBB7728, 7729 Lilliendal (Figs 31, 32, 33, 34 & 35). The significance of these two populations must be emphasized. The place is the Karringmelks River southwest of Heidelberg. This river drains into the Slangriver that flows then southwest into the Breede River. It is quite a different drainage basin to the Klip and Duiwenhoks rivers that drain the area north and west of Heidelberg and enters the sea south of Heidelberg. Matjestoon and the origin of H. ‘heidelbergensis var. toonensis’ is south of Lilliendal and on the same drainage system. The plants would at the time have made it difficult to imagine H. ‘heidelbergensis’ as a discrete system, and these plants force a direct comparison with H. ‘maraisii’, if that system could be generalized. The plants are small, dark-green, rough and with a degree of spination on the surface tubercles. Surfaces may be opaque or translucent. There is a striking resemblance of the seedlings to H. floribunda. The populations are about 1km apart. Both are on Bokkeveld but at the second population it is curious to observe that the plants are not on the ferricrete only meters away.
Fig. 31 7728 H. mirabilis. Lilliendal.Fig. 32 7728 H. mirabilis. Lilliendal.Fig. 33 7729 H. mirabilis. Lilliendal.Fig. 34 7729 H. mirabilis. Lilllendal.Fig. 35 7729 H. mirabilis. Lilliendal.
11. JDV87-3, MBB7165 Matjestoon(Figs 36, 37 & 38). This is one of the original populations known to G. G. Smith and it is an uncomfortable fit anywhere on account of the length and spiny-ness of the leaves. That the plants were placed in ‘heidelbergensis’ is a reflection of the doubtful and composite nature that the species gained by default. There is a semblance of similarity to H. rossouwii but what may be more notable is that at Dassieklip to the southeast there is a very aberrant population that is assigned to H. floribunda simply for want of a more likely category. I say this is notable because in my field observations of Haworthia there is a very strong and predictive geographic continuity between populations. This is evidenced in this general discussion.
Fig. 36 JDV87-3 ‘H. heidelbergensis var toonensis’. Matjestoon.Fig. 37 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.Fig. 38 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.
12. MBB7736 Koppies (Figs 39 & 40). These are small dark-green plants with strongly deflexed leaves and a little unlike an earlier record from 2km west (MBB6879 ex E. Esterhuizen) where the plants have more pronounced surface tubercles becoming quite robust and dark in cultivation. The habitat is among white quartz occasionally associated with ferricrete.
Fig. 39 7736 H. mirabilis. Koppies.Fig. 40 7736 H. mirabilis. Koppies.
13. MBB7723 Stuurmanskraal (Figs 41 & 42). These rank among what might be regarded as the general body of H. ‘maraisii’. Small, dark roughish plants with strong evidence of floribunda in the way the leaf-tips tend to be flat, twisted and rounded. The habitat is Bokkeveld shale.
Fig. 41 7723 H. mirabilis. Stuurmanskraal.Fig. 42 7723 H. mirabilis. Stuurmanskraal.
14. MBB7734 Stuurmanskraal N (Figs 43 & 44). This is a previously known population similar in its responses to the Koppies plants (MBB6879) that become quite robust and dark in cultivation. The habitat is white bentonitic clay that develops under the ferricrete sills from decomposure of underlying shale.
Fig. 43 7734 H. mirabilis. Stuurmanskraal.Fig. 44 7734 H. mirabilis. Stuurmanskraal.
15. MBB7731 Oudekraalkop (Figs 45, 46 & 47). Finding this population at all suggests that there may be many more evading the eye. The plants are small and dark and completely hidden among dense but short vegetation on a rocky scree-like area. As in other populations one is compelled to look at every plant and consider its departure from a hoped for norm for the plants there. Three illustrations hardly do justice to the variability and this true for all these populations discussed here.
Fig. 45 7731 H. mirabilis. Oudekraalkop.Fig. 46 7731 H. mirabilis. Oudekraalkop.Fig. 47 7731 H. mirabilis. Oudekraalkop.
Conclusions My despair over the years that I have been involved with Haworthia, is that the classification model and nomenclatural system we use, does not seem to allow me the freedom to express what I have seen and experienced. My argument has been that we are dealing with a biological system in which things that seem different are the same; and conversely things that seem the same are different. It is not a problem to apply a formal classification system except that a ranked or tiered system suggests quantum differences too, and this is simply not true. The greater problem is that most plant enthusiasts, and those who feel strongly about classification and names, confuse the reality of the sameness that suggests ‘species’ with the sameness that plants of different species may exhibit.
It is with a source of considerable curiosity that I now take note that a taxonomist is one who classifies by appearances and a systematist one who classifies by evolutionary pathways. In truth I never considered that classification was anything but an attempt to accomplish both aims so that meaningful names could be established. The issue is clouded in complexity when it is considered that neither taxonomist nor systematist may be sure when a species is a species. Probably hence the observation that species (binomials) carry a reality they do not possess. My proposal, supported by the observations in this article, is that we adopt a far more informal and flexible approach to the use of all the names we now have at our disposal. This suggestion is also supported by an experienced taxonomist/systematist/botanist who wrote: “The informal system that I prefer allows for endless forms that are identifiable by their locality and do not clog up the literature. Essentially it is a reversion to the polynomials of the pre-Linnaean era.”
My contention is that the only rational classification of the above 20 populations is within a greatly expanded concept of H. mirabilis that includes H. maraisii, H. magnifica, and H. heidelbergensis. This corroborates the observations reported in the chapters in Haworthia Updates and specifically Chapter 6, Update 2,1:50-79 “How to understand H. mutica var. nigra”, and Chapter 1, Update 3,1:1-87 “H. mirabilis and the interface with H. maraisii”.
Acknowledgements The population MBB7704 was shown to me by Ms. Tineke Kraaij of RSA National Parks Board. The concurring botanist is Dr John Manning, who has authored many botanical works and who is best known for his familiarity with the highly complex Cape bulb flora. Dr. Syd Rhamdani introduced me to the new direction that molecular biology is driving plant systematics. Gerhard Marx has been a great help in getting me to resign to the unrealities of popular taxonomy. Steven Hammer, considerably more successful than I have at adapting to the fact that our perceptions about many things are very different to that of others, helped me with the manuscript. Rhett Hiseman of Cape Nature, Jaap Viljoen, Henniie and Elma van Deventer of Koppies, the Odendaals of Dagbreek, Diamant and Dankbaar, Alfred and Andrea of Lilliendal, Dirk Papendorp of Uitvlugt, D. van As of Stuurmanskraal, Anna and Arno Steenkamp of Oudekraalskop (Anna’s Farm) are sincerely thanked for information and/or access to plants. ♦
If the name “H.enigma” applies to the plant (or plants) from east of Riversdale at Komserante, it is a name that I really do not advise to be taken seriously from a botanical point of view. It is useful at population level and to demonstrate the nature of classification difficulties but it is a minor problem in so far as those difficulties extend. The plants were first shown to me by J. Dekenah on the same day that he also showed me ”H. magnifica” in the Nature Reserve just south of Riversdale that is less than 3km away. My impression then was that it was the same element even if it did look a bit different. The plants are quite large (to 70mm diameter), fairly tubercled and often with lines in the upper retused area of the leaf face. While I originally classified “H. maraisii” under “H. magnifica”, I later separated them because it seemed so incongruous to include all the variants of the western “H. maraisii” with the few populations of “H. magnifica” then known. Also, as Essie Esterhuizen pointed out, “H atrofusca” as a variant of “H. magnifica”, seemed to be more dominant than had been realized. There were several other complications largely due to ignorance. Since my revision I have done so much more exploration and turned up so much new material that I have been forced to the conclusion that there is really one main element involved and that is H. mirabilis. This is where I believe the Komserante plants belong and the difference from the Nature Reserve population is due to a degree of infusion of H. retusa.
I revisited the site with Kobus Venter many years ago but did not look at a reported second population higher up the hill, taking it to be a little different based on plants I saw in Kobus’ collection. What was on my mind while we were recently exploring the area further east to examine the possible connection of H. mirabilis “magnifica” to “splendens” (and which we confirmed), was the fact I had never seen Kobus’ plants from Kruis Rivier northeast of Riversdale other than in Kobus’ collection. The plants I saw were also generally more robust than “H. magnifica” and more evenly tubercled. Kobus kindly took me to that Kruis River locality and much to my surprise the plants were in flower late October (see JDV92/65 Figs1). This is quite wrong for H. mirabilis, which is essentially a summer flowering species. I later went again to explore Komserante more thoroughly and to look at both the “magnifica” populations to which I believe the name “H. enigma” has been applied. The populations are in fact no more than 75m apart and cannot be considered to be genetically discrete at all (see MBB7778 Figs 2, and MBB7779 Figs 3). While it is true that the habitats are slightly different, this is reflected in the plants that at the upper slope of the hillside are vegetatively more robust and even clump forming, while those lower down in a bushier grassier habitat tend to be solitary and more withdrawn into the soil. These plants flower in summer and it is evident to me that there must have been some genetic exchange with H. retusa that grows approximately 200m away on the same hillside.
1. JDV92/65 H. mirabilis Kruisrivier.
2. MBB7778 H. mirabilis. Komserante.
3. MBB7779 H. mirabilis. Komserante.
Habitat preferences are very strict and it is curious to observe that there are four small buttresses forming the western slope of the low hillside. On the northern one we have the two H. mirabilis populations. On the second is H. retusa ’geraldii’ (see MBB7780 Figs 4) that is very clump forming, the third has no haworthias and on the fourth is a less clump forming H. retusa that is often referred to as ‘fouchei’ (see MBB7781 Figs 5) because the leaves are slightly longer and more upright than in ’geraldii’. This is not quite correct. Mr. Dekenah showed me what he took to be the true H. retusa ‘fouchei’, further south off the hillside and along the stream at Komserante. This habitat has since been cleared for farmland. The plants were very large and the leaves considerably more erect than is the case for the plants on the hillside. Curiously H. minima is present on the first and last buttresses. Flowering time for these H. retusa is spring but we did see vicarious flowering in summer so that definitely it is possible and probably a regular occurrence for out-of-season flowering to occur with the possibility of hybridization. I noted such hybrids between H. retusa and H. mirabilis in our exploration further east from Riversdale and I did think some of the clones in the Komserante plants evidenced this as well.
4. MBB7780 H. retusa. Komserante.
5. MBB7781 H. retusa cf. ‘foucherii’. Komserante.
The reality is that the Kruisriver and Komserante “magnifica” may flower at different times, but they are vegetatively very similar indeed. I consider that they belong in the same system as H. mirabilis and this conservative view of “species” as systems is the most sensible way to interpret and understand the plants. The approach can be taken further, as I tried to do in an earlier article concerning H. mutica ‘nigra’. In that article I dealt with a whole series of populations in and around Heidelberg, to show that there are three main elements viz. H. retusa (including H. turgida), H. mirabilis (including H. magnifica) and H. floribunda. I did not deal specifically with, nor resolve, the issue of H. mutica var nigra. I also noted populations along the lower Duiwenhoks river and suggested that H. floribunda is absorbed into H. mirabilis virtually south of the N2 east west highway, re-emerging on the northwestern corner of the Potberg in the south.
My most recent exploration was to check the flowering times of the original population of H. mutica var nigra at its origin at Kransriviermond and what I assumed to be the same thing on rather limited examination further north at Morning Star. Both these populations are along the Duiwenhoks river southeast of Heidelberg and in a continuum of four populations of H. retusa extending from just east of Heidelberg to still further east at an eastern Diepkloof locality (I think it has been referred to as Droekloof and Droerivier too, and the name ‘chromatica’ attached to the plants – see MBB7794 Figs 6). Less than 10km away is a population of H. retusa at Pienaarsriver (see MBB7776 figs in Chapter 3) that is very near to the whole array of populations of Kiewietskraal that I discussed in my writings about H. mutica var nigra. Two of those were distinctly H. retusa and so is the Pienaarsriver population that is different in that the plants are mostly quite dark in colour.
6. MBB7794 h. retusa ‘chromatica’. SE Heidelberg.
In those same writings I mentioned several populations of H. mirabilis along the Duiwenhoks river as well as populations of H. retusa var turgida. Here I am unashamedly relating “turgida” to “retusa” as I believe that when one calls all the populations now known to mind, as well as the variants they contain, this is the correct position of the plants in relation to a sensible and rational species concept. It is evident that H. floribunda is merged in H. mirabilis south of Heidelberg and the same thing occurs south of Swellendam. Thus I see no problem in now dealing with the population at Kransriviermond to which I applied the name H. mutica var nigra. I attached the varietal name to H. mutica because this is what G.G.Smith had done, and I believed that it was the link between H. retusa in the east and H. mutica in the west, when there were still huge gaps in the known distribution records for both species.
I have mentioned a population of ostensibly H. mutica var nigra at Morning Star, which is about 5km north of the Kransriviermond origin of the plants so named. This Morning Star population (see MBB7221 Figs 7) is very significant because while it includes virtually the same range of variants as the Kransriviermond plants, it flowers in summer as opposed to spring for Kransriviermond. Not only that, but it is 300meters away from a population of H. mirabilis (previously this would have been H. magnifica) that flowers and seeds at the same time. The habitat is marine plane ferricrete that overlies Bokkeveld shale but there are subtle differences between the two habitats.
7. MBB7221 H. retusa ‘nigra’. Morning Star.
The Kransriviermond plants (see MBB7804 Figs 8) are down in the Duiwenhoks valley in exposed Bokkeveld shale and I consider that the dark colour of the plants in the two populations is due to infusion of H. mirabilis. The rounded leaf tip would be derived from the “atrofuscoid” element of H. mirabilis while some plants also have the tuberculate roughness that that element often has.
8. MBB7804 H. retusa ‘nigra’. Kransriviermond.
Further south and west is the Slangrivier valley where there is also a suite of populations that includes smaller forms of H. mirabilis (H. heidelbergensis var. toonensis) as well a H. retusa var turgida also in an unusual form that supports the taxonomic position I now adopt.
The conclusion is that the Morning Star and Kransriviermond ‘nigra’ can be regarded as H. retusa within the same broader concept that includes ‘turgida’. The problem now arises from still another new discovery. This is from along the Buffeljags River (see MBB7801 Figs 9) about 10km southeast of Swellendam and 12km from the nearest obvious H. retusa at Goedverwagting further east; and from the less obvious H. retusa (that I have elsewhere noted as H. mirabilis) south of the Tradouw Pass. The nearest H. mutica are within about 15km to the west at Napky and Luiperdskop. The plants are the now customary “variable”. The leaf tips are very rounded. Some plain, some lined, some with curious submersed pinkish dots. The surfaces can be quite smoothish or else the surface cells terminate in minute spines that give the surfaces a glistening appearance – evident in H. mutica too. These plants flower in summer whereas both H. mutica and H. retusa are spring flowering. Note needs to be taken of the habitat. It is an extensive and old river gravel embankment and terrace, and therefore another variant on the interface of recent ferricrete deposits and the older Bokkeveld shales. The vegetation is grassy Karooid bushveld, unlike the dry fynbos to the northwest or the renosterveld (mainly cultivated) to the east.
Kobus Venter saw the photographs and suggested that the plants might be H. mirabilis’ and reminded him of ‘atrofusca’, perhaps also by virtue of the flowering time. This is of course what my “enigma” is all about. It is compounded in this case because these Buffeljags ‘muticas’ are rather odd in that the leaf ends tend to be flattened almost as though H. floribunda has also entered the fray. Kobus was much happier with the H. mutica connection when he saw the plants in the field.
The story here closes with H. mutica MBB7741 from Dankbaar about 12km west of the Breede River bridge at Swellendam. The plants here also have very glistening surfaces and thus seem to confirm the very original perception of G.G. Smith’s that the Duiwenhoks plants and the plants west of the Breede River were connected. It is certain that he never envisaged the complexity of the interrelationships between three main elements viz. H. mirabilis, H. floribunda and H. retusa.
Footnote. It is just assumed that perhaps the name “H. enigma” has been used for the Komserante H. mirabilis, because that population is indeed enigmatic as this article explains. Gerhard Marx informs that “enigma” applies to a H. mirabilis population further west where the more atrofusca variants occur. Since writing this chapter yet another similar population to the Kruis River and Komserante populations has been found about 15km SE Riversdale. ♦
James Deacon asked Bruce Bayer, “What is this thing at Brandrivier?”
James – you have no idea how significant this picture is. It is an emelyae variant and I really need to know a lot more about precise whereabouts. Multifolia comes from a few km to the east and there was a very very rossouwii-like multifiolia on Brandrivier. It is pictured in Revisited. But it is not there anymore. What is there are plants like your picture indicate. Did you perhaps observe Tulista opalina?
Jakub Jilemicky added “This plant was described by Gerhard Marx as H. obserata. It occurs mainly at Brandrivier, but as well at Springfontein. It flowers with all the magnifica type plants, not with emelyae group.”
May 19 – 7888 Brandrivier. H. emelyae ‘obserrata’. It raises an interesting issue. Names may have to change every generation as the plants may change? It used to be ‘multifolia‘. Actually my species definition includes the matter of space and time. In which case the name ‘obserrata‘ is taxonomic malfeasance.
May 19 – 7846 Between Springfontein and Brandrivier. Flowering August. Surely the type loc for “H. obserrata” that I see as H. emelyae. But this is just a distraction. When you still have commentators making comparisons to the magnifica types and flowering times, you realize that the absurdity of Haworthia classification is going to continue for the next 80 years as well.
As a point of interest. There seem to be many different ideas of what species are. From the typological concept of things that look near identical, to a list of the species one thinks makes up a genus. In the latter case the genus is considered to be what is made up from the species one imagines. My own species concept is very well defined and my map (this spider web of colours) indicates the way factors of space and time are involved in only one small group of Haworthia.
Still paradoxa Vermaaklikheid but the first few were June and this is September.
Let me try this as no 3…What we have is a post of three pictures to show three “species” paradoxa, bobii and joleeniae and to ridicule Bayer for suggesting they are the same. Firstly those pictures are NOT the species. They are specimens taken to illustrate the species but by what is recognised in botanical taxonomy as the “typological concept”, i.e. a species consists of individuals that all look the same. I wish I could copy here what I wrote in the New Haworthia Handbook (1976) where I use the name H. magnifica var. paradoxa and discuss affinity with H. turgida, H. emelyae, H. retusa and H. mirabilis. I ended the discussion there with this… Consideration of the variability of this species and distribution of variants, is very helpful towards understanding variability in the genus as a whole. What I conclude is that the species are systems with any number of populations and individuals that vary enormously. The typological concept creates mayhem in the minds of critics, growers and collectors alike. So let us start with the paradoxa, but first dismiss H. magnifica as a species. That name is a total myth. The myth ghosts around probably a single plant I collected with J. Dekenah at the type locality. It proved very amenable in cultivation and plants ended up at Sheilam nursery to go world-wide. It is actually difficult to now find a clone quite the same at the original locality where there are several discrete “populations” in quite a small area – and countless variants. It is senseless to say that H. magnifica can be a species if in every aspect of such a decision it is topologically based, as is the unfortunate case. The name in SANBI is H. mirabilis. Paradoxa is a synonym. SANBI are not my friends or my clients. Some very bright and competent people are employed there. The element paradoxa is thus in my view a variant in the species H. mirabilis and illustrated here by three populations viz. at Vermaaklikheid, Osplaas and Koenserus.
Just get something clear about type specimens and the system. A specimen is only anchor for the name. It does not prove anything about where it comes from. It is just representative. A second point is in that the nomenclatural rules uphold chronological discovery (better still – description) over reality. This mad rule of priority totally confounds the prime aim of a classification that follows and explains the genetic history (phylogeny). If in fact evolutionary theory is even valid. Generally the rules provide fertile ground for confusion, argument and publication credits that do little to promote their purpose.
Two Vermaaklikheid picture sets have been posted. This is another. A cooler west facing outlook but also on those limestone rocks. I must confess to a bit of apprehension that some may claim this is a different species when my opinion will be a great deal more conservative in what lies ahead.
What is the problem? I suggest we have an antiquated, outdated, dysfunctional classification system so that our knowledge, philosophy and methodology does not allow explanation of groups like Haworthia. So I will try and demonstrate it like this…Gerhard posted pictures of paradoxa, bobii and joleeniae. One of each! So let us just see how this can be interpreted. Here are several pictures of ‘paradoxa‘ from Vermaaklikheid east of the village. I first saw it at a site north of the village.
There is so much still to explore. Here is a map just to indicate the scale that has to be exercised. Initially I was happy to stop every 50 miles to see what might be there. Now it is every 25meters! Just note an interesting phenomenon here. 3 species and as many populations as there are recorded habitats. M = mirabilis, R retusa and F floribunda. Yes we covered most of the ground between too.
Mukesh Vaid asked – What does these four digit numbers signify
Bruce Bayer – Just my collecting numbers. This map is only intended to show intensity of search to demonstrate anything. I did have a collecting permit strictly adhered to. Often, as I do now, I do my searching with camera!
Every collector should have an accession book to record what you record and find where and when. I idiotically did not get this right to start with. Even as an entomologist I did not keep proper numbers. When I started at Karoo Garden I used the garden accession system KG with number/year. No good. In the end it is the quality of accession record that indicates the credibility of the work! I did not have the luxury of digital camera or GPS that is essential nowadays. At one time it was enough to just say “collected between the Cape and the diamond fields! With plant theft and cadaverdog-like shadows, it is a bit stressful to even produce the maps that I show! Sometimes I have mentioned something and the next moment a new species is described directly connected??
Mukesh Vaid – Where is H. turgida in this?”
Bruce Bayer – This not actually about turgida but just to show level of search necessary. But let me take the opportunity to try and get this message/hypothesis across. There is a single gene pool (a system) that produces mirabilis, retusa, turgida, pygmaea, mutica and emelyae. It can be simplified if it recognised that retusa is really a offset of turgida in among 5 main lines. So let us try and get something else straight. Science (i.e. scientism actually) is a system of reductionism that supposes that everything can be explained by breaking things down to their smallest parts. So we mess up in Haworthia by thinking the smallest parts are species when in fact it is a system of many things. Does that help?
This is where I think Mukesh rightly has a problem. That bottom pink line should not be there. That is why I refer to them as intervals. These bureaucratic scientists lay down the law that this should be seen for what it is and call it an interval – because it is a break in distribution continuity e.g.. The Knysna interval is a serious break between SE coast flora and S Cape flora. That coast line area is not easily explored and the geology is limestone with a different vegetation. There may well be Haworthias there. There is a record of a mirabiloid(?) on the sea cliffs at St Sebastian west of the Breede River mouth.
There is a really weird intimation that bureaucrats have determined what species names apply on Haworthia! It is actually a very democratic process based on 4 tenets of science. Universal truth, communality – no secrets, no private gain, and organised scepticism. If you have the data and you have the facts, you are free to organise and present your case. I have no privileged access to anything and certainly no sycophantic following with commercial interests in mind. I often wonder that not a hair sticks up from the trenches of my defense when so much garbage is thrown on attack
On this map…. There is actually no known connection across from L = longebracteata (also a mirabiloid area), to P = paradoxa, to B = bobii. Historically the shore line has changed and there may have been. Variegata seems to have jumped the gaps. So now we have populations a = Buffelsfontein, b = Sandfontein, c = Sandhoogte and d = Infanta. A and b are in the De Hoop Reserve and it is quite difficult to access so I have not been there with a digital camera unfortunately. Buffelsfontein I owe to Chris Burgers, Sandfontein to Adam Harrower and Sandhoogte to Jakub Jelimicky. Infanta I owe to mesemb research and what “bob” had to do with it is anybody’s guess. Certainly I did not remotely think it was or is a new species, as excited as I was to first see it. The overall complexity of the spider web does not do justice for the problems of variability among the floribundoids, the variegatoids and the mirabiloids. Turgida seems to hold its own. Holistically I can conceive of only the one solution that excludes adding another few species to an impossibly and unrealistic list. It would be nice if the culprits owed up to the chain of events that led to “bobii” and a few other names. Including the authorization for collecting that is so uncomfortable and dehumanizing. I make no apology for suggesting the association of bobii and paradoxa , even beyond a greater issue of the mirabiloids. I do not doubt that it is a difficult “ask”. The two pictures are of ‘a’ and do not do justice to the plants in the field at all.
Arthur Dixon posted these .. Bruce Bayer replied “All ‘paradoxa’.”
These are the plants at ‘b’ where Adam Harrower recorded them. I named the place as Sandfontein or “Sandhoogte N”. Makes me long to go and drool over them again!
These are my pictures of ‘c’ in the ‘bobii’/paradoxa/joleeniae/mirabilis milieu. Funnily enough I really do sympathise with the doubters and unbelievers and am very apprehensive about further showing you what I think is the same. (remind me if I forget!). I have not mentioned “muticoid” in this lot where I say turgida is real. I suggest that mutica does come in to the picture and I will touch on that when I get to ‘joleeniae’.
Arthur posts…”It seems to me that endless discussions and reasoned arguments supported by habitat shots whilst life-affirming and endlessly enthralling are not advancing the discussion in any way as (returns to main point) WITHOUT that definition of ‘species’ the cycle continues to revolve (Personal point)”. Yes yes yes. It has been ongoing for the last 80 years. Like I said – I get credit for my great contribution to Haworthia while effectively been told I know nothing. I am happy to accept that, if some intelligence is put in place of what I have written over the years. Contrary to general belief, I do not like offending or hurting people. Therefore I am very limited in what I can actually say and tell. What we need is a dimensional shift in how we think, how we reason and in what we are really wanting to achieve. We have a societal crisis and we see it not!
Tony Brook wrote “My apologies Bruce I am not a Botanist but surely the definition and description of Bobbi should precluded the 2 plants which are relatively hairless. Otherwise many other Haworthia would be included in the “species”.”
Bruce Bayer replied “No Tony. this is essentially the problem. I did not consider bobii to be a species. H. mirabilissii the species and the best way of defining species at the moment is by DNA sequencing but rather also by what my definition of species requires. Viz a holistic view of its relationships, similarities whatever, with possible and probable relatives.”
Here is another ‘joleneae‘ population that demonstrates how observations can be slanted to re-enforce propaganda. Someone also sent me an article on Haworthiopsis that is a piece of real modern day journalism. Unable to tell left from right or right from wrong, the writer sits on the fence like a Fiscal Shrike impaling worms on the barbs, i.e. hoping to have his believers (us ordinary worms) accept him as an all-seeing eye of truth. This has plagued me all my writing life! No joleneae is not definitely and distinctly, always glabrous. Some of these pictures even show the mutica shiny speckled appearance those plants can have. Differences between populations are in my observation as much due to the skeletal soil differences (geology) as they are to spatial isolation. I think. Some amazing individuals here.
So let us move on because there are still surprises and unpalatable truths. While we have seen mirabilis ‘bobii‘ from the sandstones of the Potberg itself, we need to see what mirabilis does on the geological confoundment of the flats north of the Potberg. First there is Melkhoutfontein and I assume that this is included in the concept of “joleneae” (that I usually misspell). It really should be H. mirabilis ‘joleneae‘ and just forget formal process. It is not possible to regulate and manage the welter of names. An attempt was made to register Aloe cultivars, but if this is practical I doubt. It is senseless to argue there is no continuity from ‘bobii‘ to ‘joleneae‘. This just upholds a nonsensical humongous heap of formal binomials to argue about and ignores the many more things that attract either no aesthetic attention or commercial interest. Joleneae is on the terrace cobbly sandstone as a simply descriptor.
It occurs to me. Joleneae is claimed to be consistently glabrous. This comment disregards the electron microscopy “work” done by Dr. David Cutler of Kew that I reported on in Updates. It is obvious from these pictures that the plants are incipiently hairy or spinose whatever is correct. we simply cannot properly assess the real nature of individual characters like this. Just for recall – there was an amazing paper written on the cladistics of the Aloids published in Taxon (about 30 years ago). Whoever peer reviewed that paper prior to publication deserves serious censure. Barely a character state is correctly stated.
What it needs to is for the thought leaders, like editors, society committee members, media notaries etc. to get their act together and seriously consider what their role is in the mess of argument and infighting. I have suggested now a solution where at least in Haworthia we can (if we so choose) admit the spider web and set-out how to communicate about it. Perhaps we need to identify the mafia who have vested interest or are just too intellectually lazy or impractical to even want agreement.
Is this and that getting too much for you? Still ‘joleneae‘. It is a useful name, but not actually the issue. At this place Tulista minima (I am gobsmacked that some over zealous mind has decided it is actually T. minor – I suppose this name change in intended to avoid confusion!!! ) as does H. variegata ‘hemicrypta’. So I include hybrid pictures. The two species do not directly share habitat. Stoffelsriver.
Here is a map of the north Potberg area to highlight the significance of geology and its impact on floristics/vegetation/plant species. It is a complex situation with deposited materials overlying basic formations and erosional effects. The white material to the higher left are eroded and decayed shales that result in banks of white clay much like elsewhere towards Riversdale and Heidelberg. Is it surprising that the retusa/mirabiloids adjust and adapt accordingly. Why do these plants differ so much from joleneae/bobii/paradoxa and switch to resemble atrofusca and floribunda?
Snakes and ladders. Background noise! I have 206 folders for populations of which most are this unphotogenic ilk. Their lack of appeal and hidden nature is why H. heidelbergensis was spawned. The more conspicuous and bigger forms were H. maraisii and H. magnifica. The only attention I know they ever got from collectors was the complaint that they never could own or get a proper idea of what H. mirabilis looked like. Shades of the great doyen of Botanical Latin and plant taxonomy W.T. Stearn who destroyed the foundation of Haworthia taxonomy with his typification of the name Aloe atrovirens, spinis herbaceus numerosus ornata and choosing the herbaceus part for the epithet. H. atrovirens should be H. mirabilis! I do not think the name mirabilis was even used in von Poellnitz and Smith’s time? So I can see that my chances of putting Haworthia taxonomy to rest are zero. No herbarium can ever hope to hold a proper record of variation within a single population. Whoever wants to quarrel with me needs to now that their classification includes all the stuff I have tried to contend with! My pictures of this background noise are disappointing and I have none for stuff seen before ca. 2005 or so. Emoji of laughter or tears? And who wants to know what a species is anyway?
Nothospecies or notaspecies? That is the big question. Why bother to ask difficult questions, just call this H. mirabilisXmutica H. hammeri instead. It consists of one population occupying an area less than 500sqm among surrounding mirabilis and mutica populations. Will it ever be a system? (Nothospecies is coined for the offspring of two other species that in the absence of a definition become a new species. It really applies to readily seed propagating and readily dispersing plants – agriculture and horticulture!)
Here is my 2009 baby I think written in 2009 before DNA sequencing proved to us that there were 3 genera where we saw only 1. I said so when? First handbook 1976? The thing that really got me is that the results of the sequencing were actually fudged (again). The discussion and conclusions were written long before the introduction and methods, to ensure that there was a match up of results to hypothesis being tested.
Yes my favorite critic had something to say about this fishtail bud as well. Or at least the fact that I considered it at all. It is actually a feature of virtually all of the retusoid mirabiloids varying from zero in the east to nearly dramatic in the west. So herbacea and reticulata have the most exaggerated version. In these two systems the flower is nearly “regular” (as in pentagonal) hence reticulata ‘subregularis’. H. herbacea is characterised by the beige colouring while reticulata flower is pinkish. The Wolfkloof Villiersdorp herbacea has an enormous (close to 3cm long) very pink flower to fudge the issue. H. herbacea ‘paynei’ near McGregor has a bicoloured flower, hence ‘luteorosea’.
6509 H. mirabilis meiringii, W Bonnievale – Flowers are problematic because the flowers are not static things. They change by the day. In this case I pictured most open flowers on every single stem from 1-5 open.
6089 NW Potberg – some of the background noise to mirabilis. Many many populations of these small black things (“klein swartetjies” according to Michael Malherbe founder of Sheilam, in imitation of the Little Brown Jobs of bird-watching)). Not as enchanting as the larger and less common variants, they are not covered by popular classification.
H. mirabilis ‘notabilis’. This is a place in the mountains midway between Robertson and Worcester. Both arachnoidea and reticulata occur here as well in different habitats. There are shale, dolomite and sandstone formations. The countryside is formidable. How does one describe the plants here and link them adequately to a host of populations running all the way east to Ashton, and to the oddities of the NE Worcester area. Not only that. There are continuities southwards as well.
Lets try something. First the chaos formula imaging. Like a DNA seq. phylogram it is two dimensional and cannot adequately represent a spacetime issue (that species are whether evolutionary or independent creation of ‘species’. Secondly imagine a continuation of the fractal image with 6 species in a particular time frame continuing on to eventually end up as 3 species in a third frame? The frames are geographic space. The brackets demonstrate how species may overlap in distribution and/or characters. The chaos formula demonstrates overlap of characters in time or space but not in both as is needed. That is called linearity as opposed to actual reticulate reality? I am no intellectual wizard but am quite sure that plant taxonomy has got things WRONG.
Guan Tan-Amo Lim wrote “Haworthias are a little bit like domesticated dogs… they come in all shapes and sizes and can breed with each other readily. Yet dogs are all of one species. Could the many Haworthia species similarly be reduced to if not one species then to only a few? With many “breeds” corresponding to the natural populations in habitat?”
Bruce Bayer replied “Guan Yes. but we shift the problem from one level to another. I have a spider web of five species (turgida/retusa, emelyae, pygmaea, mutica, emelyae that could be one. But there is another spider web in the upper left of herbacea, reticulata, maculata and pubescens. So where do we stop? Things like wittebergensis, pulchella and marxii seem totally separate but there is chaos among all the other true Haworthia. Funny I was just browsing an article on a math issue that claims that there are things that are true but they cannot be proved to be true? Life is a conundrum.”
A fairly random lot? Actually no. This is a place a few km from ‘hammeri‘. Their is a rounded hill with small nondescript LBJs (little black jobs) most places, but at one spot quite large plants like this. Atrofuscoid? The floribunda-like leaf tip is common in the small mirabiloids. I should add… a note on fieldwork. Did I do field work? Walking around with nothing to measure and itemise but an eyeball assessment and some pictures.
6638 from NNE Bredasdorp. Mirabilis. This is a place where turgida also occurs, and with rossouwii and mutica within a few km. Also home to the ‘heidelbergensis minor’ that I suggest has rossouwii genetic material. One of these clones is very reminiscent of what I observed and considered a true hybrid between mutica and mirabilis where populations were in very close association. Does anyone really have problems with names used in an informal way like this rather than behind a facade of deep science and a degree of fraud?
From 6638 pictures. This weirdo strikes me as reminiscent of more obvious hybrids retusa/mirabilis or mutica/mirabilis where they are in close enough proximity to support a hybrid presumption. Turgida is an estimated 400m away from here.
Soumen Aditya comments “I didn’t see a features of natural hybrid … Most probably the polymorphic evolution…” Here is what IS most probably hybrid. Where mirabilis and mutica are in two populations meters apart. SO I think what would be features of a natural hybrid may just be a subjective opinion? I have mentioned before how problematic the issue of hybrids is and have often been asked – How do you tell it is a hybrid? Credibility in classification barely exists at the best of times.”
What do I ask? I once was in a meeting with a group of top managers discussing a topic they had been dealing with for over 20 years viz. rangeland production. I was suggesting a method of assessing potential. The group seemed so lukewarm to my suggestion that I felt the need to establish if they really did know what I was talking about. So I sent around a problem and asked if it could be solved using the available knowledge they had been debating for over the previous 20plus years. Deadly quiet. No wonder they were so indifferent. Now I feel exactly the same about Haworthia classification. People have been arguing and debating this since Herre’s time (1930). Here in SA nothing has changed. Perhaps I also do not know what I am talking about! The spider web for the retusoid/mirabiloid plants is a small part of the whole, and fairly straightforward compared to the bigger picture. But my suggestion for a systems approach is met with the same deadly quiet, especially from where it matters. What do I do next?
These are a few pictures of 30 from a population in a vast expanse of a no go area for any but the most intrepid. What is it? It is from an aloid sparse space and while I do have pictures for another 5 populations in the area they are not more informative. From my limited (is it that bad?) experience of mountains and the way succulent exploration has been done, I shudder to think how the Haworthia community, led as it is, is going to be able to digest what is still to be learned.
I mentioned that vast no-go area (6694). Here is one of the other 5 populations I know that happens to be only about 500m away (7994).
7994 Yes the flowers are significantly different. 7994 I attributed to maculata and the fishtail bud indicates that it is an extension of the retusa/mirabiloid web. The rounded bud tips of 6694 suggest arachnoidea or nortieri, as geographic candidates and the most probable system affinities.
June 4, 2021 – Something useful I can do is amplify H. mirabilis meiringii. I described this to draw attention to very herbacea like forms of mirabilis. This is in the Bonnievale area where the mirabilis variants seem like a system within a system, within a system. Westwards they are less spinescent and similarly eastwards. 6509. The name was not to honour someone so much as to record a time when plants were perhaps first harvested and sold from the field.