24. Haworthia mutica Haw. Revis :55(1821). Scott, Aloe 11:4(1973). Bayer :139(1976). Bayer, Excelsa 8:50(1978). Bayer :48(1982). Scott :118(1985). Type: Cape. Not preserved. Lectotype: icon (K). Epitype (B&M): Bredasdorp to Swellendam, Soesriver, Bayer in KG623/69 (NBG): H. otzenii Smith, JS.Afr. Bot. 11:72(1945). Type: CAPE‑3419 (Caledon): Bredasdorp (-BD), Otzen 6 in Smith 5478 (NBG).
mutica: without a point.
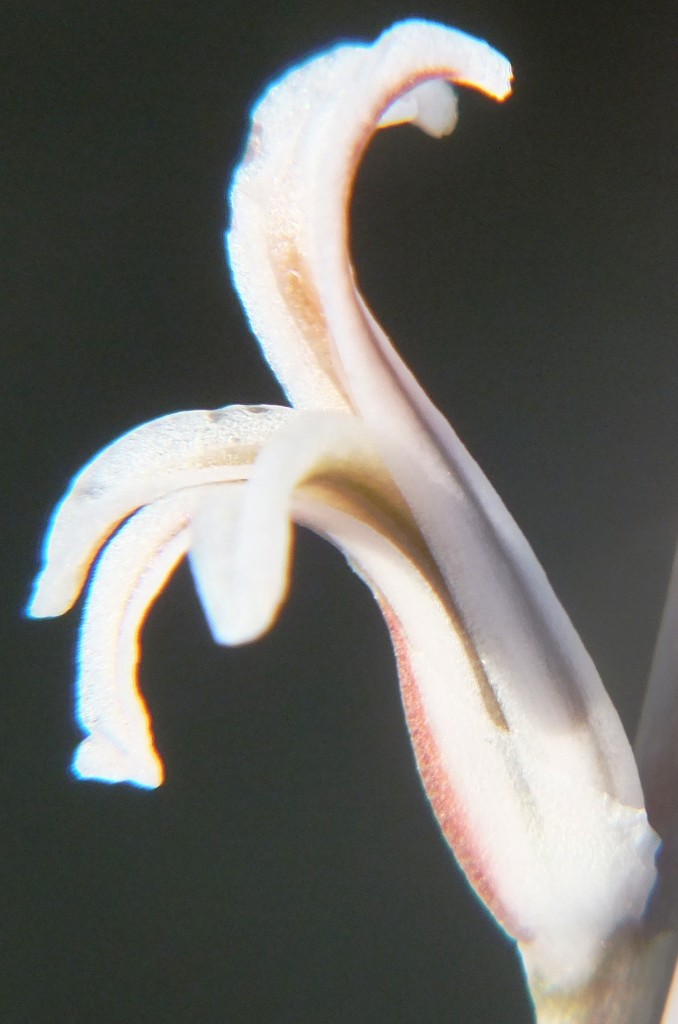
Rosette stemless. non-proliferous, 6-8cm φ. Leaves 12-15, retused, blunt-tipped, brownish-green, in habitat developing purplish cloudiness, barely pellucid with several longitudinal lines. Inflorescence simple, to 20cm. Flowers white with brownish veins.
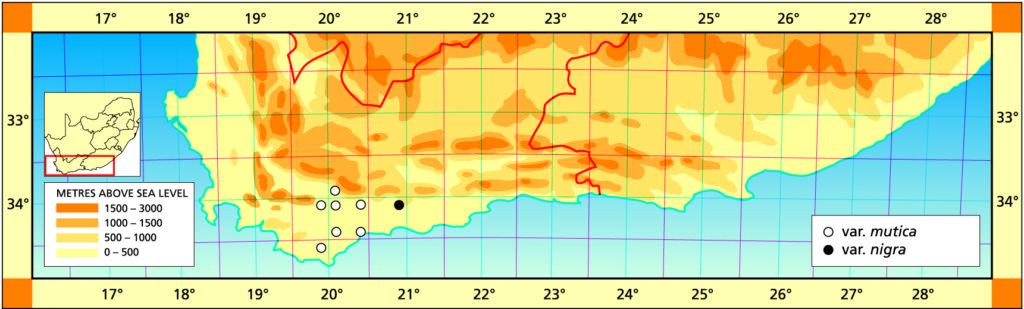
1982 – The name mutica is suggested by the leaf tips which are rounded. This species is very similar to H. retusa and if the two species do meet it can be expected to be in the area between Heidelberg and the lower Breede River. H. mutica is only known west of the Breede River south of Swellendam, but it occurs at one locality north of the river at Drew. In the field the venation and leaf colour is slightly purplish. It is interesting to note that it is very often difficult to distinguish between smooth forms of H. pygmaea and H. mutica. In one comparison of the flowers the only difference that could be detected was that the bracts were purplish‑veined in H. mutica as opposed to greenish in H. pygmaea. H. mutica leads a rather precarious existence in an intensively farmed area, and survives on rocky shale ridges. It covers nearly the same distribution range as H. mirabilis but the two species never appear to grow in the same habitats. H. mutica is generally more glaucous than H. retusa and has a characteristic bluish‑brown coloration.
1999 – A relationship has always been sought between this species and H. retusa. However, a more appropriate solution lies in the recognition that H. retusa should either be enlarged to encompass H. turgida and all its variants, or regarded as a separate species with no interaction with others in terms of its range or distribution – a difficult decision.
a. var. mutica. This is the element west of the Breede river. It was once known from northwest of Drew (north of the Breede River) from where it was collected and sold in pre-war times. That locality has since been taken in by wheat fields, and a single solitary clone survives in the Karoo Botanic garden collection. Leaves of this clone have been propagated. It is remarkable for the white opacity which has subsequently developed in the leaves, and which is also appearing in offsets of the original clone.
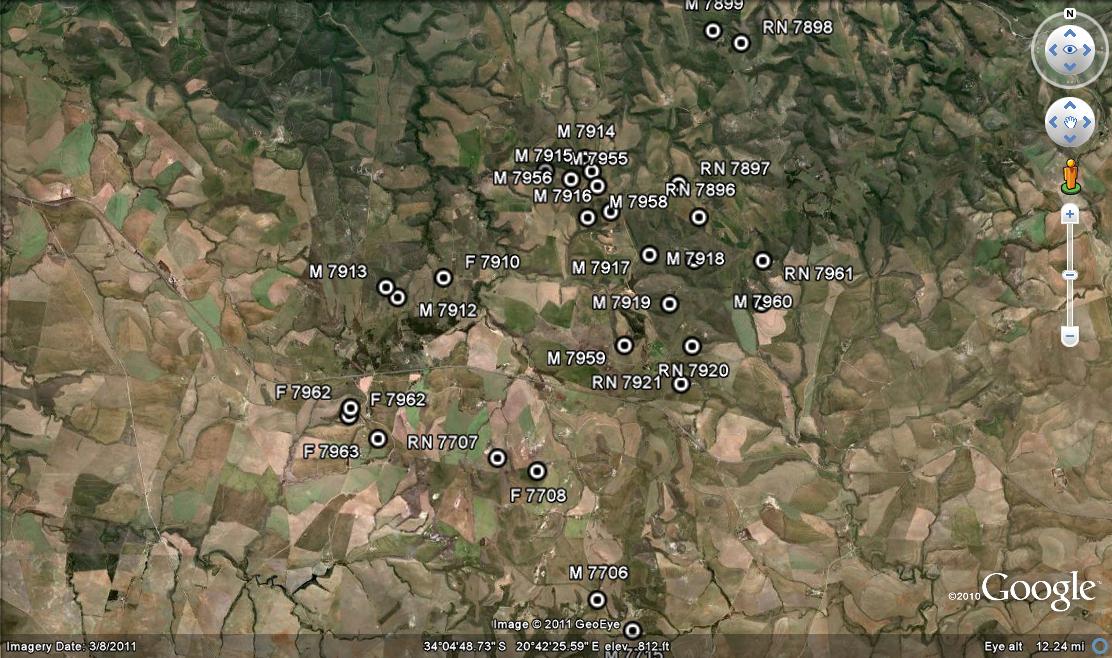
Distribution: 3320 (Montagu): N. Drew (-CC), Smith 5614 (NBG); Drew (-CC), Fouche in PRE 34916. 3419 (Caledon): E. Riviersonderend (-BB), Otzen 6 in Smith 5478 (NBG); 10km E. Riviersonderend (-BB), Smith 3270, 3460, 3970, 5478 (NBG), Bayer 4479 (NBG); Klipdale (-DB), (NBG). 3420 (Bredasdorp): E. Stormsvlei (-AA), Smith 3249 (NBG); Noukloof (-AA), Bruyns (NBG); Rietfontein (-AA), Bayer 2526 (NBG); Crodini (-AB), Bayer (NBG); Soesriver (-AC), Bayer 4473, in KG623/69 (NBG); Kykoedie (-AC), Bayer in KG83/77 (NBG); N. Kykoedie (-AC), Bayer in KG327/71 (NBG); Badjieskraal (-AC), Bayer in KG85/72 (NBG); Haarwegskloof (-AD) Venter 2 (NBG).
Inadequately located: ex hort, Malherbe, Smith 3430 (NBG); Caledon, Hurling & Neil (BOL); Bredasdorp, Theunissen (BOL).
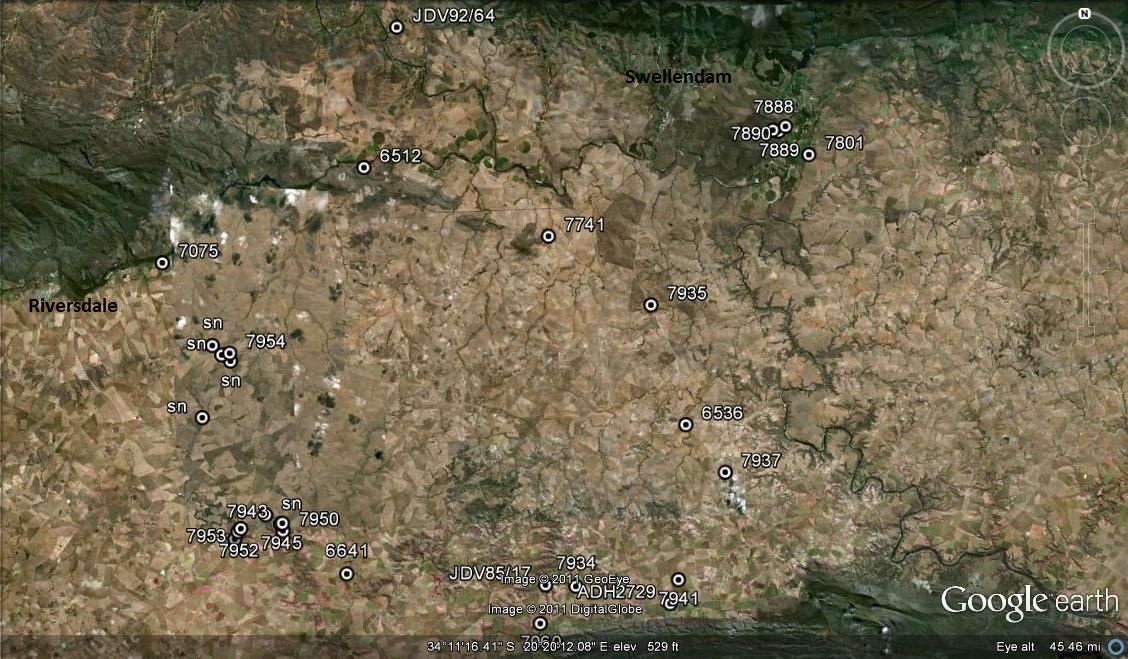
Haworthia mutica var. mutica JDV92/64 Drew. The only clone known to have survived avaricious collecting. Collected in 1970, it has developed a curious and progressive white cloudiness in the leaves – also observed, albeit rarely in other collections.
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. This species flowers earlier than H. retusa.
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. In the field the plants have a purplish waxy bloom to the leaves.
b. var. nigra var. nov. Type: CAPE-3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG, Holo.).
nigra: black.
Differs from the type in its very dark coloration and its occurrence south and east of Heidelberg. (A var. mutica colore perfuscato differt).
This variety from Kransriviermond has been known for a long time and was distributed as H. retusa. It is very dark green, although this does depend on growing conditions as under softer light it may be quite green. There are also populations, probably of this variety, just east of Heidelberg.
Distribution: 3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG); Heidelberg (-BB), Smith 5509 (NBG); E. Heidelberg (-BB), Smith 5755 (NBG); 2km W. Heidelberg (-BB), Smith 6196, 6567 (NBG); N. Heidelberg (-BB), Smith 5509 (PRE).
Haworthia mutica var. nigra JDV86/16 south of Heidelberg. Less wax-like and dark green.
Haworthia mutica var. nigra JDV90/1 east of Heidelberg. There are three such populations known which seem to link H. mutica to H. turgida and to H. retusa.
During my early years with the Karoo Botanic Garden, in fact it never really got any better, my time spent looking for Haworthia was largely my own. This meant hasty weekend trips, or looking for Haworthia secondarily to other things and more general goals. Also it was a question of scale. I needed to see populations and plants from across the entire distribution range. So those years were spent rather as reconnaissance and in checking all the herbarium and other records. I came across a letter among Major F.R. Long’s papers which put me on the track of a Mr. P.L. Meiring of Bonnievale, south-east of Robertson. This was a copy of a letter from Meiring to Triebner in Windhoek arranging for the collection and purchase of an Haworthia from Drew Station. The result was that 200 plants were sold for the huge price (for those days) of 1s each – allowing for inflation this would have been worth about R14ea in today’s currency.
The plants must have been referred to as H. mutica, or this is simply the general name I was using for what appeared to be a fairly distinctive element south of the Riviersonderend mountains and west of the Breede River. Meiring indicated to me where he had collected the plants and I stopped hastily in that general vicinity he indicated northwest of Drew station. A hurried excursion into the veld and after a short while, I picked up a branch of Renosterbos, and there was a plant. Having confirmed the site, I most uncharacteristically collected this single specimen – something I have never done again. The plant remained in cultivation at the Karoo Garden and is probably still there, while leaf propagules have found their way around the world. Kobus Venter obtained one of these and also still has it in cultivation (Fig.1 JDV92/64). When he started taking an interest in Haworthia in 1985, he set off to look for this mutica. Alas, no success. The site had been cleared and ploughed for a massive centre-point irrigation system. We have many times explored the nearby disturbed and undisturbed patches near there, finding only the odd H. maraisii / heidelbergensis plants and a few H. minima var. poellnitziana. The occurrence of these has given us some hope that H. mutica may still be surviving somewhere there.
Kobus and I discussed the issue and when his plant came into bud, we asked the new owners of Sanddrift, Mr. and Mrs. J. Humby, if we could place the plant in the veld in the vain hope that it would be pollinated. The plant was duly taken there, still in its small pot, and sunk into the ground under the protection of three scraggly Renosterbos plants. When I called there late in September the plant was looking healthy but the developing flower was rather scrawny and too shaded. I broke away a few branches to open it up a little. Late in October I called back and to my astonishment there were three seed capsules. The plant was gleefully taken back to Cape Town and I collected 120 seeds from the three capsules. In my joy I planted half of the seed at once. Alas! It was too early in the summer and too hot for this winter regime dweller. Not learning, I planted another 20 seeds a month later and now have 6 rather spindly seedlings and 20 seeds still to sow.
Sanddrift is a remarkable place as three of the four species of Robustipedunculares grow there in discrete populations, and there are hybrids between both marginata and pumila, and marginata and minima. It is unclear just what relationship heidelbergensis and maraisii enjoy there because elements of both are present too. But now we know that mutica is still there somewhere and we have seed to prove it. The nearest other population is south of the Riviersonderend River more than 15km away (Fig.2a, b & c JDV96/27). Here the plants do not have the purplish-grey tones of the species as I would generalise for it south of the Riviersonderend mountains – but then I never have with any conviction claimed that it is different to H. retusa or H. pygmaea. One of the few specimens from there also evidences the whiteness in the leaves which manifested in the Drew plant many years after its collection. The exposure of our potted plant to its native air and a deposit of dry renosterbos leaves, seem to have given the plant a rejuvenating kick and the accompanying illustration shows how beautiful it looks for its visit home. Sept. 2001 is soon upon us and methinks the White Widow needs the company of her soul mate again. (Note:- in a later Chapter, I describe the rediscovery of H. mutica at Sanddrift, Drew).
Acknowledgement To Kobus of course for his deep interest in anything going for Haworthia; and then to Beth Humby whose love for anything to do with life and with nature is extraordinary. It was wonderful to have the co-operation of persons who are so enthusiastic and interested. ♦
Fig. 1. JDV02-64 Sanddrift DrewFig. 2a. 96-27 H. mutica. SE Drew.Fig. 2b. JDV96-27 H. mutica. SE Drew.Fig. 2c. JDV96-27 H. mutica. Drew.
When I wrote the first part of this essay, I was anticipating completing it in three parts. However, I was also in the process of exploring more widely and thoroughly, and the problem and its explanation seemed to grow exponentially. The result was seven essays, and they are presented here as close as possible to their original format. The purpose is to show how a classification should have predictive value, and how an understanding of plants can develop, or fall apart, as more information accumulates. The seven parts were published in Haworthiad 17:1:24-32, 17:2:53-54, 18:1:21-33, 18:2:52-57, 18:3:92-101.
Part 1: Haworthia mutica var nigra indeed!
An early problem in my career as a Haworthia taxonomist was that I had plants of H. mutica which I could not distinguish from plants of H. pygmaea. The former is from the Caledon/Bredasdorp area in the west, and the latter from Albertinia/Mossel Bay in the east. I had also had a collection between these, from Kransriviermond south of Heidelberg, which did not fit comfortably with my perception of H. retusa at Riversdale, or with either that of H. mutica or H. pygmaea. I was also stuck with the probability that there could be other continuities by virtue of unrecorded and unknown populations. A known troublesome population was that from south of the Tradouw Pass that I eventually included in a wider interpretation of H. magnifica, however uncomfortable that was. A problem here is that in separating magnifica from maraisii as species, and in also recognising the variety H. magnifica var. atrofusca, the closer identity of magnifica gets lost in a fragmented residue. As will be seen in the following discussion, this also relates to the understanding of H. mutica var. nigra.
While there was initially a limited amount of material to go by, it might have been thought that more collections would clarify the problems. Instead it became increasingly evident after my first two handbooks that additional collections were confounding the picture even more, and they certainly have. Esterhuizen made the first of such collections from Klipdrift, northwest of Heidelberg. Then there were two populations recorded by Venter, from east of Heidelberg and southeast of Heidelberg. I did not have material of Esterhuizen’s collection but mentally included it with the Tradouw pass population as H. magnifica var. magnifica. The last I included in my concept of H. mutica var. nigra while I was not immediately certain what to do with the population just east of the town of Heidelberg, close to where H. heidelbergensis occurs. I knew these retusa-like plants from photographs and specimen in G.G. Smith’s record. They are very similar to the Klipdrift plants except the habitat is again slightly different and the plants are solitary and the leaves level with the soil surface. It should be noted that I drew attention to this aspect before, when I suggested that H. retusa was in reality a ecotypic adaptation to level ground as is the vegetatively proliferous turgida the adaptation to steep rock faces. The problem was lack of material and the best I could do then was to predict continuity between mutica and retusa. Thus the variety nigra was out of the geographic areas of either and yet not substantial enough to suggest that it was a separate SYSTEM. I have since seen Smith’s plants live as JDV90/113 in cultivation and in the field.
In the last two years I have paid more attention to H. mirabilis and the disconcerting variability of this species. A closer study of the var. sublineata has produced a dismaying degree of variability and I do not doubt that an argument could be made on this score for including the Tradouw Pass plants under H. mirabilis. This is not as improbable as it may seem if all the variants are considered such as in H. heidelbergensis, H. maraisii, H. floribunda. Experience with the Eastern Cape complexes also leads me to predict that these western species are similarly going to be inextricable from one another.
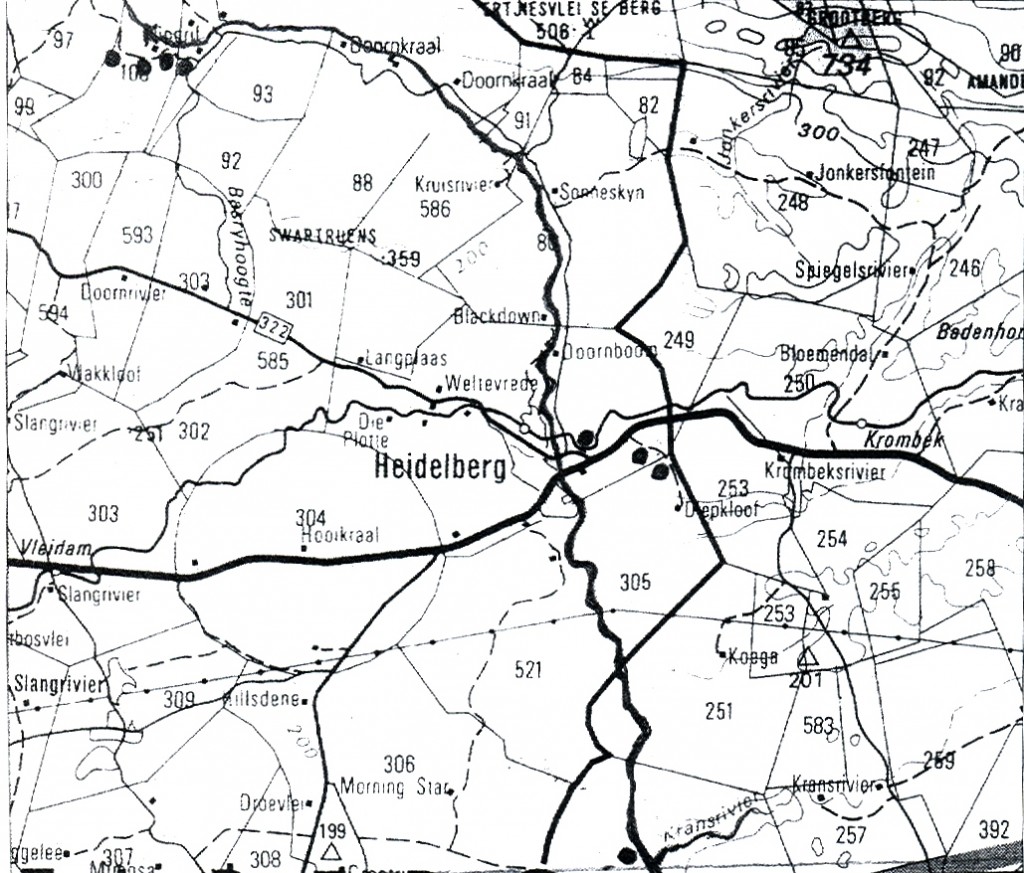
Fig. 01. Map of the Klipdrift-Heidelberg area. Note the farm Blackdown, the type locality for H. floribunda where it hybridises with H. turgida which is common in the area.
Without proposing another solution, other than for the Tradouw plants, I needed to first explain the Esterhuizen population. Here I was fortunate to have the interest and hospitality of Dr. Christie Venter who grew up on a farm just to the northwest of Klipdrift. He now has a property near Zuurbraak where he accommodated my wife and I and we were able to explore the Klipdrift area (see map, Fig.1) to good effect. We found four populations along the southern side of the Duiwenhoks River, two on Goedehoop (Figs.2 & 3) to the west of Klipdrift, one on Klipdrift itself (Figs.4, 5 & 6) and another on Doornriver to the east (Fig.7). From there we went to the two sites east and southeast of Heidelberg (Figs. 8, 9 & 10), as well as to a further population midway between these two. We also revisited the familiar locality for H. heidelbergensis east of the town.
Fig. 02. MBB7133 H. mutica var niga. Goedehoop, west of Klipdrift. The habitat is a gradual slope with little rock, partly bush covered and with Aloe ferox.Fig. 03. MBB7133 H. mutica var nigra. Goedehoop.Fig. 04. E. Esterhuizen in JDV92-29 H. mutica var nigra. Locality given as Klipdrift.Fig. 05. MBB7134 H. mutica var nigra. SW Klipdrift. Grassy habitat with riverine stone.Fig. 06. MBB7134 H. mutica var nigra. SW Klipdrift. A plant with stubbier leaves.Fig. 07. MBBsn H. mutica var nigra. E Klipdrift. A stony slope with Pteronia incana. Only a few plants on lip of road cutting.Fig. 08. MBB6650 H. mutica var nigra. Diepkloof, SE Heidelberg. Plants here are quite variable but nealry all with rounded leaf-tips.Fig. 10. MBB6650 H. mutica var nigra. Diepkloof, SE Heidelberg. With pointed leaves.
What do these populations suggest and what is the solution? These plants do not fit the bill for H. magnifica because they lack the deep dark colouration of that species or the generally shortened leaves. They do not fit with H. mirabilis because the leaves are generally more recurved, the reverse of the leaves is less spotted, and the surfaces are generally smoother. The geographical location does not agree either. The plants cannot be placed comfortably with H. turgida var. turgida, which is a smaller more spinose and clustering species in sandstones. There is some correspondence to H. turgida var. longibracteata and also to H. retusa. From the latter it differs in having fewer more recurved and less spinose leaves, while from the latter it differs in being smaller, more translucent and with narrower more erect leaves.
There actually does not seem to be a categorical answer, which is just what I would have expected. There seems to be this pointer to the new language we need, where we agree on the meaning of names in a fairly arbitrary way. I suggest that for the present this group of populations of doubtful identity should be treated as H. mutica var. nigra. I am quite conscious of the fact that there is an opposition body that wishes to re-interpret such doubtful elements as even H. asperula, and probably produce an entirely different classification and new names. I respect this point of view and it was Prof. G.F. Smith who stated in the introduction to Breuer’s book The world of Haworthia, Vol. 2, this is a golden age and we are each now able to make our own decision! For communication purposes it would be wise if we tried to agree on one set of names. Simply each of us naming plants as species as they come before us has no intrinsic meaning or message for anyone else. It will work for a closed group in close contact with one another, and all seeing and talking about that limited range of specimens. For me who has seen so many plants over so many years, it just becomes quite meaningless. It means that we have a set of names which has no practical value for anyone outside of that group and nor does it have any value for say biologists or scientists working in other disciplines where the names have to have meaning in the broader sense of biological systems as applicable to all life forms.
Fortunately there seems to be an awakening in the botanical fraternity to the fact that our classification system and the notion of a branching tree with the species dangling on the straight branch ends, is faulty. One only has to think what happens when we have a species and then describe a variety of that species. The nomenclatural system requires that a typical variety be automatically created. The implication is that the two varieties stand equal to one another – and this is patently and obviously untrue and unworkable in practise. It actually means that while the variety may be fully distinctive and even be derived from a single always fully recognisable clone, the typical variety has to accommodate every other variant in the species. Thus the more varieties that are described, the more shattered, indefinable and unrecognisable is the “typical” variety. In this is just how it should be if we properly and logically think about distribution of phenomena in space and change with time. Where subconsciously we probably think of species as very definite definable things, the reality is that it may only be individuals (individual clones and vegetatively propagated plants) that have any individual recognisable identity. It is just a myth to say that in a species, the flowers of the individuals are always all similar (identical), or that the leaves have this special character – and so on. If readers are familiar with Haworthia Update Vol.1, they will appreciate that considerably more illustrations are required to truly reflect the relationships of the populations I discuss here and extend this to the broader geographic area i.e. H. heidelbergensis, H. magnifica, H. maraisii, H. mirabilis and others.
A consideration I must add is that the question of very local adaptations to habitat complicates the picture enormously. Thus the complex geology, topography, and climate of the southern Cape are as important as reasons for the extremely rich and variable flora as may be any history of the origin of vegetation from a distant past. Here I am dealing with just the drainage systems of a small river, the Duiwenhoks, which runs from north-west of Heidelberg first towards the south-east and then south to the sea. Other “species” in the close area are H. heidelbergensis, H, turgida and H. magnifica (note that the variety atrofusca thoroughly obscures the identity of the typical variety, and this is further clouded by other varieties, what to say of some of the names now proposed for elements within this huge complex).
Part2: Consequent to Haworthia mutica var nigra indeed!
I omitted illustrating JDV90/113 and of plants from the type locality for H. mutica var. nigra from Part 1, stating simply because these two populations lead from that system to another further east that is more complex. But this is not quite true. What is true is that G.G. Smith was considering the population later collected as JDV90/113 (as a single plant too, no doubt) to be either a new species or a new variety of H. retusa. What is thus difficult to explain is that it is not just a question of how this population related to H. retusa or to H. mutica. It is a problem of how is it related to either of these complexes that have their own internal as well as other external associations (see figs 11 & 12).
Fig. 11. JDV86-16 H. mutica var nigra. KransriviermondFig. 12. JDV86-16 H. mutica var nigra. Kransriviermond.Fig. 13. JDV90-113 H. mutica var nigra. E Heidelberg.Fig. 14. JDV90-113 H. mutica var nigra. E Heidelberg.
In the case of H. mutica var. nigra (Figs. 11 & 12) itself from Kransriviermond, I did not happen to have a picture then available and in any case it is illustrated in Haworthia Revisited. With this short note, the position was rectified and pictures of both collections are provided. It can be noted that JDV90/113 (Figs. 13 & 14) is within 50 meters of the population that I took to be the type locality for H. heidelbergensis. There are not many plants and they are solitary and the leaf surfaces are level with the soil surface. This is the general pose for H. retusa where it grows exposed on level habitat (see figs 13 & 14).
Why have I then not called these plants H. retusa? It is because I considered there was a continuous range of populations that linked H. retusa and H. turgida, and the Kransriviermond population was at variance with this opinion. It is of course equally valid to say that the plants I have illustrated as H. mutica var. nigra could also be regarded as possibly continuous with either H. retusa or H. turgida, as well as include some influence from either H. magnifica or H. heidelbergensis or both.
So what more needs to be said that can be said? An explanation really only begins to make sense when all the role players are examined in detail. H. mutica cannot be represented by a single plant and neither can H. retusa or H. turgida or H. magnifica. Even when one has all these myriad collections in front of one, one wonders what more is there which will cloud the issue further! It is evident that we tend to seek an answer in an easier two-way comparison, when the solution is a much more difficult manifold or reticulate relationship.
Part 3: Still consequent to Haworthia mutica var nigra indeed!
In this part of the essay I discuss what I refer to as “the distant turgida connection”. In the two preceding notes I commented on the problem of using the name retusa in connection with H. mutica var. nigra. It was not unduly surprised to read that Esterhuizen claimed that nigra was rather a variety of retusa than mutica (Alsterworthia). This observation of his is perfectly valid and only misplaced in the context of what I had written before, and as far back as the original Haworthia Handbook. Where the difficulty lies, apart from the intrinsic difficulty of the problem, is with our so-called “typological” concept of species. Thus we form a mental image of, say, H. retusa from an illustration, either the original type illustration: or one that has been chosen to resemble that original. Then we build images around the name of other plants that we come to know as H. retusa. But these images are seldom composite ones that accommodate all the variations and are also limited by how much we have seen. Similarly we then generate images of, in this case, H. turgida and H. mutica and then try to weave them together into a single mental construct.
In this case the introduction H. pygmaea would be somewhat of a surprise. But I did say at the start that one of my very early disconcerting experiences was having plants of H. mutica and H. pygmaea, and finding that I could only separate them on the basis of known origin and the colour of the floral bracts. Without going into the lengthy detail of the variation of H. pygmaea and its associations which include H. turgida and thus to H. retusa, briefly the tale is this: It can be taken as read that the colour of the bracts is not diagnostic for any Haworthia species, where all we then have in this particular instance is geographical information. These single images we build leads to confusion and to misapplication of names. When we think we are specifying, we are actually making very general statements, which in turn help to confound attempts at identification and classification.
Therefore I want here to just illustrate some variants of H. turgida in order to provide a broad image of this species so that its role in the tale can also be guessed at.
I do this for six reasons:-
1. I started a series of articles with the grandiloquent title “Natural variation and Species Delimitation in Haworthia”, for the National Cactus and Succulent Journal way back in 1972. One of the manuscripts I started concerned H. turgida. I had a vast amount of material and I was rather stuck as to how to illustrate it all. It was also obvious that there was a great deal more to see. So the project was shelved until now when this discussion generated the need.
2. While going through my files, I found a few of the old pictures I would have used.
3. I think that readers should be made aware of this major pitfall of perception now that so many writers are in the arena.
4. In addition to H. turgida, the following cannot be positively excluded from reckoning: H. retusa, H. heidelbergensis, H. mutica, H. maraisii, nor H. magnifica. Other authors are very industriously adding new names that will cloud the issue still further.
5. I still would like to see more H. turgida and more H. retusa. I am now quite well positioned in terms of new material, to write about the other elements.
6. The literature is now fragmented beyond sensibility and it is difficult even for me to follow it all and keep a chronological account of what is written there. Other writers have the same problem while they are not even paying proper attention to what has gone before. This is of course their prerogative; and that of the editor who permits his authors to wander where they will.
I do agree with Esterhuizen’s contention that H. mutica has not been found east of the Breede River, which could suggest that the relationship of mutica var. nigra is better sought with H. retusa. However, this would make sense if the typological concept of species was sound and if the concept of the phylogenetic tree with sequential single-branching stems was true. The fact is that one can find the characters that we might use to diagnose any one “species” across the board. Thus I can find “nigra” or similarities, in populations of turgida, retusa, heidelbergensis, magnifica, maraisii and mirabilis. Esterhuizen is quite happy (I have done so myself and it is no criticism) to use the name retusa for plants from near east and even west of Heidelberg, when in fact there is exactly the same problem of “identity” at Riversdale and beyond. This is that retusa is probably an ecotypic variant of turgida. Forget that the name retusa would have precedence over turgida if the taxonomic chips were down.
The first three illustrations (figs 15, 16 & 17) are of H. turgida in a sequence from north to south towards Heidelberg. I choose (chose) this sequence because “turgida” is the sandstone ecotype, and these populations suggested to me that there was a continuous set of populations which linked “turgida” to the more solitary clay-soil, level-ground, silcrete-hill “retusa”.
Fig. 15. MBB in KG240-72 H. turgida var turgida. 9km N Heidelberg.Fig. 16. MBB in KG241-72 H. turgida var turgida. 4km N Heidelberg.Fig. 17. MBB in KG242-72 3km N Heidelberg.
Perhaps I should also emphasise the point that I am using single clones to illustrate populations and systems – where this does not actually work. It is already an approximation. Immediately north of Heidelberg, there is a problem where turgida merges with heidelbergensis. Heidelbergensis itself has its origin in a few plants found beneath one bush, 0,4km West of Heidelberg (I was wrong in following Dekenah’s contention that the type locality was the population 0,4km east, and also heidelbergensis is extraordinarily abundant and variable just west of Heidelberg). It is apparent that the concept of heidelbergensis is based on the interface of two systems viz. turgida and one other. This “other” carries the name “heidelbergensis“, and I will in due course present some of the very extensive ramifications that have emerged in the last few years concerning this particular problem.
The next three illustrations (Figs 18, 19 & 20) are from populations quite some distance from the sandstone range of the Langeberg Mountains (which is the main “turgida” domain), for which I use the varietal name longibracteata. I could not really censure Hayashi for finding new names for any of these. There are several other such non-sandstone populations that I should illustrate, and I know I have not seen them all. I add two illustrations (figs 21 & 22) of the var. suberecta for the eastern forms of “turgida“. To finally knock a nail into the coffin, I add a picture (fig.23) of H. mirabilis var. consanguinea from the Riviersonderend sandstone mountains south of McGregor. Why I name it “mirabilis” is for very good reason, and this reason is as complex as is the full “turgida” saga or that of others. It is thus very frustrating to read simple solutions that are being presented by writers who in my opinion just do not have the competencies or material to do so authoritatively.
Fig. 18. MBB in KG312-71 H. turgida var longibracteata. 16km N Bredasdorp.Fig. 19. MBB in KG94-71 H. turgida var longibracteata. 16km N Bredasdorp.Fig. 20. JDV86-71 H. turgida var longibracteata. Stilbaai, Riversdale.Fig. 21. MBBsn H. turgida var suberecta. Draaihoek, N Albertinia.Fig. 22. MBBsn H. turgida var suberecta. Gouritz Bridge, Albertinia.Fig. 23. MBB in KG332-70 H. mirabilis var consanguinea. S Die Galg, MacGregor.
Part 4: Still on about Haworthia nigra var mutica.
When the previous Part 3 of this small series was published, a few words were omitted from the title. These were, “The distant turgida connection”. This part could have been entitled the close retusa connection. What I was actually looking for was a more definitive explanation and I could not find it. In writing about the relationship of H. floribunda, both Esterhuizen and myself have assumed that the plants I attribute to the species H. variegata viz. the varieties hemicrypta and modesta are in fact that species. It is probably truer to say that they are not and that they are simply part of a much larger complex system for which classification solutions are right for one part of the distribution range and wrong for another. I say this because in this article I will be using the name heidelbergensis in the same way (in the text all references will be in terms of the typical variety. Technically there is no population of this species in which the plants all generally resemble those from which the type was supposedly derived).
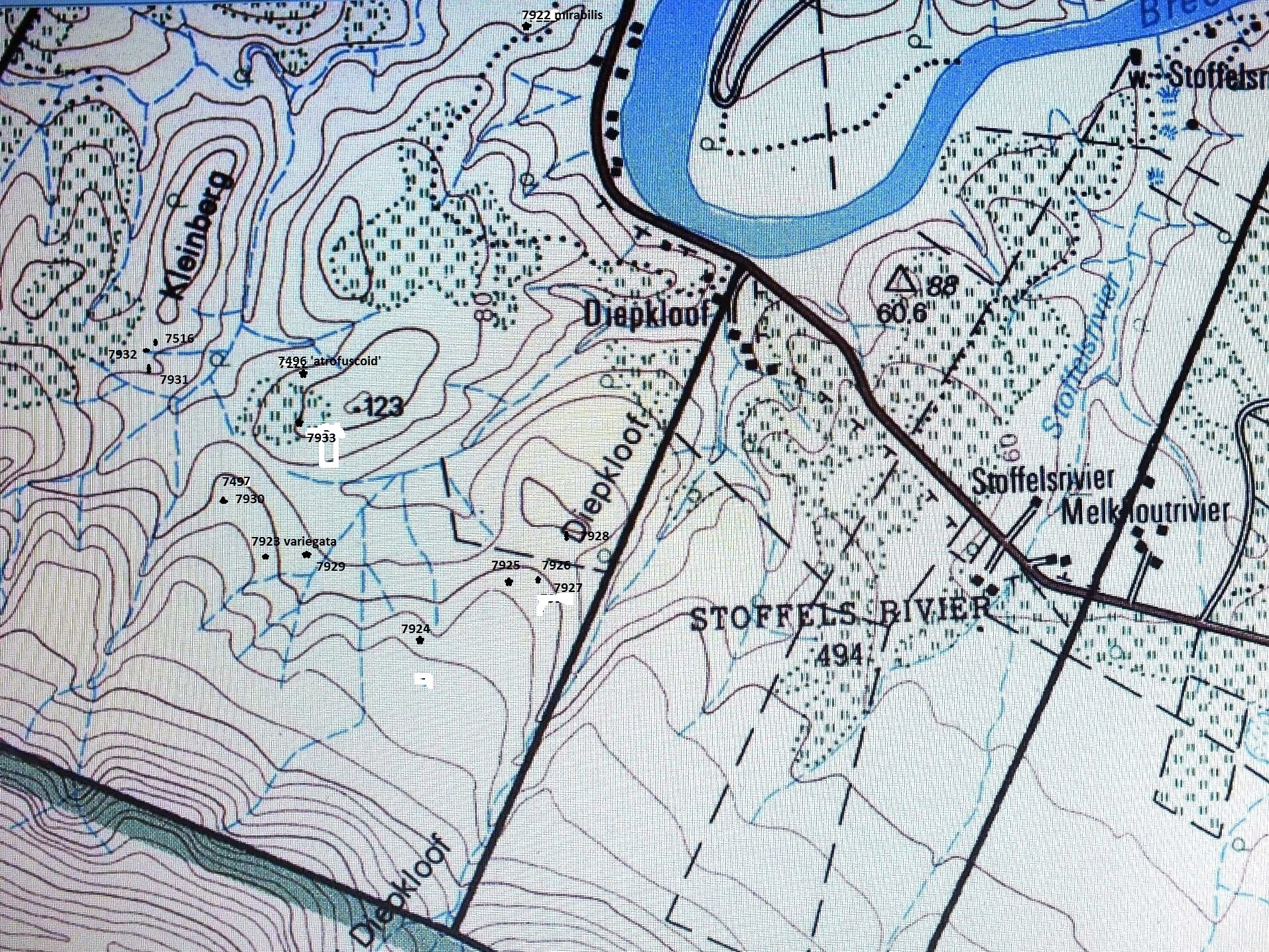
What I want to write about is a series of populations from east of Heidelberg that really throws a cat among the pigeons. There is a low flat plateau between Heidelberg and Riversdale named Kiewietskraalse Vlakte (to be referred to simply as Vlakte) at an altitude of 300m (see map Fig. 24). It is divided into three farms, Kweekkraal East, Kweekkraal West and Melkboom further west. The area to the east, north and west is very broken and generally unsuitable for cultivation. Southwards the terrain is lower-lying at about 150m and is largely cultivated. The pattern of cultivation suggests that the soils in that area are arable and hence were never really suitable habitat for Haworthia. This is an assumption based on where haworthias are found in the greater area and this is invariably on sites where the soils are very skeletal and where rock is exposed. Often these sites are on broken profiles of erosion channels or small hillsides, perhaps for the presence of harder and less erodable rock. This is not to say that there were no suitable habitats on the level areas and certainly the four species of the Robustipedunculares are/were less confined by habitat requirements than members of the subgenus Haworthia. Assumptions about occurrence and suitable habitat are important because they relate to the degree of isolation of populations, to the role of geology in the evolution of ecotypes and hence to the taxonomic relationship of those populations. My impression is that cultivation has generally not impacted on Haworthia populations.
On Vlakte the vegetation is Renosterveld. It has numerous buttresses and surrounding it are a few separate inselbergs. The upper level areas are cultivated and where the natural vegetation is still intact, haworthias are still only found in very specific and identifiable habitats. The eastern-most inselberg is Spitzkop that is the type locality for H. magnifica var. atrofusca. H. retusa in its classic form occurs there too. H. magnifica var magnifica, in its classic form, is still further east at south of Riversdale itself. H. floribunda is known at Langkloof to the north-east, but otherwise there is very little available information regarding what lies between Vlakte and Heidelberg where H. mutica var. nigra, H. heidelbergenis, and the typical true form of H. floribunda occur.
My attention was first drawn to Melkboom by Kobus Venter who told me that there was a complex interaction there between several species (retusa, heidelbergensis and magnifica). He was with J.M. Esterhuizen who has also commented on what I recall was the relationship there of H. magnifica and H. heidelbergensis. I visited Melkboom with Kobus and indeed saw what I will refer to as heidelbergenis. At one point (squared brackets indicate position on the map Fig.24) it ([1] Figs.25a, b & c MBB7108) was growing near to but discrete from H. retusa ([2]Figs.26a, b & c MBB7107). These retusa-plants have smooth leaves with very rounded tips and in this respect hence incline towards H. mutica var. nigra. But on the extreme west wing of the Vlakte plateaux there is an apparent hybrid population of retusa ([3]Fig.27a, b JDV93-53, & c MBB7110) between two populations of heidelbergensis ([4]Fig.28a & b MBB71109, c, d, e, f & g JDV93-54, and [5]Fig 29a MBB7111 b & c JDV93-52). This small population is curious because it is situated between and yet so close, to populations of only one-putative parent. The other parent, presumably H. retusa, is absent. The fact of hybridity is not proven and is only speculative based on the spatial relationships and physical appearance of the plants as judged to be “intermediate”.
Fig. 25a. MBB7108 H. heidelbergensis. Melkboom [1].Fig. 26a. MBB7107 H. retusa. Melkboom [2].ig. 27a. JDV93-53 H. Xretusa. Melkboom [3].Fig. 28a. MBB7109 H. heidelbergensis. Kleinkragga [4].Fig. 29a. MBB7111 H. heidelbergensis. Melkboom [5].Fig. 29b. JDV93-52 H. heidelbergensis. Melkboom[5].Fig. 29c. JDV93-52 H. heidelbergensis. Melkboom [5].
I had now seen H. magnifica at Kweekkraal East ([6]Fig.30a, b & c MBB6817) and West ([7]Fig.31a, b & c MBB6662), so was anxious to see what else occurred on the Vlakte. Those magnificas I had grown from seed, and I knew they consisted of a motley assemblage of forms between the more typical magnifica and atrofusca. What was the relationship then of these populations to the smaller heielbergensis plants at Melkboom? At this stage I never considered that H. floribunda would enter the picture.
Fig. 30a. MBB6817 H. magnifica. Kweekkraal E [6].Fig. 31a. MBB6662 H. magnifica. Kweekkraal W [7].
Kobus came along with my wife and I to re-visit the area and explore between Melkboom and Kweekkraal West. The Vlakte Plateau has an irregular shape and has many buttresses separated by incised small valleys. The upper edges of these buttresses often comprise fairly skeletal soils with rock that probably accounts for irregular erosion of the plateau edge. The rock also seems to overly erodable clay and this is exposed by pressure bursts which result in a low clay bank and an extended apron of bare clay soil with here and there tufts of grass or low herbaceous plants. The valleys are more thickly and densely vegetated as the soil are deeper and retain water for longer. The rock and clay sites provide the habitat for the haworthias and nearly every buttress is populated. Thus we found a confounding array of plants which all seemed to indicate a continuum from the heidelbergensis-like plants on Melkboom to the magnifica-like plants at Kweekkkraal. The populations were disturbingly variable and yet differed also from one another. The description and quantification of such “difference” is probably an ultimate challenge to botany. The collections are [8]Fig.32a & b MBB7112, [9]Fig.33a, b & c MBB7113, [10]Fig.34a & b MBB7114, [11]Fig.35. MBB7115, [12]Fig.36. MBB7116, [13]Fig.37. MBB7117, [14]Fig.38a & b MBB7118.
Fig. 32a. MBB7112 H. heidelbergensis. Kweekkraal W[8].Fig. 32b. MBB7112 H. heidelbergensis. Kweekkraal W [8].Fig. 33a. MBB713 H. heidelbergensis. Kweekkraal W[9].Fig. 34a. MBB7114 H. heidelbergensis. Kweekkraal W [10].Fig. 35. MBB7115 H. heidelbergensis. Kweekkraal W [11].Fig. 36. MBB7116 H. heidelbergensis. Kweekkraal W [12].Fig. 37. MBB7117 H. heidelbergensis. Kweekkraal W [13].Fig. 38a. MBB7118 H. heidelbergensis. Kweekkraal W [14]
My wife and I returned to the area East twice after that to further explore the plateaux. We first made these collections on Kweekkraal West [15]Fig.39a & b MBB7151, [16]Fig.40 MBB7152, [17]Fig.41a, b &c MBB7153, Fig.42a, b, c, & d MBB7154. The first two I regarded as heidelbergensis, the third inclined to magnifica, while the fourth was more positively magnifica including clones which would class as var atrofusca. On Kweekkraal East, significant finds were a population south of the Pretoriuskop inselberg consisting almost entirely of the var. atrofusca ([19]Fig.43a, b, c & d MBB7157) as opposed to several of the heidelbergensis/magnifica continuum we had seen on Kweekkkraal West. We also found H. floribunda on Pretoriuskop [21], southwest of (Fig.44 MBB7155) as well as on an unnamed inselberg to the west ([20]Fig.45 MBB7156). Exciting was that in the population of atrofusca, which was growing in a very white clay layer, was a plant of H. floribunda (MBB7158). It is extremely difficult in these field sites to recognise variants and distinguish what could have been hybrids, and we did not think there were such. However, about 50 meters away we came across a population of small heidebergensis ([22]Fig.46 MBB7159) like plants which we decided were hybrids – based on the fact that another 50 meters away was a population of floribunda. Again this is speculative.
Fig. 39a. MBB7151 H. heidelbergensis. Kweekkraal W[15].Fig. 40. MBB7152 H. heidelbergensis. Kweekkraal W[16].Fig. 41a. MBB7153 H. heidelbergenis=magnifica. Kweekkraal W [17].Fig. 42a. MBB7154 H. magnifica. Kweekkraal W [18].Fig. 43a. MBB7157 H. magnifica var atrofusca. Pretoriuskop[19].Fig. 44. MBB7155 H. floribunda var dentata. SW Pretoriuskop[20].Fig. 45. MBB7156 H. floribunda var dentata. W Pretoriuskop[21].Fig. 46. MBB7159 H. Xfloribunda. N Bosgasiekop[22].Fig. 47a. MBB7160 H. magnifica. N Kweekkraal E[23].Fig. 48a. MBB7161 H. magnifica. N Kweekkral E[24].Fig. 49a. MBB7162 H. magnifica. Far N Kweekkraal E[25].
At other sites we found several other populations that included both small ([23]Fig.47a & b MBB7160) and large magnifica ([24]Fig.48a & b MBB7161, [25] Fig.49 MBB7162) depending often on the nature of the pressure-bursts and the quality of the rock and clay. Often the clay was quite red and the rock very ferruginous (iron-stone). At other sites it was often very white and clean. The rock similarly varies from the very characteristic red-pebbly constituency of true ironstone, to the smooth white appearance of pure quartz.
Fig. 53 Site 24, showing the kind of vegetative cover over a dense population of H. magnifica.
On a still later occasion my wife and I visited Kweekkraal East again with Casper Mazurel of Holland. We had not explored the northern edge of the Vlakte plateau and needed to know more. Casper found H. retusa north of Kweekkraal East ([26]Fig.49 MBB7173) in the same kind of white clay that atrofusca occupied near Pretoriuskop – we also found floribunda again (Fig.49[23] – MBB7172). The curious thing about both the retusa populations from Vlakte, have very rounded leaf-ends (the mutica connection? – more later). The Melkboom population is relatively light-coloured and the more eastern population the deeper green/purple tones of magnifica. The small population at [25](Figs.47 MBB71620) seen on the previous visit (in white clay in which the plants were quite robust) then takes on a new face and could be imagined to be of possible “hybrid” (retusaXmagnifica) origin.
It required yet another visit to flesh out distributions and this was to the inselberg Rooikop. We were not quite sure what to expect and I did think that H. floribunda would appear again. Instead we found a series of small populations of plants I attribute to H. heidelbergensis/magnifica ([28]Fig.50 MBB7217)
Fig. 50. MBB7173 H. retusa. NW Kweekkraal E[26].
As I have noted, variability in and between these populations is enormous. It is curious that H. floribunda only appears in the northeast of the study area and almost totally maintains its integrity where heidelbergensis/magnifica is excluded. The variant H. magnifica var. atrofusca is discrete in only one of these populations (Fig.42 – MBB7157) and this is the one exception where H. floribunda is also present. There seems to be a remarkably smooth continuum from H. magnifica-like to H. heidelbergensis-like plants.
I cannot say at all that Vlakte has been fully explored. There are many more possible habitats that we have not explored. While I am busy examining the evidence we have for a connection of mutica var. nigra to H. retusa, I find myself looking at the connection of H. heidelbergensis to H. magnifica with the implications of hybridisation (introgression is not a better word or concept, as both hybridisation and introgression imply an initial difference) involving H. floribunda and H. magnifica var. atrofusca which have rounded leaf-tips. Thus the saga continued.
Note: Kobus (J.D.) Venter made a presentation involving many of the above collections at the Aloe Congress at Calitzdorp in Sept.2003. This is due to be published in Alsterworthia. In his presentation he includes a geological map of the area and this shows that Vlakte is tertiary ferricrete overlying much older conglomerate rock. It is this ferricrete formation which provides a mosaic of small inselberg’s scattered throughout the southern Cape and playing a critical role in the occurrence of Haworthia).
Fig. 51. MBB7172 H. floribunda var dentata. N Kweekkraal E[27].Fig. 52. MBB7217 H. heidelbergensis. Rooikop, N Kweekkraal.[28].
Part 5: Continuing the saga.
On the way home after our last visit to Kweekkraal, Daphne and I stopped at Morning Star south of Heidelberg. We did this thanks to our friend Dr. Christie Kloppers who in discussing the distribution of H. marginata, let on that the owner of Morning Star was a relative of his. Morning Star is a locality recorded for H. marginata and as I had seen that species in the area many years before, I was anxious to confirm its survival there. As is the case in Haworthia there are so many considerations. Morning Star is not far from the Duiwenhoks river where the steep rocky riverbanks offer a continuum of habitats from north of Heidelberg all the way to the sea. A short way to the southeast is the locality Kransriviermond where H. mutica var. nigra has it origin. It is interesting to note that G.G. Smith made the connection of that population with three different accessions of H. mutica. Which goes to show how perceptions may differ – he had no compunctions about making that association as opposed to the opinions reported in the essay. The place Dassieklip is also on the lower reaches of the Duiwenhoks and this is where a very curious Haworthia is found which I have attributed to both H. floribunda and H. chloracantha. (Fig.54 MBB7164).
Fig. 54. MBB7164 H. floribunda=chloracantha. Dassieklip.
However, there is also something else. This is the collection MBB6663 (Figs.55a also MBB in KG107/74, & b JDV97-35, also Fig.27 in Pt.1) from immediately southwest of Heidelberg. This collection is cited in my revision under H. magnifica and that identification falls very far short of explaining its true identity or significance. I wrote an article in Aloe (34:4, 1997) discussing the problem of the “species” as they occur around Heidelberg. H. magnifica is barely mentioned and this particular collection suggested some introgression of H. floribunda. However, since that time I have come across several other populations to the west that must surely relate to this one at Heidelberg and impact on the relationship of H. magnifica to H. maraisii and H. mirabilis. Thus anything of this ilk to the south of Heidelberg becomes extremely important. That collection MBB6663 was instrumental in my early decision to treat H. maraisii and H. magnifica as one species
Fig. 55a. MBB6663 H. magnifica. SW Heidelberg.Fig. 55b. JDV97-53 H. magnifica. SW Heidelberg.
At Morning Star we were, by kind favour of Mr Coetzee Uys, able to get to the Duiwenhoks River. There we found H. turgida (Fig.56 MBB7219) on the rocks of the valley banks as we expected. It was surprisingly a rather small form (var. turgida) more reminiscent of its appearance on the sandstones north of Heidelberg, rather than the bigger var. longibracteata. But significantly we also found, in the immediately adjacent south-facing shale rocks, H. magnifica/floribunda (Fig. 57a & b MBB7218) as in MBB6663. The plants were smaller than that collection and there was more evidence of floribunda AND heidelbergensis in their appearance. We stopped and searched again about 3kms away on higher-lying ferricrete and in grass tufts, where we found plants of the same ilk (Fig.58a & b MBB7220). These two populations suggest fairly convincingly that H. magnifica and H. floribunda introgress (are continuous).
Fig. 56. MBB7219 H. turgida var turgida. Morning Star, S Heidelberg.Fig. 57a. MBB7218 H. floribunda=magnifica. Morning Star, S Heidelberg.Fig. 58a. MBB7220 H. floribunda=magnifica. Morning Star, S Heidelberg.
We were not finished. As a last effort we drove a short distance and stopped next to some exposed rock where we found H. mutica var. nigra (Fig 59a & b MBB7221). These are very dark-green almost black, shiny-leaved plants with the rounded leaf-tips we associate with H. mutica. One of the clones was almost identical to a plant of H. retusa (so-named) from north of Kweekkraal (MBB7173) where one could speculate that the rounded leaf-tips and darker colouration had some connection to introgression (that word again) with H. magnifica var. atrofusca. The second plant illustrated (Fig.59b) is somewhat of an aberration that I selected to emphasise that identifications are often very tenuous.
Fig. 59a MBB7221 H mutica var. nigra, Morning Star, S HeidelbergFig. 59b. MBB7221 H. mutica var nigra. Morning Star, S Heidelberg.
In my discussion of H. incurvula in Aloe ( 36:34, 1999), I suggest we need a new language to discuss, explain and name haworthias. We need a better conceptualisation of names and what they mean. In taxonomic botany, the name of a species is linked to a “type’ which is desirably a herbarium specimen, but it could be an illustration of some kind. It is often simply assumed, perhaps even unconsciously, that this “type” is typical. We all (nearly all) assume that anything so named will be identical or near identical to that type. Conversely we assume that anything different from the type should have another name and ipso facto be another species. The concepts of subspecies and varieties are an extension of this way of thinking. The nomenclatural code which seeks uniformity and stability in the way we name plants cannot legislate or prescribe when and where names should be applied and nor at what rank. Where it does seem to fall short is the requirement for the automatic creation of a typical rank when an equivalent rank is created. Here it seems to be making the same assumption that all plants of a species are the same. Thus when a new, say, variety is described, there is a typical variety that stands in juxtaposition to the new. This is patently fallacious. What is required is a re-description of the species to include the variation of the new lesser rank, and only those individuals which meet the varietal description become identified by an added name. The residue forms the core of the species and are designated by only that name?
What we need in addition, is the adoption of a definition of a species as a system of living organisms which is named by a convention recognised as a language requirement, and not as any scientific process or achievement. The name is only the passkey to information – the key by which we generalise about a particular living system. (I have since written a chapter which suggest that species are fractal, and that we will need to adapt our thinking to this kind of geometric/mathematical pattern).
Part 6: Another twist to the story.
Until recently I had seen very little of the haworthias immediately west of Heidelberg. Kobus Venter has several collections, some by J Esterhuizen, and these included plants from Uitkyk immediately southwest of the town (Fig. 60a & b JDV89/2) as well as from further west. He also has two collections from immediately north of the town where previously I have referred to only one. Taking these first, I think it is quite clear than they both suggest the continuity of turgida with heidelbergensis, while one of the collections suggests a closer similarity to the plants from Klipdrift of Part 1. The Esterhuizen collection from northwest of Heidelberg (Fig.61 JDVsn) seems to support such a contention.
Fig. 60a. JDV89-2 H. heidelbergensis. Uitkyk, SW Heidelberg.Fig. 60b. JDV89-2 H. heidelbergensis. Uitkyk, SW Heidelberg.Fig. 61. JDVsn H. heidelbergensis. NW Heidelberg.
But I really want to deal with plants west of Heidelberg. The locality is referred to as Die Plotte but actually the area is partly township land, an area of smallholdings and sections of what was a very large farm Hooikraal. Dekenah and thus G.G. Smith original site of heidelbergensis may be non-existent now because of the creation of a rubbish-dump site immediately west of the town. Plants similar to the original heidelbergensis can still be found southwest and further west (and of course immediately east of Heidelberg which I had concluded (wrongly and influenced by Dekenah) was the type locality. Kobus’ collection from Die Plotte (Figs. 62a, b & c JDV91/142) are of smaller plants while JDV91/146 (Figs.63a & b) from the same general area, are bigger plants. These are intermediate with the Esterhuizen plants northwest of, and typical heidelbergensis from east of Heidelberg (Figs. 64a, b JDV87/1 & c MBBsn).
Fig. 62a. JDV91/143 H. heidelbergensis. Die Plotte, W Heidelberg.Fig. 63a. JDV93-146 H. heidelbergensis. Die Plotte, W Heidelberg.Fig. 64a. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 64c. MBBsn H. heidelbergensis. E Heidelberg.
When Kobus delivered his paper at the Aloe Congress 2003 at Calitzdorp he met up with Mr. J. Hoffman. I had met him at Worcester many years before and he had told me of plants west of Heidelberg. Thus I undertook to renew this contact and went to see him at Heidelberg. He took me to the northeastern corner of the farm Hooikraal which borders on Die Plotte and which is also west of the town dumpsite. Apart from showing me H. marginata, he also showed me a population of small heidelbergensis (Figs. 65a, b, c & d MBB7233) and then a population of very large plants that were quite remarkable (Figs. 66a, b, c & d MBB7234). The plants included one that even resembled a large sturdy mirabilis – and Esterhuizen commented in an article he once wrote about the spotting on the back of the leaves of “heidelbergensis” which he likened to mirabilis. Generally the plants are with darker green and purplish hues that I associate with heidelbergensis or even magnifica. There were very yellow-green forms similar to what I saw many years ago at Skeiding further west of Heidelberg. These are plants that Esterhuizen (and myself too) has referred to as retusa, and I would agree except that they fall in the query zone between retusa and turgida var longibracteata i.e. there is continuity between the two species. In fact Kobus and I revisited Skeiding and I thought the plants (Figs.67a & b MBB7240) were almost identical to H. retusa at Melkboom (Fig.26 MBB7107 in Part 4), although the latter tended to have rounded leaf-tips. H. magnifica, MBB7239, was also present in a rare situation for these two species to be so closely located within meters of each other). We also visited a population immediately north of Heidelberg (Figs. 68a & b MBB7236) that is not far distant from the two Nature Reserve population already referred to. Had I seen this population in 1975, I may have been more adamant in maintaining that H. retusa and H. turgida were but one species. The plants were generally big and very proliferous and one could not help reflecting on how similar they were the population that gave rise to Scott’s H. geraldii (H. retusa var.). The collection from northwest of the Heidelberg Nature Reserve (Figs. 69a, b & c JDV89/5) must surely suggest that there is a continuity of H. turgida with the Klipdrift plants illustrated in part 1.
Fig. 65a. MBB7233 H. heidelbefrgensis. N. Hooikraal, SW Heidelberg.Fig. 66a. MBB7234 H. heidelbergensis. S Hooikraal, SW Heidelberg.Fig. 67a. MBB7240 H. retusa. Skeiding, W Heidelberg.Fig. 68a. MBB7236 H. turgida=retusa. N. Heidelberg.Fig. 69a. JDV89-5 H. turgida=retusa. N Heidelberg.
My conclusion is that there is now a proven continuity between turgida, heidelbergensis, magnifica, retusa, mutica var. nigra, maraisii, pygmaea and even floribunda, chloracantha, rossouwii and variegata. The continuity is also with emelyae and its varieties north of the Langeberg mountains. At the same time some of these elements may occur together and so appear to be discrete. Thus a taxonomic solution for one set of collections may not be appropriate for another.
Paul Forster and Russel Scott (both from Australia) spent some time in South Africa while attending the Calitzdorp Congress, and I was privileged to have them join me in the field at Heidelberg. We explored the middle Duiwenhoks river between Dassieklip and the town of Heidelberg. I had expected that such exploration would throw some light on the strange Dassieklip population (Fig.56 MBB7164). In my Revision (1999), I have ascribed it to H. floribunda simply for the want of a better solution outside of regarding it as a discrete species based on a single population!).
Apart from seeing H. turgida we also found find populations of plants at Witheuwel (Figs. 70a, b & c MBB7227) and Somona (Figs71a, b & c). These are more like the Melkboom and Kweekkraal heidelbergensis/magnifica collections. I would have expected a similarity to MBB6663 (SW Heidelberg), referred to in part 5, figs 2 & 3) or to MBB7218 and MBB7220 which I would ascribe to floribunda/maraisii rather than to H. magnifica – tenuous as such a distinction is. Essie Esterhuizen has apparently discovered a similar population to KG107/74 along the Duiwenhoks close to of Heidelberg. It must be noted that Matjiestoon, the locality for H. heidelbergensis var toonensis (Figs.72a, b, c & d MBB7165) is only about 15km west of Dassieklip. Thus while I am writing in the context of H. mutica var. nigra, I have drifted helplessly into discussing H. heidelbergensis. This is the crux of the whole thing about Haworthia – especially the subgenus Haworthia – an intricate network of relationships that just will not resolve.
Fig. 70a. MBB7227 H. heidelbergensis. Witheuwel, S Heidelberg.Fig.-71a.-MBB7229-H.-heidelbergensis.-Somona-S-Heidelberg.Fig.-72a.-MBB7165-H.-heidelbergensis-var-toonensis.-Matjiestoon.-SW-Heidelberg.
Part 7: Completing the saga.
So far in this series of articles I have dealt with H. mutica var. nigra as a system of populations typified from the southernmost population at Kransriviermond, and extending through eight known populations to Klipdrift northwest of Heidelberg. Then I considered the role of H. turgida in part 3. In part 4 I dealt with the very complex situation that seems to occur between Riversdale and Heidelberg, followed in part 5 by discussion of populations along the lower Duiwenhoks River. In part 6 the discussion and illustrations are of plants from mostly west of Heidelberg.I have discussed to some degree the possible relationship of these populations to each other to H. turgida, H. retusa, H. heidelbergensis and H. magnifica. Each of those systems in turn interacts with others and it becomes highly doubtful that we are in fact dealing with discrete systems at all.
If we now switch the attention to H. mutica and point to some of the populations that constitute my concept of that species, it may help to crystallize aspects of my classification. I will not deal with all the material at my disposal as is too extensive for the purpose. The name H. mutica is based on an illustration in the Royal Herbarium, Kew, of a plant of unknown origin (Fig.73). I use the name for a series of populations all west of the Breede River, from near Drew in the north, to near Bredasdorp in the south. These populations all seem to be discrete from other species in that general area, which are H. mirabilis, H. maraisii, H. rossouwii, H. heidelbergensis, H. reticulata and H. turgida. Those species display their own interactions as follows:
Fig. 73. H. mutica. Kew herbarium.
H. mirabilis never occurs in any close association with any other species (of the same subgenus) including H. maraisii or with H. heidelbergensis. Where it may, is where H. mirabilis var. badia occurs in close proximity to H. maraisii at Napier. I can be suggested that here the two elements are introgressive and that var. badia is actually the consequence of this introgression (again the word implies interaction between different things). H. maraisii never grows in association with H. heidelbergensis except for the place Rooivlei north of Bredasdorp where this H. heidelbergensis var. minor could in fact be a variant of H. rossouwii. It should be noted that H. turgida var longibracteata also occurs here as a very isolated population.
If one extends these observations it will be found that very few species maintain their identity in close proximity to any other. H. magnifica may occur in close proximity to H. tugida or H. retusa, but the latter two never to each other. H. magnifica is not discrete in any sense from H. maraisii and two species are recognised simply as a device to assist discussion where we have a western (maraisii) and an eastern (magnifica) counterpart. Neither of these ever occurs near to, or with, H. heidelbergenis. We actually need to again consider this last-named species very closely. It is typified from a population just west of Heidelberg (as Esterhuizen rightly pointed out, and it is by fortunate coincidence that my assumption drawn from Dekenah that it was east, does not alter the basic perception of what the species looks like). H. heidelbergensis has by default, and from simple circumstance and complex reality, become a hodge-podge of populations which interdigitate between H. mirabilis, H. maraisii and H. magnifica. H. floribunda is a curious anomaly that is widely distributed and is known to hybridise with H. turgida, H. magnifica var. atrofusca and with H. pygmaea var. argenteo-maculosa. There is some evidence that it introgresses with (or has never segregated from) H. magnifica (synonymous maraisii) south of Heidelberg, and similarly with H. heidelbergensis (synonymous also with maraisii) south of Swellendam.
H. mutica only grows in the close proximity of H. mirabilis at Rondeheuwel south of Stormsvlei. At Sanddrift, Drew it is in close proximity to H. heidelbergensis in a form which is akin to H. maraisii. Further to the southeast it is in close proximity to H. maraisii. At Napky these two elements are also close.
My argument for maintaining H. mutica as discrete from H. retusa was based on the observation that it was never necessary to invoke H. turgida in the equation as it is when discussing H. retusa. It also is never introgressive with H. maraisii as H. retusa seems to be with H. magnifica east of Riversdale, nor with H. heidelbergensis as H. retusa seems to be at Kweekkraal. Nevertheless this decision is not indisputable. I have always borne in mind that H. mutica has some affinity with H. pygmaea. H. mutica seems to have a retusa-like origin whereas H. pygmaea has a magnifica-like origin. In neither case can one be sure that H. turgida is not prominent in the ancestry.
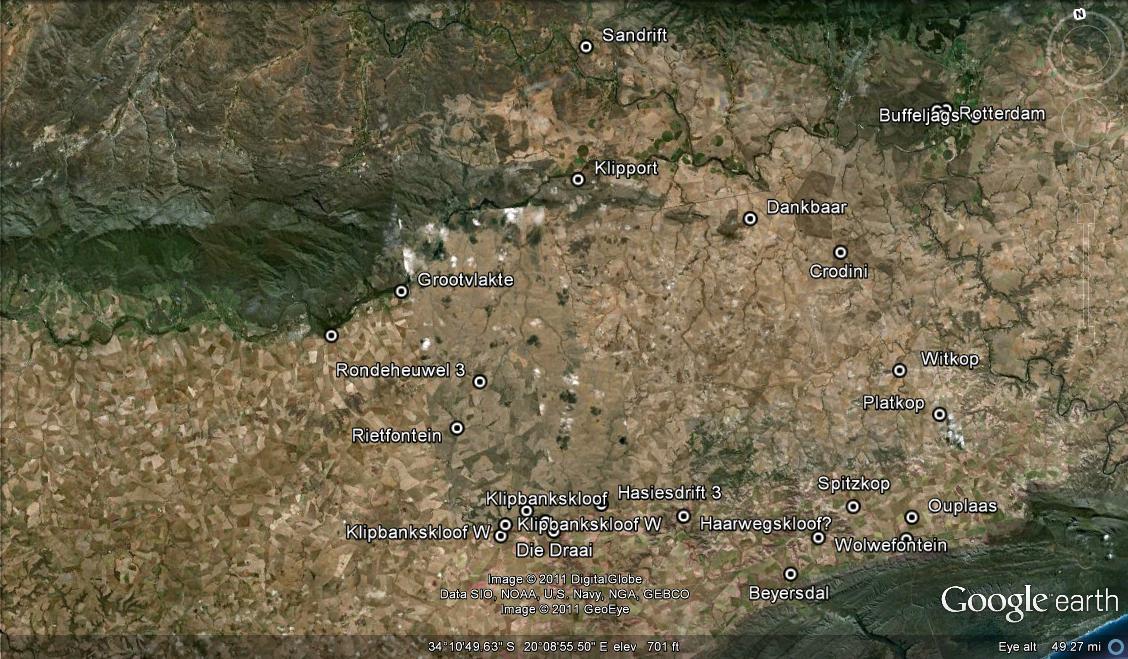
A not-so-curious thing about H. mutica is its variability too. Just like any of the other “species”, it has many faces. The one I would like to mention is the similarity of plants of H. mirabilis var. badia from the western-most known point of its distribution. This is not readily apparent. But is we look at plants at the western end of the population viz. Sandfontein Napier (Figs.74a & b MBB6987), the plants begin to depart from the more classical form with the strongly recurved leaves, and begin to resemble more typical var. mirabilis as at Mierkraal Bredasdorp (Figs.75a & B MBB7090). What has astonished (even) me is that some of the clones form a very close match with H. mutica, especially with south (Figs.76a & b JDV96-16) and north of Napky (MBB7031 respectively). Thus we are now thrown into (an admittedly obscure) relationship of H. mutica and H. mirabilis (there is also a similarity with a clone of H. magnifica from the problematic “south of Tradouw site” (Fig.77 MBB6666). To my knowledge H. mutica and H. mirabilis only occur in close association at one place. This is at Rondeheuwel south of Stormsvlei where the mirabilis is atypical and evidence of the continuity it displays in so many places with H. maraisii. My collection (Fig.78 MBB6982) from Hasiesdrift north of Bredasdorp seems to be a good representation for a concept of H. mutica. The habitat photo (Fig.79), however, raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.
Fig. 74a. MBB6987 H. mirabilis var badia. Sandfontein, Napier.Fig. 75a. MBB7090 H. mirabilis var mirabilis. Mierkraal, Bredasdorp.Fig. 76a. JDV96-16 H. mutica. S Napky, SW Swellendam.Fig. 77. MBB6666 H. magnifica var magnifica! S Tradouw Pass.Fig. 78. MBB6282 H. mutica. Hasiesdrift, N Bredasdorp.Fig. 79. MBB6282 H. mutica. Hasiesdrift habitat picture.Fig. 79. MBB6282 H. mutica. Hasiesdrift habitat picture.
There the saga for the moment must rest. The proper taxonomic place of H. mutica var. nigra sensu Bayer is simply not very significant given the huge problem which is only touched on here. This is the interplay of H. heidelbergensis with H. maraisii, H. magnifica, H. mirabilis, H. turgida, H. retusa, and others. I should say of all the populations (should I count them?), I know which can be attributed to these “species”. ♦
This essay was published in Haworthiad 16:86, 2002.
I wrote an article about Haworthia rossouwii in Aloe 38:31(2001), in which I resurrected this old name to replace that of my own H. serrata. This was necessary because I had found this plant (because of its localisation and its abundance there, it is better to say ‘this species’) at two places near Bredasdorp as opposed to where I had described my species from near Heidelberg.
One needs to know something about the geography and geology of the Southern Cape (and the Overberg as a part of it is also known) to really follow all the ramifications of any discussion about Haworthia, including this one. In fact one needs to know a whole lot more, and I will also try to explain that and its implications for the collector and Haworthiophiles. This “whole lot more”, I will call the Corporate Mind because it includes so much – so remember CM! If I regard H. rossouwii as a species, I have to consider all the plants and all the places where they grow in order to determine the nature of this particular system of living things. As I explained in my article, there is a problem with the fact that little is actually known about the Haworthia of the Overberg. They occur in small populations scattered over a fairly wide area which has been heavily impacted on by agriculture. Thus about 90% or more of the Overberg is now wheatfield or pasture. Like Gasteria carinata, which is also a Southern Cape species, Haworthia is associated with rocky outcrops and thus also with the geographical erosion and drainage systems of the area. It is quite probable that cultivation has had relatively very little impact on Haworthia in terms of actual available and suitable habitat.
The problem is to get to all the small potential habitats and explore these at a scale which will help to resolve the problems of variability, relationships and classification. While writing about H. rossouwii, I was very conscious of this problem and the fact that my explanation is not complete. There are some collections by Chris Burgers which I have not seen, and there are many potential habitats which I have not explored either. The place Kathoek and H. variegata var. modesta which grows there is a particular case in point. Kathoek is at a point midway between the Breede River and Bredasdorp. It is just east of the De Hoop wetland/stream and on the northern edge of the Coastal limestones. In the west, also on limestones, is H. rossouwii var. petrophila which I first placed with H. variegata. In the south (also on coastal limestone) is H. rossouwii var. calcarea, which I originally had with H. mirabilis. To the north is H. variegata var. hemicrypta which is on Silcrete of a quite different geological origin. But H. variegata variants, as I interpret the populations, are also on and about the Potberg to the east which is Cape Sandstone.
My problem is “How do these populations actually relate to one another?” What is missing? What can I find to better explain their similarities and their differences? So at the back of my mind is the wish to explore the Coastal limestones between Bredasdorp and the Potberg and find out more. But it is not as easy as that. I also want to know more about H. mirabilis, H. maraisii, H. heidelbergensis, H. mutica, H. minima, H. marginata and H. pumila, which all also occur in the Overberg area and each have question marks attached. They each present problems that I would like to resolve for my own satisfaction, without being confused and befuddled by others who are toying with the same body of knowledge at different levels of expertise and ignorance. In this play we are using words, terms, names and conventions. For this play we need to have a commitment to truth and use the same language or else it is all just meaningless.
So we come to what took me to the Overberg in November 2001. Was it was just my curiosity about all those inviting rocky unexplored places? Is it to learn something more myself? Is it to impress anybody? Actually it was many things including a fascination with chameleons – largely my wife Daphne’s. In March 2000, Daphne and I had come across a most remarkable chameleon at Napier. It was on a small shrub among whitish dried grass growing on a calcareous outcrop strewn with white quartzitic pebbles. The tiny reptile was ivory white, with a stunning array of small orange tubercles on sides and back. One has to know the Cape Dwarf chameleon (Fig.1) and its cousin, the Robertson Dwarf, to know how beautiful and amazing these creatures can be. Another thing was H. mutica and my fascination with collections of this species and their histories of discovery and exploration which are stories of their own. Still another was the news that the Botanical Society had funded a researcher to explore the remnants of the vegetation of the wheat areas. What had he found? Apart from Haworthia, I had once been involved in a preliminary attempt to map and identify vegetation remnants for conservation purposes, and had vainly tried to draw attention to the curiosities of the Silcrete outcrops and remnants of vegetation among the wheatfields.
This researcher was an old acquaintance, Nick Helme, who kindly told me what Haworthias he had seen in his exploration. There was not much new, except for H. mirabilis east of Caledon, and this became our first stop. The site is south of the main area for the species which is along the Riviersonderend river north of Caledon. It does, however, occur in recognisable form south-east of Caledon, also around Napier where it transmutes to some degree (inter alia to the var. badia), and then also at Bredasdorp. There it also transmutes to the var. sublineata (sandstone) and var. mirabilis (the old mundula – and there may be some other way out, but for me this represents the low cussedness of botanical nomenclature). What H. mirabilis does to the north and east of Riviersonderend actually fully explains the problem of classification of Haworthia, and who knows if anyone will ever write that explanation – and truthfully.
Nick Helme’s site is a deep valley which drains to the south. Typical of the area, are these small drainage areas which seldom have permanent water. There may be odd pools lasting into mid- or late summer. There are often exposed shale ridges which host a particular array of succulents and other species – and are often home to owls too. The first problem is access and where does one find the landowner after one has succeeded in finding the place? If one does find the dwelling, will it be the right one and will one find the landowner. Will it be a big aggressive StBernard and more aggressive and ugly Rottweiler reflecting the mindset and security fears of the owner and grossly offending my own, like once near Worcester – or will it be a lonely and over friendly sheepdog. November is a very bad time. This is when the wheat harvest is on. At the end of a bad season, when the crop is drying and it has to be cut and reaped the weather is playing its usual strange tricks, farmers cannot be expected to be approachable. But we did find a helpful soul. From the farmhouse, we drove twenty kilometres back to the main road to the roadside stall, where a family member, Herman DeKock, gave us the necessary permission to venture onto the property.
It does seem an easy business, but in fact it becomes very complicated. Nature Conservation regulations are strict, and land ownership, presence, trespass, stock theft, personality and who knows what else, influence the mindset when setting out an expedition of this kind at all. With those problems all out of the way, and only then, is one able to really enjoy being out in the field and communing with nature – which it is all about in the first place. So we found Nick’s site. We climbed through the fence (s) and tried not to disturb the forlorn sheared, miserably self-conscious sheep that clustered around the earthen dam. It was hot and dry and the most promising spots were quite a long way off. One has to plan one’s approach as the southern slopes are often quite densely vegetated and very uncomfortable to traverse. Snakes are also a consideration and especially in early summer. On this trip we never saw one, although Daphne surprised, and was surprised, by a Lynx drinking at a waterhole. So we did find the plants that Nick had said and took a photograph for the record (Fig.2). Just ordinary H. mirabilis var. triebneriana (cussedness – there should be no typical variety, just a broad general name mirabilis).
From here of course we went to the chameleon at Napier. What a laugh! We had difficulty finding it on the same day on the same bush, five minutes after first seeing it. So expecting to find any chameleon at all, and I have never seen a chameleon in the field in all my years of plant exploration, was quite the wildest dream. But with a dossier of 60 individuals recorded on our small-holding at Kuilsriver over a period of about a month, we felt entitled to hope (all contributing to CM). It is also a most wonderful excuse to be out in the country.
We headed for De Hoop. This is also quite a story. Where do we stay? The DeHoop Nature Reserve is a wonderful place with great accommodation, but it is not where we wanted to be. The staff does not know what it is we are looking for, what our CM data base contains and what our needs are. We want to see the area around the north of the wetland. This is half private land, and half state-owned. Besides there is a travellers Lodge on the edge of the reserve which we understood offered easy and official access to the reserve, presumably to near where we wanted to be. Not so. It is one thing dreaming about this exploration while sitting at home with the map, and quite another to be faced with the problems of roads, fences, ownership, dogs, snakes, flies, mosquitoes, permissions, heat, cold, wet, terrain, bush, visibility, time, distance, misinformation, local “knowledge”, incipient old-age, food, water and the spirit of inspiration and energy which drive the legs (CM). So the Lodge was an unfortunate mistake (we did look, and we did also look where we were told an Haworthia did grow – it was Gasteriacarinata), and we did end up at the office at DeHoop Reserve. You see over the years, one comes to realise that helpful and kind as people by nature are, an awful lot of time is spent chasing red-herrings and wild geese (CM). The help and information one gets is very often of a negative hindrance rather than a positive help. So my real gratitude to the manager there, Mr Ben Swanepoel, who without serious qualm, entrusted us with the key to the northern part of De Hoop where we needed to be.
Unfortunately the problems listed above rather limited our activity. We exhausted our energy climbing up and down rocky slopes in an area which should have occupied us for a week. It is an area which has to be explored by someone at sometime. We found naught, which should not in the least be interpreted to say there are no haworthias there. The terrain is formidable and also most promising. But CM called.
We moved to accommodation closer to our budget restraints and nearer to our goals. Driefontein, where the owner graciously made the necessary contacts for us. What a place – the cold vegetated coastal limestone behind, and featureless cultivated endlessness in front. I have an old record of a collection of mine of H. mutica from “Beyersdal”. Subsequent travels indicated that place, name and memory did not match. So we went to Beyersdal. We did find mutica and it was a different place and a new record (Figs.3a, b & c). Who can actually describe the wonder of plants like these? In tiny patches of rock in a sea of wheat and low-grade (by rainfall and vegetation standards) pasture. Also the quiet, the two owls, also Gasteriacarinata again, this time the Haworthias among them – unusual. The extraordinary colour of the plants. This time the bronze colours of badia. Nearby we also visited an old, by thirty years, locality for H. heidelbergensis. Still there and astonishingly in among white quartz in a small outcrop, but in two little groups occupying hardly two square meters in the approximately two hundred available (Fig. 4).
Then we went to Vanderstelkraal. We had met the owner, Mr Adam Albertyn, who very kindly let us wander over his farm. CM. How does one explore a place like that without living there? It is on the edge. The agriculturally useless coastal limestone at the back, and the productive massively utilised Bokkeveld fields at the front. If terrain had allowed, there would not be so much as a hedgerow to have conserved anything. We saw some very interesting sites and exhausted quite a number of things, like energy, time, passion for the quest, tolerance to bad weather, ostriches, tight-strung barbed-wire fences, no roads and also to the heaviness of the overwhelming destructive productive activities of man. We did not find Haworthia. But I omit a lot, including all the good farming. Mr Albertyn owns the eastern side of the De Hoop wetland where he has a 4X4 route – and there is also a sixty-year old legend of H. minima in that particular area which I would like to confirm. Not this trip though. We did try to the west later, where CM eventually prevailed against us – aided and abetted by overgrowth of alien acacias and the negative vibe they seem to generate (by the side, at Vermaaklikheid, Riversdal, H. mirabilis var. paradoxa grows/survives under Acaciacyclops).
Then we decided to climb the mountain behind Kathoek. Why? I cannot remember why I climbed there in the first place, or how I even did it. Then I found plants in a burnt out area, under the remnant blackened skeletons of Protea – natural fire cycle encouraged by farmers. It does produce a grazeable growth and also invasive aliens. The plants of H. variegata var. modesta were just discernible sticking out from loose sand. Kobus Venter later also visited Kathoek and found plants which were more robust. The plants have spreading variegated leaves (in cultivation?) and while I had no real problem associating them with Karsriver, viz. H. rossouwii var. petrophila, they seemed to have a greater affinity with variegata at the Potberg and Luiperdskop. But my collection and Kobus’ seemed at variance. So Kathoek was a must. From the road CM prevails and it looks quite an expensive option in terms of CM resources. Actually the climb was quite easy. We found the plants before we expected to. They did not look like what we expected them though. They were densely clustering (expected) and in rock crevices (Figs.5a, & b) and pans (Figs.6a & b). The rosettes were tiny and reminiscent of even H. reticulata (see Fig.7). At one place a dense cluster under and in a restioid grasslike plantlet, looked just like a yellowish-brown windblown fallen mass of dry leaf bracts (Fig.8). What impression did it make? Nothing new. It provoked the existing doubt. The population is very similar to petrophila and less so to variegata and hence this variety, with the exclusion of the Potberg population COULD BE transferred to H. rossouwii. But this does not help much, because the Potberg var. modesta is less like the northern Potberg’s two populations and the Luiperdskop population of H. variegata var. hemicrypta, and not more like petrophila! So the CM has to carry this anomaly together with the position of calcarea until whatever else to be found is found. These finds are not going to make things easier to resolve in the CM, nor to explain in a way which assuages the CM of anybody else.
All the while we were pondering other clues to the nature and relationship of things. Like the endemic Braunsiivanrensburgii; a curious specimen of Carissabispinosa so laden with large red fruits it resembled a Pyracantha; Euphorbiasilenifolia everywhere, and what a strange distribution this summer-deciduous geophyte has; and what about Asparagus; what is the relationship of A. capensis, A. mariae, A. suaveolens and A. stipulaceus? Where was Aloe brevifolia or A. saponaria? What was that pugniform Euphorbia at Driefontein or the bayeri-like one at Vanderstelkraal?
Unexpressed CM – we are using my son’s LDV as in my own car anything below well-maintained road is not negotiable. CM – the cellphone we arranged for failed. There is a call in my sub-conscious ‘vehicle needed back home’. We end the trip a day earlier and the message turns out to be true as there is a breakdown of one of the other delivery trucks.
On the way home we pick up some loose ends. We call at a spot north of Bredasdorp where in my lucky ignorance I once found mutica. In several subsequent visits I could not locate it, until one year Daphne and I together with Lawrence Loucka had a very thorough search of the small spot. We found about five plants in a spot about 1/2sq meter. At a later visit we found that the plants had seeded and there were many seedlings but crowded together with seedlings of Aloeferox. Generally Aloe and Haworthia do not mix. Aloe need more root room than Haworthia and Haworthia cannot tolerate competition from larger plants. So we thinned out both aloe and haworthia seedlings. This visit revealed the successful fruits of our activity (Fig.9). The two bigger adult plants were almost being pushed out of the ground by a burgeoning aloe seedling, but there were about 15 other vigorous and healthy plants to carry the group forward. CM – this was an interesting repeat observation of the small distances that seed and seedlings actually do commonly disperse. I have seen the same thing with hybrids where the hybrid is often within the multiple body of one of the parent species.
Mission ending, we made our last visit to Drew. This story is written more fully elsewhere. We had found the pollen source for a plant of H. mutica which I collected there more than thirty years ago and which Kobus and I had taken to the field again to see if seed would set. This surviving single plant is now a collector’s dream which I dub “Silver Widow”. No one had succeeded in finding plants around where a huge centre-point pivot installation now graces the landscape. We were sure the plants were gone, and taking the plant back there was a kind of forlorn gesture of regret that more had not been done to conserve the population – and that so much had been done to eradicate the plants (two hundred plants had been collected and sold to Triebner circa 1935). If anything can be done or could have been done at all to have saved its family? But it worked and we got seed. Where did the pollen come from The pollinator is mainly a solitary bee with a fairly limited flying range. The pollen parent had to be somewhere near. Daphne and I had made several excursions to add to the many different attempts that are known to have been made to find them. Finally I decided that we just had to look again and the plants had to be where we had failed to find them before. So what happened? We left the farm house and walked straight to the spot (getting ‘stung’ by the electric stock fence on the way) less than 300m away and probably about 150m from where we had left Silver Widow to be pollinated. There were about 35 plants (Fig.10) in an area of about 3sq m in flower under Eriocephalusafricanus. So now we returned to successfully collect seed.
The curious thing in CM and all this discussion about big things like Continental Drift, and Biomes, and Vegetation, and Habitats, and Ecotypes, is that H. mutica is not particularly concerned. Here at Drew it grows in an ancient river-boulder situation in what we can call renosterveld under Eriocephalus. At Beyersdal it is on the edge of a stream-bed on exposed Bokkeveld shale with Gasteriacarinata. At Hasiesdrift it is on a gravelly bank (Bokkeveld shale) with dense Aloeferox. Near Napky it is on silcrete with grass. West of Swellendam, it is on whitish-clay, probably associated with silcrete and growing within a restioid species and with Gibbaeum esterhuyseniae. I have also seen it on a fairly bare hillside with a scattering of quartzitic pebbles where the parent shale is too dense and shallow for cultivation or to support denser vegetation. The places hardly have anything in common other than their more general geographic containment, climate, and low biomass potential. How, ever, did they get to their widely-spaced, isolated island sites?
So the tale ends with the reflection on the criticism that I have not treated H. scabra and H. starkiana in the same way I have H. retusa and H. mutica. I have been “inconsistent”. Readers can research and educate their own individual CMs (corporate minds) to the point that they can form opinions which are consistent to the point and purpose of any communication. Aren’t we lucky that the Robertson Dwarf chameleon is a small animal (Fig.11 – by Scott Russel).
Acknowledgement: This fun work is really just about people. Daphne, who kept me company. Janet Albertyn at Nachtwacht, Herman Dekock of Jongensklip, Thys DeVilliers at Boskloof, Tom Ambrose at the cement-block operation at Napier, Rory Allardice at Buchu Rest-camp, Ben Swanepoel at De Hoop Cape Nature Conservation, Annette VanEeden of Driefontein, Mr. Dawid Beyer of Beyersdal, Adam Albertyn of Vanderstelkraal, Theo Blom of Soutspanvlakte, Jan Joubert of the adjacent farm, and Beth and John Humby of Sanddrift. Their good-will and kindness is unforgettable.
Fig.1 Cape dwarf Chameleon – Bradypodion pumilum. Markings, colour, tubercles and spines are individual characteristics. It is said that even the genus is in doubt!Fig.2 Haworthia mirabilis var. triebneriana, MBB7059 Jongensklip, Caledon. Note the reddish spines.Fig.3a Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. This plant is exposed to full afternoon sun for at least a few hours each day. Note the mucronate (pointed leaves).Fig.3b Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A more general specimen with rounded leaf-tips.Fig.3c Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A particularly compact specimen. In this population, the waxy sheen of the species seemed less conspicuous.Fig.4 Haworthia heidelbergensis var. minor, MBB7063 Beyersdal, Bredasdorp. Varietal names are just a geographical convention, as indeed even the species which I recognise, seem to be.Fig.5a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster under a small rock overhang.Fig.5b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster in an exposed crevice and visible as a dark mossy fill.Fig.6a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill.Fig.6b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill – a closer view.Fig.7 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Very reminiscent of H. reticulata say south-west of Robertson.Fig.8 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Barely recognisable to the undirected eye.Fig.9 Haworthia mutica var. mutica, MBB6982 Hasiesdrift, Bredasdorp. Nine seedlings in a tiny island of plants, in a bigger island of Aloe ferox and surviving vegetation on the edge of a wheatfield.Fig.10 Haworthia mutica var. mutica, MBB7064 Sanddrift, Drew. This lovely plant is nearly 120mm diam. and the largest of about 50 plants. They survive, and have survived in this small heavily impacted area for so long. There is no where for them to go when options must have been very limited to start with.Fig.11 Robertson dwarf – Bradypodium gutturale, photo by Scott Russel. Zoologists cannot determine if the Robertson dwarf is different from the Karoo dwarf, or from the two subspecies of the Namaqua dwarf. They use the same fuzzy logic that botanists do, plagued by low sampling and difficult application of statistical principles and probabilities.
If the name “H.enigma” applies to the plant (or plants) from east of Riversdale at Komserante, it is a name that I really do not advise to be taken seriously from a botanical point of view. It is useful at population level and to demonstrate the nature of classification difficulties but it is a minor problem in so far as those difficulties extend. The plants were first shown to me by J. Dekenah on the same day that he also showed me ”H. magnifica” in the Nature Reserve just south of Riversdale that is less than 3km away. My impression then was that it was the same element even if it did look a bit different. The plants are quite large (to 70mm diameter), fairly tubercled and often with lines in the upper retused area of the leaf face. While I originally classified “H. maraisii” under “H. magnifica”, I later separated them because it seemed so incongruous to include all the variants of the western “H. maraisii” with the few populations of “H. magnifica” then known. Also, as Essie Esterhuizen pointed out, “H atrofusca” as a variant of “H. magnifica”, seemed to be more dominant than had been realized. There were several other complications largely due to ignorance. Since my revision I have done so much more exploration and turned up so much new material that I have been forced to the conclusion that there is really one main element involved and that is H. mirabilis. This is where I believe the Komserante plants belong and the difference from the Nature Reserve population is due to a degree of infusion of H. retusa.
I revisited the site with Kobus Venter many years ago but did not look at a reported second population higher up the hill, taking it to be a little different based on plants I saw in Kobus’ collection. What was on my mind while we were recently exploring the area further east to examine the possible connection of H. mirabilis “magnifica” to “splendens” (and which we confirmed), was the fact I had never seen Kobus’ plants from Kruis Rivier northeast of Riversdale other than in Kobus’ collection. The plants I saw were also generally more robust than “H. magnifica” and more evenly tubercled. Kobus kindly took me to that Kruis River locality and much to my surprise the plants were in flower late October (see JDV92/65 Figs1). This is quite wrong for H. mirabilis, which is essentially a summer flowering species. I later went again to explore Komserante more thoroughly and to look at both the “magnifica” populations to which I believe the name “H. enigma” has been applied. The populations are in fact no more than 75m apart and cannot be considered to be genetically discrete at all (see MBB7778 Figs 2, and MBB7779 Figs 3). While it is true that the habitats are slightly different, this is reflected in the plants that at the upper slope of the hillside are vegetatively more robust and even clump forming, while those lower down in a bushier grassier habitat tend to be solitary and more withdrawn into the soil. These plants flower in summer and it is evident to me that there must have been some genetic exchange with H. retusa that grows approximately 200m away on the same hillside.
1. JDV92/65 H. mirabilis Kruisrivier.
2. MBB7778 H. mirabilis. Komserante.
3. MBB7779 H. mirabilis. Komserante.
Habitat preferences are very strict and it is curious to observe that there are four small buttresses forming the western slope of the low hillside. On the northern one we have the two H. mirabilis populations. On the second is H. retusa ’geraldii’ (see MBB7780 Figs 4) that is very clump forming, the third has no haworthias and on the fourth is a less clump forming H. retusa that is often referred to as ‘fouchei’ (see MBB7781 Figs 5) because the leaves are slightly longer and more upright than in ’geraldii’. This is not quite correct. Mr. Dekenah showed me what he took to be the true H. retusa ‘fouchei’, further south off the hillside and along the stream at Komserante. This habitat has since been cleared for farmland. The plants were very large and the leaves considerably more erect than is the case for the plants on the hillside. Curiously H. minima is present on the first and last buttresses. Flowering time for these H. retusa is spring but we did see vicarious flowering in summer so that definitely it is possible and probably a regular occurrence for out-of-season flowering to occur with the possibility of hybridization. I noted such hybrids between H. retusa and H. mirabilis in our exploration further east from Riversdale and I did think some of the clones in the Komserante plants evidenced this as well.
4. MBB7780 H. retusa. Komserante.
5. MBB7781 H. retusa cf. ‘foucherii’. Komserante.
The reality is that the Kruisriver and Komserante “magnifica” may flower at different times, but they are vegetatively very similar indeed. I consider that they belong in the same system as H. mirabilis and this conservative view of “species” as systems is the most sensible way to interpret and understand the plants. The approach can be taken further, as I tried to do in an earlier article concerning H. mutica ‘nigra’. In that article I dealt with a whole series of populations in and around Heidelberg, to show that there are three main elements viz. H. retusa (including H. turgida), H. mirabilis (including H. magnifica) and H. floribunda. I did not deal specifically with, nor resolve, the issue of H. mutica var nigra. I also noted populations along the lower Duiwenhoks river and suggested that H. floribunda is absorbed into H. mirabilis virtually south of the N2 east west highway, re-emerging on the northwestern corner of the Potberg in the south.
My most recent exploration was to check the flowering times of the original population of H. mutica var nigra at its origin at Kransriviermond and what I assumed to be the same thing on rather limited examination further north at Morning Star. Both these populations are along the Duiwenhoks river southeast of Heidelberg and in a continuum of four populations of H. retusa extending from just east of Heidelberg to still further east at an eastern Diepkloof locality (I think it has been referred to as Droekloof and Droerivier too, and the name ‘chromatica’ attached to the plants – see MBB7794 Figs 6). Less than 10km away is a population of H. retusa at Pienaarsriver (see MBB7776 figs in Chapter 3) that is very near to the whole array of populations of Kiewietskraal that I discussed in my writings about H. mutica var nigra. Two of those were distinctly H. retusa and so is the Pienaarsriver population that is different in that the plants are mostly quite dark in colour.
6. MBB7794 h. retusa ‘chromatica’. SE Heidelberg.
In those same writings I mentioned several populations of H. mirabilis along the Duiwenhoks river as well as populations of H. retusa var turgida. Here I am unashamedly relating “turgida” to “retusa” as I believe that when one calls all the populations now known to mind, as well as the variants they contain, this is the correct position of the plants in relation to a sensible and rational species concept. It is evident that H. floribunda is merged in H. mirabilis south of Heidelberg and the same thing occurs south of Swellendam. Thus I see no problem in now dealing with the population at Kransriviermond to which I applied the name H. mutica var nigra. I attached the varietal name to H. mutica because this is what G.G.Smith had done, and I believed that it was the link between H. retusa in the east and H. mutica in the west, when there were still huge gaps in the known distribution records for both species.
I have mentioned a population of ostensibly H. mutica var nigra at Morning Star, which is about 5km north of the Kransriviermond origin of the plants so named. This Morning Star population (see MBB7221 Figs 7) is very significant because while it includes virtually the same range of variants as the Kransriviermond plants, it flowers in summer as opposed to spring for Kransriviermond. Not only that, but it is 300meters away from a population of H. mirabilis (previously this would have been H. magnifica) that flowers and seeds at the same time. The habitat is marine plane ferricrete that overlies Bokkeveld shale but there are subtle differences between the two habitats.
7. MBB7221 H. retusa ‘nigra’. Morning Star.
The Kransriviermond plants (see MBB7804 Figs 8) are down in the Duiwenhoks valley in exposed Bokkeveld shale and I consider that the dark colour of the plants in the two populations is due to infusion of H. mirabilis. The rounded leaf tip would be derived from the “atrofuscoid” element of H. mirabilis while some plants also have the tuberculate roughness that that element often has.
8. MBB7804 H. retusa ‘nigra’. Kransriviermond.
Further south and west is the Slangrivier valley where there is also a suite of populations that includes smaller forms of H. mirabilis (H. heidelbergensis var. toonensis) as well a H. retusa var turgida also in an unusual form that supports the taxonomic position I now adopt.
The conclusion is that the Morning Star and Kransriviermond ‘nigra’ can be regarded as H. retusa within the same broader concept that includes ‘turgida’. The problem now arises from still another new discovery. This is from along the Buffeljags River (see MBB7801 Figs 9) about 10km southeast of Swellendam and 12km from the nearest obvious H. retusa at Goedverwagting further east; and from the less obvious H. retusa (that I have elsewhere noted as H. mirabilis) south of the Tradouw Pass. The nearest H. mutica are within about 15km to the west at Napky and Luiperdskop. The plants are the now customary “variable”. The leaf tips are very rounded. Some plain, some lined, some with curious submersed pinkish dots. The surfaces can be quite smoothish or else the surface cells terminate in minute spines that give the surfaces a glistening appearance – evident in H. mutica too. These plants flower in summer whereas both H. mutica and H. retusa are spring flowering. Note needs to be taken of the habitat. It is an extensive and old river gravel embankment and terrace, and therefore another variant on the interface of recent ferricrete deposits and the older Bokkeveld shales. The vegetation is grassy Karooid bushveld, unlike the dry fynbos to the northwest or the renosterveld (mainly cultivated) to the east.
Kobus Venter saw the photographs and suggested that the plants might be H. mirabilis’ and reminded him of ‘atrofusca’, perhaps also by virtue of the flowering time. This is of course what my “enigma” is all about. It is compounded in this case because these Buffeljags ‘muticas’ are rather odd in that the leaf ends tend to be flattened almost as though H. floribunda has also entered the fray. Kobus was much happier with the H. mutica connection when he saw the plants in the field.
The story here closes with H. mutica MBB7741 from Dankbaar about 12km west of the Breede River bridge at Swellendam. The plants here also have very glistening surfaces and thus seem to confirm the very original perception of G.G. Smith’s that the Duiwenhoks plants and the plants west of the Breede River were connected. It is certain that he never envisaged the complexity of the interrelationships between three main elements viz. H. mirabilis, H. floribunda and H. retusa.
Footnote. It is just assumed that perhaps the name “H. enigma” has been used for the Komserante H. mirabilis, because that population is indeed enigmatic as this article explains. Gerhard Marx informs that “enigma” applies to a H. mirabilis population further west where the more atrofusca variants occur. Since writing this chapter yet another similar population to the Kruis River and Komserante populations has been found about 15km SE Riversdale. ♦
These field trips, to the Potberg this time, are always made with some objective in mind in respect of new exploration. In this case I wanted to get more pictures of H. mutica as it is a species that I have few digital images of. There were also localities that I remembered from the days when I was sweeping the countryside at a fairly coarse scale and was not much bothered by detail. I confidently expected the number of real species conforming to that in other fields of botany and zoology, to be in the region of 33. I never dreamed that such divergent views would, or even could, arise from less information than even then available to me. So while 450 names were whittled down to the mid-hundreds by me, students of the genus have in recent years pushed that up to the 600 mark. My opinions have been couched in quite conservative terms but it is a problem of the nomenclatural system that an identification in respect of a Latin name evokes a reality that does not exist. I maintain that the problems we experience in Haworthia are no different to that which exists in many animal and plant genera. I think that primarily this is because of the absence of insight into, and understanding of, the actual nature of species and the two dimensional model we use to relate them. Species are very variable systems because they have to be to survive the constantly changing world they occupy. In this article I am just going to present images of plants within populations of four different species viz. H. variegata, H. minima, H. mirabilis and H. mutica.
Set 1 H. variegata H. variegata has an odd distribution in that it is primarily in the lower Goukou River drainage system southeast of Riversdale. Plants of H. monticola from the Uniondale and Willowmore areas, and even those of H. angustifolia from the Eastern Cape, can be very similar. So the distribution south and west of the lower Breede River is quite remarkable. Two populations, one high on the western end of the Potberg Mountain and another northwest of this at Luiperd’s Kop are very similar to those in the Goukou area. Chris Burgers of Cape Nature brought me plants from the lower north slopes of the Potberg that were quite different. Small single plants with outcurving leaf ends barely protruding above the pebbles the plants were growing in. I named this H. variegata ‘hemicrypta’ but not without reservation in the same way as I also named the plants in another population on the Potberg and at Kathoek as H. variegata ‘modesta’.
Chris Burgers did take me to his site many years ago and it is one of those places I cannot recall in the pebbly flats and Fynbos vegetation of that area. So the finds here seem to represent that same element but with subtle differences in respect mainly of variegation and spination of the leaf margins. I have mentioned elsewhere that there is an interaction with an element I refer to as H. floribunda although it is not clear if this indeed that species, or if it is an emergent polytopic element of H. mirabilis in consideration of the variation and distribution of these two apparently different species. We may have a truth for one area that is different from a truth in another. Polytopic means similar looking things arising in different places from a common genetic pool.
1. Map of the Diepkloof area (fig.1)
Map of the Diepkloof area.
2. MBB7923 H. variegata S Diepkloof. (figs 2)
Fig. 2.1 7923 S Diepkloof
3. MBB7924 H. variegata SE Diepkloof (figs 3)
Looking west to the Kleinberg
4. MBB7925 H. variegata SE Diepkloof (figs 4)
Fig. 4.1 7925 SE Diepkloof
5. MBB7927 H. variegata SSE Diepkloof (figs 5)
Fig. 5.1 7927 SSE Diepkloof
6. MBB7928 H. variegata SSE Diepkloof (figs 6)
View looking east to 7927.View looking southwest with Kleinberg off to the right.
7. MBB7515 H. variegata Stoffelsriver (figs 7)
Fig. 7.1 7515 Stoffelsrivier.
Applying the available varietal name, I should refer to these populations as H. variegata var. hemicrypta. However, I think this implies fixity of opinion and a certainty or reality that are neither not strictly true. I have added the Stoffelsriver two pictures just as a reference because while the first 5 populations are associated with recent Tertiary deposits and Renosterveld vegetation, the last is in sandstone gravels and in Fynbos.
Set 2 H. minima This species of course belongs with three others in the Robustipedunculares and in my opinion more closely related to Astroloba than to Haworthia. It is widespread and even occurs in the Little Karoo. None of these populations are significantly different from the general run for the species, but we were awed by the beauty of the plants in their natural settings where exposure results in compactness and beautiful colouration. They can occur in quite different environments and habitats. There are a few aberrant populations and hybrids with other members of the sub-genus.
8. MBB7496a H. minima Diepkloof S (figs 8)
Fig. 8.1 7496a Diepkloof S
9. MBB7926 H. minima SE Diepkloof (fig. 9)
Fig. 9 7926 Diepkloof S
10. MBB7929 H. minima SE Diepkloof S (figs 10)
Fig. 10.1 7929 SE Diepkloof
11. MBB7930 H. minima S Diepkloof (figs 11)
Fig. 11.1 7930 with 7947 SE Diepkloof minima
12. MBB7932 H. minima S Kleinberg (figs 12)
Fig. 12.1 7932 S Kleinberg
Set 3 H. mirabilis H. mirablis is an extremely complex group of many populations and plants and distributed from Albertinia in the east to near Caledon in the west, and from the foothills of the Langeberg to the coast. The relationship with H. retusa hangs on the slender thread of flowering time, with H retusa flowering in late spring and H. mirabilis in late summer. The relationship of these two species to H. pygmaea in the east and H. mutica to the west has been mooted by me elsewhere. A similar scenario can be suggested for the species recognized north of the Langeberg.Included in the following sets is the population at Die Kop that I surmise has its origins in interaction between H. mutica and H. mirabilis.
There are three populations in these sets (nos 13-15) that are suggestive of H. mirabilis ‘atrofusca’, while others have distinct resemblances to H. floribunda (e.g. 16). It is curious that nearly all the populations of H. mirabilis that occur in the same general geographic area as does H. mutica are of smallish plants. These I originally regarded as a separate species viz. H. heidelbergensis. This should be seen as one of those decisions based on inadequate sampling and knowledge of the field. This phenomenon now suggests to me that H. mutica is occupying middle ground between H. retusa in the ‘turgida’ form and H. mirabilis.
13. MBB7496 H. mirabilis Diepkloof S (figs 13)
13.1 7496 Diepkloof SView overlooking habitat 7496.View looking west to north of Kleinberg.
14. MBB7497 H. mirabilis S S Diepkloof (figs 14)
Fig. 14.1 7497 S S DiepkloofView looking north.
15. MBB7933 H. mirabilis S Diepkloof (figs 15)
Fig. 15.1 7933 S 7496 Diepkloof
16. MBB7922 H. mirabilis Diepkloof (figs 16) A locality established by Jakub Jelimicky. The similarities to H. floribunda is only apparent in some of the plants where the leaf tips are twisted, slightly flattened and rounded at the ends.
Fig. 16.1 7922 Diepkloof
17. MBB7931 H. mirabilis S S Kleinberg (figs 17)
Fig. 17.1 7931 SS Kleinberg.
18. MBB7531 H. mirabilis S Kleinberg (figs 18)
S Kleinberg slope. Enthusiasts at play.Figs 18.1 7531 S Kleinberg
19. MBB7935 H. mirabilis Langvlei (figs 19)
7935.1 LangvleiLangvlei, the plants are in a tiny bit of rock just beyond the dumps.
20. MBB7938 H. mirabilis SE Stoffelsriver (figs 20) This locality was recorded when Gerhard Marx and Jakub Jelimicky misunderstood my directions. The plants are incredibly cryptic and very hard to see in a not very well defined habitat. One can only follow my opinion on the classification in the context of 8 other populations in the sandstones and Fynbos, and then relate these to populations in shales, tertiary deposit and transitional vegetation.
Fig. 20.1 7938 SE StoffelsrivierSE StoffelsrivierSE Stoffelsrivier View looking east to Sandhoogte
21. JDV86/2 H. mirabilis Haarwegskloof (figs 21) These are small plants in very close association with H. mutica as discussed above and are in quartz outcrops rather than in the shale ridges where H. mutica is found.
Fig. 21.1 JDV862 Haarwegskloof
Set 4 H. mutica H. mutica is not a species that is well-known and this is probably because it is not proliferous. The name mutica means without a point and “typically” plants of H. mutica should and do have no point (end-awn). The main object of the trip was to obtain more images of this species to convey a better impression of the many variants and how it can be viewed in relation to other species.
22. MBB7075 H.mutica Grootvlakte (figs 22) I am not certain of the origins of H. otzenii G G Smith and the locality that I traced was approximately 8km east of Riviersonderend. That site is now very degraded and I could not find plants on a visit many years ago but where I did see them on my first visit there. This Grootvlakte population occupies a similar habitat in an exposed shale ridge south of the Riviersonderend River.
Fig. 22.1 7075 Grootvlakte7075 View of site at Grootvlakte
23. MBB7935 H. mutica Crodini (figs 23) The name Crodini may have been used for the following collection at nearby Witklipkop, and I say this because I did once get seed from there and Jakub Jelimiky informs he has a plant labeled ‘Crodini’. I only saw two plants here at Crodini itself when the site was severely grazed, and we found only 8 plants on this occasion. The plants appear to be in quartz, but this is a bit of an illusion as there are white quartz bands in the shale and the plants prefer the shale.
Fig. 23.1 7935 CrodiniView across site. Grootvlakte
24. MBB6536 H. mutica Witklipkop (figs 24) This is not a robust population in shale and also marginal to quartz.
Fig. 24.1 6536 Witklipkop, Napky
25. MBB7937 H. mutica Platkop, S Napky (figs 25) The substrate here seems to be silcrete but there is no underlying kaolinite and the area is cultivated, which suggests shale. So that is a bit of an unknown.
Fig. 25.1 7937 N Platkop SE LangvleiN Platkop SE LangvleiN Platkop SE Langvlei
26. ADH2729 H. mutica Ouplaas, DeHoop (figs 26)
Fig. 26.1 ADH2729 Ouplaas, De Hoop
27. MBB7934 H. mutica Wolwefontein (figs 27)
In early 1970 I took Peter Brandham and David Cutler of Kew to look at Haworthia in the field. In fig.27.12 the view is southward to Kathoek and the cottage in the foreground was occupied. It must have been abandoned soon after and the access road obscured. The result was that the place was later difficult to find, but the plants are still there. It is not far from the problematic Die Kop population to come.
Fig. 27.1 7934 WolwefonteinWolwefontein
28. JDV85/17 H. mutica Haarwegskloof (figs 28) This record goes back to 1985. We did not have landowner permission here as we did not know exactly where to go. The area was also unfenced. So we were confronted by the farm manager and never took as many pictures nor explored more widely. It certainly would have been problematic had we not simply being photographing. Anybody else may have ended up at the police station. It is a case of farm land too hilly and rocky to cultivate and bought by apparently wealthy city people with an eye to conservation. This may result in the place being game-fenced, a lodge built for the up-market; and made perhaps even less accessible.
Fig. 28.1 JDV85-17 Haarwegskloof
29. MBB6641 H. mutica Hasiesdrift (figs 29 JDV97/26) This was one of my early encounters in 1969 with Haworthia, where I fortuitously stopped at a river crossing, lifted a branch of a shrublet and saw a magnificent plant just like that illustrated and serving as the type for the name H. mutica (see fig.29.1for an example)). A year later a new road was built and if there had been a population there it was no more. In 1996 when I was writing Haworthia Revisited, I went back there and after a diligent search found a very small group of plants at the base of a small plant of Aloe ferox. The two species seldom grow among each other so this really was odd. I have periodically been back to see what was happening because there were many Aloe seedlings. The very small population has waxed and waned and the mothering Aloe seedling has died. There has been a small spread of plants to occupy an area of about 2 square meters. I know here are more plants about 2km away but have not explored the wider area more thoroughly.
Fig. 29.1 JDV97-26 Hasiesdrift.
30 MBB7500 H.muticaXmirabilis Die Kop (figs 30) So we come to the problem child that has acquired the species Latin binomial H. hammeri. I think this is unfortunate as Steven Hammer deserves a great deal better than this. It should not be necessary to have to also manage personal issues in an inquiry into what the name means or if it is a valid statement in science. This population is one I found when I was exploring widely for an understanding of H. mirabilis. It was not growing in conventional habitat and populations occur at many places in the wider vicinity. So I do not comprehend at all why this single population with absolutely nothing to support its designation as a species in respect of anything but the fact that the plants look different from something else. The plants are also very variable as the set of pictures should show, and the plant suggested by the picture accompanying the description is not typical. The proximity of H. mutica and the nuances of these plants with this species and also with that of other nearby populations of H. mirabilis, suggest to me that this population is of hybrid origin. I also use the term hybrid in the sense of genetic similarity because hybridity itself implies separate existence. My feeling is that H. mirabilis and H. retusa have separated in time from a common gene pool and that the process is not complete and nor will it be. My species definition requires that we see all these populations as one system that is spread in geographic space and is changing with time. The rate of change is a fourth dimension.
I suppose one should comment on the shiny leaf surface (fig. 30.3 especially) that is present in H. mutica, as this character is also described as a diagnostic for H. groenewaldii that is as contentious as H. hammeri.
Fig. 30.1 7500 Die Kop
Conclusion The objectives of the excursion were fully met and I am sure these pictures will contribute to a better view of how these four species vary. Perhaps this will illustrate also the illogic of Latin binomials for every variable, however desirable these names are for traders and collectors.
Acknowledgement Bennie Viljoen of Grootvlakte, Koos Badenhorst of Langvlei, Maree and Elizabeth Prinsloo of Stoffelsriver, Ferdie of Diepkloof Security Services, Adele and Vlooi DuToit for Die Kop, Martin Mijnhardt of Haarwegskloof, Neil Giliomee of Hasiesdrift. In some cases we simply could not track landowners and trust they will forgive us our trespasses remembering that invasion of ones territory is close to an unforgivable sin. ♦
A supposed new species of Haworthia viz. H. groenewaldii Breuer, is described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald. It presents the description of this supposed new species from Buffeljags east of Swellendam that I would simply have identified as another variant of H. mutica. This is not because I am confounded by the variability among the plants in the genus, or even in any one species. I recognize the species as systems of populations in which the individuals vary from one another as one would expect in any group of living things that maintains the flexibility to adapt to constantly changing world conditions. In this even time becomes a variable. I think species are very important elements if the whole of creation and not just for taxonomist and collector activity. Other people have other ideas of what species are, so my disagreement is hopefully forgivable..
Although H. mutica was described by Haworth in 1821 it was not allied to a South African field population until recognized by Col. Scott in 1985. G.G. Smith had failed to recognize it when he described his H. otzenii in 1945. The type by which the name is supported is a Kew illustration reproduced here as Fig. 1. This then is what one would expect a typical representative of the species to look alike. Now the ever present problem in Haworthia, is that no two plants in a population may look quite the same. Hence my problem with the description in which the word “typical” is rather bandied about. It falls into the first aspect of taxonomy.
Fig. 1 H. mutica. Kew herbarium. The type.
Firstly a plant is illustrated on the front cover of the respective Alsterworthia that, presumably the authors, state is a typical specimen of H. groenewaldii. Secondly, Marx is quoted as saying that the “typical H. mutica” grows only 20km west at the farm Dankbaar. Statements like these are used to strengthen and support opinions and generate a reality that Latin binomials sadly lack. It so happens that I know both these populations quite well and these statements are news to me. I do not think the specimen on the front cover is by any means typical of the population at Buffeljags, and certainly not at three sites recently discovered nearby. The plants at Dankbaar also do not in my opinion fully meet the imputed similarity to fig.1. See fig 2. for an image of a plant representative of the Dankbaar population. I would be very hesitant to say that this is typical of Dankbaar plants.
Fig. 2 MBB7741 H. mutica. Dankbaar. A representative.
There is a curious problem here in that Scott does not use the type illustration in his Revision and does not state any origin of the plant he uses to illustrate the species i.e. H. mutica. As far as I am aware the type of a synonym that G.G. Smith described viz. H. otzenii, came from east of Riviersonderend, but this is for an Otzen collection no. 6. The type was cited by Scott as Otzen 10 but this is not in the Compton herbarium where it should be. So I am not sure offhand where that came from. However, this is not really relevant to the discussion. I just want to state that finding a plant that matches the type is no mean feat and that it was by sheer chance that in 1969 I came across a population of plants at Hasiesdrift that did. See fig. 4 and 5. I selected one image and then realized that it did not have round enough leaf tips to meet need, so I selected another. In the first picture the leaves tend to have a “mucro” – a small point to the leaf that looks different to a well developed end-awn (bristle) that the leaves can also have.
Fig. 3 JDV97-26. H. mutica. Hasiesdrift. Representative.Fig. 4 JDV97-26 H. mutica. Hasiesdrift. More representative.Fig. 215 7801 H. mutica ‘groenewaldii’.
Another issue is that Gerhard Marx once argued with me that the mooted H. groenewaldii was much nearer to H. mirabilis than to H. mutica as I had suggested. What he luckily is able to ignore is my observation of the similarity of some plants of H. mutica to H. mirabilis see figs 5 and 6. I never saw a plant quite like fig. 5 in all my exploration at Buffeljags but Jannie Groenewald collected this one there. Fig. 6 is not typical of the population either and I used this same figure somewhere else in my writing to comment on the similarity to H. mirabilis ‘badia’ variants at Sandfontein (east of the “typical”). Gerhard is still more fortunate to be able to ignore the similarity of H. mutica to H. retusa see figs 7 and 8. I even surprised myself when in looking for a suitable picture, from many, I picked this fig. 7 and find it is also Hasiesdrift albeit a cultivated version. (There is such an interesting story around the Hasiesdrift site). Fig. 8 is a representive of H. retusa from Pienaarsriver pictures and I feel sure that readers will agree that the names could be switched. It was very difficult to ignore pictures from Pienaarsriver that I could have used with figs 5 and 6 in the context of H. mirabilis.
Fig. 6 JDV96-16 H. mutica. S Napky.Fig. 7 JDV97-26 H. mutica. Hasiesdrift, N Bredasdorp.Fig. 8 MBB7776 H. retusa. Pienaarsrivier.
It becomes still more interesting (I would have said complicated but my critics maintain that this variation confuses me and it is actually possible to get it all tidy and neat) when one further compares a plant of H. retusa ‘nigra’ ( fig. 9) with both H. mutica (fig. 7); and then H. retusa ‘nigra’ (fig. 10) with H. retusa (fig. 11). The latter is in fact from the population where the variant ‘geraldii’ originates.
Fig. 9 MBB7920 H. retusa ‘nigra’. Van Reenens Crest.Fig. 10 MBB7921. H. retusa ‘nigra’. Van Reenens Crest.Fig. 11 MBB7781 H. retusa. Komserante.
A last point I can make is that the leaf flecks said to characterize “H. groenewaldii” do occur in H. mutica at Klipport (see figs 12 & 13). One cannot simply ignore the extraordinary chain of similarities that extends across the entire distribution area producing only a slightly different situation at either extreme.
Fig. 12. MBB6512 H. mutica. Klipport.Fig. 13. MBB6512 H. mutica. Klipport.
Floral difference is a great issue that is misused to force an opinion. The flower is extremely difficult to study because the differences across the entire subgenus are so small. There are complications where, as an example, flowers of H. pulchella ‘globifera’ are indistinguishable from those of H. cymbifomis var. incurvula. In the subg. Hexangulares there is an incredible problem where floral differences within species exceeds that between species e.g. a flower of a plant of H. limifolia may more closely resemble that of H. coarctata rather than that of another plant of H. limifolia. It is easy to draw conclusions from small samples of a few flowers from a few populations but one very soon finds that with increasing sample size the most carefully constructed table of differences becomes senseless. Just when is an ever-aging flower on a stalk at the precise same stage of a flower you want to compare it with? How many flowers from how many plants are needed to make a valid statement? Do not forget that the observations must be made on plants on the basis of random selection too. This is a requirement mostly totally ignored when the more serious question of a species difference is being debated. That of course brings us back to this casual use of the word “typical” and its intractability.
The hardest problem to deal with is that of flowering time and on the face of it a winter flowering time versus a summer flowering time can be taken to suggest significant difference. Yet if one considers that a species needs to harbour genetic variation to ensure adaptation to any kind of environmental change and so survival, a different flowering time may be an extremely useful resource. Then we do have the reality of hybrids between species that do flower at these different times. So obviously and self-evidently populations of species may exist that has arisen from such hybridization between plants that flower at different seasons. The capacity of plants and animals to synchronize breeding periodicity is well-known.
I close with my observation that H. mutica is an assorted group of plants that seem to fall in some middle zone between H. mirabilis and H. retusa. Hence in the west we have H. mirabilis/ H. mutica/H. retusa ‘nigra’, while in the east we have H. pygmaea (=H. mirabilis+H.retusa ‘retusa’) and H. retusa ‘turgida’. In the area between there is vast variation of H. mirabilis and H. retusa that get a bit of H. floribunda thrown in too.
Map showing known distribution of H. mutica.
So am I confused, or have I confused you? As Steven kindly put it…’it’s almost as if you were being blamed for nature’s complexities’. Of course the ultimate reality is that we each have our own idea of what “species” are, and here I have used my version! ♦
It so happens that Heidi Hartmann first visited the Karoo Garden more than 35 years ago and it has been very difficult for me to pay attention both to her mesembs and all my other plant interests. In the last few years she has been working on Acrodon. This is a small genus of only 5 to 6 species that occurs in the Southern Cape with much the same distribution and habitat requirements as Haworthia. She had had some second thoughts on a species she had described as Acrodon calcicola and intimated that she needed photographs to show what proves to be detaching fruits (capsules). So off we went to get that northeast of Bredasdorp at Rooivlei. But Nick Helme had about a year before sent me an intriguing picture of a greenish soft looking plant from near the De Hoop Reserve entrance road to the east. I had considered that it might be an equivalent of the H. muticaXmirabilis population at Die Kop (MBB7500) that Ingo Breuer usefully described as H. hammeri. I use the name with great trepidation because to say what is correct usage is difficult. It could pass as a cultivar name, a varietal name or a form name. I am quite sure it has its origins in the interaction of two species and that is what a botanical name should reflect that; thus H. muticaXmirabilis or however else the nomenclaturists may require. So these journeys are never without distractions as Rooivlei itself is a remarkable site. I find that I have few images of the populations of Haworthia that occur there. The product is nearly all pictures/images.
Set 1. MBB7941 H. mutica Ouplaas, Wydgelee (figs 1-45)
This population is just 3km west of the Die Kop MBB7500 population. There are two intervening H. mirabilis populations; one small, one large and scattered, and a third a little to the north of east and more to the north. But that species is well covered in an Update article and my attention here is on H. mutica. Why Nick Helme’s plant was so green and soft-looking is a bit odd because the population of H. mutica there at Ouplaas is quite one of the largest and most dramatic of any that I have seen of the species. Daphne and I spent hours examining, reexamining and exclaiming at the sheer beauty and variation of the plants. There were plants squeezed into cracks and flattened so that one could not see any centre, others in the flat clayish areas between the exposed vertical shale ridges, but mostly in the shale. The site is north-facing and so is very hot and dry in summer. The plants had just finished flowering.
There was one particular image (fig. 24) I have used to show five different leaf-end shapes on the same plant.
Fig. 1 7941 OuplaasFig. 44 7941 View west to HaarwegskloofFig. 45 7941 View east to Die Kop
Set 2. MBB7943 H. mutica E Klipbankskloof (figs 46-54)
I saw plants and photographed one (MBB7246) south of Klipbankskloof in a valley mapped as Verbrandskloof. There is another farm, also named Klipbankskloof further west where Jakub Jelimicky recorded H. mutica too. I wanted to see that locality and registered the two Klipbankskloof places as the same. Not so. So I intended up exploring further east where we first found this in a west facing ridge. There was no sign of fruits or flower.
Fig. 46 7943 E KlipbankskloofFig. 54 7943 View west to farmhouse
Set 3. MBB7945 H. mutica SE Klipbankskloof (figs 55-80)
This was on a north facing slope and at first we were a bit taken aback because the plants we first observed had flowered poorly or some time previously. But that changed as from north facing slope we entered a very minor valley with both northeast and northwest aspects (more to follow). Here there were many plants and all just finishing flowering. There were some outstanding, dramatic plants. I wish I was a more accomplished and better equipped photographer. Some of the images would have been stunning.
Fig. 55 7945 SE KlipbankskloofFig. 80 7945 View lookin across depression to west facing slope
Set 4. MBB6638 H. mirabilis Rooivlei (KoeiseKop) (figs 81- 92)
It was fortunate for my story that I took pictures here. As in most of the greater area occupied by H. mutica,H. mirabilis occurs as these smaller, often (how often is often?) dark rough versions of H. mirabilis. Of particular note is the first image (fig. 81). This is of a plant with much smoother textured leaves and had there been and H. mutica in the near vicinity I would surely (mistakenly?) taken this for a hybrid because I know from experience elsewhere that summer and winter flowers can and do hybridize. But wait! Rooivlei is a unique site because there is a population of H. retusa ‘turgida’ nearby that is prominent in my whole explanation of relationships that my critics so dislike. Outside chance is that hybridisation is an explanation.
Fig. 81 6638 Rooivlei
Set. 5 MBB7944a H. mirabilis SE Klipbankskloof (figs 93-101)
I have numbered this as ‘a’ and ‘b’ for a reason. The two ‘populations’ are less than 200m apart. In fact the two H. mutica populations are not all that much more distant and there would be some debate as to whether they constitute populations at all. My argument would be that they occupy discrete separated habitats. This mirabilis population falls into middle ground. It is the upper slope where aspect changes from south to north. It is moister and the southern aspect of the small shale ridges provides mirabilis habitat. It is as if H. mirabilis is found where what I can describe is an escape route is present. These dark plants do not readily tolerate a full days exposure to direct sun and I speculate that this is the same kind of thing that happens on north-facing mountain slopes as around MacGregor. Plants are usually where the aspect allows escape from a full days worth of sunlight. The plants are much like at Rooivlei.
[ed.] images mislabeled as 7943, should be 7944.
Fig. 93 7943a SE Klipbankskloof
Set. 6 MBB7944b H. mirabilis SE Klipbankskloof (figs 102-110)
The different number is because the plants are now sharing both habitat and niche with H. mutica. I do not know any other site where this happens. The two species do occur in the close general area but that is then different habitat and the distance between ranges from about 50m (Haarwegskloof) to 400m (Rondeheuwel). What is now fascinating is that some of the plants resemble fig 1 of Rooivlei and we are faced with the probability that there is exchange of genetic material between H. mutica and H. mirabilis. This is what is suggested at Rooivlei too for H. retusa ‘turgida’ and H. mirabilis (see above).
Fig. 102 7943b SE Klipbankskloof
Conclusion None. This is purely instructive. What I gained from the trip was the realization that H. mutica would be a very good subject by which to explore variability and change with time. Also to look at conservation and impact of land-use on populations. Over to Jakub Jelimicky.
Acknowledgement Again a list as long as an arm. Francois Uys for access to Rooivlei. Nick Helme for information in respect of Ouplaas. Odette Curtis for interest in Renosterveld conservation and contact for Nick. Johan Dutoit of Vleitjie and a friendly combine mechanic for access to Ouplaas. Wynand Dutoit of Klipbankskloof for access to what is his wife, Madrẽ’s farm. ♦
The objective was to explore some likely habitats previously observed at Van Reenens Crest and nearby. We extended the scope to include further exploration for Haworthia mutica as I am still questioning the place of this species in the greater scheme of things. Thus here are four sets of populations that I report on viz. H. mirabilis, H. retusa ‘nigra’, H. floribunda and H. mutica. See maps Figs 1 and 2 for geographical position.
Van Reenens CrestFig. 1 Van Reenens Crest. M = H. mirabilis, RN = H. retusa ‘nigra’, F. = H . floribunda.Fig. 2 Known distribution of H. mutica
H. mirabilis
Set 1. MBB7955 Van Reenens Crest – mid west. (figs 3-11)
The plants are moderate in size to about 50mm diam. and growing on an erosional slope below tertiary gravels where there is white clay. They vary from densely tubercled to relatively smooth.
Fig. 3 7955 Van Reenens Crest
Set 2. MBB7956 Van Reenens Crest – west 7955. (figs 12-34)
Despite apparently favorable habitat these plants were about 300m west of the previous set. Variation is again apparent in respect of presence or absence of surface roughness. Generally there are plants in which some leaves are without a point (see Fig 1) while others are acute tipped (mucronate – with a short hard point) or even have a long end-awn. Margins may be smooth or very lightly spined. This seems to be more apparent in young plants.
Fig. 12 View looking east to Set 1.Fig. 13 7956 Van Reenens Crest
Set 3. MBB7957 Van Reenens Crest – west 7956. (figs 35-41)
Again the plants are very variable with dense surface tubercles to virtually none. The leaf shapes vary and in fig.39 that appears to be a group of seedlings there is a remarkable series of different leaf ends and margins. In some plants there is distinct spination of the leaf margin.
Fig. 35 View looking south to Tradouw PassFig. 36 7957 Van Reenens Crest
Set 4. MBB7958 Van Reenens Crest – south west 7957. (figs 42-51)
Fig. 43 shows two rosettes of a plant with a “crocodile-skin” surface. The surface tubercles seem to be flattened and the leaves are mucronate. The following photograph, fig.44, is of a plant with acuminate leaves ending in long end-awns, and the surfaces are relatively smooth – retusoid? Unlike the previous records for the area that are north-facing, this is a south-facing “pressure-burst” of white clay. A nearby habitat of vertical shale that might have been considered favorable for H. mirabilis elsewhere was devoid of plants.
Fig. 42 7958 Van Reenens Crest
Set 5. MBB7959 Van Reenens Crest – west homestead 7956. (figs 52-64)
Figs 53-55 are smallish plants with rounded leaf ends and relatively opaque surfaces. Fig.58 is remarkably like H. retusa, say, at Pienaarsrivier with very awned leaf ends and a high degree of translucence in the leaves.
Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS.Fig. 53 7959 Van Reenens Crest
Set 6. MBB7960 Kruiskloof, E Van Reenens Crest. (figs 65–78)
This population departs from a very broad norm for Van Reenens Crest and picks up the continuity with populations north, east and south. This is most noticeable in the surface texture where the tubercles are smaller and denser. The leaves can be highly rotate, see fig.68. A feature seen occasionally in the Van Reenens Crest populations and as far afield as the Diepkloof (Malgas) area much further south, is the occasional plant with a very stellate (five –pointed star) arrangement of the leaves that are quite horizontal see fig.70. Among the many oddities is the plant in fig.72 with floribundoid leaf ends and a leaf surface where the tubercles seem to be flattened. In fig.73 the tubercles seem to be spined. In fig.75 tubercles are almost absent.
Fig. 65 7960 Kruiskloof
H. retusa ‘nigra’
Set 7. MBB7961 Kruiskloof, E Van Reenens Crest. (figs 79-90)
These plants reinforce my perception that H. retusa and H. mirabilis are uncomfortably closely related. Figs 79 and 80 have the same general form as H. mirabilis ‘sublineata’. Fig.82 appears to me as possibly a hybrid as leaf opacity is deeper and there are marginal spines.
Fig. 79 7961 KruiskloofFig. 82
H. floribunda
Set 8. MBB7962 Niekerkshek. (figs 91-99)
Fig. 91 7962 Niekerkshek
Set 9. MBB7963 Niekerkshek. (figs 100-111)
The plants vary in size and to some degree in colour. This variation in colour from a deep purple to dark grey is very evident in fig.107. The leaves are much flattened and an occasional leaf is pointed. The plant in fig.102 is unusual in having white spots on the leaves. Marginal spination can be absent or very coarse. The leaf surfaces show no venation or translucens, as is expected in this species.
Fig.100 View looking south to Potberg.Fig. 101 7963 Niekerkshek
H. mutica
Set 10. MBB7950 DeDraai Klipbankskloof. (figs 112-123)
Fig. 112 – 7950 Die Draai, E Klipbankskloof
Set 11. MBB7951 Klipbankskloof West 1. (figs 124-126)
Fig. 124 – 7951 Klipbankskloof West 1
Set 12. MBB7952 Klipbankskloof West 2. (figs 127-129)
Fig. 127 – 7952 N Klipbankskloof West 2
Set 13. MBB7953 Klipbankskloof West 3. (figs 130-144)
Fig. 130 – 7953 Klipbankskloof West 3 View westwardsFig. 131 – 7953 N Klipbankskloof westward view of habitatFig. 132- 7953 N Klipbankskloof West 3
Fig. 175 JDV92-64 Very localised.Fig. 176 JDV92-64 Sanddrift
Set 16. JDV85/17 Haarwegskloof. (figs 188-213)
Fig. 188 JDV85-17 Haarwegskloof
There is a very wide range of variants among these 7 population sets. Leaf colour and shape vary enormously and it can only be said that generally the leaves are muticate. Almost invariably any one plant has a leaf or leaves that are also mucronate and in some cases even awned. The leaves may be abbreviated and short and squat or drawn out and elongated. Venation is also varied from many to few or only a single vein on the upper leaf surface. These veins may be straight and clearly separated or they may anastomize and link togther in a reticulate pattern. I did include two pictures of flowers, figs 173 and 174 just to indicate that simply taking two flowers at random one can see obvious differences.
Attention can be paid to figs 145 and 146 of Rondeheuwel where I consider these two plants indicate hybridization with H. mirabilis that does occur about 1km to the northwest. Fig. 147 shows a plant with a rather wrinkled surface.
Conclusion My opinion is that consideration has to be given to the odd way in which the “species” do not share habitat. It is extremely difficult to rationalize an ‘average’ plant for each population and so establish any norm for proper comparison. The populations vary by degree. It is also clear that if one examines the distribution of the species across the study area, it can be observed that H. mirabilis where it occurs with H. retusa ‘nigra’(at Heuningklip and Kruiskloof in the east), is not the same as plants to the west where H. retusa ‘nigra’ is absent. Consideration has to be given to absorption into H. mirabilis. Why H. floribunda should make a re-appearance at Rietkuil and Niekerkshek in the west, when everything points to its absorption into H. mirabilis in the east at Skeiding, is a mystery. The fact that at Niekerkshek it also has radically different forms to those found anywhere else adds to the difficulty.
I will conclude by acknowledging that the populations of H. mutica are all summer flowering but I do not think that this is as significant as the authors of “H. groenewaldii” will contend. The Van Reenens Crest complex demonstrated dramatically that H. retusa and H. mirabilis share a very close common history and an equally close contemporary relationship. My conjecture is that H. mutica is linked to that relationship in the same way the complex of populations are that I assign to H. pygmaea. It is painfully obvious that if floral differences that are so easily detected within populations, expecting to find more evidence in the flowers than exists in the vegetative structures across the genera, and especially the subgenus Haworthia is an absolute pipedream. Floral differences between species are minimal and if anything will provide support for a reduction of species rather than add to the. If any small difference is suggested to separate species, then equal weight must be given to similarities that should then unite species. The contention then that flowers are offering characters that I as an author have ignored, while already wholly untrue, is additionally nonsensical.
Acknowledgement Mr. and Mrs. Trevennan Barry of Van Reenens Crest. Mr. N. Swart of Poststal, Mr. J. duToit of Tarentaal and Wydgelee, Mr. W. duToit of Klipbankskloof, Mr. J. Mathee of Klipbankskloof West, Mr. Vlooi du Toit of Die Kop, Mr. M. Mynhardt of Haarwegskloof, Mr. and Mrs. Nelius Smith of Volmoed, Rondeheuwel, Mr. Adrian Steyn of Wankie Boerdery. Thank you to Kobus Venter and Lawrence Loucka for company and shared observation. ♦
The area concerned is the long and wide contact zone between the Limestone stretching from Bredasdorp to Potberg, and the Bokkeveld shale north of that. The soils and vegetation of the two areas are grossly different. The limestones are agriculturally almost useless, while the shales are prime wheat and pasturage producing soils although relatively low yielding. The vegetation of the shales is Renosterveld and there are very few patches left. Large areas resemble ecological deserts with nothing of the original surface intact. Here and there are shale banks and associated quartz outcrops and also some remnants of tertiary deposits that overlie the shale. Under this deposit layer the shale has decomposed to kaolin and in places there are gravel sheets of fine quartz on white clay. The skeletal nature of these remnants is the saving grace but it is unbelievable to what lengths farmers must have gone to make fields arable. Enormous amounts of stone that have been carted away and dumped to make cultivated lands. Sadly the stone is often dumped on exposed rock and prime Haworthia habitat. The remnants are still under threat and a mindset that has developed in the road construction and maintenance arena is that roads must be clean and scraped fence to fence. Similarly there are farmers who want every square inch under control and in subservience to their production needs. Dense vegetation is abhorred and burnt to control predation of sheep by jackal and lynx. Vegetation adjoining crops is treated with weedkiller to minimize crop contamination. Crops are also grown in conjunction with animal production. When crops are in, the animals are on fallow land and on whatever is left of natural vegetation. It is the harsh reality of conservation.
Haworthias exist on those remnants. I have often pondered that maybe they have not been so severely suffered by habitat destruction but now I am not so sure. There are many remnants where nothing is to be found, and others are species rich for no obvious reason. It is therefore difficult to assess.
This chapter explores the occurrences of two species at 11 localities in the Ouplaas vicinity east of Bredadsorp, and records some of their oddities.
The area in question:
This is shown in the map Fig 1 and I need to re-capitulate what I observed on Die Kop because I do not think I have given proper account for the populations I saw and the variance that I encountered in my first exploration there. This is most unfortunate as the name H. hammeri has since been applied to one or more of the variants and without context. Changes have taken place there too, that now compound the issue. I have in Vol. 3 Pt. 1 given an account of H. mirabilis in the wider area and will not repeat all of it, restricting myself just to those few populations that impact directly in these new observations. What is considerably more relevant is the chapter 13 in Part 2 where the main role players are listed and now again emerging in stark reality..
I have changed the arrangement from that in previous accounts to deal with local groups. Thus three sets, H. mirabilis at Die Kop – 5 populations and re-visits, H. mirabilis at Spitzkop, Haarwegskloof and Langvlei – 5 populations, and H. mutica at Spitzkop,- 3 populations.
Fig 1. Map of area and points covered
SET 1 – Haworthia mirabilis Die Kop
Subset 1 MBB7500 Die Kop E. Figs 2-8
Considering the wide array of populations and variants in the greater area, I concluded that these are most probably the result of interaction between H. mirabilis and H. mutica. The surfaces vary from shiny minutely tubercled to smooth, leaf tips rounded or pointed and flowering time is summer. This particular population is in a deposition area off the Silcrete inselberg.
Fig. 2 7500 Die Kop E
Subset 2 MBB7502 Die Kop N. Figs 9-10
When I first explored here in 2006 I skirted north of the first hill on Die Kop (see fig.1 Map) and observed an array of plants that were more like H. mirabilis and I dismissed them as such. However Jakub Jelimicky and Gerhard Marx both indicated that their observations contradicted mine.
Fig. 9 7502 Die Kop W
Subset 3 MBB7973 Die Kop N. Figs. 11-27
We went further north on this excursion passing through the area touched on in 2006 that was now dramatically different. The previous shrubbery was absent and the area was trampled and grassy with also Bobartia, a large grass-like Irid. It seemed unlikely that Haworthia previously there could have survived, but we had seen them before still further west and they may still be present. This new area we saw was densely populated with plants similar to but smaller than MBB7500.
Fig. 11 7973 Die Kop N
Subset 4 MBB7503 Die Kop mid. Figs. 28-34
My memory of these plants, east of those above, was that they inclined more to H. mirabilis than plants from 7500. Fig. 18 is almost identical to a clone photographed at 7500.
Fig. 28 7503 Die Kop mid
Subset 5 MBB7974 Die Kop mid. Figs. 35-46
Change due to heavy rain and erosion was more apparent here. There was less influence of H. mirabilis and the resemblance to 7500 nearly complete although the plants were generally smaller. This may be because the substrate was more skeletal and eroded than at 7500. What was really curious was the plant pictured figs 45-6 with narrow slender leaves. We did not observe any other species there that could have generated this as a hybrid but we did not have opportunity to comb the entire area small as it is.
Fig. 36 7974 Die Kop mid
Subset 6 MBB7504 N Die Kop. Figs 47-55
This population is about 1km north on an isolated inselberg. The plants are small and more typically mirabiloid with rough surfaces and spined margins and keel. Fig. 55 shows a plant with characteristics of the 7500 plants, but otherwise I would say this population perhaps relates better to Tarentaal MBB6539 (see Updates Vol 3.1 note 17).
Fig. 47 7504 N Die Kop
SET 2 – Haworthia mirabilis Haarwegskloof and Spitzkop.
Subset 7 JDV86-2 Haarwegskloof. Figs 56-63
This subset with subset 11, are included as a reminder of what other populations in the area are like. Originally when so much less was known, this was classified as H. heidelbergensis var. scabra and subsequently so many variants have been found that naming each is not deemed practical by me. It is probably 3-4km west of the following three populations and similar populations occur about the same distance further south at Kathoek and Beyersdal. There are other populations known to me that I have never sampled or photographed and the following task is to explore a large area between Bredasdorp and Spitzkop.
The plants at Haarwegskloof, at the time the pictures were taken in late spring, were both with translucence and without. The plants were small and with leaves erect.
Fig. 56 JDV86-2 Haarwegskloof
Subset 8 MBB7964 Spitzkop West. Figs 64-81
Fig. 64 indicates how difficult it is to find these small plants until you know exactly what you have to look for. One starts the search walking and eventually end upon all fours! Fig. 65 also shows how cryptic the plants can be. The plants are some of the smallest I have ever seen. Fig.72 shows an unusual plant with rounded tips to the leaves and I will show this again in another subset. Why this is so significant is because if the H. floribunda similarity in so many of the populations in the general area. The photographs disappointment me in that my first reaction on seeing the plants, was the association with images of H. parksiana. The pictures do no seem to fully reflect this. Kobus Venter has related his experience of finding H. parksiana-like plants among seedlings of H. mirabilis ‘magnifica’.
The leaves are reflexed as in H. parksiana as they of course are in many populations of H. mirabilis ‘maraisii’. But it is a curious reminder of the floribunda-like leaves in so many populations and the problem of look-alikes.
The plants were on a south slope in an area with white quartz and we had identified this as the most likely place to find them out of the direct north or northwest aspect. But we then did find them there too.
Fig. 64 7964 Spitzkop West
Subset 9 MBB7965 Spitzkop West. Figs 82-86
The plants were again hard to find over quite an extensive area. Fig. 83 again shows the floribundoid leaf tips.
Fig. 82 7965 Spitzkop West
Subset 10 MBB7966 Spitzkop West. Figs 87-103
It is curious how these small localized quartzitic areas can be so different. This was about 500m further north. There was more kaolin present as well as fine gravel. At an intervening similar habitat we found only Acrodon demidiatus, absent from the previous habitats and also from this one where Gibbaeum austricola now appeared. Again all the plants were small with dark green leaves, virtually no translucence and surfaces ranging from mildly spiculed to only minutely tubercled or just rough. Figs 99-101 are of plants with distinctly floribundoid leaves.
These are in white quartz and Aloe ferox is also present. There is moss present which is not the case at Spitzkop but true of Haarwegskloof. Curiously the pinched leaf ends as in fig. 106 have been observed in some plants of H. mutica too.
Although a small area of quartz in a cultivated field, the plants are densely concentrated in small areas. There were some dense clumps in the rocks and this is unusual for H. mutica. It is the variation in these plants that deserves close attention. Some plants have very rounded leaves, they can be quite elongate or short and squat, others have an end-awn, the leaves may be opaque or even translucent; the surfaces may be shiny and minutely tubercled or smooth. Colour may be reddish, golden, or green. Most striking is perhaps the venation. There may be anything from 3-13 straight lines or very anastomizing (veins joining). There is no sign of either true turbercles or surface spination and also none of marginal spines.
Fig. 109 7967 Spitzkop (quartz rock)Gasteria carinata is often severely animal grazed.Looking SE towards Die Kop and PotbergAlso looking SE towards Die Kop and PotbergViewing site from the north
Curiously there is a shale ridge (fig.149) between this population and the first that one would think would be suitable habitat for this species as it is very like the Ouplaas habitat. But they were absent, perhaps because the ridge was lower down the slope. But a little higher where the shale was also exposed in smaller ridges the plants were also absent. Where they did occur was among small quartz pebbles to which the quartz may fragment. Fig. 157 shows a plant that was under the protection of small shrub and shaded.
Plants absentPebbles and plants presentThis is not how one expects to see this species.Fig. 152 7968 Spitzkop (pebbles)Showing the small shale ridgesLooking to the previous habitat of larger quartz rock
Subset 14 MBB7969 Spitzkop East. Figs 160-161
This locality is about 3km east of the previous two at a slightly lower altitude. There is very little quartz but well exposed shale creating apparently suitable habitat as at Ouplaas now a short distance away. The area is well utilized by grazing animals and was very weedy, which may account for the fact that we found only two plants and these were not in the shale where we would have expected them.
Fig. 160 7969 Spitzkop East
Conclusion I must here again relate the story in respect of KG35/70 of which there is only an herbarium specimen. That was found more than 40 years ago at Verfheuwel about 15km eastwards from Spitzkop. I was scrambling up a rough animal track to the top of the Verfheuwel hill when I saw some dried seed stalk less than 10cm high. The plants were tiny, greyish leaved with marginal teeth and rounded tips. I simply did not know what to do with them and identified them tentatively along with a few other populations from that wider area as H. maraisii. I went back to look for them several times with no luck. In 1996 I was with Kobus Venter and we found a strong population of larger plants not far away on the same hill. I recognize now that those first plants I saw were floribunda-like and that this similarity is commonly seen in populations of H. mirabilis in this area.
I speculated in Update 3 about the origins of H. mutica in the context of H. mirabilis and I do contend that these three H. mutica populations show characteristics that support this view. The Die Kop populations suggest to me a more recent amalgamation of H. mutica with H. mirabilis, to virtually reverse the process. The floribunda-like characters and even the parksiana resemblance also support this image of very close common origin and that particularly H. floribunda is simply a polytopic phenomenon in this south western area (polytopic – similar elements arising independently from the same common gene pool) and its genetic resources are embedded in H. mirabilis particularly. This seems to be definitely the case in the greater Swellendam and Heidelberg areas.
What is also so confounding is that now we have H. mutica occurring virtually undifferentiated in several different kinds of habitat. This is quite unlike the Van Reenens Crest area where similar habitat does not even have the same species. The H. mirabilis at Spitzkop west is unlike the other small plants in populations of H. mirabilis in the area and does seem to be differentiated according to habitat.
Acknowledgement: Mr. Johannes and Pat Uys of Die Kop, Mr. Vlooi and Adele duToit of Die Kop, Mr. Gawie de Wet of Spitzkop (West), Mr. Franko de Wet of Spitzkop (East), Mr. Koos Badenhorst of Langvlei and Mr. Martin Mynhardt of Haarwegskloof. ♦
No conclusion is written to this appendix. It is evident to me that there just is no way for me to generate a composite picture for each set of flowers by which one can adequately compare. In general the summer flowering cf. H. retusa set have flowers that are correspondingly lighter coloured than those of the darker flower of the correspondingly darker H. mirabilis set. While gathering the plant pictures from files it struck me how similar plants are from quite different sets across both groups and persuading me that H. retusa and H. mirabilis are in fact one integral system and H. mutica and H. pygamea are western and eastern offshoots.
Acknowledgement I must again appreciate the kindness and helpfulness of landowners in allowing access. Neels and Suzanne Smit of Volmoed; Hector Odendaal of Dankbaar; Jon and Cindy Webber of Klipport; Tom,Trevennan and Hesphia Barry of Van Reenens Crest, and Coetzee and Surieta Uys of Morning Star. The present owner of the Bromberg site could not be established. The other plants photographed are not field plants and neither can they be seen as any more than flowers of one or few cultivated specimens with provenance already acknowledged.
The following additional populations and plants were observed:-
1. MBB7954 H. mutica, Volmoed. 2. MBB6512 H. mutica, Klipport. 3. MBB7741 H. mutica, Dankbaar. 4. & 5. MBB7920 and MBB7921 H. retusa (nigra), Van Reenens Crest. 6. MBB7803 H. rossouwii, Morning Star, S. Heidelberg.
In addition I include pictures from plants in cultivation:-
7. MBB7758 H. retusa, Skietbaan, S Riversdale. 8. MBB7780 H. retusa (geraldii), Komserante, E Riversdale. 9. MBB7781 H. retusa (foucheii), Komserante, E Riversdale. 10. MBB7776 H. retusa, Pienaarsrivier, W. Riversdale. 11. MBB6747 H. pygmaea, Vleesbaai. 12. MBB33 H. comptoniana, Georgida. [ed.] MBB33 doesn’t exist in Bayer’s Accession, correct reference is most likely KG114/72
The population H. rossouwii (elizeae) at Bromberg, Stormsvlei was visited but the plants were only in very early bud.
The Heidelberg H. rossouwii is a small solitary plant and only known to me from two populations. H. rossouwii (elizeae) is also small, proliferous and very problematic. It is so far only known to me as one very restricted population. I doubt if the eastern end of the Riviersonderend Mountains have ever been adequately explored to exclude it from there. Even the Bromberg alone could yield more populations.
MBB7954 H. mutica, Volmoed.
2. MBB6512 H. mutica, Klipport.
3. MBB7741 H. mutica, Dankbaar.
MBB7920 Van Reenens H. retusa (nigra), Crest.
MBB7921 Van Reenens H. retusa (nigra), Crest.
6. MBB7803 H. rossouwii, Morning Star.
7. MBB7758 H. retusa, Skietbaan, S Riversdale.
8. MBB7780 H. retusa (geraldii), Komserante, E Riversdale.
9. MBB7781 H. retusa (foucheii), Komserante, E Riversdale.
10. MBB7776 H. retusa, Pienaarsrivier, W. Riversdale.
This is an unusual population in that it is the most northwestern one known although only about 15km from Klipport nearer Stormsvlei. It is the source of the single plant that developed gross milkiness and for which I coined the name “Silver Widow”. The population was first recorded by a Mr. Meiring who worked as a nurseryman for the Bonnievale Nursery of Hurling and Neil in the years at least prior to World War 2. There is a letter in the Compton Herbarium archive which records a transaction between Meiring and Triebner of 100 plants as 1s ea. (i.e. = 10 cents in today’s currency.). I really struggled to find plants again in a very disturbed area and eventually did so after I had placed “Silver Widow” there in her pot to see if the flowers would be pollinated in situ. Indeed they were and then found about 20 plants in a very small area nearby. This visit owes to the report by Jakub Jilemicky of still more plants a very short distance again from these. Here there are about 80 plants in an area of about so many square meters.
Illustrations of the plants and flowers are given. The only comment I can make is to repeat that I never ignored the flowers out of ignorance. The fact is that we have a number of pretentious experts who have no concept of species other than an inherited vague notion that species have something to do with the capacity to interbreed. The fact that Haworthia plants do so freely does not handicap their enthusiasm for new species either. So this is a bit of irony. My contention was that if you did consider flowers there would be a lot less species than even in my conservative approach. In fact I am quite sure a competent professional taxonomist might well consider that I have been too generous with species and too kind to my critics.
(image files mislabeled as 8012, but are 8018 – ed.)
Acknowledgement I thank Gerhardt Swart for access to the site and to Jakub Jilemicky for informing me of the new find. ♦
The species definition Bayer uses is that populations are fractal and DNA is governed by mathematical non-linearity. What does that mean? We have space with two dimensions latitude and longitude, and we have time with two dimensions – calendar time, and the speed (instability) of the arrangement of the DNA base pairs. At any moment in linear calendar time there will be an arrangement of the DNA depending on the stability of the DNA. At one time there will be a clear set of ‘species’ and at another time a different set as the mix of characteristics continually change within and between the populations. ~ Lawrence Loucka
Haworthia mutica (groenewaldii) and its twisted leaves. By M.B. Bayer
There is some argument about the status of this population of plants at Buffeljags. I have explained my opinion of it based on a species definition that I use. I also have reported on three other populations a short way away at Rotterdam. Furthermore I have discussed the flowers at length in a flower report. These are all available free on Haworthiaupdates.org. Gerhard Marx does not agree. The disagreement first has its roots in what constitutes a species and Marx stays with the standard view that characters are what define species. I opt for the view that species are systems related to geographic distribution and to all the elements that drive vegetation and change (evolution + equals change from some unknown initial condition). I think these patterns of change and difference are fractal i.e. detailed pattern repeating itself. Perhaps it would be more correct to say that the organization of pattern is according to a mathematical function which is non-linear. This means that the end product has many different outcomes. But this is complexity just of argument that none of us can deal with.
What comes into the geographic nature of species is also the nature of habitat. What happens is that we have a set of apparent species in Haworthia with a known distribution range. These species are primarily H. retusa and H. mirabilis. There is clear evidence that H. pygmaea and H. mutica emerge from a milieu of populations of those two and that H. floribunda is also involved. Buffeljags is geographically central to this arrangement and the habitat (wrongly described in the description of H. groenewaldii) is very unlike those where the named species generally occur. It is thus no surprise that the plants appear different. The Buffeljags population and its habitat also differ to a small degree from the west side of the river, but both are essentially geologically fairly recent river alluvial deposit.
Marx is insistent that the plants at Buffeljags are so different as to be a discrete species and I disagree. My disagreement is based on my experience of characters in plants. In Haworthia I think these are few and obscure. Thus it is almost impossible to delineate or circumscribe a species by characters and no one has succeeded in producing an identification key that can work. All the differences of opinion and argumentation about names come down to this issue of a species definition and the characters available to recognize them.
The essential points made for H. groenewaldii as a species is the shiny leaf surface and the flowering time. H. mutica does flower four months earlier – agreed. But H. marginata in the same close area also flowers similarly out of synchronicity with H. marginata elsewhere, as H. minima was also observed to do. In elaborating differences for H. groenewaldii, Marx offered the facts that the plants had fish-tail buds and that H. mutica did not. Very soon after he stated that he did not actually know what the case was in H. mutica.
Fig. 1 to 9, MBB7801 Haworthia mutica (groenewaldii), Buffeljags.
In addition he maintained that the leaves in H. groenewaldii were different as follows…”In terms of the angled newer leaves of H. groenewaldii, have a look at the plants again and you’ll see the young leaves are consistently twisted sideways. A spiraling effect. This never shows up in H. mutica”. I find this statement very odd because such a structural difference would come down to a difference on the level of genus or even family. So I looked at the plants I have in my possession and provide illustrations here to demonstrate no significant deviation in respect of twisting. I have even included a picture of a mature plant (fig. 10) of H. mirabilis to show the same “character”. The spiralling effect is universal in the aloids and is even visible in those species with distichous leaves. In the retusoids, where H. mutica belongs, the leaves have been said to be 5-farious. More usually it is possible to see them as trifarious. In young seedlings the leaves are bifarious as the very basic spiral effect comes into play.
10. 7912 H. mirabilis, Rietkuil
I do not think this is a character one can use to distinguish a species at all. There are many cases where it is fairly possible to characterize populations by a wide range of so-called characters. My opinion is that generally in the many species (by my definition) of Haworthia this can virtually only be done at individual plant level. The Buffeljags and Rotterdam populations are simply the western counterparts of populations around Albertinia (eg H. mirabilis ‘splendens’, H. pygmaea ‘fusca’, H. pygmaea ‘esterhuizenii’) that emanate from the relationship of the prime species that I named above.
11. 7916 H. mirabilis, Van Reenens Crest.12. 7925 H. mirabilis, Van Reenens Crest13. 7945 H. muticaX mirabilis, Klipbankskloof.14. 7967 H. mutica, Spitzkop.15. 8005 H. retusa (turgida), Wegwysersrivier.
Reminiscences – a particularly nice plant of Haworthia mutica.
It seems a picture cannot ever capture the essence of the living plants in nature. These are also H. mutica down south and east of Bredasdorp.
If I were asked to explain how things are put together in creation, I would answer something like this…
Are we knowledgeable? This seems such strange question to ask until one really begins to explore the meaning of information and knowledge. We really seem to live in a world of confusion and disbelief. I ask the question in the sphere of plant classification and this exercise in arranging and organising plants by form and appearance into a system of names an classification that is supposed to make sense and lay the foundation for the basis of still more information and knowledge. But does it actually do that and serve such a profound function. The more I learn of the subject and try to understand all its ramifications, the more concerned I am that we have not addressed Prof. A. Croncquist’s question to his fellow international taxonomists at a world taxonomic summit (Miami, Fairchild Arboretum 1987?)…”Do we know what we are doing?”
Well how can we know if we do not even have a well-defined and decisive concept of what a “species” might actually be? Adding to the confusion is the Linnaean binomial genus/species system that insists there exists a natural system of creation based on idea of survival of the fittest and adaptation of living things to environmental influences. There are scientists today who question this now aged conviction and ask very pertinent questions about the nature of DNA, function and form. It is long time we reviewed belief, convention, habits and systems that do not truly fulfil the need and promises of the time.
Think for a moment what leading writers like Rupert Sheldrake, Stephen Meyer and Donald Hofmann are trying to explain to use beyond the clamour for recognition, acknowledgement and effect we feel so pushed to achieve. What for the sake of meaning are the Platonic solids and what might they have to do with formlessness and form? It is quite incredible to consider that these “solids” and their significance have been virtually ignored in the modern era. The “solids” are the five shapes with regular sides and faces that can be fitted into a cube. The term “solid” is simply a misnomer. They are the diagrams of five energy fields each representing one of the five basic elements of nature viz. earth, water, fire, air and ether. They arise from either the numerical zero or infinite zero at the centre of an imaginary cube, and the points of intensity at the apices of the triangles represent the energy matrices of an expanding cube. Such basic elements of nature were well-known in mysticism and ancient religions and their significance is quite lost in a society mislead by intellectualism and belief systems.
The significance of the energy fields is that they are formless but all forms in creation arise from the organisation and patterns of energy derived from an infinite series of such fields and all matter in creation is organised according to these energy fields.
It is so nice to be able to share memories like this. The mysterious ever different vein patterning calls to mind Dancing wu-li masters. Colour too is so enchanting where the leaves can allow or disallow light into the deep interior. Leaf shape is just as mysterious and no two leaves are ever the same. Language is such funny stuff. “Mutica” is supposed to mean without a point. When Col. Scott encountered H. springbokvlakensis he first took it to be the missing H. mutica that had not yet been identified to a field source and population. G.G. Smith had made a similar mistake and described H. otzenii without realizing that it was Haworth’s H. mutica. I really need to see some old reference material. I am sure the translation for the Latin descriptive term “retuse”, is given by Haworth as meaning “bent back like a thumb”. It is not the contorted shape of a flat leaf with the margins overgrowing the mid point of the leaf as in present day use. In H. mutica there is in fact every degree of “non-pointedness” of the leaves. Perhaps this is one of the significant ways in which Haworthia is mistaken to be different from other plant genera and species. Smith followed Reynolds in how he had collected and described “species”. Now Aloe and Haworthia ARE in the way in which a single specimen of, say, Aloe ferox in any one population can in a general sense taken to represent what Aloe ferox looks like anywhere in its distribution range. This sure does not apply in the haworthiads.
Another 3 from the same population as the previous 5 mutica. Emile Heunis once commented that this was his favourite H species, which coming from the plant wizard he is, is a real recommendation. I can never get enough of these magical nuances of texture colour and shade.
More mutica pictures to muse over. Check individual leaf shapes and especially the dancing flames of the veins as they leave the protected leaf base.
I should make a collage of individual leaves to illustrate venational differences acrooss the spectrum, from plants with very few, many, very windowed and then simply straight. It is the fun stuff that attracts us?
These mutica plants I have posted recently are all from the same place and here are two clones that demonstrate huge differences in venation. Incidentally, some of the clones in this population were very proliferous.
This was such a strong population. It seems so strange musing about variability and how we each deal with it. I sometimes just examine the leaf shapes and reflect that often no two leaves have the same outline and end shape.
I am at the end of my active life and realise there is still an important statement to make about these plants. Not only do the plants change in respect of where and under what conditions, they change in respect of our own personal perceptions and attitudes too. Do our perceptions and attitudes change like this as well. There are the last 4 mutica pictures I have of this one population. It is extraordinary to think of it as something permanently etched on the screen of time. No matter who or what does what, it can be taken to exit and it is just a matter of having valued that experience of contact and its existence.
I have often taken people into the field – or at least accompanied them, as they make real contact with nature for virtually the first time. Also I have been taken into the field and shown things many times, remembering new experience and plants and animals I had never seen before. Often being amazed by being told that things that look different are the same, and things that look the same are different – despite far too much experience to be so easily fooled. Plant and animal life is just incredibly complex and I think we have all been suckered into looking at things in the wrong way. Gordon Rowley was an amazing man and I am so glad to have met him and benefitted from his opinions and expertise. He was wanting me to maintain the plethora of names against my own wish to simply try and understand the system that was the science of the times suggested was correct. I think Gordon was right and I am sorry I did not have the means and abilities to maintain all those many names in meaningful way. I will look around and see what comes to mind. But largely I will unfairly lay the blame at the magazines and journals that serve collector and grower interests. The prime function of these societies, their committees and activities should be to manage the language and names that make up information and knowledge. ♦