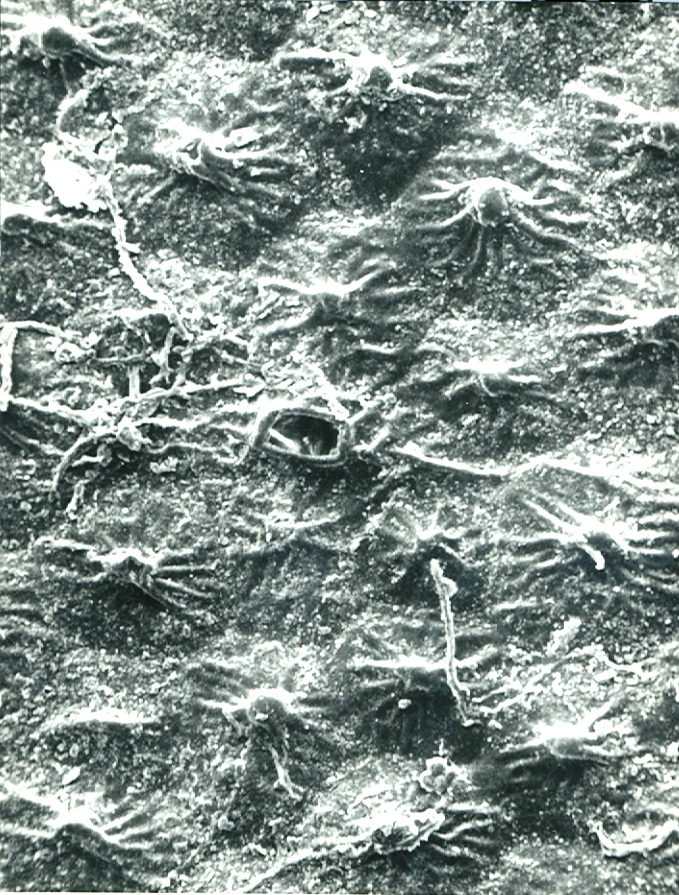
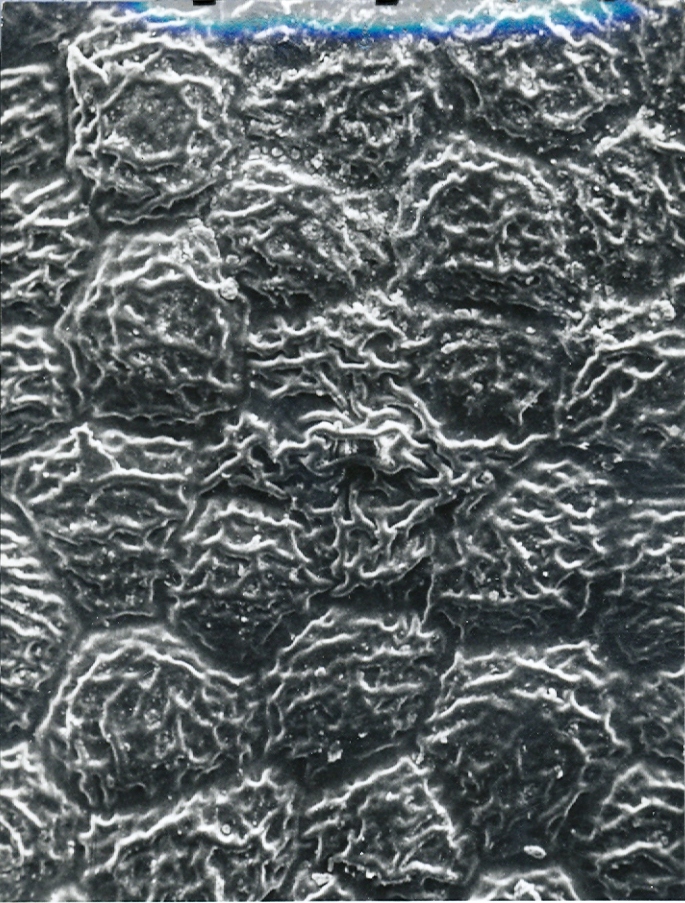
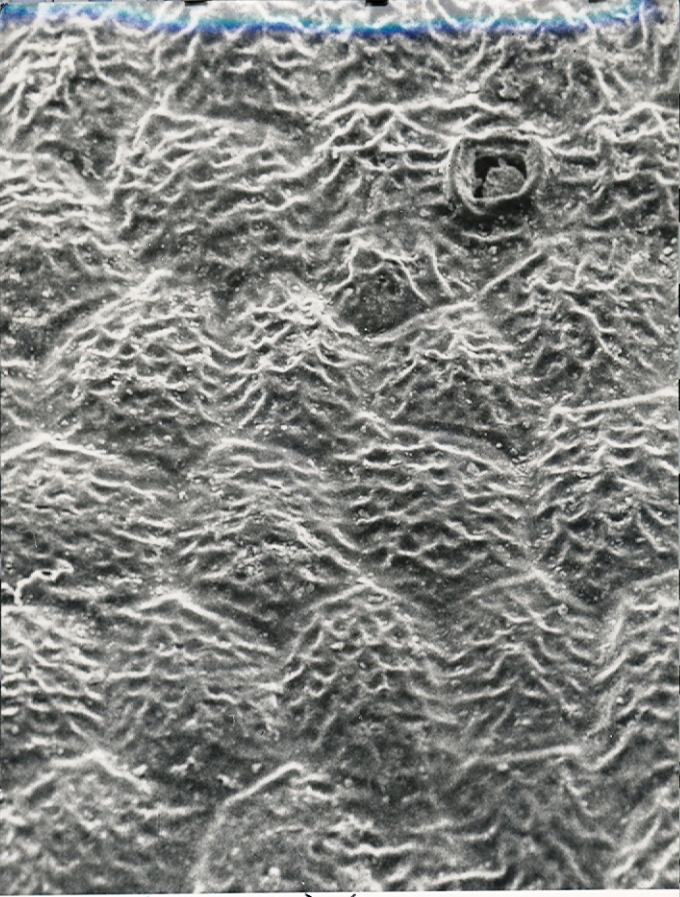
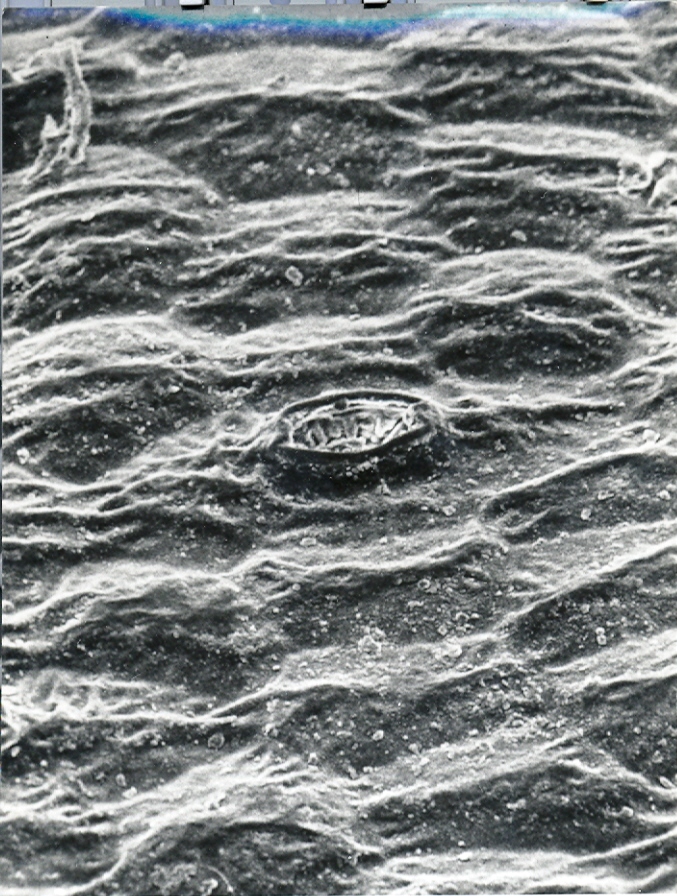
Foreward
Much of the early work on Haworthia was based on fairly limited observations in nature. These were essentially point collections (often very close to roads) from which seemingly discrete elements were recognised as species. This is akin to a survey made from a plane flying at low altitude assuming that broader patterns are being more easily identified at this illusory distance. The obvious danger of such incomplete observations is that simplistic or incorrect conclusions might be drawn. Indeed, the classification often seems easy from such selective sampling.
When Bruce Bayer started his studies of the genus Haworthia circa 1970, a new era of more methodical collection and thorough recording of the material resulted. Due to the enormity of the task, Bruce’s fieldwork focused on the Worcester/Robertson Karoo, where he built up an excellent understanding of the natural variation of and complex interaction between the recognised species of the area. His resulting model was then extrapolated to lesser-known areas further east. This work culminated in the landmark publications The New Haworthia Handbook in 1982 and Haworthia Revisited in 1999, where a classification framework for understanding the genus was proposed. This framework has been extensively field-tested by a number of people, including myself, and found to be workable.
Bruce remained doubtful about some of his solutions which were based on insufficient collection data, and some new collections which were difficult to place within the framework. During the course of the past few years he has studied further some of the problem areas, the results of which are presented in this update. New insights were gained which are used to propose an improved framework, in the form of a hypothesis, for the major Eastern Cape species complexes. As Bruce points out, this is not the only possible solution as there seem to be several alternatives, each with some reservation, when attempting to explain the observed natural variation. Some problem areas in the Eastern Cape and Little Karoo warrant further detailed investigation. Nevertheless, this work presents a valuable extension of our knowledge of Haworthia.
In his foreword to The New Haworthia Handbook in 1982, Prof. H. B. Rycroft said “the wide range of species and their endless variability keep the professional taxonomist fully occupied and sometimes confused”. This account shows how local continuities exist between adjacent species, only to be contradicted by discontinuities elsewhere between the same species. It is a pleasure for me to introduce you to this fascinating story.
Kobus Venter
Still Bay
December 2001
Foreword by the Publisher This collection of essays deals with some of the more intractable problems in the classification of Haworthia, and demonstrates just how ambiguous types can be. It also demonstrates the scale and nature of the problem in a far clearer way than Bayer’s revision. Bayer himself has said that his revision is still probably premature, and that a lot more needs to be known about Haworthia. This up-to-date account shows where some of the difficulties lie and that it is difficult to determine discontinuities in the species of the genus. Bayer pointed out that there seemed little sense in the emphasis on, and pre-occupation with, types and names, when the elements to which they are being applied are simply misunderstood or even unknown.
The process of the classification of plants is ongoing and it is correct that work should be published before the final solution is reached, as such solutions are probably unattainable. It is therefore appropriate that Haworthia Revisited should be followed by the present series of essays. It is the intention to continue publishing further collections of essays, at present four sets are envisaged. When the first four are complete, they will be bound into “collectors” editions and made available to purchasers of the original books.
Contents 1.Introduction. 2.Haworthia mucronata and its new variety. 3.Haworthia gracilis, H. cymbiformis and H. cooperi in the greater Baviaanskloof area. 4.The case of Haworthia incurvula – an up-date. 5.Haworthia cooperi and H. bolusii var. blackbeardiana. 6.The Haworthias of Kaboega. 7.Summation.
Introduction When I undertook to re-write my New Haworthia Handbook, and present it as an authoritative revision, I did so with some apprehension. However, I did so because in the intervening years my classification had proved personally satisfying. Also I saw little no evidence of the “adequatio” for the task anywhere else, either in circumstance, or willingness, or field experience, to do so. I am an aspirant mystic with none of the transport experience which should have given me the vision I sought. The hermetic and intuitive approach is all I have really been able to bring to my work other than a passion for truthfulness and understanding.
Now, three years after the initial drafting, Haworthia Revisited is published. The initial shock and ‘post-natal’ depression is over and I can look at it more dispassionately. My self-doubts had driven me back into the field and through many bouts of self-examination. There are no doubt many minor mistakes in the book, one of which is the appearance in my preface, of the word “difference” instead of the word “deference”. The latter was intended to convey the sentiment “courteous regard for others”.
After drafting the book, I tried to divorce myself from the manuscript and even from the plants themselves. However, nagging doubts and uncertainties remained with me, and these papers are the product of some intensive field “work”. I use the word in inverted commas for two reasons. Firstly the word “work” in this regard is very loosely used. Wandering around in the field, stumbling on plants here and there and anxiously uprooting them by hands sticky with excitement at “new’ things, does not constitute “work”. Secondly, it is in fact the most rewarding and stimulating way of coming into contact with a most extraordinary and marvelous natural environment. The actual work entails the making of the tangible physical record that places a specimen on the scientific map. I also wrote in my revision (p.12) “A revision.. stands and falls.. on .. the dry and uninteresting herbarium record”. The work therefore also entails deliberating about and examining the herbarium record with a good knowledge of its content. The implications that any new collection may have for that record and the way in which it is arranged, have to be considered and evaluated. That constitutes the “work”, and helps define an important aspect of the “adequatio” required for it to be proven as such.
I have not been able to reduce all the new collections I have made since 1996 to such a record. The plants are in cultivation and are being propagated by seed and offset, and are being recorded in this written record and by photographs. There is still a great deal to be done. Unlike T.L. Salter, who closed his revision of Oxalis with words which suggested that he had had enough, or like G.G. Smith who retired invalidated by the criticism he felt directed towards him, I hope I can continue to “work” on Haworthia.
This book consists of five essays discussing specific problems in the classification and identification of Haworthia. The two subjects are inseparable and unfortunately it is simply a case in which the species cannot be identified unless they are known. My life’s experience suggests to me that this is a sad truth for many things. Thus I rely very heavily on photographs to provide a comprehensive picture description of all the items discussed. The relevant photograph is numbered and this number preceded by a hyphen, usually follows the first reference to a collection number. Each essay has its own lesson and its own conclusion. A final sixth essay attempts to summarise and suggest a solution to a difficult problem.
Acknowledgement I would particularly like to thank the following who have been so supportive and helpful during the exploration and writing of these papers:
W. Schwegmann, J. D. Venter, D. M. Cumming, E. J. van Jaarsveld, P. V. Bruyns, T. Dold, D. Weekes, J. G. Marx, E. Heunis, E. Aslander, D. Clark, L. Loucka and C. Marais.
Farm owners have also been particularly helpful and I am very grateful indeed for access to such properties. In this case I am indebted to Mr and Mrs. N. Sparg, Mr. and Mrs. Killian, Mr. P. Moolman and Mr. J. Truter.
Lastly I am very indebted to Ian and Sandy Ritchie of Kaboega for their hospitality and kindness, and for sharing their own Eden and love of nature so generously with Daphne and myself. I must also record my gratitude and respect to the owners of Kaboega, and note the contribution they are making towards conservation – Mr. and Mrs. Andre Bezuidenhout.
I am grateful to Umdaus Press, and particularly to Mr. J. A. Retief, for their encouragement, support and effort to bring this work to fruition.
Acknowledgement is hereby given to Cape Nature Conservation and to the Eastern and Northern Cape Provincial Nature Conservation Authorities for the permits which were issued for the necessary but limited collecting required for this work. In the Western, Eastern and Northern Cape Provinces of the Republic of South Africa, all species of the genus Haworthia are classified as “protected flora” in terms of the Nature and Environmental Conservation Ordinance, 1974 (Ordinance 19 of 1974), as it is applicable to each of the afore-mentioned Provinces. Material for this book was gathered and collected on terms of the spirit and letter of this Ordinance. Nothing in the book should be construed as an encouragement to acquire these plants from their natural habitat or to suggest they are better located anywhere but in that situation. ♦
M B Bayer, 16 Hope Str., 8001 Cape Town.
View from the upper slopes of the Zuurberg at Wilgerfontein, with Darlington Dam just visible in the distance.
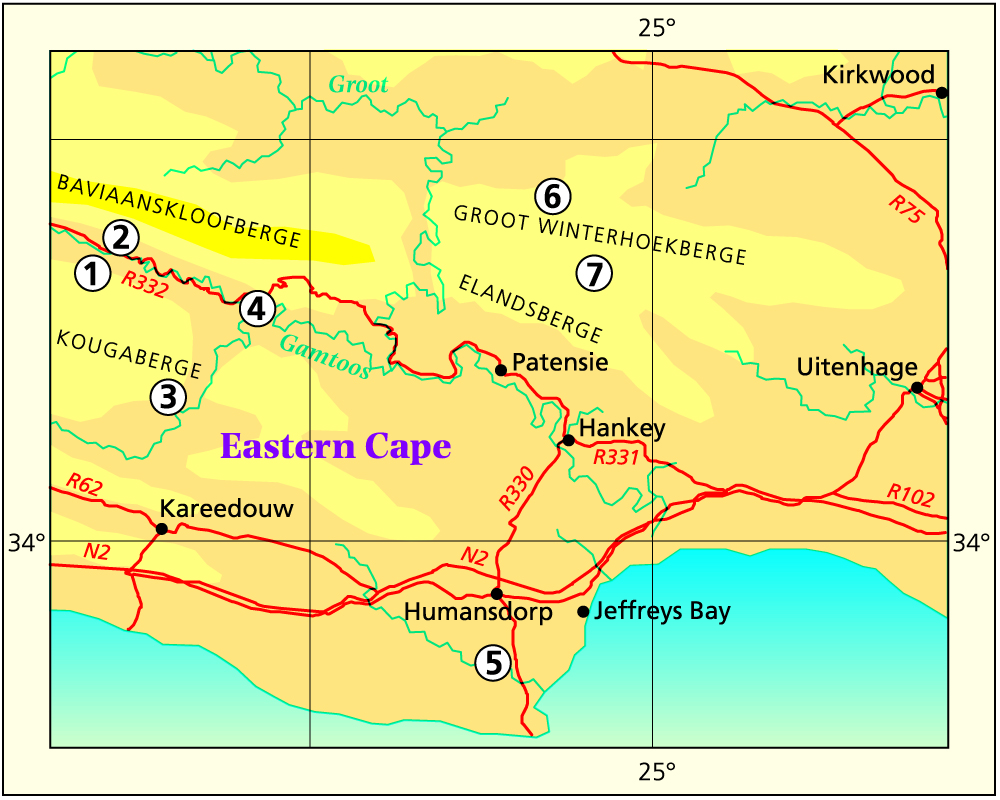
I will be disappointed if anyone had concluded I had any fixed ideas on the classification of these three species and their relationship. It has a problem which has long been on my mind. What happened recently (Nov.1998) is that I was offered the use of a time-share apartment at Jeffrey’s Bay, near the mouth of the Gamtoos River. I used this opportunity to spend six days in the field testing my hypothesis concerning the species Haworthia cymbiformis, Haworthia cooperi and Haworthia gracilis, and this is what I would like to record. Subsequent to that trip (Mar.1999) I planned and executed an excursion through the Baviaanskloof to Grahamstown and Stutterheim in March 1999, and repeated the exploration in Sept. and Oct, 1999.
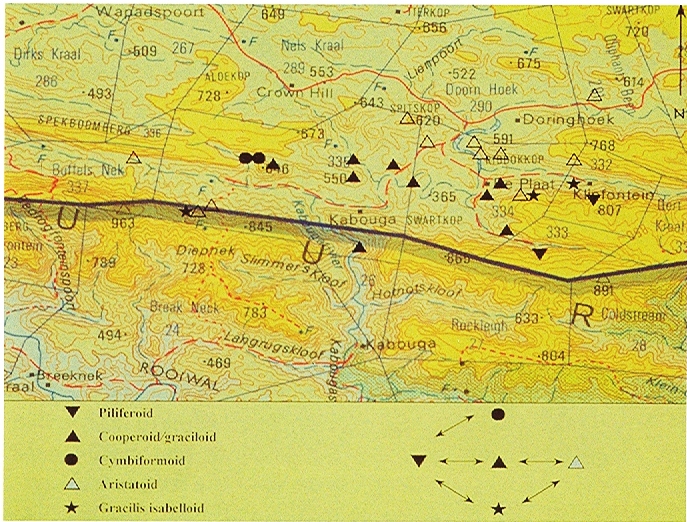
Baviaanskloof and environs: 1-Engelandkop, 2-Rooikloof, 3-Moordenaarskloof, 4-Rooihoek, 5-Ripon, 6-Ouplaas, 7-Houtkloof
First I must point out again that there is a problem with plant names in that they do not necessarily reflect a set of objects which can be seen and easily recognised. The naming and typification process is partly to blame. Typification is simply the process of ensuring that a name is based on some substantial recognisable item, preferably a specimen rather than an illustration or photograph. (A specimen is preferred because it lends itself to some other analysis e.g. a tissue sample). The type is used to secure and permanently attach a name to for reference purposes. A type specimen and hence a name need not be, and often is not, typical of the taxon which the specimen represents. This does create difficulties particularly when varieties are at issue. In Haworthia there are an inordinate number of populations of uncertain pedigree. This is not because of some obscure evolutionary process, and nor is it just because there is an absence of characters by which to characterise and circumscribe each population. It is what we should expect. Everything is in a state of change and Haworthia is simply following the same unsteady path. I have shown that in Oxalis, in which there are an embarrassment of riches where characters are concerned, that these are of surprising little help in circumscribing the species. I was interested to read this sentence in a book about chameleons, “Without collecting data many chameleons are difficult to identify.” One would expect chameleons to be less difficult to identify than plants. Darwin stated that geographic distribution is the doorway to understanding species, and this is all we have in Haworthia which I can see to make sense.
We simply use ordinary visual observation and sensibility to make judgements about similarities. The less objects we have to examine, the easier it is to assess similarities among them. The problems that then arise are no different from those that develop even with complicated and convoluted discussions and explanations that arise from sophisticated techniques and analysis of characters. In my experience it is very often the direct visual judgement which is used to test the worth of technology rather than the converse. These comments are made because readers seem to think that plant names are as centrally defined as their own unexpressed concept they have of species being groups of things with well-defined limits and readily compartmentalised. Thus they imagine that there is a solution and it is just a question of how it is found. These comments are made because several times in the literature, wild statements have been made about the use of some character or technique or other throwing great light in the darkness. It just does not happen that easily and simply. The more sophisticated the technique, the smaller the sample and the less ‘peer’ review and replication. In statistics it is also pointed out that you do not undertake sophisticated statistical analyses when the figures already show the obvious. Yet how many times do analysts not persevere with test after test until a result does appear significant – itself a matter of probability. Technique may become and end in itself and the purpose for which it is devised lost to view.
There is a nomenclatural code for plant names which regulates about everything except what the binomial stands for. What should be pointed out with some vigour, is that there is a tendency to think that species can be described from single specimens in the paradigm (this is a most useful word because it suggests a pattern, a habit and a model of a time period) of Salm Dyck, Von Poellnitz, G.G. Smith and company. The single plant (and its description) and specimen that is used to typify a name, does not constitute a circumscription of the species. These people described specimens – NOT species. Their work was thus in some measure comparable with stamp collecting. It has little to do with present time description, organisation and understanding of the living variable and changing systems which species are. The different elements in Haworthia named as ‘species’ have been determined by early writers on the basis of vegetative characters such as colour, rosette shape, leaf number and shape, translucence and spination. Nothing much has changed except the scope and extent of the problem. A difficulty with characters is stated in Vavilov’s Law of Homology which states that a set of characters in a taxon will also be present in related taxa. Thus if one finds awned and obtuse rounded leaf tips in the genus, it is possible and indeed probable, that this character set will also occur in lesser ranks. If a set of characters is present in one species, they will possibly and probably occur in related species. This article is to demonstrate the difficulty of classifying plants on this simple basis and in an era when so much more material is there to classify.
It is necessary to discard the name translucens as I have used it in my earlier books. Originally the name really is associated with what is obviously H. herbacea, and I only used it to follow a tradition of misuse. The misfortune of this is that the name gracilis then occupies the nomenclatural hierarchy, but not the same geographic position in my understanding of the species. The consequence is that an outlying variant (Hellspoort, Grahamstown) will become the species name (i.e. gracilis) and a name has to be unearthed for the core of the species in the Gamtoos valley viz. H. gracilis var. isabellae. This in my new book replaces H. translucens var. translucens. Similarly H. cooperi var. cooperi is not central to the plot as is the var. pilifera, and I unearthed the latter name in my 1999 revision for this reason, as also the name gordoniana. I opted in 1985 to use the rank ‘subspecies’ under H. translucens for reasons which should be obvious – a weaker geographical relationship of the known variants of the species associated with a broader distributional phenomenon.
This following discussion is done in terms of Darwin’s statement about the importance of geographic distribution, and the evidence I have in my mind for a wide range of species. The essential problem with the species considered in this article is that we have a set of plant populations from Grahamstown, replicated to a degree by a set of populations in the Humansdorp area. In neither area is the relationship in the set well understood, and this does not imply that they can be so understood either. I set out to deal only with recent limited observations in the Humansdorp area but like a stone thrown into a pond, the ripples soon spread.
The link between the Grahamstown and Humansdorp areas is a curious one. It can be seen in the vegetation types of Acocks, which is a broad view of species associations. Valley Bushveld occurs discontinuously from Natal through to Swellendam. Alexandra Thickets are coastal, east of Port Elizabeth. Knysna forest, west of Humansdorp. Dry fynbos dominates the immediate Humansdorp area. Karoid vegetation dominates north of the Groot Winterhoek Mountains. It is at species level that really interesting facts emerge that impact on our analysis of Haworthia. What happens in the genera Aloe, Gasteria, Astroloba etc.? I leave this as a largely unanswered question because unless the reader can respond, he or she is unlikely to appreciate that it may have implications for understanding Haworthia. Astroloba does not occur in the Humansdorp area. There is instead the curious Haworthiapungens. The Baviaanskloof is home to several endemic Gasterias as well as to Aloe pictifolia. Haworthiaviscosa is ubiquitous. H. nigra with an otherwise fairly similar distribution is not. The distribution of the genus Encephalartos (cycads) is particularly interesting because of its known antiquity and the similarity of present day species to evidence in the fossil record. Aloe must also have an ancient history as evidenced by presence of species of this genus on Madagascar – a continental drift pre-history. It would in fact be instructive for would-be authors to read what R.A. Dyer wrote about the classification of cycads, before trotting out any banal aphorism about ‘active evolution’. There is an interesting break (interval) between the cycad species, east of the Sundays and Bushmans River valleys which separate Grahamstown and Humansdorp. This break can be observed in the distributions of other genera and species and evaluated both for compliance and non-compliance. Haworthia has both conditions e.g. cymbiformis and pilifera happily cross the interval. Angustifolia and coarctata only just do so. Marumiana, and nigra do so in the inland areas. Attenuata does so. Viscosa is almost ubiquitous. Fasciata does not and neither do sordida, longiana nor xyphiophylla. No species jump the Knysna forest interval and the genus as a whole jumps the Transkei interval northward only via limifolia and koelmanniorum (+mcmurtryi). These east-west breaks are interesting because they thus indicate speciation pressures. They may be helpful in deducing possible or probable relationships in a two-directional way i.e. distribution and variation in one set of plant variants provides a tool for the examination of other sets. In the case of H. gracilis, one can argue simply that it complies with the interval, in which case the var. tenera east of Grahamstown is discrete from the Uitenhage, Baviaanskloof look-alikes. Alternatively one can hypothesise non-compliance and that continuity will be found between Grahamstown and Uitenhage. Hellspoort (the site of the var. gracilis) is somewhat supportive of this view, but their is still a wide gap. Perhaps this is only in the known distribution record.
Known collections and observations were the basis of my 1985 classification hypothesis. Many of these were either dry herbarium specimens perhaps supported by photographs of single plants grown in cultivation. This hypothesis for the species in question here was arrived at from specimens from the immediate Grahamstown area and from the lower and eastern Baviaanskloof (Humansdorp) area. For the 1999 revision I have broadened the range of varieties, and relevant to this discussion are:
Haworthia cooperi
var. cooperi
var. leightonii
var. pilifera
var. gordoniana
and included Scott’s two new species as varieties:-
H. joeyii (= var. dielsiana)
H. venusta (var. venusta)
Haworthia gracilis
var. gracilis
var. tenera
var. isabellae
Haworthia cymbiformis
var. cymbiformis
var. incurvula
var. transiens
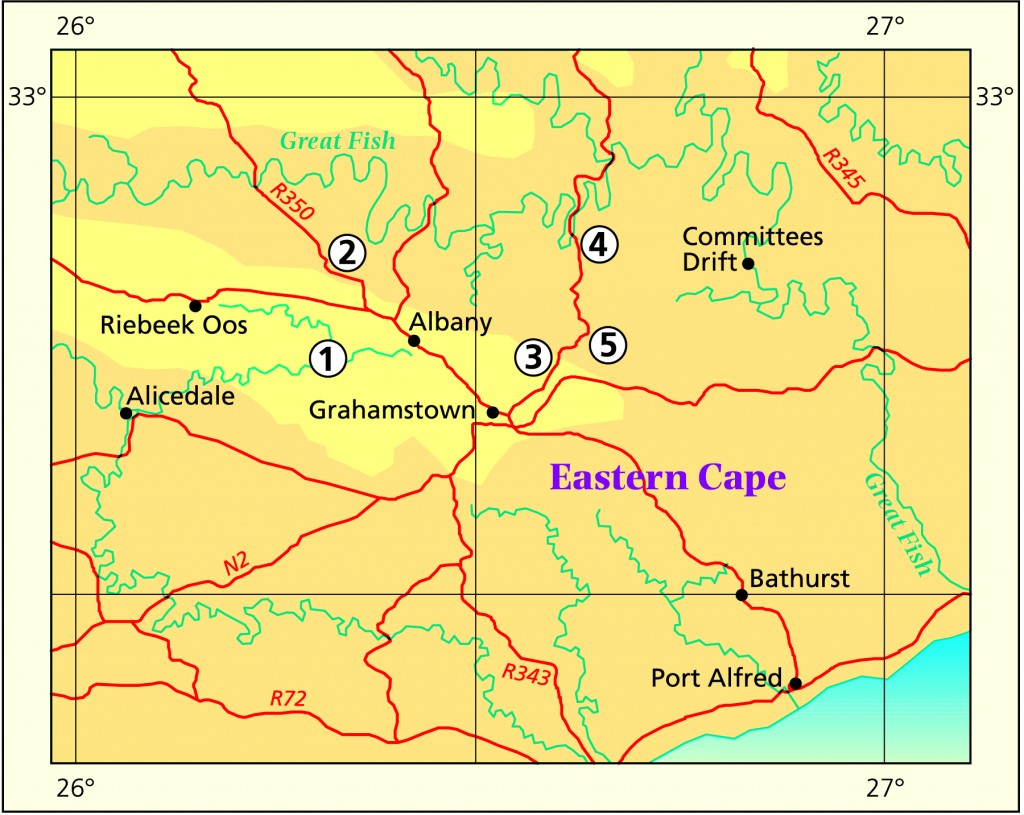
In this discussion I will largely avoid the issue of both H. bolusii var. blackbeardiana, H. cooperi var. cooperi and H. decipiens although they are closely involved. I will deal primarily with the Humansdorp (Hankey-Patensie); and to lesser degree with the immediate Grahamstown (Plutosvale) area.
I will use the term look-alikes because it is frequently used in the fynbos vegetation to refer to plant species from different genera, which are superficially so similar that close examination and full flowering material is often needed to establish their real identities. In Haworthia this detail is simply not available and thus there are specimens from different species which are geographically and thus probably genetically aeons apart, but which are visually identical.
Grahamstown (Albany) H. cymbiformis – is well represented and discrete. Yellow-green plants with spreading broader, flatter, ovate leaves. Clump forming plants on steep rocky faces. The var. incurvula – is known from many collections but only one locality (Plutosvale). It has look-alikes in the Patensie area particularly. Its relationship as a variety of cymbiformis really follows G.G. Smith who made the combination on the basis of a continuity which he claims to have seen. I recall (!) that its flowers are more reminiscent of tenera and this article may expose this as a possibility. Pale semi-opaque green, small clump-forming, with ovate, obtuse incurved leaves.
H. cooperi – is well represented by the var. pilifera. Blue-green plants tending to purplish, with incurved thicker, shorter ovate leaves, often spined. Solitary plants withdrawn into the soil. (The var. cooperi is not discussed in this appraisal although the concept presents a substantial problem on which I am preparing a manuscript).
H. gracilis – is known only from Hellspoort north-east of Grahamstown, with a second population of larger more cooperi-like plants from nearer Plutosvale (MBB6603-99, Glen Craig – a Gerhard Marx collection). Gracilis does have look-alikes in the Kirkwood area (Paardepoort, and a greater Humansdorp area). Paler grey-green plants tending to clump, with more leaves. The leaves are incurved narrow attentuate, and with or without spines. My opinion, based on my most recent collection from Hellspoort is that the var. tenera is actually the element concerned and is thus a superfluous name for gracilis. This var. tenera – is very well known from several populations north-east and east of Grahamstown and these include a glabrous population. It has look-alikes in the Uitenhage and Hankey areas of Humansdorp. Grey-green, small solitary or clump-forming, with many narrow, spined incurved leaves.
(Note:- the original description of gracilis seems to be of a moderately spined species, and von Poellnitz concept was apparently drawn from several disparate collections. The conception of the species from an illustration in Desert Plant Life, and indeed my own collection from Hellspoort in early 1970s, is of a moderately spineless plant. The recent collection of mine has forms which are relatively spineless as opposed to forms which are indeed similar to the smaller, densely spined, tenera. Thus we again have the problem where a name is neither central to the geographic element nor typical. The name tenera could be dispensed with and replaced with var. gracilis – this is evident from my discussion).
Humansdorp H. cymbiformis – represented by the typical variety in the small Baakens River valley and discrete. It may occur north-east of the Baakens River in the Rocklands area of Port Elizabeth. There is also the population at Hell’s Gate north-west of Uitenhage which I am regarding now as the var. transiens (It needs to be re-examined – hastening to add ‘by someone competent to do so’). This var. transiens – is only recently known to be very well represented in the upper Longkloof and in the Baviaanskloof. Its relationship with cymbiformis may not be as close as varietal rank, and the linking population at Hellsgate east of Uitenhage may be misunderstood. Leaves are incurved, pale-green and less opaque than in cymbiformis. Some of the variants resemble plants of the var. incurvula from Plutosvale.
H. cooperi as the var. pilifera – well represented and largely discrete. However, the typical variety with the necrosing-end area seems to have a western limit just outside Port Elizabeth and also near Uitenhage (the furthest west being at Perseverance, JDV90/40). It is then replaced by a variant which is often smaller, having the same blue-green colour darkening to purplish, but the leaf tips remain slender acuminate even in light conditions. This is what I have regarded as the var. gordoniana which in its typical form is not well represented.
H. gracilis as the var. isabellae – is well represented but apparently variable. The leaves tend to be spreading, pale greyish-green and spined. Some plants resemble gracilis from either Hellspoort (less-spined) or from Plutosvale (densely spined – tenera).
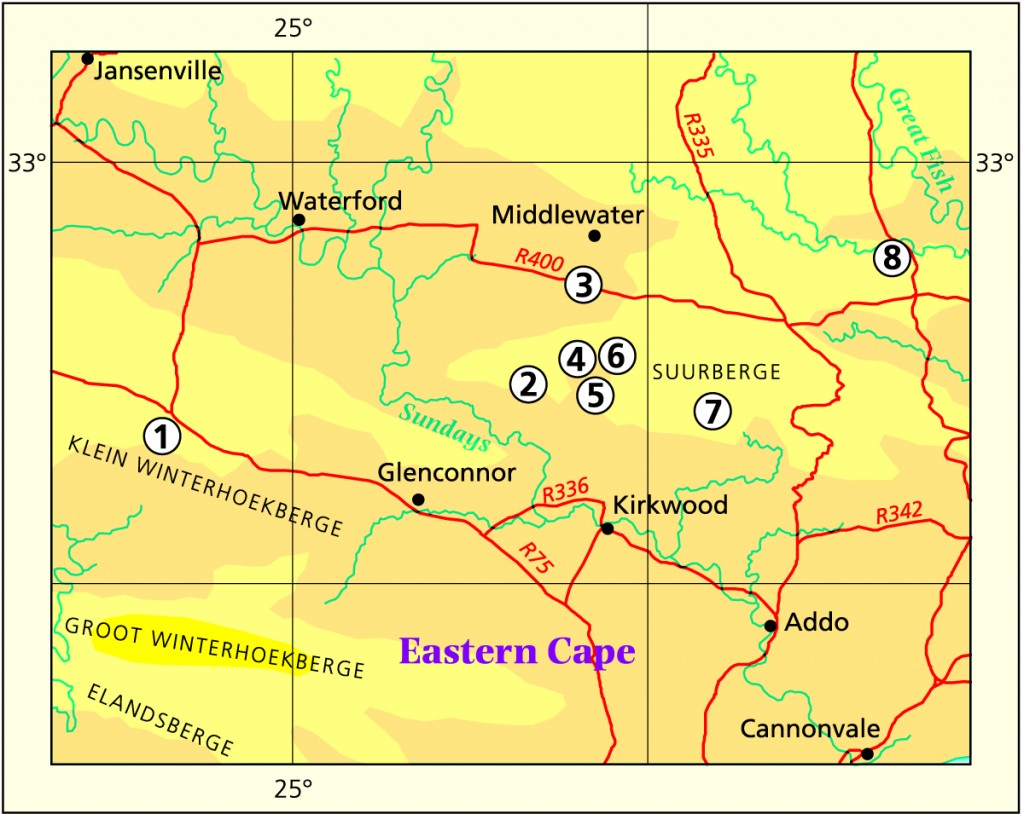
General It is known to me that in the area from Kirkwood, westward to the Little Karoo (north of the Baviaanskloof), there is an interaction between gracilis and decipiens. H. cooperi (not designating any variety) also occurs in that area. There is a problem in the Addo area, where another element (aristata – a separate weak hypothesis) may intrude. It has also been suggested, and this has always been in my own mind, that unicolor (mucronata) from the western Little Karoo may be continuous with either cymbiformis, bolusii or cooperi. It is continuous with arachnoidea, lockwoodii and with decipiens, and I am treating it as a separate issue because there are also other reasons for its exclusion here. The discussion could descend to the point where one says simply that there are not many species in the sub-genus Haworthia.
Results
Grahamstown I first looked at Hellspoort early in 1970 to collect and form an impression of H. gracilis. This was of a largish plant with leaves exceeding 3cm long, fairly spineless and weakly clustering. I also saw tenera in Plutosvale and at Hunts Drift. The former, small, densely spined and strongly clustering. At a later occasion I collected tenera again in Plutosvale as the conventional spiny element, and close by as a glabrous element of the same size. On that occasion I neither looked for nor saw incurvula or cymbiformis in the area. I revisited Hellspoort with J.D. Venter and G.D. Marx in 1996 and Venter made a re-collection again in 1998 (JDV89/42-117). My 1996 collection (MBB6614-118) reminded me forcefully of Plutosvale tenera, the plants were smaller than my previous collection and also spinier. Venter’s 1998 collection (all slightly different points within Hellspoort) was of the conventional gracilis that I visualise from the Von Poellnitz illustration, and from my early collection. These were quite large plants. During my visit of March 1999, I observed and recorded systematically from the bottom of Plutosvale, to the top (Roff’s Rock). On the north side of the kloof from the Cotswold farmhouse to the middle of the kloof which is Plutosvale, there was tenera both large clusters and relatively solitary plants. I could not locate or identify the site where I had collected the glabrous form before. The area has been degraded by the grazing of goats and it is possible the plants are now gone. From there we explored the south side of Plutosvale as the road climbed out of the kloof. Roffs Rock is very accessible and this no doubt accounts for the large number of herbarium records for incurvula. We found if from virtually the top of Plutosvale, to the lower and eastern point across from where we had observed tenera at our western-most search point. It is possible that the collection G.D. Marx in JDV93/73-93 was from further to the south-west than our starting point at Roffs Rock, at the upper point of Plutosvale. Nevertheless we found the same very pointed leaved plants of that collection, tending to be clump-forming. As we moved eastward, so the plants tended have leaves which were less pointed and more incurved. The colour was yellowish-green rather than the greyer or bluer-green of the Baviaanskloof incurvula look-alikes. The leaves seemed to lack the more swollen upper and end margins that may characterise cymbiformis.
My impression from this repeat visit to the Grahamstown area, is that the elements gracilis and tenera may be better considered as the same taxon. This may lay the basis for a better understanding, or be more compatible, with the range of look-alikes in the west around Humansdorp and Uitenhage.
The incurvula problem is now largely resolved and although it may still be necessary to find other links between incurvula and cymbiformis which Smith claimed and which is not supported by herbarium evidence. To the contrary, the collections of the gracilis-gordoniana plants from Glen Craig and Roffs Rock (viz. -99, -89, -93)), as well as the interplay of species in question in the Baviaanskloof, suggest that incurvula may equally be interpreted as a variant of H. gracilis or even H. cooperi. There is a further possibility based on new observations in respect of H. bolusii var. blackbeardiana. It may be possible to restructure the classification to relate the var. bolusii with H. semiviva; and to consider the element blackbeardiana as a variety of cooperi, (as nomenclaturally correct but geographically weak) as the species, with pilifera and gordoniana as principle variants. Nomenclatural considerations in ‘the strict terms of the code’, might confuse the issue, but I see no reason why reasoned and logical argument could not be used to dismiss names which are confusing because of their geographical connotation.
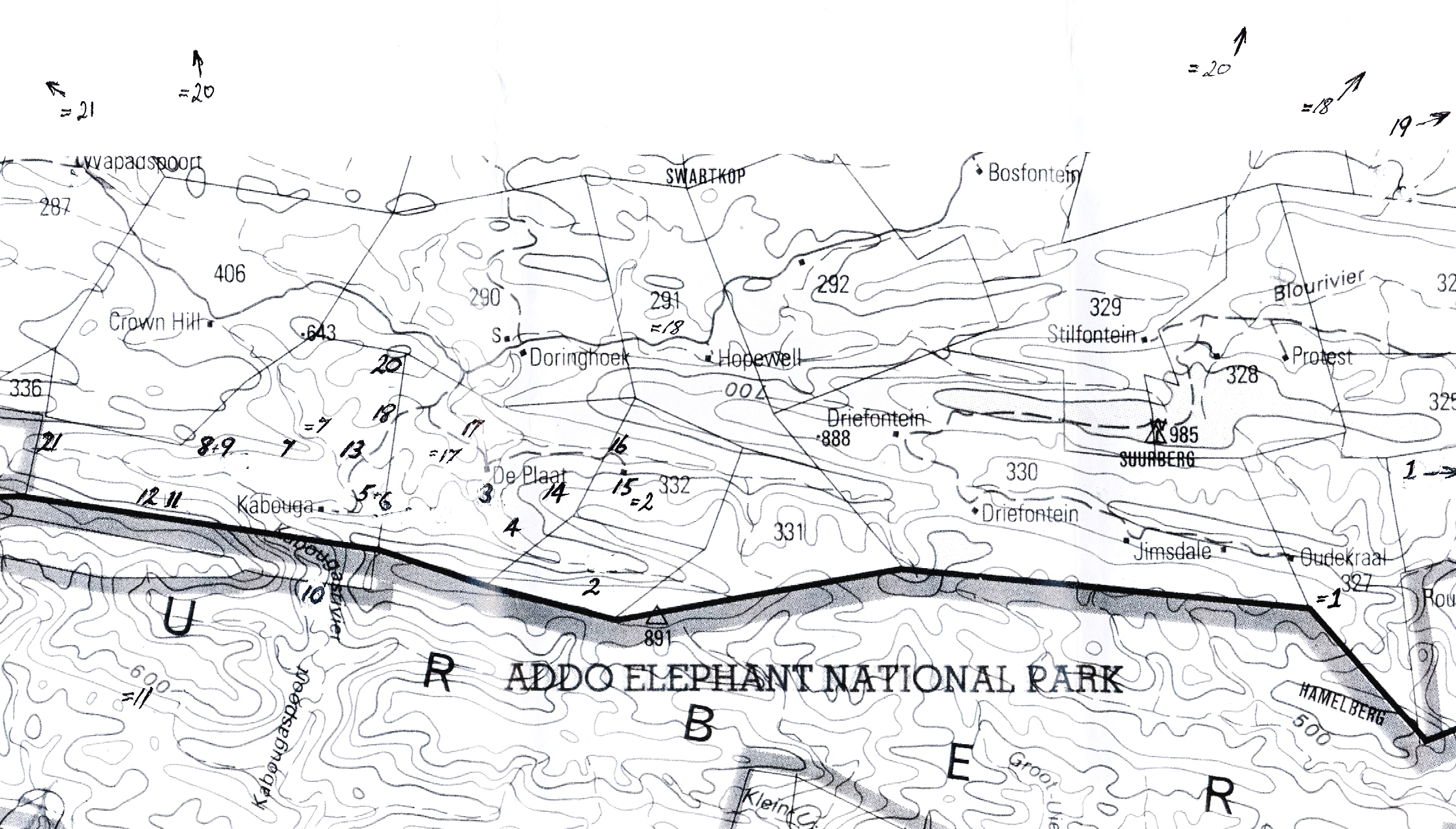
Humansdorp I looked at populations at Jeffrey’s Bay, Zuurbron between Humansdorp and Hankey, two at Draaihoek to the south-west of Patensie, three near Andrieskraal, one further west in the Groot River Poort, and three still further west in the Baviaanskloof, one at Patensie, one at Houtkloof in the Elandriver valley, one at Hankey, one north of Hankey, two south-west of Hankey, and one at the Gamtoos Bridge. Some of these populations were known to me before, and many of them are already represented in the Herbarium. I will also refer to other collections (mostly multiple plants) in my discussion which I have either not seen in the field at all, or in the more distant past. These are collections by credible field botanists and naturalists such as P.V. Bruyns, G.D. Marx, E.J. van Jaarsveld, T. Dold and D. Clarke. This thus reviews some 60 populations, but does exclude some significant ones in the Kleinwinterhoek mountains, where other considerations apply.
MBB6792-20 Jeffrey’s Bay. Fynbos. Very short vegetation on level sandstone outcrop. Early bud stage. Small single plants to 20mm diam. Leaves incurved at tips, no marginal spines and bluish-green, darkening to purplish hue (3) – gordoniana-picturata.
MBB6933-21 E. Humansdorp. Grassy fynbos. Rocky sandstone knoll. Small single plants, bluish-green with coarser spination than MBB6793 (2) – gordoniana.
MBB6553-22 and MBB6793-22 Zuurbron. Valley Bushveld. Scrub vegetation on level alluvial stream bank with loose sandstone rocks. Flowering and seeding. Small single plants to 30mm diam. Leaves thicker, leaves incurved at tips, many small closely spaced marginal spines, bluish- green (2) – gordoniana (possibly close to type locality).
MBB6784-23 SW Patensie. Valley bushveld, west sloping conglomerate. Seeding. Small single plants to 30mm diam. Leaves incurved at tips, forms with and without marginal spines, and blue-green (1). Smaller plants of this order were collected by Tony Dold at Drinkwaterskloof in the Baviaanskloof near Geelhoutboskloof – gordoniana.
MBB6786-24 SW Patensie. Valley bushveld, steeper south facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaves incurved at tips, mostly plants with marginal spines, blue-green (2) – gordoniana-isabellae.
MBB6827-27 Rooihoek. East of Geelhoutboskloof. Steep east facing slope. Plants single, greenish, very small – picturata-transiens.
EvJ14680-28 Vetmaakvlakte, S Rooihoek – isabellae (as for a clone of -41, resembling H. aristata).
MBB6789-29 Groot River Poort (Komdomo). Valley bushveld, very steep west-facing shale cliff. Flowering. Small clustering plants to 30mm diam. Leaves with spreading tips, slightly obtuse, no marginal spines, pale-green (5) – transiens-picturata.
MBB6830-30 E Komdomo. Riverine margins. Large clustering plants on steep soil-covered rock- gracilis.
J.N.Reddi in JDV93/86-31 2km NW Andrieskraal, is a collection which may be the same as -30 – gracilis.
EvJ15927-32.1 N. Groot River Poort. Steep south-facing cliffs. Clustering or single. Fairly similar to -29 – transiens-isabellae, the clone 32.2 to decipiens var. minor?
MBB6790-33 Andrieskraal. Valley bushveld, very steep south-facing conglomerate. Flowering and seeding. Small clustering plants to 30mm diam. Leaf tips moderately incurved, slightly obtuse, no marginal spines, pale yellow green (4+Y) – picturata.
MBB6791-34 Andrieskraal. Valley bushveld, steep east-facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaf tips incurved, slightly obtuse, no marginal spines. Pale yellow green (4+y) – picturata.
MBB6930-35.1 E. Andrieskraal, Nuwelande. Steep south facing conglomerate. Small plants down to 16mm diam. As above (4+y). – picturata. JDV90/55-35.2 from nearby westward, is also small plant with very pronounced windows.
MBB6928-36 Nuwelande. Upper steep conglomerate cliff. Plants nearly spineless and the leaves more slender and terete. Greenish (4). These are similar to JDV94/115-37 from the Paul Sauer Dam wall and also from the site for Gasteria glomerata where it was collected by E.van Jaarsveld (EvJ11076 in JDV90/118-38.1&2). A plant from that collection (-38.3) also resembles the Ripon plants (MBB6932-39) which have broader more deltoid, and more spinose leaves – isabellae.
J.G. Marx298 in JDV94/30-40, also Paul Sauer Dam, include both spined and spineless plants which also appear to be (4) – isabellae.
MBB6799-41.1 also (-9&-10) NW Patensie. Valley bushveld. South facing conglomerate slope. Seeding. More robust single plants to 45mm diam. Leaf tips incurved, with and without marginal spines. Pale green and blue-green (1-3) – gracilis-isabellae. One clone in this quite variable collection, strongly resembled a large specimen of H. aristata (-125,-142,-143).
MBB6801-42 SW Hankey. Early flowering. Valley bushveld. Vertical conglomerate. Small clustering plants to 20mm diam. Leaf tips incurved, with marginal spines. Pale blue-green (2-3) – isabellae.
MBB6802-43 SW Hankey. Valley bushveld. Steep east facing conglomerate. Early flowering. Plants mostly single to 30mm diam. Leaf tips tending to spread, with marginal spines. Pale blue-green (2-3) – isabellae.
PVB7128-44 Holriver, far S Patensie. Steep north-facing cliffs. Similar to above – isabellae.
D.Clark1050-45 Kouenek, Geelhoutboskloof. No detail, similar to above. – isabellae.
MBB6773-46 N Kareedouw. Fynbos. Conglomerate. South facing. Flowering late Dec. Plants single small, leaves incurved, very spiny and spines on leaf surfaces too. Blue-green (2) – isabellae.
MBB6771-47 Moordenaarskloof (N. Kareedouw). Steep south facing slope. Plants in small clusters or single. Relatively spineless. Leaves incurving. Green (4) – picturata.
MBB6805-48 NE Hankey. Valley bushveld. Gradual west facing slope. Seeding. Plants single to 50mm diam., leaf tips incurved, with and without marginal spines. Blue-green and pale-green and (1-4) – gracilis-isabellae.
MBB6804-49 N Hankey. Valley bushveld. Dense vegetation on alluvial stream bank. Seeding. Plants single to 50mm diam., leaf tips incurved, without marginal spines. Pale-green and blue-green (1-4) – gracilis-isabellae.
JDV97/3-50 E Hankey. Valley Bushveld. Plants similar to above – (1-2) – gracilis-isabellae.
MBB6808-51.1 Gamtoos Bridge. Valley bushveld. Steep west facing conglomerate. Seeding. Clustering plants to 50mm diam. Leaf tips spreading, with marginal spines. Blue-green (2) – isabellae. There are very similar plants at Longmore, east of Loerie (MBB6555-51.2).
MBB6798-52 Houtkloof (Upper Elandsriver valley). Fynbos. Rocky valley in sandstones. Neither flower nor seed (flowered in cultivation in Jan.). Plants single to 35mm diam. Leaf tips incurved, very spiny. Blue-green (2) – isabellae.
C. Marais in JDV96/95-53 Forest Glade and JDV92/136-54 Oaklands, both Elandsriver, are similar but more coarsely spined and paler green (4) – isabellae.
MBB1404a in JDV86/13-55 is an old collection of a tenera-like plant from the lower Elandriver valley (Groendal Dam) – cf. tenera.
D.M. Cumming6831 is from nearby and is more typically isabellae-like).
PVB7040 in JDV97/8-57 and PVB7944 in JDV97/7-58 Ouplaas (E. Cockscomb) are very similar to the above from the Elandsriver, but the plants are more robust, with much wider and more obtuse leaves. Particularly interesting because similar plants also occur in the Kleinwinterhoek eg JDV93/41-59 N Campherpoort, JDV87/180-60 S Campherpoort, JDV91/136-61 N Wolvefontein, JDV92/140-62.1 and JDV94/45-62.2 both Wolven (all decipiens var. minor). There is also the collection from the northern Groot River Poort (EvJ15927 – 32.2) linking these, and perhaps also the Cockscomb and Elandsriver plants (ie. isabellae) with transiens.
JDV97/6-63 W Braamrivier. Dry Fynbos. Steep W. facing cliff. Clump-forming, short obtuse incurving leaves. Pale green (5) – transiens. EvJ15342-64 Dieprivier, NE Kareedouw; EvJ17548-65 Horee, Saptou, SE Uniondale; PVB7077 in JDV97/5-66.1 Oshoek, E of Uniondale; MBB6729-66.2 S Uniondale, and Reddi in JDV93/54-67 Kabeljouwsriver, near Jeffrey’s Bay (a doubtful collection), are also this element – transiens.
The latter is similar to -29 Groot River Poort (transiens-picturata), ie. small clustering plants to 30mm diam. Leaves with spreading tips, obtuse, no or very few marginal spines, pale-green. I could not locate this population and it would prove that gracilis, transiens and cooperi co-exist. E. Aslander1247-68 made a collection from the place reported by Reddi, and these plants are unquestionably gordoniana-like.
PVB7093 in JDV97/1-69 Skrikrivier, north of -63 is similar to that collection, but leaves more elongate acuminate and some lightly spined. Flowered late Jan. Very reminiscent of gracilis var. gracilis and particularly a brighter green variant from Paardepoort (MBB6600-70, var. viridis in the new revision) in the Kleinwinterhoek mountains to the north-east. PVB5402 in JDV97/20-71.1 Palmietrivier and MBB6589-71.2 Dorschfontein, both E. Steytlerville, are similar to this latter collection but better related to decipiens var. minor. A problem arises here in trying to limit the scope of the paper because there are similar collections from north of Glenconnor (JDV91/17-72.1), and south of Lake Mentz (JDV91/116-72.2, which grows with gordoniana JDV91/115-73).
MBB6810-74 (also -11&-12) Joubertina. Fynbos. South facing, rocky sandstone slope. Neither flower nor seed. Plants single to 50mm diam. Leaf tips incurved, without marginal spines. Blue-green (1) – gordoniana.
MBB6811 & JDV90/80-75 (also -13) Uniondale Pass. Renosterveld. Rocky defile, steep sandstone. Early bud stage. Plants single to 25mm diam. Leaf tips incurving, with and without marginal spines. Blue-green (1) – gordoniana. PVB7079-76 Saptou, (Upper Longkloof), PVB7062-77 Redcliffe (NE Willowmore), JDV91/80-78 Engelandsekloof (Baviaanskloof), JDV94/95-79 Nuwekloof (W. Baviaanskloof), are all in this class. A collection G. Marx194 in JDV91/81-80 from Apieskloof (Baviaanskloof) is unusually pale green but otherwise also seems to compare with these gordoniana-like plants).
In completing this review of collections I would like to mention five which seem to touch on this issue, but which are even more relevant to the H. cooperi var. cooperi and H. bolusii var. blackbeardiana issue. These are G. Marx in JDV91/14-145 De Plaat (NW Kirkwood), PVB5002 in JDV92/33-147 Kaboega Gorge (Suurberg), JDV96/89-116 Gladhurst (S Adelaide), J.G. Marx in JDV96/4-98 and MBB6603-99 NE Grahamstown (Glen Craig), and JDV93/73-93 upper Plutosvale. These are all plants with elongate acuminate leaves about as wide as thick, and almost or completely without spines. They could be confused with H. cooperi var. leightonii, or with clones of H. bolusii var. blackbeardiana – and of course with H. gracilis var. gracilis.
Discussion In trying to circumscribe each collection I have been aware of my deficient descriptive skills. Can one rationalise such similar things one observes in writing in a way and which can lead others to identifications? I have elsewhere pointed out major weaknesses in the capacity of persons to compare what is written with what is seen, and also to compare plants and illustrations. Apart from the problem of variation in the populations, there is a problem of different habitat conditions (light, soil, temperature and moisture) which will cause phenotypic differences. It should be possible to define these if the plants could all be grown under the same conditions. However, local variation and sample size also becomes a problem. Colour must be critical. There are four main classes in this set of species under discussion – the opaque yellow-green of cymbiformis, the darker blue-green of cooperi which can develop reddish veins and a purplish colour under stress, the mid pale (or light) grey-green of translucens and the subdued pale green of transiens. Under stress the latter becomes very pale. In the descriptions above I have categorised colour on a scale of 1-5 from blue-green through to pale green, and added Y to indicate a more opaque yellow colour. In my book I recognise a variety (i.e. MBB6791-34 picturata from Andrieskraal) in which the transition from opacity to translucence is abrupt. This collecting trip does not strongly support such a geographic entity although their may be populations in that area which do. Certainly in the var. transiens, some plants are also very conspicuously patterned. Flowering time in early summer for all these collections appears to be very much the same. Curiously the Jeffrey’s Bay collection was still at bud stage when most of the other collections were seeding already. When I visited this population six weeks earlier in the previous year, the plants were already in flower.
The off-setting, clustering character is also worth commenting on. As in the case of H. turgida and H. retusa (remembering also an expressed contention of mine that the latter can be regarded as a variant of the former), clustering is usually associated with very steep rocky situations as opposed to plants on level sites being solitary. In this particular study the slopes varied from level through moderate to vertical. Clustering was most pronounced on vertical sites and at Andrieskraal where vertical and sloping sites were nearly adjacent, the plants were clustering on the vertical and solitary on the sloping. Clustering may be accompanied by the capacity of the plant stems to elongate. It is significant that populations around Hankey and Patensie which seemed to be var. gracilis, were invariably on moderate sloping or level ground (NOTE. I have previously always identified these collections as gordoniana), and using the name gracilis now is derived from the product of the papers in the compendium. My later papers suggest that transferring these elements to H. cooperi is worth consideration).
The above collections confirm my observation that Haworthia in the Baviaanskloof are very difficult to classify and circumscribe. It does seem on the basis of collections by Bruyns, Van Jaarsveld, and Venter that transiens is a very substantial element to the west, which may not have a strong connection with cymbiformis. Gordoniana is apparently a much stronger element than I had previously supposed and I am comfortable with the decision to now recognise it as a valid variety of cooperi. The above collections in the Longkloof, and some other recorded collections, seem to show convincingly that it effectively co-occurs with transiens. In the area just west of Patensie, three collections described above seem to indicate that gordoniana intergrades directly with transiens and this transition gives rise to plants which resemble incurvula and which I identify as H. gracilis var. picturata. Similarly south-west of Patensie gordoniana (-24) seems to alter to isabellae. South west of Hankey this latter variety (-42) resembles tenera. The resemblance to tenera is also apparent in the Groendal area of Uitenhage. The Elandsriver collection (-52), and a similar one by Clifton Marais in the same valley (-53), appear to be intermediate in appearance between tenera and isabellae. Isabellae as a separate entity is also represented by the Gamtoos Bridge collection (-51), and an old collection by W.R. Branch from the Krom River estuary at Ripon (-39).
A major difficulty is the Jeffrey’s Bay population (-20). These small solitary plants seem, by virtue of their growth habit and the near co-occurrence with isabellae (perhaps transiens too, if Reddi’s collection -31, was confirmed), to be the local equivalent of cooperi var. pilifera. However, if they had been observed at Grahamstown, they would be taken to be gracilis. In cultivation they have come to bear a stronger resemblance to the -47 which is closer to picturata than to gordoniana. The Zuurbron population (-22) passes for the equivalent of pilifera with some difficulty, and this does transpose to isabellae via Draaihoek, SW of Patensie (-24). The gracilis-like plants around Hankey (-48,-49,-50) and Patensie (-41) too, often present an isabellae facies. Thus we are faced with the possibility of a strange inversion. The upper Longkloof has cooperi as the var. gordoniana, but in the lower Longkloof similar plants may be derived from gracilis. A similar problem is presented by the similarity of gordoniana-like plants to H. bolusii var. blackbeardiana at several different places in the wider Eastern Cape.
The collection -46 is particularly interesting (I am indebted to Ernst van Jaarsveld for its discovery). This is a very small isabellae which has the same colour and (spininess) hairiness of the leaf surfaces as has Scott’s venusta. This may be very significant in providing supporting for an hypothesis that prior to inundation of the continental shelf (12-18000yrs ago) there was less of a vegetation interval east and west of the Sundays and Bushmans Rivers. I have suggested that this is reflected in the relationship and distribution of H. coarctata and H. reinwardtii, and will suggest elsewhere that H. fasciata may also be evidence of such a situation. However, this is very speculative. The surface spination is also evident in some collections from north and south-west of Jansenville (-129,-130,-131,-132 and -133 which I have identified with either decipiens vars. minor or pringlei), although the spination is sparse.
Conclusions These specific observations suggest that my classification hypothesis (the classification as presented in my two handbooks and in my recent revision) is a sound communication medium for the two areas in question, without implying thereby that it cannot be improved. What is problematic is that in the greater Baviaanskloof area, the three elements never seem to occur together. I repeat what I have written in respect of H. cooperi and H. bolusii var. blackbeardiana, and in my handbooks and revision. It is also stated indirectly in the preamble to this paper. “The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain good will.” In this particular article, the key question would have been:-
“Do three elements we can identify as separate living systems representing the species cooperi, cymbiformis and gracilis grow in the same close geographic proximity?”
Evidence available to me and in the herbarium record, is that they do not. There are a few known instances where two of these species are in such proximity – but not all three. What is curious is that it is rare for any two species of the subgenus Haworthia to be in immediate proximity to the extent that they share any specific habitat.
Observations seem to confirm my opinion that there is a connection between the three species in their occurrence east and west of the Sundays and Bushmans Rivers. What does concern me is the relation between cymbiformis and transiens, and the implications these observations have for incurvula, gracilis and cymbiformis around Grahamstown. The set of plants around Grahamstown favours cooperi, cymbiformis and tenera, while the mirror set of the Humansdorp area, favours gordoniana, transiens and isabellae. The relationship of incurvula needs to be explored. In the eventual integration of all these collections, sight should not be lost of the complex situation around Fort Beaufort, where the same set of species and variants seems to be present. Nor should it be forgotten that in the Kleinwinterhoek mountains H. decipiens becomes involved. The situation north of the Groot River is complex and there are substantial records from the Perdepoort and from Willowmore and Steytlerville which indicate a close interaction between decipiens and gracilis as well as gordoniana.
There are a good many other collections from the upper Baviaanskloof, but mostly these are represented only by herbarium records and I would like to see this material in its living state.
A final thrust of this article is to emphasise that Haworthia is not going to be understood by botanists or enthusiasts sitting on some other continent. This is particularly if they depend on the convolutions of the Code of Nomenclature, literature and limited collections for their information and the opinions they feel compelled to express. Less so if they do not and cannot, read and write the English language sufficiently well to communicate properly. Still less so if they have no knowledge and feel of South African geography and vegetation, and are operating in an outdated paradigm where specimens represent entire species. Explanations which only account for a few plants in cultivation are not going to be very satisfactory. The fact must be faced that classification is difficult and more information makes it more difficult. This is not a problem peculiar to Haworthia. It only seems so because the genus has received so much attention. The question now is:
“How is this information recorded and presented in a way that can be accessed, understood and appreciated?”
My conclusion is that we now need to resort to computerisation and digital imagery, recognising that dealing with single plants is not the same as dealing with populations. It is not easy to assess populations objectively as the sample sizes become unmanageable. We should progressively assess and re-assess populations and build up good herbarium documentation. In this way a proper revisionary process is set in motion, which limits Micky Mouse decisions about hypotheses, classification and nomenclature. ♦
1 a. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.1 b. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.2 a. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 b. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 c. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.3 a. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 b. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 c. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 d. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.4 a. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.4 b. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.5 a. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.5 b. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.6 a. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.6 b. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.7. MBB6825 H. cymbiformis var. transiens to H. gracilis var. picturata, lower Geelhoutboskloof.8. MBB6827 H. gracilis var. picturata to H. cymbiformis var. transiens, east of Geelhoutboskloof.9. EvJ14680 H. gracilis var. isabellae, Vetmaakvlakte, south of Rooihoek.10 a. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 b. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 c. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 d. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 e. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 f. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).11 a. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 b. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 c. MBB6830 H. gracilis var. gracilis, east of Komdomo.12. JDV93/86 H. gracilis var. gracilis, 2km northwest of Andrieskraal.13 a. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 b. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 c. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 d. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 e. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 f. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.14 a. MBB6790 H. gracilis var. picturata, Andreskraal.14 b. MBB6790 H. gracilis var. picturata, Andreskraal.14 c. MBB6790 H. gracilis var. picturata, Andreskraal.15 a. MBB6791 H. gracilis var. picturata, Andrieskraal.15 b. MBB6791 H. gracilis var. picturata, Andrieskraal.15 c. MBB6791 H. gracilis var. picturata, Andrieskraal.15 d. MBB6791 H. gracilis var. picturata, Andrieskraal.15 e. MBB6791 H. gracilis var. picturata, Andrieskraal.16. 1 a. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 b. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 c. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 d. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 2 JDV90/55 H. gracilis var. picturata, also east of Andrieskraal.17 a. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 b. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 c. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.18. JDV94/115 H. gracilis var. isabellae, Paul Sauer Dam Wall.19.1. EvJ13809 H. gracilis var. isabellae, upper Paul Sauer Dam.19.2. EvJ11076 in JDV90/118 H. gracilis var. isabellae (smooth), upper Paul Sauer Dam.19.3. EvJ11076 in JDV90/118 H. gracilis var. isabellae (= MBB6932, Ripon), upper Paul Sauer Dam.20. MBB6932 H. gracilis var. isabellae, Ripon, Krom River.21 a. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 a. MBB6799 H. gracilis var. isabellae, northwest of Patensie.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 c. MBB6799 H. gracilis var. isabellae, northwest of Patensie.22.2. MBB6799 H. gracilis var. gracilis to H. gracilis var. isabellae, northwest of Patensie – a clone resembling H. aristata.23 a. MBB6801 H. gracilis var. isabellae, southwest of Hankey.23 b. MBB6801 H. gracilis var. isabellae, southwest of Hankey.24 a. MBB6802 h. gracilis var. isabellae, southwest of Hankey.24 b. MBB6802 h. gracilis var. isabellae, southwest of Hankey.25 a. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.25 b. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.26. D.Clark1050 H. gracilis var. isabellae, Kouenek, Geelhoutboskloof.27 a. MBB6773 H. gracilis var. isabellae, north of Kareedouw.27 b. MBB6773 H. gracilis var. isabellae, north of Kareedouw.28 a. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 b. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 c. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 d. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.29 a. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 b. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 c. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.30 a. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 b. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 c. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.31 a. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.31 b. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.32.1 a. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.1 b. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.2 a. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.32.2 b. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.33 a. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.33 b. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.34 a. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.34 b. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.35 a. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 b. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 c. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.36 a. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.36 b. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.37. D.M.Cumming6831 H. gracilils var. isabellae, south of Groendal Dam.38 a. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 b. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 c. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.39. PVB7043 in JDV97/7 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.40 a. JDV93/41 H. decipiens var. minor, north of Campherpoort.40 b. JDV93/41 H. decipiens var. minor, north of Campherpoort.41. JDV87/180 H. decipiens var. minor, south of Campherpoort.42 a. JDV91/136 H. decipiens var. minor, north of Wolvefontein.42 b. JDV91/136 H. decipiens var. minor, north of Wolvefontein.43.1. JDV92/140 H. decipiens var. minor, Wolven near Wolvefontein.43.2. JDV94/45 H. decipiens var. minor, Wolven near Wolvefontein.44 a. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 b. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 c. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.45 a. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 b. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 c. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.46. EvJ15748 H. cymbiformis var. transiens, Horee southeast of Uniondale.47.1 a. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 b. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 c. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 d. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 e. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.2. MBB6729 H. cymbiformis var. transiens, De Vlugt, Prince Alfred’s Pass, south of Uniondale.48. J.N.Reddi in JDV93/54 H. cymbiformis var. transiens, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.50 b. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].50 c. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].51 a. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 b. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 c. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.52.1 a. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.1 b. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.2 a. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 b. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 c. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.53.1. JDV91/17 H. gracilis var. viridis, north of Glen Connor.53.2. JDV91/116 H. graciis var. viridis, south of Lake Mentz.54 a. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.54 b. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.55 a. MBB6810 H. cooperi var. gordoniana, Joubertina.55 b. MBB6810 H. cooperi var. gordoniana, Joubertina.56 a. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.56 b. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.57. PVB7079 H. cooperi var. gordoniana, Saptou upper Longkloof.58. PVB7062 H. cooperi var. gordoniana, Redcliffe northeast of Willowmore.59. JDV91/80 H. cooperi var. gordoniana, Engelandkop, Baviaanskloof.60 a. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.60 b. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.61 a. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.61 b. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.
A new variety, Haworthiamucronatavar.rooibergensis is described in Haworthiad 13:5 (1999). It raises many questions, and leaves as many unanswered.
What do the authors, Esterhuizen and Battista, mean by Haworthiamucronata? A very curious picture emerges. There are two sources which must be considered recent and hopefully authoritative. These are C.L. Scott, and M.B. Bayer. Bayer does not use the name mucronata and therefore Scott must be presumed to be the authority followed. But Scott regards H. habdomadis as a separate species whereas Esterhuizen and Battista treat it as a variety of H. mucronata. The two authors also say that their new variety, where it occurs east of Vanwyksdorp (and presumably also their mention of its occurrence south of Calitzdorp), is on the southern boundary of the H. mucronata complex. Who do they follow? If they are using Scott (or Von Poellnitz for that matter) they seem to have mistaken the given distribution. Scott’s distribution map gives five points for H. mucronata which must by south of Vanwyksdorp, and one of these is even west of Mossel Bay. There are also two points north and east of Queenstown.
Curiously, Von Poellnitz is considered to have taken the two Haworth species viz. altilinea and mucronata as synonymous. Scott uses the first for what I called H. cooperi (var. pilifera in my revision), where he also recognises H. pilifera with practically the same distribution. He places his two species altilinea and pilifera in different sections. He does cite the direct synonym H. altilinea var. mucronata = H. mucronata var. mucronata, but Von Poellnitz does not. Scott cites Feddes Repert. 45:169(1938) as his source. I have a handwritten translation in which H. altilinea var. typica is distinguished from H. altilinea var. mucronata. The distributions for these two elements is extraordinary viz. typica from Stockenstroom, Seymour, Redhouse, Grahamstown, Cathcart, Zwartkops, Prince Albert, Hankey, Longmore (Loerie), and mucronata from Adelaide, Albany, (Van)Wyksdorp and Wolwefontein (Jansenville). Effectively H altilinea var. typica was left in taxonomic limbo, and I am not sure if my new revision has followed all these devious ramifications. I can only hope it does. Triebner and Von Poellnitz transfer the varieties of altilinea to mucronata in Feddes Repert. 49:23, 1940 but seem to omit both var. mucronata and var. altilinea.
I can now only try to explain the incongruity of the situation in terms of what I know. Firstly, I know that both Von Poellnitz and Scott were confusing H. mucronata and H. bolusii (at least as far as the variety blackbeardiana is concerned). This confusion is itself indicative of the validity of a new variety which “at fist sight looked very much like H. bolusii“! Secondly I know that Scott also recognised H. aristata, H. mcclarenii and was also noting plants from west of Barrydale (mistaking them for my H. serrata), which I currently think belong with his concept of H. mucronata. This is where I think H. habdomadis also belongs. One has to know that the variety H. bijliana var. joubertii (which I believe came from immediately north of Ladismith per communication with A.J. Joubert) comes into contention. This is in relation to the comparison Esterhuizen and Battista make with H. altilinea var. brevisetata. That variety did in fact originate at Vanwyksdorp, but Von Poellnitz also cites H. altilinea var. mucronata from there. He later cites the var. brevisetata from also Albany and Port Elizabeth. This is in Feddes Repertorium 49:29 (1940) where he also cites H. altilinea var. inermis from Halesowen, Queenstown, Ladismith and Hankey. A better comparison could have been made with this concept of inermis or with joubertii of Von Poellnitz, rather than with brevisetata. I personally think either choice is irrelevant given either:-
1. the various localities cited by Von Poellnitz, or
2. the variation that occurs, for one example, just west if Ladismith where the element habdomadis ‘moves’ from sandstone to shale.
In describing the variety in the detail that they do, and in stating that it “at first sight looked very much like H. bolusii“, the authors invite this comment. I personally very much doubt if any descriptive detail has been given and could be given, which could be used by anyone, to identify a plant of this variety with any certainty from variants of the species H. bolusii, H. cooperi, H. gracilis or H. decipiens, let alone from the many variants of H. mucronata itself. These species are themselves interlinked and I have stated that they are only real as systems in their geographical context.
A curious aside is that, I cannot find the transfer of the name var. brevisetata from H. altilinea to H. mucronata. Von Poellnitz made all the other changes in Feddes Repert.49:29 (1940), and Scott cites this reference for the change to H. mucronata var. brevisetata. I cannot find this in my photocopy of Von Poellnitz’ paper, and Breuer (in his outstanding compilation of the literature) also loses the epithet, as a “Nomen dubium” under H. altilinea. I think he also has lost the var. typica. My statement in my Handbook (1983).. “Von Poellnitz in the same year reduced this variety to synonymy with H. mucronata“, is fallacious and I cannot explain it.
There is a further problem with the citation by Esterhuizen and Battista which reads “North of Uniondale plants are found which may also be H. mucronata.” I consider that this is the same citation for H. decipiens which they give earlier in the paper.. “this species..is growing about 20km north west of Uniondale”. They also relate this to their new variety as “showing (some of the same) characteristics”. The problem is that this is all a vast understatement of its true nature. In my newest book which constitutes a formal revision, I have re-described the situation of the species arachnoidea and mucronata. Here I wholly alter the order I had in my previous works and try to accommodate my respected friend Col. Scott as well. The Uniondale references above are not resolved. There is already a massive interaction or interplay between H. arachnoidea and H. mucronata. The same is evident for H. decipiens and H. mucronata north of the Kleinswartberg, between Calitzdorp and Oudtshoorn, and for H. mucronata and H. lockwoodii south of the Rooiberg Pass and east of the Anysberg. The Uniondale area is grossly under-explored but is known to be touched by the interplay of H. cymbiformis, H. gracilis, H. decipiens, H. bolusii and H. cooperi. This is all fairly evident in Hayashi’s speculative analysis of Haworthia in the same edition of Haworthiad, and it is also evident from the localities cited by von Poellnitz. Unfortunately that work, although its intrinsic message is excellent, also has severe limitations. I have my doubts as to whether Esterhuizen and Battista are aware of any of the above ramifications relating to their new taxon.
Since I first drafted my revision in 1996, I have recognised the reversal in the fortunes predicted by Bruyns in Kew Magazine (4:148, 1987) where he wrote of Haworthia taxonomy being set back 40-50 years. Consequently I have felt the need to do further exploring and recording to expand my observations and possibly verify the predictiveness of even my latest work. I can here confirm that there is a better solution in at least one area and that relates to H. bolusii var. blackbeardiana. I am preparing a separate extensive explanation of that solution. The illustrations for this present article also contribute to that new solution.
The photographs submitted, show the variation which extends continuously from Aliwal North (H. bolusii var. blackbeardiana, JDV97/62-1), southwards to Queenstown (JDV92/22-2), to Stutterheim (JDV94/60-3), across westwards to Somerset East (H. decipiens var. pringlei, MBB6561-4,-108, H. aristata MBB6852-5,-126), to Jansenville and Willowmore (H. decipiens var. pringlei MBB6582-6,-131, MBB6583-7,-132, H. bolusii var. bolusii! MBB6856-8). Then the illustrations show the continuity from H. cooperi var. gordoniana (it could be H. gracilis var. isabellae) from Hankey and Patensie (MBB6799-9,-10,-41) through Joubertina (MBB6810-11,-12,-74) going westward to Uniondale (JDV90/80-13,-75). From here I switch to H. decipiens var. cyanea in the Beaufort West and Merweville districts (JDV90/105-14, MBB6885-15), H. decipiens morphing to H. mucronata west of Prince Albert (H. decipiens var. decipiens! JDV93/71-16,-17), and finally just two of many H. mucronata variants (JDV86/84-18, JDV91/74-19). ♦
62. JDV97/62 H. bolusii var. blackbeardiana, southeast of Aliwal North.63. JDV97/22 H. bolusii var. blackbeardiana, Fincham’s Neck, Queenstown.64. JDV94/60 H. bolusii var. blackbeardiana, Stonefell, northeast of Stutterheim.65. MBB6561 H. decipiens var. pringlei, Patryshoogte, east of Somerset East.66. MBB6852 H. aristata, Stonefountain, south of Somerset East.67. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.68. MBB6583 H. decipiens var. pringlei, Waaipoort, southeast of Mt. Stewart.69. MBB6856 H. bolusii var. bolusii (!), northeast of Fullarton.70. MBB6799 H. cooperi var. gordoniana, northwest of Patensie.71. MBB6799 H. cooperi var. gordoniana, northwest of Patensie.72. MBB6810 H. cooperi var. gordoniana, north of Joubertina.73. MBB6810 H. cooperi var. gordoniana, north of Joubertina.74. JDV90/80 H. cooperi var. gordoniana, south of Uniondale.75. JDV90/105 H. decipiens var. cyanea, east of Merweville.76. MBB6885 H. decipiens var. cyanea, Trakaskuilen, south of Beaufort West.77. JDV93/71 H. decipiens, Gamkadam, west of Prince Albert.78. JDV93/71 H. decipiens, an H. mucronata var. inconfluens look-alke, also Gamkadam.79. JDV86/84 H. mucronata var. inconfluens, east of Calitzdorp.80. JDV91/74 H. mucronata var. inconfluens, northwest of Seweweekspoort, Ladismith.
This article was first published inAloe 36:34, (1999) and is now rewritten in part to accommodate some new collections and a conclusion modified accordingly.
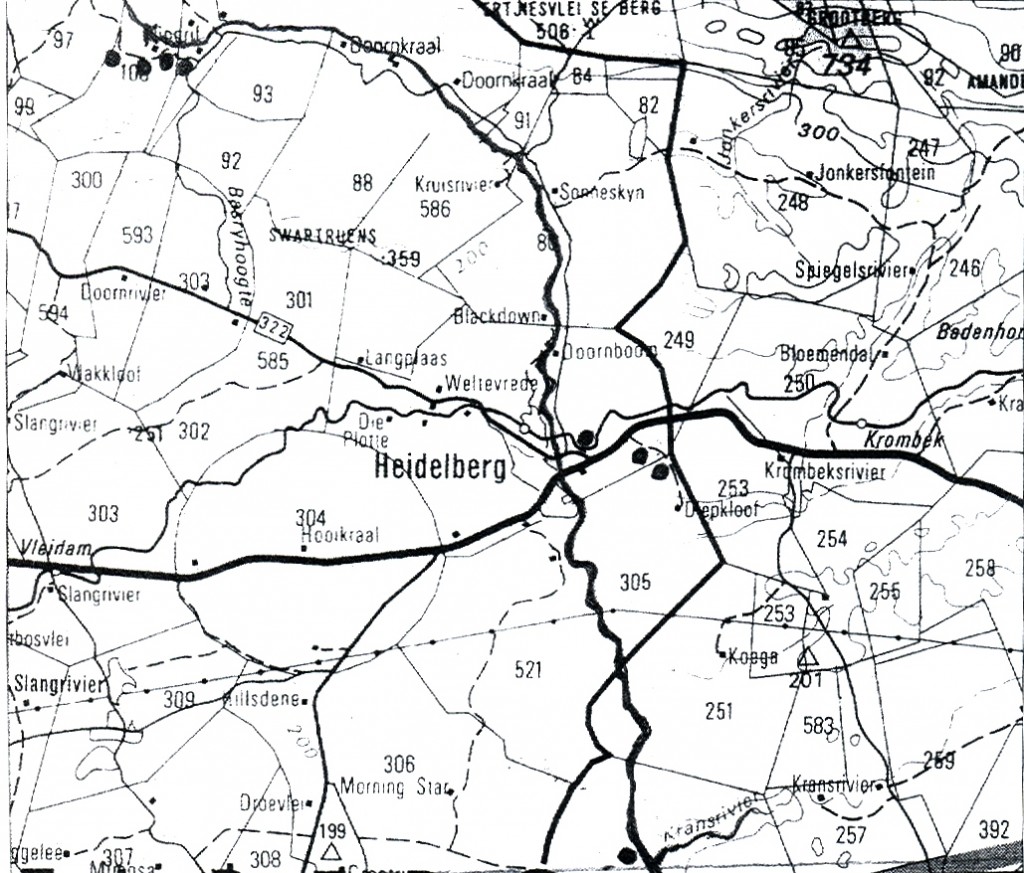
The problem von Poelnitz described Haworthia incurvula (Feddes Repertorium 31:85, 1932), from specimens ostensibly from Grahamstown, sent to him by Mrs. E. Ferguson. The important elements of the description are “leaves about 40, barely 20mm long, up to 12mm wide, broad ovate-oblong, without teeth, hairs or spines, on the face almost flat and inconspicuously turgid towards the apex, on the back convex and towards the apex with a rounded keel, very rarely inconspicuously double keeled, pale green and traversed by greenish longitudinal lines, which are somewhat anastomosing, some of which reach the pellucid tip, which is by no means abruptly pellucid”. von Poellnitz (Fed.Repert. 41:203, 1937) acknowledges that this is the plant illustrated in Flowering Plants of Southern Africa 9:t356 (1929), which is there identified there as H. cymbiformis var. planifolia (Haw.) Baker. Again (Fed. Repert.44:233, 1938) he acknowledges the illustration given by J.R. Brown (Desert Plant Life 8:45, 1936). Here he gives “… the exact locality is Pluto’s Vale in Albany district, between rocks and bushes.” citing R.A. Dyer as the source (Botanical Survey of South Africa, Memoir 17:98, 1937). von Poellnitz adds here that.. “in vigorous cultivated plants the leaves are less incurved or almost not at all”. He was uncertain what its affinities were and placed it in the section Muticae. However, it should be noted that von Poellnitz placed the synonymous H. planifolia (Obtusatae) and H. cymbiformis (Planifoliae) in different sections too. This was despite his confession and obvious in his writing, that he did not know the difference between them.
Smith (SA Journal of Botany 16:1, 1950) transferred the species H. incurvula to the section Obtusatae on the contention that it was continuous with H. cymbiformis which he said was also in the close Pluto’s Vale area. I wrote in my Haworthia Handbook (p124, 1976) “Smith’s field book untypically bears no reference to other collections of this kind in that specific area, and the writer’s limited field experience does not bear this out either. There is however, an apparent if obscure relationship with H. translucens subsp. tenera which bears further investigation. Forms very similar if not identical to H. cymbiformis var. incurvula also occur at Andrieskraal north of Humansdorp, which is a long way to the west.” In my New Haworthia Handbook I could add nothing new, and neither could I add anything substantial in an article I wrote for Cactus & Co 2:11 (1998). Col. C.L.Scott maintained H. incurvula as discrete, writing that “it has been gathered at a number of localities along Pluto’s Vale near Grahamstown”. His riposte is “H. incurvula is a distinct species, and is readily separated from H. cymbiformis“. It must be noted that he does not separate H. cymbiformis and H. planifolia either.
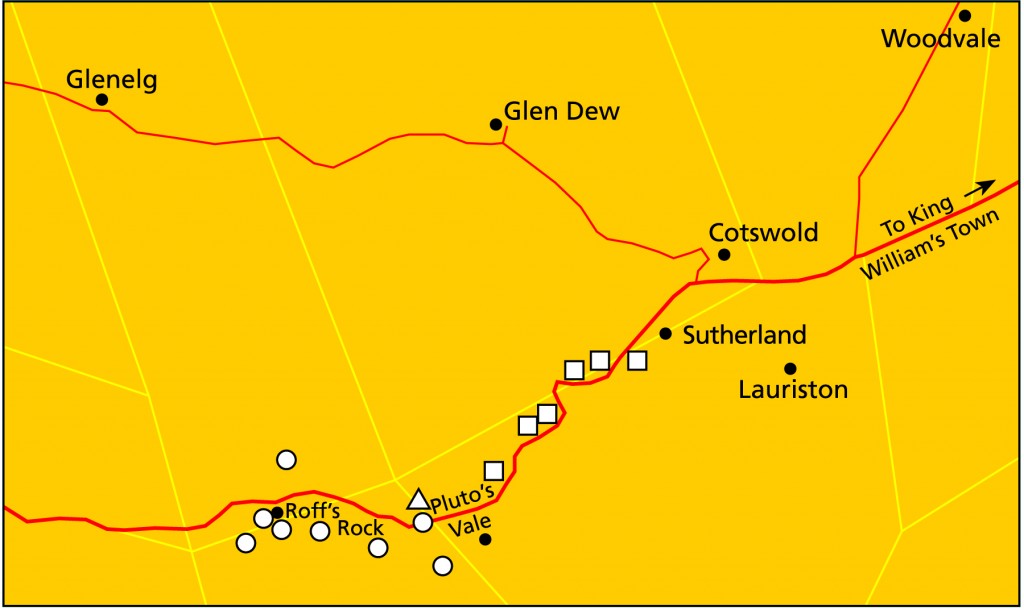
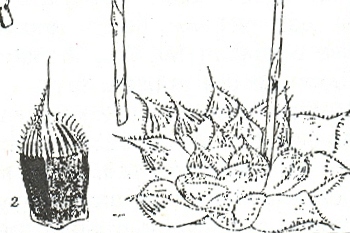
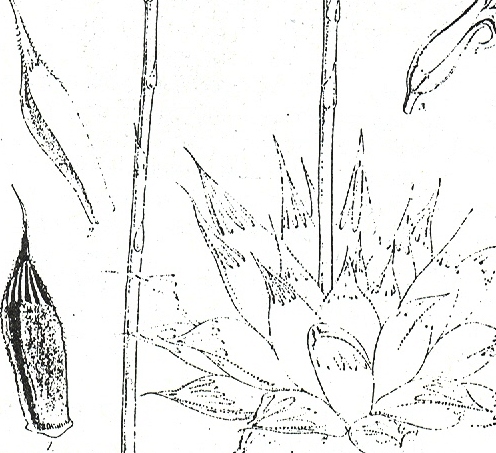
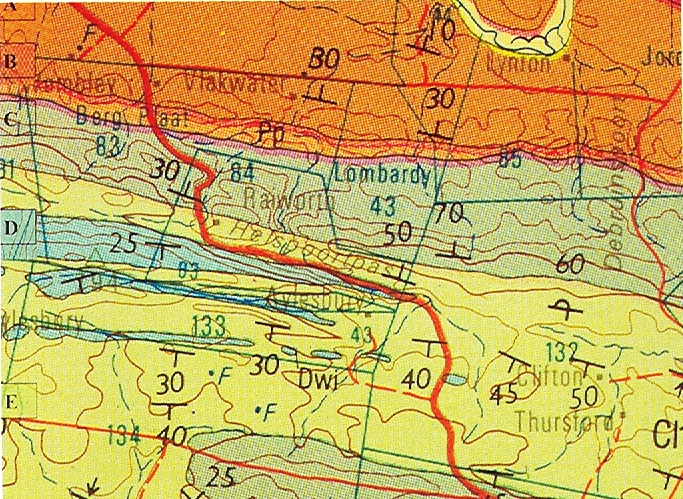
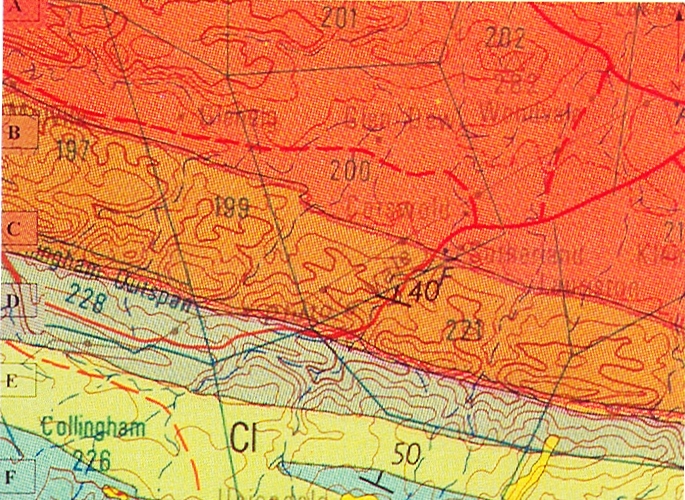
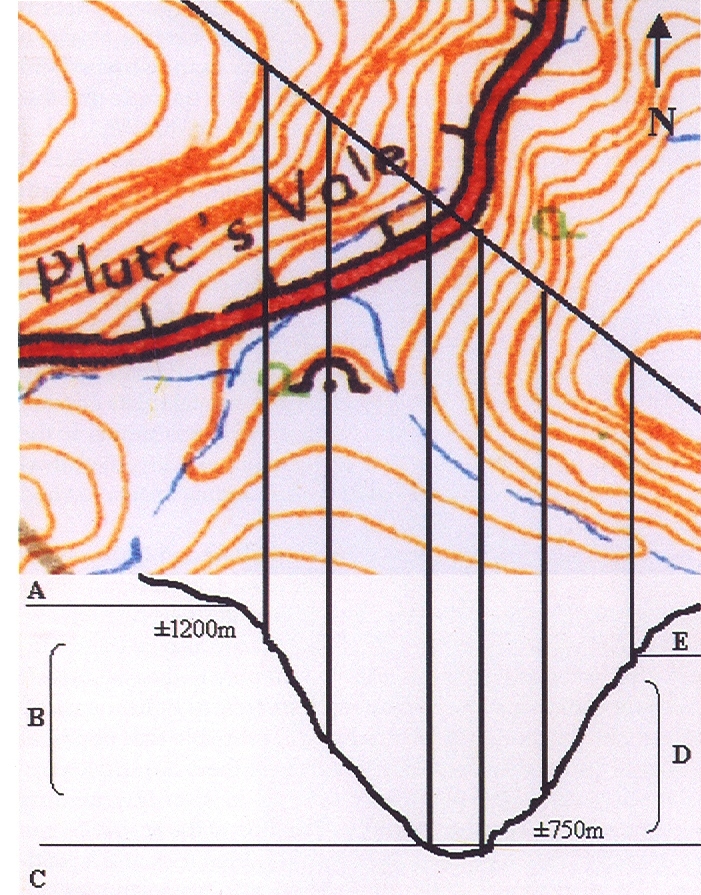
Pluto’s Vale area. The squares represent the sites for “tenera”, the circles for “incurvula” and the triangle marks the site of glabrous “tenera”. “tenera” has also been collected at Glenelg to the north, and H. cymbiformis at Bosky Dell just off the map to the northwest. The origin of H. cymbiformis var. lepida is still further north and west.
Pluto’s Vale is also the origin of H. tenera von Poellnitz (Fed.Repert. 31:86, 1932). The collector was Miss Blackbeard. The site is incorrectly given as “Plinters Vale” and apparently, by courtesy Albany Museum staff, the origin is “Cloete’s Vale”. This is a smaller more proliferating species with more, slender and less densely spined leaves. However, I made a collection from Pluto’s Vale myself circa 1972. In fact two collections (KG47/72a &b), within 75 meters of one another. In one the plants were spinescent, and in the other glabrous. The plants were too small, with the leaves too spinescent and too slender to be incurvula, but the potential for connection was there and that is the predictive stance that was taken. In my New Haworthia Handbook (1983), the point was also made of look-alikes in the Humansdorp area for both tenera and incurvula.
At this point I must state that in my revision (1999) I depart from the name translucens and replace it with gracilis. My thinking subsequent to this change and prior to current writing is, that the name gracilis (from Hellspoort, NW Grahamstown) is in fact a variant of tenera (or vice versa). Scott’s solution is not greatly different except that he applies the name tenera more widely to include elements in the Humansdorp area; and he limits the name gracilis (his use of the name translucens has no bearing on this discussion) to its point of origin.
The hypothesis I now pursue is my own, which is that the element incurvula does not constitute a species and that it can be explained in the context of two other systems (species) as I first suggested in 1976. These are H. gracilis (H. translucens sensu Bayer 1976, 1983: and including the variety H. gracilis var. tenera) and H. cymbiformis (hence H. cymbiformis var. incurvula). Lest anyone again question what I mean by species as do Crouch et al (Aloe 36:8, 1999) in their allusion to “very seldom have some of these authors suggested what might constitute a species”; let me spell out what is implicit in all my classification. Thus – “A species is a system of living organisms which are genetically and morphologically continuous in space and time”. Unfortunately Crouch et al pursue their allusion with considerable vigour and extraordinary circumlocution and tautology. They write of the “amateurish attempts of interested lay persons and taxonomists alike to force an absolute and linear and dichotomous order on the genus”. Unfortunately it is exactly this flawed reasoning and logic that force me to wonder if it is even worth pursuing knowledge anymore (C&SJ.US 70:183, 1998). Our classification system is essentially a dichotomous one and the very concept “genus”/”species” is itself a dichotomy. I will try to explain new findings without attempting to invent a new classification system, although we undoubtedly need one.
New records While re-writing my Handbook and formally revising Haworthia, I was required to cite all herbarium specimens. There are 15 records for incurvula alone and they are all from Pluto’s Vale. None give any detail outside of that geographical information statement. Thus there is no evidence to support Scott’s contention of.. “a number of localities along Pluto’s Vale”. J.D. Venter has a collection (JDV93/60-81 [81 a-d]) by Scott’s colleague, the indefatigable Dr. Reddi, which states “middle of Pluto’s Vale”. This collection was said to have been made north of the road.
There is of course a whole range of specimens of H. cymbiformis from various places in the Eastern Cape. These should really be discussed, analyzed morphometrically and illustrated. These would show why the elements planifolia and cymbiformis can be treated as synonymous. This synonymy is largely due to the origination of the names, and suggests simply that two such morphological elements could have been from a single local system. The herbarium evidence would indicate why the concept “incurvula” is reasonably included there among the many varieties of H. cymbiformis described at one time or another. These are ramosa, reddii, transiens, obtusa (recognised by myself), and further names such as lepida, angustata, brevifolia, alta, agavoides, incrassata, calochlora, longifolia, compacta, exulata, setulifera, obesa and olivacea (which are not).
There appear to be several populations from the greater Grahamstown area in which the plants are smallish and with bluntish incurved leaves (E. Fort Brown, Blaaukranz, Vaalvlei, Fernkloof, Kaffirdrift). There are also populations in which the plants are larger with acuminate, flatter leaves in a more spreading rosette (Committees, Howiesonspoort, Kapp-Fish confluence) and populations with obtuse turgid leaves (Alicedale, Swartwaterpoort).
Here we are expressly concerned with Pluto’s Vale and the plants which occur there. Curiously incurvula is at its closest and most accessible to collection, near the top of the steep descent into the valley, Pluto’s Vale. There is a gate there into the field and a dwelling at a point here called Roff’s Rock. In the tradition of plant collecting in South Africa, as exemplified by the discovery of the first Lithop species by Burchell in the 1800s, collections are invariably alongside a road, or within very short walking distance of some obvious stopping point. The Roff’s Rock site is just such a place and incurvula is less than 50 meters from the roadside. There is a also a vast population of Haworthia attenuata (var. britteniana), and also the plant which the collectors probably actually and repeatedly came to view ie. Encephalartos trispinosus. Incurvula also occurs mid-way down the steepest descent of Pluto’s Vale, north of the road and immediately adjacent to it. Tenera is also known from the farm Glenelg to the north, to the east near Committees and also to the south-east at Hunt’s Drift. H. cymbiformis is a separate element in those same areas and I did cite specimens of incurvula from south of Hunt’s Drift too (Smith5741, Smith6508). I have not seen these populations to confirm that they are, as living specimens, in fact comparable.
In November 1998, I specifically visited Pluto’s Vale again to explore the area a little more thoroughly and to re-locate my glabrous H. tenera. My wife, Daphne visited the area again during July 1999 because of the failure of the previous visit to locate that form. We visited the site again in September 1999 together with Steven Hammer and with Tony Dold of the Schonland Herbarium. A sketch map is provided of the area to demonstrate what we found where. The first visit was collectively quite an extensive search, but by no means adequate to establish the whole truth.
Tenera was found to be common in the Vale itself i.e. east of Roff’s Rock (MBB6832 [82 a, b], MBB6833 [83], MBB6834 [84], MBB6835 [85], MBB6840 [86]). Venter collected at the exit from the valley (JDV90/36 [87 a-c]) and also at Hunt’s Drift (JDV91/103 [88 a, b]).
Incurvula was abundant from the vicinity of Roff’s Rock and extends along that ridge south of the road to where the ridge ends at the base of the Vale (MBB6836 [89 a-f], MBB6837 [90 a-f], MBB6838 [91.1 a, b], MBB6839 [92 a-c]). It was also collected by Marx and Venter (JDV93/73 [93 a, b]) on one occasion slightly towards the south-west of Roff’s Rock, and we also collected it at the second visit (MBB6893 [94 a, b]) still further west. I could not locate the original site of my glabrous tenera (KG47/72a) north of the road and in the Vale. The normal spinose form was on all the south facing rocks. The vegetation seems to have been modified by extensive grazing of the two farms and management systems now evident (the one area is now a game farm and goats have been introduced on the other). However, on the most recent visit we did again find the glabrous form (MBB6891 [95 a, b]) in profusion almost directly above the incurvula population (MBB6892 [96 a, b]) north of the road. In the first version of this report (Bayer, 1999) I incorrectly cited JDV93/60 [81 a-d] as Dr. Reddi’s collection of incurvula from south-west of Roff’ Rock. Apparently it was actually collected north of the road in the middle of the Vale. I can now associate it with my collection [96 a, b] of Pluto’s Vale which is however, of smaller plants than Reddi’s.
The Marx/Venter [93 a, b] collection is of plants with longer (to 50mm) sub-erect leaves which are very lanceolate as opposed to the ovate-oblong shape and incurvature of the leaves in the original description. Traditionally and conceptually incurvula has leaves which are obtuse at the tip. The description does not circumscribe this, and this could have become a very important point. The Marx/Venter plants have such dramatically lanceolate leaves that they throw a slightly new aspect onto the scene. My first collection [89 a-f] between the given Marx/Venter site and Roff’s Rock, also is of plants with such lanceolate and pointed leaves. My wife turned up a similar population north-west of Roff’s Rock (MBB6884 [97 a-d]) in which the plants were smaller and proliferous on a rockier, steeper, south face. There is a stronger tendency to spination. The colour is towards the grey rather than the green. The leaves are more gracile and acuminate. There is no turgidity or swelling of the leaf as one finds in pilifera or in cymbiformis var. obtusa, and given as “inconspicuously so” in Von Poellnitz’ description. But this collection is not the same as the two glabrous ones which so obviously relate to tenera. It does perhaps relate better to Hellspoort gracilis, although it is fairly similar too to [89 a-f], [93 a, b] and [94 a, b].
There is a collection by Marx and Venter (JDV96/4 [98] also by myself MBB6603 [99 a-d]), from north-east of Grahamstown at Glen Craig. H. cooperi var. pilifera also occurs there but about 2km to the east. This collection is of largish plants, greener rather than the bluish/purplish coloration of the cooperi complex. The leaves of the plants are ovate-lanceolate and not turgid above the mid-point. There are many collections of look-alikes from as far afield as Adelaide, the Zuurberg and the Humansdorp area. There is also a very interesting record by R.A. Bayliss from north-east of Grahamstown that needs following-up. These collections impinge on the interplay of H. cooperi var. cooperi, H. bolusii var. blackbeardiana, H. decipiens var. pringlei and H. aristata, which is the subject of another manuscript.
Again: the four collections made from a point south-west of Roff’s Rock to approximately 2 kilometers along the same southern face of the ridge down into the Vale. The first collection was the one described above [89 a-f], of plants with the more lanceolate and acuminate leaves of the Marx/Venter collection, the second [90 a-f] was about 200 meters eastwards which included plants with acute and awned, and plants with obtuse, leaf-tips. One specimen of the third collection [91.2] is identical to a clone in a collection from Alicedale (MBB6847 [100]), a significant collection of H. cymbiformis var. obtusa which I will write about on another occasion. There are close similarities to a collection of mine of this same element from NW Grahamstown (the Bayliss site?, Thornkloof, MBB6895 [101]). That collection was made in pursuit of a R.D. Bayliss collection KG382/76. The progression at Pluto’s Vale to the foot of the slope, was to obtuse leaves and the cymbiformis-like appearance which is also evident in collections of mine from greater Humansdorp (eg. [15 a-e]). The plants were often single and occasionally with more than one rosette in a cluster.
An interesting point is that of the relationship between marginal spination and pointedness of the leaf-tip. One might expect, as indeed I did, that the two situations would be correlated. To a degree they are. Incurvula has leaves which are seldom spined but often blunt-tipped, in tenera the leaves are generally spined and as in glabrous tenera, always with a pronounced point to the leaves. In the case of H. cooperi var. pilifera from Slagtersnek [104 a-e], there are clones in which the leaves are very truncated as in Col. Scott’s joeyii (the var. dielsiana). There is no tip to the leaf at all, and yet the margins do have spines.
The bluntness of the leaf-tips cannot thus be taken to be diagnostic. Here we have pungent and blunt leaf-tips in a continuum of specimens from a relatively extensive area. The “Flowering Plants” illustration appears to have no tip to the leaf per se, whereas J.R. Brown writes “Occasionally a leaf may be found with a very short pellucid point.” His photograph shows leaves with a distinct short abrupt blunt mucro (point), but nothing like the tapered sharp extended point evident in many of the plants I saw. This variation alternative is also evident for H. cymbiformis, for H. cooperi and for H. gracilis within the interactive systems that they are. The phenomenon is also evident in the south-western Cape complex where for example we have H. magnifica which has leaves which are acute with long end-awn, and the var. atrofusca in which the leaves are blunt (obtuse). This is evident within populations.
Tenera was collected in the Vale from a point north of the road [86] from where my search for incurvula terminated. We found it on all of the five rock faces we searched from this point to the eastern end of the Vale. There our earlier expedition ended without real conclusion. It is proliferous and rosettes are usually in clusters. The two elements tenera and incurvula thus grow in close proximity. While in Aloe (1999) I wrote that there was no direct evidence of obvious introgression, or morphological or genetic continuity. My recent collections of small incurvula[96 a, b] and glabrous tenera[95 a, b] require a more committed opinion. The connection is thus through the glabrous populations of tenera directly to incurvula, and also via the three most western and northern pointed-leaved plants from glabrous tenera to incurvula.
In the case of my latest exploration I returned to the Andrieskraal area (SW Patensie) in the eastern Baviaanskloof. I wanted to get some idea of what G.G.Smith had recorded in the Mistkraalkloof area. I already had collected H. gracilis var. picturata at Andrieskraal itself [14 a-c,15 a-e]. This time I collected a little further east [16.1 a-d, 16.2] and these are smaller, non-proliferous plants. One clone was only 16mm in diameter with 35 leaves, while another was 25mm diameter with less than 25 leaves. These plants bear a surprisingly similarity to incurvula, and apart from the blunt leaf-tips, also to the glabrous tenera. Curiously I collected from another population [17 a-c] a little further to the east. This was a glabrous H. gracilis var. isabellae which is very similar too, to glabrous tenera.
In this particular paper I do not consider H. cooperi, which can elsewhere found to be continuous with both gracilis and cymbiformis. H. cooperi var. pilifera is present to the south-west of Pluto’s Vale at Glen Craig, to the north at Fort Brown, and at Committees to the north-east. It does not appear to be involved as such at Pluto’s Vale.
The conclusion There is an ambience to the literature on Haworthia which makes it very difficult to write meaningfully and truthfully about the genus. This ambience reaches a climacteric in the recent article by Crouch et al cited earlier. The review of Haworthia Revisited by the botanist Paul Forster is also so much in the stereotyped and conventional mould of taxonomic botany, that there is little useful to direct scepticism and inquiry in a direction which can possibly advance understanding of classification. This is not true only for Haworthia. Speciation in Haworthia (if not also in many other plant genera, as I have said elsewhere) is chaotic. This can be construed in various ways and I hope it will one day seen to be in a proper sense of order rather than disorder. A search, very thorough by the standards one is generally led to observe, has resolved the relationship between incurvula and tenera to a greater degree than the first publication of this report. Yet, it cannot be stated categorically that incurvula belongs in H. cymbiformis nor that tenera belongs in H. gracilis. A pattern is only evident here for which I have now evidence and this repeats itself in several other areas. Exploration and understanding of these patterns is necessary but is not going to make description and classification in the traditional way, any easier.
One of my observations about natural hybridisation in Haworthia is that hybrids are always very conspicuous, and usually more so because of their close proximity to both parents. On several occasions I have collected hybrids in physical contact with one or other of the putative parents and seldom in the absence of both. The word “putative”, means by inference, but my conclusion is that the use of the word in relation to natural Haworthia hybrids is that it is too passive. The inference I make is that genetic interchange between populations is very limited and populations are able to develop and maintain their own integrity over very short distances. Thus we have glabrous tenera and small incurvula existing as discrete entities in close proximity (75 meters), with a potential third element representing possibly gracilis as the four western collections of plants with the pointed leaves. A parallel situation with three main role players is also evident in the Baviaanskloof and it has always been observance of the degree of co-occurrence of these role players which has led me to my classification. I do not think that these role players really represent “systems” and thus species in themselves, but they can be explained in terms of my classification and revision. I would suggest that my classification is valid as an hypothesis for further inquiry along these same lines.
There are still many outstanding doubts and a decision to “force” change in the dichotomous order will remain very inviting to the uninitiated. There is a very wide interplay between many “species” in what is a very widely spread and intricate system or systems. We are dealing with visual imagery and trying to create order in this. The fact is that we are really dealing with unquantified miniscule differences of size, colour and leaf shape, with a tendency to seek and attach importance to even more miniscule differences of spination, translucence etc. in individual plants. There is also a tendency to attach significance to sophisticated technology applied to limited samples. The problem with this is that it is difficult then for adequate sampling, repetitive testing and casual review.
As mentioned, a similar manuscript which deals with the interplay of the species gracilis, cymbiformis and cooperi, and another which deals with the interplay of aristata, bolusii var. blackbeardiana and cooperi. These show that taxonomic solutions which were thought to be easy to present for some small areas and for small samples are not. Such solutions also break down in other areas. It is very difficult indeed to draw all these “species” into a single discussion, but I feel confident that my classification and revision can be used for this purpose. It has been successfully used like this since 1976. The “group” species solutions as given by T. Salter for Oxalis and by O. Hillyard for Streptocarpus are not very satisfying. It is not a question of forcing the imposition of a linear system. This IS the system in general use. We need another one. I am impressed with a bifurcation diagram given in Glieck’s book “Chaos, the Making of a New Science”. This particular diagram suggests that, made three-dimensional, with asynchronous and asymmetric bifurcation, it could model speciation in plants. The model would be presumably be described by a non-linear differential equation. We need a new language and/or a new paradigm, if we are to name and communicate about plants in a harmonious way, understandable and acceptable way. I hope we will be able to understand that when it comes, because we are not doing very well in the current language we use. ♦
81 a. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 b. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 c. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 d. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.82 a. MBB6832 H. gracilis var. tenera, east of Roff’s Rock.82 b. MBB6832 H. gracilis var. tenera, east of Roff’s Rock.83. MBB6833 H. gracilis var. tenera, east of Roff’s Rock.84. MBB6834 H. gracilis var. tenera, east of Roff’s Rock.85. MBB6835 H. gracilis var. tenera, east of Roff’s Rock.86. MBB6840 H. gracilis var. tenera, east of Roff’s Rock.87 a. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.87 b. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.87 c. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.88 a. JDV91/103 H. gracilis var. tenera, Hunt’s Drift.88 b. JDV91/103 H. gracilis var. tenera, Hunt’s Drift.89 a. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 b. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 c. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 d. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 e. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 f. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.90 a. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 b. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 c. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 d. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 e. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 f. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.91.1 a. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock.91.1 b. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock.91.2. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock cf.[100, 105a-f].92 a. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.92 b. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.92 c. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.93 a. JDV93/73 H. gracilis var. gracilis, southwest of Roff’s Rock, Pluto’s Vale.93 b. JDV93/73 H. gracilis var. gracilis, southwest of Roff’s Rock, Pluto’s Vale.94 a. MBB6893 H. cymbiformis var. incurvula, 400m west of Roff’s Rock.94 b. MBB6893 H. cymbiformis var. incurvula, 400m west of Roff’s Rock.95 a. MBB6891 H. gracilis var. tenera, smooth form at foot of Pluto’s Vale.95 b. MBB6891 H. gracilis var. tenera, smooth form at foot of Pluto’s Vale.96 a. MBB6891 H. cymbiformis var. incurvula, foot of Pluto’s Vale, north f the road.96 b. MBB6891 H. cymbiformis var. incurvula, foot of Pluto’s Vale, north f the road.97 a. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 b. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 c. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 d. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.98. Marx & Venter in JDV96/4 H. gracilis var. gracilis, northeast of Grahamstown.99 a. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 b. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 c. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 d. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.100. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.101. MBB6895 H. cymbiformis var. obtusa, Thornkloof, Grahamstown.
One of the greatest difficulties in Haworthia is that of trying to recognise discrete species. This translates into confusion which can be attributed to writers. The initial source of confusion is without doubt the nature of the plants themselves, and this is not a problem confined to Haworthia. The species are often not easily recognisable and discrete entities. I abhor the statement that the genus is in a state of active evolution, but this does at least seem to convey a message that readers understand, even if it is somewhat hackneyed. My observations on Haworthia are based on a definition of species as a system of living organisms which are continuous in time and space. In my New Haworthia Handbook, I suggested that a primary problem lay in separating H. bolusii and H. cooperi, and for the purposes of that work I largely discounted the secondary problems. My first concern was to identify core areas and names as working postulates. This did not mean I was unaware of lesser problems contained within the recognition of those two species. The purpose of this paper is to present my current understanding of the problem.
In my opinion the publication of Scott’s book and whatever merit it had, broke the foundation for understanding. As Bruyns said (Kew Magazine, 1986), it set taxonomy in Haworthia back by 40-50 years. This is because Scott did not attempt to examine what I had done, and was doing, in any objective and cognitive way. Thus there was no progression from a common hypothesis, or from a common concept of ‘species’. In my opinion his work was written in the paradigm of Reynold’s work on Aloe, or of Von Poellnitz and G.G. Smith, and was thus anachronistic. My work was in the intellectual climate of the day, was influenced by researchers of the time and was based on a definition of ‘species’. Scott’s thoughts and actions were both understandable and excusable. His classification reflects the same problems, but differently to mine. Where I recognised H. bolusii he recognised H. bolusii. Where I recognised the var. blackbeardiana he recognised H. cooperi and H. batteniae. Where I recognised cooperi he recognised H. pilifera and H. altilinea. Except for his contention that these latter are separate species, there is really concordance. We recognised different markers along the same continuum.
Essentially I considered that there were two species with two major facies, thus:-
1. H. bolusii – var. bolusii the smaller very densely spined typical variety which seems to diffuse into the species H. semiviva in the west of its distribution range.
– var. blackbeardiana a larger less densely spined element to the south and east which seems to diffuse into H. cooperi. Scott used the name cooperi for this element. There is merit in this, but he had to describe another species, batteniae, because of the inherent tension in the solution (I must point out here that Scott’s problem may have been the inadequate herbarium record that he used for his work, plus the fact that he made few specimens. Thus there is no way of assessing his decisions. Had he had to physically file specimens as I have done, he might have had great difficulty with batteniae and his cooperi, and also in separating his altilinea and pilifera). I regarded bolusii as a more spinose species than cooperi, with thinner and wider leaves. Cooperi then generally less spinose, squatter and with thicker leaves.
2. H. cooperi – var. cooperi itself, with relatively erect slender leaves, and including all the forms with more truncated abrupt leaves.
– var. leightonii – the coastal form with even more attenuate leaves than the typical form, and also very proliferous.
The solutions offered by either myself (1982) in respect of my cooperi and blackbeardiana, or in the case of Scott (1985), altilinea and pilifera, cooperi and batteniae, do have problems. These arise from the nature of the terrain in the area between Graaff Reinet to Queenstown and down to Fort Beaufort. Rugged broken terrain difficult to explore. Like the Baviaanskloof it offers many different classes of habitat, and thus potential ecotypes. The relationship between the varieties is complex and compounded by continuities with other species e.g. gracilis, cymbiformis, and decipiens. It should also always be remembered that any decision is a product of the collecting record and must emphasise that this is the context in which this article is written. Initially I did think that the typical variety of H. cooperi may possibly prove to be well-defined geographically. It has not proved indisputably so, but my classification nevertheless does allow good expression of the continuities which occur. I used the name to cover forms with erect slender leaves as well as forms with relatively blunt truncated leaves. Such plants do occur within the same populations and this is evident in the J.G. Marx collection from Fort Brown. The corresponding Scott names were altilinea and pilifera. To make my system more workable, in my new book (1999) I recognised H. bolusii and its var. blackbeardiana as before, but in H. cooperi, several more varieties, thus:-
– var. dielsiana – a more truncated version of pilifera, the leaves often without end-awns. (Scott’s “joeyii” is synonymous).
– var. truncata – a eastern lighter coloured, proliferous and smaller version of dielsiana.
– var. leightonii – the pinkish, proliferous, slender leaved form growing on granites at Kaisers Beach (Scott maintained this as a separate species).
– var. gordoniana – the erect, slender leaved form in the Hankey and Patensie area, which is very like the typical variety cooperi. In truth var. gordoniana from the type locality is a small very compact plant with short incurved and finely spined leaves.
– var. venusta – a very localised hairy variant from the coast near Alexandria.
I hoped this would explain the variations and resolve the tensions in the solution in an economical way. There is no doubt that Scott’s argument of cooperi sensu Bayer, and blackbeardiana sensu Bayer being the same (and he used only the name cooperi in this context), is correct in the nomenclatural sense. The Thomas River specimen he cites in his book as representative (although Scott does cite the Kew specimen as type, which I have not seen), would have been attributed by Bayer (myself, using the Saunder’s Refugium illustration as type) to my concept of H. cooperi, as also the doubtful gracilis-like elements for the Adelaide and Kingwilliamstown citations. I would have named the Cradock specimen H. bolusii var. blackbeardiana. To bridge the difficulties inherent in his solution, Scott had to later recognise three species viz. batteniae, pringlei and joeyii, with the potential for many more. I have said elsewhere, that a classification which grows with the description of new species, is indicative of a weak system. As in the case of bolusii, the Scott solution is not as economical as mine. His concept of an altilinea and pilifera is essentially the image of my problem in recognising a cooperi/blackbeardiana interface. It would have helped had I initially recognised pilifera as a variety. However, Scott’s interpretation in the sense of the geographical relationships of his various species is problematic throughout his revision, where specimens are not cited and full use has not also been made of the available herbarium record.
I was predicting that an element, namely H. cooperi var. cooperi occurred which could be defined in a geographic context. The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain goodwill. This article thus is written to expand on the problem and indicate where the difficulties lie. These are not difficulties that I was oblivious of, or tried to obfuscate. I have many times said that we can find solutions which work in one area, but they may not work in another. This will again be evident in this paper.
In my revision (Haworthia Revisited, 1999) I made the combination H. decipiens var. pringlei and resurrect the old species name H. aristata. These two elements, and an explanation for the tensions which these two names create and try to address, will be touched on here but will follow in more detail in a later article. As in this immediate case, new material has been seen and collected since the revision was drafted, and a better explanation can now be made.
For this particular issue, the key questions to the hypotheses of Bayer and Scott are difficult to formulate because they are confounded by the different use of names. Therefore I frame the questions in terms of my own classification like this:-
Does an element we can identify as cooperi grow in the same close geographic context as blackbeardiana? (Bearing in mind the assumption that the hypothesis regarding continuity between bolusii and blackbeardiana in fact is valid. Also noting that to maintain the classification hypothesis that there is a difference at the rank of species, the answer must be ‘yes’).
Does the element we can identify as cooperi grow in the same close geographic context as pilifera? (Bearing in mind that to maintain the classification hypothesis of a difference only at the rank of variety, the answer must be ‘no’).
Results The answer to question 1:- has never been a strong point of my classification because I saw cooperi in a broader sense to include pilifera (and altilinea of Scott). The recognition of varieties now strengthens the classification in one respect but weakens it in another. It strengthens the classification in respect of recognising the extent of the variability within my concept of H. cooperi. It weakens it in suggesting both a strong geographical separation of the varieties and a weaker relation with H. bolusii var. blackbeardiana. In Nov.1996, I went with Peter Bruyns to the Eastern Cape and we spent some time in the greater Somerset East area. In Dec.1996 I travelled with Kobus Venter. In Dec.1997, Dec.1998, in Sept. (with Tony Dold, Dez Weekes and Steven Hammer) and again in Oct.1999, I travelled with my wife to these areas. We made many significant collections.
The first of these concerns Scott’s species, H. joeyii and H. pringlei. In the case of joeyii my contention that it is continuous with pilifera barely needs discussion, and I do not think the presence or absence of an end-awn is necessarily diagnostic for such an element – hence synonymy with var. dielsiana. We found three discrete populations within a small radius around Eastpoort (MBB6558-102, 6559 and 6560-103) which support this observation. I do not think there are strong grounds for separating it from pilifera, although I have now done so. At Bedford (there are plants with the similar abrupt leaf-tip as well as plants without. I currently have four batches of seedlings which I want to examine for variation. One from Slagtersnek, south-east of Somerset East (MBB6778-104) is particularly variable. I even have a collection of H. cymbiformis (MBB6847-100,-105) which has the same truncated, awnless leaftips as Scott’s joeyii. Regarding Scott’s pringlei, my conclusion that it is related to decipiens is possibly geographically and otherwise incorrect. (It should be noted that no specimen of pringlei has to date been deposited in the Pretoria herbarium. My understanding of the species is from the description, from two plants sent to J.D. Venter by J.N. Reddi (JDV93/46-106) who collected the plants for Scott, and from two plants given to G. Marx (JDV93/52-107) also by Dr Reddi). It is a problem of ‘look-alikes’ and I will deal with that a little later as a separate issue.
Further south and east of Eastpoort, at Patryshoogte (MBB6561-108 also -4), we found a different population not to my knowledge co-occurrent with a cooperi variety, which must be comparable with Scott’s species i.e. pringlei. It has narrower and more elongate leaves than blackbeardiana generally has, and the keel may be more pronounced. Nevertheless I am sure it has a connection with H. bolusii var. blackbeardiana. In terms of a much wider knowledge of other populations, I decided to link pringlei to decipiens, to emphasise the evident and probable continuity between that species and H. bolusii. This is particularly so in the area north and east of Jansenville.
To the south-west at Somerset East (MBB6776-109, Glen Avon), we found a population of bluer green plants with rather more attenuate leaves than in the vars pilifera and dielsiana. These plants are very like an Adelaide (Koonap Bridge, MBB6563-110) collection, these both satisfy my concept of the var. cooperi. It was not much further to the south (-104), and also not far south of Eastpoort, where we found forms of the vars pilifera and dielsiana in the same population. Thus it seemed that a hypothesis maintaining the species cooperi and H. bolusii var. blackbeardiana, was well supported, but not so the contention that pilifera and dielsiana were good varieties.
The most significant discovery up to Dec.’97 was of two different elements growing together towards the south, near Ripon Station. By together, I mean as populations occupying different niches in the same close proximity – not growing as a common medley of individuals. One population was clearly the typical blunt-leaved pilifera (MBB6557-111) which included dielsiana, but the other (MBB6556-112) was with more slender and erect leaves viz pringlei, and possibly the leaves being broad and spinose enough to even represent blackbeardiana. Thus the added complexity is that it is not possible to separate those two elements. After two years in cultivation as field collected clones and as seedlings, the difference from pilifera (as -111) is maintained. The collected clones grown in Kobus Venter’s collection convincingly demonstrate that the one population is identifiable with pringlei/alias blackbeardiana and the other with cooperi var. pilifera. Thus the answer to the first key question is “Yes, they do grow together in the same geographical context, because there is that obvious connection of blackbeardiana with pringlei. It seems probable that both these elements could, and perhaps should, be interpreted as varieties of H. cooperi. It is a complex problem which cannot be separated from consideration of other elements such as H. gracilis and H. aristata, and these are the issues I am addressing in these papers.
There is not much further evidence that cooperi var. cooperi is a strong, discrete and valid element. There is a collection by J.G. Marx of plants in the Hogsback area (in JDV91/82-113, Woburn) which may be identified as this variety, as can the two collections already mentioned viz. -109 Glen Avon, and -110 Koonap Bridge (originally a Marx collection). However, there are collections generally (one is D.M. Cumming8489-114 of plants south of the Waterdown Dam, Upper Chilton), where the plants can be confused with H cooperi var. gordoniana (in the context of the problem in the Baviaanskloof), whereas geographically they should be blackbeardiana. I also made a collection at Waterdown Dam itself (MBB6569-115), of a plant which is very like a spineless smooth gracilis. These plants tend to be much more blue in colour than the southern look-alikes. There is also JDV96/89-116 Gladhurst, Adelaide; and JDV96/4-98 and MBB6603-99 from Glen Craig, north-east Grahamstown, which pose a similar conundrum. These are gracilis-like and while exploring that problem, the issue became very much more prominent, and is even expressed in the discussion regarding H. cymbiformis var. incurvula. Kobus Venter and I had both re-collected H. gracilis var. gracilis at Hellspoort (JDV89/42-117, MBB6614-118), although we had not concerned ourselves with a search over the whole of the valley. What is now becoming more evident is that there is an archetype which is gracilis-like. Thus Hellspoort needs to be examined as closely as Pluto’s vale.
A later collection of mine (Oct.’99, MBB6927-119, W Ripon) influences the picture dramatically and may further prove a statement of mine true. This is that there instances where there are no real boundaries between species. My collection is further west of -111 (pilifera/dielsiana) and -112 (pringlei/blackbeardiana). Here in -119, the plants include individuals which could be either identified as the vars cooperi or pilifera, with the former collection equally well representing pringlei/blackbeardiana.
The answer to question 2:- has not really been a problem. What we found was what we expected to find and more. South of Adelaide one finds populations of plants which satisfy the tendency towards slender leaves ie. var. cooperi (-109 Glen Avon or -110 Paardefontein, Koonap Bridge); and populations which satisfy the tendency to short obtuse leaves ie. var. pilifera eg. MBB6564-120 Chancery Hall or MBB6591-121 The Tower, S. Fort Beaufort. The same applies just east of Somerset East where in the broader geographical context we can identify var. cooperi (-109, Glen Avon), var. dielsiana (when mostly or wholly without end-awn, MBB6565-122, W. Somerset East, -111 W. Ripon Statio, and vars dielsiana and pilifera as geographically complementary and even in admixture (-104, Slagtersnek). Thus “No, the varieties cooperi and pilifera do not grow together in the same close geographical context as discrete entities.” In fact they are often represented as forms in the same collections as in -119 (far west of Ripon) where both varieties are present as single plants as well as a longer leaved form representing blackbeardiana as decipiens var. pringlei.
The answers given to these two questions thus do still not solve the problem as there remain populations which can not confidently be ascribed to either of the names here used. There are populations which may thus be assigned to cooperi or to blackbeardiana or pringlei. An example is the Marx collection from north-east of Grahamstown (W Fort Brown, in JDV91/85-123) which is ascribable to pilifera although some of the clones are inseparable from blackbeardiana. JDV98/39-124 from Brakkloof, northwest of Grahamstown poses a similar problem with the leaves of the plants tending to lengthen in cultivation. The ultimate difference then between cooperi and blackbeardiana becomes a very subtle one of degree of spination and leaf bulk. The issue is further complicated by the fact that four other elements are involved in the total geographical context, namely H. cymbiformis, H. gracilis, H. decipiens, H. aristata and H. arachnoidea var. xiphiophylla.
My most recent field work, was in following up information and collections concerning H. gracilis. Tony Dold of Rhodes University gave specimens of an Haworthia from the Annsvilla area to Gerhard Marx (MBB6851-125). He thought they may have some bearing on the existence and reality of my interpretation of H. aristata, and this is indeed the case. The plants are small, the darkish blue-green of the cooperi/blackbeardiana elements, and with more, smaller, and quite spinose leaves. Annsvilla is close to three localities for that species cited in my revision viz. Verdun, Stonefountain (re-collected in MBB6852-126 also -4) and Kommadagga. Following Dold’s find, recollection in the vicinity of each of those sites, produced plants which can be interpreted and identified in the light of the Ripon collections. While they equate in some way to H. bolusii var. blackbeardiana, they also do to H. cooperi and to H. decipiens var. pringlei. An (unfortunately doubtful) Kommadagga collection (EvJ sn.-127) is very similar to the long-spined gordoniana-like plants from the eastern Joubertina -74, Uniondale -75, Longkloof -76 and Baviaanskloof (Geelhoutboskloof -78, Nuwekloof -79). The leaves of the plants are rather flatter and broader, and so closer to blackbeardiana. In Sept.’99 Dold and Hammer found a small plant with shorter and squatter leaves than my concept of aristata, south-east of Kommadagga (in MBB6897-128). We also saw plants from the area east of Jansenville which seem to link the greater H. bolusii (i.e. var. blackbeardiana) complex with either H. decipiens or H. arachnoidea var. xiphiophylla. Included in that broad statement is an explanation for the specimens which I have cited as H. decipiens var. pringlei in my revision. These include recent collections from north of Jansenville (MBB6580-129), north-west (MBB6581-130), and south-east of Mt Stewart (MBB6582-131 also -6, 6583-132, also -7). When I first saw a specimen of pringlei my first reaction was recall of plants I had collected at Mt Stewart, and of plants said to have been from Jansenville. Thus I created H. decipiens var. pringlei, expecting to find this east-west continuity between blackbeardiana and decipiens. I did not think that the connection between Middleton (pringlei) and Jansenville would be found to be as evident and as strong as it is. There was already some evidence that there is south to north-east connection between Jansenville and, say, Cradock. This is of a connection between H. decipiens and H. bolusii.
A complicating issue is that MBB6587-133 (further SE Mt Stewart than -132) is a population which is H decipiens var. minor. A small plant with a tendency for incurving leaves but with the widely spaced large spines I associate with the more classical view of H. decipiens. The next collection was MBB6589-71.2 (a little south-east of -133) and it throws us into the H. gracilis context of smaller plants, clustering on cool south-facing rock faces. There are many collections which continue this trend to gracilis, and in fact I suspect to xiphiophylla too, and that will be dealt with as a separate issue. The field-work also revealed populations (J.G. Marx in MBB6845-134, east of Alicedale, and MBB6847-105, also Alicedale) which link cooperi to cymbiformis. This also can be dealt with together with two collections from near Kagasmond (MBB6562-135, south of Adelaide and JDV96/89-116, Gladhurst) as a slightly different issue.
Regarding H. aristata, I resurrected this name for several collections from the Eastern Cape, and somewhat justified by the Dold collection at Annsvilla (-125). However, it seems quite certain that the issue is clouded. Re-collections at the Soutkloof (JDV96/90-136, Addo, Dead Man’s Gulch) were more like pilifera, but there is evidence of deviation. Here D.M. Cumming (DMC3870-137 collected pilifera-like plants, whereas Venter, Marais and Bayer (-136) collected a variant – there was a more typical bluish-green pilifera and a plant tending to the opaque yellow-green of xiphiophylla. I returned there in Oct.1999 because I still had not seen anything like the plants I had seen there on an earlier visit, or like W.R. Branch’s original (WRB459 in JDV87/53-138), and the other cited collections. On this occasion I did find a small population (MBB6920-139) of this kind, and these confirm a relation to the more northern collections mentioned above. At the nearby village of Addo itself, there is the fairly normal pilifera (JDV86/117-140). The Annsvilla collection (Dold and Marx -125) and the recent collection of my own from Stonefountain (-126 also -5) seem also to be in the context of the Ripon collection (-112) of pringlei. These are, however, collections of much smaller plants than from Ripon. Thus it confirms for me that the Haworth names aristata and denticulata could easily have had their origins in plants from this area between Ripon and the Zuurberg, or further south to Soutkloof. My collections MBB6916-141, MBB6917-142 and MBB6901-143, from Kaboega and Hopewell are of the same order.
Discussion and conclusions There is still a very large area unexplored. There are several like collections from the greater area Kirkwood to Uniondale which I have generally ascribed to H. cooperi var. gordoniana. None of these are the short-leaved pilifera type. The teasing probability is that it is in fact blackbeardiana, pringlei, gracilis or aristata (as variants of H. cooperi) which are the main role players. They differentiate (clearly?) in the east to pilifera or cymbiformis and in the west into decipiens. In the south they pass to xiphiophylla, cymbiformis and varieties of gracilis. North-westwards it is to bolusii. My opinion now is that we have an archetype which is in the mould of H. gracilis and this is the root of all the elements I have named here. Curiously Tony Dold has recently sent me specimens from Chalumna (T.Dold3961 in MBB6921-144) which suggest that an aristata-like element is also associated with H. cooperi var. leightonii. Other Chalumna collections (MBB1621 and G.G.Smith 514) bear a very close resemblance to plants (-99, -116) which I have said are gracilis-like.
The above statements all have to be seen in the overall statement about continuity. It barely seems practical (nor legally possible) to sample and analyze plants on the scale that will be necessary, using whatever technique, to get a more definitive answer. The best answer will be continual exploration of the simple kind reported here, which explains occurrences on a smaller and smaller scale. There are herbarium records which need to be corroborated in the field but these do not suggest to me a better solution. Ultimately each recorded population will have to be assigned unequivocally to a taxon (or taxa!) and this is the next necessary step in Haworthia classification. Another revision based on less exacting field observations and a lesser record, will simply exacerbate the regression in time which Scott’s work precipitated, and the consequences which we now suffer. My recent collections are only preserved as living collections and photographically. Specimens need to be made.
I am very conscious of other tensions in my classification and I think it is imperative that we stay with one nomenclatural arrangement and hypothesis to resolve these. The confusion which arises from nomenclatural changes which are nothing but pretentious and cosmetic, is not worth any price. Any difficulties in respect of nomenclature can be resolved by a process of explanation and conservation without formal name change. These changes can be made when it is expeditious to do so and when changes can really offer a better explanation of the genus and hence better communication.
What this discussion should demonstrate is the problem of really observing and discussing variability in Haworthia. There are many possible arrangements of names which can be presented as conclusions in themselves – but done like this they simply cloud and destroy any hope of a broader understanding and good communication. Haworthia is now so much in the public domain, that I would suggest to editors that they move in the direction of encouraging authors to adopt a conserved nomenclature. I regret to be so straightforward and blunt. I see nothing but further confusion if persons feels that they can contribute to an understanding of the situation in the field without:
Consensus on the issue of species definition.
Consensus on the issue of a set of names.
Familiarity with herbarium records and what these represent in terms of fact and fixed reference points.
Familiarity with the written record.
Familiarity with South African geography and the ability to interpret populations in that context.
♦
102 a. MBB6558 H. cooperi var. dielsiana, north of Eastport.102 b. MBB6558 H. cooperi var. dielsiana, north of Eastport.103 a. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 b. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 c. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 d. MBB6560 H. cooperi var. dielsiana, south of Eastport.104 a. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 b. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 c. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 d. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 e. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.105 a. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 b. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 c. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 d. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 e. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 f. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.106 a. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.106 b. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.107 a. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.107 b. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.108 a. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 b. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 c. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.109 a. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 b. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 c. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 d. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 e. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.110 a. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.110 b. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.111 a. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 b. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 c. MBB6557 H. cooperi var. pilifera, west of Ripon station.112 a. MBB656 H. decipiens var. pringlei, west of Ripon station.112 b. MBB656 H. decipiens var. pringlei, west of Ripon station.112 c. MBB6556 H. decipiens var. pringlei, west of Ripon station.112 d. MBB6556 H. decipiens var. pringlei, west of Ripon station.113 a. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.113 b. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.114 a. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.114 b. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.115. MBB659 H. bolusii var. blackbeardiana, Waterdown Dam.116 a. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 b. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 c. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 d. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 e. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.117 a. JDV89/42 H. gracilis var. gracilis, Helspoort.117 b. JDV89/42 H. gracilis var. gracilis, Helspoort.117 c. JDV89/42 H. gracilis var. gracilis, Helspoort.118 a. MBB6614 H. gracilis var. gracilis, Helspoort.118 b. MBB6614 H. gracilis var. gracilis, Helspoort.119 a. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 b. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 c. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.120. MBB6564 H. cooperi var. pilifera, Chancey Hall, south of Adelaide.121 a. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.121 b. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.122 a. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 b. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 c. MBB6565 H. cooperi var. pilifera, west of Somerset East.123 a. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.123 b. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.124 a. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.124 b. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.125 a. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.125 b. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.126 a. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 b. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 c. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 d. MBB6852 H. aristata, Stonefontain, s. Somerset East.127. EvJ H. aristata, Aalwynspoort, east of Commadagga.128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.129 a. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 b. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 c. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 d. MBB6580 H. decipiens var. pringlei, north of Jansenville.130 a. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 b. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 c. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 d. MBB6581 H. decipiens var. pringlei, northwest of Janesville.131 a. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.131 b. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.132 a. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.132 b. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.133 a. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.133 b. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.134 a. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 b. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 c. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.135 a. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 b. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 c. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 d. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.136 a. JDV96/90 H. cf. aristata, soutkloof, Addo.136 b. JDV96/90 H. cf. aristata, soutkloof, Addo.136 c. JDV96/90 H. cf. aristata, soutkloof, Addo.137. D.M.Cumming3870 H. cf. pilifera, Soutkloof, Addo.138 a. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.138 b. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.139 a. MBB6920 H. aristata, Soutkloof, Addo.139 b. MBB6920 H. aristata, Soutkloof, Addo.140. JDV86/117 H. cooperi var. pilifera, Addo.
Introduction Kaboega (also spelt Kabouga) is now an assemblage of farms (De Plaat, Wilgerfontein, Vygeboomfontein, Klipfontein) nestled against the north slopes of the Zuurberg mountains, north of Kirkwood. It is only about 15km away from Kirkwood as the crow flies, but 150km away by road. Oudekraal is about 20km east and it is the source of Haworthia angustifolia var. baylissii and Gasteria baylissiana. There are several records of Haworthia for the Kirkwood area, and von Poellnitz named H. stiemiei (Regarded as insufficiently known and not recognised by Col. C.L. Scott or myself) from there. He also identified plants from Kaboega and Uyepoort, both described as “at Kirkwood”) as H. altilinea var. denticulata (Haw.) v. Poelln. These plants are all in the melange that I attribute to H. cooperi var. gordoniana (the subject of another long essay). The Kaboega farm lies on the Kaboega river which drains an area of about 1m ha and then flows through the long Kaboegapoort into the Sundays River just north-west of Kirkwood. The terrain is very broken with the sandstone Zuurbergs themselves dominating the southern boundary at about 850 to 950m above sea level. The lowest point on the farm is at about 300m and the northern lesser shale or dolerite peaks reach 550 to 650m. The vegetation on the sandstones is Dry Mountain Fynbos. North of this is Karoo Valley Bushveld. Thus Kaboega is at an ecotone of the karoid veld, Eastern Cape grassland and the Noorsveld (Euphorbia thicket) of the Jansenville area.
The name Kaboega means ‘the Big Hole’, referring to the deep gorge which the Kaboega River cuts through the Zuurberg. This river joins the Sundays River where it skirts the eastern end of the Kleinwinterhoek Mountains. Thus Kaboega Gorge is about 20km east of the Sundays River Gorge and 20km west of Oudekraal where the Witrivier also cuts through the Zuurberg flowing southwards.
The only known Haworthia collections from Kaboega prior to this report are two collection by Gerhard Marx (JDV91/14-145, JDV91/15-146) from the easterly part of the farm (DePlaat, north and south aspects), and a similar collection by Peter Bruyns (PVB5002 in JDV92/33-147) from Kaboegapoort itself. Discounting the strange (expected) internal variation in the latter, these three collections are fairly similar. Plants with the brighter yellow-green of H. cymbiformis, but with more terete and slender sub-erect leaves. The plants are quite robust and in cultivation reach about 80mm diam. with leaves up to 90mm long. Look-alikes are found in the gracilis, cooperi and cymbiformis var. transiens complex of the Baviaanspoort, and I have generally referred these all to H. cooperi var. gordoniana. However, that variety is actually quite a distinctive one from the Hankey Pass, north of Humansdorp and perhaps I should never have adopted it for general use in the way I have. Thus in my discussion about H. cooperi and H. bolusii var. blackbeardiana, I speculate that gracilis is an archetypal form which may lie at the root of the Eastern Cape species here being discussed.
The species H. aristata poses similar problem, and so does H. decipiens var. pringlei (Scott) Bayer as well as two collections which I and Bruyns made at Ripon station which is north-east of Kaboega. One of these latter collections is H. cooperi var. pilifera (-111) and the other is H. decipiens var. pringlei (-112). Largely because of that collection, I felt pressured into believing that the latter would best be coupled with H. bolusii var. blackbeardiana rather than with H. decipiens, and I was contemplating a major change of this kind. This would also have involved subsuming that element in an enlarged H. cooperi var. cooperi. There are, however, some other collections from the greater Darlington Dam (Lake Mentz) area to the west, which are relevant to this problem. These include older ones which suggested the link of pringlei with decipiens which I was thus also predicting, and new ones which confirm that this does in fact happen.
Because of the extent and complexity of the problem, this report deals specifically with the Kaboega area. In conjunction with it, a manuscript regarding H. cymbiformis var. incurvula, was written to give another indication of the nature and scope of the problem of classification of Haworthia. However, the chief reason for the visit to Kaboega was somewhat fortuitous. I was intending visiting Pluto’s Vale again, also the farm Thornkloof where Col. R. Bayliss had collected; the place Aalwynpoort to check on an Ernst van Jaarsveld collection and also hoping to cast light on a collection from the Bosberg at Somerset East. Peter Bruyns was hoping I would recollect a Stapelia aff. kougabergensis which he had seen on the Zuurberg, and there are also some other Haworthia records in that general area which need verification. What also materialised was a visit by Steven Hammer to South Africa, and contact with Tony Dold of the Schonland Herbarium at Grahamstown. The best of all was contact by my wife Daphne, with the gracious managers of the Kaboega farming enterprise, Sandy and Ian Ritchie. Through their kindness and hospitality we came to spend four days on the farm and briefly explored what it has to offer. One day there was spent with Steven Hammer, and with Tony Dold and Dez Weekes of Grahamstown. My wife and I returned for a second visit in Sept. ’99, when we also went to Oudekraal via a direct farm road from Kaboega to the east.
Results On our first visit we first explored the western side of the farm known as Wilgefontein. Tony and Dez went up the slopes of the Spekboomberg on the north side of the valley, and the remainder of the party climbed to near the top of the Zuurberg. We climbed straight to the grassveld where the grass was very long and thick. We soon found a solitary-growing greyish-green plant in flower (MBB6904-148), and then higher and on vertical rocks, a less translucent clump-forming plant with a velvety texture – also in flower (MBB6905-149). The plants looked rather different and we were quite excited about it being something apparently out of the ordinary. It is possible in the context of later collections, that these two collections are ecotypes. Looking at similar rocks about 200m to the south-east, we found what at first was obviously H. coarctata var. adelaidensis (MBB6907). This turned out to be a big population of plants which can, as such, be collectively regarded as intermediate between H. glauca and H. coarctata. This is a very significant collection because of the occurrence of glauca in its typical form at Zuurberg Pass, and nearer at Oudekraal, both to the east. Var. coarctata itself is not known nearer than at Patterson 70km eastwards and var. adelaidensis from east of Riebeek East which is still further away. (see also ‘Haworthia Revisited’ p179). What we did hope to see was H. angustifolia known at its western limit from Oudekraal. It must be on Kaboega and we just have not seen it yet.
Across the valley Dez Weekes had collected three specimens from a south facing steep cliff (MBB6903-150). These had long stems with bright green terminal rosettes and I have identified these as H. cymbiformis (with reservations! as I think this could again be a local ecotypic adaptation) without seeing the plants in habitat myself. This species is also not known from nearer than Hell’s Gate 50km to the south, but – it almost certainly has affinities with the plants collected by Marx and Bruyns. Steven, Tony and Dez had to leave after the first day, but Daphne and I continued the exploration the second day with a long climb up the hill east of DePlaat. We soon found Gerhard Marx’ (MBB6909-151) plants at the base of the mountain and continued eastwards and upwards. We came across a very extensive and dense mass of H. glauca (MBB6908). The plants were variously tubercled and lacked the distinctive grey colouration of the species. Any affinity with “coarcata” was less obvious than at Wilgefontein. Another interesting plant there was a dwarf form of Aloe tenuior. We crossed over to the steep north slopes and on the way down came across three plants (MBB6910-152) of what appeared to be similar to the velvety plant of the previous day (-149). It was in the same rocky situation. We looked further for it, but failed to find it again probably only for lack of concerted effort. The terrain was very difficult and we were getting a little hot and tired. We came across the Marx plants again. These were further down the hill and looking rather bleached in the sun.
149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.
On the way home we were travelling across some very stony ground covered with scattered bush and a low-growing Felicia, probably filifolia. Underneath these, in algae and moss, and with Euphorbia stellata and Tylecodon reticulatus were single plants of a cryptic small blue-green species (MBB6917-142). This I relate to collections from Stonefountain and Verdun cited in my revision under H. aristata, again collected as MBB6852-126 and Dold in MBB6851-125 (Modderfontein).
142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
The following day, accompanied by Sandy Ritchie, we ventured into the Kaboegapoort itself. We walked to the boundary with the Addo National Park. On the way we had seen inaccessible clumps of an Haworthia on a very steep cliff and we tried to reach these on the way back. We were lucky to be able to sample four clumps on the first of the rock faces where some plants had established themselves off the face (MBB6911-153). On the other cliff we could not reach anything. The plants were not cymbiformis but relate rather to the Marx plants except that they were clump-forming and bleached. A better comparison is with (-148). They were also in flower.
148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.
Peter Bruyns collection (-147) is of six clones from this poort and each of these plants is different. However, one clone (-147.1) resembles gracilis var. viridis from Perdepoort (-70) and also resembles a collection of Bruyns of cymbiformis var. transiens (-69) from the Langkloof. (This latter collection is involved in the issue of that species or gracilis var. picturata). Kobus Venter also has a collection of similar plants from south of Lake Mentz (-72.2). A second clone (-147.2) was an aberrant plant with very terete, abruptly mucronate leaves which were also unusually translucent. Another clone (-147.3) was of a plant identical to the big gracilis var. isabellae of the Krom River Estuary (Ripon, WRB1, recollected by myself -39), and comparable with a collection (MBB6855-154) from Waterford, east of Jansenville. That collection could be identifiable as H. arachnoidea var. xiphiophylla and perhaps hinting at a link of that element with H. decipiens var. minor. Two clones (147.4, 147,5) are the same as the DePlaat collections (-145, -146), resembling the Gladhurst (-116) and Glen Craig (-98,-99) forms of gracilis. Thus representing my view of an archetype.
147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.
On our last day, Ian Ritchie kindly took us on a drive to territory which had looked quite visitor friendly from the top of the Zuurberg. On closer contact they are anything but so. On the Spitzkop, which is on the northern boundary of the farm, we found aristata (-141) growing on bluish shale in a situation favoured by H. cooperi var. cooperi. It was a little bigger than our previous collection, but smaller than plants we subsequently collected northwards on the road between Riebeek East and Jansenville (Paddafontain, MBB6899-155), which I refer to pringlei). Driving to the east of Spekboomberg we saw plants (MBB6914-156) similar to the Marx plants of DePlaat (-145) in profusion, some of them without the softer translucence of the Marx collection. The leaves were occasionally much flatter and ovate and distinctly reminiscent of cymbiformis. Daphne and I walked down the hill from that point seeing the plants for most of the descent. Ian in the meantime drove back and further to the south-east and then found very similar plants on a steep slope also facing south-east (MBB6915-157). These plants can also occur in large clusters.
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.
Daphne and I drove to Somerset East to reconnoitre the Bosberg and on the way back saw aristata again about 10km north-east of Kaboega (-143). We had looked at the Paddafontein (-155) plants on the way out and they quite obviously can be linked to aristata too. Unlike the other collections of aristata, however, the Paddafontein plants have a large robust inflorescence with many flowers open (usual in decipiens) as opposed to say the Commadagga (-128), aristata, but with fewer and stumpier leaves) dwarfs with only 8-10 flowers per stalk and one open at a time (thus more pilifera-like).
128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.
During our second visit, we again went to the top of the mountain at Wilgefontein, after Ian had shown us H. glauca in the Kaboegapoort itself. This population was not typical of the species and also more like the De Plaat plants. We revisited the site of -148 and -149. Both were in flower and on this occasion we found the grassy ecotype within about 75m of the stone-face plants. The latter had flowered but seed-set was very poor, as opposed to the grass element which had well-formed capsules. From there we went to the Dez Weekes’ slopes via a different route and collected (MBB6925-158) plants ranging from the same greenish cymbiformis-like plants of -156 to specimens which could be nothing else but typical of true cymbiformis. We saw the same plants again at the dam to the north-west of the homestead, and again on the south slopes (-159) behind the previous De Plaat collection (-151). We completed the stay with a visit to Soutkloof where we saw again the true aristata (-139), and also to the office of the Addo National Park on top of the Zuurberg. Here we saw specimens of H. cooperi var. pilifera from that vicinity, and similar to a collection by Ernst Van Jaarsveld from Oudekraal (I had seen these plants when I collected H. angustifolia var. baylissii there many years ago). On our visit to Oudekraal we stopped to the west of my previous visit and probably also west of where Ernst had collected. We found the cooperi-like plants (MBB6922-160) again growing among rocks in dense grassland. The plants had very pronounced reddish-lines in the leaves and this was evident in all the Kaboega collections.
158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.
Curiously a post-graduate botany student busy with a study of succulent endemism brought in a number of plants for identification. This is P. Desmett, and among his collections, two are relevant. One is PD2310-161 from Boplaas. This is north-east of Kirkwood where the Kaboega meets the Sundays River. The plant is a small spinose specimen which could relate to the arachnoidea-like (as von Poellnitz compared it) stiemiei. It could alternatively, and because of its colour and translucent patterning of the leaves, be more probably compared with H. decipiens var. minor represented by several MBB collections from Sapkamma (MBB6618-162, MBB6619-163, MBB6620-164) to the west. The other is PD2309-165 from the southern end of Kaboegapoort. It is the apparently puberulous-like element resembling -148, and also -153. Kobus Venter also collected and aristata-like plant from the Sundays River Poort (-73) which I think compares very favourably indeed with a large number of collections from afar afield as Redcliffe (north-east of Willowmore), the Baviaanskloof, Uniondale, down the Longkloof to Humansdorp and Hankey/Patensie. These are all collections which I have identified as H. cooperi var. gordoniana, and considered in the context of another paper.
161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
Discussion It is apparent to me that there are can be only two elements (species) of the sub-genus Haworthia present on Kaboega. These are from either of the geographical elements cymbiformis, aristata, gracilis and cooperi, and they are directly continuous. In cultivation it is apparent and obvious to me, that aristata from Spitzkop (-141) is mirrored by the gracilis-like -152, which is continuous with the more gracilis-like -151. This latter element leads through several collections to the cymbiformis-like plants in -150, -158 and -159. Similarly a very cooperi-like element in -148 is the apparent ecotype of the very gracilis-like -149. But -148 (and -165) must also be compared with -152 and to -153, which take us back to the gracilis-like archetypes. More significantly these seem to be the elements which best relate to the collections from Oudekraal, and with what occurs still further east at Zuurberg. These collections are considered to be H. cooperi var. pilifera.
There is no doubt that the Spitzkop aristata (-141) must be compared with -155 at Paddafontein and thus connecting aristata to the greater Jansenville area, and to the western elements of H. decipiens var. pringlei. There is the Waterford collection north of Lake Mentz (MBB6855-154) which is problematic as it does not have the opacity nor darker blue-green colouration of pringlei. It is better compared with xiphiophylla (or perhaps this is H. decipiens var. minor) in 72.2 south of Lake Mentz. It also bears a remarkable likeness to the Krom River collection of gracilis var. isabelllae (-39) as indicated above.
Conclusions I conclude that at Kaboega we have a situation where cooperi is excluded by the fact that the archetypal gracilis is represented by an advanced version of gracilis from which aristata and cymbiformis are extended. This pattern of identifications and classification true for one area, are not true for another. Already fully aware of the complex interaction between species like H. bolusii, H. cooperi and H. decipiens, and fast becoming even more aware of the extension of this complexity to H. gracilis, H. cymbiformis, H. arachnoidea var. xiphiophylla and even H. marumiana, I have to express conclusions very guardedly. Any classification of Haworthia will undoubtedly have tensions within it. It has been long apparent to me that sophisticated technology is unlikely to prove of much value in dealing with the nuance of variation between populations. If it is, it has going to have to first take into account the kind of variation one sees at the scale covered by this article. My belief was, and is now confirmed, that this is indeed the scale at which observation is now required. It can still be a lot closer. We did not spend sufficient time at Kaboega to explore the area thoroughly, and neither have we yet made any permanent record of our observations other than this report and accompanying illustrations. The point may now have been passed at which casual generalisation from a memory bank of images is possible. Extensive photographic and herbarium records are going to be essential to create a physical record which can be studied and manipulated. There is a series of eight mountain ranges from near the coast, with the Zuurbergs being the last of these in the north-east. If I calculate how long it would take to explore that area on the scale of our limited survey of Kaboega, I reckon on at least three years of continual search.
At this point I realise that the expectations of “Haworthia Revisited” are not going to be met. There are already snivels and meuls because there is no “data” in my revision. My experience tells me that this is not because the average reader would in fact take any cognisance of such data – but it is part of the illogical and faulty paradigm of modern “science” (“materialism”, the Theosophist would say). My conviction is that I have in my revision presented there a very comprehensive picture of the genus. This can definitely meet the time-worn wishful thought of the platitudinous foreword that “this book will stimulate/encourage/direct/guide further research”. Classification of Haworthia is not simply sorting a few single specimens as they are represented in collections or on herbarium sheets. It is trying to understand a complex system of closely interrelated and similar looking elements, as populations, which do not fit a classic and static image of a genus and so-many discrete species. My contention is that this is not only the case for Haworthia, but for many other genera too. ♦
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.144 a. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. arista, Chalumna. (H. cooperi var. doldii – ed.)144 b. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. aristata, Chalumna. (now H. cooperi var. doldii – ed.)145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.146. J.G.Marx in JDV91/15 H. cooperi var. cooperi, De Plaat, south aspect, Zuurberg.147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
M. B. Bayer, Cape Town. Ian Ritchie, Somerset East. ♦
The five papers in this book represent analyses and discussion of a large number of collections revolving principally around the problems of the Eastern Cape Haworthia species. The paper regarding H. mucronata does provide some insight into the overall problem. The citations by Von Poellnitz for varieties of H. altilinea and H. mucronata indicate just how difficult it is to differentiate between species such as H. bolusii and H. mucronata, from their respective geographic locations. In the case of H. cymbiformis var. incurvula; then in the case of H. cooperi, H. bolusii var. blackbeardiana, H. aristata and H. decipiens var. pringlei; then H. cooperi, H. gracilis and H. cymbiformis; and finally in the case of Kaboega; it is evident that a better solution can be sought.
During the course of the analyses, it became evident to me that a gracilis-like element is present in every one of the cases. It is less evident in the case of H. mucronata, because I have not presented the data and also because I have not collected sufficiently either. Certainly H. mucronata is closely allied to cooperi, cymbiformis, blackbeardiana and to decipiens. However, it is also very closely intertwined in H. arachnoidea and a lot more field observations are required to determine what its archetypal kinships may be. They may be the same as for cooperi and cymbiformis. Regardless whether or not this is so, the question still arises:- If gracilis is the archetype for so many species which the evidence leads me to suggest it is, what then is this element I call gracilis?
I have always had problems of clearly separating variants of cooperi and variants of gracilis particularly in the Baviaanskloof. Examination of the Middleton, Baviaanskranz (pringlei) and Ripon collections demonstrates the tension that exists in the classification. The two former collections seem to constitute an element which can be differentiated from blackbeardiana and from cooperi (represented mainly by its variety pilifera), albeit with difficulty. That is, it has more slender leaves than either of those species. Thus I must point out that cooperi is problematic because in my 1999 revision, the main geographic component is assigned to its varieties. The typical form of cooperi occupies a no-man’s-land between blackbeardiana and pilifera. A similar situation arises with the discarding of the name translucens for gracilis. The elements which I had previously included as typical for translucens, became the var. isabellae of gracilis. Gracilis itself came to occupy a relative no-man’s-land and possibly only truly represented at its origin at Hellspoort. These problems are discussed in some detail in Haworthia Revisited.
The fact that none of the elements discussed in the preceding papers form clear discontinuities, suggests that I need to seek a better way of expressing the situation and the problem. The Kaboega situation is particularly difficult, and the fact is that we have recognisable cymbiformis, aristata and gracilis var. isabellae, connected by an array of forms which seem to be archetypal of gracilis. At Pluto’s Vale, we have gracilis var. tenera and cymbiformis connected by a small array of a similar archetypal gracilis. In the Baviaanskloof, we have variants of gracilis, and variants of cooperi which are linked through cymbiformis-like picturata to transiens. But there do not there seem to be three discrete species any more than at Kaboega, or at Pluto’s Vale. The same is true of the Ripon (Somerset East) area. There, there are only two elements. In terms of my 1999 revision, one is cooperi (MBB6557) and the other is either blackbeardiana or pringlei (MBB6556). It cannot be both. Furthermore, these two elements seem to be ecotypes of the same species, and proven by their co-occurrence in a third collection (MBB6927) from a differing habitat.
I have suggested that a more technically orientated botanical revision would result in a reduction of the number of species.These previous papers suggest that such species reductions may even be necessary at the present level if we are to maintain coherent communication. The reference to a more technically botanical solution immediately gives me problems, because I do not from my experience see the adequatio to undertake such a task. I here try to summarise, distill and synthesise the conclusions arrived at in my other papers in my own way. The questions I ask are:-
Is there are common solution to the classification of the variants of Haworthia over the geographic areas of the middle Fish River, the upper Sundays River and the Longkloof (Couga River) and Baviaanskloof (lower Groot River and Baviaanskloof rivers).
Can any classification solution be offered which provides a better explanation for the variation and the continuities we see in the field?”.
Results The problem relating to Ripon, makes it quite obvious that we have cooperi var. cooperi, cooperi var. pilifera, cooperi var. dielsiana, bolusii var. blackbeardiana, decipiens var. pringlei and aristata involved in a common discussion.The problem at Kaboega is that we have cymbiformis, aristata and possibly gracilis as emergent elements from a core which could be gracilis. Similarly at Pluto’s Vale, tenera and incurvula emerge from a gracilis core. In the Baviaanskloof, there is a single core which leads either to gracilis, cooperi or cymbiformis variants. In nearly each case I have mentioned two key collections.These are JDV96/89 from Gladhurst, south of Adelaide, and MBB6603 from Glen Craig, north-east of Grahamstown. To this can be added the collection by Gerhard Marx in MBB6845 just east of Alicedale. These are all difficult to reconcile with my 1999 classification concepts of cooperi or gracilis. If we re-examine gracilis, we find that it is only truly represented at Hellspoort, north-east of Grahamstown. Kobus Venter collected there (JDV89/42) and again in 1988. I collected there together with him (in MBB6614) in 1966. Looking at these plants in cultivation and in the light of the classification problem, it becomes obvious that pringlei via particularly the Ripon collection MBB6556, is comparable with gracilis from Hellspoort. With this consideration in mind, it becomes evident that if the concept of cooperi is enlarged to incorporate gracilis, a radically new and more practical solution is presented. The element which I have called gracilis var. gracilis becomes cooperi var. cooperi.
The implications are as follows:
H. cooperi becomes a substantial element which absorbs all the varieties of the obsolete gracilis. The variety dielsiana is abandoned as a variant within the variety pilifera. Cooperi also absorbs pringlei from decipiens in relation only to the collection from the type locality viz. Scott in PRE8970 and two others mentioned in my revision viz. MBB&PVB6561, Baviaanskrans, Patryshoogte, and MBB&PVB6556 from Ripon, both cited under bolusii var. blackbeardiana. It should be noted that no specimen of pringlei was in fact deposited and a letter promising such a specimen occupies the herbarium file. My perception and understanding of the element is from two plants sent to J.D.Venter by Dr Reddi, and also from two plants obtained from Gerhard (J.G.) Marx, all from the ostensible type locality.
Thus cooperi would virtually comprise a superspecies which excludes cymbiformis and its varieties with some difficulty. It would also be difficult to exclude bolusii. Cooperi would in fact absorb the variety picturata of gracilis and could absorb transiens of cymbiformis. A further implication is the variety viridis of gracilis. The evidence suggests that it belongs with cooperi, but that it is interwoven with decipiens and perhaps with xyphiophylla. In this respect, there is not enough data to build a clearer picture of what occurs in the field. There is a similar problem with aristata. There can be little doubt that it is the geographic complement to gracilis and pringlei, and while it is distinctive at Kaboega, there is also the evidence there that it is continuous with cooperi and cymbiformis. There is also the question of the remaining specimens which are cited in my revision under pringlei. These are from the area around Jansenville, where we already know (Lootskloof) that bolusii var. bolusii intrudes, and where we also have a series of greenish plants (Mt. Steward area principally) with numerous erect spinose leaves rather green in colour. They differ from decipiens in having smaller narrower and elongate leaves. These elements need to be further researched and perhaps re-named. Aristata can be linked with these as it does bear a striking resemblance to the darker and bluer forms of decipiens viz. var. cyanea from south and west of Beaufort West.
Conclusion The first question can be answered with some confidence. There is a solution which can be offered for the portions of the river drainage systems mentioned above. This is to merge the concepts of cooperi var. cooperi, gracilis var. gracilis and part of decipiens var. pringlei.
The second question cannot be answered because of the scope and extent of the problem in relation to the available data. It is quite obvious that small scale collecting and observation is needed. From what has been done, it is very evident that decipiens, bolusii, cymbiformis, arachnoidea and mucronata are very poorly represented in terms of their actual variability. This is particularly true of decipiens. If one considers its geographic central role between mucronata and cooperi, it is predictable that research will disclose some very interesting variants. There are vast areas (in relation to the scale of exploration done at Pluto’s Vale and at Kaboega) which can be regarded as wholly unknown. Particularly void are the areas between Oudtshoorn and Willowmore, and the between Willowmore and Graaff Reinet and southwards to the Cockscomb. The many ranges of hills between the Longkloof and the Zuurberg must hold many populations too, while the mountains to the general north of Somerset East must do the same.
I have done quite a degree of detailed exploration in the south-western Cape which has exposed some extraordinary populations. Some of the variability and endless difference borders on the bizarre, without detracting from the beauty and fascination of the genus. Thus I do not think there is a classification system in the classic sense of dichotomy that characterises taxonomic botany. The variability within Haworthia appears to be highly reticulate and ductile. Many different solutions can possibly be provided which will all be influenced by the material available, and the form in which it is seen by the viewer.
Ironically, my definition of species is that they are systems which are genetically and morphologically continuous in space and time. In my revision I wrote that “I was trying to identify nodes in a complex interlinked web”. This may suggests that the nodes are more important than the matrix in which they occur. I also wrote “… when a variety is selected (described) the balance of variability is loaded towards the typical”. What happened with the recognition by myself of pilifera after I had in my previous Handbooks placed it in synonymy with cooperi, is that it suggested cooperi to have much more substance. Similarly, recognising varieties of translucens and abandoning the name in favour of gracilis, created a similar problem there. Varieties occur as nodes in the matrix of the parent species, just as species do in the matrix of the genus. At some point the discontinuities (in either case) may be obvious and perhaps absolute. I suggest that in the sub-genus Haworthia these continuities are largely obscure. It seems to me that we need to examine the herbarium specimens in terms of a sampling record. It is by now quite substantial, and should thus be considered as a very largely representative of the matrix of the genus. I have become increasingly confident that my classification based on that record, is a good one. Furthermore, that it is has a sufficiently predictive element to accommodate the greater number of new records that will be made.
While writing these papers, I have been carrying some ideas in the back of my mind. These relate to the species in the southern and south-western Cape. I have many new records from there as well, and I am aware of several collections which are otherwise outside of my ken. What I have seen suggests to me that it may be possible to identify the background matrix of an archetype such as I in this work suggest for the eastern Cape (viz. cooperi). ♦
Photos from a field trip to the farm Van Reenes Crest (half way between Swellendam & Heidelberg along the N2). See an earlier article in Haworthia Updates Vol. 7 Chapter 6.
I must explain why I wrote and why I write. Reading and writing are inextricably related. I observed Haworthias, I read about them, and I wrote about them. This was to organise and record what I had learned, to refute what I deem to be wrong and to seek verification of the new. I never did feel adequate to the task of formally revising Haworthia. The fact that I did so was because I was asked to. It was quite clear to me that however inadequate I was, there was nobody else who was going to do it any better. The work (Haworthia Revisited, 1999) was premature in the light of what really needs to be known if a good classification is going to emerge. It is a considerable disappointment to me that much of my writing has not served to guide other writers as I hoped it might. The forewords to Breuer’s two volumes by messrs. Ihlenfeldt and Smith confound my sense of what is true that I truly wonder if there are readers who are anxious to share that same ideal. Sometimes I wonder if my writing has served any greater purpose for which it may have been intended other than to help me organise my own ideas and activities.
Species identification in Haworthia is difficult. This is not a new statement, but simply now a self-evident truth which is becoming increasingly so as I extend my field experience. Recently someone commented on the odd flowering time of H. pulchella var. globifera, commenting that the flower was even unusual. The species pulchella was not yet showing signs of flowering, whereas the odd H. monticola var. asema was budding. The similarities of that variety to globifera in terms of leaf colour and shape, proliferation and flowering time are striking. I later looked more closely at the flower of globifera and found that, in the clones I had available, there was no visible difference whatsoever, between the flowers of globifera and H. cymbiformis var. incurvula from Pluto’s Vale. My collections of the latter were also not all in flower, and quite clearly flowering time (useful as it is) is no more an absolute than can be stated in Stephen Gould’s paraphrased … “The strongest statement that one can possibly make in biological science is ‘hardly ever’.” What to say of the similarity of the flowers in two elements which could hardly be further apart in the sense that classification is applied in the genus.
There is an almost universal conviction among laymen that taxonomists are a quarrelsome lot who do no good but argue about names, and change them in the process. This is unfortunately largely a justified observation, but the justification arises from an inexcusable degree of nescience on the part of both botanist/taxonomist and involved observer. As my essays in recent years have substantiated, plant classification seems to offer the lowest level of intellectual entry to science writing. Anyone, without having any qualification or degree of competence, has access to plant description. Nothing more is needed than an existing template and some kind soul to write some Latin words for the required synoptic sentence. Without any kind of overview or experience of biological systems and thus a more general feeling for classificatory criteria, anybody may step forward and claim authority.
These claims cannot be questioned or challenged by the average or even above average botanist, and not even by those odd few who specialise in classification. The adage “experience is what you have acquired after the need has passed” is too true for comfort. Too few botanists know enough outside of their specialised fields to question what is happening in other families and genera. They are too busy defending the integrity of their own beliefs and systems. Publications in prestigious, rigorously refereed botanical journals fail to neuter some really poor science writing. These observations are all substantiated in the following essays or elsewhere.
In Haworthia, the situation is not that botanists have failed to involve themselves. They have. The results are no better than that of the foolhardy amateur. The consequence is that we now have a fair body of interested growers and collectors who cannot know what is true and what is not. These people can make any derogatory statement about taxonomy that they wish to make, with the confidence that they may be right. Some equally ignorant botanist will step forward to agree and may blithely enter the writing arena too. Mutual backslapping is the order of the day with scant regard for the facts and for what might be true.
The question can again be asked … “Is Haworthia a problem genus?” I wrote an article entitled “Haworthia as a problem genus”, which was published in the Cactus and Succulent Journal (6:251,1971). After so many years and so many written words, I am constrained to ask myself how I see the position now.
I recently wrote an article discussing the question “Is classification science or art?”. This was published in Asklepios (77:3, 1999). Then I also wrote a collection of essays published as “Thoughts on Haworthia”, in which I try to show that Haworthia is not peculiarly difficult. Rather, that the problem is how we as writers and readers perceive our respective roles, and how much we know about plants and their plasticity.
My conclusion is that individually and collectively we may not have a very clear idea of what we are doing or trying to do. If one looks really objectively at the issue of classification and all that is written about plants and the names we use for them, it must be apparent that we are not being very sensible.
The binomial naming system in which groups of similar plants are gathered into genera, with subgroups of similar plants as species, is all well and good. But do we understand the actual nature of these groups. If we look at dictionary definitions of the terms genus and species, we find them wholly inadequate.
What appears to be the situation is that each of us has a concept and construct in our personal consciousness of what we regard as a species. Whether or not this is the same for all of us, and if our worded definition for “species” is the same wherever it is used, does not seem to matter. Generally we may use a word with a fairly liberal interpretation of what it may actually mean, in the framework of a limited vocabulary and knowledge.
One of my essays in this volume is an analysis of the book “Rock of Ages”, written by Stephen Jay Gould. Here I try to show that we are experiencing problems with the fullness of our lives. We as human beings are not doing in our limited spheres of consciousness, what we ought to be doing if we wish to understand creation – of which Haworthia is a very tiny part. My wish thus in submitting these essays for publication is that some silent majority does exist and that my work will in a small way perhaps satisfy their own inner wish to know what is true and real. A deeper wish is that someone better equipped will pick up from where I will leave off.
Acknowledgement I acknowledge that a better and more satisfying discussion and explanation of Haworthia is desired. Unfortunately the competences, interest and opportunity do not seem to be vested in anyone else that I would gladly submit to. I am extremely grateful to the many people who have simply taken me on trust. There are many individuals who could expect to find themselves listed here as a simply matter of fact rather than a measure of personal satisfaction. Most of them are unlikely to ever see this publication and I thus record my enormous appreciation to a faceless multitude for kindnesses and consents that made access to the plants possible. I find it curious that Conservation bodies demand to be acknowledged when their contribution is largely passive and permit aquisition a demeaning process. I am grateful to the law enforcement officer who in a kind aside commented that their regulations were not for people like my self. I think it is necessary that Conservation recognizes a constitutional right of individuals to engage themselves in study of nature, and not condescension on their part to allow it. I have tried to comply with Conservation legalities and have certainly done so in respect of intent. I am not sure that authorities are achieving what they may aspire to as I have seen too much degradation of this great natural resource of ours and alienation of people to be convinced they are properly directed or motivated.
I would particularly like to thank Kevin Belmont for his interest in the manuscript, and to Harry Mays for much more than just the labour of bringing it to press. We have had a long association in which he has never been less than helpful and consistent in his view that the platform should be accessible to all. Bob Kent, Steven Hammer, and Gerhard Marx have patiently endured my ranting. Other contributors are acknowledged at various places in the essays. I truly regret that Kobus Venter has not had the time to participate as fully as he would have liked, as the work would have been better for the benefit of his wisdom, interactive personal skills and interest. ♦
Introduction Haworthia Revisited was drafted in 1996, and since then the first author has undertaken a number of field excursions in an attempt to clarify uncertainties. The putative nature of species of Haworthia as recognised by Bayer (listed in Haworthia Revisited, Umdaus 1999) and the importance he attached to geographic distribution are stressed in all his publications. This is because these so-called species seem to vary continuously with one another in that context of geography. Classification seeks to portray relationships and origins. Hence when a species has been recognised, a cognitive attempt has been made to speculate on phylogenetics, where distribution must be significant. In the case of Haworthia floribunda this proves rather difficult, and this article is a discussion of the relationship of this species to its possible relatives. The point we do make is that the Linnaean binomial system, as well as cladistic methods, seem neither to deal with nor portray the problem of reticulate relationships. In other words, the nomenclatural system and the way we classify plants and analyse their relationships assumes linear dichotomy in those relationships.
Considerations H. floribunda was described by von Poellnitz in 1938. It was preceded by H. parksiana in 1936. Other species which need to be considered are H. chloracantha Haw. (1821), H. variegata Bol. (1929), H. magnifica VPoelln. (1933) and H. maraisii VPoelln. (1935). Each of these species, although H. parksiana to a lesser degree, has complex variability within assignable geographic space. The difference with H. floribunda is that it is peripheral to all of them.
Bayer, in his Haworthia Handbook (1976), writes of H. floribunda ” … It is not certain what the relationship is between this species and H. chloracantha var. denticulifera which is found north and south of Albertinia on the west of its distribution range. Thus H. chloracantha abuts on H. floribunda and may be geographically continuous with it.” In the New Handbook (1982) Bayer writes “There is a known population north of Albertinia in which the plants have more and shorter leaves, as well as another similar population near Gouritzmond. These two populations may suggest an affinity with H. chloracantha (here an unpublished comment by A.E Speechley is added which probably has its origins in Bayer’s note of 1976, or in private communication between the two persons). There may be such a relationship, but it seems likely that H. floribunda and H. parksiana are in fact related. They both tend to grow well‑shaded and in moss and lichen.”
The problem is not resolved in Bayer’s more recent revision (1999). He does recognise three varieties of H. floribunda. These are:
Fig. 01. MBB158 H. floribunda. Type locality, N Heidelberg.
a. the typical one comprising a single population north of Heidelberg (Fig.1 MBB158).
b. a similar single population near Swellendam representing the var. major (Figs.2a & b MBB6859).
Fig. 02a. MBB6859 H. floribunda var major. S Swellendam.Fig. 02b. MBB6859 H. floribunda var major. S. Swellendam.Fig. 03b. JDV89-17 H. floribunda var dentata. NE Riversdale.Fig. 04. MBB2311 H. floribunda var dentata. N Albertinia.
c. the remainder, the var. dentata (Figs.3a & b JDV89/17).
In the discussion of H. chloracantha, it is also stated that a population north of Albertinia (i.e. Draaihoek, Fig.4 MBB2311) considered as chloracantha in 1976, considered non-commitally as chloracantha in 1982; ” … includes plants which resemble H. parksiana and it may best be related to H. floribunda“. This collection is cited under H. floribunda var. dentata. Thus there is a degree of uncertainty for which the author offers an apology, and here attempt to summarise the situation a little more comprehensively.
Fig. 04. MBB2311 H. floribunda var dentata. N Albertinia.
There are some red herrings in various comments made by Bayer. These include reference on p48 of Haworthia Revisited to the lower Gouritz population (Fig.5 MBB5460) “previously being assigned to H. floribunda“, possibly referring to a Smith identification on a photograph and cited under H. chloracantha var. denticulifera. There is a similar population nearer Albertinia (Fig.6 MBBsn). Also there is a citation of a specimen from the Duiwenhoks Causeway (Muller-Doblies 89/098) under H. chloracantha var. denticulifera, which may be the same population sampled by Venter and Esterhuizen (Figs.7a & b JDV92/31) from a site named as Dassieklip also on the Duiwenhoks River (the geographical grid references differ and these may be incorrect), but cited under H. floribunda var. dentata.
Fig. 05. MBB5460 H. chloracantha var denticulifera. NW Gouritzmond.Fig. 06. MBBsn H. chloracantha var denticulifera. S Albertinia.Fig. 07a. JDV92-31 H. floribunda var dentata. Dassieklip, S Heidelberg.Fig. 07b. JDV92-31 H. floribunda var dentata. Dassieklip, S Heidelberg.
Haworthia parksiana is known from at least five different sites of which two are recorded by Mr Jan Vlok, viz. at Outeniqua Siding, and north-east of Brandwacht, Mossel Bay. No herbarium specimens of these are known to have been lodged. J.D Venter has commented that parksiana-like seedlings were grown from H. magnifica seed (parent plants ex near Heidelberg). It is doubtful if this can be used to support a claim that this is the origin or relationship of the two species. Bayer has also grown small stunted forms of magnifica from seed of an H. magnifica population south of Riversdale. Such dwarfed individuals have been observed among seedlings derived from other populations. The connection of H. magnifica to the Mossel Bay area is just too tenuous. At Albertinia itself we have the variation to H. magnifica vars. dekenahii and splendens, while to the south west we have the var. acuminata. This is assuming that Bayer is correct in making these combinations which are known to be complicated by consideration (among many) of H. emelyae north of the Langeberg mountains.
H. chloracantha is treated in Haworthia Revisited, as three varieties. The typical variety is considered to consist of the known single population north of Herbertsdale (Bayer in KG411/75). However, Paul Vorster drew my attention to a population at the Wolwedans Dam north of Great Brak which is almost identical and which invalidates varietal distinction defined by geographical considerations (Fig.8 MBB7425). A specimen of this collected by C Burgers is also cited for the Gouritz Gorge where it exits the Langeberg Mountains. P.V.Bruyns and E.A.van Jaarsveld have also made collection from further north in the same gorge, which are uncited. These three populations cannot be categorically included in the typical variety and could be just as at home in the var. denticulifera. The only consideration is probably size and the greenish colour – as opposed to the usually smaller and purplish-green of the var. denticulifera. The var. sub-glauca also does not have a clear geographical base in terms of difference or distribution range, and may just be a much localised ecotype in the vicinity of Great Brak. It occurs close to the Wolwedans population both to the south and to the west. The recognition of these varieties makes it a little difficult also to include the Albertinia and Cooper Siding populations as cited in Haworthia Revisited under H. chloracantha as they are so closely similar, and located, to H. floribunda. The geographical considerations, however, argue for their inclusion.
Fig. 08. MBB7425 H. chloracantha var chloracantha. Wolwedans, Great Brak. MBB7425
The Great Brak/Mossel Bay area contains some endemic species which suggest the operation of a mechanism which could have isolated the vegetation of that area at some time in the past. H. parksiana, H. kingiana, H. pygmaea, H. chloracantha and species like Euphorbia bayeri and Duvalia immaculata suggest this. The Langeberg mountains are an east/west barrier separating the more arid Little Karoo and its succulent Karoid vegetation from the Southern Cape where Renosterveld and Karoid Valley Bushveld are more strongly represented. The Gouritz River Gorge may have been, or is, a similar north/south divide. Westwards, the Swellendam area seems to provide a vegetation interval that H. turgida seems to bridge with difficulty, and which also marks some kind of a break for the H. magnifica/H. maraisii complex. This divide may be a function of inadequate collecting or the lack of adequate habitat. The Breede River valley may, like the Gouritz, present a north/south barrier. It is the home for H. venosa subsp. venosa and also for an endemic asclepiad Stapeliopsis stayneri. West of Swellendam is also the start of the Worcester Robertson Karoo which is vegetatively much closer to the Little Karoo than is the eastern part of the Southern Cape. This area also has its own Haworthia endemics or near-endemics.
The physical commonalities of the three species floribunda, chloracantha and parksiana, are that the leaves are in a stemless rosette, having from 20-40 leaves per rossete. The lowest numbers are in floribunda with parksiana and chloracantha generally having more. The leaves are firm and slightly scabrid. H. chloracantha is more spinose than the other two species and the leaves are triangular in cross-section with a conspicuous keel. In H. parksiana the leaves tend to be short and sharply recurved. In H. floribunda the leaves are more erect, although spreading. Characteristically the leaves are twisted and there is no keel, so the leaf is more effectively strap-shaped particularly towards the apex. The leaf-tip is also rounded with a short point. In H. chloracantha, the leaves are erect and spreading, not twisted. They are keeled and thus triangular in cross section. The leaf tip is attenuated and pointed. The leaf surfaces of the three species are variable. Both H. floribunda and H. parksiana can exhibit quite tuberculate surfaces, although parksiana never has the relatively glabrous surface that may occur in either chloracantha or floribunda. Colouration is also variable across the three species, although parksiana does not occur in the lighter colours that the other two species may exhibit. A hatched pattern of the under-leaf surface may appear. All three species make offsets although parksiana is slower and more reluctant to do so – an observation which may just be peculiar to the individual grower and the clones he has under his conditions.
New material Regarding H. parksiana, the only new records are those of Jan Vlok mentioned above. The writers assume that the identifications are correct and that they establish the discrete identity of that species. Regarding H. chloracantha, there is only one significant new collection. This is one of the var. denticulifera made by Bayer, Kent and Venter. It was made at the same time as a visit to the site of the typical var. chloracantha north of Herbertsdale. Here the plants are on a very steep clay slope (in fact recent rains had caused massive mud-slides). The plants are quite large and form immense chlorotic-green clumps. We were at Herbertsdale to verify a collection of H. pygmaea made by Ernst van Jaarsveld in the same vicinity. This is a short distance away on a steep conglomerate slope. H. pygmaea was present on the dry northern edge. But in the moss and lichen of the cooler eastern overhang were the small single plants of H. chloracantha (Figs.9a, b & c JDV97/136). These were so evidently different from the typical form that we accepted (with reservation) the identification as H. floribunda. One reason for such identification is simply one of informal communication. A similar “floribunda“-like plant grows with H. pygmaea and H. parksiana virtually within Great Brak Township (Fig.10 JBouwer sn). This was collected and referred to as H. floribunda by both messrs. H. Gie and J. Bouwer. It is possible, and probable, that it is in fact H. chloracantha var. denticulifera, as is the new population JDV97/136.
Fig. 09a. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 09b. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 09c. JDV97-136 H. chloracantha var denticulifera. Herbertsdale.Fig. 10. JBouwer sn H. chloracantha var denticulifera. Great Brak township.
The Dassieklip population (Fig.7) remains somewhat of an enigma. A good comparison can even be made with collection of H. monticola from Trompetters Poort, north of Willowmore, as the plants also have an apparently smoother epidermis than appears evident in chloracantha. This is a highly improbable relationship, but it is necessary to make this comparison in view of what has been speculated elsewhere about the possibility of a continuation between H. chloracantha, H. variegata and H. monticola (H. divergens Bayer!). This speculation has been by Bayer and is not significant enough to cite in detail. What is important is that the Dassieklip population may in fact be closer to H. variegata. Hence its citation under both H. chloracantha and H. floribunda is unavoidably misleading. However, that can only be considered in a detailed evaluation of H. variegata in its relationship to the coastal limestones and re-occurrence in shales both west and south of Swellendam.
The newest finds relating to H. floribunda are in the broad area southwest of Heidelberg, and west of Bredasdorp. However, there is one new record by E. Aslander of plants from northeast of Albertinia (Figs.11a, b & c EA1238) in which plants very clearly have the characteristics of H. floribunda. There are also individuals with the leaves of H. chloracantha. There is another collection by Aslander from Snymanskraal west of Albertinia (Figs.12a & b JDV92/2) which resembles the Draaihoek sample and thus may also be H. chloracantha var. denticulifera. A further collection by J.D. Venter is midway between this collection and Riversdale (Figs.13a, b, & c JDV93/56), and is clearly H. floribunda. A feature of this collection is the striated rugosity of the leaf surface in some clones and also the occurrence of short, almost terete, leaves that are evident in a clone of H. parksiana in a JDV collection.
Fig. 13c. H. floribunda var dentata. Plattekop, E Riversdale.Fig. 15a. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 11a. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 11b. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 11c. EA1238 H. floribunda var dentata. NE Albertinia.Fig. 12a. JDV99-1 H. floriibunda var dentata. Snymanskraal, E Riversdale.Fig. 12b. JDV99-1 H. floribunda var dentata. Snymanskraal, E Riversdale.Fig. 13a. JDV93-56 H. floribunda var dentata. Plattekop, E Riversdale.Fig. 13b. JDV99-1 H. floribunda var dentata. Plattekop, E Riversdale.
Collections from the western areas include:- from a population reported by E. Esterhuizen on the farm Koppies, southeast of Swellendam (Figs.14a, b & c MBB6879) where the plants tend to H. maraisii; from slightly to the east and south of this at Oudekraal (Figs.15a, b, c & d MBB6881) the plants are very like H. floribunda var. dentata as it occurs at Buffeljachts and at the Bontebok Park south of Swellendam. Esterhuizen commented on the appearance of the twisted and flattened leaf-end of H. maraisii north of Bredasdorp. This is confirmed in populations at Napky (MBB7030) and at Adoonskop (Figs.16a, b & c MBB6640) Adoonskop (maraisii) where the plants do indeed look like very robust forms of floribunda. The same characteristic twisted and rounded leaf-tip is evident in maraisii at Napier (Figs.17a, B & c MBB6973) and especially in the seedlings.
Fig. 14a. MBB6879 H. floribunda var dentata. Koppies, Heidelberg.Fig. 14b. MBB6879 H. floribunda var dentata. Koppies, HeidelbergFig. 14c. MBB6879 H. floribunda var dentata. Koppies, Heidelberg.Fig. 15a. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15b. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15c. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 15d. MBB6881 H. floribunda var dentata. S Oudekraal, Heidelberg.Fig. 16a. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 16b. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 16c. MBB6640 H. maraisii var maraisii. Adoonskop, N Bredasdorp.Fig. 17a. MBB6973 H. maraisii var maraisii. N Napier.Fig. 17b. MBB6973 H. maraisii var maraisii. N Napier.Fig. 17c. MBB6973 H. maraisii var maraisii= floribunda var dentata! N Napier.
Several other collections confound the picture completely either because of intrinsic variability or because they make it difficult to uphold any geographical recognition of variation. Firstly there is a collection attributable to P.V. Bruyns, north of DeHoop (Figs.18a, b & c MBB6539) which has strong resemblance to H. magnifica var. atrofusca.
Fig. 18a. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.Fig. 18b. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.Fig. 18c. MBB6539 H. maraisii var atrofusca! Tarentaal, N DeHoop.
Secondly there is a series of populations from the northwestern end of the Potberg, south of Swellendam and east of Bredasdorp. These are:- Juliesfontein (Figs.19a, b, c & d MBB6882); Brakfontein (Figs.20a, b, c, d & e MBB6886); northern Potberg slopes (Figs.21a, b & c MBB6889); and north of Brakfontein (Figs.22a & b MBB6890). These complement collections Burgers 2506, Bayer in KG35/70 and Bruyns in KG49/76 cited in Haworthia Revisited under H. maraisii, as well as MBB(PVB)6544 (Figs.23a & b), and MBB6545 (Figs.24a & b) cited under H. heidelbergensis var. scabra.
Fig. 19a. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19b. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19c. MBB6882 H. floribunda var dentata. W Juliesfontein, S Swellendam.Fig. 19d. MBB6882 H. floribunda var dentata! NW Potberg, S Swellendam.Fig. 20a. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20b. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20c. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20d. MBB6886 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 20e. MBB6882 H. floribunda var dentata. Brakfontein, S Swellendam.Fig. 21a. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 21b. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 21c. MBB6889 H. floribunda var dentata. NW Potberg.Fig. 22a. MBB6890 H. heidelbergensis var scabra! Witklipkop, S Swellendam.Fig. 22b. MBB6890 H. heidelbergensis var scabra! Witklipkop, S Swellendam.Fig.23a. MBB6544 H. heidelbergensis var scabra NW Potberg.Fig. 23b. MBB6544 H. heidelbergensisv var scabra NW Potberg.Fig. 24a. MBB6545 H. heidelbergensis var scabra! Juliesfontein, Potberg.Fig. 24b. MBB6545 H. heidelbergensis var scabra! Juliesfontein, Potberg.
The influence of other species such as H. mirabilis, H. variegata and H. serrata are in evidence. It can be noted that KG35/70 was of very small plants at Juliesfontein. Returning to the same site 30 years later I could not find these small plants at the original site, and instead found the bigger plants of MBB6882 a short distance away. There are three more collections to be considered:- by Denis DeKok near Swellendam (Figs25a, b & c MBB6644), about 10km west of Swellendam (MBB6861), and from the farm Rondeheuwel south of Stormsvlei (Figs.26a, b & c MBB6882, Bayer in KG326/71). This latter collection has previously been reported (Haworthia Handbook 1976) under one of the populations intermediate with H. mirabilis, and is cited in Haworthia Revisited under H. maraisii. These populations, as well as that southwest of Heidelberg (KG107/74 cited under H. magnifica (Figs.27a,b & c MBB6663, Bayer in KG107/74) confound the issue enormously as we actually have four populations which cannot with confidence be allied with either H. magnifica, H. maraisii or with H. mirabilis. The reality is that neither H. heidelbergensis nor H. floribunda can be excluded from the consideration of these populations. The case for each “species” needs to be dealt with separately. In this case, H. floribunda is reflected in the nature of the leaf shapes. In all the four latter populations given, there are individuals which have the same characteristic leaf shape of H. floribunda, although not necessarily the elongated strap-like leaves of the typical form. In addition to this, seedlings of many different collections of H. magnifica (Kweekkraal, Figs.28a & b MBB6817 – and the reader should refer to the chapter where interplay between H. magnifica and H. floribunda is reported), H. maraisii, H. heidelbergensis, H. mirabilis (Goudini, Caledon, Fig.29 MBB6537), and even H. mutica (Hasiesdrift, Fig.30 MBB6982), have the floribunda-like shape evident in the young leaves. Kobus Venter was especially struck by this phenomenon while photographing H. mirabilis on the Bromberg Mountain near Stormsvlei. These rounded obtuse leaf-tips are maintained until at least the three to five-leaf stages and then largely disappear. Thus there seems to be a distinct sign that the leaf-type is juvenile and that H. floribunda represents a “species” with retained juvenile characters. This may extend to the fact that the seedlings of this species also seem to remain slightly distichous for longer than is the case in other species. In the case of H. floribunda var. major from southwest of Swellendam, the leaves in some individuals may be fully pointed and triangular in cross-section and thus more closely resemble plants in the population of H. variegata var. hemicrypta west of Swellendam. This is the same kind of variation found in the populations about Albertinia, where the speculated differentiation to H. chloracantha occurs.
Fig. 25a. MBB6644 H. maraisii=mirabilis! SW Swellendam.Fig. 25b. MBB6644 H. maraisii=mirabilis! SW Swellendam.Fig. 26a. MBB6862 H. maraisii=mirabilis. Rondeheuwel, Stormsvlei.Fig. 26b. MBB6862 H. maraisii=mirabilis. Rondeheuwel, Stormsvlei.Fig. 27. MBB6644 H. maraisii=mirabilis. SW Swellendam.Fig. 27a. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 27b. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 27c. MBB6663 H. magnifica var magnifica. SW Heidelberg.Fig. 28a. MBB6817 H. magnifica var atrofusca. Kweekkraal, W Riversdale.Fig. 28b. MBB6817 H. magnifica var atrofusca. Kweekkraal, W Riversdale.Fig. 29. MBB6537 H. mirabilis var triebneriana. Goudini, Caledon.Fig. 30. MBB6982 H. mutica. Hasiesdrift, N Bredasdorp.
Flower morphology and flowering time does not appear to offer any solutions. As it is, the flowers have so far been shown in Haworthia to be useful only in so far as recognising the sub-genera is concerned. It can be shown that even at that level, the distinction causes problems for botanists. Most of the species discussed above flower in late summer. While the epithet ‘floribunda‘ was chosen to suggest many-flowered, this is true for other populations considered to be H. magnifica.
Conclusion The circumscription of H. floribunda remains obscure and the situation is in fact exacerbated by new samples which indicate the labyrynthine relationships with several species previously excluded from debate. H. floribunda may have juvenile characters. It does have distinctive populations in a recognisable geographic area. It does not directly share habitat with any species although it does occur very near to H. turgida (north of Heidelberg, where hybrids are also recorded). Where the chloracantha-like equivalent occurs with H. pygmaea var. argenteo-maculosa at Cooper Siding, hybridisation is also evident. It occurs very near to H. magnifica var. atrofusca northwest of Riversdale, growing on a cool southern slope as opposed to the latter on a hotter north slope. The situation northwest of the Potberg, where as many as seven otherwise apparently discrete species need to be included in the discussion, will be very difficult to explain.
It should be noted in closing that the Heidelberg population MBB6663 (as KG107/74) was instrumental in the initial decision by Bayer in 1976 when H. maraisii and H. magnifica were treated as one under H. maraisii. This was repeated in 1982 when H. magnifica was given its chronological priority over H. maraisii. In Haworthia Revisited these two elements are separated as discrete species and the Heidelberg collection is cited under H. magnifica. This decision was made rather to accommodate the varieties which are attached to them. Thus the additional three collections noted in connection with it viz. MBB6860, MBB6861 and MBB6862 all fall within this same indecisive category.
M.B. Bayer, Cape Town, South Africa R.W. Kent, Poway, California
Introduction After writing Haworthia Revisited in 1996, I became aware of just how inadequate readers seem to be to the task of assimilating all the available literature on Haworthia, in the botanical and intellectual climate in which we live. It seems as though the more information we have the more confused we become. In order to generate the material needed to disprove or fortify my classification hypothesis, I have spent a further considerable amount of time in the field and in cultivating plants from seed. Unfortunately the editorial support and speed of publication has not kept pace with my own effort and much of my writing and my evidence is still in manuscript form. This short essay was therefore to put forward only a little more evidence to show just how complex plant species are – not necessarily only in Haworthia.
In my first Handbook (1976), I anticipated H. maraisii and H. magnifica to be separate species – the former west of, and the latter east of Heidelberg, Cape. In the second book (1982), I felt that I was dealing with a single species and referred to a population just west of Heidelberg as H. magnifica var. magnifica (instead of as H. maraisii). I did this transposition quite deliberately in order to suggest that the distinction between the two species was very arbitrary. In my later revision (1999) I separated H. magnifica and H. maraisii again. The rationalisation is given there and I just need to explain that I thought this was a better way of communicating the nature of the variable populations attached to each of two elements. It was then evident that populations relevant to magnifica and maraisii were proving to be more radically different than a single species hypothesis could comfortably accommodate. Also new evidence was accumulating for the nature of:
1. H. heidelbergensis, which was not even included in the previous debates. 2. H. floribunda, also proving to be more variable than predicted. 3. H. mirabilis.
The overall relationship of these three species is far closer to both H. magnifica and H. maraisii than even my original conservative approach suggests.
In the essay which deals with H. floribunda I make two points particularly relevant to this article:
1. The geographic facts, viz..”Westwards (from Gouritz River), the Swellendam area seems to provide a vegetation interval which H. turgida seems to bridge with difficulty, and which also marks some kind of a break for the H. magnifica/H. maraisii complex. This divide may be a function of inadequate collecting or the lack of adequate habitat”.
2. The populations known to me. To quote from the floribunda manuscript “There are three more collections to be considered:- by Denis DeKok near Swellendam (Figs.25a, b & c MBB6644), about 10km west of Swellendam (MBB6861 not illustrated), and from the farm Rondeheuwel south of Stormsvlei (Figs.26a, b & c MBB6882, (Bayer in KG326/71). This latter collection has previously been reported (Haworthia Handbook 1976) under one of the populations intermediate with H. mirabilis, and is cited in Haworthia Revisited under H. maraisii. These populations, as well as that southwest of Heidelberg (Figs27a, b & c MBB6663, Bayer in KG107/74 – cited under H. magnifica, Bayer, 1999) confound the issue enormously. Thus we actually have four populations which cannot with confidence be allied with either H. magnifica, H. maraisii or with H. mirabilis. The reality is that neither H. heidelbergensis nor H. floribunda can be excluded from the consideration of these populations. The case for each “species” needs to be dealt with separately.”
Primarily illustrated here are plants from the population MBB6644 which occurs west of Swellendam. The population is not remarkable for the variation observable there among the individual plants, and I have taken it to demonstrate that there was, and is, predictive value in my classification hypotheses. Much of my recent writing has been directed at the very weak concepts and perceptions that appear in our general understanding of “species” and their variability. This has an impact on individual credibility, plausibility and the truth with which we examine our human condition and pursue our hobbies.
Results The photographs depicted are all taken at virtually the same distance so that the size of the plants is relatively correct; with the largest plants being near 90mm diam. (The pots are 90mm square.)
The first illustration depicts a conventional form of H. maraisii from a population from north of McGregor Fig. 1 MBB6646 H. maraisii var. maraisii). The species was first described from Stormsvlei where the plants are a little larger and more tuberculate than these pictured. Actually it has become difficult to say just where this species stops and starts, and this will become obvious. There is a degree of translucence in the leaves and there is a conspicuous vein down the centre of the leaf. This is by no means a ‘character’ for the species, and an article could be written about the variation in venation as well as about this single population (MBB6646) too.
The Swellendam plants have been named as H. maraisii=mirabilis in the preceding chapter and are illustrated as follows:
Fig.2, 3 and 4 are the clones 3, 8 and 17 of MBB6644. The surfaces are a little less tuberculate than the McGregor plant shown, but the only significant difference is actually that they are slightly bigger plants, the spination is more obvious and the leaves tend to be fewer and more erect. These are also by no means diagnostic characters. There is little translucence and the venation is inconspicuous. The identification of the population as H. maraisii thus seems obvious.
However, clones 2 and 4 illustrate a narrowing of the leaf (Figs.5 & 6 MBB6644). Clone 15 is a small plant also with smaller narrower leaves (Fig. 7 MBB6644). Were its leaves more erect instead of so recurved, it could be perhaps be taken to be H. heidelbergensis var. scabra.
Clones 1 and 3 (Figs.8 & 9) of MBB6663 are from west of Heidelberg and would easily be lost among the Swellendam plants if unlabeled. In the preceding chapter the population is identified as H. magnifica var magnifica. Clone 2 of this population (Fig.10 MBB6663) is a plant with more erect leaves – but …
Clones 7, 10, 11, 14 and 16 of MBB6644 (Figs.11 to 15) have a similar narrowing and erect bearing of the leaves, not to say that the plants are otherwise identical. This condition of the more erect leaves seems to be the more general one in both populations, MBB6644 and MBB6663. This is almost the crux of the problem in the classification of Haworthia, and in my experience it is not a problem unique to Haworthia. In all of biology there is this variability that requires statistical method to establish what “average” is. When one is dealing with different growing condition, and the nuances of texture, colour, shape, leaf recurvature and number, it is in fact not possible to generate such a “mean”.
Clones 9 and 13 of MBB6644 (Figs.16 & 17) are unusual in either H. maraisii or H. magnifica. The leaves tend to narrow quite considerably, but the leaves can also be unusually narrow and erect in populations of H. maraisii as in a population west of Robertson (Fig.18 MBB6647.8).
I include an illustration of a plant from a population of H. floribunda var. dentata from south of Swellendam (Fig. 19 MBB6881), – a population that tends to link floribunda with either maraisii or heidelbergensis to expose the reality that the link extends to H. mirabilis. (Apparently in a Dutch journal a writer has suggested that H. floribunda var. dentata is a form between H. floribunda and H. chlorocantha var. subglauca! This would be a complete misrepresentation of my classification hypothesis which has been fully explained and there is no reason for such a mistake. The var. dentata is represented by many populations and Chapter 3 details the possible relationship of the species floribunda and chlorocantha without any frivolity about the varieties and forms.)
Fig. 19. MBB6881.6 H. floribunda var dentata. Oudekraal, Heidelberg.
Also illustrated is a plant from a population very close (NW Kweekkraal), both geographically and in appearance, to H. magnifica var. atrofusca west of Riversdale (Fig.20 MBB6817). I have included it here because general colouration and texture are the same and it evidences the typical round-tipped leaves of floribunda which occurs so frequently in maraisii and magnifica (see older leaves in Fig.2 or Fig.8).
Fig. 20. MBB6817.1 H. magnifica var atrofusca. NW Kweekkraal, Riversdale.
Clones 1, 5 and 18 of MBB6644 (Figs.21, 22 & 23) are small rather nondescript specimens, while is clone 64 of MBB6639 (Fig.24) is a plant of H. mirabilis var. sublineata from south of Bredasdorp. These last four illustrations depict a series of smaller plants which bear a resemblance to H. heidelbergensis. The difference in the latter is the lighter green colouration, slightly more translucence and more conspicuous spination. These are significant in the context of these collections and these photographs. However, put into the context of all the populations of H. heidelbergensis, H. floribunda, H. mirabilis, H. maraisii, H. magnifica, also H. emelyae and the total variability among these, it is impossible to stipulate difference.
The four problem populations, MBB6644, MBB6861, MBB6882 and MBB6663 are not as distinctive as suggested by the way in which I have specified them. They have also to be seen in the light of the variability of the individuals in those populations, and the many other populations which abut geographically onto them. Thus where I suggest they confound a clear difference between the two species H. maraisii and H. magnifica, this is not only how it is to be understood. The essay concerning H. floribunda will illustrate how extensive the problem actually is. These four populations can be discussed in terms of each of the species named in the previous paragraph.
One has to come back to biological variation and consider how one is to circumscribe a species in such a way as to facilitate identification. Here we have a single population in which the basic features such as colour, size, texture, spination, venation and translucence, are so variable that it is impossible to establish what the mean or average plant is. The problem is compounded by the fact that the variables can also not be broken down to discrete quantities. I have shown only the whole plant and thus the vegetative features. The flowers are less variable both within and between populations and ‘species’. It is already clear from the literature that species in the sub-genera of Haworthia can barely be separated on the basis of their flowers. To expect that the flower can then provide a ‘signature’ or character by which population variability can be understood, is fallacious. Floral character in fact must persuade one that there are less species than we wish to find from the vegetative characters which attract us to the plants in the first place. Flowering time is useful but it is also not diagnostic. Generally flowering time in the species mentioned in this article lies between September and May. It is no more useful than the geographic location of the population. H. maraisii tends to flower from March to May and H. mirabilis is generally earlier. However, to establish the actual flowering times for each population in terms of different seasons and years, and as a mean for the individuals in each population, must be a herculean task.
What I have done is to take geographic distribution as a key element in the classification. My experience is that distribution reflects the relationship between populations and between species. This is just a logical extension of the processes we should expect from speciation processes associated with genetic interbreeding, and isolation which obviates it. “Genetic drift” should reflect the probabilities associated with pollination across distance and with the nature of the pollinating agent. Similarities between populations should reflect this, as well as the probabilities of distribution by whatever agent such as wind or water. Naturally these will not be absolute, and the classification hypothesis will only be as good as the information allows, with the proviso that it is more information we are going to require if a better hypothesis is to be formulated.
What is striking in all my formal and informal communication with interested parties is the failure to appreciate what judgmental factors may underlie the vexing thing we regard as a “species”. I need to make it quite clear that I take a species to be a system of individuals judged on a very wide variety of factors – including geographical relationships and the influence these seem to have on other similar systems belonging to other genera.
In Haworthia, I do not deceive myself that these systems are as discrete as my classification may suggest. That is why I wrote that I was recognising nodes in a complex interconnected array. Someone put forward their own solution to one such array by suggesting that the intermediate element be recognised as a species. This is particularly unacceptable when one considers the full implication. All the intermediates could be recognised as species and the present species then as intermediates! The position is that my classification was undertaken as a proper revision when a considerable amount of new data was available; sufficient to consider it really representative and adequate on which to base an overview of the genus. For persons now to suggest an alternative classification without a proper and thorough insight into all that old material, and without adding adequately to the known record, will border on the irresponsible and inconsiderate of the purpose of classification.
[1] Breckenridge (Haworthiad 19:4, 2005) for an example among others, admits to being wholly lost in respect of a remark I made (Haworthiad 18:44, 2004) in respect of varietal names. I wrote “… (variety) is a useful rank only if there is no typical variety, and if it is recognized that the species name is used for all the variants both formally named AND unnamed.” I was referring to the practice in nomenclature where the description of an inferior rank (variety in this case) automatically creates a “typical” rank. It is obvious that if a species is described from limited material, it is improbable that the description will include all the variants. Thus any new material of that species may differ from the described. Formally giving this new material a name should in fact broaden the initial description. If the ramifications of this fact are not apparent to the reader, then no amount of writing and explanation will lift the fog of misunderstanding. ♦
This chapter was published in Aloe 40:2:41-52, 2003, and I have added an addendum with further explanation and discussion.
My first experience of this species Haworthia limifolia, was with a plant brought from somewhere in Zululand by my late uncle Frank Bayer. He was involved with malaria control in then Zululand and Northern Natal. He must have given the plant to my father while we were staying at New Hanover during the period 1950 to 1954. I have always been attracted to Haworthia, but I did nothing more than admire the plant as it flourished in my mother’s care. I never really concerned myself with the origin of the plant other than recalling my uncle telling that he had stopped for lunch by the roadside, and his Zulu assistant had come back to the car with the plant. I had thought he said “near Nkandhla”, while my father later doubtfully remembered “Nongoma”. It could have been “Ithala”. I recall now that my uncle was stationed at Vryheid during that period and this is a small way inland from either Ithala or Bivane Dam, which feature later in this article.
It was only when I had left University that my interest really developed further. By this time the plant had divided and there were two very big plants each measuring 23cm in diameter. I was working at the Cedara Agricultural College when I decided I should try to learn more about this species and perhaps describe the plant(s) now in my possession. It was obviously quite unlike anything pictured that I was aware of, and certainly the leaf surface did not match that illustrated with the original description of H. limifolia by Marloth in 1908 (see Fig.1). His species was supposed to have come from simply “west of Delagoa Bay”, leaving it to the imagination as to when one switched to “east of Walvis Bay”. Certainly in the way Haworthia were regarded as species, these plants of mine were decidedly different from what I perceived to be H. limifolia. It must be pointed out that no one has ascertained just where Marloth’s species originated, or decided just what should constitute the geographic origin or range of the typical variety. This is a classification problem which is not adequately catered for, and Brummit’s paper (2002) may indicate why.
Fig. 1. H. limifolia var limifolia Marloth as illustrated in Trans. Royal Society S Africa.
A colleague of mine was good enough to accompany me on a trip to look for the species in 1962. We started at the Hluhluwe and Mkuzi Game reserves. We had a very adventurous journey that included being shown a place where a plant had been known to grow. Another experience was to be directed into the veld to occupy us while the police were called to query our presence. What puzzles me now in retrospect is that in my later description of the large H. limifolia var. gigantea (figs.2 & 3), which we never found), is the record in that description of a collection I made at Ngeba just north of the Hluhluwe reserve. I have no recollection of such a plant whatsoever.The big disadvantage my friend and I suffered from on our journey was the inability to communicate with locals. A bigger disadvantage was my sheer ignorance of field collecting and the preparation and effort needed to produce results. During our fruitless journey we went as far north as Stegi (now Isiteki) where I met Capt. D.R. Keith. We also saw and collected plants on the Blue Jay Ranch on the Umbuluzi River to the north. Capt. Keith had collected a plant named for him as the var. keithii (fig.4 – note: a clone, fig.5 in cultivation in J.W. Dodson’s collection as a topotype of the var. keithii seems to differ quite radically from the illustration accompanying the description by G.G. Smith. It is a plant with shorter squatter leaves than the illustrated type). Again I did not record properly the origin of that variety – my notes state “Palata Farm”. This is approximately 12km southeast of Stegi, and the farm Ravelston a little further on. Capt. Keith was also the collector of H. ubomboensis (later H. limifolia var. ubomboensis, figs.6 & 7) from south-east of Stegi. This could have been a single clone in an otherwise tubercled population, as several dried specimens from that general area are in the PRE herbarium.
Fig. 2. H. limifolia var gigantea, MBB112 Nongoma, as illustrated in Journal of SA Botany 1962.Fig. 3. H. limifolia var keithii GGSmith, as illustrated in Journal SA Botany 164, 150.Fig. 4. Haworthia limifolia var gigantea currently in cultivation. Note spreading leaves with discrete tubercles, dark coloration and thin leaves.Fig. 5. J.W.Dodson 2081 H. limifolia var keithii topotype. Note fewer broader and shorter leaves.Fig. 6. H. limifolia var ubomboensis. Clonotype in cultivation. It is not known if this variety has any population basis.Fig. 7. H. limifolia var ubomboensis. The same clone when stressed.
Over the years I simply left H. limifolia to history – for which there is some reason. Nature Conservation authorities made it painfully obvious that they were the only true custodians of nature. Nevertheless I did see several plants of H. limifolia in collections, such as:
a. the Mozaan River plant collected by Bruyns-Haylett (fig.8), illustrated with my description of the var. gigantea.
b. a plant from Barberton collected by Frank Stayner (fig.9).
c. a plant from Gollel mentioned in my varietal description (see fig.10, which is ex hort. Mr. Roux),
Fig. 8. Bruyns-Haylettsn. H. limifolia var gigantea (striata nom. nud.) Mozaan River, Pongola.Fig. 9. F.J.Staynersn. H. limifolia var limifolia. Barberton.Fig. 10. Rouxsn H. limifolia var limifolia. Gollel.
d. a few clones kindly sent to me by Mrs Tony Pooley then at Ndumu which I was sure she said had come from near the entrance gate – subsequently refuted (personal communication).
f. a plant in the garden of Mrs Mairne Hulme (fig.11) near Richmond, Natal. This was a very dark-green plant with distinctive raised wavy undulations across the leaves. These undulations are unquestionably not tubercles in the sense of any departure from the colour or texture of the surrounding leaf surface. Thus we have evidence that Marloth’s assertive “non-tuberculata” is a reality. In my revision (1999), I illustrate a plant from the Umbuluzi gorge which can be seen also to be “non-tuberculate”.
Fig. 11. H. limifolia var limifolia ex Mrs Mairne Hulme, Thornhill, Richmond
I saw plants reported to have been collected at Louwsburg which were rather like the big var. gigantea with the same tubercles on the leaves, rather than the transverse ridges we usually associate with the species. These plants have been distributed as var. gigantea, but they do differ in having thicker leaves which are sub-erect so that the rosette is more upright.
At one stage I also had a plant from the place Goba in the north-east of Swaziland again collected by someone from Johannesburg. There were also other odd plants with no history whatsoever. I made one further trip to the Pongola area in about 1980 to be shown H. limifolia by Denis DeKok some 20km west of the town. This is a rather tuberculate plant, the tubercles somewhat whitish and the leaves form a fairly closed, almost rounded rosette (fig.12).
Fig. 12. MBB sub Denis DeKoksn H. limifolia var gigantea. 20km W Pongola.
A collection with no good history also surfaced ostensibly from near Malelane east of Nelspruit. The same collection has been cited as from Komatipoort and Hectorspruit. I saw it in the Pretoria National Botanic garden in 1996 (cited from Hoedspruit!), and plants obviously from the same place have been sent to me from the USA – having arrived there by courtesy of ‘friends of a collector’. This population is still extant and known to Nature Conservation. The plant has been described as H. limifolia var. arcana (figs.13, 14 & 15) by G.F. Smith and N. Crouch (2001). A population nearer Barberton was said to have been annihilated by medicine collectors.
Fig. 13. H. limifolia var arcana. Hectorspruit, Ex Pretoria NBI.Fig. 14. H. limifolia var arcana. Ex J Hurter, Nelspruit.Fig. 15. H. limifolia var arcana. Ex Bridgevale Nursery.
From the USA (Peter Alonso), I received a plant from a collection by John Lavranos from Bumbeni “on the southern end of the Lebombo mountains” (fig.16).
Fig. 16. J. Lavranos8387 H. imifolia var gigantea. Bumbeni, Hluhluwe.
This is a very handsome plant with striking white markings. It appeared from John Lavranos’ own communication that the population was now extinct, as was said to be true for another collection he once made near Barberton. These are the extracts of letters by John Lavranos concerning these collections of his:-
re Bumbeni…
It is thrilling to know that a plant collected May 31, 1971 is still in cultivation and proof, if this were really needed, of what contributions the hobby can make to conservation, in the true sense of the term! This plant was collected near a place called Bumbeni, on the southern end of the Lebombo Hills, a range 300 miles or so long, which forms the border between Mozambique and S. Africa (Swaziland) and extends south into Natal. It was growing on very rocky east-facing slopes. The climate is very warm with occasional light frosts at night, in winter. Plants were quite common in 1971. However, all have now disappeared it seems. The reason for this was certainly not commercial collecting for the hobby but rather commercial collecting for medicinal purposes, as the plant is prized by African traditional medicine. Your plant, I am afraid, is now a true museum specimen the more so as the form does not seem to sucker. At the time it was identified as var. keithii. The plants varied substantially. The only thing they had in common was their large size and the fact that I never saw one that suckered. One used (upto the mid-60’s) to be able to buy such plants in the African medicine shops. But I have seen none in the last 30 odd years. They were collected out for the medicine trade.
1. The S. end of the Lebombo hills, near Bumbeni is composed of Rhyplit lava, which is acid. The soil is a scanty mixture of quartz sand and humus.
2. Plants in the wild were solitary. We found none with offsets or suckers. My collection number refers to a number of clones. These were all growing in a limited area.
3. I only collected the species once more, in 1956, east of Barberton, but doubt if it is surviving in cultivation.”
re Barberton…
* “The Barberton limifolia was as you describe it. It was collected on 10 June 1956 above Barberton, on the way to Havelock Mine. I remember finding one single large clump of it. I searched again many times in ensuing years but never saw it again. But of course those hills have been heavily grazed and frequently burned since 1956″.
Charles Craib (writes in the literature of plants he collected east of the Mashongololo mountains near the Paris (Bivane) Dam, north-east of Vryheid (figs.17 & 18). Crouch et al. (1999) record “three small populations”, two of which are within the Mkhuze game reserve. They would have been aware through Geoff Nichols of the third at the Bivane Dam.
Fig. 17. C.Craib H. limifolia var gigantea. Mashongololo Mts. environment. There appear to be plants with discrete tubercles as well as with bars.Fig. 18. C. Craibsn. H. limifolia var gigantea. Mashongololo Mts environment. In this plant he tubercles are concolorous.
A dramatic find was by Fanie Venter, ostensibly from near Barberton (Three Sisters) of also a smooth-surfaced, lime-green coloured plant (fig.19) very like the var. ubomboensis. The leaves are seemingly more glaucous, and the plant is bigger – also producing more stoloniferously rather than the close offsetting of var. ubomboensis.
Fig. 19. F.Venter13700 H. limifolia var glaucophylla. Three Sisters, Barberton.
What is now most disconcerting about H. limifolia is the fact that it is regarded as highly endangered. Crouch and Smith (1999) report an unbelievable 22.5 tonnes of H. limifolia traded through Durban markets per annum – an estimated 479 000 plants. Although these authors give places like Kanyayo, Inanda, and Espofu as sources for these plants, no attempt seems to have been made to locate plants in the field. So the “recovery contribution” in the title of their article is limited to “three small populations” which are all within protected areas. This is not in the least surprising considering the logistic problems of searching other areas thoroughly, and probably the greater problem being of moving freely through a politically and socially divided, and troubled area. Actually it is very difficult to make a search at all when working from established roads rather than according to terrain. Also there is no conventional structure of land ownership in a large part of Kwazulu Natal.
Crouch et al. (1999) suggest that the local and inconspicuous nature of the species may have resulted in under-collection and poor representation in herbaria. This is in stark contradiction to the volume of plants that is reported to be collected each year by the locals. Actually the species is fairly well represented in herbaria by the ordinary standard for succulent plants which do not lend themselves to making herbarium specimens – and better than for many other Haworthia species. It is probably only the fact that H. limifolia is away from the traditional succulent-rich areas, that collectors have not paid more attention to it. It is additionally curious that the species is so poorly represented in collections, particularly in regard to label documentation. I never found it an easy species to grow, although I must have propagated many offsets from the leaves of my original gigantea, and still have such a clone grown from the original collection. Some of the older limifolia collections were identified as varieties of H. pumila or H. minima, and similarly a plant of Aloe aristata from the Drakensberg was once thought to be a relative and given the manuscript name of H. natalensis.
Where a species like this is regarded as critically endangered, it certainly seems unwise to disclose known localities. However, whether this is true or not, I actually think it is counterproductive and against a basic ethic of science – no secrecy. Applications for permits to collect plants are granted with reluctance, and many collectors simply ignore the whole process. The consequence is that information from people who are interested in the plants and who can and do locate them in the field is not fed to where it matters. There is confidentiality about locality records and there should perhaps be more discussion about the merit of this secrecy. On the one hand it does nothing to preclude people from looking for the plants, and on the other everything to keep new information concealed. It overlooks and ignores the fact that it is motivated individuals who have contributed so much to the knowledge of our flora. It also minimises the fact that the plant is known virtually only in nature reserves.
Nature Conservation is perceived to keep an extremely strict and close control over permits in a “hands-off” approach. The encouragement for private individuals, or anyone interested in nature for that matter, to pursue any interest is almost non-existent. The converse is probably true, and so severely so that it must encroach on constitutional rights. A consequence may thus be that there appears to have been no tangible or noticeable effort to locate and assess the conservation status of H. limifolia outside of existing nature reserves. My own report of the species in the Hluhluwe/Umfolozi corridor in 1962 has gone unquestioned although this area is now included in the enlarged consolidated reserve. Ironically while permits have been so parsimoniously granted, field collected plants have been flooding the medicine markets. It does not seem to make sense. For forty years it has been almost impossible for anyone to have legally collected plants of H. limifolia, and now they seem to be regarded to occur only in Nature reserves! Is, and was, anything achieved? It may have been far more sensible to have rather embarked on an educational mission to emphasise the value of sustainable activity and the desirability of maintaining a genetic bank of material in cultivation which is adequately recorded and curated. Thus conservation could be seen as for people, and not a mission to dissuade them from nature as happens now. It should be said that as an impersonal public institution, Conservation Authority is perceived to be heavy and high-handed. The absolute converse applies when one is able to deal with individuals in such organisations and speak on a basis of common interest.
At present the known approximate locations for H. limifolia appear to be these.
Fig. 23. J. Bronkhorstsn H. limifolia var giganea. Nr Pagamisa, Magud, SW Pongola.
Armed with some of this information I began to contact interested parties to see if I could set about verifying the continued existence of these populations, less hopefully expand the known record, and build on the report in Aloe (1999). I am rather critical of this last cited paper where I thought the authors had failed to even have made an effort to examine the position in the field, if only in respect of the localities cited above, which are all within the general knowledge of the species. My interest was also driven by the fear that the threat to the continued existence of the species in the wild is real, and that there is a need for ex situ conservation and the establishment of a gene bank of some kind. My past experience has not led me to believe that such ex situ conservation actually is workable – and it will not be into the future unless it is done with very strong attention to identification of the genetic elements in any collection and assurance of continuity. This applies also to institutions such as recognised Botanic gardens where my long experience has shown that their collections are too closely tied to temporary incumbents, and that management has had no real direct interest in specific management of endangered species.
Geoff Nichols kindly reported on his experience with the species in respect of the medicine trade and the plants known to him. Dr Pete Goodman and Rob Scott Shaw of Ezemvelo KZN Wildlife were also most forthcoming with information regarding known populations at Mkhuze, Paris Dam and Ithala. Similarly Mervyn Lotter and Gerhard Strydom of the Mpumalanga Conservation authority offered their co-operation in respect of the Lowveld. A surprise was to learn of Errol Harrison who had collected the specimen cited for “West of Somkhele” near Hluhluwe in 1956 and who was alive and well at Mtubatuba.
Contrary to what I have written about Conservation attitudes above, the Ezemvelo and Mpumalanga authorities issued me with the necessary permits and my excursion was assured. Several other factors precipitated the decision to go, such as the support of my family, offers of accommodation at Barberton, and a 50-year school reunion back in Natal.
My wife Daphne, and I, started our Haworthia epic at the Queen Elizabeth Park headquarters of Ezemvelo KZN Wildlife at Pietermaritzburg. Here Rob Scott Shaw kindly explained the position regarding the only plants they had in cultivation there. This was part of a consignment confiscated from poachers into the Ithala Game reserve north of Louwsberg. Some of these plants had been planted in the open at the Park in Pietermaritzburg where they had flowered well and set seed (figs. 24 & 25). Brigette Church had very successfully germinated and grown that seed. I was allowed to take offsets off the mother plants and, in retrospect, I foolishly did not ask for any of the seedlings. None of the plants were quite like the var. gigantea and I should have considered the variability of the cultivated seedlings more closely.
Fig. 24. MBB7148 H. limifolia var giganea. Queen Elizabeth Park ex Ithala Game reserve.Fig. 25. MBB7148 H. limifolia var gigantea. Queen Elizabeth Park ex Ithala Game Reserve.
At Durban I visited their municipal Bridgevale Nursery in the company of Geoff Nichols. Mark Gillmer was the propagator who had done a marvelous job with plants bought at the local herb market. The plants were rather similar to the Ithala plants and I obtained six clones (figs.26, 27, 28, 29, 30 & 31). Only one of these (fig.31) could I really relate to my original var. gigantea in having the thin leaves, many small concolorous individual tubercles and potential for size. However, the leaves are thicker and more erect than I would have accepted. My conclusion was that, like the Louwsberg (immediately south of Ithala) plants referred to above, the var. gigantea could have originated in that general vicinity but the material I have seen is not sufficiently convincing.
Fig. 26-31. MBB7136 H. limifolia var gigantea. Bidgevale Nursey, Durban Municipality.
From Durban we traveled to Mtubatuba to meet Errol Harrison who had told us that the Somkhele area had now been vastly built up and it was improbable that plants would still be there. We did thus not explore the possibility of plants there or in the now greater Hluhluwe game reserve. This should be done. Errol took us to Phinda where we were disappointed to find that this Resource Reserve did not include the area we needed to see. However, by really good fortune we did find the owners of Pumalanga Game Ranch at home, Mr. and Mrs. Mannie van Rooyen. With the goodwill and kindness that characterised all the people we met on our journey, they took us to a site bordering on the expanded Mkhuze Game Reserve. We found the plants there without too much trouble, but there were only about 20 plants with a total of about 30 rosettes. There was an old ruin nearby which we were told was Bumbeni (a name derived from “bumba” for mud) and the stream which was the source of that mud for building. Thus it appears that this is in fact the same place where John Lavranos had collected more than 30 years before. (See figs.32 & 33 as well as fig.16).
Fig. 32. MBB7137 H. limifolia var gigantea. Pumalanga (Bumbeni), Hluhluwe.Fig. 33. MBB7137 H. limifolia var gigantea. Pumalanga (Bumbeni), Hluhluwe.
From Pumalanga, Daphne and I went on to Mkhuze where we were fortunate to find Mr M.Gumbe, Technical Assistant, who took us immediately on a long and high-speed walk in the failing light (hence no photograph) to find plants on Inxwala Hill. The plants are comparable with those at Bumbeni. Inxwala is one of the localities on Mkhuze reported in the Aloe (1999) article. From Mkhuze we travelled to Pongola where we again met most helpful people, but which did not materialise in the form of many collections despite our conviction that there must be many populations in the area. Why this conviction? From my experience with landowners in the Cape where there are many Haworthia species, people are simply not aware of small cryptic plants. Even with the local Zulus we found on our first trip in 1962, that generally people are just unaware of the plants. The same applies in the Cape where few farmers or their employees are conscious of what actually occurs on their property. Botanists have confused things like Aloe aristata and H. limifolia, and it is hardly surprising that less knowledgeable people make worse mistakes. This is what happened at Pongola and my opinion is that it is just a general level of nescience rather than ignorance, which clouds our perceptions of rarity. I make the distinction between two ignorances viz. when one is expected to know, and that where one is not expected to know.
Fig. 23. J. Bronkhorstsn H. limifolia var giganea. Nr Pagamisa, Magud, SW Pongola.
Johan Bronkhorst reported that there were plants (fig.23) along the road from Pongola westward to Pakamisa and Louwsberg. We stopped at a likely farm where we contacted the owner who in turn referred us to his foreman. This foreman took us to his home where he had a half-drum filled with soil and was growing “umathithibala” – the colloquial name. The plants consisted of H. attenuata, H. attenuata var. clariperla, H. radula and two Gasteria species. I must explain that at Pumalanga there was some confusion too, as apart from two lonely H. limifolia on the rockery, a huge container was filled with H. attenuata var. clariperla (better referred to as a look-alike of which more later). When we explained that we wanted to see the local “umathithibala”, the foreman said that was easy and directed one of the rangers to guide us up the hill to see it. This the ranger did most competently but the plants we were shown was a proliferous and abundant crassula!
We then ventured along the Pakamisa road where the terrain was most promising but simply inaccessible from the point of view of access and ownership – it would have taken more time than we had available to explore the area and determine ownership. Back at Pongola we were lucky to meet Kathy Cameron, a dynamic ex-teacher who was running the information centre. She put us in touch with two people. The first was the local herbalist selling her wares on the town pavement. There we acquired a guide who offered to take us to the township were the plants were. At this village of Ncotshane we were again shown a container, with six plants of H. limifolia consisting of at least three clones carrying seed. We were graciously allowed to take three offsets. But we asked to be shown where the plants had come from. In response a finger was pointed to the nearby hill and we set out, with some apprehension, to climb it. It was also to no avail and we saw nothing in rather unlikely terrain just as we had feared. Ncotshane would class as “north of Pongola”, and I have no doubt that on those many hills, the species does in fact still occur.
Fig. 34. MBB7140 H. limifolia var gigantea. Daaiwater, Pakamisa, Magud.Fig. 35. MBB7140 H. limifolia var gigantea. Draaiwater, Pakamisa, Magud.Fig. 36. MBB7140 H. limifolia var gigantea. Draaiwater, Pakamisa, Magud.
Through Kathy Cameron we also contacted Rynold and Rolene Steenkamp at Magud. Rolene very kindly guided us to where she had known plants on their farm Grootdraai near Pakamisa west of Pongola. There we found 10 plants among rock in a very grassy and somewhat wooded area (figs.34, 35 & 36). Rolene said that the population was far less vigorous than she remembered it and it did seem as if the surrounding vegetation was too competitive for H. limifolia. Time was now very limiting and we left Pongola with much left simply undone, and we travelled to Isiteki. Again we were assisted by really kind and gracious people – Jonathan and Helen Pons who own Mabuda farm. We initially tried on our own to explore the Palata/Ravelston area. This was frustrating in the extreme as the road passes along the escarpment but never approaching near enough to initiate an excursion. Often the vegetation was overgrown with aliens forming an impenetrable thicket while there was the same problem of land ownership and authorization to enter, presented by such communal area. Palata and Ravelston farms had been expropriated by the British Govt. to hand to the Swaziland National Trust on the granting of independence. So our visit there turned out to be no more than a preliminary reconnaissance and we were not able to assess var. ubomboensis. Later Jonathan Pons obtained for me two plants from the approximate area (figs. 37 & 38). Curiously the former does mirror the var. keithii in profile whereas Fig.38 does so the leaf surface.
Fig. 37. J.Ponssn H. limifolia var keithii. SE Isiteki, Swaziland.Fig. 38. J.Ponssn H. limifolia var keithii. SE Isiteki, Swaziland.
From Isiteki we travelled to Barberton where we were met by Mervyn Lotter of the Mpumalanga Parks Board. He took us to see the locality for var. arcana, which unfortunately for us had been fenced and we were faced by a locked gate. However, this was not a serious issue as the plants are well-described and there is adequate material in cultivation to ensure ex situ conservation (in fact it is being grown at Bridgewater nursery and I did acquire an additional clone there for my own record). We went then instead to Kaapmuiden and ended up on the farm Boondoks. We obtained permission to explore a little, and after one unsuccessful excursion Mervyn found a very small population of 84 rosettes in an area of about 2sqm (figs.39 & 40). The area had been burnt and the plants were quite severely singed. The plants do seem to prefer tight rocky habitat with sparse grass cover and so it would take a very severe fire to kill them. It was just very difficult to form an opinion regarding appearance of the plants and I could only surmise a resemblance to var. arcana.
Fig. 39. MBB7143 H. limifolia var limifolia. Boondoks, Kaapmuiden. Plants severely singed, suggest var arcana.Fig. 40. MBB7143 H. limifolia var limifolia. Boondoks, Kaapmuiden.
At Barberton we met Rhona Milstein, who has lived in Barberton all her life and is extremely knowledgeable about all things pertaining to the place. She guided us to a nearby hillside were it was thought the population had been collected out. Fortunately this was not true and we found many plants albeit in a small area again in closely fragmented rock and sparse cover (figs.41). The plants were again badly scorched. We saw the plants again at May Mine on the Mountainlands Game Reserve alongside the Havelock Road. This was in the presence of a joint owner Mr. Nico Oosthuizen, who had very kindly agreed to our entry to the site. The plants were very abundant over an area of about 150m x 10m but again in a very rocky area (figs.42 & 43) and also fire scorched. It was interesting to note the presence in Rhona’s garden of a very stoloniferus form of H. attenuata cf var. clariperla which I have not seen before.
Fig. 41. MBB7141 H. limifolia var limifolia. Rymer’s Ceek, Barberton.Fig. 42. MBB7142 H. limifolia var limifolia. May’s Mine, Barberton.Fig. 43. May’s Mine scene.
Our next excursion was to locate Fanie Venter’s (13700, fig.19) collection at Three Sisters. Again we were overwhelmed by the kindness and helpfulness of John and Bernadette Roux, owners of Three Sisters. They took great pains to show us where to find the plants and stayed in telephone communication while we drove along the tortuous rough road to the near summit of the Sisters (three peaks, with intimidating steep slopes which we would never have negotiated on foot). We found the plants on a nearer locality than indicated to us and there were many in a very small area. The vegetation was almost alpine and very dense. Fire here would surely result in the death of many plants, with survival only in rockier more sparsely vegetated mini-refugia (figs.44 & 45). From here we could look down on the Boondoks site and also identified a likely hillside to the southwest of this. We went there on our descent of the mountain and after a long walk we did find a very small population in an area of 0.5sqm. I refer to this population as Manders Dam. It may have originated from a single clone, which may be true for the Boondoks plants too. The plants were growing in the excavated old soil of a termite mound and badly scorched (figs.46 & 47). Their exposure to fire could not be considered to be more or less severe than had they been anywhere else in that terrain and my opinion is that fire per se is not a hazard to the species. In general if the fuel load is high enough to present a threat to survival of the plants, they are anyway subject to threat by exclusion of light and unfavourable growing conditions.
Fig. 44a. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton. 029Fig. 44b. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton.Fig. 45. MBB7144 H. limifolia var limifolia. Manders Dam, Barberton.Fig. 47. MBB7144 H. limifolia var limifolia. Manders Dam, Barberton.
However, there were odd inflorescences about and it was evident that no seed was set. Fire may affect the pollinators and naturally will also destroy inflorescences and seed if untimely. But this surely is a seasonal factor and there will be years when seed is produced. What one can speculate is that the populations are derived from vegetative propagation as the species does proliferate readily from stolon. If this is so they will probable be non-seed producing as plants are usually self-sterile.
It was sad to leave Barberton with so much to do and such promising unexplored terrain. The drive to Vryheid was also frustrating for that reason. There must be populations of the species in the area Badplaas, Osplaas (Gerhard Strydom of Mpumalanga Parks Board reported rumours of its presence there) on to Lochiel, Amsterdam and Piet Retief. At Vryheid we were taken to the Paris (Bivane Dam) where, pushed for time, we actually saw only one population (figs.48, 49 & 50), surrounded by most promising terrain. We went to Nongoma where we were introduced to a local herbalist and shown “Umathithibala”. This turned out to be a container with Aloe aristata from the Ngome Forest area, and also a plant of H. attenuata. The herbalist explained the use of “umathithibala” to defuse tension and disarm intruders with harmful intention. At Pongola we had been brought three other Aloe viz. bainesii, sessiliflora and komatiensis …) which were said to be grown around the home to tranquillise atmosphere. Thus the usage of the plants is not directly medicinal, but embedded in a paradigm of esoteric values. She did not know H. limifolia. To Jonathan Pons, the Swazi name for the plant is “sidodobala” and they apparently also use it to ward off lightning or else simply explain its use in this way as the Zulus do. The proliferation suggests its use too to encourage fertility in livestock and for that purpose it is grown around kraals.
Fig. 48. MBB7146 H. limifolia var gigantea. Bivane Dam, Vryheid.
Our last stop was at Ithala, which I had not intended to visit because the plants were familiar to us from the Park Headquarters at Pietermaritzburg. There we met the Conservation Manager, Mr. Rob Blok, and were shown plants in the office rockery. What was in the rockery were two plants which I think should not have been there. One was Aloe squarrosa (an exotic) and the other was H. attenuata. This latter plant could perhaps hybridise with H. limifolia. But the plants we were shown were Aloe aristata and said to occur in the Reserve too. As the site for the originally poached plants was on a distant area of the reserve, I asked if I could see and photograph A. aristata. Without the spines, these specimens were remarkably similar in colour and size to H. limifolia as it occurs in the greater Pongola area. We went to the area indicated and must have missed the specific point where the Aloe may well have been and saw instead some plants of H. limifolia which were very like the Paris Dam plants. Our impression was that the species must well be very common at Ithala and surely so between Vryheid and at least Pongola.
Conclusion The prime object of the trip was based on the assumption that the species H. limifolia was critically endangered and hence that there is proper motive to ensure an ex situ gene bank. Thus we collected within the limits of our permit authorisation for this purpose, and these plants will be distributed under accession numbers in due course as they proliferate. The second object was to try to extend knowledge of known localities from the impression given in Aloe (1999), at least to the record cited in Haworthia Revisited (1999) and evident in RSA herbaria. The third object was to derive some kind of estimate of conservation threat. The fourth was to arrive at a new opinion on classification for known and unknown material. Thus:
1. Material of 20 accessions is now held by me of which 12 can be regarded as seed reproducible. It would be very desirable to increase this.
2. The cited record clearly still exists and is largely confirmed. At least five of these new collections can be regarded as new. Hence if one considers the product of a virtually blind-driven effort, there must be many more populations which are still unknown.
3. The conservation threat from fire which has been cited seems to be fallacious, and the same may even be said for the ‘herbal’ trade. The species does not have a particular unique role in herbal medicine or mythology. Where the threat does lie is common to any living thing and far more applicable to any number of animals and other plant species – this is the density of the population living off the land and widespread degradation of the environment due to fire being used to generate grazing for livestock or simply to clear the landscape from vegetation which may harbour vermin, snakes and human intruders. Thus all organisms in communally owned land are at risk. There is a conservation plus in the large number of game ranches and reserves which are being game fenced and patrolled to stop poaching.
4. Classification of plants at least in respect of Haworthia is now bordering on the ridiculous. There are any number of would-be taxonomists all with their private versions of what constitutes species, how old names can be interpreted, what name changes can be implemented and what new names can be invented. As Stevens (2002) stated, botany has been taken over by nomenclature. The peer test of any taxonomic work is largely based on observance of the nomenclatural rules, the viability of the Latin diagnosis, and the protocol of literature and specimen citations. Brummit (2002) also writes that the binomial system does not really work. This has been evident in Haworthia for the last 40 years although here it could be largely the fault of the writers.
My opinion regarding H. limifolia is based on my view of how a species should be defined. This is as a dynamic system of living organisms which are morphologically and genetically continuous in time and space. In Haworthia I have found that the geographic element is essential to the understanding of taxonomic elements. In this respect there appear to be no real discontinuities between all the records cited above despite a wide variation between say var. arcana (in the absence of a population which we can say authoritatively is exclusively and decisively var. limifolia) and var. gigantea, which also has no true provenance. The var. striata could apply to any of the Pongola collections where white striations (as the continuous concolorous bars in var. limifolia) occur as frequently as there are small discrete tubercles as in var. gigantea. What is probable is that the name striata originates in the plants collected at Bumbeni by John Lavranos which were introduced into cultivation. There is no evidence that the single plant from Mozaan which I photographed in Bruyns-Haylett’s possession was ever propagated and distributed. I had simply suggested to John Pilbeam while he was writing his book (1983), that based on what I had seen, a varietal name “striata” was reasonable. I am of the opinion that the Mozaan plants are closer to my variety gigantea than they are to the Bumbeni (and Mkhuze) white-banded variants. Thus the varietal name gigantea could apply to the species as it is known south of the Swaziland border, and the name striata would apply to specific forms and cultivars which have confluent white tubercles or raised white bands across the leaves.
The var. ubomboensis has not been appraised as a population based entity and neither has the old var. keithii. What is evident is that there is another disjunct population of a similarly glabrous element at Three Sisters, Barberton, as FVenter13700 and MBB7145. This latter element is definitely population based and the two collections cited can be recognised as fairly discrete. Unlike the var. ubomboensis which is a smooth yellow-green colour in good cultivation, this Barberton element is a glaucous bluish-green and the name glaucophylla is given to it as follows:
Haworthia limifolia var. glaucophylla (Asphodelaceae: Alooideae) – var. nov.
Type: Mpumalanga, 2531(Barberton): Three Sisters (CB); F.Venter 13700 (Holo. NBG)
Very similar to the var. ubomboensis but glaucous green and geographically discrete.
In habitat small pale yellowish-brown in colour, rosette seldom exceeding 3cm diametre, 8-12 leaves reaching 6cm in length. Surfaces smooth, occasionally with some rounded or barred tubercles. Slowly proliferous. In cultivation very proliferous and off-setting, rosettes to 12cm in diam. with 25-45 leaves, colour glaucous bluish-green, hence the epithet. Differs from the typical in being without raised transverse lines, and from the var. ubomboensis in being bluer in colour and geographically discrete.
There appear to be a number of discrepancies in the description of the var. arcana. In the protologue the leaf surfaces are described as undulate but generally lacking distinct tubercles as in the typical variety. This is not correct. The typical variety was named limifolia, meaning file-like, for the very reason that there were these raised undulate transverse ridges on the leaves – not discrete tubercles. The var. arcana that I have seen generally has such broken ridges which reduce to individual and quite large tubercles. This is quite different to the many smaller discrete tubercles in say var. gigantea where there is no evidence of transverse bars at all. In many of the collections I obtained in the greater Pongola area, there were both continuous bars across the leaves as well as plants in which the leaves were tubercled instead. In these collections it was common for the leaves to be either erect, incurved, sub-erect or spreading. Regarding flowering time – Smith and Crouch state that in contrast to other varieties of the species, the var. arcana flowers in mid- to late winter. This is not true. The collections at both Queen Elizabeth Park and Durban had flowered prior to spring and flowering was completed at the time of our visit in late September. Similarly all the evidence we saw of flowering on our trip was that it had been completed before our visit. We saw late flowers at some sites and collected seed at Draaiwater and at Bivane’s Dam.
This raises the point that we seem to have a convention for how taxa should be described. In the var. arcana as for other Haworthia taxa, there is a long description of the flowers which is quite valueless in the context of the structure and variability of the flowers in the whole subgenus. It lends to the elaboration of the process to give it a validity which it earns from just the fact that the taxon is worth identifying by name for practical communication and comprehension. Furthermore it feeds a perception that this fine detail enhances the credibility and validity of the element when it does nothing more than lengthen the description. Similarly, rather than use an epithet which is derived from a skewed viewpoint of what is known and what is not known (viz. arcana – mysterious, secret) why not a descriptive one such as olivacea, highlighting the extraordinary colours this taxon can get in cultivation? There are still two other things. The mapped distribution of var. arcana is entitled “distribution range” and suggests an area of circa 2600 sq km. when in fact there is one known population occupying perhaps 0.25ha. The suggestion that it is one of more links to H. koelmanniorum is window-dressing with no other truth in that var. aracana is geographically the closest known of the limifolia variants to a not so climatically stressed H. koelmanniorum at Groblersdal and Loskop Dam.)
Fig. 51. Map of Mpumalanga, Swaziland, Kwazulu-Natal to show general distribution of H. limifolia.
Acknowledgement I have seldom made all the acknowledgements in my writing that I might have done. This particular trip of mine was made under special circumstances without which I would not even have thought of going.
Firstly my son Warwick, who suggested I should go and who facilitated the trip by offering a vehicle.
Then to:-
Ms. Adri Henn, a business associate of Warwick’s at Natal Drift Nursery, Barberton. Her offer of accommodation was a vital factor in deciding to go.
Mr. Gordon Morrison and my school contempories who agreed to a 50-year reunion at Ixopo. A most moving occasion.
Ms. Michele Pfaff of the Gauteng Conservation Authority for contacts and discussion on conservation issues.
Mr. & Mrs. L.Alleman of Link Hills, Kloof for hospitality and encouragement.
Mr. Charles Craib for important information and assistance
Mr. Geoff Nichols, a conservation consultant at Durban. Also a naturalist and plant enthusiast par excellence. Also to his acquaintance at Bridgewater Propagation Nursery of the Durban Municipality, Mr. Mark Gillmer.
Ezemvelo KZN Wildlife for the necessary permit and accreditation of my effort, and to Dr. Pete Goodman and Rob Scott-Shaw for valuable co-operation and information. Mr. M.I. Gumbe, technical assistant at Mkhuze Game reserve. Mr. Rob Blok, Conservation Manager, Ithala game reserve. Mr. Alex Wood, Regional manager, Vryheid. Ms. Bridget Church, Horticulturist, Queen Elizabeth Park.
Mr. Errol Harrison of Mtubatuba.
Mr & Mrs. Mannie van Rooyen of Pumalanga Game Farm.
Mr. Kevin Pretorius and Mr. Matheus Wessels of Phinda Resource Reserve.
Ms. Kathy Cameron of Casamia, Pongola.
Mr. & Mrs. Rynold Steenkamp of Magud.
Mr. &Mrs. Jonathan Pons, Mabuda farm, Isiteki.
Mr. Mbabane Matsenjwa, Mabuda farm.
Mpumalanga Conservation Authority for permit and accreditation, and particularly to Mr. Mervyn Lotter and Mr. Gerhard Strydom.
Mr. Johan Hurter of the Lowveld Botanic Garden.
Ms. Rhona Milstein of Barberton.
Mr. Nico Oosthuizen of Mountainlands Game Reserve, Barberton.
Mr. Luke von Johnston of Boondoks Farm, Kaapmuiden.
Mr. & Mrs. John Roux and their son Etienne of Three Sisters, Kaapmuiden.
Mr. & Mrs. Hugh Foster of Rosthwaite farm, Vryheid.
Mr. Barry Killian of Bivane Dam, Vryheid.
Mr. & Mrs. Graham Root of Ntibane Bushveld Hideaway, and herbalist Erica.
Mr. Bob Kent, and Mr. Steven Hammer who were kind enough to comment.
Dr. Paul Vorster was also so good as to offer me his opinions that are not reflected in the article.
Lastly to my wife Daphne whose unfailing trust and friendship defy any evaluation.
To all these and a few whose names I did not record, I must record my appreciation for the courtesy and kindness in listening to my story and offering me so much assistance and advice. There are also many people peripheral to this special journey who played their part and who I omit mentioning. Had I gained no new knowledge or insight into Haworthialimifolia, at least I gained the association of many people whom I would like to call my friends.
Addendum Having written so much about Haworthia in the process of learning about them and knowing them better, I find it incomprehensible that other writers seem to ignore completely what I have had to say. While this may appear to be a very arrogant attitude, it simply mirrors what I consider “science” to be; primarily a review of what is known or thought to be known, and then a statement of intent, action and conclusion. At present there exists a primary field of formal science which resides in students and graduates from schools of learning (professionals), and then the field of informal science where formally unqualified persons (amateurs) try to forward their perceptions and observations. My frustration is that the hearts and boundaries of these two fields in respect of Haworthia have been and now are, so confounded, that “knowledge” and “understanding” are wholly concealed. My frustration is expressed in the following statement sent to the Editor of Alsterworthia…”The actual position is that classification and the naming of plants is not an activity calling for any special skills and it does not class as ‘science’. It can be for science. Brummit, in Taxon, has written that the binomial nomenclatural system does not truly work. If this is true, it means that for all these years taxonomic botanists have been slavishly applying a system which is flawed. Their failure to have properly applied their minds to the task has led to incongruities which are not that difficult to detect. Where leaders in the field have had their doubts and indecisions, nomenclatural niceties have become more important than the goal. Biology being the inexact science that it is has meant that totally amateur classifications have come to be lauded, praised and accepted when they have no intrinsic intellectual credibility at all. Anyone has free entry to botanical classification whether or not they can even sensibly discuss the issue of the definition of the word ‘species’ which is the basic unit of biological classification.
It is in this respect that I quote Lyall Watson, from his book, Dark Nature, where he writes about the similarity science now has to religion, and it is worth recording that the Dalai Lama has said the same thing. In Watson’s opinion scientists are largely defending a position in which they have a vested interest and they say things they wish to hear and what they expect others to want to hear too. Science according to Watson is thus in the same trap as religion … ‘an establishment intent more on protecting its own interests than in looking at the world with curiosity and honesty’.
This is the thrust of many things I have written and my objection to statements made by leading botanists in respect of classification of Haworthia. Offence will be given if I point to these individuals and their statements, but if ‘curiosity and honesty’ are employed, they can easily be found. One manuscript of mine in respect of just one of the papers which I would use to exemplify my point, reads … “it would be more honest just to acknowledge the impracticality of unispecific genera, than parade a host of fallacious arguments to obscure them. The other even more unfortunate aspect is that all this fault‑ridden argument passes over the heads of the unsuspecting community that is interested in the plants and wants to use the names? If this is what professionals do, what can be expected from amateurs? The foundation is secured for the most uninformed and ignorant to parade as taxonomists, and they do. All that actually happens is that credibility becomes a very rare commodity in plant classification and nomenclature.’ Many of us generate opinions and comment which actually have roots in ignorance.”
I therefore wish to encapsulate my observations in this addendum which addresses the above issues in respect of one object viz. Haworthialimifolia.
To do so I have first to recapitulate slightly the issue of definition of the word species. I will not repeat again anything I have written in this regard nor discuss again the paper of Brummit and its implications again. I merely say that I regard the Latin binomial as a reference to a system of living organisms as a group or groups of organisms which are genetically and morphologically continuous in space and time. The binomial itself is derived from a sample and this is formally attached to a single specimen lodged in a recognised herbarium.
What then is the species Haworthialimifolia? We have to go first to the sample and the description. The sample seems to have been a single specimen (perhaps more, the description states… “the plants”) received by R. Marloth in the year 1910 from Medley-Wood, Director of the Botanic Gardens at Durban. He is stated to have obtained (note: not “collected” as Breuer, 2000, interposes) them “originally from the country west of Delagoa Bay”. The description is extremely brief and it is accompanied by an illustration of a single leaf (fig.1). It can be said of the description regarding the flower that it is so vague that one could not even recognise to which of the three major groups in Haworthia to which it belongs. Here there is already room for confusion in the failure of various writers, either amateur or professional, to recognise what these are. This is especially so when there is a distinct probability that the evolutionary roots of those three flower types could be polyphyletic. This means in essence that classification by vegetative or any other “structure” may not necessarily indicate the same grouping.
It is quite clear that Marloth was right in his assumption that he had to deal with a species “quite different from those of any other species” and “sufficient to constitute a new section”. But he was not quite right in respect of the reason or ultimately the rationale. He was right because H. limifolia is a species system in the northeast of Southern Africa (with H. koelmanniorum) and thus widely separated geographically from any of the other species. It is not because the leaves have a specific surface character or marking and this is a point I will have to labour on later. Apart from some similarity to the geographically contiguous H. koelmanniorum, there is H. nigra in the central Karoo which bears some similarity in respect of leaf “markings”. I put “markings” in inverted commas because it is not correct. The “markings” in H. limifolia are specifically transverse undulations on the leaf surfaces giving it the file-like appearance from which the Latin epithet is derived. Marloth stated quite explicitly that the “markings” were not tubercles or tuberculate.
It is really curious that G.G. Smith (1950) – and he was amateur – had no qualms whatsoever in using the word “tubercles” for the leaf surface character. This was when he described a new variety, keithii and also reduced H. ubomboensis Verdoorn to varietal rank in H. limifolia. Var. ubomboensis with a few longitudinally arranged tubercles on the upper and lower leaf surface was in Smith’s opinion, together with the similar longitudinally tubercled var. keithii, recognised to be part of the system “west of Delagoa Bay”, namely H. limifolia. Smith introduces some new perceptions. He describes the var. keithii as lighter in colour, with less rigid and narrower leaves and less prominent tubercles, but similar in its stoloniferous habit, arrangement of back and face tubercles and the longitudinal lines on the face. Referring back to the original description, the only character here which Smith could have correctly taken (he did get the leaf surface wrong) was perhaps in respect of the width of the leaf.
After Smith there was a bizarre contribution by Flavio Resende (1943) in which he described essentially four varieties of H. limifolia, if it is at all necessary (and possible) to try and follow his logic. None of these were of known origin and my assessment of Resende’s total contribution to Haworthia, and specifically to H. limifolia, leads me to counsel complete disregard thereof. Nothing is added to knowledge of the system that can in anyway be currently applicable and useful. All that it does is further confound and confuse the issue in an environment that I am trying to show is already very difficult to function in. It could for example be argued that a variety with the proposed new name viz. gideonii could in fact be Resende’s f. diploidea (= var. marlothiana Res.). The next author to venture information by way of a new epithet was myself in describing the var. gigantea. This was based on a single specimen that for reasons cited in the preceding section, can be reasonably considered to have its origin in the Pongola valley. The last significant contribution was by Pilbeam, possibly as a consequence of my writing, tied to the collection by the very distinguished collector/explorer J. Lavranos, of the var. striata.
Breuer (2000) has subsequently made an excellent compilation of descriptions and illustrations. Other than that it is an excellent compilation which brings all the literature together, it adds neither further information nor new insight. In respect of H. limifolia, it does curiously add a Berlin herbarium photograph to illustrate the “typical” variety. This photograph could just as readily been used to illustrate the var. schuldtiana or one of the other varieties such as even gigantea. There is nothing to suggest that Breuer could have and did consider all the above, nor the three papers which I deal with below. Breuer (2002) has proposed his system for Haworthia in which there is a new varietal structure for H. limifolia. However, I am familiar with his other outstanding compilations and the many less rational imputations and statements which he attaches to names and his perceptions of what those names are as “species”. These often call for the kind of response which Smith and Hilton-Taylor (1996) made to Breuer’s “contribution” to the phytogeography of the genus Haworthia (1996). Breuer clearly cannot know more about H. limifolia than either Smith et al. (1997), nor Crouch et al. (1999). I feel constrained to say that Breuer is not competent to even attempt such taxonomic rearrangements. The title of his 2002 paper includes the words “species concept update”. It is in fact nothing of the kind. Breuer (2002) does not produce any evidence of what his “species” concept is and his list is only his concept of what species there are in Haworthia. Having produced a revision, I am fully aware of many of its limitations and what is needed to verify it and resolve outstanding problems. Among the problems is the major one of the species concept which has to be resolved at polymath level as it is in the realm underlying the whole of taxonomic botany. Breuer has given me no indication of his capacity and willingness to undertake the intellectual activity this impels, and has dismissed most of my attempts as “polemically things” (private communication). He is also not positioned to undertake the arduous and extensive field-work that is required to even verify my revision, let alone add to it or make changes.
The next attempt of note to explain H. limifolia is that by G.F. Smith et al. (1997). Smith et al. paper purports to illustrate and describe the typical variety limifolia on the basis of two specimens both of uncertain geographic origin. The significant thing is that the paper ignores Marloth’s specific statement that the species was not tuberculate. They repeat the curious aberration that G.G. Smith introduced. Despite the key the paper provides, the var. gigantea is misidentified as var. limifolia as can be gleaned from the illustration. I do not doubt that to a degree I am also culpable here because I know a draft of the paper was sent to me for comment. Unfortunately I do not have any recollection of what my comment was and any contribution I may have made is not acknowledged. Certainly it has not corrected this error. I would have thought the authors would have come to realise their error when they became involved in the next paper.
This is the contribution by Crouch and Smith (1999). It must be noted that the title refers to the typical var. limifolia. This paper does acknowledge a contribution from me, but I must say I think this is misplaced and could refer to the earlier paper by Smith (1997). I cannot imagine that this second paper, and certainly not the final draft, by Crouch and Smith would have passed my critical eye. The paper essentially deals with plants either obtained from the Durban herbal market, or from plants shown to the authors by Dr. Peter Goodman, then resident botanist at the Mkhuze Game Reserve. It also contains some quite specious comments about classification. Clearly no effort at all went in to considering and exploring known records or to generating new ones. From what I have discussed in the earlier part of this work, the variety the paper deals with is either gigantea or striata and definitely not the var. limifolia. I have suggested that these two former names are probably synonymous, and I have also suggested that the recognition of a typical variety is somewhat of a problem in botany. The species as a system is typified by ALL its variants. I take Crouch et al. for being remiss in emphasising, as they do, the importance of understanding taxonomic identity and then making such a weak effort at doing this. Here they are dealing with a variable element distributed over an extremely wide area. They have information that the area still extends considerably southward of the known records which they have in addition not properly registered (I will explain this later in this note). No effort was thus made to question distribution. One can hardly take Breuer remiss for thinking he could revise the genus from Nederzier, when local professional approach the “importance of taxonomic identity” in this way.
Now we come to Smith and Crouch (2001). The paper opens with the words “After consideration of available evidence…”. In my opinion very little of the evidence that I have presented here, of that which was then available, has actually been considered. Among my criticisms of the paper is that of distribution. If Smith and Crouch had properly considered this, they would have been struck with the fact that H. limifolia has been reported near Komatipoort as “SildoSuavo to Moamba”, and that the one small locality known for arcana lies between this record and records south-west of Malelane and at Barberton itself. They should also have considered Venter’s collection at Three Sisters which is only about 30km distant. It is beginning to appear to me that as my plants collected from these sites grow in cultivation that these will also class as var. arcana. The area is between Delagoa Bay and the Reef and it is thus highly probable that the var. arcana is a superfluous name for limifolia. Despite their extensive discussion of classification and population variability, the authors simply accept plants of unknown origin to represent Marloth’s species. These plants are the var. gigantea and NOT var. limifolia.
There is very little in Marloth’s description than can be put to good account. The colour is given as “obscura-viridia” which can be taken for dark-green and otherwise it is the transverse non-tuberculate undulate raised lines on the upper leaf surfaces which are the major point of the description. The detail of the flower is quite worthless and it is not possible even to determine the subgenus. Despite this paucity of information, there is little difficulty in saying that all the plants included in the preceding discussion are H. limifolia. What is it then that makes this species recognisable among this welter of words and the confusion they contain?
In their diagnosis for the var. arcana, Smith and Crouch make the following points:- it is “characterised by a combination of vegetative morphological features. It has a distinctive dark-brownish green colour and the leaf-shape is somewhat more reminiscent of H. limifolia var. ubomboensis than of typical H. limifolia. The leaf surfaces are prominently undulate, but generally lack the distinct tubercles characteristic of the typical variety. Furthermore, it differs from the other varieties in the flowering time from the middle to the end of winter.”
Colour I would agree may be distinctive as this is what struck me when I first saw plants at Pretoria. However, colour is a fractious character and largely influenced by growing medium and exposure. My three collections from the close area where arcana occurs (i.e. Boondoks, Manders Dam and Three Sisters), show quite marked differences in colour with the Boondoks plants at present very dark green and Manders Dam plants simply dark green. The glabrous Three Sisters plants are a glaucous bluish-green. At present I have clones of var. arcana which are olive-brown (fig.13 ex PRE), purplish-brown (fig.14 ex J.Hurter) and dark green (fig.15 ex Bridgevale). Smith and Crouch’s remark concerning leaf-shape is misleading as comparison with the Marloth illustration will show. In any case the var. ubomboensis is based on a single clone in which the leaves are quite long and slender. What the Marloth illustration shows is a short-leaf with a reflexed end-area. This is not apparent in all my plants of var. arcana, but there are clones which exhibit this markedly and others which do not. Similarly in plants of var. gigantea from Bridgewater, I have one specific clone in which the leaves are all very reflexed. When it comes to the question of surface character which is the most significant part of the Marloth description outside of origin, then the whole argument falls apart. Marloth’s diagnosis makes the point that the leaf surfaces are prominently undulate whereas Smith et al. say of arcana …”but generally lack the distinct tubercles characteristic of the typical variety”. What Smith et al. seem to have done is to consider that the typical variety limifolia is as they illustrated and described in Bothalia. This is where they have taken the var. gigantea to be the typical, and thus their detailed description does not bear them out as it is incorrect. I have already commented on flowering time.
The conclusion I must reach is that if the var. arcana is properly researched it will be found to contain all the ingredients of the typical variety and prove to be a superfluous name.
I wrote (1962) that “A great deal more information on the distribution and variability of H. limifolia is required before the true relationship (i.e. a good classification) of the various forms can be determined”. Despite this warning and a good many more since, classification is still used as an easy vehicle for publication. I think we have here a series of three publications which are based on a minimum of field-work and knowledge of the subject. It all suggests that my contentions expressed in my letter to the Editor of Alsterworthia, quoted above, are unfortunately true. A new name may be useful as a language tool for collectors and growers even it is quite misleading. It may also be just a source of revenue to traders. On the other hand it maybe worthless as a tool for the meaningful organisation of information and knowledge while it introduces further complexity and confusion into classification. If a name is not rooted in sound reason and science, it can only be such a source of confusion and misunderstanding. In Haworthia there is a constant inflow of poorly researched dubious comment about classification which provokes negation or negative response. The end result is that the literature becomes simply contentious and there is nothing there for the information of the general reader. This is no doubt why nomenclature and name changes are so vilified by the plant growing community. We need a new ethic in classification.
As an end note I must explain the probable source of the statement by Crouch et al. that the distribution of limifolia extends much further south than suggested in any previous reference. While preparing for my exploratory trip to search for the species, I wrote a letter with illustrations for publication in Veld and Flora (The journal of the Botanical Society of South Africa) asking if any readers could help with information. I received seven responses of which only one properly identified the species and this was a photograph of a potted specimen as used in an advertisement by a clothing store. Three re asking if any readers could help with information. I received seven responses of which only one properly identified the species and this was a photograph of a potted specimen as used in an advertisement by a clothing store. Three responses were in respect of Aloe aristata and the others were for H. attenuata. Extraordinarily two of the latter respondents sent me plants or photographs of plants received by them from people living in the Isipofu (Espofu of Crouch et al.) area of Kwazulu-Natal which is in the Sottburgh district south of Durban. The one respondent informed me that his plant had been identified by a botanist as “H. limifolia”. This is clearly the source of the Crouch et al. statement and should be considered in the light of my experiences and encounters with H. attenuata further north related in the main text. ♦
Literature cited
Bayer, M.B. 1962. A new variety of Haworthia limifolia from Natal. Journal of South African Botany 28:215.
Bayer, M.B. 1970. Haworthia as a problem genus. Cactus and Succulent Journal (U.S.) 42:251.
Bayer, M.B. 2000. Thoughts on Haworthia. Spiderwalk Services, Durbanville.
Breuer, I. 1996. A contribution to the phytogeography of the genus Haworthia Duv. Haworthiad 10:35.
Breuer, I. 1998. The World of Haworthia. Vol.1. I. Breuer, Nederzier.
Breuer, I. 2000. The World of Haworthia. Vol.2. I. Breuer, Nederzier.
Breuer, I. 2002. An Haworthia species concept update. Alsterworthia International – Special Issue No.1.
Brummit, R.K. 2002. How to chop up a tree. Taxon 51:41.
Craib, C. 1999. The ecology and status of H. limifolia east of the Masongololo mountain in South Africa’s Kwazulu Natal. Haworthiad 13:49.
Crouch, N.R., Smith, G.F., Nichols, G., Burden, J.A. & Gillmer, M. 1999. A species recovery contribution for H. limifolia var. limifolia, the umathithibala of the Zulu. Aloe 36:8.
Marloth, H.W.R. 1908-10. Some new South African Succulents. Part.2. Liliaceae. Transactions of the Royal Society of South Africa 1:403.
Pilbeam, J.W. 1983. Haworthia and Astroloba, a Collectors Guide, London.
Resende, F. 1943. Succulentas Africanas 3. Memorias de la Sociedade Broteriana 2:94.
Smith, G.F., Crouch, N.R. & Condy, G. 1997. Haworthia limifolia var. limifolia (Asphodelaceae: Alooideae). Flowering Plants of Africa 55:24.
Smith, G.F. & Crouch, N.R. 2001. Haworthia limifolia var. arcana (Asphodelaceae: Alooideae): a new variety from eastern South Africa. Bradleya 19:117.
Smith, G.F. & Hilton-Taylor, C. 1996 A contribution to the phytogeography of the genus Haworthia Duv. – response to Breuer. Haworthiad 10.
Smith, G.G. 1948. Views on the naming of Haworthias. The Journal of South African 14:55.
Smith, G.G. 1950. Some new species and varieties in the genus Haworthia: VII. The Journal of South African Botany. 16:3.
Stevens, P.F. 2002. Why do we name organisms? Some reminders from the past. Taxon 51:11.
Haworthialimifolia is something of an enigma in that both it and Haworthiakoelmanniorum are geographical rather isolated from the rest of the genus. The latter is confined to a small area around Loskop Dam and Groblersdal in what was the old Transvaal province. H. limifolia is much more widespread and occurs from the southern Kruger National Park southwards through Swaziland in to Northern Kwazulu Natal. It was described by Rudolph Marloth from a specimen very loosely said to have come from “west of Delagoa Bay”. No one has ever managed to tie the species down in terms of geographic origin, and the Flowering Plants of Africa (55:24-29, 1997) description of the typical var. limifolia, depicts it as the var. gigantea. My own fieldwork and observations of material of known field provenance show that this var. gigantea and the var. striata are surely synonymous. Neither of them could truthfully said to be from “west of Delagoa Bay”.
The var. arcana is another matter and a very curious one too. It does come from a place very close to the old wagon routes serving the goldfields of the Transvaal viz. near Hectorspruit. However, plants from there do not seem to match Marloth’s original description, and there is some confusion about the question of the “tuberculate” versus the “non-tuberculate” of that original description. Having seen plants which I think are truly non-tuberculate and yet with the file-like ridges across the leaves, I do not think var. arcana fits the bill for the typical variety. When I first saw a specimen of this variety in the Pretoria Botanical Garden while David Hardy was still there, I would also have plumped for a new variety based on its odd colouration. However, that specimen I saw was very olivaceous and I would have named the variety accordingly. The plants which Crouch and Smith collected of var. arcana are more brown coloured.
I was fortunate enough to be able to spend some time in Swaziland and Mpumalanga (Transvaal Lowveld) in 2002 looking for H. limifolia. One of my aims was to see the var. arcana in habitat, and the other to re-locate F. Venter’s collection of a glabrous specimen from very near the same place. The material of Venter’s was illustrated in Haworthia Revisited as var. ubomboensis.
Fig.1 is P.J. Hurter’s 105 of var. arcana from the type locality and Fig.2 is a specimen of Neil Crouch’s ex Bridgewater Nursery, Durban, also from type locality (approximately Hectorspruit – the locality has been variously given as Komatipoort and Malelane. It should also be noted that there are rumours of the occurrence of the species northwards within the Kruger Park, and there is a firm record for Suado Suavo across the border in Mocambique). I photographed the latter Crouch specimen because it is a less tubercled glabrous plant among a few others. It is quite evident from these two pictures that the prominences on the leaves would qualify as tubercles with a little evidence to suggest that they could become concolorous with the rest of the leaf surface and also merge to form the ridges described by Marloth.
Fig. 1. P.J.Hurter102 H. limifolia var arcana. HectorspruitFig. 2. N.Crouch H. limifolia var arcana. Ex Hectorspruit, at Bridgevale Nursery, Durban.
Fig.3 is my MBB7144 from south of Malelane and about 30km west of the Hectorspruit population. Although there were about 100 rosettes they were in a very restricted area and I suspect that they were all of one stoloniferous clone. The three samples I collected are identical. They are the same size as the arcana specimens but the colour is darker and the brownish tint is absent. Fig.4 is my MBB7144 from about 2km to the south-west and again there were only about 15 rosettes in a situation which suggested plainly they were clonal. The plants are larger, the tubercles concolorous and there is a stronger tendency to banding. My MBB7145 (fig.5) is of Fanie Venter’s glabrous element (I have named it elsewhere as var. glaucophylla) from the top of the Three Sister’ mountain. This is about 1,5km from MBB7145. My collection seems to not be from the same precise locality as that of Venter’s and I do suspect, although the plants were plentiful and scattered over a small area in very rough terrain, that the population is derived from vegetative proliferation. My attempts at pollination have so far not been successful and I need to try and pollinate the Venter plants with my own.
Fig. 3. MBB7143 H. limifolia var arcana. Boondoks, Barberton.Fig. 4. MBB7144 H. limifolia var arcana. Manders Dam, Barberton.Fig. 5. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton.
Lastly I have MBB7142 (fig.6) from very close to Barberton itself and MBB7141 (fig.7) even closer. These I refer to the typical variety. It is highly improbable that these are related clones because of the distance and terrain separating the populations. However, the plants are so consistently alike within and between the populations that it is a possibility that cannot be ruled out. In the previous chapter I illustrated a F. Stayner collection from near Barberton which is very similar to these two collections of mine. Stayner gave the locality as “Barberton Airfield”. This, given the nature of the terrain, is highly unlikely and it appears to me that Stayner was in fact referring to an airstrip near Kaapmuiden, and very close to Three Sisters. The leaves of the plants are more barred and file-like, but there is till a strong tendency for the coalesced tubercles on the upper leaf surface to be broken down the mid-line. This could impact on the integrity of the plants in that area being a geographical entity. I have not been able to successfully pollinate between plants within those Barberton collections and have not tried between them. Although I have plants of var. arcana from three sources, I have also not been able to get them to set seed and it is possible that they are all clonal in origin. This may be true for the original locality too, although it is reported that there are many plants there, and the clone I photographed from Crouch’s sample appears to be different from the other specimens I have.
Fig. 6. MBB7142 H. limifolia var limifolia. May’s Mine, Barberton.Fig. 7. MBB7141 H. limifolia var limifolia. Rymer’s Creek, Barberton.
I have no conclusion to draw other than to say that the problem of genera in the Alooideae is a reflection of the problem in the lower ranks. Alternatively it should be stated in the converse. This may help cladists to better appreciate that building a phylogenetic classification from only the top down probably will not satisfy ordinary people who will use the classification. Where the subgenera are unequivocally (notwithstanding anything Dr. Hayashi has to say in this regard) recognisable on the basis of the floral structure, the species are not. Thus even in the subgenus Hexangulares it is difficult to know if all the species satisfy the requirements of definition. For example H. coarctata and H. reinwardtii, or H. attenuata and H. glabrata, may be synonymous. Even H. koelmanniorum and H. limifolia may be so. A species such as Haworthia pungens (fig.8) also creates problems because of the similarity to Astroloba on the basis of the leaf spirals approaching 5-farious, and the smoothness and rigidity of the leaves. It may well be that vegetative is more compelling than floral structure, and this has to be very carefully considered when speculating about true phylogenetic relationships. ♦
Fig. 8. JDV96-100 H. pungens. Braamriver, Joubertina.
During my early years with the Karoo Botanic Garden, in fact it never really got any better, my time spent looking for Haworthia was largely my own. This meant hasty weekend trips, or looking for Haworthia secondarily to other things and more general goals. Also it was a question of scale. I needed to see populations and plants from across the entire distribution range. So those years were spent rather as reconnaissance and in checking all the herbarium and other records. I came across a letter among Major F.R. Long’s papers which put me on the track of a Mr. P.L. Meiring of Bonnievale, south-east of Robertson. This was a copy of a letter from Meiring to Triebner in Windhoek arranging for the collection and purchase of an Haworthia from Drew Station. The result was that 200 plants were sold for the huge price (for those days) of 1s each – allowing for inflation this would have been worth about R14ea in today’s currency.
The plants must have been referred to as H. mutica, or this is simply the general name I was using for what appeared to be a fairly distinctive element south of the Riviersonderend mountains and west of the Breede River. Meiring indicated to me where he had collected the plants and I stopped hastily in that general vicinity he indicated northwest of Drew station. A hurried excursion into the veld and after a short while, I picked up a branch of Renosterbos, and there was a plant. Having confirmed the site, I most uncharacteristically collected this single specimen – something I have never done again. The plant remained in cultivation at the Karoo Garden and is probably still there, while leaf propagules have found their way around the world. Kobus Venter obtained one of these and also still has it in cultivation (Fig.1 JDV92/64). When he started taking an interest in Haworthia in 1985, he set off to look for this mutica. Alas, no success. The site had been cleared and ploughed for a massive centre-point irrigation system. We have many times explored the nearby disturbed and undisturbed patches near there, finding only the odd H. maraisii / heidelbergensis plants and a few H. minima var. poellnitziana. The occurrence of these has given us some hope that H. mutica may still be surviving somewhere there.
Kobus and I discussed the issue and when his plant came into bud, we asked the new owners of Sanddrift, Mr. and Mrs. J. Humby, if we could place the plant in the veld in the vain hope that it would be pollinated. The plant was duly taken there, still in its small pot, and sunk into the ground under the protection of three scraggly Renosterbos plants. When I called there late in September the plant was looking healthy but the developing flower was rather scrawny and too shaded. I broke away a few branches to open it up a little. Late in October I called back and to my astonishment there were three seed capsules. The plant was gleefully taken back to Cape Town and I collected 120 seeds from the three capsules. In my joy I planted half of the seed at once. Alas! It was too early in the summer and too hot for this winter regime dweller. Not learning, I planted another 20 seeds a month later and now have 6 rather spindly seedlings and 20 seeds still to sow.
Sanddrift is a remarkable place as three of the four species of Robustipedunculares grow there in discrete populations, and there are hybrids between both marginata and pumila, and marginata and minima. It is unclear just what relationship heidelbergensis and maraisii enjoy there because elements of both are present too. But now we know that mutica is still there somewhere and we have seed to prove it. The nearest other population is south of the Riviersonderend River more than 15km away (Fig.2a, b & c JDV96/27). Here the plants do not have the purplish-grey tones of the species as I would generalise for it south of the Riviersonderend mountains – but then I never have with any conviction claimed that it is different to H. retusa or H. pygmaea. One of the few specimens from there also evidences the whiteness in the leaves which manifested in the Drew plant many years after its collection. The exposure of our potted plant to its native air and a deposit of dry renosterbos leaves, seem to have given the plant a rejuvenating kick and the accompanying illustration shows how beautiful it looks for its visit home. Sept. 2001 is soon upon us and methinks the White Widow needs the company of her soul mate again. (Note:- in a later Chapter, I describe the rediscovery of H. mutica at Sanddrift, Drew).
Acknowledgement To Kobus of course for his deep interest in anything going for Haworthia; and then to Beth Humby whose love for anything to do with life and with nature is extraordinary. It was wonderful to have the co-operation of persons who are so enthusiastic and interested. ♦
Fig. 1. JDV02-64 Sanddrift DrewFig. 2a. 96-27 H. mutica. SE Drew.Fig. 2b. JDV96-27 H. mutica. SE Drew.Fig. 2c. JDV96-27 H. mutica. Drew.
When I wrote the first part of this essay, I was anticipating completing it in three parts. However, I was also in the process of exploring more widely and thoroughly, and the problem and its explanation seemed to grow exponentially. The result was seven essays, and they are presented here as close as possible to their original format. The purpose is to show how a classification should have predictive value, and how an understanding of plants can develop, or fall apart, as more information accumulates. The seven parts were published in Haworthiad 17:1:24-32, 17:2:53-54, 18:1:21-33, 18:2:52-57, 18:3:92-101.
Part 1: Haworthia mutica var nigra indeed!
An early problem in my career as a Haworthia taxonomist was that I had plants of H. mutica which I could not distinguish from plants of H. pygmaea. The former is from the Caledon/Bredasdorp area in the west, and the latter from Albertinia/Mossel Bay in the east. I had also had a collection between these, from Kransriviermond south of Heidelberg, which did not fit comfortably with my perception of H. retusa at Riversdale, or with either that of H. mutica or H. pygmaea. I was also stuck with the probability that there could be other continuities by virtue of unrecorded and unknown populations. A known troublesome population was that from south of the Tradouw Pass that I eventually included in a wider interpretation of H. magnifica, however uncomfortable that was. A problem here is that in separating magnifica from maraisii as species, and in also recognising the variety H. magnifica var. atrofusca, the closer identity of magnifica gets lost in a fragmented residue. As will be seen in the following discussion, this also relates to the understanding of H. mutica var. nigra.
While there was initially a limited amount of material to go by, it might have been thought that more collections would clarify the problems. Instead it became increasingly evident after my first two handbooks that additional collections were confounding the picture even more, and they certainly have. Esterhuizen made the first of such collections from Klipdrift, northwest of Heidelberg. Then there were two populations recorded by Venter, from east of Heidelberg and southeast of Heidelberg. I did not have material of Esterhuizen’s collection but mentally included it with the Tradouw pass population as H. magnifica var. magnifica. The last I included in my concept of H. mutica var. nigra while I was not immediately certain what to do with the population just east of the town of Heidelberg, close to where H. heidelbergensis occurs. I knew these retusa-like plants from photographs and specimen in G.G. Smith’s record. They are very similar to the Klipdrift plants except the habitat is again slightly different and the plants are solitary and the leaves level with the soil surface. It should be noted that I drew attention to this aspect before, when I suggested that H. retusa was in reality a ecotypic adaptation to level ground as is the vegetatively proliferous turgida the adaptation to steep rock faces. The problem was lack of material and the best I could do then was to predict continuity between mutica and retusa. Thus the variety nigra was out of the geographic areas of either and yet not substantial enough to suggest that it was a separate SYSTEM. I have since seen Smith’s plants live as JDV90/113 in cultivation and in the field.
In the last two years I have paid more attention to H. mirabilis and the disconcerting variability of this species. A closer study of the var. sublineata has produced a dismaying degree of variability and I do not doubt that an argument could be made on this score for including the Tradouw Pass plants under H. mirabilis. This is not as improbable as it may seem if all the variants are considered such as in H. heidelbergensis, H. maraisii, H. floribunda. Experience with the Eastern Cape complexes also leads me to predict that these western species are similarly going to be inextricable from one another.
Fig. 01. Map of the Klipdrift-Heidelberg area. Note the farm Blackdown, the type locality for H. floribunda where it hybridises with H. turgida which is common in the area.
Without proposing another solution, other than for the Tradouw plants, I needed to first explain the Esterhuizen population. Here I was fortunate to have the interest and hospitality of Dr. Christie Venter who grew up on a farm just to the northwest of Klipdrift. He now has a property near Zuurbraak where he accommodated my wife and I and we were able to explore the Klipdrift area (see map, Fig.1) to good effect. We found four populations along the southern side of the Duiwenhoks River, two on Goedehoop (Figs.2 & 3) to the west of Klipdrift, one on Klipdrift itself (Figs.4, 5 & 6) and another on Doornriver to the east (Fig.7). From there we went to the two sites east and southeast of Heidelberg (Figs. 8, 9 & 10), as well as to a further population midway between these two. We also revisited the familiar locality for H. heidelbergensis east of the town.
Fig. 02. MBB7133 H. mutica var niga. Goedehoop, west of Klipdrift. The habitat is a gradual slope with little rock, partly bush covered and with Aloe ferox.Fig. 03. MBB7133 H. mutica var nigra. Goedehoop.Fig. 04. E. Esterhuizen in JDV92-29 H. mutica var nigra. Locality given as Klipdrift.Fig. 05. MBB7134 H. mutica var nigra. SW Klipdrift. Grassy habitat with riverine stone.Fig. 06. MBB7134 H. mutica var nigra. SW Klipdrift. A plant with stubbier leaves.Fig. 07. MBBsn H. mutica var nigra. E Klipdrift. A stony slope with Pteronia incana. Only a few plants on lip of road cutting.Fig. 08. MBB6650 H. mutica var nigra. Diepkloof, SE Heidelberg. Plants here are quite variable but nealry all with rounded leaf-tips.Fig. 10. MBB6650 H. mutica var nigra. Diepkloof, SE Heidelberg. With pointed leaves.
What do these populations suggest and what is the solution? These plants do not fit the bill for H. magnifica because they lack the deep dark colouration of that species or the generally shortened leaves. They do not fit with H. mirabilis because the leaves are generally more recurved, the reverse of the leaves is less spotted, and the surfaces are generally smoother. The geographical location does not agree either. The plants cannot be placed comfortably with H. turgida var. turgida, which is a smaller more spinose and clustering species in sandstones. There is some correspondence to H. turgida var. longibracteata and also to H. retusa. From the latter it differs in having fewer more recurved and less spinose leaves, while from the latter it differs in being smaller, more translucent and with narrower more erect leaves.
There actually does not seem to be a categorical answer, which is just what I would have expected. There seems to be this pointer to the new language we need, where we agree on the meaning of names in a fairly arbitrary way. I suggest that for the present this group of populations of doubtful identity should be treated as H. mutica var. nigra. I am quite conscious of the fact that there is an opposition body that wishes to re-interpret such doubtful elements as even H. asperula, and probably produce an entirely different classification and new names. I respect this point of view and it was Prof. G.F. Smith who stated in the introduction to Breuer’s book The world of Haworthia, Vol. 2, this is a golden age and we are each now able to make our own decision! For communication purposes it would be wise if we tried to agree on one set of names. Simply each of us naming plants as species as they come before us has no intrinsic meaning or message for anyone else. It will work for a closed group in close contact with one another, and all seeing and talking about that limited range of specimens. For me who has seen so many plants over so many years, it just becomes quite meaningless. It means that we have a set of names which has no practical value for anyone outside of that group and nor does it have any value for say biologists or scientists working in other disciplines where the names have to have meaning in the broader sense of biological systems as applicable to all life forms.
Fortunately there seems to be an awakening in the botanical fraternity to the fact that our classification system and the notion of a branching tree with the species dangling on the straight branch ends, is faulty. One only has to think what happens when we have a species and then describe a variety of that species. The nomenclatural system requires that a typical variety be automatically created. The implication is that the two varieties stand equal to one another – and this is patently and obviously untrue and unworkable in practise. It actually means that while the variety may be fully distinctive and even be derived from a single always fully recognisable clone, the typical variety has to accommodate every other variant in the species. Thus the more varieties that are described, the more shattered, indefinable and unrecognisable is the “typical” variety. In this is just how it should be if we properly and logically think about distribution of phenomena in space and change with time. Where subconsciously we probably think of species as very definite definable things, the reality is that it may only be individuals (individual clones and vegetatively propagated plants) that have any individual recognisable identity. It is just a myth to say that in a species, the flowers of the individuals are always all similar (identical), or that the leaves have this special character – and so on. If readers are familiar with Haworthia Update Vol.1, they will appreciate that considerably more illustrations are required to truly reflect the relationships of the populations I discuss here and extend this to the broader geographic area i.e. H. heidelbergensis, H. magnifica, H. maraisii, H. mirabilis and others.
A consideration I must add is that the question of very local adaptations to habitat complicates the picture enormously. Thus the complex geology, topography, and climate of the southern Cape are as important as reasons for the extremely rich and variable flora as may be any history of the origin of vegetation from a distant past. Here I am dealing with just the drainage systems of a small river, the Duiwenhoks, which runs from north-west of Heidelberg first towards the south-east and then south to the sea. Other “species” in the close area are H. heidelbergensis, H, turgida and H. magnifica (note that the variety atrofusca thoroughly obscures the identity of the typical variety, and this is further clouded by other varieties, what to say of some of the names now proposed for elements within this huge complex).
Part2: Consequent to Haworthia mutica var nigra indeed!
I omitted illustrating JDV90/113 and of plants from the type locality for H. mutica var. nigra from Part 1, stating simply because these two populations lead from that system to another further east that is more complex. But this is not quite true. What is true is that G.G. Smith was considering the population later collected as JDV90/113 (as a single plant too, no doubt) to be either a new species or a new variety of H. retusa. What is thus difficult to explain is that it is not just a question of how this population related to H. retusa or to H. mutica. It is a problem of how is it related to either of these complexes that have their own internal as well as other external associations (see figs 11 & 12).
Fig. 11. JDV86-16 H. mutica var nigra. KransriviermondFig. 12. JDV86-16 H. mutica var nigra. Kransriviermond.Fig. 13. JDV90-113 H. mutica var nigra. E Heidelberg.Fig. 14. JDV90-113 H. mutica var nigra. E Heidelberg.
In the case of H. mutica var. nigra (Figs. 11 & 12) itself from Kransriviermond, I did not happen to have a picture then available and in any case it is illustrated in Haworthia Revisited. With this short note, the position was rectified and pictures of both collections are provided. It can be noted that JDV90/113 (Figs. 13 & 14) is within 50 meters of the population that I took to be the type locality for H. heidelbergensis. There are not many plants and they are solitary and the leaf surfaces are level with the soil surface. This is the general pose for H. retusa where it grows exposed on level habitat (see figs 13 & 14).
Why have I then not called these plants H. retusa? It is because I considered there was a continuous range of populations that linked H. retusa and H. turgida, and the Kransriviermond population was at variance with this opinion. It is of course equally valid to say that the plants I have illustrated as H. mutica var. nigra could also be regarded as possibly continuous with either H. retusa or H. turgida, as well as include some influence from either H. magnifica or H. heidelbergensis or both.
So what more needs to be said that can be said? An explanation really only begins to make sense when all the role players are examined in detail. H. mutica cannot be represented by a single plant and neither can H. retusa or H. turgida or H. magnifica. Even when one has all these myriad collections in front of one, one wonders what more is there which will cloud the issue further! It is evident that we tend to seek an answer in an easier two-way comparison, when the solution is a much more difficult manifold or reticulate relationship.
Part 3: Still consequent to Haworthia mutica var nigra indeed!
In this part of the essay I discuss what I refer to as “the distant turgida connection”. In the two preceding notes I commented on the problem of using the name retusa in connection with H. mutica var. nigra. It was not unduly surprised to read that Esterhuizen claimed that nigra was rather a variety of retusa than mutica (Alsterworthia). This observation of his is perfectly valid and only misplaced in the context of what I had written before, and as far back as the original Haworthia Handbook. Where the difficulty lies, apart from the intrinsic difficulty of the problem, is with our so-called “typological” concept of species. Thus we form a mental image of, say, H. retusa from an illustration, either the original type illustration: or one that has been chosen to resemble that original. Then we build images around the name of other plants that we come to know as H. retusa. But these images are seldom composite ones that accommodate all the variations and are also limited by how much we have seen. Similarly we then generate images of, in this case, H. turgida and H. mutica and then try to weave them together into a single mental construct.
In this case the introduction H. pygmaea would be somewhat of a surprise. But I did say at the start that one of my very early disconcerting experiences was having plants of H. mutica and H. pygmaea, and finding that I could only separate them on the basis of known origin and the colour of the floral bracts. Without going into the lengthy detail of the variation of H. pygmaea and its associations which include H. turgida and thus to H. retusa, briefly the tale is this: It can be taken as read that the colour of the bracts is not diagnostic for any Haworthia species, where all we then have in this particular instance is geographical information. These single images we build leads to confusion and to misapplication of names. When we think we are specifying, we are actually making very general statements, which in turn help to confound attempts at identification and classification.
Therefore I want here to just illustrate some variants of H. turgida in order to provide a broad image of this species so that its role in the tale can also be guessed at.
I do this for six reasons:-
1. I started a series of articles with the grandiloquent title “Natural variation and Species Delimitation in Haworthia”, for the National Cactus and Succulent Journal way back in 1972. One of the manuscripts I started concerned H. turgida. I had a vast amount of material and I was rather stuck as to how to illustrate it all. It was also obvious that there was a great deal more to see. So the project was shelved until now when this discussion generated the need.
2. While going through my files, I found a few of the old pictures I would have used.
3. I think that readers should be made aware of this major pitfall of perception now that so many writers are in the arena.
4. In addition to H. turgida, the following cannot be positively excluded from reckoning: H. retusa, H. heidelbergensis, H. mutica, H. maraisii, nor H. magnifica. Other authors are very industriously adding new names that will cloud the issue still further.
5. I still would like to see more H. turgida and more H. retusa. I am now quite well positioned in terms of new material, to write about the other elements.
6. The literature is now fragmented beyond sensibility and it is difficult even for me to follow it all and keep a chronological account of what is written there. Other writers have the same problem while they are not even paying proper attention to what has gone before. This is of course their prerogative; and that of the editor who permits his authors to wander where they will.
I do agree with Esterhuizen’s contention that H. mutica has not been found east of the Breede River, which could suggest that the relationship of mutica var. nigra is better sought with H. retusa. However, this would make sense if the typological concept of species was sound and if the concept of the phylogenetic tree with sequential single-branching stems was true. The fact is that one can find the characters that we might use to diagnose any one “species” across the board. Thus I can find “nigra” or similarities, in populations of turgida, retusa, heidelbergensis, magnifica, maraisii and mirabilis. Esterhuizen is quite happy (I have done so myself and it is no criticism) to use the name retusa for plants from near east and even west of Heidelberg, when in fact there is exactly the same problem of “identity” at Riversdale and beyond. This is that retusa is probably an ecotypic variant of turgida. Forget that the name retusa would have precedence over turgida if the taxonomic chips were down.
The first three illustrations (figs 15, 16 & 17) are of H. turgida in a sequence from north to south towards Heidelberg. I choose (chose) this sequence because “turgida” is the sandstone ecotype, and these populations suggested to me that there was a continuous set of populations which linked “turgida” to the more solitary clay-soil, level-ground, silcrete-hill “retusa”.
Fig. 15. MBB in KG240-72 H. turgida var turgida. 9km N Heidelberg.Fig. 16. MBB in KG241-72 H. turgida var turgida. 4km N Heidelberg.Fig. 17. MBB in KG242-72 3km N Heidelberg.
Perhaps I should also emphasise the point that I am using single clones to illustrate populations and systems – where this does not actually work. It is already an approximation. Immediately north of Heidelberg, there is a problem where turgida merges with heidelbergensis. Heidelbergensis itself has its origin in a few plants found beneath one bush, 0,4km West of Heidelberg (I was wrong in following Dekenah’s contention that the type locality was the population 0,4km east, and also heidelbergensis is extraordinarily abundant and variable just west of Heidelberg). It is apparent that the concept of heidelbergensis is based on the interface of two systems viz. turgida and one other. This “other” carries the name “heidelbergensis“, and I will in due course present some of the very extensive ramifications that have emerged in the last few years concerning this particular problem.
The next three illustrations (Figs 18, 19 & 20) are from populations quite some distance from the sandstone range of the Langeberg Mountains (which is the main “turgida” domain), for which I use the varietal name longibracteata. I could not really censure Hayashi for finding new names for any of these. There are several other such non-sandstone populations that I should illustrate, and I know I have not seen them all. I add two illustrations (figs 21 & 22) of the var. suberecta for the eastern forms of “turgida“. To finally knock a nail into the coffin, I add a picture (fig.23) of H. mirabilis var. consanguinea from the Riviersonderend sandstone mountains south of McGregor. Why I name it “mirabilis” is for very good reason, and this reason is as complex as is the full “turgida” saga or that of others. It is thus very frustrating to read simple solutions that are being presented by writers who in my opinion just do not have the competencies or material to do so authoritatively.
Fig. 18. MBB in KG312-71 H. turgida var longibracteata. 16km N Bredasdorp.Fig. 19. MBB in KG94-71 H. turgida var longibracteata. 16km N Bredasdorp.Fig. 20. JDV86-71 H. turgida var longibracteata. Stilbaai, Riversdale.Fig. 21. MBBsn H. turgida var suberecta. Draaihoek, N Albertinia.Fig. 22. MBBsn H. turgida var suberecta. Gouritz Bridge, Albertinia.Fig. 23. MBB in KG332-70 H. mirabilis var consanguinea. S Die Galg, MacGregor.
Part 4: Still on about Haworthia nigra var mutica.
When the previous Part 3 of this small series was published, a few words were omitted from the title. These were, “The distant turgida connection”. This part could have been entitled the close retusa connection. What I was actually looking for was a more definitive explanation and I could not find it. In writing about the relationship of H. floribunda, both Esterhuizen and myself have assumed that the plants I attribute to the species H. variegata viz. the varieties hemicrypta and modesta are in fact that species. It is probably truer to say that they are not and that they are simply part of a much larger complex system for which classification solutions are right for one part of the distribution range and wrong for another. I say this because in this article I will be using the name heidelbergensis in the same way (in the text all references will be in terms of the typical variety. Technically there is no population of this species in which the plants all generally resemble those from which the type was supposedly derived).
What I want to write about is a series of populations from east of Heidelberg that really throws a cat among the pigeons. There is a low flat plateau between Heidelberg and Riversdale named Kiewietskraalse Vlakte (to be referred to simply as Vlakte) at an altitude of 300m (see map Fig. 24). It is divided into three farms, Kweekkraal East, Kweekkraal West and Melkboom further west. The area to the east, north and west is very broken and generally unsuitable for cultivation. Southwards the terrain is lower-lying at about 150m and is largely cultivated. The pattern of cultivation suggests that the soils in that area are arable and hence were never really suitable habitat for Haworthia. This is an assumption based on where haworthias are found in the greater area and this is invariably on sites where the soils are very skeletal and where rock is exposed. Often these sites are on broken profiles of erosion channels or small hillsides, perhaps for the presence of harder and less erodable rock. This is not to say that there were no suitable habitats on the level areas and certainly the four species of the Robustipedunculares are/were less confined by habitat requirements than members of the subgenus Haworthia. Assumptions about occurrence and suitable habitat are important because they relate to the degree of isolation of populations, to the role of geology in the evolution of ecotypes and hence to the taxonomic relationship of those populations. My impression is that cultivation has generally not impacted on Haworthia populations.
On Vlakte the vegetation is Renosterveld. It has numerous buttresses and surrounding it are a few separate inselbergs. The upper level areas are cultivated and where the natural vegetation is still intact, haworthias are still only found in very specific and identifiable habitats. The eastern-most inselberg is Spitzkop that is the type locality for H. magnifica var. atrofusca. H. retusa in its classic form occurs there too. H. magnifica var magnifica, in its classic form, is still further east at south of Riversdale itself. H. floribunda is known at Langkloof to the north-east, but otherwise there is very little available information regarding what lies between Vlakte and Heidelberg where H. mutica var. nigra, H. heidelbergenis, and the typical true form of H. floribunda occur.
My attention was first drawn to Melkboom by Kobus Venter who told me that there was a complex interaction there between several species (retusa, heidelbergensis and magnifica). He was with J.M. Esterhuizen who has also commented on what I recall was the relationship there of H. magnifica and H. heidelbergensis. I visited Melkboom with Kobus and indeed saw what I will refer to as heidelbergenis. At one point (squared brackets indicate position on the map Fig.24) it ([1] Figs.25a, b & c MBB7108) was growing near to but discrete from H. retusa ([2]Figs.26a, b & c MBB7107). These retusa-plants have smooth leaves with very rounded tips and in this respect hence incline towards H. mutica var. nigra. But on the extreme west wing of the Vlakte plateaux there is an apparent hybrid population of retusa ([3]Fig.27a, b JDV93-53, & c MBB7110) between two populations of heidelbergensis ([4]Fig.28a & b MBB71109, c, d, e, f & g JDV93-54, and [5]Fig 29a MBB7111 b & c JDV93-52). This small population is curious because it is situated between and yet so close, to populations of only one-putative parent. The other parent, presumably H. retusa, is absent. The fact of hybridity is not proven and is only speculative based on the spatial relationships and physical appearance of the plants as judged to be “intermediate”.
Fig. 25a. MBB7108 H. heidelbergensis. Melkboom [1].Fig. 26a. MBB7107 H. retusa. Melkboom [2].ig. 27a. JDV93-53 H. Xretusa. Melkboom [3].Fig. 28a. MBB7109 H. heidelbergensis. Kleinkragga [4].Fig. 29a. MBB7111 H. heidelbergensis. Melkboom [5].Fig. 29b. JDV93-52 H. heidelbergensis. Melkboom[5].Fig. 29c. JDV93-52 H. heidelbergensis. Melkboom [5].
I had now seen H. magnifica at Kweekkraal East ([6]Fig.30a, b & c MBB6817) and West ([7]Fig.31a, b & c MBB6662), so was anxious to see what else occurred on the Vlakte. Those magnificas I had grown from seed, and I knew they consisted of a motley assemblage of forms between the more typical magnifica and atrofusca. What was the relationship then of these populations to the smaller heielbergensis plants at Melkboom? At this stage I never considered that H. floribunda would enter the picture.
Fig. 30a. MBB6817 H. magnifica. Kweekkraal E [6].Fig. 31a. MBB6662 H. magnifica. Kweekkraal W [7].
Kobus came along with my wife and I to re-visit the area and explore between Melkboom and Kweekkraal West. The Vlakte Plateau has an irregular shape and has many buttresses separated by incised small valleys. The upper edges of these buttresses often comprise fairly skeletal soils with rock that probably accounts for irregular erosion of the plateau edge. The rock also seems to overly erodable clay and this is exposed by pressure bursts which result in a low clay bank and an extended apron of bare clay soil with here and there tufts of grass or low herbaceous plants. The valleys are more thickly and densely vegetated as the soil are deeper and retain water for longer. The rock and clay sites provide the habitat for the haworthias and nearly every buttress is populated. Thus we found a confounding array of plants which all seemed to indicate a continuum from the heidelbergensis-like plants on Melkboom to the magnifica-like plants at Kweekkkraal. The populations were disturbingly variable and yet differed also from one another. The description and quantification of such “difference” is probably an ultimate challenge to botany. The collections are [8]Fig.32a & b MBB7112, [9]Fig.33a, b & c MBB7113, [10]Fig.34a & b MBB7114, [11]Fig.35. MBB7115, [12]Fig.36. MBB7116, [13]Fig.37. MBB7117, [14]Fig.38a & b MBB7118.
Fig. 32a. MBB7112 H. heidelbergensis. Kweekkraal W[8].Fig. 32b. MBB7112 H. heidelbergensis. Kweekkraal W [8].Fig. 33a. MBB713 H. heidelbergensis. Kweekkraal W[9].Fig. 34a. MBB7114 H. heidelbergensis. Kweekkraal W [10].Fig. 35. MBB7115 H. heidelbergensis. Kweekkraal W [11].Fig. 36. MBB7116 H. heidelbergensis. Kweekkraal W [12].Fig. 37. MBB7117 H. heidelbergensis. Kweekkraal W [13].Fig. 38a. MBB7118 H. heidelbergensis. Kweekkraal W [14]
My wife and I returned to the area East twice after that to further explore the plateaux. We first made these collections on Kweekkraal West [15]Fig.39a & b MBB7151, [16]Fig.40 MBB7152, [17]Fig.41a, b &c MBB7153, Fig.42a, b, c, & d MBB7154. The first two I regarded as heidelbergensis, the third inclined to magnifica, while the fourth was more positively magnifica including clones which would class as var atrofusca. On Kweekkraal East, significant finds were a population south of the Pretoriuskop inselberg consisting almost entirely of the var. atrofusca ([19]Fig.43a, b, c & d MBB7157) as opposed to several of the heidelbergensis/magnifica continuum we had seen on Kweekkkraal West. We also found H. floribunda on Pretoriuskop [21], southwest of (Fig.44 MBB7155) as well as on an unnamed inselberg to the west ([20]Fig.45 MBB7156). Exciting was that in the population of atrofusca, which was growing in a very white clay layer, was a plant of H. floribunda (MBB7158). It is extremely difficult in these field sites to recognise variants and distinguish what could have been hybrids, and we did not think there were such. However, about 50 meters away we came across a population of small heidebergensis ([22]Fig.46 MBB7159) like plants which we decided were hybrids – based on the fact that another 50 meters away was a population of floribunda. Again this is speculative.
Fig. 39a. MBB7151 H. heidelbergensis. Kweekkraal W[15].Fig. 40. MBB7152 H. heidelbergensis. Kweekkraal W[16].Fig. 41a. MBB7153 H. heidelbergenis=magnifica. Kweekkraal W [17].Fig. 42a. MBB7154 H. magnifica. Kweekkraal W [18].Fig. 43a. MBB7157 H. magnifica var atrofusca. Pretoriuskop[19].Fig. 44. MBB7155 H. floribunda var dentata. SW Pretoriuskop[20].Fig. 45. MBB7156 H. floribunda var dentata. W Pretoriuskop[21].Fig. 46. MBB7159 H. Xfloribunda. N Bosgasiekop[22].Fig. 47a. MBB7160 H. magnifica. N Kweekkraal E[23].Fig. 48a. MBB7161 H. magnifica. N Kweekkral E[24].Fig. 49a. MBB7162 H. magnifica. Far N Kweekkraal E[25].
At other sites we found several other populations that included both small ([23]Fig.47a & b MBB7160) and large magnifica ([24]Fig.48a & b MBB7161, [25] Fig.49 MBB7162) depending often on the nature of the pressure-bursts and the quality of the rock and clay. Often the clay was quite red and the rock very ferruginous (iron-stone). At other sites it was often very white and clean. The rock similarly varies from the very characteristic red-pebbly constituency of true ironstone, to the smooth white appearance of pure quartz.
Fig. 53 Site 24, showing the kind of vegetative cover over a dense population of H. magnifica.
On a still later occasion my wife and I visited Kweekkraal East again with Casper Mazurel of Holland. We had not explored the northern edge of the Vlakte plateau and needed to know more. Casper found H. retusa north of Kweekkraal East ([26]Fig.49 MBB7173) in the same kind of white clay that atrofusca occupied near Pretoriuskop – we also found floribunda again (Fig.49[23] – MBB7172). The curious thing about both the retusa populations from Vlakte, have very rounded leaf-ends (the mutica connection? – more later). The Melkboom population is relatively light-coloured and the more eastern population the deeper green/purple tones of magnifica. The small population at [25](Figs.47 MBB71620) seen on the previous visit (in white clay in which the plants were quite robust) then takes on a new face and could be imagined to be of possible “hybrid” (retusaXmagnifica) origin.
It required yet another visit to flesh out distributions and this was to the inselberg Rooikop. We were not quite sure what to expect and I did think that H. floribunda would appear again. Instead we found a series of small populations of plants I attribute to H. heidelbergensis/magnifica ([28]Fig.50 MBB7217)
Fig. 50. MBB7173 H. retusa. NW Kweekkraal E[26].
As I have noted, variability in and between these populations is enormous. It is curious that H. floribunda only appears in the northeast of the study area and almost totally maintains its integrity where heidelbergensis/magnifica is excluded. The variant H. magnifica var. atrofusca is discrete in only one of these populations (Fig.42 – MBB7157) and this is the one exception where H. floribunda is also present. There seems to be a remarkably smooth continuum from H. magnifica-like to H. heidelbergensis-like plants.
I cannot say at all that Vlakte has been fully explored. There are many more possible habitats that we have not explored. While I am busy examining the evidence we have for a connection of mutica var. nigra to H. retusa, I find myself looking at the connection of H. heidelbergensis to H. magnifica with the implications of hybridisation (introgression is not a better word or concept, as both hybridisation and introgression imply an initial difference) involving H. floribunda and H. magnifica var. atrofusca which have rounded leaf-tips. Thus the saga continued.
Note: Kobus (J.D.) Venter made a presentation involving many of the above collections at the Aloe Congress at Calitzdorp in Sept.2003. This is due to be published in Alsterworthia. In his presentation he includes a geological map of the area and this shows that Vlakte is tertiary ferricrete overlying much older conglomerate rock. It is this ferricrete formation which provides a mosaic of small inselberg’s scattered throughout the southern Cape and playing a critical role in the occurrence of Haworthia).
Fig. 51. MBB7172 H. floribunda var dentata. N Kweekkraal E[27].Fig. 52. MBB7217 H. heidelbergensis. Rooikop, N Kweekkraal.[28].
Part 5: Continuing the saga.
On the way home after our last visit to Kweekkraal, Daphne and I stopped at Morning Star south of Heidelberg. We did this thanks to our friend Dr. Christie Kloppers who in discussing the distribution of H. marginata, let on that the owner of Morning Star was a relative of his. Morning Star is a locality recorded for H. marginata and as I had seen that species in the area many years before, I was anxious to confirm its survival there. As is the case in Haworthia there are so many considerations. Morning Star is not far from the Duiwenhoks river where the steep rocky riverbanks offer a continuum of habitats from north of Heidelberg all the way to the sea. A short way to the southeast is the locality Kransriviermond where H. mutica var. nigra has it origin. It is interesting to note that G.G. Smith made the connection of that population with three different accessions of H. mutica. Which goes to show how perceptions may differ – he had no compunctions about making that association as opposed to the opinions reported in the essay. The place Dassieklip is also on the lower reaches of the Duiwenhoks and this is where a very curious Haworthia is found which I have attributed to both H. floribunda and H. chloracantha. (Fig.54 MBB7164).
Fig. 54. MBB7164 H. floribunda=chloracantha. Dassieklip.
However, there is also something else. This is the collection MBB6663 (Figs.55a also MBB in KG107/74, & b JDV97-35, also Fig.27 in Pt.1) from immediately southwest of Heidelberg. This collection is cited in my revision under H. magnifica and that identification falls very far short of explaining its true identity or significance. I wrote an article in Aloe (34:4, 1997) discussing the problem of the “species” as they occur around Heidelberg. H. magnifica is barely mentioned and this particular collection suggested some introgression of H. floribunda. However, since that time I have come across several other populations to the west that must surely relate to this one at Heidelberg and impact on the relationship of H. magnifica to H. maraisii and H. mirabilis. Thus anything of this ilk to the south of Heidelberg becomes extremely important. That collection MBB6663 was instrumental in my early decision to treat H. maraisii and H. magnifica as one species
Fig. 55a. MBB6663 H. magnifica. SW Heidelberg.Fig. 55b. JDV97-53 H. magnifica. SW Heidelberg.
At Morning Star we were, by kind favour of Mr Coetzee Uys, able to get to the Duiwenhoks River. There we found H. turgida (Fig.56 MBB7219) on the rocks of the valley banks as we expected. It was surprisingly a rather small form (var. turgida) more reminiscent of its appearance on the sandstones north of Heidelberg, rather than the bigger var. longibracteata. But significantly we also found, in the immediately adjacent south-facing shale rocks, H. magnifica/floribunda (Fig. 57a & b MBB7218) as in MBB6663. The plants were smaller than that collection and there was more evidence of floribunda AND heidelbergensis in their appearance. We stopped and searched again about 3kms away on higher-lying ferricrete and in grass tufts, where we found plants of the same ilk (Fig.58a & b MBB7220). These two populations suggest fairly convincingly that H. magnifica and H. floribunda introgress (are continuous).
Fig. 56. MBB7219 H. turgida var turgida. Morning Star, S Heidelberg.Fig. 57a. MBB7218 H. floribunda=magnifica. Morning Star, S Heidelberg.Fig. 58a. MBB7220 H. floribunda=magnifica. Morning Star, S Heidelberg.
We were not finished. As a last effort we drove a short distance and stopped next to some exposed rock where we found H. mutica var. nigra (Fig 59a & b MBB7221). These are very dark-green almost black, shiny-leaved plants with the rounded leaf-tips we associate with H. mutica. One of the clones was almost identical to a plant of H. retusa (so-named) from north of Kweekkraal (MBB7173) where one could speculate that the rounded leaf-tips and darker colouration had some connection to introgression (that word again) with H. magnifica var. atrofusca. The second plant illustrated (Fig.59b) is somewhat of an aberration that I selected to emphasise that identifications are often very tenuous.
Fig. 59a MBB7221 H mutica var. nigra, Morning Star, S HeidelbergFig. 59b. MBB7221 H. mutica var nigra. Morning Star, S Heidelberg.
In my discussion of H. incurvula in Aloe ( 36:34, 1999), I suggest we need a new language to discuss, explain and name haworthias. We need a better conceptualisation of names and what they mean. In taxonomic botany, the name of a species is linked to a “type’ which is desirably a herbarium specimen, but it could be an illustration of some kind. It is often simply assumed, perhaps even unconsciously, that this “type” is typical. We all (nearly all) assume that anything so named will be identical or near identical to that type. Conversely we assume that anything different from the type should have another name and ipso facto be another species. The concepts of subspecies and varieties are an extension of this way of thinking. The nomenclatural code which seeks uniformity and stability in the way we name plants cannot legislate or prescribe when and where names should be applied and nor at what rank. Where it does seem to fall short is the requirement for the automatic creation of a typical rank when an equivalent rank is created. Here it seems to be making the same assumption that all plants of a species are the same. Thus when a new, say, variety is described, there is a typical variety that stands in juxtaposition to the new. This is patently fallacious. What is required is a re-description of the species to include the variation of the new lesser rank, and only those individuals which meet the varietal description become identified by an added name. The residue forms the core of the species and are designated by only that name?
What we need in addition, is the adoption of a definition of a species as a system of living organisms which is named by a convention recognised as a language requirement, and not as any scientific process or achievement. The name is only the passkey to information – the key by which we generalise about a particular living system. (I have since written a chapter which suggest that species are fractal, and that we will need to adapt our thinking to this kind of geometric/mathematical pattern).
Part 6: Another twist to the story.
Until recently I had seen very little of the haworthias immediately west of Heidelberg. Kobus Venter has several collections, some by J Esterhuizen, and these included plants from Uitkyk immediately southwest of the town (Fig. 60a & b JDV89/2) as well as from further west. He also has two collections from immediately north of the town where previously I have referred to only one. Taking these first, I think it is quite clear than they both suggest the continuity of turgida with heidelbergensis, while one of the collections suggests a closer similarity to the plants from Klipdrift of Part 1. The Esterhuizen collection from northwest of Heidelberg (Fig.61 JDVsn) seems to support such a contention.
Fig. 60a. JDV89-2 H. heidelbergensis. Uitkyk, SW Heidelberg.Fig. 60b. JDV89-2 H. heidelbergensis. Uitkyk, SW Heidelberg.Fig. 61. JDVsn H. heidelbergensis. NW Heidelberg.
But I really want to deal with plants west of Heidelberg. The locality is referred to as Die Plotte but actually the area is partly township land, an area of smallholdings and sections of what was a very large farm Hooikraal. Dekenah and thus G.G. Smith original site of heidelbergensis may be non-existent now because of the creation of a rubbish-dump site immediately west of the town. Plants similar to the original heidelbergensis can still be found southwest and further west (and of course immediately east of Heidelberg which I had concluded (wrongly and influenced by Dekenah) was the type locality. Kobus’ collection from Die Plotte (Figs. 62a, b & c JDV91/142) are of smaller plants while JDV91/146 (Figs.63a & b) from the same general area, are bigger plants. These are intermediate with the Esterhuizen plants northwest of, and typical heidelbergensis from east of Heidelberg (Figs. 64a, b JDV87/1 & c MBBsn).
Fig. 62a. JDV91/143 H. heidelbergensis. Die Plotte, W Heidelberg.Fig. 63a. JDV93-146 H. heidelbergensis. Die Plotte, W Heidelberg.Fig. 64a. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 64c. MBBsn H. heidelbergensis. E Heidelberg.
When Kobus delivered his paper at the Aloe Congress 2003 at Calitzdorp he met up with Mr. J. Hoffman. I had met him at Worcester many years before and he had told me of plants west of Heidelberg. Thus I undertook to renew this contact and went to see him at Heidelberg. He took me to the northeastern corner of the farm Hooikraal which borders on Die Plotte and which is also west of the town dumpsite. Apart from showing me H. marginata, he also showed me a population of small heidelbergensis (Figs. 65a, b, c & d MBB7233) and then a population of very large plants that were quite remarkable (Figs. 66a, b, c & d MBB7234). The plants included one that even resembled a large sturdy mirabilis – and Esterhuizen commented in an article he once wrote about the spotting on the back of the leaves of “heidelbergensis” which he likened to mirabilis. Generally the plants are with darker green and purplish hues that I associate with heidelbergensis or even magnifica. There were very yellow-green forms similar to what I saw many years ago at Skeiding further west of Heidelberg. These are plants that Esterhuizen (and myself too) has referred to as retusa, and I would agree except that they fall in the query zone between retusa and turgida var longibracteata i.e. there is continuity between the two species. In fact Kobus and I revisited Skeiding and I thought the plants (Figs.67a & b MBB7240) were almost identical to H. retusa at Melkboom (Fig.26 MBB7107 in Part 4), although the latter tended to have rounded leaf-tips. H. magnifica, MBB7239, was also present in a rare situation for these two species to be so closely located within meters of each other). We also visited a population immediately north of Heidelberg (Figs. 68a & b MBB7236) that is not far distant from the two Nature Reserve population already referred to. Had I seen this population in 1975, I may have been more adamant in maintaining that H. retusa and H. turgida were but one species. The plants were generally big and very proliferous and one could not help reflecting on how similar they were the population that gave rise to Scott’s H. geraldii (H. retusa var.). The collection from northwest of the Heidelberg Nature Reserve (Figs. 69a, b & c JDV89/5) must surely suggest that there is a continuity of H. turgida with the Klipdrift plants illustrated in part 1.
Fig. 65a. MBB7233 H. heidelbefrgensis. N. Hooikraal, SW Heidelberg.Fig. 66a. MBB7234 H. heidelbergensis. S Hooikraal, SW Heidelberg.Fig. 67a. MBB7240 H. retusa. Skeiding, W Heidelberg.Fig. 68a. MBB7236 H. turgida=retusa. N. Heidelberg.Fig. 69a. JDV89-5 H. turgida=retusa. N Heidelberg.
My conclusion is that there is now a proven continuity between turgida, heidelbergensis, magnifica, retusa, mutica var. nigra, maraisii, pygmaea and even floribunda, chloracantha, rossouwii and variegata. The continuity is also with emelyae and its varieties north of the Langeberg mountains. At the same time some of these elements may occur together and so appear to be discrete. Thus a taxonomic solution for one set of collections may not be appropriate for another.
Paul Forster and Russel Scott (both from Australia) spent some time in South Africa while attending the Calitzdorp Congress, and I was privileged to have them join me in the field at Heidelberg. We explored the middle Duiwenhoks river between Dassieklip and the town of Heidelberg. I had expected that such exploration would throw some light on the strange Dassieklip population (Fig.56 MBB7164). In my Revision (1999), I have ascribed it to H. floribunda simply for the want of a better solution outside of regarding it as a discrete species based on a single population!).
Apart from seeing H. turgida we also found find populations of plants at Witheuwel (Figs. 70a, b & c MBB7227) and Somona (Figs71a, b & c). These are more like the Melkboom and Kweekkraal heidelbergensis/magnifica collections. I would have expected a similarity to MBB6663 (SW Heidelberg), referred to in part 5, figs 2 & 3) or to MBB7218 and MBB7220 which I would ascribe to floribunda/maraisii rather than to H. magnifica – tenuous as such a distinction is. Essie Esterhuizen has apparently discovered a similar population to KG107/74 along the Duiwenhoks close to of Heidelberg. It must be noted that Matjiestoon, the locality for H. heidelbergensis var toonensis (Figs.72a, b, c & d MBB7165) is only about 15km west of Dassieklip. Thus while I am writing in the context of H. mutica var. nigra, I have drifted helplessly into discussing H. heidelbergensis. This is the crux of the whole thing about Haworthia – especially the subgenus Haworthia – an intricate network of relationships that just will not resolve.
Fig. 70a. MBB7227 H. heidelbergensis. Witheuwel, S Heidelberg.Fig.-71a.-MBB7229-H.-heidelbergensis.-Somona-S-Heidelberg.Fig.-72a.-MBB7165-H.-heidelbergensis-var-toonensis.-Matjiestoon.-SW-Heidelberg.
Part 7: Completing the saga.
So far in this series of articles I have dealt with H. mutica var. nigra as a system of populations typified from the southernmost population at Kransriviermond, and extending through eight known populations to Klipdrift northwest of Heidelberg. Then I considered the role of H. turgida in part 3. In part 4 I dealt with the very complex situation that seems to occur between Riversdale and Heidelberg, followed in part 5 by discussion of populations along the lower Duiwenhoks River. In part 6 the discussion and illustrations are of plants from mostly west of Heidelberg.I have discussed to some degree the possible relationship of these populations to each other to H. turgida, H. retusa, H. heidelbergensis and H. magnifica. Each of those systems in turn interacts with others and it becomes highly doubtful that we are in fact dealing with discrete systems at all.
If we now switch the attention to H. mutica and point to some of the populations that constitute my concept of that species, it may help to crystallize aspects of my classification. I will not deal with all the material at my disposal as is too extensive for the purpose. The name H. mutica is based on an illustration in the Royal Herbarium, Kew, of a plant of unknown origin (Fig.73). I use the name for a series of populations all west of the Breede River, from near Drew in the north, to near Bredasdorp in the south. These populations all seem to be discrete from other species in that general area, which are H. mirabilis, H. maraisii, H. rossouwii, H. heidelbergensis, H. reticulata and H. turgida. Those species display their own interactions as follows:
Fig. 73. H. mutica. Kew herbarium.
H. mirabilis never occurs in any close association with any other species (of the same subgenus) including H. maraisii or with H. heidelbergensis. Where it may, is where H. mirabilis var. badia occurs in close proximity to H. maraisii at Napier. I can be suggested that here the two elements are introgressive and that var. badia is actually the consequence of this introgression (again the word implies interaction between different things). H. maraisii never grows in association with H. heidelbergensis except for the place Rooivlei north of Bredasdorp where this H. heidelbergensis var. minor could in fact be a variant of H. rossouwii. It should be noted that H. turgida var longibracteata also occurs here as a very isolated population.
If one extends these observations it will be found that very few species maintain their identity in close proximity to any other. H. magnifica may occur in close proximity to H. tugida or H. retusa, but the latter two never to each other. H. magnifica is not discrete in any sense from H. maraisii and two species are recognised simply as a device to assist discussion where we have a western (maraisii) and an eastern (magnifica) counterpart. Neither of these ever occurs near to, or with, H. heidelbergenis. We actually need to again consider this last-named species very closely. It is typified from a population just west of Heidelberg (as Esterhuizen rightly pointed out, and it is by fortunate coincidence that my assumption drawn from Dekenah that it was east, does not alter the basic perception of what the species looks like). H. heidelbergensis has by default, and from simple circumstance and complex reality, become a hodge-podge of populations which interdigitate between H. mirabilis, H. maraisii and H. magnifica. H. floribunda is a curious anomaly that is widely distributed and is known to hybridise with H. turgida, H. magnifica var. atrofusca and with H. pygmaea var. argenteo-maculosa. There is some evidence that it introgresses with (or has never segregated from) H. magnifica (synonymous maraisii) south of Heidelberg, and similarly with H. heidelbergensis (synonymous also with maraisii) south of Swellendam.
H. mutica only grows in the close proximity of H. mirabilis at Rondeheuwel south of Stormsvlei. At Sanddrift, Drew it is in close proximity to H. heidelbergensis in a form which is akin to H. maraisii. Further to the southeast it is in close proximity to H. maraisii. At Napky these two elements are also close.
My argument for maintaining H. mutica as discrete from H. retusa was based on the observation that it was never necessary to invoke H. turgida in the equation as it is when discussing H. retusa. It also is never introgressive with H. maraisii as H. retusa seems to be with H. magnifica east of Riversdale, nor with H. heidelbergensis as H. retusa seems to be at Kweekkraal. Nevertheless this decision is not indisputable. I have always borne in mind that H. mutica has some affinity with H. pygmaea. H. mutica seems to have a retusa-like origin whereas H. pygmaea has a magnifica-like origin. In neither case can one be sure that H. turgida is not prominent in the ancestry.
A not-so-curious thing about H. mutica is its variability too. Just like any of the other “species”, it has many faces. The one I would like to mention is the similarity of plants of H. mirabilis var. badia from the western-most known point of its distribution. This is not readily apparent. But is we look at plants at the western end of the population viz. Sandfontein Napier (Figs.74a & b MBB6987), the plants begin to depart from the more classical form with the strongly recurved leaves, and begin to resemble more typical var. mirabilis as at Mierkraal Bredasdorp (Figs.75a & B MBB7090). What has astonished (even) me is that some of the clones form a very close match with H. mutica, especially with south (Figs.76a & b JDV96-16) and north of Napky (MBB7031 respectively). Thus we are now thrown into (an admittedly obscure) relationship of H. mutica and H. mirabilis (there is also a similarity with a clone of H. magnifica from the problematic “south of Tradouw site” (Fig.77 MBB6666). To my knowledge H. mutica and H. mirabilis only occur in close association at one place. This is at Rondeheuwel south of Stormsvlei where the mirabilis is atypical and evidence of the continuity it displays in so many places with H. maraisii. My collection (Fig.78 MBB6982) from Hasiesdrift north of Bredasdorp seems to be a good representation for a concept of H. mutica. The habitat photo (Fig.79), however, raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.
Fig. 74a. MBB6987 H. mirabilis var badia. Sandfontein, Napier.Fig. 75a. MBB7090 H. mirabilis var mirabilis. Mierkraal, Bredasdorp.Fig. 76a. JDV96-16 H. mutica. S Napky, SW Swellendam.Fig. 77. MBB6666 H. magnifica var magnifica! S Tradouw Pass.Fig. 78. MBB6282 H. mutica. Hasiesdrift, N Bredasdorp.Fig. 79. MBB6282 H. mutica. Hasiesdrift habitat picture.Fig. 79. MBB6282 H. mutica. Hasiesdrift habitat picture.
There the saga for the moment must rest. The proper taxonomic place of H. mutica var. nigra sensu Bayer is simply not very significant given the huge problem which is only touched on here. This is the interplay of H. heidelbergensis with H. maraisii, H. magnifica, H. mirabilis, H. turgida, H. retusa, and others. I should say of all the populations (should I count them?), I know which can be attributed to these “species”. ♦
This problem of continuity is one I seem to have difficulty in conveying to my readers and listeners. The difference between one species and another is a discontinuity and, if we believe in evolution, it is the resultant of a break-up of continuity in its ancestral parent species. The “model” we have in our minds, is of progressive change from one recognisable entity to another by evolution. Geographic distribution and re-distribution are key elements in this process. But we do not seem accept this in the way we try to classify plants or interpret classifications. Apart from recognising that change could be gradual and therefore manifest continuity, the change may be from a complex variable system which contains different levels of continuity within itself, and not from a simply understood uniform ‘ancestor’.
The result is that in a genus like Haworthia, which is by no means exceptional, the differences between species i.e. the discontinuities between “species”, may be very difficult to either recognise or rationalise. It in fact becomes a statistical operation in which all the characters should be involved i.e. multiple variate analysis. If all the characters could be measured and quantified it is statistically possible to subject all the data so obtained by one of several statistical methods to measure “distance” and “significant difference” between groups of plants which we want to ascertain are species, varieties or even just hybrids. The process of “cladistics” is the use of a system to generate a branching “tree” of relationships base on characters which are also evaluated and loaded for chronological priority (primitive versus advanced). In using such a mathematical package, it is pretended that the classification becomes “objective” and hence replicable to satisfy the scientific requirement. In my estimation, the cladistic process assumes that a two-dimensional “tree” adequately represents the spatial and temporal changes of evolutionary processes, and it does not work.
Somebody might one day try to apply such methods to Haworthia and I say “Good luck to you”. My experience of characterisation and variation in the biological systems I have experience of, and including Haworthia, suggest to me that sensible, practical, experienced “eye-balling” will prove the better bet. Ultimately in Haworthia, I expect that technology and cladistic methods will be testable on the result of my classification. This is not a conceited and arrogant claim. It is a simple reflection on what classification actually is and what it is for. Much of botanical classification has been done by amateurs with no, or minimal, specific training and qualification for that field at all e.g. G.W. Reynolds, L.C. Leach, T.L. Salter, J. Lavranos, C.L. Scott, G.G. Smith, M.B. Bayer etc. Their classifications form the basis of many scientific observations, sometimes by scientists who have no conception of the significance, or insignificance of the names they use or what they may actually mean. The classifications may have little to recommend them except the fact that they appear to conform to the approved nomenclatural style.
Putting that all aside I want to just demonstrate by means of a series of illustrations, what occurs in Haworthia. I have described several situations before but what follows is another account of the continuity which occurs in the southern districts of Somerset East, Eastern Cape. The main area concerned is Kaboega at the north and western Zuurberg near where it fades away north of the Klein Winterhoek Mountains (see map Fig.1). One thing should be engraved on the mind. The following comparisons are made on field impressions and on multiple plant samples. This at least takes note of statistical requirements and considerations and it would be wrong to try and use the individual illustrations to arrive at some other conclusion.
Fig. 1. Map of the western Zuurberg, Kaboega Farms.
Determining a starting point is quite difficult because there simply is none unless a second problem is addressed. This is the one of typification in which names are fixed to a specimen of some kind. After this is done there is the question of how the name is applied to the body of the plants from which it comes. In this article, the question of the application of the names cooperi, pilifera, vittata and blackbeardiana is involved and so I reproduce the illustrations accompanying the origins of these names (Figs.2, 3, 4 & 5). It is my experience that in the field, it is possible to find resemblances to these illustrations in a large number of geographically disparate populations, and hence quite a problem to apply them in a consistent and sensible way. Only the name blackbeardiana can unequivocally be said to have been derived from any particular field population and this is at Bowe’s Farm near Imvani, south of Queenstown, and the single illustration by no means represents all the variants of the many populations which can be said to belong together here. Some of those variants invoke the use of the other names. How all these names are used is something of a personal choice and mine is laid out in the book Haworthia Revisited which I wrote in 1999.
Fig. 2. H. cooperi Baker. Refugium Botanicum, 1871.Fig. 3. H. pilifera Baker. Refugium, Botanicum 1871.Fig. 4. H. vittata Baker. Refugium Botanicum, 1871.Fig. 5. H. Blackbeardiana VPoelln.,1937.
In this chapter I will take a collection (originally by G. Marx) from east of Alicedale and call it H. cooperi var. cooperi ([1]…denoting position on map Fig.6. MBB6845). In doing so I will have to point out again that there are further intrinsic difficulties in the requirements of nomenclature, which is that there is such an entity as a “typical” variety. My belief is that any plant which is not directly assignable to a variety of a species should simply be referred to as that species. Thus the Alicedale collection is H. cooperi var. cooperi and geographically in the distribution area of H. cooperi var. pilifera. The plant illustrated is of a single clone, but the plants in this particular population relate better to populations in the geographical area of the typical var. cooperi. The plants occurs here together with H. angustifolia and H. cymbiformis (in the same indeterminate varietal format as the H. cooperi, but for geographical reasons mainly, this population is referred to as H. cymbiformis var. obtusa. A similar form of H. cooperi depicted recurs on the Zuurberg and I saw it at Oudekraal when I explored there for H. angustifolia var. baylisii ca 1982. I have plants in cultivation from there which were collected by Ernst van Jaarsveld, and I made a new collection MBB6922 slightly west of there in 1999. I saw it again at Vyeboomfontein ([2]Fig.7 MBB6949) much further west and again just north of that at Klipfontein [2] MBB7049. I am unsure if the identification should be var. cooperi or var. pilifera but choose the latter.
Fig. 6. MBB6845 H. cooperi var cooperi. E Alicedale.Fig. 7. MBB6949 H. cooperi var pilifera. Vyeboomfontein.
Slightly further west at DePlaat, the problem starts with plants which are greener (particularly on the south slopes) and with longer more attenuate leaves. There is first the collection from north slopes ([3]Fig.8 JDV91/14) that I attribute to H. cooperi var. gracilis, and then mine from the south slopes ([4]Fig.9 MBB6935). Near the Kaboega farmhouse at Koksedam there is also a greenish element and two plants are illustrated ([5,6]Figs.10a & b MBB6915) to show that it is partly cooperoid and partly cymbiformoid. I propose calling these, with MBB6935, H. cooperi var. viridis because it is complicated (it could invoke the name gracilis of von Poellnitz, but I described a smaller green variant from near west as var. viridis, and this name seems more appropriate). I have, in Haworthia Update Vol.1, described that there is this greenish archetypal form which is quite widely spread and often in an indeterminate zone, between forms of H. cooperi and H. cymbiformis. On Kaboega these plants take three elemental directions:
Fig. 8. JDV-14 H. cooperi var gracilis. DePlaat.Fig. 9. MBB6915 H. cooperi var viridis. De Plaat.Fig. 10a. MBB6915 H. cooperi var viridis. KokseDam.Fig. 10b. MBB6915 H. cooperi var viridis. KokseDam.
1. Directly west to Spekboomberg ([7]Fig.11 MBB6914) and then to Wilgerfontein ([8,9]Fig.12a & b MBB6925) at. At Fig.11 the plants are consistent with Fig.9. At Fig.12a & b the plants resemble H. cymbiformis. Some clones are more similar to that species than others while odd clones are entirely indistinguishable from it. I find it difficult to determine what species it is, never mind add a varietal name, because of its obvious intergradation with H. cooperi variants and its geographical disassociation with H. cymbiformis. My suggestion is that this is where the case calls for the use of the species name, without the addition of the typical variety epithet. But even that is unsatisfactory here because I think the plants, despite their outside similarity with cymbiformis, are actually in the same genetic bag as cooperi var. viridis. It is worth stating that there is a transition from sandstones at DePlaat and eastwards, to Dwyka tillite westwards to Wilgerfontein.
Fig. 11. MBB6914 H. cooperi var viridis. Spekboomberg.Fig. 12a. MBB6925a. H. cooper var viridis=cymbiformis. Wilgerfontein N.Fig. 12b. MBB6925 H. cooperi var viridis=cymbiformis. Wilgerfontein N.
2. Slightly to the south-west, entering the Kaboega gorge is JDV92/33 ([10]Fig.13). This is indeterminate and I would label it “graciloid”, although there is evidence of other variants in the gorge which do not agree with the concept of var. gracilis. That name itself presents problems as described in Update Vol.1. Fig.14 is in sandstones, and on the steep north facing slopes (also sandstone) of Wilgerfontein a little to the west and south of MBB6925, is MBB6904 ([11]Fig.14). I would suggest that this resembles H. cooperi var. gordoniana (subsequent to drafting this manuscript I did a limited exploration south of the Zuurberg, and found a fairly similar populations occurs at [=11] in the Addo National Park. But it is an astonishing ecotypic variant too, as 25 meters away is MBB6905 ([12]Fig.15) which is “isabelloid“. I have coined the name “puberula” for it. Bear in mind that these pictures represent only samples of many plants in small discrete habitats.
Fig. 13. JDV93-33 H. cooperi ‘graciloid’. Kaboega Gorge.Fig. 14. MBB6904 H. cooperi ‘graciloid’. Wilgerfontein S.Fig. 15. MBB6905 H. cooperi ‘puberula’. Wilgerfontein S.
3. In a north-westerly direction from DePlaat is MBB6943([13]Fig.16), which is slightly spinose and indeterminate like the MBB6915 (Fig.10) collection. But it gets very curious. If we ascend the DePlaat hillside towards Klipfontein and [2]MBB7049, we find first MBB6910 ([14]Fig.18) almost indistinguishable from MBB6905 and close to MBB6904. A little further, and less than 250meters from MBB7049 is MBB7017 ([15]Fig.19), a small graciloid=puberula population in which the plants are very small and the leaves have surface spination. If we cross the valley and go now north of Klipfontein to the upper slopes, we find MBB6940 ([16]Fig.19). It is indistinguishable from MBB6904! If instead we stick to the low lying areas and move north of Koksedam, and northwest of DePlaat, there is first MBB6942 ([17]Fig.20) and then MBB6917 ([18]Fig.21). This latter is a relief because I have several collections from widely dispersed points [=18] to suggest these are my version of H. aristata. Not forgetting that the initialisation of that concept at Soutkloof near Addo, also involves H. cooperi. If we look at one collection from Commadagga ([19]Fig.22 MBB6897) we see a plant which is a variant of H. aristata (it could be cooperi var. pilifera!) that has look-alikes at Kirkwood in the arena of H. cooperi var. pilifera. But at Kaboega, the plot thickens further. Going still northwest from DePlaat and now north of MBB6943, is MBB6916 ([20]Fig.23). There is a problem again, and one has to go quite a way (about 20km) to find MBB6899 ([=20]Fig.24). The latter is quite a large plant in the mould of H. aristata. To find a better home for it among several other collections, including some dubious ones to the east and north at Ripon, and including Col Scott’s H. pringlei, I added it there while reducing Scott’s species to H. bolusii var. pringlei. This problem is elucidated elsewhere but I have not decided where best to place MBB6916 I think for practical and geographic reasons they could be placed under H. aristata, but they could equally be put under H. bolusii var. pringlei. I described H. decipiens var. virella to lessen tension on my use of the name pringlei and its relation to H. bolusii var. blackbeardiana and to H. cooperi var. dielsiana (includes Scott’s H. joeyi). While it does so in some areas of the classification, some tension does remain.
Fig. 16. MBB6943 H. cooperi var viridis. Dassieklip.Fig. 17. MBB6910 H. cooperi ‘puberula’. DePlaat.Fig. 18. MBB7017 H. cooperi ‘graciloid=puberula’. Klipfontein.Fig. 19. MBB6940 H. cooperi ‘graciloid’. Klipfontein N.Fig. 20. MBB6942 H. cooperi’graciloid=aristata’. DePlaat Weir.Fig. 20. MBB6942 H. cooperi’graciloid=aristata’. DePlaat Weir.Fig. 21. MBB6917 H. aristata. Kaboega.Fig. 22. MBB6987 H. aristata=cooperi var pilifera. Commadagga.Fig. 23. MBB6916 H. bolusii var pringlei=aristata. Spitzkop, Kaboega.Fig. 24. MBB6899 H. aristata. Paddafontein.
This link to the above variants is enhanced by MBB7012 ([21]Fig.25) from Buffelsnek, overlooking Lake Mentz and west of Wilgerfontein. This collection is also labelled H. cooperi ‘graciloid’ as I have done for MBB6904, but it could just as well be H. aristata or even bolusii var. pringlei (because of the nature of continuity offered by other populations to the northwest [=21]. To show just how complex the situation is, I illustrate JDV92/44 (Fig.26). This was initially a P.V. Bruyns collection from Inverbolo, far distant and northeast of Cathcart. I specifically went there to confirm (MBB6942) that this is H. bolusii var. blackbeardiana. I have said elsewhere that there is a problem of this question of continuity and that H. decipiens can be indistinguishable from H. bolusii var. blackbeardiana. To add insult to injury I illustrate MBB6730 (Fig.27). No! It is not H. aristata, it is H. mucronata var. habdomadis from Seweweekspoort in the Little Karoo, and from an entirely different set of populations. It is in fact extremely disconcerting to have to admit that the green plants of Figs. 9 or 10 can similarly shown to be indistinguishable from H. mucronata var. morrisiae, also from the Little Karoo.
Fig. 25. MBB7012 H. cooperi ;graciloid’. Buffelsnek, W Kaboega.Fig. 26. JDV92-44 H. bolusii var blackbeardiana. Bolokei, NE Stutterheim.Fig. 27. MBB6730 H. mucronata var habdomadis. Seweweekspoort, E Ladismith.
In writing this, I hope that readers will now get to understand that in apparently attacking the work of aspirant Haworthia taxonomists, I am not trying to defend myself or my system. I simply recognise that these persons do not have the competences or the insight to follow and understand the difficult choices that have to be made any better than myself. The main point is, that I know that if we explore still further west of Kaboega (and there are collections which prove this), and then into the Klein Winterhoek mountains to the south-west, still further problems are going to unravel (us) as we roller-coaster between H. decipiens and H. cooperi all the way to Uniondale and beyond. These same problems bedevil most of the species systems in the southern and western Cape. Unless one understands and respects the intricate geographic relationships, the use of the names in the way I recognise them will perhaps not make sense and I truly see no other way in which a nomenclatural system and classification, which will work in the field, can be derived for Haworthia. ♦
With Tony Dold, Selmar Schonland Herbarium, Grahamstown. This essay was published in Aloe 40:10, 2003. Minor additions are made here.
Introduction
By way of introduction it is fitting to repeat Dyer’s (1937) note on the Karroid Scrub of the Albany Division … “Pluto’s Vale and Hell’s Poort, names suggesting places of an unenviable reputation, are on the roads from Grahamstown through the Fish River valley to Kingwilliamstown and Bedford respectively. Certainly very hot passes during the summer, they are nevertheless the homes of many botanical treasures, particularly succulents. Several species of Haworthia are present (in the Albany division) but the soft leaved ones are usually found under the protection of rock ledges or scrub. Pluto’s Vale is the home of two rarities, H. incurvula and H. tenera, both described recently by von Poellnitz”.
The genus Haworthia is fairly difficult to classify, but Dyer’s words reflect the uncritical way in which any new species or variety is accepted into the literature. Bayer (Aloe 36:34, 1999) discusses the two ‘species’ noted by Dyer and the complexity of their relation in Pluto’s Vale. Bayer did not deal with the geological factors which might underlie their relationship, but this is implicit in the various books and papers he has authored and the reliance he has placed on geographic considerations. In a new publication (see chapter 7) Bayer has also explored the relationship of Haworthia species on the northern slopes of the Zuurberg in the Kaboega area. Here it is evident that geology and habitat are key factors in determining relationship between similar elements to H. incurvula (H. cymbiformis var. incurvula) and H. tenera (H. gracilis var. tenera). In both cases the same key species viz. H. cooperi (to include H. gracilis) and H. cymbiformis, are involved. As in the case of Pluto’s Vale, the species, H. gracilis comes into question too. This latter species has a questionable taxonomic history, but it was at the last considered to have originated in Helspoort (Afrikaans) to the north-west of Grahamstown. While mentioning here, based also on recent publication of a possible type illustration (Breuer, 2000), that it may not have come from Helspoort at all, and may in fact be synonymous with what Bayer (1999) has taken to be H. aristata. As discussed in Haworthia Update, there is a problem with the recognition of the Helspoort “species” in the face of the broad range of variant populations which constitute H. cymbiformis, H. cooperi, H. bolusii, H. gracilis and H. aristata and all the varieties associated with them. A recent excursion was made to Helspoort to establish if the variation of the Zuurberg, Pluto’s Vale and the Baviaanskloof Haworthias was also evident.
We speculate that the difficulty in dealing with genera like Haworthia, is that such small succulents favour skeletal sites where inter-plant competition is low because of shallow soils and surface rock. It has been noted several times that a species may change facies depending on site and this has been reported for H. mucronata var. mucronata as it occurs on shales, and H. mucronata var. habdomadis as an ecotype of the same species occurring in sandstones (Bayer, 1999). A similar situation has been reported for H. cooperi var. pilifera on Ripon Sandstone, as opposed to H. bolusii var. blackbeardiana on shale near Ripon Station (Bayer, in ms.). Broad indications are that elements as different as H. cooperi var. pilifera and H. cymbiformis var. cymbiformis may be ecotypic, because the former is essentially a flat level grassland, and the latter a riverine-cliff element.
Apropos to Dyer’s note, it is only the two Haworthia species and H. attenuata var. britteniana that lend any particular significance to Pluto’s Vale, however diverse and extraordinary the vegetation there may be. There is no single plant species that lends any particular significance to Helspoort. The same in fact can probably be said for the whole Kaboega area of the Zuurberg, where four biomes are said to meet (Bruton and Gess, 1988). Recently there was a report on Faucaria nemorosa (Aloe 38:37, 2001) from Swartwaterspoort. My immediate response is to ask … “What is peculiar about Swartwaterspoort and how does this relate to Haworthia?” A “species” in a landscape as “ordinary” as Swartwaterspoort should not be expected to harbour something uniquely discrete. The answer is that Haworthia cymbiformis does occur there and its variants are notable (Fig.8), but barely outside of the context of the variability of the genus itself. Thus the broader question of vegetation can and should be examined when establishing the classification of Haworthia, or in fact any other genus. The object thus of this paper is to report findings at Helspoort and discuss, together with the already reported observations at Pluto’s Vale and Kaboega (Zuurberg Mountains), the taxonomic implications relative to vegetation and geology.
Observations The details of the Haworthia populations observed for Pluto’s Vale and Kaboega are given in Haworthia Update. It was only possible to spend one morning at the Helspoort site and four collections of H. gracilis were made. These are:
MBB7051, on an upper south-west facing slope on the edge of Bush thicket, west of the road
MBB7052 (Figs.3, 4, 5), in a low lying level area at the base of the valley, under Pteronia incana.
MBB7053 (Figs 1, 2), on a rocky north facing slope also low down in the valley, with Euphorbia polygona.
MBB7054 (ex T. Dold), in rocks above the valley on the north-eastern side.
The plants are all very similar, unlike the Pluto’s Vale populations and certainly not as diverse as the Kaboega ones. None were as spinose as the unpublished Berlin illustration (Fig. 6), but they do accord with the Desert Plant Life illustration (Fig.7) which has otherwise been taken to represent the species (unfortunately more significance is attached to, and more is written about, frivolous aspects of nomenclature rather than the use and application of names). The plants are pale green in colour and moderately to almost non-spinose. In the population MBB7052 under Pteronia incana, the plants were more blue-green in colour with slightly more obtuse and thicker leaves suggesting closer similarity to H. cooperi var. pilifera. This element is quite common in the area and occurs in more typical form both to the east at Piggots Bridge, nearer to Grahamstown at Table Farm and also at Vaalkrans, Thornkloof and Brakkloof farms. H. cymbiformis is not known closer than Swartwaterspoort west of Riebeek East (Fig.8), at Alicedale, at the farms Thornkloof in the south, and The Fort to the east. There are no recent confirmed collections of a gracilis-like element in the Grahamstown area, other than as mentioned in Bayer (1999). The closest affinities are a glabrous element from Gladhurst and a similar element from Elandskop, both south of Adelaide. These are similar to the cooperi-like elements described for the Zuurberg (Fig.9). H. aristata is nearest at Kommadagga, west of Riebeek East. This is certainly a product of under-exploration and unconfirmed collections from the Brakkloof ridge running east and west of Helspoort, as well as from the Fish River Rand heights north of the Fish River all need to be explored.
Geology The basic geology and topography is illustrated in Figs 10-14 – as accurately as the original maps permit. The geological formations at each area are essentially the same, particularly for Pluto’s Vale and Helspoort.
These are Dwyka, Ecca and Witteberg groups, which are characterised as follows:
Dwyka – tillite, consisting of dark blueish‑grey, very poorly sorted, massive diamictite of glacial origin.
Ecca – shale, carbonaceous shale & tuff, Fort Brown shale formation, Ripon sandstone & shale formation.
Witteberg – quartzite of the Witpoort formation and Witteberg shale, quartzite, sandstone & diamictite of the Lake Mentz subgroup.
Soils The underlying geology ultimately determines the soil type although climatic factors in the weathering process would have considerable effects on the result. Because the underlying geology of all three study sites is very broken and incised, the soils are directly influenced by geology than in the case of more mature formations that are less incised or topographically less rugged and broken. Thus some of the habitats are practically skeletal and parent rock, and what soil there is overlies rock or partially weathered rock (Hartmann 1988). Pluto’s Vale (Figs.11,13,16,17) is characterized by weakly developed lime-rich soils, while Helspoort (Figs.10,12, is similarly characterized by weakly developed soils of rocky veld with a strong sandstone contribution. The western Haworthia locality in Helspoort is on Witteberg Quartzite, where the soil is thus sandy, nutrient poor and well‑drained. The central locality is on shale while the eastern locality is on Dwyka tillite that both result in a clay rich soil with more nutrient and greater water holding capacity. The plants from Pluto’s Vale are also on Dwyka tillite and on Ecca shale, sandstone is absent. The Kaboega area (Figs 14,15) is at another scale and almost any aspect and soil type concordant with the geological formation and broken topography can be encountered. In addition, at Kaboega there is a declining rainfall gradient from east to west, as well as rain-shadow effects.
The subdivisions and variants of the parent geology make it extremely difficult to generalise about the soil and the scale of the geological maps also make it be quite inappropriate for comment on soil. Furthermore there is the question of aspect too. South slopes are cooler and moister than north slopes and soils tend to be deeper. Deep rocky ravines result in sites which are shaded and cooler for different periods of the day. On the scale of Kaboega, altitude is also critical and a rainfall gradient is pronounced as opposed to Helspoort and Pluto’s Vale localities where site moisture is very locally determined.
Rainfall Rainfall can be related to distribution of Haworthia species, but we do not think it is significant except in respect of the scale at which we are dealing. The study sites fall in a zone of approximately equal summer and winter rainfall, often with good late summer rain in March, which have a carrying over effect well into the winter period. However, the higher ridges and hills receive a greater rainfall than the lower valley levels, which also have higher temperatures. Rain coming from the coast is generally intercepted by Botha’s Ridge at Grahamstown and thereafter rainfall decreases greatly decreases towards Committees Drift and Fort Brown (Dyer 1937). The average annual rainfall at Pluto’s Vale is 500mm while that of Helspoort 482mm. The average annual rainfall at Kaboega is significantly lower at 289mm (Dent, Lynch & Schultze 1987).
Plants on the higher mountain slopes with a south or south-eastern aspect will also benefit greatly from condensation from mists from the sea, mostly during summer months. The Haworthia population at the higher western‑most locality at Helspoort would certainly be affected by these incoming mists whereas those on the central and eastern localities, being on the leeward side of the catchment, would not. Moreover the latter would experience higher temperatures than the former. At Kaboega, the entire Zuurberg range intercepts rain and mist from the south-east, and there is a rainfall gradient declining from east to west.
Vegetation Haworthia and many smaller plants are elements of the broader vegetation and usually constitute that element which deviates from the characteristics of any recognised phytochorion. This is probably because they cannot compete against the larger plants, grass, trees and shrubs which constitute ‘vegetation’ as seen by the vegetation ecologist. They occupy habitats which are hostile to larger and deeper rooted plants.
The flora of the Albany district reflects a meeting point of various phytochoria (Cape, Tongoland/Pondoland, Karoo/Namib) and a great diversity of vegetation types can be found within a radius of 150km of Grahamstown (Lubke et al 1988). The vegetation type of Pluto’s Vale is clearly Valley Bushveld (Acocks veld type no. A23) but that of Helspoort comprises Valley Bushveld on the steep slopes of the poort with False Fynbos (Acocks veld type no. 70, Grassy Fynbos of Low & Rebelo 1996) on the flat summits and karoo influences (False Karroid Broken Veld, Acocks veld type no. 37) on flat, low lying areas.
The vegetation as described by Low & Rebelo (1996) for the Zuurberg is Grassy Fynbos (Acocks veld type No. 65). This is endemic to the Eastern Cape from the Kouga Mnts to Port Elizabeth, and on the Grootrivierberge from Steytlerville to Grahamstown and Bushmans River Mouth, mainly on the mountain tops. Grassy Fynbos replaces Mountain Fynbos in areas where the component of summer rainfall increases; the Restios are replaced by grasses and the Proteoid elements replaced by small-leaved shrubs and succulents (e.g. Aloe). Due to the increase in grass‑fuel, fires are more frequent in Grassy Fynbos than Mountain Fynbos.
Factors influencing Haworthia populations The western locality of Helspoort is on the margin of False Fynbos, which coincides with the well-drained, nutrient-poor, quartz‑derived soils, incoming coastal mists and cooler temperatures. The central locality is on shale that is richer in nutrient and has a higher moisture holding capacity. The vegetation is False Karoid Broken Veld that does not receive additional moisture from coastal mist and has higher temperatures. This locality is locally dominated by Pteronia incana, a pioneer shrub that indicates poor veld management and disturbance, particularly in this veld type. It is unlikely that this is a factor in the occurrence of the Haworthia in MBB7052. What is evident in nearly all Haworthia populations anywhere is that the habitats are disturbance resistant – either from fire or from physical animal trampling and grazing. In the absence of any management at all, however, rocky sites and ravine, cliff and valley margins, do become severely degraded with a negative impact on Haworthia on such sites.
The eastern locality at Helspoort is on Dwyka tillite that has a high moisture holding capacity. The vegetation is consequently Valley Bushveld and does not receive the coastal mists or cooling winds which results in drier, hotter conditions similar to the central lower locality. However, the vegetation is less disturbed and the plants are more protected by surrounding vegetation and protruding rocks.
The rainfall difference between Pluto’s Vale and Helspoort is insignificant. The plants at Pluto’s Vale, south of the road, grow in weakly developed, shallow, lime-rich soils overlying Dwyka tillite in dense Valley Bushveld. The plants are also on the southern aspects. North of the road and all the way to the eastern exit from the valley, are the steeper solid rack faces of Ripon sandstone. The only significant difference is probably that of parent material and scale of exposed rock. Where there is a single population of H. cymbiformis var. incurvula, it is on fragmenting blue Ecca shale. The very nearby population of the H. gracilis var. tenera intermediate is in an area with tumbled rock and a product of a disturbance event of some kind. The north-western cooperi-graciloid element is again on a Dwyka formation.
At Kaboega, it is clear that the cymbiformis and cooperi-graciloid elements are predominantly on Dwyka Tillite and on the southern slopes. The cooperi-piliferoid elements are in the higher lying margins between Fynbos and either Valley Bushveld or remnant forest. The aristata elements are evident in the dryer, either low-lying, or relatively high north-facing dry sites. However, it is quite evident that substrate is only contributory and the clinal nature of the variation between populations dramatically portrays the spatial arrangement. Within this spatial arrangement it is also evident that polytopic similarities occur. The cooperi-piliferoid element is here at its westernmost, but the cymbiformoid element is polytopically a distinct element at the very western-most population indicated on the map.
Similarly there are two widely separated, almost teneroid, populations of plants which are highly site specific on vertical ferruginous quartzite in one case, and white purer quartz in the other. At the western most of these two sites, two populations occur within 25 meters of each other. They flower and seed simultaneously and the most probable relationship is that of ecotype as deduced from the general continuity between all the Kaboega populations of this ilk. One population is the teneroid (or graciloid or isabelloid) element) and the other is piliferoid/aristatoid. The latter is in dense grass over boulder-like terrain and the other (Fig.9 – ‘puberula’ in the preceding chapter) is on several small outcrops of vertical shaly-quartzite – also the home of Euphorbia polygona. The other population consists of plants that are smaller, fairly proliferous and very spinose with tiny spines too.
A few very similar plants to the last mentioned, were found to the east where they seemed to grade locally westward to a cooperoid element, but eastward to a small population on whiter quartzite, where the plants are unusual in having surface spination as well. Usually such spination is on leaf margins and keels. This surface spination does occur in a single population of H. cooperi (i.e. var. venusta from near Alexandria) and also in a single (known) populations of H. gracilis var. isabellae from the Longkloof. However, sparse surface spination is fairly common in H. decipiens, in H. nortieri and to a small degree in H. arachnoidea.
The species H. cymbiformis, which is what the western-most of several populations clearly resembles, is only known again at Enon, east of Kirkwood and south of the Zuurberg. The particular site of this “look-alike” is a much bolder rockface than any of the other south-facing Dwyka slopes.
The populations on Kaboega (apart from H. glauca) are not widely distributed and populations can be highly site-specific. It should be noted that H. angustifolia var. baylissi, occurring a few miles east of Kaboega, does not appear in the study area. There is no indication of the speculated connection of this species to more western counterparts viz. H. zantneriana and H. monticola. H. glauca var. glauca (in a form suggesting intergradation with H. coarctata at the western-most of the populations observed) occurs very abundantly on the rocky ridges on the upper north slopes of the Zuurberg, and also on the west facing slopes of the ridge south of Klipfontein farm. H. sordida occurs (as known) at a single site under Elytropappus rhinocerotis (renosterbos), and H. nigra is also present at a single, similarly highly localised site on ancient boulder debris under largely Pentzia incana. H. viscosa is absent. Curiously, Astroloba foliolosa is also represented by one known population on what must be the remnant of ancient river terrace near to, but several meters above, the present course of the Kaboega river. There are many other such species singularites on Kaboega which could be described in terms of geology, aspect and altitude but information which would be extremely difficult to synthesis in any particular way. For example, Euphorbia tetragona is present in highly localised places and the plants are all very tall and old with no evidence of recruitment. Euphorbia stellaspina, E. polygona, E. silenifolia, Faucaria felina, Aloe microstigma, A. pluridens, A. speciosa, A. tenuior, A. ferox, A. striata, Piaranthus disparilis, Duvalia caespitosa, Tridentea (Stapelia) longii, Quaqua pillansii, Huernia brevirostris, H. guttata, and H. campanulata are all present and largely as site-specific populations independent of the Haworthia populations. Cyrtanthus angustifolius, Haemanthus humilis and Dioscorea elephantipes are also present and localised. So while these elements may all have specific habitat requirements, it will be an awesome task to explain all the minutae which may account for their disparate occurrences.
Conclusions Haworthia species are invariably found where there is exposed rock and where the vegetative cover is low and sparse. Rainfall, soil and vegetation are of general significance but we consider that it is geology which may play the predominant role in explaining variability and hence ecotypification. We explain this as follows:-
a. Plutos Vale – evidence is that Dwyka seems to support H. cymbiformis var. incurvula. On the more quartzitic Dwyka series it transforms to a more cooperoid-graciloid element, while to the Ripon sandstones it becomes teneroid.
b. Helspoort – on the heavier low-lying soils the graciloid element becomes more cooperi-piliferoid.
c. Kaboega – the cooperoid and graciloid elements are in the quartzitic formations, the cymbiformoid in the Dwyka tillite, and the aristatoid in the Ecca and Ripon shales.
The cooperi-graciloid elements of Pluto’s Vale and Kaboega are morphologically continuous with the cymbiformoid, and are indistinguishable from many of the graciloid, and cymbiformoid (H. cymbiformis var. transiens) elements of the Baviaanskloof.
We repeat what Bayer wrote in 1999, that the basis of classification of Haworthia must be the geographical component. We add that the tendency in both the professional and academic ranks to recognise “species” on the flimsiest of morphological pretences, and to rank nomenclature wholly disproportionately to the function of identification, will never help to resolve a problem which seems to exist uniquely in Haworthia, itself a complete misconception. Classification in Haworthia has simply been dumbed-down by a wide community of writers who seem to have no empathy with the literature of the subject. The classification should be of such a nature as to have some significance for others who may seek to know something about the driving forces and products of change relating to plants. The converse should also be true. In this case, it is apparent that the Haworthia species and their variability on Kaboega, must have some relation to the broad statement … “where four biomes are said to meet”.
References
Dent, M.C., Lynch, S.D. & Schulze, R.E. 1987. Mapping mean annual and other rainfall statistics over southern Africa. Agricultural Catchments Research Unit Report No. 27. Department of Agricultural Engineering, University of Natal, Pietermaritzburg.
Dyer, R.A. 1937. The vegetation of the divisions of Albany and Bathurst. Botanical Survey of South Africa Memoir 17. Government Printer, Pretoria.
Hartmann, M.O. 1988. In Lubke, R.A., Bruton, M.N. & Gess, F.W. (eds). Towards and Environmental Plan for the Eastern Cape. Rhodes University, Grahamstown.
Lubke, R.A., Tinley, K.L. & Cowling, R.M. 1988. In Lubke, R.A., Bruton, M.N. & Gess, F.W. (eds). Towards and Environmental Plan for the Eastern Cape. Rhodes University, Grahamstownn
Bayer, M.B. The Haworthias of Kaboega. In Haworthia Update. Umdaus in ms.
Bayer, M.B. The Case of Haworthia incurvula. Aloe 36:34, 199
Dolt, T & Hammer, S. (2001). Notes on the Faucaria from the Forest: Faucaria nemorosa L.Bolus ex L.E.Groen. Aloe, 38:37-38.
Low, A.B. & Rebelo, A.G. (eds.). (1996). Vegetation of South Africa, Lesotho and Swaziland. In M.N. Bruton & Gess (eds.) Towards an Environmental Plan for the eastern Cape. Rhodes Univ., Grahamstown.
This essay was published in Haworthiad 16:62, 2002.
Introduction
Subsequent to my revision Haworthia Revisited (1999), I have done much more fieldwork, particularly in the Eastern Cape. This has revealed even more striking evidence of the intense inter‑relatedness of the so‑called species in Haworthia. Classification and revisionary classification is a sampling process. As this progresses and more material is collected, so the classification firms up. A extensive discussion explaining the following combinations and two new varieties is provided in Haworthia Update Vol.1 and an insight into the taxonomic problems to be solved is provided by the illustrations with another article, “Small Hairy Things”.
My classification had some problem areas that were anticipated to a degree in Haworthia Revisited. The following sentence appears in the discussion of H. cymbiformis var. transiens: “Thus H. mucronata can be allied with equal facility to either H. cymbiformis or H. cooperi, when in fact in the field it is more intimately related to H. decipiens. The location of this note is a powerful reminder that distinctions between species are highly blurred and that alternative solutions are possible.” I also make repeated references to the nature of the relationship of species and variants. Many of those are specific to, or apply to, or are predictive of the following changes. The basis of the following combinations is thus laid in Haworthia Revisited.
Although collections are cited, these are not always now represented by herbarium specimens. The reason for this is simply one of resources: herbarium space, the impracticality of trying to represent all the variants in such herbarium state, and the effort required to manage living collections and their preservation as specimens. A photographic record is being maintained in lieu of dried specimens in herbaria, where the record extant is deemed to be otherwise inadequate.
The following new names and combinations are published below:
1. Re‑arrangement of Haworthia pringlei and H. xiphiophylla within H. bolusii and H. decipiens H. bolusii var. pringlei (C.L.Scott) M.B.Bayer comb. nov. H. decipiens var. xiphiophylla (Baker) M.B.Bayer comb. nov. H. decipiens var. virella M.B.Bayer var. nov.
2. Goodbye to Haworthia gracilis H. cooperi var. gracilis (v.Poelln.) M.B.Bayer comb. nov. H. cooperi var. isabellae (v.Poelln.) M.B.Bayer comb. nov. H. cooperi var. tenera (v.Poelln.) M.B.Bayer comb. nov. H. cooperi var. picturata (M.B.Bayer) M.B.Bayer comb. nov. H. cooperi var. viridis (M.B.Bayer) M.B.Bayer comb. nov. H. cooperi var. doldii M.B.Bayer var. nov.
3. A familiar new species, Haworthia transiens H. transiens (v.Poelln.) M.B.Bayer stat. nov.
4. Where does Haworthia helmiae belong? H. helmiae transferred to H. arachnoidea var. nigricans in synonymy .
1. Re‑arrangement of Haworthia pringlei and H. xiphiophyllawithinH.bolusii and H. decipiens
H. bolusii var. pringlei (C.L.Scott) M.B.Bayer comb. nov.
H. pringlei C.L.Scott, Bradleya 12: 103 (1994). H. decipiens var. pringlei (C.L.Scott) M.B.Bayer, Haworthia Revisited: 67 (1999) in respect of the type only. Type: CAPE‑3224 (Graaff Reinet): Adelaide district (‑DD), Scott in PRE8970 (holotype).
Collections:
3224 (Graaff Reinet): Adelaide district (‑DD), Scott in PRE8970.
All the other specimens and collections cited in Haworthia Revisited under H. decipiens var. pringlei are transferred to a new variety, H. decipiens var. virella.
My collection from east of Somerset East (Baviaanskranz) MBB6561 supported by the two Ripon collections above, and by several other collections pertaining to H. aristata, indicates that H. pringlei C.L. Scott is in fact better related to H. bolusii var. blackbeardiana. This is suggested by its interactions with H. cooperi var. dielsiana at Ripon, and complicated both by interaction with H. aristata south of that and extending to the new variety H. decipiens var. virella. This new combination is explained further in the discussion following var. virella.
H. decipiens var. xiphiophylla (Baker) M.B.Bayer comb. nov.
Haworthia xiphiophylla Baker, Fl.Cap. 6: 354 (1896). H. arachnoidea var. xiphiophylla (Baker) M.B.Bayer, Haworthia Revisited: 36 (1999). Type: Cape-3325 (Port Elizabeth); near Uitenhage, Howlett s.n. cult. Kew (holotype): icon (B). Epitype: CAPE‑3325 (Port Elizabeth): N. Coega Station (‑DA), Mrs E.B. King (NBG).
My earlier transfer of the element xiphiophylla (Figs.1, MBB6604 2a, b & c MBB6616) to H. arachnoidea in Haworthia Revisited was done with some reluctance and based on two main collections by Venter (Fig.3 JDV91/122 and JDV91/117, both vicinity of Mentz Dam). (Note: I do not regard an earlier transfer by J.J. Halda to H. arachniodea as valid. Halda put forward many nomenclatural changes which are so ridiculous that I do not accept their publication as serious botanical work – Halda’s folly is summarised in Haworthiad 14:35, 2000. How decisions are made can be questionable at the best of times and mine are made from a deep instinctive response arising from wide experience. Mistakes are excusable in view of the inherent difficulty of making decisions in Haworthia, but not excusable if made in total ignorance).
This change now follows my own fieldwork and particularly the collection MBB7028 (Figs 4a & b). from Darlington Dam (Lake Mentz. Other recent collections by principally Philip Desmet of University of Cape Town, and by me, show that xiphiophylla must point in the direction of H. decipiens as H. arachnoidea is present as a very distinctive dark green entity with minimum translucence, in the Steytlerville area. On the other hand there are many collections from the Groot Winterberg linking the Uitenhage/Coega xiphiophylla with H. decipiens variants in the Jansenville and Steytlerville areas. This also impacts on the question/problem of the relationship of H. gracilis var. viridis (transferred to H. cooperi in this paper) and H. cooperi to H. decipiens. To facilitate communication and discussion, I thought at first that it would be advisable to widen the application of the name xiphiophylla to incorporate a wide range of collections from the vicinity of Pearston, extending to north‑west of Jansenville and southwards to Willowmore and back to Kirkwood. These are decipiens‑like plants. Instead, I have reluctantly described these as the new variety H. decipiens var. virella, with the var. xiphiophylla transferred from H. arachnoidea to H. decipiens.
From var. decipiens, it differs with broader flatter leaves, incurved resembling H. arachnoidea and with inter‑veinal translucence. Includes populations transitional to H. decipiens var. minor M.B.Bayer and to H. cooperi var. viridis M.B.Bayer. (A var. decipiens differt foliis latioribus, planioribus, incurvis similis H. arachanoidea et interveniis translucidis. Includet populi transitionum ad H. decipiens var. minor M.B.Bayer et H. cooperi var. viridis M.B.Bayer.)
Collections:
3223 (Rietbron): S. Aberdeen (‑DC), Perry 659 (NBG).
3325 (Port Elizabeth): Lake Mentz (‑AA), Bayer 7028 (Figs.4a & b)
I have included Bayer & Bruyns 6580 from Meerlust, previously cited under H. decipiens var. decipiens and MBB6583 from Waaipoort, previously cited under H. decipiens var. minor. These changes may seem flippant and frivolous, but they should be seen as evidence of complex continuities that may not ever be resolved.
These plants have longer more attenuate leaves than H. decipiens, but with the brighter green of the typical var. xiphiophylla. The spination is generally coarser and firmer than H. bolusii var. blackbeardiana. These are the plants I had in mind when I decided to absorb Scott’s H. pringlei in H. decipiens. I wish to stress that doing so was not as mindless as may be suggested, because there is still a greater inherent problem in this new arrangement. There are populations, particularly south of Cradock, of H. bolusii var. blackbeardiana, which cannot be distinguished from populations of H. decipiens var. virella. Ironically, such latter populations near Pearston, grow with and discrete from H. bolusii var. bolusii (Figs.11 a & b). I also include illustrations of three collections from E. Steytlerville (Figs.13 JDV5-68, 14 JDV91-118 & 15 JDV93-40).
I do recognise that H. decipiens is a species that I do not know well enough; despite all the material I have seen. It perhaps occupies a geographical pivotal role in the interpretation of particularly H. cooperi, as I have now constituted that species. Pivotal in that it occupies the geographic centre stage between the south‑western Cape, the Karoo and the Eastern Cape; and is to some degree continuous with “species” in those areas. I have not seen any evidence to suggest that H. mucronata is actually directly involved with either H. cymbiformis or H. cooperi. There is no doubt that there is visual similarity, but I expect and regard the geographic association to be via H. decipiens.
2. Goodbye to Haworthia gracilis
It is difficult to reconcile recent field observations with my Haworthia Revisited concepts of H. cooperi and H. gracilis. However, there is a solution that can be offered for the classification problems, as discussed in depth in Haworthia Update (in press). If the concept of H. cooperi is enlarged to incorporate H. gracilis (a relatively recent von Poellnitz name), a more practical solution is presented.
This new concept of H. cooperi as a super‑species only excludes H. cymbiformis and H. bolusii var. blackbeardiana with some difficulty. These are important issues, but which I think are largely addressed by the changes made in this paper. It should be recognised implicitly that H. bolusii var. pringlei and H. decipiens var. virella, in terms of their history and present treatment, point directly at a connection between H. cooperi, H. bolusii and H. decipiens, as well as at wider associations.
H. cooperi var. gracilis (v.Poelln.) M.B.Bayer comb. nov.
H. gracilis v.Poelln., Feddes Repert.Spec.Nov. 27: 133 (1929). Idem 41: 201 (1937). Non C.L.Scott, The Genus Haworthia: 69 (1985). M.B.Bayer, Haworthia Revisited: 75 (1999). Type: Graaff‑Reinet, Amalienstein, Willowmore, Stellenbosch. Not preserved. Lectotype (Breuer, World of Haworthias 1:150 (1998): unpublished photographic icon :H. gracilis v.P. in (B). Epitype: CAPE‑3326 (Grahamstown): Hellspoort (‑BA), Britten (PRE).
H. cooperi var. isabellae (v.Poelln.) M.B.Bayer comb. nov.
H. isabellae v.Poelln., Feddes Repert.Spec.Nov. 44: 226 (1938). Non C.L.Scott: 76 (1985). H. gracilis var. isabellae (v.Poelln.) M.B.Bayer, Haworthia Revisited: 77 (1999). Type: Cape, near Port Elizabeth, Mrs I. King. Not preserved. Neotype: CAPE‑3325 (Port Elizabeth): Humansdorp, Gamtoos Bridge (‑CC), H. Hall in NBG 68799.
H. cooperi var. tenera (v.Poelln.) M.B.Bayer comb. nov.
H. tenera v.Poelln., Feddes Repert.Spec.Nov. 31: 86 (1933). C.L.Scott: 76 (1985). H. gracilis var. tenera (v.Poelln.) M.B.Bayer, Haworthia Revisited: 77 (1999). Type: Cape, Pluto’s Vale, Grahamstown, Miss Blackbeard 15. Not preserved. Neotype (Breuer & Metzing, Taxon 46(1):3 (1997): CAPE‑3326 (Grahamstown): Glenelg (BA), G.G.Smith 5416 (NBG).
H. cooperi var. picturata (M.B.Bayer) M.B.Bayer comb. nov.
H. gracilis var. picturata M.B.Bayer, Haworthia Revisited: 78 (1999). Type: CAPE‑3325 (Port Elizabeth): Enon (‑BC), Thode 21507 (NBG, Holo.).
H. cooperi var. viridis (M.B.Bayer) M.B.Bayer comb. nov.
H. gracilis var. viridis M.B.Bayer, Haworthia Revisited: 79 (1999). Type: CAPE‑3325 (Port Elizabeth): Perdepoort (‑AC), G.G.Smith 6867 (NBG, holotype).
Rosettes small to 30mm diameter with 25‑30 slender, attenuate leaves to 50mm long, erect spreading, dark purplish green and with firm white marginal spines 2‑3mm long. (Rosulae parvae 30mm diametro, foliis 25‑30 gracilibus, attenuatis, 50mm longis, erecto‑expansis, atropurpureo‑virdibus et spinis marginalibus firmis albis 2‑3mm longis.)
Collections: Only known from type locality.
This new, small form of H. cooperi is in the geographical orbit of the var. leightonii at Kaiser’s Beach and the more graciloid forms of the latter at Payne’s Hill, also nearby. It is named for Tony Dold of the Schonland Herbarium, Rhodes University, Grahamstown, who first collected it. I am recognising it as a distinct variety because of its geographic separation from other isabellae‑like plants and because it also has coarser spination and a darker purplish‑colour than the other small varieties of H. cooperi.
3. A familiar new species, Haworthia transiens
H. transiens (v.Poelln.) M.B.Bayer stat. nov.
H. planifolia var. transiens v.Poelln., Feddes Repert.Spec.Nov. 45: 163 (1938). H. cymbiformis var. transiens (v.Poelln.) M.B.Bayer, Haworthia Handbook: 162 (1976). M.B.Bayer, New Haworthia Handbook: 36 (1982). Type: Cape, Prince Alfred’s Pass, Archibald 327. Not preserved. Lectotype (Breuer & Metzing, Taxon 46(1):3 (1997): icon 5479 “H. planifolia var. transiens v.P.Typ.” (B). H. cymbiformis var. translucens Triebner et v.Poelln. idem 45:166 (1938). C.L.Scott: 94 (1985). Type: Cape, Prince Alfred’s Pass, Lategan in Triebner 1137. Not preserved. Neotype: CAPE‑3323 (Willowmore): Prince Alfred’s Pass, G.G.Smith 5709 (NBG).
Additional collections: (private photographic record, and living plants, not necessarily herbarium specimens):
The collection 3324 (Steytlerville): N Komdomo (‑DA), MBB 6789 includes plants which are this element, and the population intergrades to H. cooperi var. picturata and H. cooperi var. gordoniana. Similarly 3324 (Steytlerville): Grootriverpoort (‑DA), EvJ15927 is approximately transitional to H. cooperi var. isabellae. This intergradation is not only according to physical similarity but also geographic and ecotypic.
In raising this element to specific status, I am now suggesting that specimens cited in Haworthia Revisited as H. cymbiformis vars. brevifolia Triebner et v.Poelln. and multifolia Triebner from Uitenhage (Hellsgate) do indeed belong in H. cymbiformis.
All the variants of H. transiens that I am aware of, link this species with variants of H. cooperi as now constituted. That is where the continuity is. Thus I am recognising the fact that H. transiens is more directly connected to a broad concept of H. cooperi and less to any such one for H. cymbiformis. H. transiens is most closely continuous with H. cooperi var. picturata, which can be shown to be also continuous with the vars. gordoniana and isabellae. This is said within the context of my report on the variation of Haworthia at Kaboega in Haworthia Update (in press), where H. cooperi and H. cymbiformis are also said to be continuous. The difference from the case of those ex‑gracilis varieties, is in the distribution and number of populations in the Baviaanskloof which can be reasonably identified as H. transiens.
4. Where does Haworthia helmiae belong?
In M.B. Bayer, Haworthia Revisited: 117 (1999), both H. helmiae and H. integra were cited under H. mucronata, although the word ‘integra’ was erroneously omitted, and thus also omitted from the index. In a manuscript with Aloe (in press), I correct my citation of H. mucronata and let H. integra revert to the status of excluded names, because it was so poorly known and confused for so many years. I did however, leave the citation of H. helmiae under H. mucronata, which is a mistake ‑ especially as I cite the specimen and discuss the population as H. arachnoidea var. nigricans. My interpretation is still based on Scott’s statement that Mrs. Helm had collected this at a specific site in Schoemanspoort, near Oudtshoorn, where I subsequently collected. Had it not been for Scott, H. helmiae would also have been assigned to the ranks of the excluded and insufficiently known. The correct citation for H. helmiae v.Poelln. sensu Bayer is:
H. arachnoidea var. nigricans (Haw.) M.B.Bayer
Syn. Haworthia helmiae v.Poelln., Feddes Repert. Spec. Nov. 41: 201 (1937). C.L.Scott: 99 (1985). Type: Cape, Heidelberg, Smithers in Triebner 891, Great Brak River, Mrs Helm in Triebner 901. Not preserved. Lectotype: icon “H. helmiae v.P.”, Great Brak, Triebner 898 (B). Epitype: CAPE‑3322 (Oudtshoorn): Schoemanspoort (‑AC), M.B.Bayer 171 (NBG).
I have to point out that formal nomenclature is not my forte and that I have little enthusiasm for it. One of the reasons is that, in my opinion, it spawns authors and spurious authority recklessly and needlessly. Breuer, World of Haworthias 2: xiii (2000), states that I do not demonstrate ambiguity when I have cited epitypes where lectotypes had been selected. The fact is that in Haworthia just about any type is self‑evidently ambiguous and it seems hardly worth repeating the entire literature to prove this obvious point.
In the case of H. helmiae, von Poellnitz cites four Triebner numbers for four different collections. These were cited as from Heidelberg, Great Brak, George and Brandwacht (Brandwacht could also be said to be Great Brak ‑ Mrs. Helm lived actually nearer Little Brak at Brandwacht). I said in Haworthia Handbook: 60 (1982), that there is one photograph for Triebner 898 (Great Brak!) that could be taken to agree with plants from Schoemanspoort. Breuer (Idem: 512) repeated this as lectotypification. The fact is that it could be taken for anything. Breuer (Idem: 511, Haworthiad 15(3): 84 (2001) and private communication) now suggests (apparently on the basis of a photograph of Fourcade 5407 and one by Brown in Cact. Succ. J. (US) 18: 39 (1946)) that my H. outeniquensis is in fact this species. This is not convincing evidence in the paradigm of the Haworthia literature, nor in the face of the Fourcade photographs. My contention is that it is misleading to attempt to rewrite literature and interpret types without an adequate understanding of biological diversity and the problems of variation and species delimitation.
Where next for Haworthia description?
It must again be stressed that these new combinations and varieties will facilitate discussion, but will not eliminate the intrinsic problem of continuity. Although H. cooperi here seems to become a huge unwieldy entity, this better expresses the field situation, while still falling short. The recognition of species often requires the extension of a concept, which is the nomenclatural one derived from a single plant or from limited sampling. This may or may not be really representative of anything. It is very destructive and confusing if this nomenclatural aspect overrides the functional way in which names come to be used and are meant to be used.
There are real problems in attempting to classify Haworthia. In Haworthia Revisited, I stated that I tried “to practically identify nodes in a complex interlinked web”. The definition of “node” in physics is “a point of minimum displacement in a standing wave” ‑ and this is the meaning I transfer to Haworthia. The species should be seen as a series of standing waves superimposed over one another and they generally do not separate. My names are intended to be used to identify the principal groups of plants and populations that may be meaningful for identification. These names also relate those plants to geographic factors and to other plant species and vegetation.
The element H. transiens has always been problematic for me, as it does merge into H. cooperi var. picturata. There is a similar mergence of H. cooperi var. gracilis to H. cymbiformis var. incurvula at Pluto’s Vale, but which is localised. At Kaboega, the mergence of H. cooperi with H. cymbiformis is similarly complex, but it is here where there is real benefit from incorporating H. gracilis into the former. Thus the brighter green forms of the continuum are thus equated with H. cooperi var. viridis, the smaller spined forms with H. cooperi var. isabellae, and others either with H. cooperi var. gordoniana or H. cooperi var. cooperi. The difficulty is at the departure from the tight geographical boundaries of those variants, as well as from the implied or actual concomitant genetic variation. It is impossible to find consistent degrees of difference. There is a difficulty in explaining the relationship of all the Baviaanskloof elements and particularly the way H. cooperi var. gordoniana relates to H. decipiens from the Zuurberg in the east, to Uniondale in the west.
It is also not possible to find a classification solution that satisfactorily explains the extension of the relationships of H. cooperi and H. cymbiformis to H. decipiens and H. aristata. To do so would mean incorporating them within either species, and this would only lead to still further combinations and problems. I must personally resist any further proposals to combine the species that I regard as discrete. In the opposite direction, adding names for all the existing variants of already recognised elements will result in an unmanageable plethora, with the prospect of new names for further variants still to be found and this also cannot be supported.
In such a horticulturally popular group as Haworthia, there is an unfortunate pressure for classification which may have nothing to do with botanical realities. Often this has been done early in a learning process, rather than as a consequence of accumulated knowledge. Thus different opinions may be expressed with insufficient experience and sampled material. I think that it will be foolish in the extreme for anyone to make pronouncements or attempt to alter what has now been done without extensive additional field data and a greater understanding than my own.
I hope this discussion will drive other authors towards a more considered attitude to classification and the application of names.
Acknowledgements Many people have contributed in one way or another to the above. Among them, I would like to express appreciation to J.D. Venter for ongoing support; to Mr. and Mrs. G. Hobson of Ebenezer for their kindness and hospitality; P.V. Bruyns and Gerhard Marx for locations; to Tony Dold for the Chalumna find and for other assistance and support. Also, to Alan Butler, Gretchen Loucka and Derek Tribble for assistance with this text. ♦
Fig. 01. MBB6604 decipiens var xiphiophylla. CoegaFig. 02a. MBB6616 H. decipiens var xiphiophylla. Bauerskraal, UItenhage.Fig. 02b. MBB6616 H. decipiens var xiphiophylla. Bauerskraal, Uitenhage.Fig. 02c. MBB6616 H. decipiens var xiphiophylla. Bauerskraal, Uitenhage.Fig. 03. JDV91-122 H. decipiens var virella. S Darlington Dam.Fig. 04a. MBB7028 H. decipiens var virella. S Darlington Dam.Fig. 04b. MBB7028 H. decipiens var virella. S Darlington Dam.Fig. 05. JDV87-81 H. decipiens var virella. Ebenezer, Pearston.Fig. 06a. MBB7023 H. decipiens var virella. Ebenezer, PearstonFig. 06b. MBB7023 H. decipiens var virella. Ebenezer, Pearston.Fig. 07. MBB6581 H. decipiens var virella. Welgelegen, NW Jansenville.Fig. 08a. MBB7043 H. decipiens var virella. Llangollen, NE Jansenville.Fig. 08b. MB7043 H. decipiens var virella. Llangollen, NE Jansenville.Fig. 09. MBB7047 H. decipiens var virella. DeRust, E Jansenville.Fig. 10. MBB7041 H. decipiens var virella. Palmietfontein, Jansenville.Fig. 11a. MBB7022 H. decipiens var virella. SE Pearston.Fig. 11b. MBB7022 H. decipiens var virella. SE Pearston.Fig. 12. MBB7582 H. decipiens var virella. SE Mt Steward.Fig. 13. JDV85-68 H. decipiens var virella. E Steytlerville.Fig. 14. JDV91-118 H. decipiens var virella. Sandpoort, Steytlerville.Fig. 15. JDV93-40 H. decipiens var virella. Baroe, E Steytlerville.Fig. 16a. T.Dold3961 H. cooperi var doldii. Chalumna.Fig. 16b. T.Dold3961 H. cooperi var doldii. Chalumna.
This article was published in Haworthiad 16:43, 2002. Since then I have implemented name changes and I indicate these in bold type.
When I have written about Haworthia, I have generally taken as a subject a particular species, in the sense that people regard a species as a kind of thing universally and unmistakably recognisable. It is not always easy to find such things in the lower life forms, and this is also true for the sub‑genus Haworthia. Here I am just writing about a few odd plants, without going into the many ramifications that are actually involved.
I am also using the classification, and system, rationalised and explained as best I could in my book “Haworthia Revisited” (1999). Since that was written, I have been on many more exploratory journeys and have learnt a lot more. Much of this new information has been published in Haworthia Update Vol.1. There are several essays there, one devoted to the Baviaanskloof and one to the northern Zuurberg (Kaboega). I explain that the name H. gracilis is probably redundant. (I limit its use to H. cooperi var. gracilis as it occurs at Helspoort, Grahamstown.) It may actually be better to regard most of the Baviaanskloof populations all as one species ‑ variants of a greater species that will be H. cooperi. I will implement the necessary name changes in another paper (This was done in, and the article is copied, in a preceding essay).
Thus one of the dominant variants will be the old H. gracilis var. isabellae as H. cooperi var. isabellae, along with H. cooperi var. picturata, plus H. cymbiformis var. transiens as H. transiens. I will also use here the name H. decipiens var. pringlei which will transfer to H. bolusii. There are populations that can then be identified as:
* var. gracilis ‑ plants with erect incurving simple leaves with few marginal spines * var. picturata ‑ plants with relatively opaque, green incurving leaves in a compact rosette * var. gordoniana ‑ plants with blue‑green stubby leaves also relatively opaque * var. isabellae ‑ plants with slender erect leaves which are fairly densely spinose, usually only on the margins * transiens ‑ plants with more stubby, relatively translucent and reticulate leaves
The difficulty with the existing system is that the continuities which we see in the Baviaanskloof and Longkloof, are also evident in surrounding areas. For example, at Uniondale there is difficulty in excluding H. mucronata from the discussion, although there is no evidence to convince me that there is geographical continuity between these Uniondale plants and H. mucronata west of Calitzdorp. The problem is more in respect of H. decipiens and the variation of this species north and north‑east of Uniondale, then westwards north of the Klein Swartbergs (to meet both H. lockwoodii and H. mucronata from Prince Albert westward), as well as south of the Swartberg (with H. mucronata) north‑east of Calitzdorp. Similarly the Baviaanskloof populations have affinities across the Springbokvlakte area east of Steytlerville, meeting with cooperoid elements from north‑east of Uniondale, all the way to the Zuurberg around the greater Kirkwood area. These cooperoid elements have continuities with the H. decipiens complex of that area.
I write this article to show that neither “lumping” nor “splitting” offers any solution to how we talk about, explain and communicate about these plants. Opinions have been expressed that certain Haworthia varieties and species should be “lumped”. We want to talk about distinctive individual recognisable things, but this is simply not possible, however desirable it may be. When I say a plant name, I have to make it clear that I am usually talking in general terms of many individuals, which collectively have created an image in my mind, and this is not the way the nomenclatural code works.
If one attaches varietal or species names to every local variant, it means that so very many names will be needed. It is a difficult decision ‑ many names which are independent, untrackable and meaningless, or fewer names which are connected, interdependent and usable. The variants I mention comprise a range of networked forms that are different from one another to a greater or lesser degree. This then means there is no formula, diagnostic or descriptive key or method by which these can all be documented and classified and identified with any degree of certainty. There never will be.
Here I am taking some of these small hairy plants and showing how similar they can be and how they may link up to each other.
Commentary ‑ on Figures 1a & b: H. cooperi var. gordoniana.
Fig. 1a. MBB6553 H. cooperi var gordoniana. N Zuurbron.Fig. 1b. MBB6553 H. cooperi var gordoniana. N Zuurbron.
These are small plants which have the characteristic blue‑green colouration of H. cooperi but the spines are smaller, more numerous and more closely spaced. It is very distinctive where it occurs in the Hankey Pass near Zuurbron. There is intergradation in the Baviaanskloof from var. gordoniana, to H. gracilis var. isabellae, to var. picturata and on to H. cymbiformis var. transiens. It does not end there. I have suggested that a solution, which would also solve problems in the greater H. cooperi arena, would be to absorb H. gracilis into H. cooperi, and perhaps H. bolusii var. blackbeardiana as well. However, it does not end there either, and we would logically have to extend this to include H. cymbiformis and H. decipiens. This also does not exhaust the possibilities, exacerbated by the fact that field exploration is by no means complete.
on Figures 2 to 9: H. gracilis (cooperi) var. isabellae.
Fig. 2. JDV97-58 H. cooperi var gordoniana. N Zuurbron.Fig. 3. MBB6773 H. cooperi var isabellae. N Joubertina.Fig. 4. PVB7128 H. cooperi var isabellae. Holriver, S Hankey.Fig. 5. EvJ14080 H. cooperi var isabellae. Vetmaakvlakte, Baviaanskloof.Fig. 6a. MBB6826 H. cooperi var isabellae. Baviaanskloof.Fig. 7. H. cooperi var isabellae. Philips Tunnel, Hankey.Fig. 8a. MBB7021 H. bolusii var bolusii. SE Pearston.Fig. 8b. MBB7021 H. bolusii var bolusii. SE Pearston.Fig. 9. MBB6583 H. bolusii var bolusii. Lootskloof, E Jansenville.
There are a number of populations that are fairly close in appearance, usually becoming paler in colour and with more translucence and longer and coarser spines. I also include a number of populations in this category that have relatively few leaves, which are often without spines and are too robust to be regarded as H. gracilis (hence driving change to the name cooperi). Curiously, what happened was this. I produced and grew some seed of a collection by Ernst van Jaarsveld from north of Joubertina. Ernst’s find is H. graciliscooperi) var. isabellae by virtue of the many slender leaves and dense spination. It is also special in that the leaves have surface spination. Then I lost the seedlings and was puzzled by a batch labelled JDV97/158, which looked like H. cooperi var. gordoniana (fig.2 –note JDV97-58) without surface spination. That was until I discovered that JDV97/158 is Kobus Venter’s accession number for my MBB6773 (fig.3), which is also Ernst’s find but collected by myself. At the time of writing, I am still not sure that these seedlings really are MBB6773, because at this stage they have fewer, shorter, less‑spined leaves than their parents. Perhaps there is a mistake, although I have grown several other collections in the same way. While disconcerted by the difference of the seedlings from the parents, it has never been to this degree. (There was in fact a mistake – a transposition of digits – the seedlings were JDV97-58)
MBB6773 has a few look‑alike collections that are very similar e.g. PVB7128 (fig.4) from south‑west of Hankey. This is a very blue‑green finely spined version of H. gracilis (cooperi) var. isabellae. There is also EVJ14080 (fig.5) from Vetmaakvlakte along the Couga River, where it turns south from the Baviaanskloof to the Longkloof. Then there is MBB6826 (figs.6a & 6b) from Rooikloof, further west along the Baviaanskloof, or MBB6802 from south‑west of Hankey at Philip’s Tunnel (fig.7). Triangulated between these three, are populations which are either better placed with H. cooperi var. gordoniana, or with H. gracilis (cooperi) var. picturata, or with H. cymbiformis var. transiens (transiens).
In the case of MBB6826, I have illustrated two plants; one is half the size of the other, to show that there is a great deal of variation within populations too. Often many plants in cultivation are vegetatively propagated clones which obfuscate the degree of variability. Using one plant does not help very much, because all too often one finds that, although the overall appearances of populations may be different, there are individuals common to both which are very similar. In the field, it also happens that the plants may look different, whereas in cultivation these differences are lost.
on Figures 8 to 10: H. bolusii var. bolusii and H. decipiens var. virella.
Fig. 8a. MBB7021 H. bolusii var bolusii. SE Pearston.Fig. 8b. H. bolusii var bolusii. SE Pearston.Fig. 9. MBB6583 H. bolusii var bolusii. Lootskloof, E Jansenville.Fig. 10. MBB6583 H. decipiens var virella. Waaipoort, NE Steytlerville.
If one takes, say, the Philip’s Tunnel illustration (fig. 7), one can compare it very favourably to MBB7021 (fig.8a & b) from far away Pearston, which is a completely different species, viz. H. bolusii var. bolusii. The reservation is of course that the above‑mentioned chain of continuity extends geographically through the Eastern Cape from Hankey across several mountain chains into the interior. I have again given two illustrations to show that even in relatively homogenous populations, one can still find distinctive variants. MBB7046 (fig.9) is a smaller version of var. bolusii from east of Jansenville (Note: Breuer has described this as a new species viz. H. odetteae, done in ignorance of the collection MBB7021 and also the fact that similar plants occur at Hinchinbrook close to the northeast. These with other records, strongly suggest continuity with bolusii var. bolusii at Graaff Reinet itself.) There it grows with another element, which I have included in my extended version of H. decipiens. This is actually a variety occurring in many populations, which I need to explain links H. cooperi, H. decipiens and H. aristata. While occurring in a distinctive form and populations together with H. bolusii var. bolusii (three such localities have been seen by me), it also has an extension MBB6583 (fig.10) from north‑east of Steytlerville, which is a look‑alike except for a deeper green colour. This population is presently classified with H. decipiens var. pringlei (decipiens var. virella), which I am planning to re‑structure.
on Figures 11 to 14: H. aristata and H. gracilis (cooperi) var. isabellae.
Fig. 11. MBB6902 H. aristata. Hopewell, N Zuurberg.Fig. 12. MBB6917 H. aristata. Kaboega, Zuurberg.Fig. 13. MBB6905 H. cooperi ‘puberula’. Wilgerfontein, Kaboega.Fig. 14. MBB7017 H. cooperi ‘puberula’. Klipfontein, Kaboega.
One can then again take the Philip’s Tunnel illustration (fig. 7), always remembering that it is one of a variable population, and compare it with MBB6901 from Hopewell (fig.11) or MBB6917 from Kaboega (fig.12), both from north of the Zuurberg. These are both variants of the geographic complex that constitute H. aristata. That complex has extensions such as MBB6905 (fig.13) from Wilgerfontein and MBB7017 (fig.14) from Klipfontein, which are both from on the northern edge of the Zuurberg (I discuss these in preceding chapters and refer to them as H. cooperi ‘puberula’). The former is remarkable for its fine spination and the fact that twenty‑five metres away, it has an ecotype, which has the fewer stumpier leaves of what could be taken for more typical H. cooperi var. gordoniana. The latter (i.e. Fig.14) is remarkable because its small size likens it to H. gracilis var. isabellae; firstly to MBB6826 (fig.6a & b) and secondly to MBB6773 (fig.3), because it also has unusual surface spination. The plants illustrated in figs.13 &d 14 are several kilometres apart and occur in two very small, localised populations. They are related to each other through many other populations that in many respects suggest a cooperoid or cymbifomoid archetype. (see Chapter 7 Continuity…) ♦
If any two species present a particular problem for the taxonomist in terms of their perceived variation, it must be these two. I wrote a short article discussing this in respect of one particular population of Haworthia arachnoidea (Aloe 38:76, 2001). I also wrote on the same problem in my book Haworthia Update Vol.1 (Umdaus Press, 2002). Despite some of the things I said there, there is still some misunderstanding expressed in informal communication regarding my approach, not only to the recognition of varieties, but to the interplay between what have been perceived to be different species. One of the complaints is that if I do not recognise some of the new varieties described by other authors, my own varietal taxa should not be taken seriously. In my Aloe article I wrote… “I have used varieties simply as a communication and descriptive tool to suggest lesser nodes and connections between what I think may be species.” In my revision (Haworthia Revisited, 1999, Umdaus Press), I wrote…”Lesser ranks should not be taken too seriously.”
In about 1972 I was exploring west of Ladismith (Winkelplaas, MBB2716) when I for the first time came upon a problematic population which was neither H. arachnoidea nor H. mucronata. Subsequently the problem this presents for classification has grown with virtually every outing I have made into the Little Karoo. I now have a vast amount of material which is, however, still inadequate in terms of geographic coverage to form the basis of a fully comprehensive report. This short note simply arises out of a curious report kindly sent to me by Ingo Breuer who stated that H. arachnoidea grew together with H. mucronata at Grootrivier, west of Ladismith. I was very dubious about this statement because his locality is at Buffelsdrif which is virtually at east of Winkelplaas.
To explain that I first would like to briefly explain and illustrate my interpretation of what H. mucronata is. The name (note just the name) that designates the species is typified by an illustration in the Kew herbarium (Fig. 2) to which no exact origin is attached. In my earlier works I regarded the name as insufficiently typified and chose rather to use von Poellnitz’ name H. unicolor based on plants from south of Barrydale (see Figs.3a & b MBB7216). Scott preferred to uphold these Barrydale plants as the species H. mclarenii and use the name H. mucronata for a Dekenah collection from Calitzdorp. To accommodate Col. Scott I unwisely used his application of the name mucronata to the system of populations that I considered to be the same one species for which I applied the name unicolor. I should have stayed with the name H. unicolor based on a geographically designated collection and left the name H. mucronata as of doubtful origin and hence uncertain (see Endnote). This is now water under the bridge. I use the name H. mucronata for a system of populations which extends from west of Montagu to east of Oudtshoorn (see also endnote). I should designate an epitype which would be … Cape(3320DC): Barrydale, J. Dekenah 234 (NBG) = MBB7216, to make it quite clear from which field population it is derived and to which I attach the name. But this is also not so simply resolved. My plants in Figs.3a & b (MBB7216) are of large glabrous plants on a warm and dry north slope at Barrydale. There are smaller plants on the cool, wetter south slope only about 100 meters away (Fig.4 MBB7213). Fig.5 (MBB7212) shows two larger, spined plants from Tradouwshoek, a locality about 1.5km north-east of Barrydale on a moderate south slope. In the Tradouw pass a few kilometers away there are plants which could just as well be H. arachnoidea or extensions of these variable plants of H. mucronata from Barrydale itself.
Fig. 02. H. mucronata. Kew herbarium.fig. 03a. MBB7216 H. mucronata var mucronata. S Barrydale.Fig. 03b. MBB7216 H. mucronata var mucronata. S Barrydale.Fig. 04. MBB7213 H. mucronata var mucronata. S Barrydale.Fig. 05. MBB722 H. mucronata var mucronata. Tradouwshoek, NE Barrydale.
Having said this, I must point out that this article does not seek to explain or illustrate the full extent of the variability of either species. H. arachnoidea is illustrated in Fig.6 (MBB7207). This is of a plant from the site at Buffelsdrif [2] presumed to be that indicated to me by I. Breuer. Had I seen this plant alone I would have said “Yes, this is clearly H. arachnoidea.” I would have been embarrassed to have to add “var. setata” which I use to denote the Little Karoo variants of this species, because there is really no good basis for such a varietal distinction and I never fully believed that there was. The problem is, that as Breuer suggested, there is something else present. This is a smaller fairly glabrous plant with some marginal translucence to the leaves (Fig.7, also MBB7207). Does this then mean that H. arachnoidea and H. mucronata are there growing sympatrically (co-occurring, growing together in one habitat)? Unfortunately the spot we explored was rather confined and time did not permit further exploration. I am not certain this is in fact the exact site referred to by Breuer, but there were no other options in terms of his referral. What does therefore motivate me to write is the information peripheral to the report and what we saw elsewhere on this particular excursion. My old Winkelplaas collection tells me that we have “hybridisation” of the two species. But other collections suggest to me that this “hybridisation” is so extensive throughout the Little Karoo that it cannot be dismissed so simply.
Fig. 06. MBB7207 H. arachnoidea. Buffelsdrif.Fig. 07. MBB7207 H. arachnoidea=mucronata. Buffelsdrif.
In Haworthia Update Vol.1, I indicated how intensively one has to explore to properly account for field variability. The Buffelsdrif/Winkelplaas area has many potential habitats that would have to be searched to fully explain what is there. Just on this one outing we stopped at several places. At Vensterkrans a short distance to the south-west, we explored four hillsides. Figs.8a & b(MBB7205) are of spinose and glabrous plants from west of Vensterkrans [3]. They are paler in colour than H. arachnoidea and they exhibit a degree of translucence. These are actually the two characters by which I would suggest the two species could be generally distinguished i.e. arachnoidea – very dark-green and with no translucence to the leaves, mucronata – paler green and with translucence especially to the leaf margins. The two illustrations by no means indicate the degree of variation at this one site. Fig.9(MBB7206) is of a plant from between Vensterkrans and the first noted Buffelsdrif locality [4]. It is mid-green, spination is reduced, there is no obvious translucence and the plant has very firm margins and keels to the leaves. This gives slightly greater rigidity in the rosette and is a common feature in plants from many populations in the greater area. Also the plant is but one from a highly varied assemblage – none of which resemble either H. mucronata or H. arachnoidea. It is for populations of this ilk that I adopted the varietal name H. arachnoidea var. nigricans, explaining that it was only a device to accommodate plants (largely as populations) that were neither H. arachnoidea nor H. mucronata, but something in-between. One could equally have applied the varietal epithet for these plants under H. mucronata. As proposed with H. cooperi/gracilis, this is once again a case where a series of intermediate populations (in this case called H. arachonidea var. nigricans) blur the distinction between the ‘core’ species (largely because of the vagaries of nomenclature and application of types), H. arachnoidea and H. mucronata, thus invalidating any neatly definable species boundaries.
Fig. 08a. MBB7205 H. arachnoidea var nigricans. Vensterkranz.Fig. 08b. MBB7205 H. arachnoidea var nigricans. Vensterkranz.Fig. 09. MBB7206 H. arachnoidea var nigricans. W Vensterkranz.
Figs.10a & b (MBB7210) are of two smaller plants from east of Vensterkrans [5]. Plants from map site [6] are comparable with sites [3], [4] and [5]. These are similar to the preceding plants illustrated as H. arachnoidea var. nigricans, but tend to be more compact, still less spinose, and on some plants there is no marginal translucence. When I first saw plants of this ilk from about 8 kilometres to the east, I speculated that maybe this is where H. marumiana had originated as judged from an old Luckhoff collection from Ladismith designated by Von Poellnitz as that species. The plants from east of Vensterkrans do in fact have some superficial resemblance to H. marumiana var. batesiana. Stressed young plants do suggest the typical variety of that species and I suggest that this accounts for Von Poellnitz’s erroneous reference (note the plant in Fig.7).
Fig. 10a MBB7210 H. arachnoidea var nigricans. E Vensterkranz.Fig. 10b. MBB7210 H. arachnoidea var nigricans. E Vensterkranz.
I conclude this note with Figs.11a & b (MBB7211) of plants from Springfontein some distance to the southeast and nearer to the Langeberg. The illustrations are of a spinose and a glabrous plant, neither of which have marginal translucence. I would identify them as H. arachnoidea var. nigricans, and say that this is what I used to denote by the name H. unicolor var. venteri in my 1982 work, The New Haworthia Handbook, Kirstenbosch. I then wrote … “It is not in the least clear what the relationship is between H. unicolor (now H. mucronata) and H. arachnoidea. I make this point again and again to explain that it is pointless trying to name and describe an endless array of taxonomic varieties in the mistaken belief that a better grasp has been obtained of this incredible interplay of natural systems. I think it is fairly useless to suggest that some unfortunate taxonomist in the future will be saddled with the task of deciding the validity of such efforts, when the evidence here in the present is already indeterminate.
Fig. 11a. MBB7211 H. arachnoidea var nigricans. Springfontein.Fig.-11b. MBB7211 H. arachnoidea var. nigricans. Springfontein.
Endnote: It is perhaps useful to give Scott’s interpretation relating to the above:
1. He had H. mcclarenii in a section Loratae as a name specifically for Barrydale plants.
2. He he had H. mucronata in the section Denticulatae as a name for an array of specimens scattered across the landscape. Included in a wide array of “selected (herbarium) specimens are plants from:
Middelburg, Sterkstroom and Cradock, that are H. bolusii var. blackbeardiana), Adelaide, Grahamstown, Uitenhage and Humansdorp, that are H. cooperi var. gracilis.
Napier, that is H. rossouwii.
Ladismith, Oudtshoorn, that are H. mucronata var. inconfluens.
3. He had H. aristata in a section Planifoliae on its own, also for plants from Barrydale and comingling with his distributions for H. mucronata.
4. He recognized two species viz. H. helmiae and H. integra from single localities in the Little Karoo, in a section Muticae.
All these five elements of Scott’s, in terms of names and origins, fall into the system/s to which I apply the names arachnoidea var. nigricans or mucronata. Nothing suggests to me that other writers have a better interpretation of very difficult problem in determining the degree of interfacing of this system with that of arachoidea, lockwoodii, decipiens and cooperi (the latter as discussed in Haworthia Update Vol.1).
Acknowledgement I would like to record the credibility afforded to me by Western Cape Nature Conservation Board in extending permit approval for the limited collecting this kind of investigation requires. Mr. Johan Meyer of Barrydale was generous enough as to accommodate my wife and I, arrange landowner contact, add his own enthusiasm and interest, and accompany us on the excursion. I particularly would like to acknowledge the improvements and alterations to the manuscript suggested by Paul Forster of Queensland, Australia. It is clear that writing in the sphere of Haworthia now requires critical and informed comment before publication, and I appreciate this from Paul. ♦
This appeared as an article in ALOE 38:31 (2001). Unfortunately there was a problem with illustrations and captions and these are corrected here. A comment is also added as an addendum to respond to criticism by I. Breuer published in Alsterworthia 1:13(2002).
Introduction:
I described Haworthia serrata in 1973 (Jl S.AFr.Bot.39:249, see Figs.1) from Oudekraal, southwest of Heidelberg. I commented then on the wisdom of describing a new species when “the recognition, estimation of taxonomic rank and circumscription of elements in Haworthia…” was so problematic. The new species was said to resemble H. emelyae var. multifolia (Figs.2). In respect of its distribution, I said it was closest to H. heidelbergensis at Heidelberg (Figs.3 JDV87/1) and as at Matjestoon (Fig.4 JDV87/3), and also to H. sublimpidula at Swellendam (now known to be H. floribunda var. major (Fig.5 MBB6859, taxonomically with little connection to H. rossouwii). The implication was that it could have been taxonomically related to those elements in terms of geographic distribution. I was still puzzled by the relationships of H. serrata when I wrote (New Haworthia Handbook :55, 1982) that collections by C. Burgers from the coastal limestones might throw more light on the matter (Fig.6 MBB6985 H. mirabilis var. calcarea).
Fig. 01a. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.rossouwii 1 043Fig. 01b. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.Fig. 01c JDV87-47 H. rossouwii, Oudekraal, Heidelberg.Fig. 01d. JDV88-53 H. rossouwii. Oudekraal, Heidelberg.Fig. 02a. JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 02b JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 03a. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03b. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03c. MBBsn H. heidelbergensis. E Heidelberg.Fig. 04. JDV87-3 H. heidelbergensis var toonensis. Matjestoon, SW Heidelberg.Fig. 05. MBB6859 H. floribunda var major. S. Swellendam.
In his revision of Haworthia (1983), C.L. Scott recognised my species. He confused it, and illustrated it, with a photograph of H. mucronata var. mucronata sensu Bayer (1999), which he called H. mclarenii. This does exemplify the problem of look-alikes in the subgenus, and this curiously does have some connection to the actual problem.
When I wrote Haworthia Revisited (1999), I was still not sure what the real affinities of H. serrata were. I mentioned that my 1982 observations were vague because of the unusual elements which occurred in the coastal limestones. I described these as new: H. mirabilis var. calcarea from De Hoop Nature Reserve (see Fig.6), H. variegata var. petrophila (CBurgers2158) from Karsriver and H. heidelbergensis var. minor from N Bredasdorp (Figs.7 JDV86/5). In my 1999 classification, I placed H. magnifica var. paradoxa (Figs.8 JDV86/78 & 94/89) in H. mirabilis, suggesting that it may have a connection there via the var. calcarea and an element from the Potberg (CBurgers 2018) which I have not yet seen in the field myself (since I wrote the article I have in fact seen this population, MBB7356, and it is discussed in the Chapter re H. mirabilis). The connections of paradoxa must be viewed in its relationship to H. emelyae var. major, in the same way that serrata is compared with H. emelyae var. multifolia. I also suggested that H. serrata had a very close resemblance to some forms of H. mirabilis var. sublineata from south of Bredasdorp. This present paper addresses some of these observations again.
Fig. 17b. Fig. 06a. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 06b. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 07a. JDV86-5 H. heidelbergenis var minor. Rooivlei, Bredasdorp.Fig. 07b. JDV86-5 H. heidelbergensis. Rooivlei, N Bredasdorp.Fig. 07c. JDV86-5 H. heidelbergensis var minor. Rooivlei, Bredasdorp.Fig. 08a. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08b. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08c. JDV98-89 H. mirabilis var paradoxa.
New Records In June 1999, I revisited H. serrata at Oudekraalkop (MBB166) again, this time together with J.D.Venter. Despite rumours of over-collection, it was still very abundant, although localised. It was also still present at Koppies (Fig.9 JDV99/6) a short distance to the west, but in very low numbers at a badly burnt site. P.V. Bruyns (priv. comm.) has reported it at a more northerly site as well. This could be where J.N.O. Uys also showed it to me at a second site on Oudekraal when he first introduced me to this species. It struck me again how similar the plants were to H. mirabilis (particularly the var. sublineata, Figs.10 MBB6639, bearing in mind the vast variation that exists there) and to H. heidelbergensis var. minor. The visit in June ’99 was a general excursion to explore the reported presence of H. maraisii at Koppies (ex E. Esterhuizen), in relation to that species and heidelbergensis as I know it in that general area. Our finds and observations led us later in the year to explore the area north of the Potberg. The extraordinary finds there led me to further explore westward to Bredasdorp itself and to see for myself the var. calcarea in the De Hoop. Two visits for that purpose were fruitless, but thanks to Dennis DeKok, I did locate it in the company of Lawrence Loucka in July 2000 (MBB6985-fig.11). This latter excursion was particularly fruitful because, by great good fortune, we discovered H. serrata in unmistakable form north of Bredasdorp (Soutkloof, Fig.12a & b MBB6983), and Nooitgedacht, MBB6984). Both sites are west of Rooivlei where heidelbergensis var. minor occurs. We also collected variegata var. petrophila (Fig. 13 MBB6986) west of the Karsriver limeworks. For that variety I have C. Burger’s record (CB2158) from east of the limeworks, and a collection I made at Renosterfontein (Fig.14 MBB6632) on the east bank of the Karsriver in 1997. This latter collection, in particular, reminded me in both form and colour of H. emelyae var. multifolia (see fig.2).
Fig. 10a. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10b. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10c. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10d. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10e.Fig. 10f.Fig. 10g.Fig. 11a. MBB6985 H. rossouwii var calcarea. DeHoopFig. 11b.Fig. 12a. MBB6983 H. rossouwii. Soutkloof. Fig. 12b.Fig. 13a. MBB6986 H. petrophila KarsrivierFig. 13b.Fig. 14. MBB6632 H. rossouwii var petrophila. Renosterfontein, Karsriver.
The conclusion from seeing H. serrata north of Bredasdorp, and both var. calcarea and var. petrophila, is that this is the prime connection. It seems now more improbable that the var. petrophila is as close to H. variegata as I had supposed, and that the var. calcarea is closely connected to both H. serrata and to var. petrophila. The other observations remain valid viz. that there is a strong resemblance between serrata and mirabilis var. sublineata; but particularly that serrata may be connected to heidelbergensis via the var. minor. However, heidelbergensis in its general relationships is extremely complex. It is interfused with H. maraisii, it may be the direct (if complex) geographic extension of H. mirabilis in the context of the relationship between that species and maraisii, and it may have affinities and continuity with both H. variegata and H. floribunda. There is still another problem: H. variegata var. modesta at Kathoek (Figs.15 JDV97/24) and Potberg (Figs.16 MBB6542). My collection of this element at Kathoek (Figs.17 MBB2551) suggests that it is possibly a small very proliferous sand dwelling serrata. J.D. Venter’s collection from the same farm is a more robust element with rather longer, more erect leaves which suggests H. variegata. My Potberg collection (fig.16) is another small element which I relate to variegata although the leaves are shorter and more turgid. It must be borne in mind that H. variegata var. hemicrypta occurs north of the Potberg on the gravelly flat so that there is ecotypic variation in evidence. Further work will have to be done to elucidate what could be an unresolvable situation i.e. the Potberg is home to variants which link all the elements so far named. The evidence for this is already available, but insufficient to crystallise a solution.
Fig. 15a. JDV97-24 H. vaiegata var modesta. Kathoek.Fig. 15b.Fig. 16a. MBB6542 H. variegata var modesta. Potberg.Fig. 16b.Fig. 16c.Fig. 17a. MBB2551 H. variegata var modesta. Kathoek.Fig. 17b.
There will be difficulty in considering the extent and the degree of similarity of H. serrata to other populations. Some of these which include serrata-look-alikes are:
Fig. 18. JDV96-52 H. heidelbergensis var . N Kathoek.Fig. 19. JDV93-51 H. heidelbergensis. Melkboom.Fig. 20a. JDV87-4 H. heidelbergensis var scabra, Leeurivier, Drew.Fig. 20b.Fig. 20c.Fig. 20d.Fig. 21. JDV97-49 H. mirabilis var consanguinea. Dwarswaterkloof, MacGregor.Fig. 22. JDV88-83 H. mirabilis var consanguinea. Dwarswaterkloof, Macgregor.Fig. 23a. JDV96-38 H. maraiss var notabilis. Agtervink, Robertson.Fig. 23b.Fig. 23c.Fig. 23d.Fig. 24a. JDV98-1 H. maraisii var notabilis, Wolfkloof, Robertson.Fig. 24b.Fig. 25. JDV97-30 H. cf. heidelbergensis. N Ashton.Fig. 26. JDV86-1 H. mirabilis var triebneriana. Stormsvlei.
Some of these populations and their variants are dealt with in other chapters (Particular note must be made of H. elizeae which has to be considered in relation to H. rossouwii).
H. serrata was coming into bud in July a good five months after mirabilis had started to flower. This shows that my predictive thoughts about the relationship with calcarea and petrophila may not be particularly sound if flowering time is considered. Calcarea flowered a little before mirabilis var. sublineata in January, petrophila flowered in February while Roovlei heidelbergensis and Leeurivier (Drew) heidelbergensis flowered in December and March respectively. Flowering time for other collections needs to be observed, noting that this is not necessarily definitive either (for example while H. mirabilis var. badia is essentially late-summer flowering, single plants may flower in winter.
When I consider how much time I have spent in the area, it was really surprising to find serrata so far west. It means that something very interesting can still be found in the large triangle Swellendam/Bredasdorp/Riviersonderend, and that exploration will have to be done on a very small scale. The Karsriver and Renosterfontein petrophila are just smaller more spinose versions of Oudekraal, and the leaves are more spotted on the reverse. Calcarea was previously known from two clones which had rather shorter leaves and, perhaps, fewer leaves per rosette. This, the locality, and the spotting on the back of the leaves would have been the reason for opting for a relationship with mirabilis. We only found it with quite detailed directions because it was such a small population and so cryptic, consisting of three tiny locations under bush with a total of about 15 clumps. One location could have been the product of vegetative propagation with six to seven clumps. The second was similar with only two clumps and evidence of porcupine activity around the clumps. The third covered about 15sqm and there were another 15 or so plants as clumps or as single rosettes. These did vary and only one clump really resembles the two I have seen in cultivation viz. the Karoo Garden plant of C. Burgers, and a plant I received from Dennis DeKok. Burgers, however, appears to have made two collections from De Hoop; this needs to be confirmed. The plants seen by me in habitat do vary quite a bit and the range of variation equates that of both serrata ex Oudekraal (the Koppies population is reduced to about five plants) and north of Bredasdorp. The mean length of the leaves and leaf number per rosette may be shifted to the shorter and fewer, respectively.
The two Bredasdorp populations of H. serrata are remarkable for the similarity of the plants to serrata at the type locality. The habitat is a little different and at the westernmost of the populations, the plants cluster much more freely than the Heidelberg populations. At Koppies the plants are on a Witteberg substrate, whereas at Oudekraal and N Bredasdorp, it is Bokkeveld shale. We may have a slightly biased image of serrata with turned out leaves; the plants can be quite globose, and I did illustrate a plant like that in my original description when I drew the comparison with herbacea.
Where serrata really differs from mirabilis is by the greater number of leaves per rosette, the thinness of the leaves, narrowness at their bases, lack of a mid‑leaf swelling and by the incurving of the leaf tip; that character is fairly significant. H. mirabilis var. sublineata grown recently from seed has proved to be remarkably variable, but none of the plants have incurving leaves. Some of the clones are very similar to serrata, but the flowering time is earlier in the year and more in keeping with H. mirabilis in that respect.
Discussion H. variegata var. petrophila and H. mirabilis var. calcarea should be transferred to the species H. serrata. The same may be true for H. heidelbergensis var. minor. However, H. heidelbergensis has a complex distribution and there are other complexities in variation that preclude a definitive statement. It is quite probable in the light of the findings discussed in Haworthia Update Vol.1, that there is no discontinuity which satisfactorily and convincingly can be claimed to distinguish species. This is probably just another example of a more general problem which I have experienced with the identification and classification of many plant genera.
I am conscious that I have not dealt fully with the problem in omitting mention of co-occurrence (I mean sympatry – when two obviously different elements grow together at the same site or very closely adjacent). I see this as the best measure of discontinuity. Thus if two elements grow in close proximity, even if not sharing the same habitat, they are probably discrete. This does not preclude the possibility that they are simply ecotypes and this is taken into consideration. However, I have before pointed out that a taxonomic solution which can be deduced in this way for populations might not be true for the entire distribution of the species concerned. It is evident that serrata generally occurs alone. At Oudekraal and at Koppies, there is a floribunda/maraisii element in the vicinity. In the case of heidelbergensis var minor, both turgida and maraisii are present. I have always been struck by the similarity of mirabilis and serrata, and by the fact that they do not co-occur. Mirabilis, in the context of this paper, is presently not known east of Napier except at Bredasdorp and Mierkraal south of Bredasdorp. This is as the var. sublineata and var. mirabilis respectively, which differ from serrata in respect of flowering time, less leaves per rosette, and the non-incurving leaves. Curiously, it appears that mutica, at least in the Bredasdorp area, grows generally alone too, although at Haarwegskloof (Bredasdorp) it does grow with heidelbergensis. At Drew it is in the close company of both maraisii and heidelbergensis, and south of Stormsvlei is near to a maraisii/mirabilis intermediate. Thus even an element as apparently different as H. mutica may presumed to be, cannot be ignored.
However, another consideration now enters the picture with the discovery of serrata so far west. There are three poorly understood elements which were reported to have come from Bredasdorp and Napier respectively. The first of these was H. bijliana von Poelln. that does, however, seem to belong rather with H. arachnoidea than with anything else. G.G. Smith appears to have had a plant of this ilk which he received from Maj. Venter 9, from Caledon. The other two, however, are H. altilinea var. bicarinata Triebner and H. rossouwii v.Poelln. They were both described from the same collection viz. Napier at Bredasdorp, Rossouw in Triebner 1059. The former was never illustrated and the description is rather brief … “Backs of leaves always with two keels and with teeth. Leaves 3-5cm long, 6-8mm broad, with 4-6 dark lines in lighter part, with two keels at back, keels with up to 2mm long denticula”. This description is in the joint paper by Triebner and von Poellnitz, but authorship is attributed to Triebner alone. von Poellnitz later retained this variety when replacing the name altilinea with mucronata (note that in the context of the names which I use, Scott confused my serrata with mucronata as well). This is very curious because it is evident from the description of H. rossouwii, which is very much more detailed, that another plant from the same collection is in fact again described. The differences are essentially that von Poellnitz gives the leaves as 5-6cm long, and 8-10mm wide as opposed to the respective 3-5cm and 6-8mm by Triebner; and with three lines on the ‘end surface’ as opposed to four to six by Triebner. This says a great deal about the detailed descriptions which are routinely called for, and which in Haworthia seldom amount to much because of the problem of look-alikes, and the variability within and between populations.
I repeat the von Poellnitz description here to illustrate that it is a very good description that nevertheless omits some key information:
H. rossouwii von Poellnitz spec.nov. (Stemless rosette, spirally many-leaved. Leaves rather pale green, nearly oblong lanceolate, acuminate, on the face somewhat retused near the apex thus forming a triangular face; on the back, towards the apex, with sub-pellucid tubercle-like spots, denticulate on the margins and keels; the triangular face strongly acuminate, sub-translucent, not tubercled, but carrying some sub-pellucid small teeth, and traversed by a few longitudinal lines).
Stemless many-leaved rosette about 5-6cm in diameter. Leaves more or less erect, 5-6cm long, 8-10mm wide, about oblong-lanceolate, tapering gradually, pointed, slightly widened at the base, almost straight or a little bent to one side near the tip, rather light green, dull. On the face flat or usually somewhat convex, near the tip slightly bent backwards (retused) at an angle of 20 degrees; on the back, from the middle outward or near the tip obliquely keeled, or somewhat more rarely with two keels, near the tip with a few, often elongated, very often raised slightly pellucid spots, and frequently with a few leaf-coloured tubercles. The end-surface half-pellucid, 20mm long or somewhat longer, 7-9mm wide at the base, triangular, tapering very gradually, pointed at the tip often slightly curved inward, somewhat convex, smooth, rarely with a few indications of tubercles, but with some semi-translucent teeth (these teeth loosely arranged in a lengthwise middle line, now and then also in another line nearer the margin and very rarely scattered about in small numbers) with three long, occasionally interrupted, green lengthwise lines, of which only the middle one may reach almost to the apex, and usually with a few very short line; leaf margins and keel with pale teeth, or near the base often leaf-coloured, teeth, up to barely 2mm long; end-bristle pale, with small side bristles, 5-7mm long.
… On account of the teeth on the end surface stands near to H. schuldtiana and to H. paradoxa (I treat these respectively as H. maraisii and H. mirabilis var. paradoxa) but is easily distinguished from both. Its flowers are thus far unknown”.
The omissions are the flower of course, and there is no stipulation of the approximate number of leaves, and leaf thickness. The weakness is that it is a single plant description, evidenced by the fact that another plant from the same collection is accepted as a variant of a totally different complex.
In the past I have I regarded rossouwii speculatively as a synonym of H. mirabilis, due largely to the fact that it was recorded from the Bredasdorp area (and to the ambiguity of the type). Another reason was the similarity I observed between the illustration of H. rossouwii in Kakteenkunde with H. heidelbergensis var. minor and with H. mirabilis var sublineata. I opted for the latter (with which Scott was in accord) and the similarities are also recorded in the discussion above. I dismissed bicarinata as insufficiently known because I assumed that Triebner may have had some better reason to have equated his specimen with the Little Karoo H. mucronata. Possibly there has been a mistake in the Triebner numbers. It is this discovery of serrata so close to Bredasdorp, and equidistant from Napier, which revives and also casts new light on the problem. I conclude that my H. serrata is a later synonym of H. rossouwii of von Poellnitz.
There are still two other considerations. In my 1999 revision, the fact is that I placed paradoxa as a variety of mirabilis while discussing the possible connection of these elements via calcarea. In the light of this paper it is evident to me that paradoxa could also be linked to serrata rather than with mirabilis. The same is true for emelyae var. multifolia. I would not like to commit myself to this probable solution at this stage, as I think more evidence should be obtained. It is also improbable that, however attractive, the solution will not avoid impacting on H. emelyae, H. magnifica and H. heidelbergensis. It is also evident that the requirements of the ICBN are that should paradoxa indeed need to be incorporated into serrata, more name changes will result, as paradoxa will have priority over rossouwii. My opinion is that this is a fundamental weakness in the code, which inhibits speculative treatments, and also which results in smaller taxa becoming the typical main element of a species. My further opinion is that it is a weakness of the Code that the description of an element automatically creates an equivalent taxon. This may be acceptable and desirable where the elements are clearly separable and geographically defined, but not where the typical element is required to absorb any other variants which are not so defined. Thus H. paradoxa var. paradoxa could become the main component of a species where it is actually a geographical isolate. In a system which does not have an adequate general definition for ‘species’, this probably does not matter; whereas it might if there is a definition which suggests that the ‘species’ is an operating living system (see Bayer in Asklepios 77:3, 1999).
The formal statement of this paper, which would better be regarded as an hypothesis subject to test, is thus as follows:
H. rossouwiivon Poelln. in Kakteenk.7:75 (1938). Type: Napier near Bredasdorp, collected by Mr Rossouw, South Africa Police in Napier, Triebner 1059. Not preserved. Lectotype: designated by Breuer & Metzing 1998, icon in Kakteenk. non syn. H. mirabilis sensu Scott 1985, Bayer 1976, 1982, 1999.
H. altilinea var. bicarinata Triebn. in Triebner & von Poellnitz in Feddes Repert.Spec.Nov. 45:170 (1938). Type: Napier at Bredasdorp leg. Mr Rossouw (=Triebn.1059). Not preserved.
H. serrata Bayer, Jl S.Afr.Bot. 39:249(1973). Bayer :55(1976). Bayer :147(1999). non Scott :62(1985). Type: CAPE‑3420(Bredasdorp): Oudekraalkop, Heidelberg (‑BA), Bayer 166 (NBG).
H. rossouwii var. calcarea (Bayer) comb.nov. H. mirabilis var. calcarea Bayer in Haworthia Revisited :110(1999). Type: CAPE-3420(Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG, Holo.).
Material seen:- 3420(Bredasdorp): NNE. Buffelsfontein, Potberg (-BC), Burgers 2018 (NBG). De Hoop (-AD), C. Burgers 1648 (NBG), Bayer 6985.
H. rossouwii var. petrophila (Bayer) comb.nov. H. variegata var. petrophila Bayer in Haworthia Revisited:159(1999). Type: CAPE-3420(Bredasdorp): Renosterfontein (-AC), Burgers 2158 (NBG, Holo.).
The provisions of this solution are that the epithet paradoxa may be instated preferentially, and that emelyae var. multifolia and variegata var. modesta also be included.
A second provision is that the species H. bijliana v.Poelln. be rejected as insufficiently described and sourced, and that the type (the illustration in the Berlin herbarium) is ambiguous – as indeed are many Haworthia types and descriptions based on single plants and not correctly tied to geographical origin. This is demonstrated by the citation by VPoelln. (Feddes Rep. 1936) of this species viz. bijliana from Klawer and Eenriet and also Feddes Rep. in 1937 from Springbok – which cannot be anything other than H. arachnoidea. In this last publication he cites a specimen from Bonnievale, another from Ezeljacht Oudtshoorn and also sinks fergusoniae here which in fact was supposed to have come from ‘near’ Grahamstown.
H. bijliana Poelln. in Feddes Repert. 27:134 (1929); Bredasdorp, Mrs va der Bijl. Type van derBijl s.n. in (B). Not preserved. Lectotype desig. Breuer 1999, unpublished photo. icon. (B). Species rejecienda.
Acknowledgement I am grateful for the company on the field trip of Lawrence Loucka of Connecticut (USA), and that of my wife, Daphne. The assistance of Paul Botma of Cape Nature Conservation is acknowledged over and above the general acknowledgement to that Department. Mr. C.L. Neethling, Mr. Thys Swart and Mr. Pieter deKok were most kind and helpful in respect of access to sites. I am also grateful to Kobus Venter for support, advice, and constructive exchange of ideas. Not least to Steven Hammer, with comment from Bob Kent, who was so kind as to read this manuscript critically and make corrections. Residual errors will be mine.
Addendum Alsterworthia Vol 1:13(2002) “A comment on Bayer’s latest taxonomic contribution” by Ingo Breuer. [link] With reference to the comments in Alsterworthia 1:13(2002) regarding the identity of H. rossouwii and H. serrata:
Notes
Fig. 27 H. rossouwii. Vpoelln. type.Fig. 28a. MBB6984.1 H. rossouwii. Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.Fig. 28b.Fig. 29a. MBB6984.1 H. rossouwii. Colour photo originals of the preceding. Fig. 29b. Fig. 30. MBB6984.2 H. rossouwii. Nooitgedacht, Bredasdorp. A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second. Fig. 31. MBB6983 H. rossouwii. Soutkloof, N Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might. Fig. 33 Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Fig. 27 A copy of the von Poellnitz lectotype of H. rossouwii.
Figs 28a & b Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.
Figs 29a
& b. The colour originals of the preceding.
Fig.30 A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second.
Fig.31 A photograph of a plant of MBB6983 Soutkloof, north of Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might.
Figs 32a
& b. Scanned and manipulated copies of figs 28a & b.
Fig.33
Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Figs.7a, b & c. These are clones of JDV86/5 H. heidelbergensis var. minor, Rooivlei, NNE Bredasdorp. This is a small population, of also small plants, from northeast of the collection MBB6984, Nooitgedacht. The plants partly resemble H. rossouwii from there, and partly the var. petrophila from Karsriver. Classifying them as a variety of H. heidelbergensis is not necessarily correct. Considering habitat and substrate, one must consider that ecotypification is playing a role, and that was habitat available, there would be further populations which would confirm continuity with these known collections.
Regarding Figs.28a & b. A bit of play with light and focus and the keel and under-leaf spotting could be varied. I would like to suggest that they be considered them faded, less sharply focused, printed with less contrast and on a coarser grained paper. Figs.32a & 32b are in fact such scanned copies of Figs.29a & b. I do not think it even necessary to consider that the original rossouwii of Fig.1 endured a long journey in the post to a foreign land. The assertion by Breuer in Alsterworthia is that not only are the plants I figured as H. rossouwii different from Fig.1, but that this Fig.1 lectotype is “probably” of the maraisii-complex. This is at variance with…
1. the Triebner description of the same collection as a variety of H. mucronata;
2. with Col. Scott who regarded it as H. mirabilis. What to say of von Poellnitz’ own assessment – the man who described both species i.e. maraisii and rossouwii in the first instance.
3. with the facts of the pictures before you.
What should an observer make of maraisii growing very near to rossouwii at Oudekraal and near to rossouwii at Napier? Can we really see a closer resemblance of Figs 27 & 28 with those long attenuate, double-curved, thin leaves – to H. maraisii rather than to H. serrata. I truly do not see that such a judgement can be seriously considered in the light of Breuer’s interpretations. It is of course valid when one considers all the variants that I illustrated in my article above and in terms of my over all classification.
Generally these plants of the SW Cape tend to have the double curvature of the leaves as is evident in the VonPoellnitz lectotype, but one can find in the same population and/or taxon, clones with incurved, outcurved or erect leaves (see fig.30). Broadly speaking H. maraisii has thick recurved leaves. But this is a non-argument even if one (outside of a “lumping vs. splitting” red-herring) considers the complexity of the so-called maraisii-complex, and I am sure that Breuer does not. I frankly think that most observers cannot even dream how complex it is and that it is just braggadocio to write as though anyone can. Breuer implies that one can make the inference of similarity between rossouwii and maraisii from the illustration (Fig. 27), the description and its given locality. This is patently and self-evidently false as is evident from the fact that I used this same information to reasonably deduce the similarity of rossouwii and serrata.
I did not originally submit a copy of the von Poellnitz lectotype, perhaps because I felt that the response “you will see that rossouwii is quite a different plant from serrata“, is what I could have expected from an uninformed audience! The fact is that all the evidence, circumstantial and descriptive, show that they are the same. Including the lectotype would have just been “overkill” and I have never before felt required to write for a readership with less than common sense or basic knowledge, nor for obstructive readers.
It should not be necessary to say this, but the character of the flowers cannot be specified in the way that writers are inclined to suggest. They have this same enormous range of variability and it is just fraudulent to say that there are floral characters by which these taxa (the varieties of H. rossouwii which I now recognise) can be unmistakeably differentiated. It is the typological concept which drives people to think there is a single description which can encompass the description of a taxon. This is nonsense. It is a statistical problem and we have the difficulty of establishing the probabilities associated with “decision”. Breuer writes “… von Poellnitz’ description probably depends only on 1 or very few selected plants and no other records are reported for this taxon”. This is circa 1945 circumlocution, it is unnecessary and it is untrue. There was never a question of “selection”, and furthermore we have the Triebner description of the same collection as H. altilinea var. bicarinata. Why this sudden doubt about the interpretation of types. It is a shifting of goalposts.
There are few collections in flower now in April, but I could take this picture of three flowers from the one collection MBB6672 H. maraisii – Koningsriver, SW Robertson (Fig.33). It is not an exceptional case or an exceptional collection. But one can see the differences in thickness of peduncle and variation in the individual flowers. It can happen like this, albeit to lesser degree, on the same plant. Colour is also frequently quite variable between and often within collections. Many years ago I undertook to illustrate all the flowers in my maraisii collections. But they were so variable that I just gave it up. The variability in the flowers mirrors the variability of the rosette and the latter is just a little easier to deal with. One has the added problem with flowers, of the changes which take place during the opening and aging process. So then how does one define the flower, say, of H. mirabilis. It cannot even be done for the range of leaf rosettes, and this has now been shown in the literature many times.
May I also point out that one of my understandings of botany and plant classification is that there is a tendency to overreach ones talents. I have freely admitted that I am there because of the paucity of talent. I frankly do not think the environment is enriched by the particular article I now refer to in these added notes.
I can point out that there are writers who really persist in flogging dead horses. The result is phantasmagoric fanfaronades. The article concerning the name H. correcta (Alsterworthia 1:6, 2001) is not particularly accurate either and indeed it may be my own fault. Smith explained that Mrs. Blackburn did not in fact collect correcta ‑ it was a Mrs. le Roux and the plants came from near Vanwyksdorp. Later Mrs. Ferguson sent a plant from 26 miles east of the town. Thus from this information H. correcta has to be a later synonym for H. emelyae.
Regarding types:- the painful realisation was actually expressed by Schelpe in the distant past, and brought home to me many times. These old types are doubtful and ambiguous. I was faced with that option long ago – just forget them all, or try to reach a compromise with the understanding and information available at the time. H. rossouwii is a case where there is real new information which would have made me use the name rossouwii in 1976 and before, had it been available. But here, had I persisted with the name serrata for this new material MBB6983 and 6984, there is absolutely no doubt in my mind that I would have been criticised for failing to see the connection to rossouwii. Not that this would concern me unduly, for I think there is actually very little benefit in this mindless re-interpretation of types which takes us nowhere. Notwithstanding, and without compromising, anything so far written:- the really disconcerting thing about the whole matter is that without the locality data, it will be possible to generate an argument bolstered by illustrations, to show that rossouwii was in fact H. decipiens, H. cooperi or H. herbacea (among others), of the option less improbable than H. maraisii.
The closing paragraphs of the article are a rather hackneyed parody sung by most “wannabe” taxonomists. Of course it is right when Breuer says if we lump it means we would have to lump many other things. It is just as true to say, if we split we have to split many other things. That does not make sense either. This is ancient stuff of 60 years ago and it seems strange that it still provides so much fodder for writers, who have in fact no rational species concept to begin with. The Linnaean system was based very largely on floral characters and hence taxa at all levels are based on “related floral characters”. To suggest that this pertains in some special way to “series” is not correct. This sums up the article in Alsterworthia. ♦
This essay was published in Haworthiad 16:86, 2002.
I wrote an article about Haworthia rossouwii in Aloe 38:31(2001), in which I resurrected this old name to replace that of my own H. serrata. This was necessary because I had found this plant (because of its localisation and its abundance there, it is better to say ‘this species’) at two places near Bredasdorp as opposed to where I had described my species from near Heidelberg.
One needs to know something about the geography and geology of the Southern Cape (and the Overberg as a part of it is also known) to really follow all the ramifications of any discussion about Haworthia, including this one. In fact one needs to know a whole lot more, and I will also try to explain that and its implications for the collector and Haworthiophiles. This “whole lot more”, I will call the Corporate Mind because it includes so much – so remember CM! If I regard H. rossouwii as a species, I have to consider all the plants and all the places where they grow in order to determine the nature of this particular system of living things. As I explained in my article, there is a problem with the fact that little is actually known about the Haworthia of the Overberg. They occur in small populations scattered over a fairly wide area which has been heavily impacted on by agriculture. Thus about 90% or more of the Overberg is now wheatfield or pasture. Like Gasteria carinata, which is also a Southern Cape species, Haworthia is associated with rocky outcrops and thus also with the geographical erosion and drainage systems of the area. It is quite probable that cultivation has had relatively very little impact on Haworthia in terms of actual available and suitable habitat.
The problem is to get to all the small potential habitats and explore these at a scale which will help to resolve the problems of variability, relationships and classification. While writing about H. rossouwii, I was very conscious of this problem and the fact that my explanation is not complete. There are some collections by Chris Burgers which I have not seen, and there are many potential habitats which I have not explored either. The place Kathoek and H. variegata var. modesta which grows there is a particular case in point. Kathoek is at a point midway between the Breede River and Bredasdorp. It is just east of the De Hoop wetland/stream and on the northern edge of the Coastal limestones. In the west, also on limestones, is H. rossouwii var. petrophila which I first placed with H. variegata. In the south (also on coastal limestone) is H. rossouwii var. calcarea, which I originally had with H. mirabilis. To the north is H. variegata var. hemicrypta which is on Silcrete of a quite different geological origin. But H. variegata variants, as I interpret the populations, are also on and about the Potberg to the east which is Cape Sandstone.
My problem is “How do these populations actually relate to one another?” What is missing? What can I find to better explain their similarities and their differences? So at the back of my mind is the wish to explore the Coastal limestones between Bredasdorp and the Potberg and find out more. But it is not as easy as that. I also want to know more about H. mirabilis, H. maraisii, H. heidelbergensis, H. mutica, H. minima, H. marginata and H. pumila, which all also occur in the Overberg area and each have question marks attached. They each present problems that I would like to resolve for my own satisfaction, without being confused and befuddled by others who are toying with the same body of knowledge at different levels of expertise and ignorance. In this play we are using words, terms, names and conventions. For this play we need to have a commitment to truth and use the same language or else it is all just meaningless.
So we come to what took me to the Overberg in November 2001. Was it was just my curiosity about all those inviting rocky unexplored places? Is it to learn something more myself? Is it to impress anybody? Actually it was many things including a fascination with chameleons – largely my wife Daphne’s. In March 2000, Daphne and I had come across a most remarkable chameleon at Napier. It was on a small shrub among whitish dried grass growing on a calcareous outcrop strewn with white quartzitic pebbles. The tiny reptile was ivory white, with a stunning array of small orange tubercles on sides and back. One has to know the Cape Dwarf chameleon (Fig.1) and its cousin, the Robertson Dwarf, to know how beautiful and amazing these creatures can be. Another thing was H. mutica and my fascination with collections of this species and their histories of discovery and exploration which are stories of their own. Still another was the news that the Botanical Society had funded a researcher to explore the remnants of the vegetation of the wheat areas. What had he found? Apart from Haworthia, I had once been involved in a preliminary attempt to map and identify vegetation remnants for conservation purposes, and had vainly tried to draw attention to the curiosities of the Silcrete outcrops and remnants of vegetation among the wheatfields.
This researcher was an old acquaintance, Nick Helme, who kindly told me what Haworthias he had seen in his exploration. There was not much new, except for H. mirabilis east of Caledon, and this became our first stop. The site is south of the main area for the species which is along the Riviersonderend river north of Caledon. It does, however, occur in recognisable form south-east of Caledon, also around Napier where it transmutes to some degree (inter alia to the var. badia), and then also at Bredasdorp. There it also transmutes to the var. sublineata (sandstone) and var. mirabilis (the old mundula – and there may be some other way out, but for me this represents the low cussedness of botanical nomenclature). What H. mirabilis does to the north and east of Riviersonderend actually fully explains the problem of classification of Haworthia, and who knows if anyone will ever write that explanation – and truthfully.
Nick Helme’s site is a deep valley which drains to the south. Typical of the area, are these small drainage areas which seldom have permanent water. There may be odd pools lasting into mid- or late summer. There are often exposed shale ridges which host a particular array of succulents and other species – and are often home to owls too. The first problem is access and where does one find the landowner after one has succeeded in finding the place? If one does find the dwelling, will it be the right one and will one find the landowner. Will it be a big aggressive StBernard and more aggressive and ugly Rottweiler reflecting the mindset and security fears of the owner and grossly offending my own, like once near Worcester – or will it be a lonely and over friendly sheepdog. November is a very bad time. This is when the wheat harvest is on. At the end of a bad season, when the crop is drying and it has to be cut and reaped the weather is playing its usual strange tricks, farmers cannot be expected to be approachable. But we did find a helpful soul. From the farmhouse, we drove twenty kilometres back to the main road to the roadside stall, where a family member, Herman DeKock, gave us the necessary permission to venture onto the property.
It does seem an easy business, but in fact it becomes very complicated. Nature Conservation regulations are strict, and land ownership, presence, trespass, stock theft, personality and who knows what else, influence the mindset when setting out an expedition of this kind at all. With those problems all out of the way, and only then, is one able to really enjoy being out in the field and communing with nature – which it is all about in the first place. So we found Nick’s site. We climbed through the fence (s) and tried not to disturb the forlorn sheared, miserably self-conscious sheep that clustered around the earthen dam. It was hot and dry and the most promising spots were quite a long way off. One has to plan one’s approach as the southern slopes are often quite densely vegetated and very uncomfortable to traverse. Snakes are also a consideration and especially in early summer. On this trip we never saw one, although Daphne surprised, and was surprised, by a Lynx drinking at a waterhole. So we did find the plants that Nick had said and took a photograph for the record (Fig.2). Just ordinary H. mirabilis var. triebneriana (cussedness – there should be no typical variety, just a broad general name mirabilis).
From here of course we went to the chameleon at Napier. What a laugh! We had difficulty finding it on the same day on the same bush, five minutes after first seeing it. So expecting to find any chameleon at all, and I have never seen a chameleon in the field in all my years of plant exploration, was quite the wildest dream. But with a dossier of 60 individuals recorded on our small-holding at Kuilsriver over a period of about a month, we felt entitled to hope (all contributing to CM). It is also a most wonderful excuse to be out in the country.
We headed for De Hoop. This is also quite a story. Where do we stay? The DeHoop Nature Reserve is a wonderful place with great accommodation, but it is not where we wanted to be. The staff does not know what it is we are looking for, what our CM data base contains and what our needs are. We want to see the area around the north of the wetland. This is half private land, and half state-owned. Besides there is a travellers Lodge on the edge of the reserve which we understood offered easy and official access to the reserve, presumably to near where we wanted to be. Not so. It is one thing dreaming about this exploration while sitting at home with the map, and quite another to be faced with the problems of roads, fences, ownership, dogs, snakes, flies, mosquitoes, permissions, heat, cold, wet, terrain, bush, visibility, time, distance, misinformation, local “knowledge”, incipient old-age, food, water and the spirit of inspiration and energy which drive the legs (CM). So the Lodge was an unfortunate mistake (we did look, and we did also look where we were told an Haworthia did grow – it was Gasteriacarinata), and we did end up at the office at DeHoop Reserve. You see over the years, one comes to realise that helpful and kind as people by nature are, an awful lot of time is spent chasing red-herrings and wild geese (CM). The help and information one gets is very often of a negative hindrance rather than a positive help. So my real gratitude to the manager there, Mr Ben Swanepoel, who without serious qualm, entrusted us with the key to the northern part of De Hoop where we needed to be.
Unfortunately the problems listed above rather limited our activity. We exhausted our energy climbing up and down rocky slopes in an area which should have occupied us for a week. It is an area which has to be explored by someone at sometime. We found naught, which should not in the least be interpreted to say there are no haworthias there. The terrain is formidable and also most promising. But CM called.
We moved to accommodation closer to our budget restraints and nearer to our goals. Driefontein, where the owner graciously made the necessary contacts for us. What a place – the cold vegetated coastal limestone behind, and featureless cultivated endlessness in front. I have an old record of a collection of mine of H. mutica from “Beyersdal”. Subsequent travels indicated that place, name and memory did not match. So we went to Beyersdal. We did find mutica and it was a different place and a new record (Figs.3a, b & c). Who can actually describe the wonder of plants like these? In tiny patches of rock in a sea of wheat and low-grade (by rainfall and vegetation standards) pasture. Also the quiet, the two owls, also Gasteriacarinata again, this time the Haworthias among them – unusual. The extraordinary colour of the plants. This time the bronze colours of badia. Nearby we also visited an old, by thirty years, locality for H. heidelbergensis. Still there and astonishingly in among white quartz in a small outcrop, but in two little groups occupying hardly two square meters in the approximately two hundred available (Fig. 4).
Then we went to Vanderstelkraal. We had met the owner, Mr Adam Albertyn, who very kindly let us wander over his farm. CM. How does one explore a place like that without living there? It is on the edge. The agriculturally useless coastal limestone at the back, and the productive massively utilised Bokkeveld fields at the front. If terrain had allowed, there would not be so much as a hedgerow to have conserved anything. We saw some very interesting sites and exhausted quite a number of things, like energy, time, passion for the quest, tolerance to bad weather, ostriches, tight-strung barbed-wire fences, no roads and also to the heaviness of the overwhelming destructive productive activities of man. We did not find Haworthia. But I omit a lot, including all the good farming. Mr Albertyn owns the eastern side of the De Hoop wetland where he has a 4X4 route – and there is also a sixty-year old legend of H. minima in that particular area which I would like to confirm. Not this trip though. We did try to the west later, where CM eventually prevailed against us – aided and abetted by overgrowth of alien acacias and the negative vibe they seem to generate (by the side, at Vermaaklikheid, Riversdal, H. mirabilis var. paradoxa grows/survives under Acaciacyclops).
Then we decided to climb the mountain behind Kathoek. Why? I cannot remember why I climbed there in the first place, or how I even did it. Then I found plants in a burnt out area, under the remnant blackened skeletons of Protea – natural fire cycle encouraged by farmers. It does produce a grazeable growth and also invasive aliens. The plants of H. variegata var. modesta were just discernible sticking out from loose sand. Kobus Venter later also visited Kathoek and found plants which were more robust. The plants have spreading variegated leaves (in cultivation?) and while I had no real problem associating them with Karsriver, viz. H. rossouwii var. petrophila, they seemed to have a greater affinity with variegata at the Potberg and Luiperdskop. But my collection and Kobus’ seemed at variance. So Kathoek was a must. From the road CM prevails and it looks quite an expensive option in terms of CM resources. Actually the climb was quite easy. We found the plants before we expected to. They did not look like what we expected them though. They were densely clustering (expected) and in rock crevices (Figs.5a, & b) and pans (Figs.6a & b). The rosettes were tiny and reminiscent of even H. reticulata (see Fig.7). At one place a dense cluster under and in a restioid grasslike plantlet, looked just like a yellowish-brown windblown fallen mass of dry leaf bracts (Fig.8). What impression did it make? Nothing new. It provoked the existing doubt. The population is very similar to petrophila and less so to variegata and hence this variety, with the exclusion of the Potberg population COULD BE transferred to H. rossouwii. But this does not help much, because the Potberg var. modesta is less like the northern Potberg’s two populations and the Luiperdskop population of H. variegata var. hemicrypta, and not more like petrophila! So the CM has to carry this anomaly together with the position of calcarea until whatever else to be found is found. These finds are not going to make things easier to resolve in the CM, nor to explain in a way which assuages the CM of anybody else.
All the while we were pondering other clues to the nature and relationship of things. Like the endemic Braunsiivanrensburgii; a curious specimen of Carissabispinosa so laden with large red fruits it resembled a Pyracantha; Euphorbiasilenifolia everywhere, and what a strange distribution this summer-deciduous geophyte has; and what about Asparagus; what is the relationship of A. capensis, A. mariae, A. suaveolens and A. stipulaceus? Where was Aloe brevifolia or A. saponaria? What was that pugniform Euphorbia at Driefontein or the bayeri-like one at Vanderstelkraal?
Unexpressed CM – we are using my son’s LDV as in my own car anything below well-maintained road is not negotiable. CM – the cellphone we arranged for failed. There is a call in my sub-conscious ‘vehicle needed back home’. We end the trip a day earlier and the message turns out to be true as there is a breakdown of one of the other delivery trucks.
On the way home we pick up some loose ends. We call at a spot north of Bredasdorp where in my lucky ignorance I once found mutica. In several subsequent visits I could not locate it, until one year Daphne and I together with Lawrence Loucka had a very thorough search of the small spot. We found about five plants in a spot about 1/2sq meter. At a later visit we found that the plants had seeded and there were many seedlings but crowded together with seedlings of Aloeferox. Generally Aloe and Haworthia do not mix. Aloe need more root room than Haworthia and Haworthia cannot tolerate competition from larger plants. So we thinned out both aloe and haworthia seedlings. This visit revealed the successful fruits of our activity (Fig.9). The two bigger adult plants were almost being pushed out of the ground by a burgeoning aloe seedling, but there were about 15 other vigorous and healthy plants to carry the group forward. CM – this was an interesting repeat observation of the small distances that seed and seedlings actually do commonly disperse. I have seen the same thing with hybrids where the hybrid is often within the multiple body of one of the parent species.
Mission ending, we made our last visit to Drew. This story is written more fully elsewhere. We had found the pollen source for a plant of H. mutica which I collected there more than thirty years ago and which Kobus and I had taken to the field again to see if seed would set. This surviving single plant is now a collector’s dream which I dub “Silver Widow”. No one had succeeded in finding plants around where a huge centre-point pivot installation now graces the landscape. We were sure the plants were gone, and taking the plant back there was a kind of forlorn gesture of regret that more had not been done to conserve the population – and that so much had been done to eradicate the plants (two hundred plants had been collected and sold to Triebner circa 1935). If anything can be done or could have been done at all to have saved its family? But it worked and we got seed. Where did the pollen come from The pollinator is mainly a solitary bee with a fairly limited flying range. The pollen parent had to be somewhere near. Daphne and I had made several excursions to add to the many different attempts that are known to have been made to find them. Finally I decided that we just had to look again and the plants had to be where we had failed to find them before. So what happened? We left the farm house and walked straight to the spot (getting ‘stung’ by the electric stock fence on the way) less than 300m away and probably about 150m from where we had left Silver Widow to be pollinated. There were about 35 plants (Fig.10) in an area of about 3sq m in flower under Eriocephalusafricanus. So now we returned to successfully collect seed.
The curious thing in CM and all this discussion about big things like Continental Drift, and Biomes, and Vegetation, and Habitats, and Ecotypes, is that H. mutica is not particularly concerned. Here at Drew it grows in an ancient river-boulder situation in what we can call renosterveld under Eriocephalus. At Beyersdal it is on the edge of a stream-bed on exposed Bokkeveld shale with Gasteriacarinata. At Hasiesdrift it is on a gravelly bank (Bokkeveld shale) with dense Aloeferox. Near Napky it is on silcrete with grass. West of Swellendam, it is on whitish-clay, probably associated with silcrete and growing within a restioid species and with Gibbaeum esterhuyseniae. I have also seen it on a fairly bare hillside with a scattering of quartzitic pebbles where the parent shale is too dense and shallow for cultivation or to support denser vegetation. The places hardly have anything in common other than their more general geographic containment, climate, and low biomass potential. How, ever, did they get to their widely-spaced, isolated island sites?
So the tale ends with the reflection on the criticism that I have not treated H. scabra and H. starkiana in the same way I have H. retusa and H. mutica. I have been “inconsistent”. Readers can research and educate their own individual CMs (corporate minds) to the point that they can form opinions which are consistent to the point and purpose of any communication. Aren’t we lucky that the Robertson Dwarf chameleon is a small animal (Fig.11 – by Scott Russel).
Acknowledgement: This fun work is really just about people. Daphne, who kept me company. Janet Albertyn at Nachtwacht, Herman Dekock of Jongensklip, Thys DeVilliers at Boskloof, Tom Ambrose at the cement-block operation at Napier, Rory Allardice at Buchu Rest-camp, Ben Swanepoel at De Hoop Cape Nature Conservation, Annette VanEeden of Driefontein, Mr. Dawid Beyer of Beyersdal, Adam Albertyn of Vanderstelkraal, Theo Blom of Soutspanvlakte, Jan Joubert of the adjacent farm, and Beth and John Humby of Sanddrift. Their good-will and kindness is unforgettable.
Fig.1 Cape dwarf Chameleon – Bradypodion pumilum. Markings, colour, tubercles and spines are individual characteristics. It is said that even the genus is in doubt!Fig.2 Haworthia mirabilis var. triebneriana, MBB7059 Jongensklip, Caledon. Note the reddish spines.Fig.3a Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. This plant is exposed to full afternoon sun for at least a few hours each day. Note the mucronate (pointed leaves).Fig.3b Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A more general specimen with rounded leaf-tips.Fig.3c Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A particularly compact specimen. In this population, the waxy sheen of the species seemed less conspicuous.Fig.4 Haworthia heidelbergensis var. minor, MBB7063 Beyersdal, Bredasdorp. Varietal names are just a geographical convention, as indeed even the species which I recognise, seem to be.Fig.5a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster under a small rock overhang.Fig.5b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster in an exposed crevice and visible as a dark mossy fill.Fig.6a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill.Fig.6b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill – a closer view.Fig.7 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Very reminiscent of H. reticulata say south-west of Robertson.Fig.8 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Barely recognisable to the undirected eye.Fig.9 Haworthia mutica var. mutica, MBB6982 Hasiesdrift, Bredasdorp. Nine seedlings in a tiny island of plants, in a bigger island of Aloe ferox and surviving vegetation on the edge of a wheatfield.Fig.10 Haworthia mutica var. mutica, MBB7064 Sanddrift, Drew. This lovely plant is nearly 120mm diam. and the largest of about 50 plants. They survive, and have survived in this small heavily impacted area for so long. There is no where for them to go when options must have been very limited to start with.Fig.11 Robertson dwarf – Bradypodium gutturale, photo by Scott Russel. Zoologists cannot determine if the Robertson dwarf is different from the Karoo dwarf, or from the two subspecies of the Namaqua dwarf. They use the same fuzzy logic that botanists do, plagued by low sampling and difficult application of statistical principles and probabilities.
Recently a cybernet note suggested that someone had a good understanding of the taxon/species, H. intermedia v.Poelln. That same writer has been a bit casual in describing new varieties, and in explaining what he actually means when he has used the species epithet accompanying them. Understanding is a very relative term. It may be extraordinary in terms of people who do not deal in the subject at all, high in relation to the knowledge level of the people with whom one is in ordinary contact and it may be very low in relation to people deeply involved in the subject. It may also be quite negligible in terms of truth and ultimate reality. With this in mind, I am going to take an opportunity to explain my own use/misuse of the epithet “intermedia”, and change it. For several years I have been plagued by articles and statements in Haworthiad (and elsewhere), which are not strictly true. In fact this distress goes back to correspondence with, and publications by, Col. C.L. Scott starting in 1965. So when Dr Urs Eggli recently kindly stated to me his approach as follows:
….”Classification (systematics) for me has always been an ATTEMPT to understand how biological diversity could have arisen. In contrast to mathematics and physics, biology is not what we in German term “exact science”, since we cannot prove anything except what we currently see (presence or absence). Any question which starts with “why” defies proofs. It may have one and only one correct answer, but in my opinion, we have no way to decide which of several possible answers is the only correct one. Accordingly, I never place much weight on a classification, be it mine or that of somebody else. I approach the problem from a more practical side: ‘Does the classification help me to get some understanding of the observed diversity and variation, and does it appear plausible.'”
His comment exposed for me the very roots of my discontent and unrest concerning the approach of the succulent enthusiast, and his community, to classification in general and of Haworthia in particular. Also stirring my curiosity about the way the “scientific community” seems to regard my publications on Haworthia. In fact Dr. Eggli is saying that in not knowing Haworthia himself, he is required to adopt a practical approach. My curiosity regarding this subject is always aroused in informal contact with biologists, with ecologists and with taxonomists, and also with writers who have reviewed and commented on my publications. It is also piqued by the responses I receive from editors in my communication with them about publications as well as about my own manuscripts. So I replied to Dr. Eggli as follows:
….”I would like your help in unravelling this issue of classification. Haworthia has been wracked with problems since von Poellnitz, Smith, Brown, and Uitewaal. It is not a different problem to that in other groups ‑ it is just that Haworthia is more visible in the way that it is collected and grown, and in the number of people who try to write about them? Plausibility has always been a problem and I have never understood my colleagues at the Compton, Bolus and PRE herbaria who have this common laissez faire attitude to classification. None of them could speak to me about the problem I had with Col. Scott’s classification. Even now, my classification is ignored by them because they cannot actually determine plausibility of my “opinions” against Scott’s. I have written extensively about the question of plausibility using other words for it, but no one seems to take the issue seriously. Reviews of my revision by botanists have been fatuous and all they can seem to really contribute is pseudo‑intellectual comments on the nomenclature and the requirements of the code. In my address to Succulenta 2000 (Congress held at Kirstenbosch) I pointed out the absence of a definition of the “species”. Actually it is a fundamental problem which comes down to Darwinism and science and elementals ‑ which is what my review (see later chapter) of Gould’s book (Rock of Ages, Jonathan Cape, 2001) is about.
“If you have seen my analysis of Prof. Vosa’s cytological work on Gasteria, you will see the style of botanical science which I experience regularly. Yet there is this same laissez faire attitude because the average botanist does not actually know what is being written about regarding subjects which are not in their specific field of work even although they may impact on what they write. What actually happens down at the lower ranks (where I labour), is that we have a huge group of uninformed readers who get the kind of leadership which botanists in forewords to Breuer’s two books World of Haworthias Vols 1 & 2 provide. Thus an environment is brewed which makes it extremely difficult to achieve credibility and to know what is plausible and what is not, and to whom. Breuer also gets credibility for a publication in Taxon which is a chronological mishap, where he has done typifications without really knowing what the names were for, or knowing what names are going to be needed. Nor has he persuaded me that he has really understood what has already been written.
“All this may have been manageable 40 years ago ‑ but we cannot go on with all these meaningless changes of name, or creation of new names, based on weak opinion and the absence of sound or reasonable peer review. Our botanical leadership is just non‑existent, and it frustrates me no end to find myself trying to play this role against the tide of people who should be doing it. To say you have not followed my review of Gould’s book is something of an apology, but you should know what Gould and Linde say in the (Scientific America Book of the Cosmos, MacMillan, 2000).
“Where I would like some help is on this issue of “how?” as opposed to “why?”. Newton surely asked ‘Why did that apple fall?’ ALL questions have answers and are thus valid when one seeks to know. Science seeks to know. Science does not belong to physicists or botanists. And what physicists know, botanists must know, even if it is only in general terms. How are we going to make progress unless botanists, and those posturing as botanists, as a whole think at the level of scientists in other disciplines? Botany and biology appear to me to be in a dark age of a new religion of “science”. This is what Linde is hinting at in “Book of the Cosmos”.
“My recent work on Haworthia confirms what I said long ago. I just put it now in other words. Species are fractal. Darwinism is a partial answer. Species vary and change “chaotically”, and there may well be (is, is my honest view) purpose and meaning. Botanists seem to happily go along with the old view that reductionism will produce answers, when in the hard sciences it has been fairly well proven that it will not. Where do biologists stand on this issue? They do not seem to want to know?”
That was my letter.
A point I now make, is that the question “how biological diversity could have arisen”, might be validly and better answered by the question …”why is it like this?” Another question is this about the philosophy of science itself…”Can any hypothesis be proven?” In statistics, the null-hypothsis is chosen on the basis that you can never prove a hypothesis but you can disprove it.
Without repeating any of my arguments I placed before Succulenta Congress 2000, I want to say that writers are claiming an understanding of taxa when in fact they do not actually know what the significance of any taxon may be. I wrote in Haworthia Revisited (1999) … “The basis of classification of Haworthia must be the geographical component.” The tendency in both the professional and academic ranks is still to recognise “species” on the flimsiest of morphological pretences, and to rank nomenclature wholly disproportionately to the function of identification and communication. This is never going to help to resolve a problem which is falsely seen to exist uniquely in Haworthia. This is a complete misconception.
The case of H. intermedia may help to clarify a few points that I have made.
In writing Haworthia Revisited, I adopted an even more concise style than usual. In the case of H. intermedia, I knowingly strayed, but I did not quite appreciate that my disrespect for the cosmetics of nomenclature would lead me to deliberately obstruct its objective. I in fact needed a name for an element I collected in KG106/70 (later recollected as MBB6514), from a site, Buitenstekloof between Robertson and Worcester.
What actually happened is this, von Poellnitz described H. intermedia in Kakteenkunde 1937 p.134, citing “Cape, Robertson, McGregor: Mr. G.J. Payne sn. (coll. Triebner 956)”. There was no accompanying illustration. He used the epithet “intermedia” because he felt that the plant (probably a single plant) was intermediate between H. integra (which, with all the confusion associated with that epithet, can perhaps safely be assigned to H. mucronata – i.e. safely for anyone with actual understanding of the uncertainty of Haworthia) and with H. haageana and its var. subreticulata (both v.Poelln.).
Let us just examine this. H. haageana, based on the photograph in Berlin (of which I saw a copy in 1969), chosen by Breuer as lectotype too, is safely ascribable to H. reticulata despite being said to be from Grahamstown. The variety subreticulata, also said to have come from Grahamstown, is arguably ascribable to H. reticulata judging from the available photograph. It could actually be something else.
When I met George Payne in 1970, I specifically asked him about H. intermedia and he informed me that he had collected it at Buitenstekloof. That is where I found H. reticulata (KG105/70) and which is why intermedia appears in my earlier publications as a synonym of H. reticulata (see Fig.1). This Fig.1 is an unpublished picture recently dredged out of the Berlin Herbarium as the type of H. intermedia, and I think it can be taken to confirm this first diagnosis. This is despite the comment by von Poellnitz that the leaves were “marked differently”! Some troubles appear when it is shown that von Poellnitz added a reference to a plant collected by Eric King at Scottburgh Farm (in Long474, Port Elizabeth, to his 1938 Feddes Repert. citation of H. intermedia (Fig.2, also an unpublished herbarium illustration). This plant could be one of those dubious things common in that area (particularly Hankey/Patensie/Humansdorp) which is neither H. cooperi(gracilis) var. picturata nor H. cooperi var. gordoniana. It is not possible for geographical reasons to suggest that the Buitenstekloof and the Port Elizabeth plants can be synonymous. When I came to write Haworthia Revisited, I was frankly just frustrated and irritated with the silly requirements of nomenclature and need for the frivolity of a latin synopsis. I had this second element from Buitenstekloof (and I am not talking about a single plant) which was not H. reticulata, and I doubted that Payne had ventured to where I found it (Fig.3 MBB6514). Unlike the reticulata there which is on the dolomite, this element is on shale. It may be neither H. maraisii var. notabilis (in that element’s many guises), and nor is it H. maculata which has its own problems. So I lifted the name intermedia and used it here in relating the plant to H. maculata. I really did not think this would ultimately give any sensible revisioner who really bothered to explore that mountaineous and geologically complex terrain for better answers, any serious problems. In fact my intention was that my treatment would demand a thorough exploration of the area. As it is, I know of three unconfirmed collections which need to be followed up if any more acceptable answer is going to be obtained. I really did not think that buffoonery of a greater order than my own was going to prevail, but so it has. Figs 1, 2 & 3 are from World of Haworthia, Vol.2 pp529-30, by I. Breuer. Here in juxtaposing the illustration of what could be three quite different species, Breuer is suggesting that these are all intermedia.
A plant illustrated by Breuer as H. maculata var. intermedia (Fig.3) is from my collection MBB6514, and it is correct for my use of the epithet. But it is not necessarily H. intermedia of von Poellnitz, and seeing Fig.1 for the first time I would have stayed with my conviction that this could have been H. reticulata. So, while explaining again that von Poellnitz was dithering between reticulata, mucronata and “gracilis”, that J.R. Brown was doing the same at Sectional level, that Breuer implies that they are all the same, and that Bayer has a different opinion; it is quite obvious that someone else now claiming understanding of this taxon, is making a very wild statement. To correct the situation and preclude a minor disaster, I correct the deliberate and regrettable misuse of an epithet as follows:
Haworthia maraisi var. notabilis (v.Poelln.) Bayer syn. Haworthia maculata var. intermedia Bayer in Haworthia Revisited 1999 p.91, non v.Poelln. in Kakteenkunde 137, 1937. An incorrect name under Art. 11.4 of the code, because of the principle of priority (with gratitude to Dr Urs Eggli).
H. reticulata Haw. syn. Haworthia intermedia v.Poelln. in Kakteenkunde 137, 1937. Lectotype: Breuer et al Cape, McGregor, Robertson, G.J. Payne (Triebn.956), B.
Thus my use of the epithet intermedia was for a population-based variant possibly of …
1. H. maculata, despite having a later flowering time. This is quite extensively discussed in Haworthia Revisited. H. maculata is recognised as it occurs in a series of closely spaced populations north of the Brandvlei Dam at Worcester, and as two populations approximately at Sandhills, north-east of Worcester.
2. H. maraisii var. notabilis, despite having a flower more typical of H. maculata. This variety is recognised in the population at Buitenstekloof discussed above, a population north of Agtervink west of Robertson, three closely spaced and variable populations at Wolfkloof, Robertson (the type locality for H. notabilis v.Poelln.) this variety, and three populations in the Klaasvoogds Valley east of Robertson (Figs.4 to 9 and map Fig.10). Note that the plants illustrated are individual clones only and do not convey the variability of the respective populations. The flowering times of the populations other than Buitenstekloof one (Nov.) is approximately Sept. but the Kranskop population had flowering specimens as early as May. H. maraisii mostly flowers early winter viz. late March to May.
I have now chosen the latter option because it does not require any nomenclatural manipulation. It should be quite evident by now that a species concept for Haworthia (actually any species) has to consider that the species is a complex system of individuals that vary with place and time. Should anyone be misguided enough to claim any knowledge of von Poellnitz’s H. intermedia, they must please go further than my attempt to do so either here or in my revision. They must also competently demonstrate that they have read and understood at least this explanation.
Fig.1. Triebner 956 H. intermedia V.Poelln. as illustrated in Breuer, World of Haworthia, Vol.2:529 = H. reticulata.Fig.2. Long 474 “H.intermedia” ex Scottburgh, Port Elizabeth (Breuer, World of Haworthia, Vol.2:530) = H. cooperi var. picturata/gordoniana.Fig.3. Breuer 1999(MBB6514) “H. maculata var. intermedia“, Buitenstekloof, Robertson = H. maraisii var. notabilis.Fig.4. JDV86/108 H. maraisii var. notabilis also Buitenstekloof.Fig.5a. JDV96/38 H. maraisii var. notabilis, Agtervink (East of Buitenstekloof).Fig. 5bFig. 5cFig.6a. MBB7097 H. maraisii var. notabilis, Kranskop, Klaasvoogds,Fig. 6b.Fig.7. MBB7098 H. maraisii var. notabilis, Bergplaas, Klaasvoogds (note the similarity to either H. reticulata or to H. turgida in this exposed and clumping state.Fig.8a. MBB7055 H. maraisii var. notabilis, Rietvlei, Klaasvoogds. A cryptic form.Fig. 8bFig.9. JDV97/49 H. mirabilis var. consanguinea, Dwarswaterkloof, SW McGregor. A “look-alike”!Fig.10. Map of the Robertson area showing:- 1- Buitenstekloof, 2- Agtervink, 3- Wolfkloof, 4- Bergplaas, 5- Kranskop, 6- Rietvlei, 7- Dwarswaterkloof far to SW.
References
Bayer, M.B. Haworthia Revisited, Umdaus 1999.
Bayer, M.B. Natural Variation and Species Delimitation in Haworthia Duv. Pt.1. H. reticulata Haw. Nat.C.S.Jl.27:10(1972).
Breuer, I. The World of Haworthia Vol.2, Breuer & Arb. Mamm. 2000.
Key to illustrations Fig.1. Triebner 956 H. intermedia V.Poelln. as illustrated in Breuer, World of Haworthia, Vol.2:529 = H. reticulata.
Fig.2. Long 474 “H.intermedia” ex Scottburgh, Port Elizabeth (Breuer, World of Haworthia, Vol.2:530) = H. cooperi var. picturata/gordoniana.
Fig.3. Breuer 1999(MBB6514) “H. maculata var. intermedia”, Buitenstekloof, Robertson = H. maraisii var. notabilis.
Fig.4. JDV86/108 H. maraisii var. notabilis also Buitenstekloof.
Fig.5a, b & c. JDV96/38 H. maraisii var. notabilis, Agtervink (East of Buitenstekloof).
Fig.6a & b. MBB7097 H. maraisii var. notabilis, Kranskop, Klaasvoogds.
Fig.7. MBB7098 H. maraisii var. notabilis, Bergplaas, Klaasvoogds (note the similarity to either H. reticulata or to H. turgida in this exposed and clumping state.
Fig.8a & b. MBB7055 H. maraisii var. notabilis, Rietvlei, Klaasvoogds. A cryptic form.
Fig.9. JDV97/49 H. mirabilis var. consanguinea, Dwarswaterkloof, SW McGregor. A “look-alike”!
Fig.10. Map of the Robertson area showing:- 1- Buitenstekloof, 2- Agtervink, 3- Wolfkloof, 4- Bergplaas, 5- Kranskop, 6- Rietvlei, 7- Dwarswaterkloof far to SW.
Many years of experience have left me very skeptical of botany and botanical science, and particularly of writers who masquerade as taxonomists. This is reflected in much of my writing and in these essays. One of the difficulties I have had is in trying to understand why botanists tend to regard species as quantum units which are distinguishable from one another unless some taxonomists has made some statement to suggest that they are not. Non-botanists are further beyond the pale. This brief essay is to demonstrate that I have not found vaunted sophistication and technology of much assistance in better understanding the relationship of Haworthia populations to one another. Often references to such technology are used as subterfuge by authors who should not be engaging themselves in classification at all.
Early in 1970 Dr David Cutler (Royal Botanic Gardens, Kew) was kind enough to have made many photographs of the leaf surfaces of a large number of Haworthia specimens representing a wide range of seemingly interconnecting populations. See the attached list for the plants examined. I ended up with about 350 of these photographs which I have been unable to explain. Looking at them as one would look at the plants themselves, I am again and again struck by the fact that in the end, single specimens were taken to represent populations. This is despite a specific request to have at least 10 leaves photographed from at least one population. The photographs were taken at various magnifications, powers and angles. Most significant would probably have been 100X, but only about 10 such pictures were taken. The most were at 300X magnification and this article is illustrated with these, excepting the final fig.24 which is 100X).
I could not, and can still not, find any pattern which helps me to resolve the relationships any better than explained in many of my publications or in my revision of 1999. No population can be described from a single plant, and in Haworthia no meaningful description can be written to exclude plants from other populations – such is the nature of the continuities that exist between populations. To expect that leaf surfaces (and flowers and seed too, as has so often been implied by my critics) will provide characters to resolve these continuities into recognisable discrete entities, is actually very far-fetched. Suggesting that it will, borders on the fraud. The most I would expect is that leaf surface morphology as depicted in an SEM photograph, may be very useful for distinguishing between, say, H. viscosa and H. pungens, where nothing is actually needed; but it is not going to resolve the problems I described, say, in H. rossouwii and the complexities of its relationships with as many as seven other elements. These elements can be seen to be discrete at some point in their distribution ranges, but not everywhere. I mention H. viscosa and H. pungens particularly because there is a writer who has already proclaimed the latter to be a re-description of H. beanii, despite the very obvious differences that are visible to any eye. They are not the same thing, and the latter is in fact a variant of H. viscosa. These inaccurate diagnoses and ridiculous statements are all too common and P. Halda made a similar error. He described a new variety of H. viscosa from the Baviaanskloof, not far from the localities for H. beanii and its var. minor (both G.G. Smith). However, he also went so far as to also reduce H. nigra to the rank of subspecies under H. viscosa. These are major issues which are difficult to refute in the absence of incontrovertible evidence, despite the fact that they are so easily resolved by reference to the field situation and gross visual examination. One can separate H. viscosa (H. beanii) and H. pungens in the dark simply by feel.
In the subgenus Haworthia, the problems are wholly of a different magnitude. The various species cannot be differentiated by any method and never will be – until the resolution comes to pass of its own in geological time.
Looking at some of the SEM photographs:
Fig.1 is KG226/70 H. maculata from Moddergat, south of Worcester. In terms of the photographs I have, it is very comparable with fig.2 KG48/70, which is H. maraisii from Stormsvlei, and it may also be comparable with fig.3 KG20/70 which is also H. maculata but from the Brandvlei Dam south of Worcester. But then I am confronted with four other photographs for this species, viz. figs.4 and 5 also Moddergat, and figs.6 and 7 both Brandvlei. One can easily agree on some or other common feature and perhaps decide that we are dealing with one species. This is not the case.
We can take fig.8, which is also KG20/70 H. maculata from Brandvlei, and juxtapose it with fig.9 KG163/71 H. emelyae var. multifolia from Springfontein in the Little Karoo – very comparable. Figs.10 KG178/70 H. reticulata and Fig.11 KG 179/70 H. notabilis are both from Wolfkloof north of Robertson. They are barely distinguishable and especially so in the context of the degree of difference between photographs of the same specimens at different power and angle.
Figs.12 and 13 KG36/70 are H. heidelbergensis from Rooivlei north of Bredasdorp. This is a problem population because it may have (it does have) ties with fig.14 KG318/71 H. rossouwii (was H. serrata), Oudekraal, Heidelberg, and with fig.15 KG709/60 H. mirabilis var. sublineata, south of Bredasdorp. The latter must be compared with fig.16 which is the latter element again, but from north of Bredasdorp. Two other figures are fig.17 and fig.18, KG81/71 H. heidelbergensis, from Matjestoon south of Heidelberg and KG210/71 H. heidelbergensis from just east of Heidelberg. There is very little evidence of discrete pattern.
If one attempts to formulate pattern from the above figures, then look at fig.19 KG134/70 H. mirabilis var. triebneriana (the forma diversifolia) from east of McGregor, or fig.20 KG213/70 H. maraisii from the sandstones south of McGregor. Then I include fig.21 KG35/70 H. maraisii from north of Bredasdorp and compare with fig.22 which is H. mucronata var. inconfluens from west of Ladismith.
I emphasise that these are just single samples from an extremely limited range of populations and I cannot imagine that anyone can afford the time and the expense of analysis of enough material, to arrive at any definite and better conclusion. It is a simple phantasm to suggest that technology is going to resolve a conceptual problem. Where aspiring writers are unable on macromorphological grounds to resolve issues as obvious as that of H. pungens and H. viscosa or H. viscosa and H. nigra, technology and sophistication are potential weapons of folly.
From all the pictures, it emerges that H. maraisii and H. mirabilis may have a commonality in the raised nipple at the centre of the surface cells. However this is absent in fig.23 KG400/61 H. maraisii (a variant – naturally) from Dublin west of Robertson, while it is present in H. arachnoidea and specifically fig.22. Fig.23 represents quite a major departure from most of the photographs and it will attempt someone to suggest this must be a different species. This will be ridiculous. My species concept and definition is that geographical distribution is the major criterion for the determination of the species. Thus the plant from which fig.23 is derived is firstly seen as from a single population which is just a geographical part of an extended complex. Without any strain on logic or reason, its characteristics can be taken to extend from, and also contribute to, that complex. Thus if leaf surfaces are to be studied in pursuing understanding of the relationships in Haworthia, a good approach would be to first take such a single complex and take an overview. The photographs suggest that 100X would have been a good magnification at which to do this (see fig.24 H. notabilis and compare with the same in fig.11).
What needs to be done is for my revision and explanation of Haworthia to be taken as a hypothesis and tested as such. Even an article of this kind requires this basic hypothesis. The presentation of alternate hypotheses and nomenclatural solutions based on less material, inadequate conceptualization and poor intellectual preparedness is a disaster for the collector and enthusiast. It is also a disaster for the botanist and scientist who fail to understand, as so many do, that the authority of a classification is not just a question of acceptance or observance of a nomenclatural code and the product of anyone who is enthusiastic enough to produce an identification key and a classification. There is adequatio involved in the case of both the taxonomist and the acceptance of the product.
Fig. 1. MBB in KG226-70 H. maculata. Moddergat, S Worcester.Fig. 2. MBB in KG48-70 H. maraisiivar maraisii. Stormsvlei.Fig. 3. MBB in KG20-70H. maculata. Brandvlei.Fig. 4. MBB in KG226-70 H. maculata. Moddergat.Fig. 5. MBB in KG226-70 H. maculata. Moddergat.Fig. 6. MBB in KG167-70 H. maculata. Brandvlei.Fig. 7. MBB in KG 167-70 H.maculata. Brandvlei.Fig. 8. MBB in KG20-70 H. maculata. Brandvlei.Fig. 9. MBB in KG163-71 H. emelyae var multifolia. Springfontein.Fig. 10. MBB in KG178-70 H. reticulata. Wolfkloof, Robertson.Fig. 11. MBB in KG179-70 H. maraisii var notabilis. Wolfkloof, Robertson.Fig. 12. MBB in KG36-70 H. heidelbergensis var minor. Rooivlei, Bredasdorp.Fig. 13. MBB in KG36-70 H. heidelbergensis, Rooivlei, north of Bredasdorp.Fig. 14. MBB in KG318-71 H. rossouwii var rossouwii. Oudekraal.Fig. 15. MBB in KG709-60 H.mirabilis var sublineata. S Bredasdorp.Fig. 16. MBB in KG311-71 H. mirabilis va sublineata. N. Bredasdorp.Fig. 17. MBB in KG81-71 H. heidelbergensis var toonensis. Matjestoon.Fig. 18. MBB in KG210-71 H. heidelbergensis vr heidelbergensis. E Heidelberg.Fig. 19. MBB in KG134-70 H. mirabilis var triebneriana. Olifantsdoorn, SE Macgregor.Fig. 20. MBB in KG213-70 H. maraisii var maraisii. S Macgregor.Fig. 21. MBB in KG35-70 H. maraisii var maraisii. Rooivlei, N Bredasdorp.Fig. 22. MBB in KG 287-72 H. arachnoidea va nigricans. S Calitzdorp.Fig. 23. MBB in KG400-61 H. maraisii var maraisii. Dublin, Worcester.Fig. 24. MBB in KG179-70 H. maraisii var notabilis. Wolfkloof, Robertson.
Figure captions Fig.1 KG226/70 H. maculata, Moddergat, south of Worcester. Fig.2 KG48/70 H. maraisii, Stormvlei, south of Bonnievale. Fig.3 KG20/70 H. maculata, Brandvlei Dam, south of Worcester Fig.4 KG226/70 H. maculata, Moddergat, south of Worcester. Fig.5 KG226/70 H. maculata, Moddergat, south of Worcester. Fig.6 KG167/70 H. maculata, Brandvlei Dam, south of Worcester. Fig.7 KG167/70 H. maculata, Brandvlei Dam, south of Worcester. Fig.8 KG20/70 H. maculata, Brandvlei Dam, south of Worcester. Fig.9 KG163/71 H. emelyae var. multifolia, Springfontein, Little Karoo. Fig.10 KG178/70 H. reticulata, Wolfkloof, north of Robertson. Fig.11 KG179/70 H. notabilis, Wolfkloof, north of Robertson. Fig.12 KG36/71 H. heidelbegensis, Rooivlei, north of Bredasdorp. Fig.13 KG36/71 H. heidelbegensis, Rooivlei, north of Bredasdorp. Fig.14 KG318/71 H. rossouwii, Oudekraal, south-west of Heidelberg. Fig.15 KG709/60 H. mirabilis var. sublineata, south of Bredasdorp. Fig.16 KG311/71 H. mirabilis var. sublineata, north of Bredasdorp. Fig.17 KG81/71 H. heidelbergensis, Matjestoon, south of Heidelberg. Fig.18 KG210/71 H. heidelbergensis, east of Heidelberg. Fig.19 KG134/70 H. mirabilis var. triebneriana (the forma diversifolia) from east of McGregor. Fig.20 KG213/70 H. maraisii (var. whitesloaneana), south of McGregor. Fig.21 KG35/70 H. maraisii, north of Bredasdorp. Fig.22 KG287/72 H. arachnoidea var. nigricans from south of Calitzdorp. Fig.23 KG400/61 H. maraisii, Dublin west of Robertson. Fig.24 KG179/70 H. notabilis, Wolfkloof, Robertson.
Appendix List of specimens submitted and scanned (bracketed numbers are the laboratory codes):- 520/60 (40) H. maraisii, Cogmanskloof. 709/60 (54) H. mirabilis var. sublineata, S. Bredasdorp. 400/61 (31) H. maraisii, Dublin. 624/69 (51) H. mirabilis var. mirabilis, Mierkraal. 625/69 (58) H. mirabilis var. badia, Napier. 629/69 (47) H. maraisii, W. Swellendam. 638/69 (39) H. maraisii var. notabilis, Klaasvoogds. 661/69 (61) H. maraisii var. notabilis, Wolfkloof, Robertson. 679/69 (62) H. mirabilis var. badia, Napier. 681/69 (59) H. mirabilis, N. Napier. 688/69 (32) H. maraisii, Oliva, Robertson. 689/69 (30) H. maraisii, SW Robertson. 692/69 (53) H. mirabilis var. triebneriana, W. Genadendal. 20/70 (26) H. maculata, Brandvlei West. 26/70 (55) H. mirabilis, Stormsvlei Pass. 2?/70 (64) H. mirabilis, S Greyton. 30/70 (60) H. mirabilis, SE. Greyton. 31/70 (63) H. mirabilis, S. Greyton. 32/70 (57) H. mirabilis var. beukmannii, Skuitsberg. 35/70 (20) H. maraisii, Rooivlei, Bredasdorp. 36/70 (6) H. heidelbergensis var. minor, Rooivlei, N.Bredasdorp. 48/70 (18) H. maraisii, Stormsvlei. 106/70 (45) H. maraisii var. notabilis, Buitenstekloof. 112/70 (13) H. pubescens, Sandberg, Worcester. 134/70 (28) H. mirabilis, Olifantsdoornkloof. 167/70 (24) H. maculata, Brandvlei. 178/70 (42) H. reticulata, Wolfkloof, Robertson. 179/70 (43) H. maraisii var. notabilis, Wolfkloof. 180/70 (44) H. maraisii var. notabilis, Wolfkloof. 202/70 (7) H. magnifica var. atrofusca, Spitzkop, Heidelberg. 212/70 (27) H. maraisii, SW McGregor. 213/70 (29) H. maraisii, Houtbaaikloof. 213/70 (29b)H. maraisii, S McGregor. 223/70 (33) H. maraisii var. meiringii, W Bonnievale. 224/70 (34) H. maraisii var. meiringii, E Bonnievale. 226/70 (25) H. maculata, Moddergat. 402/70 (2) H. springbokvlakensis, Springbokvlakte. 2/71 (35) H. maraisii var. meiringii, W Bonnievale. 9/71 (37) H. maraisii var. meiringii E Bonnievale. 13/71 (38) H. maraisii var. meiringii, Goudmyn. 80/71 (11) H. maraisii, Napky. 81/71 (4) H. heidelbergensis var. toonensis, Matjestoon. 83/71 (16) H. magnifica, Riversdale Nature reserve. 84/71 (17) H. mirabilis var. paradoxa, Vermaaklikheid. 118/71 (19) H. emelyae var. major, Muiskraal. 163/71 (1) H. emelyae var. multifolia, Springfontein. 170/71 (41) H.maraisii, N. Ashton. 210/71 (5) H. heidelbergensis, NE Heidelberg. 310/71 (52) H. mirabilis var. triebneriana, Skietpad. 311/71 (56) H. mirabilis var. sublineata, N. Bredasdorp. 318/71 (3) H. rossouwii, Oudekraalkop. 326/71 (12) H. mirabilis/maraisii, Rondeheuwel. 335/71 (8) H. emelyae, Rooiberg. 344/71 (46) H. maraisii, Langvlei. 481/71 (10) H. mirabilis var. triebneriana, Leeurivier (ex van Breda). 94/72 (21) H. heidelbergensis, Leeurivier. 111/72 (14) H. bayeri, S Oudsthoorn. 114/72 (15) H. emelyae var. comptoniana, Georgida. 125/72 (49) H. arachnoidea var. nigricans, Gamka Rd. 125/72 (50) H. arachnoidea var. nigricans, Gamka Rd. 138/72 (36) H. emelyae, Sandkraal. 287/72 (48) H. mucronata, W. Ladismith. 107/74 (22) H. magnifica, Tradouw Pass. MBB1128 (23) H. pubescens var. livida, Lemoenpoort. ‑ ‑ (9) H. mirabilis var. mirabilis, Mierkraal.