7. Haworthia chloracantha Haw., Revis. :57(1821). Bayer :106(1976). Bayer :32(1982). Scott :52(1985). Aloe chlorocantha Roem. et Schultes, Syst.Veg. 7:641(1929). Salm-Dyck, Monogr. 13:f1(1836). Type: Not preserved. Neotype: icon, 13:f1 Salm-Dyck, Monogr. Epitype (ex B&M, designated here): N. of Herbertsdale, Bayer in KG411/75 (NBG).
chloracantha: green-thorned.
Rosette from 25-40mm φ, proliferous. Leaves: erect spreading, firm to slightly scabrid, triangular in cross-section, spines on margins and keel. Inflorescence simple raceme, lax. Flowers small.
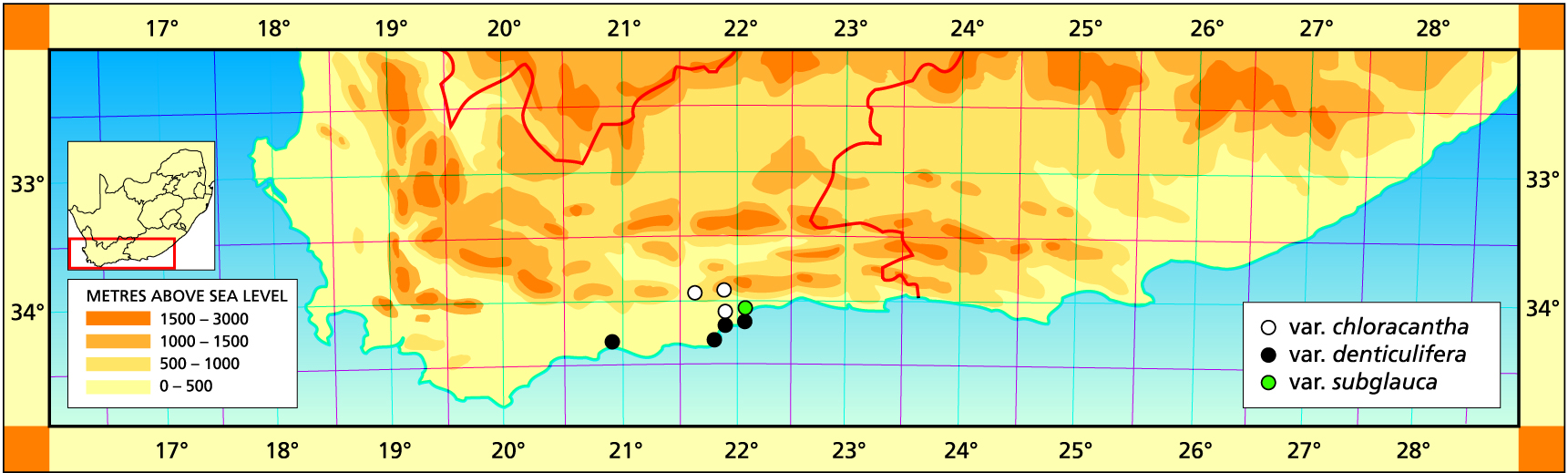
1982 – H. chloracantha is a fairly localised species occurring in the Herbertsdale, Mossel Bay and Great Brak area. It occurs as three main varieties. Firstly the variety chloracantha from north and west of Herbertsdale which is a relatively robust light green form. The var. denticulifera is a smaller, usually purplish-green form found in and around Mossel Bay, while var. subglauca is a more robust waxygreen form from the granitic soils at Great Brak. Von Poellnitz, particularly, confused this species with H. angustifolia but it is slightly more scabrid and the marginal teeth are larger and wider apart. Although more robust, the var. subglauca may reach up to 60-70 mm tall with leaves up to 10 mm broad at their widest. The var. denticulifera may be as small as 30 mm tall with leaves less than 3 mm broad at their widest. The relationship between H. chloracantha and H. floribunda to the west is obscure. The Gouritz River valley effectively divides the two species but there are dubious populations north and south-east of Albertina which may suggest a relationship between these two species (see H. floribunda). However, it is unlikely that either H. floribunda, H. divergens or H. variegata will be confused with H. chloracantha.
1999 – This species was related to H. angustifolia even prior to the recognition of H. monticola ( H. divergens Bayer, 1982). H. chloracantha is more probably more directly related to H. floribunda and H. variegata, but it is also possible that the relationship of the varieties given here is incorrect and that the typical variety has weaker links with the southern Cape than the other two varieties. The new H. monticola var. asema from Calitzdorp, as well as the discovery of H. outeniquensis, also need to be taken into consideration.
It appears that the dark-green erect plants from southeast and east of Albertinia (Cooper Siding) should be regarded as H. chloracantha and not as H. floribunda. The population at Draaihoek to the north, includes plants which resemble H. parksiana and it may best be related to H. floribunda. Plants with erect leaves growing with H. parksiana at Groot Brak have been regarded as H. floribunda and it is more probable that they are in fact also H. chloracantha. It seems improbable that three similar species, in which the affinities with one another are in question, can co-occur.
a. var.chloracantha. The typical variety is taken to be the very proliferous green forms around Herbertsdale and along the Gouritz River as it passes through the Langeberg mountains. While Breuer and Metzing do not use the Salm Dyck illustration to typify this name, that illustration and the icon in Bergers revision in Fas Pflanzenreich, are really the main sources for the application of the name.
Haworthia chloracantha var. chloracantha JDV87/80 Hertertsdale. In cultivation, this variety often appears pale and chlorotic.
Haworthia chloracantha var. chloracantha JDV97/138 Herbertsdale. The typical variety occurs only in this area and is always a plainer and yellower green.
b. var. denticulifera (V.Poelln.) Bayer :112(1976). Bayer :32(1982). H. angustifolia var. denticulifera V.Poelln., Feddes Repert.Spec.Nov. 41:194(1937), ibid. 44:228(1938). Type: Cape, Montagu, Mrs Helm. Not preserved. Lectotype (B&M): icon (B).: H. angustifolia var. lilliputana Uitew. Sukkulenta 43(1953). Type: Not preserved. Neotype (designated here): CAPE-3422 (Mossel Bay): Hill above Mossel Bay (-AA), Courtenay-Latimer in Smith 5223 (NBG).
denticulifera: bearing small teeth.
This variety is dark-green and occurs around Little Brak and Mossel Bay. It also occurs along the lower Gouritz Valley where it has been previously assigned to H. floribunda. Von Poelnitz (1938) cited a number of improbable localities for this element as a variety of H. angustifolia. These included Montagu, Calitzdorp, Riversdale and Great Brak. It is generally concluded that the plants originated from the greater Great Brak area. Uitewaal’s smaller plants – var. lilliputana – are comparable with the population which occurs within the town of Mossel Bay itself. The specimen cited from the Duiwenhoeks causeway is also one with erect pointed leaves which draws the relation with H. floribunda into the discussion.
Distribution: 3420 (Bredasdorp): Duiwenhoks Causeway (-BD), Muller-Doblies 82/098 (NBG). 3421 (Riversdale): Cooper Siding (-BB), Bayer 3586 (NBG); 3km N Gouritzmond (-BD), Smith 7519 (NBG), Bayer 3586 (NBG). 3422 (Mossel Bay): 2km N. Mossel Bay (-AA), Smith 2896 (NBG); 1km N. Mossel Bay (-AA), Smith 3958 (NBG); Mossel Bay (-AA), Smith 5223, 5750 (NBG); Little Brak (-AA), Smith 5751 (NBG); Great Brak (-AA), Bouwer (NBG).
Inadequately located: ex hort. Armstrong in Smith 2830 (NBG).
Haworthia chloracantha var. denticulifera JDV88/17 Hartenbos. Usually a dark green plant with fine spination.
Haworthia chloracantha var. subglauca JDV87/79 west of Great Brak. Sometimes almost bicoloured with reddish tips above the dark green leaves.
c. var. subglauca V.Poelln., Kakteenkunde 9:135(1937). Bayer :106(1976). Bayer:32(1982). Type: Cape, Great Brak, Mrs Helm. Not preserved. Neotype (B&M): Great Brak, Hurling & Neil (BOL).
subglauca: nearly glaucous.
As stated above, this variety differs in coloration from the other two varieties and in the larger sparser spines. There is a specimen cited rather vaguely as from near Zebra which could perhaps be H. outeniquensis.
Distribution: 3422 (Mossel Bay): Great Brak (-AA), Fourcade 18 (NBG), Hurling & Neil (BOL), Ferguson 1 (BOL), Smith 2889 (NBG); E. Great Brak (-AA), Smith 2885, 3957 (NBG); Bayer in KG(98/71 (NBG); George (-AA), Malherbe in NBG304/40 (NBG), Fourcade in NBG2617/34 (NBG).
Inadequately located: ex hort, Pillans (BOL). Heyn’s farm near Zebra, van der Bijl 474 (BOL).
Haworthia chloracantha var. subglauca JDV84/58 south of Great Brak. Larger leaves which are a waxy bluish colour.
Haworthia chloracantha var. subglauca JDV94/3 north-east of Brandwacht. This population seems to be intermediate between vars. subglauca and denticulifera. Very dark coloured.
Haworthia chloricantha var. subglauca JDV84/58 south of Great Brak. Plants can be very withdrawn into the soil.
Introduction Haworthia Revisited was drafted in 1996, and since then the first author has undertaken a number of field excursions in an attempt to clarify uncertainties. The putative nature of species of Haworthia as recognised by Bayer (listed in Haworthia Revisited, Umdaus 1999) and the importance he attached to geographic distribution are stressed in all his publications. This is because these so-called species seem to vary continuously with one another in that context of geography. Classification seeks to portray relationships and origins. Hence when a species has been recognised, a cognitive attempt has been made to speculate on phylogenetics, where distribution must be significant. In the case of Haworthia floribunda this proves rather difficult, and this article is a discussion of the relationship of this species to its possible relatives. The point we do make is that the Linnaean binomial system, as well as cladistic methods, seem neither to deal with nor portray the problem of reticulate relationships. In other words, the nomenclatural system and the way we classify plants and analyse their relationships assumes linear dichotomy in those relationships.
Considerations H. floribunda was described by von Poellnitz in 1938. It was preceded by H. parksiana in 1936. Other species which need to be considered are H. chloracantha Haw. (1821), H. variegata Bol. (1929), H. magnifica VPoelln. (1933) and H. maraisii VPoelln. (1935). Each of these species, although H. parksiana to a lesser degree, has complex variability within assignable geographic space. The difference with H. floribunda is that it is peripheral to all of them.
Bayer, in his Haworthia Handbook (1976), writes of H. floribunda ” … It is not certain what the relationship is between this species and H. chloracantha var. denticulifera which is found north and south of Albertinia on the west of its distribution range. Thus H. chloracantha abuts on H. floribunda and may be geographically continuous with it.” In the New Handbook (1982) Bayer writes “There is a known population north of Albertinia in which the plants have more and shorter leaves, as well as another similar population near Gouritzmond. These two populations may suggest an affinity with H. chloracantha (here an unpublished comment by A.E Speechley is added which probably has its origins in Bayer’s note of 1976, or in private communication between the two persons). There may be such a relationship, but it seems likely that H. floribunda and H. parksiana are in fact related. They both tend to grow well‑shaded and in moss and lichen.”
The problem is not resolved in Bayer’s more recent revision (1999). He does recognise three varieties of H. floribunda. These are:
Fig. 01. MBB158 H. floribunda. Type locality, N Heidelberg.
a. the typical one comprising a single population north of Heidelberg (Fig.1 MBB158).
b. a similar single population near Swellendam representing the var. major (Figs.2a & b MBB6859).
Fig. 02a. MBB6859 H. floribunda var major. S Swellendam.Fig. 02b. MBB6859 H. floribunda var major. S. Swellendam.Fig. 03b. JDV89-17 H. floribunda var dentata. NE Riversdale.Fig. 04. MBB2311 H. floribunda var dentata. N Albertinia.
c. the remainder, the var. dentata (Figs.3a & b JDV89/17).
In the discussion of H. chloracantha, it is also stated that a population north of Albertinia (i.e. Draaihoek, Fig.4 MBB2311) considered as chloracantha in 1976, considered non-commitally as chloracantha in 1982; ” … includes plants which resemble H. parksiana and it may best be related to H. floribunda“. This collection is cited under H. floribunda var. dentata. Thus there is a degree of uncertainty for which the author offers an apology, and here attempt to summarise the situation a little more comprehensively.
Fig. 04. MBB2311 H. floribunda var dentata. N Albertinia.
There are some red herrings in various comments made by Bayer. These include reference on p48 of Haworthia Revisited to the lower Gouritz population (Fig.5 MBB5460) “previously being assigned to H. floribunda“, possibly referring to a Smith identification on a photograph and cited under H. chloracantha var. denticulifera. There is a similar population nearer Albertinia (Fig.6 MBBsn). Also there is a citation of a specimen from the Duiwenhoks Causeway (Muller-Doblies 89/098) under H. chloracantha var. denticulifera, which may be the same population sampled by Venter and Esterhuizen (Figs.7a & b JDV92/31) from a site named as Dassieklip also on the Duiwenhoks River (the geographical grid references differ and these may be incorrect), but cited under H. floribunda var. dentata.
Fig. 05. MBB5460 H. chloracantha var denticulifera. NW Gouritzmond.Fig. 06. MBBsn H. chloracantha var denticulifera. S Albertinia.Fig. 07a. JDV92-31 H. floribunda var dentata. Dassieklip, S Heidelberg.Fig. 07b. JDV92-31 H. floribunda var dentata. Dassieklip, S Heidelberg.
Haworthia parksiana is known from at least five different sites of which two are recorded by Mr Jan Vlok, viz. at Outeniqua Siding, and north-east of Brandwacht, Mossel Bay. No herbarium specimens of these are known to have been lodged. J.D Venter has commented that parksiana-like seedlings were grown from H. magnifica seed (parent plants ex near Heidelberg). It is doubtful if this can be used to support a claim that this is the origin or relationship of the two species. Bayer has also grown small stunted forms of magnifica from seed of an H. magnifica population south of Riversdale. Such dwarfed individuals have been observed among seedlings derived from other populations. The connection of H. magnifica to the Mossel Bay area is just too tenuous. At Albertinia itself we have the variation to H. magnifica vars. dekenahii and splendens, while to the south west we have the var. acuminata. This is assuming that Bayer is correct in making these combinations which are known to be complicated by consideration (among many) of H. emelyae north of the Langeberg mountains.
H. chloracantha is treated in Haworthia Revisited, as three varieties. The typical variety is considered to consist of the known single population north of Herbertsdale (Bayer in KG411/75). However, Paul Vorster drew my attention to a population at the Wolwedans Dam north of Great Brak which is almost identical and which invalidates varietal distinction defined by geographical considerations (Fig.8 MBB7425). A specimen of this collected by C Burgers is also cited for the Gouritz Gorge where it exits the Langeberg Mountains. P.V.Bruyns and E.A.van Jaarsveld have also made collection from further north in the same gorge, which are uncited. These three populations cannot be categorically included in the typical variety and could be just as at home in the var. denticulifera. The only consideration is probably size and the greenish colour – as opposed to the usually smaller and purplish-green of the var. denticulifera. The var. sub-glauca also does not have a clear geographical base in terms of difference or distribution range, and may just be a much localised ecotype in the vicinity of Great Brak. It occurs close to the Wolwedans population both to the south and to the west. The recognition of these varieties makes it a little difficult also to include the Albertinia and Cooper Siding populations as cited in Haworthia Revisited under H. chloracantha as they are so closely similar, and located, to H. floribunda. The geographical considerations, however, argue for their inclusion.
Fig. 08. MBB7425 H. chloracantha var chloracantha. Wolwedans, Great Brak. MBB7425
The Great Brak/Mossel Bay area contains some endemic species which suggest the operation of a mechanism which could have isolated the vegetation of that area at some time in the past. H. parksiana, H. kingiana, H. pygmaea, H. chloracantha and species like Euphorbia bayeri and Duvalia immaculata suggest this. The Langeberg mountains are an east/west barrier separating the more arid Little Karoo and its succulent Karoid vegetation from the Southern Cape where Renosterveld and Karoid Valley Bushveld are more strongly represented. The Gouritz River Gorge may have been, or is, a similar north/south divide. Westwards, the Swellendam area seems to provide a vegetation interval that H. turgida seems to bridge with difficulty, and which also marks some kind of a break for the H. magnifica/H. maraisii complex. This divide may be a function of inadequate collecting or the lack of adequate habitat. The Breede River valley may, like the Gouritz, present a north/south barrier. It is the home for H. venosa subsp. venosa and also for an endemic asclepiad Stapeliopsis stayneri. West of Swellendam is also the start of the Worcester Robertson Karoo which is vegetatively much closer to the Little Karoo than is the eastern part of the Southern Cape. This area also has its own Haworthia endemics or near-endemics.
The physical commonalities of the three species floribunda, chloracantha and parksiana, are that the leaves are in a stemless rosette, having from 20-40 leaves per rossete. The lowest numbers are in floribunda with parksiana and chloracantha generally having more. The leaves are firm and slightly scabrid. H. chloracantha is more spinose than the other two species and the leaves are triangular in cross-section with a conspicuous keel. In H. parksiana the leaves tend to be short and sharply recurved. In H. floribunda the leaves are more erect, although spreading. Characteristically the leaves are twisted and there is no keel, so the leaf is more effectively strap-shaped particularly towards the apex. The leaf-tip is also rounded with a short point. In H. chloracantha, the leaves are erect and spreading, not twisted. They are keeled and thus triangular in cross section. The leaf tip is attenuated and pointed. The leaf surfaces of the three species are variable. Both H. floribunda and H. parksiana can exhibit quite tuberculate surfaces, although parksiana never has the relatively glabrous surface that may occur in either chloracantha or floribunda. Colouration is also variable across the three species, although parksiana does not occur in the lighter colours that the other two species may exhibit. A hatched pattern of the under-leaf surface may appear. All three species make offsets although parksiana is slower and more reluctant to do so – an observation which may just be peculiar to the individual grower and the clones he has under his conditions.
New material Regarding H. parksiana, the only new records are those of Jan Vlok mentioned above. The writers assume that the identifications are correct and that they establish the discrete identity of that species. Regarding H. chloracantha, there is only one significant new collection. This is one of the var. denticulifera made by Bayer, Kent and Venter. It was made at the same time as a visit to the site of the typical var. chloracantha north of Herbertsdale. Here the plants are on a very steep clay slope (in fact recent rains had caused massive mud-slides). The plants are quite large and form immense chlorotic-green clumps. We were at Herbertsdale to verify a collection of H. pygmaea made by Ernst van Jaarsveld in the same vicinity. This is a short distance away on a steep conglomerate slope. H. pygmaea was present on the dry northern edge. But in the moss and lichen of the cooler eastern overhang were the small single plants of H. chloracantha (Figs.9a, b & c JDV97/136). These were so evidently different from the typical form that we accepted (with reservation) the identification as H. floribunda. One reason for such identification is simply one of informal communication. A similar “floribunda“-like plant grows with H. pygmaea and H. parksiana virtually within Great Brak Township (Fig.10 JBouwer sn). This was collected and referred to as H. floribunda by both messrs. H. Gie and J. Bouwer. It is possible, and probable, that it is in fact H. chloracantha var. denticulifera, as is the new population JDV97/136.
Fig. 09a. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 09b. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 09c. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 10. JBouwer sn H. chloracantha var denticulifera. Great Brak township.
The Dassieklip population (Fig.7) remains somewhat of an enigma. A good comparison can even be made with collection of H. monticola from Trompetters Poort, north of Willowmore, as the plants also have an apparently smoother epidermis than appears evident in chloracantha. This is a highly improbable relationship, but it is necessary to make this comparison in view of what has been speculated elsewhere about the possibility of a continuation between H. chloracantha, H. variegata and H. monticola (H. divergens Bayer!). This speculation has been by Bayer and is not significant enough to cite in detail. What is important is that the Dassieklip population may in fact be closer to H. variegata. Hence its citation under both H. chloracantha and H. floribunda is unavoidably misleading. However, that can only be considered in a detailed evaluation of H. variegata in its relationship to the coastal limestones and re-occurrence in shales both west and south of Swellendam.
The newest finds relating to H. floribunda are in the broad area southwest of Heidelberg, and west of Bredasdorp. However, there is one new record by E. Aslander of plants from northeast of Albertinia (Figs.11a, b & c EA1238) in which plants very clearly have the characteristics of H. floribunda. There are also individuals with the leaves of H. chloracantha. There is another collection by Aslander from Snymanskraal west of Albertinia (Figs.12a & b JDV92/2) which resembles the Draaihoek sample and thus may also be H. chloracantha var. denticulifera. A further collection by J.D. Venter is midway between this collection and Riversdale (Figs.13a, b, & c JDV93/56), and is clearly H. floribunda. A feature of this collection is the striated rugosity of the leaf surface in some clones and also the occurrence of short, almost terete, leaves that are evident in a clone of H. parksiana in a JDV collection.
Fig. 13c. H. floribunda var dentata. Plattekop, E Riversdale.Fig. 15a. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 11a. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 11b. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 11c. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 12a. JDV99-1 H. floriibunda var dentata. Snymanskraal, E Riversdale.Fig. 12b. JDV99-1 H. floribunda var dentata. Snymanskraal, E Riversdale.Fig. 13a. JDV93-56 H. floribunda var dentata. Plattekop, E Riversdale.Fig. 13b. JDV99-1 H. floribunda var dentata. Plattekop, E Riversdale.
Collections from the western areas include:- from a population reported by E. Esterhuizen on the farm Koppies, southeast of Swellendam (Figs.14a, b & c MBB6879) where the plants tend to H. maraisii; from slightly to the east and south of this at Oudekraal (Figs.15a, b, c & d MBB6881) the plants are very like H. floribunda var. dentata as it occurs at Buffeljachts and at the Bontebok Park south of Swellendam. Esterhuizen commented on the appearance of the twisted and flattened leaf-end of H. maraisii north of Bredasdorp. This is confirmed in populations at Napky (MBB7030) and at Adoonskop (Figs.16a, b & c MBB6640) Adoonskop (maraisii) where the plants do indeed look like very robust forms of floribunda. The same characteristic twisted and rounded leaf-tip is evident in maraisii at Napier (Figs.17a, B & c MBB6973) and especially in the seedlings.
Fig. 14a. MBB6879 H. floribunda var dentata. Koppies, Heidelberg.Fig. 14b. MBB6879 H. floribunda var dentata. Koppies, HeidelbergFig. 14c. MBB6879 H. floribunda var dentata. Koppies, Heidelberg.Fig. 15a. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15b. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15c. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15d. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 16a. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 16b. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 16c. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 17a. MBB6973 H. maraisii var maraisii. N Napier.Fig. 17b. MBB6973 H. maraisii var maraisii. N Napier.Fig. 17c. MBB6973 H. maraisii var maraisii= floribunda var dentata! N Napier.
Several other collections confound the picture completely either because of intrinsic variability or because they make it difficult to uphold any geographical recognition of variation. Firstly there is a collection attributable to P.V. Bruyns, north of DeHoop (Figs.18a, b & c MBB6539) which has strong resemblance to H. magnifica var. atrofusca.
Fig. 18a. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.Fig. 18b. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.Fig. 18c. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.
Secondly there is a series of populations from the northwestern end of the Potberg, south of Swellendam and east of Bredasdorp. These are:- Juliesfontein (Figs.19a, b, c & d MBB6882); Brakfontein (Figs.20a, b, c, d & e MBB6886); northern Potberg slopes (Figs.21a, b & c MBB6889); and north of Brakfontein (Figs.22a & b MBB6890). These complement collections Burgers 2506, Bayer in KG35/70 and Bruyns in KG49/76 cited in Haworthia Revisited under H. maraisii, as well as MBB(PVB)6544 (Figs.23a & b), and MBB6545 (Figs.24a & b) cited under H. heidelbergensis var. scabra.
Fig. 19a. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19b. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19c. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19d. MBB6882 H. floribunda var dentata! NW Potberg, S Swellendam.Fig. 20a. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20b. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20c. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20d. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20e. MBB6882 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 21a. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 21b. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 21c. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 22a. MBB6890 H. heidelbergensis var scabra! Witklipkop, S Swellendam.Fig. 22b. MBB6890 H. heidelbergensis var scabra! Witklipkop, S Swellendam.Fig.23a. MBB6544 H. heidelbergensis var scabra NW Potberg.Fig. 23b. MBB6544 H. heidelbergensisv var scabra NW Potberg.Fig. 24a. MBB6545 H. heidelbergensis var scabra! Juliesfontein, Potberg.Fig. 24b. MBB6545 H. heidelbergensis var scabra! Juliesfontein, Potberg.
The influence of other species such as H. mirabilis, H. variegata and H. serrata are in evidence. It can be noted that KG35/70 was of very small plants at Juliesfontein. Returning to the same site 30 years later I could not find these small plants at the original site, and instead found the bigger plants of MBB6882 a short distance away. There are three more collections to be considered:- by Denis DeKok near Swellendam (Figs25a, b & c MBB6644), about 10km west of Swellendam (MBB6861), and from the farm Rondeheuwel south of Stormsvlei (Figs.26a, b & c MBB6882, Bayer in KG326/71). This latter collection has previously been reported (Haworthia Handbook 1976) under one of the populations intermediate with H. mirabilis, and is cited in Haworthia Revisited under H. maraisii. These populations, as well as that southwest of Heidelberg (KG107/74 cited under H. magnifica (Figs.27a,b & c MBB6663, Bayer in KG107/74) confound the issue enormously as we actually have four populations which cannot with confidence be allied with either H. magnifica, H. maraisii or with H. mirabilis. The reality is that neither H. heidelbergensis nor H. floribunda can be excluded from the consideration of these populations. The case for each “species” needs to be dealt with separately. In this case, H. floribunda is reflected in the nature of the leaf shapes. In all the four latter populations given, there are individuals which have the same characteristic leaf shape of H. floribunda, although not necessarily the elongated strap-like leaves of the typical form. In addition to this, seedlings of many different collections of H. magnifica (Kweekkraal, Figs.28a & b MBB6817 – and the reader should refer to the chapter where interplay between H. magnifica and H. floribunda is reported), H. maraisii, H. heidelbergensis, H. mirabilis (Goudini, Caledon, Fig.29 MBB6537), and even H. mutica (Hasiesdrift, Fig.30 MBB6982), have the floribunda-like shape evident in the young leaves. Kobus Venter was especially struck by this phenomenon while photographing H. mirabilis on the Bromberg Mountain near Stormsvlei. These rounded obtuse leaf-tips are maintained until at least the three to five-leaf stages and then largely disappear. Thus there seems to be a distinct sign that the leaf-type is juvenile and that H. floribunda represents a “species” with retained juvenile characters. This may extend to the fact that the seedlings of this species also seem to remain slightly distichous for longer than is the case in other species. In the case of H. floribunda var. major from southwest of Swellendam, the leaves in some individuals may be fully pointed and triangular in cross-section and thus more closely resemble plants in the population of H. variegata var. hemicrypta west of Swellendam. This is the same kind of variation found in the populations about Albertinia, where the speculated differentiation to H. chloracantha occurs.
Fig. 25a. MBB6644 H. maraisii=mirabilis! SW Swellendam.Fig. 25b. MBB6644 H. maraisii=mirabilis! SW Swellendam.Fig. 26a. MBB6862 H. maraisii=mirabilis. Rondeheuwel, Stormsvlei.Fig. 26b. MBB6862 H. maraisii=mirabilis. Rondeheuwel, Stormsvlei.Fig. 27. MBB6644 H. maraisii=mirabilis. SW Swellendam.Fig. 27a. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 27b. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 27c. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 28a. MBB6817 H. magnifica var atrofusca. Kweekkraal, W Riversdale.Fig. 28b. MBB6817 H. magnifica var atrofusca. Kweekkraal, W Riversdale.Fig. 29. MBB6537 H. mirabilis var triebneriana. Goudini, Caledon.Fig. 30. MBB6982 H. mutica. Hasiesdrift, N Bredasdorp.
Flower morphology and flowering time does not appear to offer any solutions. As it is, the flowers have so far been shown in Haworthia to be useful only in so far as recognising the sub-genera is concerned. It can be shown that even at that level, the distinction causes problems for botanists. Most of the species discussed above flower in late summer. While the epithet ‘floribunda‘ was chosen to suggest many-flowered, this is true for other populations considered to be H. magnifica.
Conclusion The circumscription of H. floribunda remains obscure and the situation is in fact exacerbated by new samples which indicate the labyrynthine relationships with several species previously excluded from debate. H. floribunda may have juvenile characters. It does have distinctive populations in a recognisable geographic area. It does not directly share habitat with any species although it does occur very near to H. turgida (north of Heidelberg, where hybrids are also recorded). Where the chloracantha-like equivalent occurs with H. pygmaea var. argenteo-maculosa at Cooper Siding, hybridisation is also evident. It occurs very near to H. magnifica var. atrofusca northwest of Riversdale, growing on a cool southern slope as opposed to the latter on a hotter north slope. The situation northwest of the Potberg, where as many as seven otherwise apparently discrete species need to be included in the discussion, will be very difficult to explain.
It should be noted in closing that the Heidelberg population MBB6663 (as KG107/74) was instrumental in the initial decision by Bayer in 1976 when H. maraisii and H. magnifica were treated as one under H. maraisii. This was repeated in 1982 when H. magnifica was given its chronological priority over H. maraisii. In Haworthia Revisited these two elements are separated as discrete species and the Heidelberg collection is cited under H. magnifica. This decision was made rather to accommodate the varieties which are attached to them. Thus the additional three collections noted in connection with it viz. MBB6860, MBB6861 and MBB6862 all fall within this same indecisive category.
M.B. Bayer, Cape Town, South Africa R.W. Kent, Poway, California
In Chapter 1 of Haworthia Update Vol. 2, I discussed H. chloracantha and H. parksiana in the context of H. floribunda. Fig. 8 in that publication is labelled “North of Herbertsdale” when in fact it is MBB7425 from the Wolwedans Dam north of Great Brak (see fig.1). This was deliberate and not seriously misleading as the plants from the two respective populations are virtually identical. The correct images for that “north of Herbertsdale” are in Haworthia Revisited and labelled JDV87/80 and 97/138.
Recently I had an opportunity to visit Herbertsdale in connection with chameleons and took the opportunity to visit a locality also north of Herbertsdale, but a lot closer (see Figs 2.1-2.11. MBB 7825) reported to me by Mr. Jaap Viljoen of Swellendam.
The plants there are smaller and much less clump-forming than JDV87/80=97/138 and are more reminiscent of H. parksiana. Unlike JDV97/136 (see figs. 3) that are solitary, small, dark-green plants growing in the lichen on a steep east-facing conglomerate slope, the plants are quite green in colour and often the tips of the leaves are slightly rounded and flattened as in H. floribunda. The habitat is west-facing on an old gravel river terrace and much drier and warmer than for JDV97/136, in sharp contrast to the very steep north-facing clay cliff where KDV87/80=97/138 occurs. In neither of these latter two sites are other plants significant, while at MBB7825 the plants are under renosterbush (Elytropappus) and related vegetation. I should mention that my wife on seeing the plants commented on the similarity to H. parksiana and this is not an observation that can simply be dismissed.
About 3km south of Herbertsdale (see fig. 4, MBB7827) we again found very similar plants, perhaps a little more spinose and slightly more clump forming. Neither of these two new populations fit comfortably in the existing formal varieties, as indeed is true for so many populations in most of the other species. I do not think it is rational to attempt formalizing names for them as there is quite substantial variation in each population. Others may do so if these plants ever get into general cultivation from vegetative propagation and onto lists.
From Herbertsdale we went to Klein Brak to investigate a set of four populations on the farm Rooiheuwel as indicated to me by Gerhard Marx. One of the species supposedly there was H. pygmaea and it was rather distressing to find that the Water Affairs Department has seen fit to run a bulldozer over the area with no indication that there was any need for doing so. However, we did find the other three species indicated by Gerhard. One is H. kingiana (see figs. 5.1-5.4 MBB7835).
The others were H. parksiana (see figs 6 MBB7828)
and H. chloracantha (see Figs 7 MBB7829), but this requires a little bending of the truth.
This H. parksiana site is about 400m southeast of an old locality of mine on an outcrop of rock virtually jutting into the road and now largely obliterated. At Gerhard’s site (viz MBB7828) the plants are small and while there is little doubt that they are generally alike to H. parksiana, there is definitely some degree of hybridization and some of the plants may actually be H. chloracantha. This is a rather difficult decision to make on the basis of stressed plants in habitat. This habitat is very restricted to a patch of fairly level aggregated conglomerate so that the plants are virtually in tiny shallow soil and lichen encrusted patches. A little distance further north there is another population to which we assign the name H. chloracantha. This is above a steep loose conglomerate cliff with a little more soil and immediate bush. The plants have rather abbreviated leaves and a darkish colour. There must be some doubt that they are discretely H. chloracantha. The habitat is not so restricted and one is compelled to ask why the plants then so are.
I try to avoid the unanswerable question of how these two species that flower together and seem to be able to hybridize quite freely, maintain their identity and come to occupy such specific, localized and widely separated sites? It would be nice to think that the technology is now seemingly available in DNA sequencing to begin probing these questions, and I will point, with reference to chameleons (see next chapter), to what I think is a disturbing reliance on that technology without the necessary knowledge and experience of what actually can be seen in the field. ♦
Variability – sounds so hackneyed now but still it does not seem as if anybody “gets it”. These are plants from two populations of chloracantha east of Herbertsdale. Two very different soils. One is shale and I show only a single plant from there – the left one of the three unearthed plants. There the plants were “ordinary” chloracantha. But the other locality was exposed “pressure burst” kaolinite such as you get around Riversdale and westwards. Some of the plants reminded me of floribunda and even parksiana. I know of only 6 populations around Herbertsdale and there the plants are different too.
112. 2020.03.26 – The trickster – H. floribunda. This is the Draaihoek Albertinia site where I was not sure (ca 1970) if it was floribunda or chloracantha. I went back to check ca 2001. The place was grazed and trampled to a frazzle – sheep and/or ostriches. I found a few specimens right at the far end. They pass for floribunda and especially if you recognise that floribunda has a good many faces. The last picture is from west looking east – so this is the top rocky edge of a south facing drop. The water is the Valse River. Dekenahii is across the way.
So in these very distressing times, lets see where we go next.
113. 2020.03.26 – This is H. chloracantha from east of Tweekuile along the Valse River north Albertinia, so also further east from Draaihoek. A small dense patch of plants under bush at the upper end of a south facing slope – escaping full exposure to sunlight. A few leaves show the round leaf tip of floribunda but the plants are also much more proliferous than I have ever seen in that ‘species’. It is very useful to consider all the variants in this complex to see how and why I arrive at my opinions on their classification. This is not a glamour complex but as systems they work in exactly the same way. I really regret not having good pictures and especially since I first saw this species at Great Brak in 1969.
114. 2020.03.27 – Ouvloere is east of the previous as that is from Tweekuile but the plants are really odd. Unlike the previous these plants are in and under low ground hugging plants at the top of a slope. This must be about 8km west of the Gouritz River.
How would you know this is H. floribunda?
The answer to … How would you know this is H. floribunda is because I say so? So please forgive my apparent arrogance. I would dearly love to show you all the plants and populations that lead to this conclusion because you would surely be able to then make that decision for yourself. These two pictures are also floribunda from very near to ‘splendens’ at Dekriet (Snymanskraal) west of Albertinia. A strange habitat where the rocky ferricrete is exposed but with a collapsed eroded depression in which the plants grow around the edges. When this time of tribulation is over, I hope to go back and get habitat pictures. Reminiscent of parksiana?
115. 2020.03.30 – The Valse Rivier joins the Gourits at Die Eiland where there is now H. chloracantha and eastwards. It supplants floribunda entirely – or is it truly just the same species?
116. 2020.03.30 – The last post was of plants from N Die Eiland (Buisplaas). From the top edge of a west facing shale cliff. These are also Buisplaas but from a low north facing slope of alluvium. I never imagined chloracantha could be so abundant and so resistant to the impact of habitat. But it becomes one of the most remarkable of the species on closer scrutiny.
The great awakening will be when one can truthfully and openly write about, discuss, the reality of basic life forms i.e. species. When we are free of the prejudices, beliefs and misconceptions of science as practiced and religions as preached. The great awakening will be when one can truthfully and openly write about, discuss, the reality of basic life forms i.e. species. When we are free of the prejudices, beliefs and misconceptions of science as practiced and religions as preached. ♦
117. 2020.03.31 – This is just north of Herbertsdale under renosterbos on shale. A serious lack is pictures of a population nearby on a very steep, north-facing crumbling clay ‘cliff’ with karoid bushveld plants growing wherever they can fit. Here the plants make enormous clumps and themselves are individually large to leaves 60-70mm. Also missing are pictures from a short way to the south-west on an east-facing cliff of conglomerate. There the plants are small, solitary and dark-colour red 😊. These demonstrate the impact of habitat. Of course also seriously lacking is a species definition and agreement on the issue. Systems, systems, systems.
118. 2020.03.31 – This is chloracantha further south at Johnson’s Post with ‘pygmaea’. Large dense clumps seemingly holding clay pedastals together. These are associations of systems and in my opinion very significant in recognising where species stop and start. Not that this simplifies the problem. Incidentally chloracantha in a small green form also occurs in the Gouritz River gorge through the Langeberg.
119. 2020.04.01 – Chloracantha occupies an incredible range of habitats and varies accordingly. These are it at Cooper Siding in feldspathic sandstone. Not far away it is also in transitional zone to shale and the plants also looking a bit different. In both cases it is with “pygmaea” and I have see hybrids despite the season difference in flowering times. Curiously I was once asked … “how do you know it is a hybrid?” Well I do not know how to answer that. It is like asking, “How do you know you which is your left foot?”
Lawrence Loucka – Please say some more about hybrids. ‘I know it when I see it.’ may be true but how to confirm? What do you see – parent proximity, morphology, flowering overlap, habitat? If not hybrid, then what?
Bruce Bayer – I think you are asking me to identify my left foot. What can it possibly be if it is not derived from the plants before your eyes? I forget now the name of the hybridity test based on mensuration and statistical analysis. I tried to use it on what were such obvious hybrids between herbacea and reticulata – and as you well know in H. mirabilis var sublineata, getting significance requires an inordinate sample size. So of course it is parent proximity. whereabouts of the “hybrid”. And what is difference in flowering time where there is 4-5 months between peak flowering time of each putative parent and a flowering time that can be acted on the the seasonal rainfall – and individual plants that simply do their own thing. With DNA we cannot even separate the species so how do we use that? It comes down to nitpicking and finding a way to discount the obvious. How do you confirm that all the plants before your eyes are two species when you have no definition of what the word means and cannot specify the actual criteria that are common to the respective entities? That is why it is possible to just come out with names like hammeri, joleeniae, vincentii etc. and find an innocent accepting audience? 😄 Best way of looking at the issue.
Jakub Jilemicky – Aren’t these floribunda v. dentata?
Bruce Bayer – Jakub, I would like you to detail the differences between floribunda and chloracantha. There seems to be a serious misconstrual of classification here. First you have to arrive at the species name and then at the next level down. In Haworthia I have virtually abandoned “nextleveldown” because the species level is already confounded. You cannot get a variant of one species in a population of another where the populations and species are arranged and derived as they are in Haworthia. Can someone explain it better? Prof. Cronquist once asked of his taxonomic proteges and fellow experts…”Do we know what we are doing?” By and large haworthia taxonomists do not even seem to know what taxonomists even should be doing. 😄
Jakub Jilemicky – True, this group of plants are very challenging. I’m using your books as a source of taxonomy. I have seen quite a few population from Hartenbos in the East to Riversdale in the West. And I’m lucky to grow quite a big collection from many sources of this plants, so I can compare them both in cultivation and in nature. H. floribunda v. dentata seems for me to continue into chloracantha v. denticulifera. Around Gouritz river it is almost impossible for me to distinguish them. Towards Herbetsdale the same happens with chloracantha. There are a few characteristic true chloracantha populations north of town, others are resembling floribunda. Yes, there can be selected true TL plants, which are characteristic, but all between seems for me to be one big continuity🤷♂️
Bruce Bayer – Yes Jakub, it is not just this group. I think I actually generated the name floribunda var. dentata. In retrospect it was a stupid thing to do because there is no such a taxonomic entity IF you realize that “variety” should mean a population entity and not “variant” meaning individual variation – in which case “form” would be better? My posts are doing what you describe – showing that there is 1/2/3? big continuity/ies.
Cooper Station – Jakub Jilemicky
Bruce Bayer – Thanks Jakub – I also very recently posted toothed chloracantha at Copper Siding. I say chloracantha only because there are none of the stronger floribunda features i.e. smoother surfaces, rounded leaf-tips, keel absent for large part of leaf end, and greyish green. The outward turning leaves are more floribundoid than chloracanthoid and we cannot forget parksiana. If we could just get over this hurdle of talking sensibly about these obvious mismatches and evidences of continuity, we might begin to properly look at real problems like the Hex valley plants. the Prince Albert problems, grasp what Kaboega populations tell us and many more.
120. 2020.04.01 – Stay calm, be patient, all things will pass. I found I had pictures of ‘pygmaea vincentii‘ from Welgevonden = Lodewykskraal east Albertinia. Was there with Kobus and there were countless plants then – August 2008. They were not well drawn into the soil and perhaps very vulnerable to close grazing? In this crisis it is worth considering that nature does not need us and can smile at our puny efforts, destructive ways and thoughtless actions. Also bless us for the few things we may be doing right. It is all under control.
Frantisek Vesely – Hello Bruce, all these plants were growing on the same spot?
Bruce Bayer – Hello Frantisek. It is a north facing rocky slope about 1km wide and I remember many plants all along the slope in about a 50m wide band. That was 2008. This March we could only find 3 plants very shrunken into rock cracks. It is possible that others are simply sand and debris covered in a very desiccated state and rain may bring them out.
Jakub Jilemicky – I was there in 2011 and after few hrs search we found only 5-6 plants in rock crevices…
Frantisek Vesely – Thank you. I would say there could be some interaction of var. esterhuizenii with var. turgida/suberecta. That would explain why the plants which are considered to be H. vincentii just look so odd but on the other hand they are somewhat intermediate between these two.
Bruce Bayer – Thank you Jakub – useful info
Bruce Bayer – Frantizek good observation. I wish I could express or, firstly, understand it better. It is not necessarily “interaction” but rather a question of distribution of genetic material on the basis that there is a single gene pool interconnected in some incomprehensible way and expressing itself on the basis of localised habitats. It is difficult to believe that at one time there were a great deal more populations and more gene exchange between them than there is now. Perhaps that was the case. The nearest “turgida” to vincentii is “rodinii” but we have the problem that here in the east the total gene pool is reducing from a mirabilis/retusoid (incl turgida) to retusoid (incl turgida)/ pygmaeoid (incl retusa/emelyaoid). Vincentii would represent the fade out of mirabilis via splendens and esterhuizenii. Dekenahii and fusca the fade out of retusa and fade in of pygmaea. It is something that needs exploration both literally and figuratively in the mind.
121. 2020.04.02 – This is a population from east of Herbertsdale. Here they grow in kaolinite and are buried up to their leaftips. Very much like those decaying white ‘pressure burst’ common in the Riversdale area and of course westwards. In the first picture of three clones, the one on the left is a plant from S Herbertsdale. How can you now name this population? There is something very wrong in the way we use and apply names? What do we mean by the word “species”. To say this population 3km E Herbertsdale has elements of parksiana and floribunda implies that they truly are different species. How would we know? ♦
122. 2020.04.07 – Like with the present pandemic we are coming to something difficult to comprehend. I went to Botlierskop, Rooiheuwel farm, north of Klein Brakrivier hoping to see a slender long-leaved form of Haworthia chloracantha (a JDV record). Instead I came across this population of rather stubby plants. Keep this population in view – two close populations to follow and then an issue of proximities!
Also on a rock slab about 70 meters away. Is this still chloracantha or is it parksiana?
Frantisek Vesely – We saw the similar situation – on slope there was chloracantha (+ hybrids), on flat a few dozens meters away there was parksiana and its hybrids with chloracantha – not difficult to recognize what happened there as they were so close and mixed up but on one spot most of them were chloracantha-like and the other spot were more parksiana-like plants.
600M away there is a population of uniform parksiana. But how can we say hybrids for the previous? How long have these populations been in existence? Is transfer of pollen a problem and over what distance? There seem to me to be a host of unanswerable questions in general about hybridization as an explanation for variability. It is linked to the more general question of co-occurrence – of mixing or non-mixing of species. Ring clines? Also something we should really try to document – i.e. the proximity of the various species to one another. Add to that the occurrence of hybrids.
123. 2020.04.09 – This population, east of Riversdale, is an aspect that really calls for attention – proximity of kinds (species) and hybridization. At this locality there is H. retusa and H. floribunda in contiguous/adjunct/closely neighbouring populations. Among the floribundas there is a single putative hybrid? How often does this happen and why are kinds (species) so seldom growing together? When is an entire population comprised of hybrids? What comes in between? Let me just add that I see very little in the taxonomic literature dealing with the problems of variability that exist in many genera other than Haworthia. It is simply obscured.
I am quite aware that my comments are often cryptic and often deliberately so. As a society we are very misled and never more so than in respect of the present pandemic. I do not see how we can put that aside and think that we can at the same time depend on our collective intelligence to solve and understand anything else e.g. taxonomy of Haworthia. It is a conscious creation and that consciousness has to be lifted to a level that we see things for what they really are. Hybridity is at the root of species definition and as such a critical step to recognising species. At the same time geographical location is also critical. To to think anything can be explained while there is a paranoia about revealing it, is wishful thinking.
2020.04.12 – There is a problem with classification where priority creates difficulty. The name Haworthia retusa has nomenclatural priority over the name turgida, when in actual fact the turgida form is the main one and retusa is a variant of that. It is much easier to then understand, describe and explain the idea that retusa becomes mutica in the west, retusa and mirabilis become pygmaea in the east, while turgida and mirabilis become emelyae in the Karoo. That might be where I should now leave things. There are a hoard of complexities that just get lost in the needs and activities of the majority who have no need of much more than a lot of desiderata. ♦
Haworthia chlorocantha is a name I use to cover a whole hoard of populations of plants that occur in the Southern Cape south of the Langeberg mountain range between George and Albertinia. Of course, as is the despairing fact in Haworthia, the plants in the different populations or even within populations, often do not look the same at all. It is only from a distance and from an overview of all the available (note ”available” – to me?) evidence, that I concluded that there are two sets of plants in the area. One is the group that I refer to as the Retusoids. It comprises H. retusa (=H. turgida), H. mirabilis, H. pygmaea, H. mutica and H. emelyae), although technically only H. pygmaea and “H. turgida” are in the designated area. The other group is the floribundoids that are H. floribunda,H. chlorocantha, H. parksiana, and H. variegata. Quite rightly and understandably, opinions about this as regards species names, differ humungously. I can only explain what my experience and information suggest. I lump the names like this because there are undeniable problems about regarding them as made up of different as fully and clearly different systems we like to name as species. Furthermore I must add that the problem is compounded when one is faced with the reality that for example H. floribunda, interacts in the field with the retusoids as well as with its fellow floribundoids.
Here I would like to just show and discuss one population of Haworthia chlorocantha that I recently came across east of Albertinia in a twilight zone where its relationship to H. floribunda is questionable. I also grew plants from seed and I will show what they look like after two years. This is particularly interesting because Essie Esterhuizen has done some remarkable Haworthia exploration over the last 40 years and commented to me upon his inability also to not find any real dramatic and clear differences within the same two groups I mention. His is a totally independent observational route to my own and he might well express his thoughts quite differently and understandably even choose to do so. Unfortunately I just do not have a picture record of all the Haworthia chlorocantha populations (and of course the others too) that I have personally seen. I would so like enthusiasts to have that available to them. So this short essay is restricted to just this one single locality and disregards others.
I have not even allocated an accession number to this population north east of Albertinea along the Valsch river. There is another even more complex population still further east that Etwin Aslander found and on which I commentated on in my Haworthia Updates. My first five figures are of adult plants in a small localized population occupying about 15sq m in Renosterveld. As you can see the plant are quite variable and there is undoubtedly I floribunda similarity in the leaves. Thinking that maybe flower characters may hold some significance is just a joke as I explained in Vol 8 of Updates. This applies nearly equally to flowering time that is March for this population.
Fig. 6 is of two pots of seedlings. Repotted at one year, they separated into two size groups. This is in itself an indication of internal population difference and my personal experience of growing these Haworthias from seed is that there is a natural inclination to first pick out the more robust seedlings and work down to the small weakest, sometimes even discarding the last! Even in this case I have discarded seedlings. What did I lose? So at two years we have a pot with 4 plants in the size range of diameter spread up to 4.5cm (figs. 7 and 8) and another pot with the spread up to only 3.5cm (fig. 9 showing a plant with a 2cm leafspread. There are differences in colour and leaf spination. With active growth in shadow, the leaves are recurved. Some of the leaves show the characteristic twisted and rounded tips of H. floribunda. The smallest plant is indicative of the H. parksiana-like size that suggests the same thoughts of species similarity that come to mind when exploring other populations.
Fig. 1Fig. 2Fig. 3Fig. 4Fig. 5Fig. 6. Two year old seedlingsFig. 7. Seedling 3.5cm diam. spreadFig. 8 Seedling 4.5 dia, spreadFig. 9. 2cm diam spread
published in Haworthiad v36.1 pp 14-16, February 2022 ♦