Printed in British Cactus and Succulent Journal 5:13 (1987)
Part 6. HAWORTHIA MACULATA (v. Poelln.) Bayer.
M. B. Bayer, National Botanic Gardens of South Africa, Karoo Gardens, Worcester.
Haworthia maculata (v. Poelln.) Bayer. Haworthia Handbook: 130 (1976); New Haworthia Handbook: 43 (1982); J. W. Pilbeam, Haworthia and Astroloba: 89 (1983). H. schuldtiana var. maculata v. Poelin. in Feddes Repert. Spec. Nov. 49: 25 (1940). Type: Cape; in the neighbourhood of Worcester, Swellendam, Bredasdorp and Caledon. Major H. Venter No. 6a. No specimen preserved. Neotype: Cape- 3319 (Worcester): Worcester District, H. Venter No. 6a in G. G. Smith 3912 (NBG)
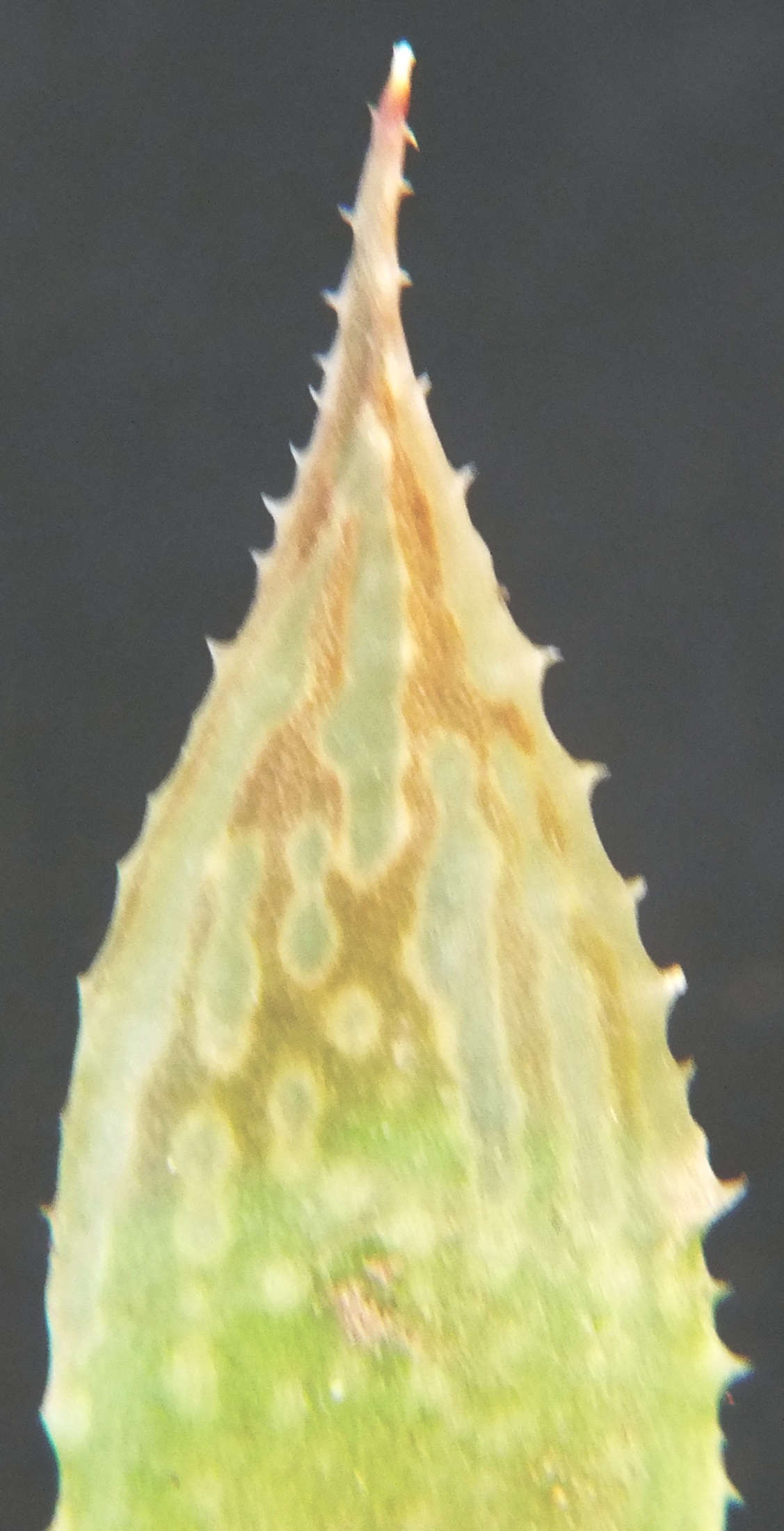
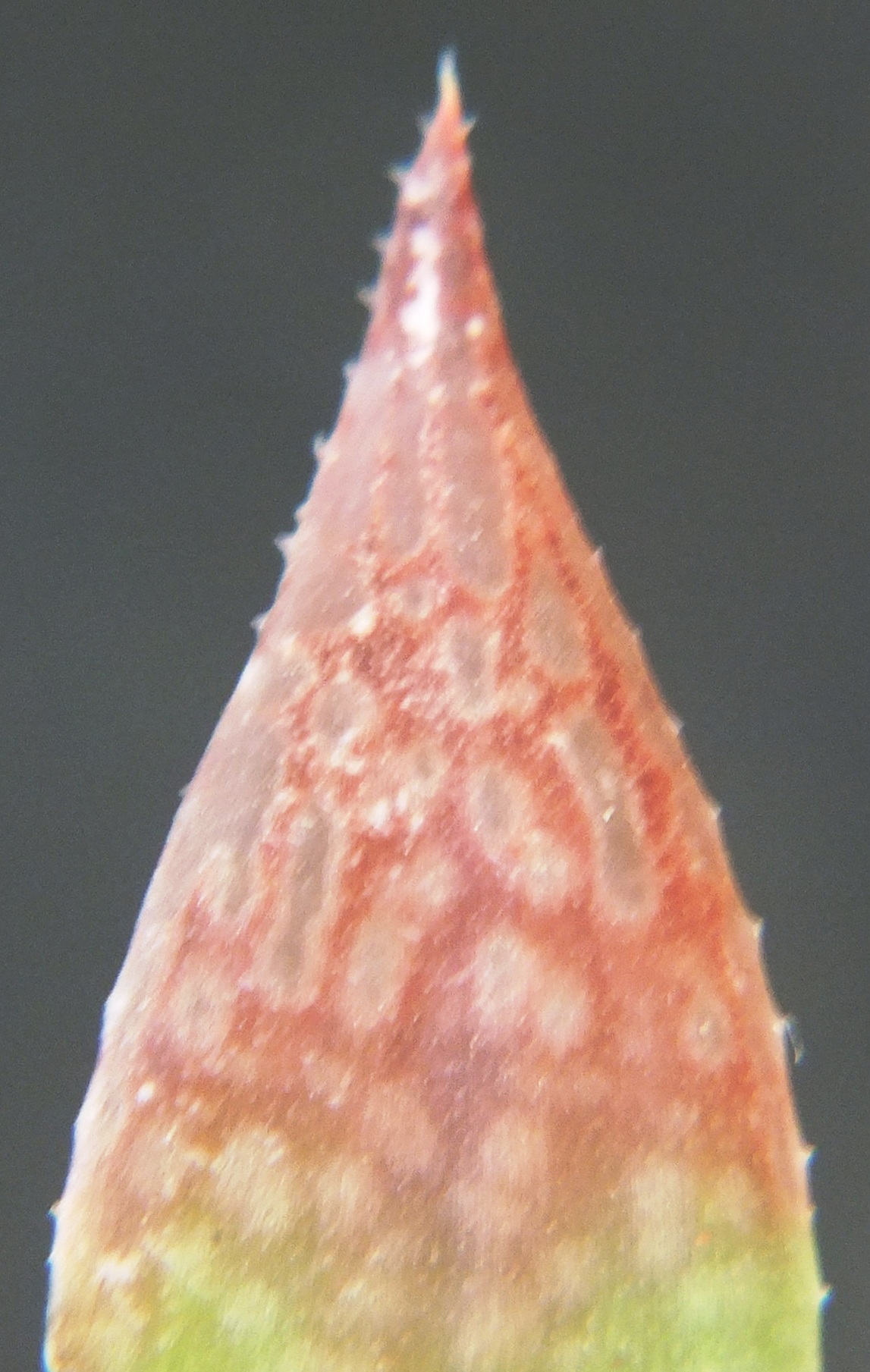
Rosette stemless, 30-70mm in diameter, up to 60 leaves. Stem thick, white-fleshed, non-fibrous, 1/3 diameter of rosette, never elongate, slowly proliferous from base. Roots thick, white-fleshed, non-fibrous. Leaves erect, spreading, slightly incurved at tips, up to 60mm long, 12mm broad, 8mm thick, ovate-lanceolate acuminate, aristate with bristle up to 5mm long, seldom setiferous, green to grey-green, purplish- green in sun, lines distinct with pronounced small longitudinal transparent areas on upper surface, lower surface heavily mottled or with longitudinal markings, spots occasionally with small hairs, face slightly concave at base, convex-turgid towards upper 1/4, back convex, frequently with second keel or with double row of spines on keel, margins sub-acute, lightly spined, spines white, up to 0.5mm long to 1mm apart. Perianth white, yellowish-green inside, nerves pinkish-brown outside, pinkish inside at tips, tube ascending curved, up to 16mm long, obclavate, bluntly triangular at base, 4.5mm across reducing to 3mm, segments free, regularly stellate, inner lower segments incurved along upper margin. Buds biarcuate, bifid at tips. Flowering Oct-Dec.
17. Haworthia maculata (V.Poelln.) Bayer :130(1976). Bayer :43(1982). H. schuldtiana var. maculata V.Poelln., Feddes Repert.Spec.Nov.49:25(1940). Type: Cape, Worcester, Swellendam etc. H. Venter 6a. Not preserved. Lectotype (B&M): Worcester, Venter 6 (BOL).
maculata: spotted.
Rosette stemless, proliferous, to 8cm φ. Leaves many, sub-erect to spreading, purplish-green, spotted, short spines on margins and keel. Inflorescence simple, slender. Flowers 15-20, few open, white, yellowish in throat, green veined.
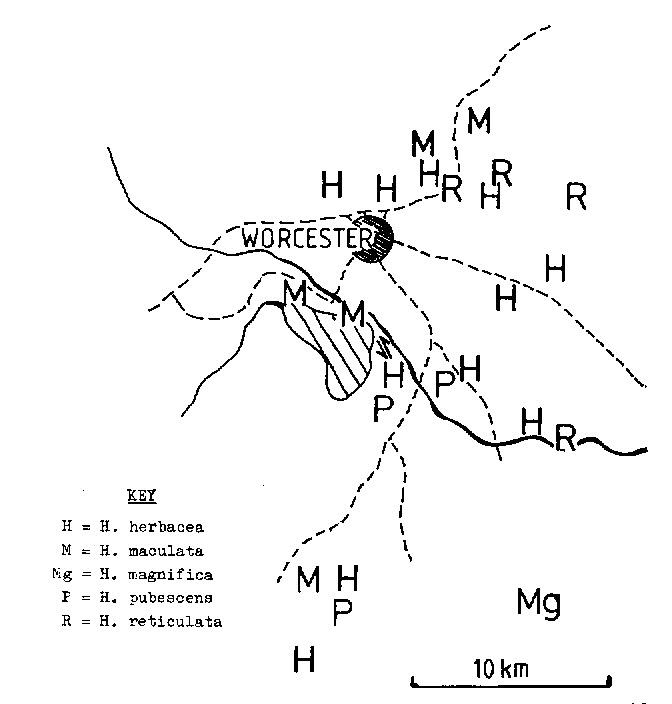
1982 – The original locality for this species was not accurately recorded but it was collected by Major H. Venter, and so it is possible from Smith’s and Long’s records to trace it to the Brandvlei Dam south of Worcester. H. maculata flowers in October/November and the form of the flower is like that of H. herbacea and H. reticulata, rather than like that of H. magnifica (H. schuldtiana). There is a clear intergradation with H. herbacea in the area, and the plants are very similar to H. herbacea except that there are fewer and more turgid leaves. H. maculata also occurs further south in a similar quartzitic rock formation to that at Brandvlei Dam. A problem is the occurrence of populations apparently of H. maculata in the mountains to the north of Worcester (both high altitude ‑ Audenberg Peak, and low altitude ‑ at Sandhills). A similar element occurs eastward towards Robertson at Buitenstekloof, distinguished again from H. magnifica by an earlier flowering time and the wide spread of the tips of the upper perianth lobes.
1999 – It is difficult to know just what is present on the higher mountains. Few succulentophiles are also mountaineers, and besides the plants could be expected to be on rocky north faces which may not attract the conventional high altitude botanist. H. nortieri bears some resemblance to H. maculata and that species is found as far south as Opdieberg (Ceres). It is quite probable that populations may occur elsewhere in the area between Worcester and Citrusdal.
a. var. maculata This variety seems to be linked to H. herbacea with possible ties to H. nortieri. The similarity to other high mountain forms (eg H. vlokii and H. turgida) cannot be overlooked and this has been repeated at several places in this book.
Distribution: 3319 (Worcester): Brandvlei Dam (‑CB), Bayer 164 (NBG), Smith 3912 (NBG); Bayer in KG669/69 (NBG); NE. Brandvlei Dam (-CB), Bayer 2591 (NBG); Audensberg Peak (-CB), Esterhuysen 16706 (BOL), Bayer 1119 (NBG); Moddergat (-CD), Bayer 1145 (NBG); Keeromsberg, Boskloof (-DA), BOL28719; S. Sandhills (-DA), Bayer 1120 (NBG).
Haworthia maculata var. maculata JDV88/29 east of Brandvlei. Growing in quartzitic stones, the plants become very cryptic
Haworthia maculata var. maculata JDV86/46 Brandvlei. The plants usually have a deep purplish hue and are very cryptic.
Haworthia maculata var. maculata JDV86/47 Brandvlei. Each localized population is a little different from its neighbours.
Haworthia maculata var. maculata JDV88/29 east of Brandvlei. At the eastern end of the range the plants form clusters and turn quite bright red during the dry hot summer.
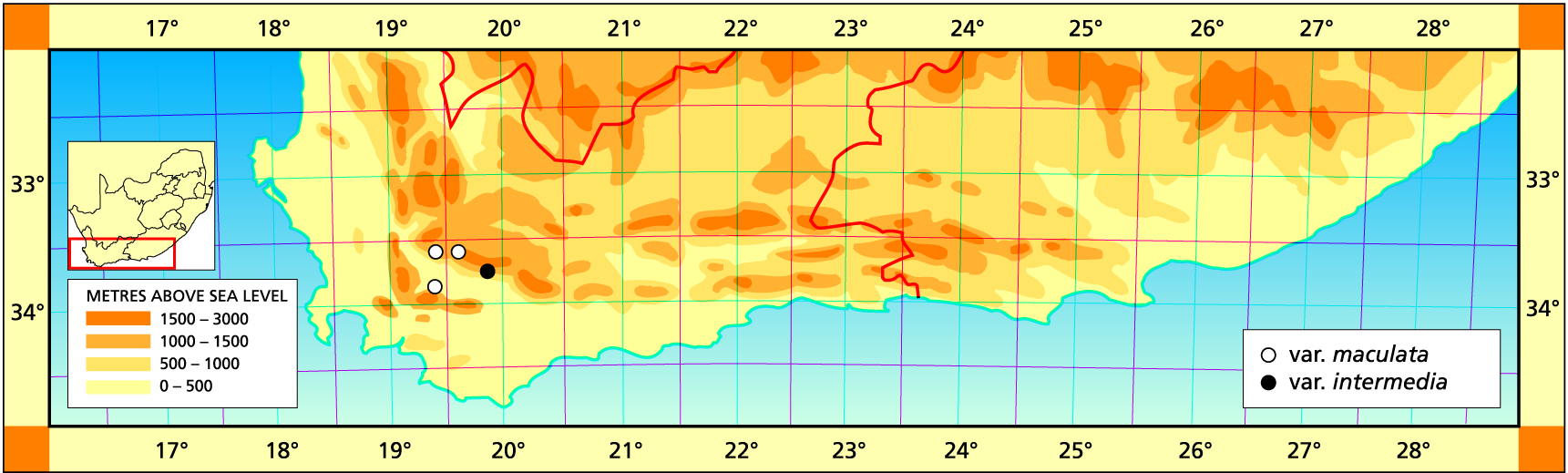
b. var. intermedia (V.Poelln.) Bayer stat.nov. H. intermedia V.Poelln., Kakteenkunde 9:133(1937). V.Poell., Feddes Repert.Spec.Nov. 44:233(1938). Type: Cape, Robertson, McGregor, G.J. Payne Not preserved. Lectotype (designated here): Epitype (designated here): CAPE-3319 (Worcester): Buitenstekloof (-DC), Bayer 4461 (NBG).
intermedia: between.
In the case of this variety, Payne (priv. comm.) did indicate the actual origin at Buitenstekloof west of Robertson. Von Poellnitz’ later citation for Scottburgh, Port Elizabeth, as well, is indicative of the close resemblance of even very different species and the difficulties which arise in trying to identify them consistently and correctly. In his discussion von Poellnitz concluded that while the plants had the long end-awn of H. mucronata, the reticulated patterning of the leaves was that of the H. reticulata group. The name suggests the difficulty in deciding just what to do with this element. It co-occurs with H. reticulata and with H. arachnoidea and bears a very close resemblance to the shale form of H. maraisii var. notabilis. As already noted it has a different flower and flowering time to that variety. Nevertheless it may be correct to place them together in one species as there is also a population recorded mid-way between the two at Agtervink. Possibly a more direct link with H. turgida should be sought as the plants do bear a close resemblance to the montane forms of that species. Certainly it is possible that there may be a connection somewhere in the mountains between Robertson and Swellendam.
Haworthia maculata var. intermedia JDV86/108 Buitenstekloof. The plants grow amongst Restionaceae. There is a close connection with H. maraisii var. notabilis.
Haworthia maculata var. intermedia JDV86/108 Buitenstekloof. The leaves are softer, more slender and upright than in the typical variety.
It is always assumed that botanists have a good grip of their subject, as one supposes for the scientists in other disciplines. Any science is the advance of knowledge by observation, hypothesis and testing by systematic enumeration and experiment. This is furthered by replication and review by other scientists. Botany is far behind the exact sciences, because of the very nature and complexity of living things and systems, and also behind because zoology where animal life is obviously more organized than is the case with plants and where more attention is focused.
1. Looking down the north face Die Nekkies east to west2. Same spot looking east northeast
It has always been recognized that South Africa has a unique flora and traditionally the difference from other floras was attributable to the extraordinary richness, diversity and endemism of the so-called Fynbos that came to be regarded as the Cape Flora. Just what the Cape Flora comprised was extraordinarily vague and the early limits were stated in terms of latitude and longitude. In the mid-20century it was recognized that South Africa should be divided into biomes, as major components of the vegetation, and these were defined as units with wide and expansive boundaries. So while the Fynbos biome was recognized among others, no connection seemed to exist between its boundaries and those of the major geological systems. The essential problem is that the boundaries of this biome are by no means wide and expansive, because one can literally step from Fynbos vegetation into Renosterveld, Sandveld and even Karoo. This is no more dramatically demonstrated than at Die Nekkies where it is obvious that the vegetation differences are primarily a result of geological substrate fortuitously linked to rainfall gradients determined by continental land-form, latitude and topography.
3. Same as figure 1, but from further east4. Still further east looking west northwest5. Looking north with M marking the location of Haworthia maculata, K – Karoo Garden6. Near east end with P marking the site of Haworthia pubescens, R indicating Rooiberg to the east7. View south to Villiersdorp
The significance of Die Nekkies is that it is formed from a narrow band of the Witteberg rock formation distorted in two transverse planes of the folded Cape Sandstone systems. Die Nekkies hills are directionally east to west and tilted down from the south. Northwards they are lost under the flood plain of the Breede River. Eastwards and southwards there is a wedge of Witteberg and Karoo formations that drives against the Cape Sandstones all the way south west to Villiersdorp. The consequence is a dramatic vegetation barrier related to the very different soil chemistry of Witteberg quartzites as opposed to Cape Sandstone, which is as dramatic where Bokkeveld Shale and Dwyka Tillite meet.
What is in effect the case, and apparently not really conceded by botanists, is that there are primarily two floras in a single winter rainfall biome. Some botanists insist that vegetation differences are simply fire driven. Here it is again difficult to separate the consequences of different fuel loads due to the substrate characteristics and the plants adapted to each substrate from the effects purely due to the fire, but it is nevertheless quite obvious that the major differences are substrate.
This is all a very broad view of a complex phenomenon that emerges when one begins to look at the individual species components of the vegetation. Vegetation classification is really based on dominant species and seldom has any attempt been made to understand species distributions that do not coincide with broad general pattern. When this has been attempt-ed it has been very seriously flawed by the problem of identification. This is a matter that botanists seriously underestimate and consistently fail to address.
It is simply assumed by general botanists that taxonomists in herbaria are competent and able, and that revisionary work and primary taxonomy have kept pace with technological progress and research in all other fields. This is not the case. Taxonomy has for many years been a forgotten science and less and less resources are allocated to either revisionary research or identification. It is simply not possible to collect, mount, store and record the variation of even a fragment of the species flora within any herbarium structure. A related fact is that there is no definition of what a species actually is. Very often it is just assumed that a definition is in place that is satisfied when some, usually morphological, criterion suggests that a specimen and sometimes more than one specimen does not agree with some ill-defined norm dependent on the taxonomist and the herbarium in which he/she is stationed.
It is self-evident that in the nature of evolutionary processes that are presumed to take place, that variation must be intrinsic to any species. This means that species need to be considered as systems that respond and change according to events that also change substrate and climate. It also means that species systems cannot be the same and a model (definition) needs to be in place that recognizes that each species (system) may be in an individual relation to time and space.
Two examples from Haworthia can be given for the significance of Die Nekkies in terms of species diversity and distribution and the difficulties that poor taxonomy introduces to obscure pattern. The first is for Haworthia maculata. Where there is already considerable doubt as to a proper classification of the Alooideae in the Asphodelaceae, Haworthia is persistently seen to be a single natural group. While there is no definition in place for the species category, the genus category is assumed to have some reality. It in fact does not. It is quite evident that some species do have very close affinities that would satisfy the perception that these form good genera, there are many species that elude such simple categorization. In Haworthia, as it is traditionally constituted, there are three distinct sets of species based on a range of characters both vegetative and floral. It is an extraordinary reflection on botanical classification that this has never been properly accepted. The genus Aloe is widely distributed in South Africa and more than 150 species are recognized. However, only two species can be seen as strictly Cape Sandstone and hence Fynbos “biome” species, although several are closely associated. There is only one species of Haworthia subgenus Haworthia that can truly be said to be a Fynbos species and that is H. nortieri. This subgenus is poorly represented north and west of the Fynbos “biome”, but it is represented by a vast array of variants in the Eastern, Southern Cape and Little Karoo that are difficult to aggregate into species. The variants seem to be ecotypes and it is such ecotypes that do adapt to sandstone habitats. There are none such variants in the sandstone mountains south and west of Die Nekkies where only one “species” viz. H. maculata occurs in localized population at several points along the range. This “species’ is very closely related to H. herbacea that is very common on all the other skeletal soils to the east, both north and south, but very rarely penetrating sandstone habitats. Where H. maculata occurs on the eastern limits of Die Nekkies, there is a subtle change of rock and shallow soil to a more Karoid habitat, and the plants tend to be more clump forming and spinose, suggesting a shift to H. herbacea. This similarity shift is replicated in H. herbacea where it occurs east of the brickworks near the Brandvlei Dam wall. Ecotypes similar to, and classified as, H. maculata occur on the Witteberg formation hills southwards to Villiersdorp, but never in close association with H. herbacea. Curiously, a very similar variant to H. maculata occurs again high in the sandstones north of Worcester on the Audensberg peaks and also lower down in the Sandhill area. Thus H. maculata is a significant marker in the relation of the Asphodelaceae and Haworthia to the distinction of “Succulent biome” and “Fynbos” biome.
8. MBB 7526 Haworthia maculata Die Nekkies, Brandvlei Dam9. MBB 7526 Haworthia maculata Die Nekkies, Brandvlei Dam
Haworthia subgenus Hexangulares is represented by only one species in the south western Cape. This is Haworthia venosa that occurs only in rock faces flanking the lower Breede River and Gouritz River. Its related species H. tessellata is a Karoid element. Haworthia subgenus Robustipedunculares is tightly allied to a winter rainfall “biome”, but consists of only four species. Three of them are endemic to the Southern Cape and three of these do not extend further west than Ashton and Bonnievale. H. pumila is abundant in the Worcester/Robertson Karoo and extends as far afield as Laingsburg, Anysberg and Barrydale, but south of the Langeberg it is not nearer to Swellendam than at Drew Station. South of the Riviersonderend it is only at Stormsvlei, near Protem and at Vaandrigsdrift. It does not occur on the Witteberg formations south of Die Nekkies, but it does occur on the eastern lower slopes of Die Nekkies itself. So here again Die Nekkies is the evident limit of this species too – the second example.
Possibly the very dramatic case of Drosanthemum halli and D. bellum more clearly suggests the significance of local diversifying factors. D. halli is yellow flowered and abundant whereas D. bellum is pink and localized. They do not co-occur. It can be dramatically shown that D. hallii is nothing but a geographic variant of the bi-coloured D. micans that occurs north and west of Worcester, and that D. bellum can only be different because of very site specific factors.
H. maculata at the western end of Die Nekkies appears to be a solitary plant but, as it occurs more eastwards, it becomes proliferously clumping as the vegetation also becomes increasingly Karoid. Why this should be so is not clear.
The many accompanying illustrations (MBB7526 figs. 8-38) show the scenery and topography and the range of variation in H. maculata. One can see that it is at home in both the Restiods of the Fynbos and under Pteronia paniculata of the Karoid.
All the following photos are MBB 7526 Haworthia maculata. Die Nekkies, west end.
Acknowledgement I would like to thank Anso Le Roux of Pokkraal, Rawsonville for drawing my attention to H. maculata much further west along Die Nekkies than previously recorded and for inspiring a walk along the full eastern limb. ♦
(Haworthiamaculata var. livida (Bayer) Bayer, comb.nov. H. pubescens var. livida Bayer in Haworthia Revisited, p.134, 1999, Umdaus) Type: Cape-3319 (Worcester): S Lemoenpoort (-CD), Bayer 1128 (NBG, Holo.).
I described Haworthiapubescens var. livida in Haworthia Revisited (Umdaus, 1999), in the full knowledge that it was in a twilight zone of inadequate information. It is a good example of how Latin names give plants a false reality. The system forces decision making without any slack being cut for doubt. This is thus a good opportunity to demonstrate what inexperience and ignorance add to the process of classification. In the small area along the Breede River north of the Brandvlei Dam near Worcester, the species H. herbacea, H. maculata and H. pubescens grow in close proximity. H. herbacea is ubiquitous throughout the Worcester/Robertson Karoo, while H. maculata has a curious distribution in that area. It occurs at widely separated localities on the western fringe of H. herbacea and I have wondered about its relationship to that species because of the similar flowers and flowering time. H. pubescens is only known from a small set of low ridges east of the Brandvlei Dam where it grows in close proximity to H. herbacea. It also has similar flowers but it flowers a little later in late spring as opposed to early spring.
When I first discovered plants at Lemoenpoort (Hammansberg) about 20km further south I found very few plants and the only thing I could do with them was regard them as in the same context of the Breede River species and hence their description as a variant of that H. pubescens.H. herbacea is very abundant in this area too but almost exclusively in Bokkeveld Shales and Dwyka Tillite. The doubtful plants at Lemoenpoort occur in Witteberg Sandstones. A population of plants was also known from the top of the Moddergat Mountain (Ouhoekberg) to the west that created some problems for me as it seemed to relate to the absence of H. mirabilis from both the Hammansberg and Ouhoekberg. Not only that but it also looked very similar to H. maculata. H. mirabilis is known at its closest east of these two mountains at several localities along the eastern ridges of the Ouhangsberg and Droogerivierberg mountains that are in turn east of Hammansberg. H. maculata is very abundant along the long ridge of low hills north of the Brandvlei Dam. Very similar plants occur high in the Audensberg peaks north of Worcester and also lower down in the Kanetvlei (Sandhills) area west of Heatlie Peak. These hills are very difficult to explore and I speculate a connection to a population at Buitenstekloof to the east. There is a population here that obfuscates the interpretation of the name H. intermedia VPoelln. I lumped this population under H. mirabilis var. notabilis. I have to rely on memory here because having set flower morphology and flowering time aside (without completely ignoring them), I did not have the methodology in place to record detail. The flower was rather similar to H. herbacea and H. maculata and it also flowered late spring. I was not, and am not, sure that there is not a connection between the Buitenstekloof plants and the Kanetvlei plants.
It is unfortunate that Gerhard Marx has expressed his opinion that I have ignored flowers in my classification of Haworthia. While there is no doubt that this is partially true, there are very good reasons why I did this. The main reason is the historic one. It was understood and accepted that the flowers in the species as they were then recognized, were very similar. Even the subgeneric differences were not seen to be significant. My own early attempts at classification also indicated that the flowers were not going to be of assistance at the level at which differences between populations and species were being recognized. In fact I had observed similarities of the flower in populations that were clearly different taxonomic entities. Conversely I observed big floral differences in populations that I imputed to be the same “species”. Because of the added seasonality of flowers, I thus put them aside to focus on exploration and explanation on the basis of the vegetative structures and geographic distribution. Marx’ observation therefore comes at a time when that work is substantial and flowers can now be used to clarify and verify relationships. My conviction is that those relationships are best understood in relation to distribution and we are only now in a position to assess what value the flowers may add to those interpretations.
This article thus reports on the exploration of the Ouhangsberg, Hammansberg and Moddergat (Ouhoekberg) mountains as well as some work done in the Brandvlei Dam area. Again I put flower and flowering time aside – for the moment. This is in the same way I am inclined to the considered view that geographic information and field observation are the essential ingredients whereby ANY method has to provide explanation that satisfies physical experience. The alternative is that what we see and experience physically and mentally, is just the illusion that metaphysicists ascribe to creation.
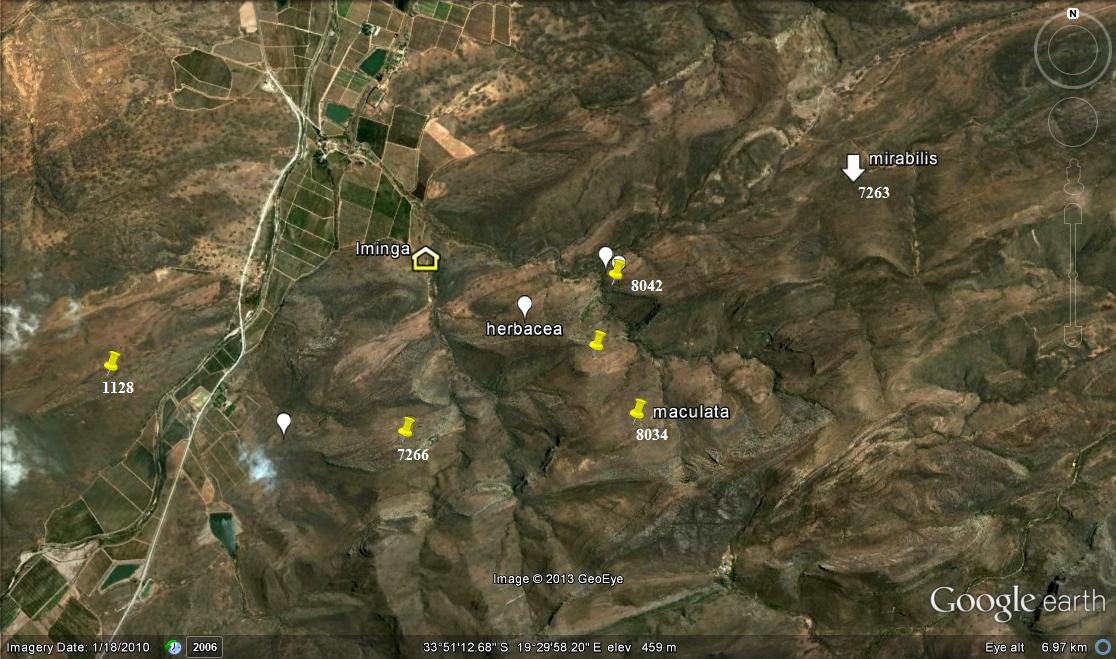
Populations recorded
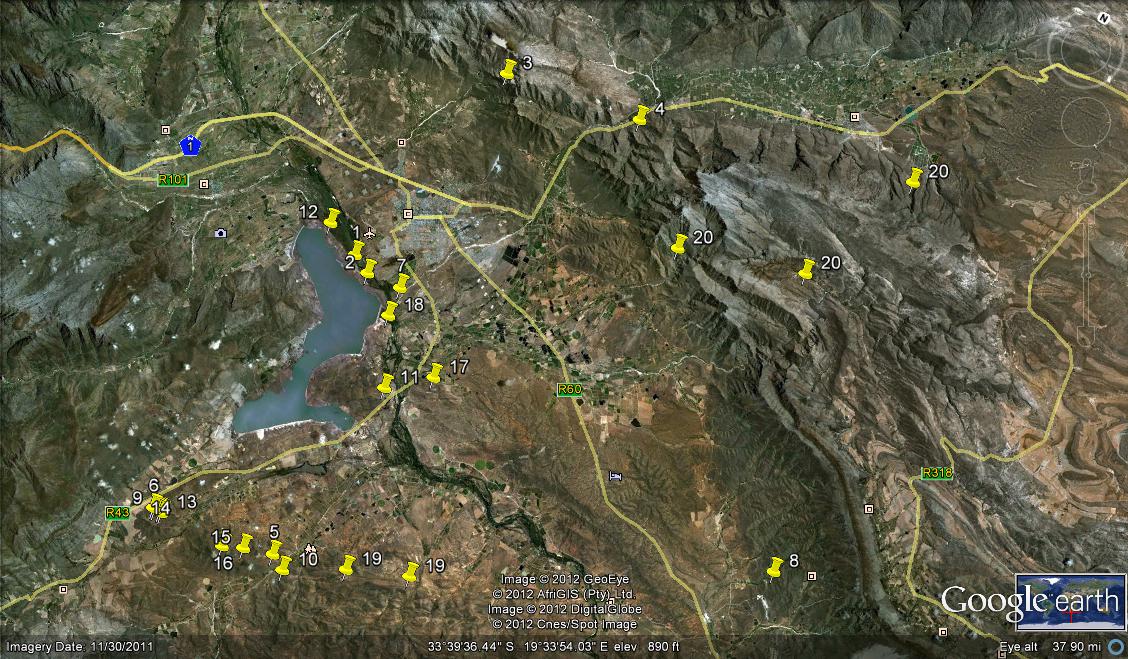
The following populations (see map) were found and observed:-
1. KG669/69 N Brandvlei Dam – set 1 2. MBB164 N Brandvlei Dam – set 1 3. MBB1119 (and 6815) Audensberg – set 2 4. MBB1120 (and 7994) Sandhills, Kanetvlei – set 2 5. MBB1128 (and 7066) W Lemoenpoort (type locality var. livida) – set 3 6. MBB1145 Ouhoekberg, Moddergat – set 5 7. MBB2591 NE Brandvlei Dam – set 1 8. MBB4461 (and 6514) Buitenstekloof – set 6 9. MBB7270 Ouhoekberg W – set 5 10. MBB7266 E Lemoenpoort – set 4 11. MBB7271 Cilmor Winery, Dewetsberg – set 7 12. MBB7526 Die Nekkies W. Brandvlei Dam – set 1 13. MBB7991a Ouhoekberg E – set 5 14. MBB7991b west Ouhoekberg E – set 5 15. MBB7992 Hammansberg midpoint – set 8 16. MBB7993 Hammansberg W – set 8 17. MBB7997 Sandberg H. pubescens, type locality – set 9 are for MBB7997 cf H. pubescens N of point 11 18. MBB7995 and 7996 Brickfield H. herbacea ‘submaculata’ – set 10 19. MBB7984, 7985 and 7988 Droogerivierberg, H. mirabilis – set 11 20. Unknowns – set 12
Map references for Haworthia maculata.
There are 4 records for the Brandvlei Dam but in Update 6, I report on H. maculata from the pump station east of the Resort, very abundant to the eastern point of Die Nekkies. Three of the records are for west of the Resort while I have not confirmed Etwin Aslander’s observation that he had seen it at the extreme western end of Die Nekkies. G.J. Payne in 1970 communicated to me that he had seen a Haworthia south of the Brandvlei Dam above and west of the Brandvlei Hot Spring. I was unable to confirm that in a single brief visit there ca. 1976. I have not been able to relocate plants at Sandhills but have no doubt that it will be found there if more thorough and extensive fieldwork is done there. In fact a very important consideration is that there is still enormous scope for exploration.
On each visit to the Ouhoekberg I have found plants in different local populations and none of the last four records is the same as seen in my first visit. At the original Lemoenpoort I always struggled to find plants at all, and the same applies to the locality to the southeast. At my last visits to these two places there have been many plants but relatively difficult to find.
H. herbacea is abundant in the Hammansberg and Ouhangsberg area as well as east of Cilmore. It has not been observed near the Ouhoekberg. It occurs between Cilmor and Die Nekkies in a larger form that acquired the name H. submaculata V.Poelln.; and I did suspect that there was a continuity with H. maculata that does not now seem to be the case. I exclude H. pubescens from this report. It occurs with H. herbacea east of Cilmor, but it also occurs on the northeastern slope of Dewetsberg north of the H. maculata recorded habitat. There are/were plants of H. mirabilis vegetatively very similar to H. pubescens at the Breede River Bridge southwest of Roberson. Their flowers and flowering times (March) correspond with H. mirabilis in that area. H. pubescens flowers in November (later than H. maculata at Die Nekkies or of H. herbacea – Sept/Oct) and the flower is similar to that of H. herbacea. It seems fairly obvious that H. pubescens fills the gap for H. mirabilis and the flower and flowering time are problematic.
The plants illustrated:
Set 1. Map points 1, 2, 7, 12. Die Nekkies Figs. 1.1 to 1.5 7526 H. maculata. Die Nekkies. Figs 1.6 to 1.10, sn* H. maculata, Die Nekkies – East.
A chapter in Update 6 deals with H.maculata – Die Nekkies Biomes, and Haworthia maculata These few images give scant insight into the variability in respect of rosette and leaf form, colour and marking, and proliferation.
Set 2. Map points 3, 4. Audensberg and Sandhills Figs. 2.1 to 2.5 MBB6185 H. maculata. Audensberg. Figs. 2.6 to 2.36 H. maculata, Vreesniet, Kanetvlei.
Unfortunately I found non-digital photography to be costly, cumbersome and unmanageable. So I have no field images for either population on the high Audensberg or at my original Sandhills site. However, I revisited the area and found the plants quite abundant a little further north at Vreesniet that is also Kanetvlei/Sandhills. The plants were almost confined to narrow rock cracks and their presence suggests to me that there is a very probable connection now eastwards to the Unknowns and even further on to the Osplaas H. arachnoidea, Etwin Aslander’s Hex Pass plants, and then still further to the H. marumana ‘dimorpha’ questionable.
Figs. 2.1 to 2.5 MBB6815 H. maculata. Audensberg.
Figs. 2.6 to 2.36 H. maculata, Vreesniet, Kanetvlei.
Set 3. Map point 5. Figs. 3.1 to 3.34 MBB7066 H. maculata, Lemoenpoort.
This is the type locality (i.e. MBB1128, W. Lemoenpoort) for H. maculata var. livida that I think is largely untenable or unnecessary fragmentation. Lemoenpoort is the valley that separates Hammansberg from Ouhangsberg. The first few plants seen were in exposed situations and had the purplish or bluish-grey colour that prompted the Latin name. This was maintained in cultivation. I linked it at the time to H. pubescens that seemed more probable at the time than to H. maculata and then simply because of the perceived demands of a relatively inflexible nomenclatural system. There are now seven new populations that contribute and improve understanding of H. maculata as a species. All these localities are in Witteberg Sandstone and the type locality is only slightly different in that the stratum of rock is less feldspathic (i.e. mineralized).
Set 4. Map point 10. SE Lemoenpoort Figs. 4.1 to 4.28 MBB7266 H. maculata SE Lemoenpoort.
This is a more west facing slope and also relatively low down on the mountain side. The plants were in rocks bordering an eroded softer shale band. They probably also occur higher on the slope in less mineralized sandstone. Generally the plants were very cryptic and tended to be solitary.
MBB7266 SE Lemoenpoort, view to southwestMBB7266 SE Lemoenpoort, looking up at locality.
Set 5. Map points 6, 9, 13, and 14. Ouhoekberg. Figs. 5.1 to 5.24 MBB7991a H. maculata, Ouhoekberg E. Figs. 5.25 to 5.28 Panoramic views. Figs. 5.29 to 5.33 MBB7991b H. maculata, Ouhoekberg.
These populations were first observed as one on a higher eastern point of the Ouhoekberg above Moddergat in about 1975. George Lombard accompanied Kobus Venter and me there in 1996 and we found them on the western high point. I located them nearby in 2004. We found them again recently as two more populations on the eastern heights near where I must have observed them first. Note the reticulation in the dried fruit capsule. It can be very much more evident in species like H. pulchella but I seriously doubt its diagnostic value. I have included views to… (a) north to show the water of the Brandvlei Dam in the far distance. The area between has not been adequately explored. What is interesting is the geology. The mountains on the left are the Table Mountain Sandstones, nearer is the valley where the soft Bokkeveld Shale has been eroded away, and then comes the Witteberg Sandstone with a neck of soft shale, and then low down is Dwyka tillite. H. herbacea is present on the Dwyka outcrop barely visible in the middle right. (b) The second view is looking east at first the Hammansberg and beyond that Ouhangsberg with H. mirabilis on its eastern flanks. The view looking southeast is over low Bokkeveld shale ridges with an abundance of H. herbacea. The view south is to Villiersdorp where Wolfkloof (not the Robertson Wolfkloof) is a deep valley behind the Table Mountain Sandstone left of the gap through the Rooihoogte Pass. Here is where H. herbacea ‘lupula’ occurs, unusually in sandstone. Altitude and skeletal soils of different origins contribute hugely to the genetic mosaic. Arable depositional soils exclude Haworthia and obviously inhibit contact between populations.
Ouhoekberg view to north.Ouhoekberg view to east.Ouhoekberg view to southeast.Ouhoekberg view to south.
Set 6. Map point 8. MBB4461 and 6514 H. mirabilis ‘notabilis’. Buitenstekloof
I am sorry that I have probably contributed to the confusion here. It started with von Poellnitz with a species he described as H. intermedia from this locality where both H. mirabilis ‘notabilis’ and H. reticulata occur, but he included E. Cape H. cooperi in the synonymy so it is not quite clear what he meant. While H. reticulata can be easily recognized for what it is at Buitenstekloof, the other population I ascribe to H. mirabilis ‘notabilis’. But I am not certain because I recall it flowering in November and with a flower type nearer to H. maculata than to H. mirabilis. It appears to me that further exploration may expose continuity westwards to the Sandhills/Audensberg populations of H. maculata and even northwards to unknowns.
H. mirabilis ‘notabilis’ (with or without the inverted commas) occurs from Klassvoogds between Ashton and Robertson, westwards through Wolfkloof, Robertson to Agtervink and further westwards still to Buitenstekloof. This all along the great Worcester fault line and intrusive igneous rock. The dolomite is apparently difficult to explain geologically and is associated with the granite-like extrusions in the area.
Habitat at BuitenstekloofView further into Buitenstekloof
Set 7. Map point 11. Cilmor. Figs. 7.1 to 7.3 MBB7271
An interesting point here that H. herbacea (map point 18) occurs between this point and all the Die Nekkies populations. I am not sure that the area between can be fully explored although I have been into it.
Set 8. Map points 15 and 16. Hammansberg. Figs. 8.1 to 8.21 MBB7992 H. maculata, Hammansberg.
The first locality 7992 midway between the eastern and western ends of the mountain was very exposed and very sparsely vegetated. The sandstone was very mineralised and fractured. The second locality to the west was well vegetated. In the view northward, the low hill in the near middle is the Draaivlei mountain that is Dwyka Tillite and hosts an abundance of H. herbacea as does the Dwyka strata on the northern side of the Hammansberg. The south side is Witteberg Sandstone with Bokkeveld shale on the lower south-facing slope. The view from the third locality southeastwards, shows massive valleys running into the Table Mountain Sandstone of the Riviersonderend mountains on the right. If one takes in mind that this is a short way east of the Wolfkloof Valley where H. herbacea ‘lupula’ occurs, then this area begs exploration. I do have a recall of H. herbacea in low altitude sandstone either near there or at a similar site at MacGregor to the east. The interesting view is the one looking eastwards that really gives the scale of exploration needed. The furthest mountains are the Langeberg Table Mountain Sandstsones and Buitenstekloof is nested in foothills there and in intrusive igneous rock. The coming closer is the Rooiberg, then Gemsbokberg, then Gannaberg, then Ouhangsberg and Hammansberg itself. Rooiberg is fairly well explored although the significance of H. herabcea ‘flaccida’ is not explained. I doubt if a anything new will crop up on the Hammansberg. But the intervening three definitely hold key populations marginal to these covered in this posting. The Langeberg? I only have the set 12 unknowns and cannot see myself resolving those.
view from Hammansberg eastwardsview from Hammansberg northwardsview Hammnsberg to the southeast
Set 9. Map point 17. H. pubescens. Sandberg. Figs. 9.1 to 9.20 MBB7997 H. pubescens, Dewetsberg.
I long ago observed H. pubescens southwest of Sandberg and south of the Breede River on the Dewetsberg. This is not much further than 500m north of a population of H. maculata at Cilmor Cellar. Visiting the site again I observe that the plants are slightly different to the Sandberg plants. While they are equally small, cryptic and dark coloured, they have less surface spinuliferation that gives rise to the species name. Also some plants are distinctly spotted in the way that H. maculata are and these spots may give way to translucens when the plants grow in less light. See the images showing leaf surface detail of H. pubescens compared to the two collections of the Brickfields plants (images 21-27) of H. herbacea in Set 10, map point 18.
Set 10. Map point 18. H. herbacea ‘submaculata’. Brickfield, Brandvlei Dam. Figs. 10.1 to 10.20 MBB7995 H. herbacea, S Brandvlei Brickfield. Figs. 10.21 to 10.53 MBB7996 H. herbacea, E Brandvlei Brickfield.
I have associated this locality with von Poellnitz H. submaculata and treated it as a synonym of H. herbacea. However, here I first illustrate a population about 600m south of that where the plants are in the usual size range for the species ie.30-40mm diam. At the locality east of the Brickfield and next to the Breede River, the plants are 1/3 to 1/2 as large again and can form huge clumps. North-west from this is a population of H. maculata at the extreme end of Die Nekkies and only about 300m distant, and this population I have always regarded as somewhat intermediate. What is interesting to note is the huge variation in leaf shape and armature within each population. In both cases the plants are in Witteberg Sandstones and at the first site this is both in a very shale-like stratum as well as in a highly quartzitic one. This is unusual for H. herbacea. Both populations wedge in geographically between H. maculata populations and in no known case do both occur.
7995 Habitat looking northwards
Set 11. Map point 19. H. mirabilis. Droogerivierberg. Figs. 11.1 to 11.11 MBB7984 H. mriabilis, Droogerivierberg. Figs. 11.12 to 11.24 MBB7985 H. mirabilis, Sandberg S. Figs. 11.25 to 11.31 MBB7988 H. mirabilis, Trappieskraalkloof.
In this area H. herbacea is common in the Dwyka Tillite but not in the Witteberg Sandstone. But instead of H. maculata, H. mirabilis is present there and commonly so. Where at one time I considered that H. pubescens was very probably an extension of H. mirabilis to a western and northern limit, this does not seem to be the case as it is so closely linked to H. maculata and H. herbacea. On the other hand I have wondered about the similarity of the Moddergat H. maculata to H. mirabilis. H. mirabilis is in the eastern areas of the Ouhangsberg and now we know that H. maculata occurs not far away on the western slopes. The centre area remains unexplored and this needs to be done as it may also help understand the idea of two principle role players viz. H. retusa and H. mirabilis as precursors of a very complex Southern Cape assemblage.
Set 12. Map points 20. Unknowns. Figs. 12.1 to 12.7 MBB7865 H. cf. arachnoidea. Keurkloof, SE Dedoorns. Figs. 12.9 to 12.15 EA1441 unknown, Hex Pass.
I have only two sets of pictures for this set. These are…(a) MBB7865 that Ernst van Jaarsveld drew to my attention at Keurkloof, south east of De Doorns in the Hex River Valley. There is a continuum of suitable habitat between there and Sandhills/Kanetvlei; (b) EA1441 and this number may be incorrect, but it is at the near-base of the Hex River Pass still further east than Keurkloof. In between is Osplaas Station where there are dramatic forms of H. arachnoidea with very spotted leaves, not to forget the nortieri-like form of H. arachnoidea at Kanetvlei about 200m north of MBB7994 H. maculata.
H. cf. arachnoidea. Keurkloof, SE Dedooorns 0087865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 0067865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 005
Conclusion The nature of the populations of H. maculata on Die Nekkies varies quite considerably in that there is an area east of the Resort where the plants form huge clumps. Individual rosettes can be quite large. But there are also points where the plants tend to be solitary, hidden in rock cracks or even truncated into the soil. The plants in the Southern populations are smaller and tend to be solitary. There may be differences in flowers and flowering time but this will really be significant in relation to H. mirabilis rather than between the populations recorded here or to H. herbacea. Attention must be paid to the way in which H. herbacea is geographically wedged in between populations of H. maculata. Equally significant is the character of H. pubescens southwest of Sandberg (Dewetsberg) where the plants have less spinuliferous leaf surfaces and also spotting (maculation). This is one of the best pieces of evidence I have for the geographic continuities that exist throughout the genus both in respect of species of how I think species can be recognized and of how difficult decision making is. In essence this report only dismisses the idea that the Lemoenpoort population can be assigned to H. pubescens. I doubt if it is rational to even formally recognize it as different to H. maculata as “variety” itself is a mythical status. Fragmentation by names is a rather archaic approach to classification that serves hardly anything but a semi-commercial need.
Acknowledgement Many people contributed in one way or another to this report … Mr Hentie deWet of Moddergat, Messrs Poffie and Hettie Conradie of Sandberg (South), Mr Pieter Naude of Vreesnicht, Mr Johan and Marie Fourie of Buitenstekloof, Mr Nico Marais of Worcester (Brandvlei) Brickfield, Mr A. Groenewald of Cilmor Cellar and Stefan Hugo of River Farm. Messrs PD and Anso leRoux provided much logistic support, interest and company as did Kobus Venter, Etwin Aslander and Werner Voigt. Lawrence Loucka has managed and facilitated the archiving of publications and pictures.
MBB1128 (MBB7066 W. Lemoenpoort) is the type locality for H. maculata var. livida. Plant pics are available in Haworthia Update Vol. 9. A myth corrected to, population 5, set 3. Here are flower photos from 8 December 2012.
MBB4461 (and (MBB6514) H. mirabilis ‘notabilis’ Buitenstekloof
In Haworthia Update Vol. 9 and a recent posting on Haworthia nortieri Bruce Bayer makes reference to this Haworthia maculata population, MBB7994 from Kanetvlei.
MBB7991 Moddergat above Ouhoekberg E, ex hort. single clone
From Updates Volume 9 Set 5 Bayer writes, “These populations were first observed as one on a higher eastern point of the Ouhoekberg above Moddergat in about 1975. George Lombard accompanied Kobus Venter and me there in 1996 and we found them on the western high point.”
These photos were taken 20 December 2012 from a single ex hort. clone and hence consistency, but still illustrative.
8034 Haworthia maculata ‘livida’, Ouhangsberg.
These plants occur in Pteronia communities on very dry and sparsely vegetated north slopes. The geology is Witteberg Sandstone but the particular strata the plants prefer seems to be a softer more erodible and less quartzitic stratum than they occupy at Lemoenpoort to the west. The plants appear to have flowered early in December coinciding with the flowering at Lemoenpoort and southeast Lemoenpoort. The two localities where we observed the plants are approximately midway between H. mirabils at Droogerivierberg to the east and Lemoenpoort to the west. There is no suggestion that they form any continuum and the flowering times are different.
The name of the farm is Iminga Mountain Reserve, a large tract of quite non-arable countryside that is appreciated by the owner as her garden.
Acknowledgement – I deeply appreciate the freedom to explore Ouhangsberg granted by Ms Suzi Broomberg.
Following on from the 8034 record, we returned to explore still further east in a direct line from the 8034 localities to the H. mirabilis population on Droogerivierberg (see map). We found plants at the same altitude as we had observed H. herbacea on the previous visit. The plants were again very cryptic and hard to find. Flowering was also long past and the few capsules present contained very little seed. The situation with respect to habitat was a little different and these plants were just off a truly sandstone stratum and in a narrow band of renosterveld. H. herbacea was within 50m of the nearest H. maculata in a more karoid vegetation similar to that where we had seen H. maculata on the previous visit. This is difficult to understand. Southeast of the Brandvlei Dam, H. herbacea occupies habitat where one would expect to find H. maculata. But it does not do so in the Ouhoekberg nor Hammansberg. Here north of the Ouhangsberg there does not seem to be a visible difference in the choice of habitats for the two species but they cannot be said to truly co-occur i.e. grow together in such close proximity that they intermingle.
The spatial relationships of the various populations are now quite significant. The spread from localities east to west on the map is 5.23km. The distance between known H. mirabilis and nearest known H. maculata is nearly 1.5km. There is no indication of transition and present evidence is that there is no continuity between these two species. There is however still that space between to explore as well as a massive valley south between the two populations in question.
Fig. 1 Image of Iminga Mountain Reserve.Fig. 2 Westwards towards homestead.Fig. 3 Below locality at fig 2, southward.
Bruce Bayer has now been back to Die Nekkies and returns with more detail to add to the broader Haworthia maculata picture, and considerable detail for H. maculata at its home range located north of the Brandvlei Dam on a low range of hills named Die Nekkies. (See Haworthia Updates Volume 9, A myth corrected to – Haworthia maculata var. livida (Bayer) Bayer – and flowers ignored). These low hills extend for 10-12km east to west along the north shore of the Dam. They are geologically Witteberg Sandstone and H. maculata is not known off this formation except in the lower Hex River Pass where it is found in Table Mountain Sandstone. H. herbacea (and H. reticulata) are more abundant on the Dwyka Tillite, and on Bokkeveld and Ecca shales. The Wiiteberg is sandwiched between Bokkeveld Shale and Dwyka Tillite, and Ecca Shale of the Karoo System overlies Dwyka Tillite. Because of considerable faulting and folding the Cape Terrain is geologically and topographically very broken and there are a multitude of skeletal rocky habitats – the raw rock is very exposed and the surfaces are erosional. Haworthia are rarely found on depositional sites. The fact that there is also a stable and dependable winter rainfall also contributes greatly to the success of small succulents in situations that cannot maintain high biomass levels. This all drives home the fact that Darwin’s dictum that distribution is the lintel to understanding species is doubly true for Haworthia where morphological differences are minimal.
In Die Nekkies and Biomes and Haworthia maculataHaworthia Updates Volume 6, Chapter 6 Bruce Bayer explained and presented data in connection with the variation and distribution of what he considers to be the species Haworthia maculata over the whole range of its distribution. He long ago wrote explanations like this for H. reticulata and H. herbacea that are related species. Bayer has already shown that in genera like this within the very diverse Southern African system, simply do not resolve on the basis of simplified morphological characterization. The new data presented here, especially with the observations on the varied flowering time of H. maculata, shows that the view expressed, that H. retusa and H. mirabilis really form one integrated system, is true. This further underscores the view long ago held by Bayer that H. herbacea, H. reticulata and H. maculata are further extensions of that single system. Even the floral morphology points to this.
– Lawrence Loucka [photographs and map by Bruce Bayer]
7996 as H. herbacea is very close to 8020 that is H. maculata and the nature of the plants at 8020 is that the population is transitional to herbacea too. In fact the same is almost true for 7996.
Haworthia maculata MBB7526, Die Nekkies
What a difference a few weeks can make … above photos taken November 1, 2012. Below were taken December 8th.
And an interesting variant…
MBB8019 – Haworthia maculata, Yacht Club Gate, Brandvlei
This is a picture taken from a very late flowering population of H. maculata near Die Nekkies resort south of Worcester. Looking north to where H. maculata grows at about 4000ft on the Audenberg peak, and to the right at about 1000ft in the lower Hex River pass. That is where it occurs close to H. aff nortieri.
Die Nekkies – a selection of Haworthia maculata plants with their leaf faces and backs showing variability over this extensive area.
I took the following photographs originally just to show how much the leaves varied in the species. The only locality data record was Die Nekkies with no differentiation into populations. The variability was firmly established and became apparent even within populations where the degree of variation surprised even me. It was also extrapolated across the genus.
As an afterthought, this turned my mind to Farden and Smith’s observation about the idiocy of trying to make varieties in Haworthia attenuata on the basis of leaf characters where the individual leaves varied so much that several varieties could be detected from the same plant. This is exactly what Breuer, Marx and Hayashi are actually doing when they get carried away by their justification for new species, drawing conclusions from single plants because of the inability of the mind to hold many images. This is exactly the way in which “H. magnifica” is maintained as a species when the imagery is based on the original single plant originally described. There can be no such thing as “H. magnifica” because the variation across the range of populations from Riversdale to Riviersonderend is so immense and complex.
The following photographs show a random selection of plants and their leaf faces and backs from Die Nekkies. These were taken some time ago when I did not record actual populations. They do record the variability of plants and leaves over this wide area which I classify as Haworthia maculata. For subsequently photographed plants I have recorded my population collection numbers which are indicated on the map found in Die Nekkies Haworthia – A further visit and updated information.
In Haworthia Update Vol 9 there is a report of a population MBB7997 identified as Haworthia pubescens from north of the Cilmor wine cellary. This is approximately 2-3km southwest of the type locality for the species. I noted that the plants have less spinuliferous leaf surfaces and there is a degree of surface translucens and maculation (spotting). I also presented 3 pictures of MBB7271 of what I identified as H. maculata from south of the Cilmor cellar. When I first visited this locality I had no problem identifying the few plants I saw as H. maculata on account of their marked spotting. However, on a recent visit we struggled to find plants at all and the few plants we found were too embedded in rock cracks to make any worthwhile identification. So we revisited the site to explore more extensively and located a large number of plants higher up and slightly west of our first sightings. These plants are illustrated here. They incline more to H. maculata than the plants at MB7997 and I have accessioned the population as MBB8002. There is the usual expected large variation in respect of superficial and observable characters. The plants can be proliferous and cluster, more so than at MBB7997. Similarly the leaves can have more translucens and even less spinuliferousness of the surfaces. Some plants have few and quite thick swollen leaves while others may have more and very slender pointed leaves. I have not observed the flowers and really do not expect them to make any difference to the problematic classification of populations that again are neither here nor there in a narrow concept of species. H. herbacea occurs at all four geographic positions at a radius of about 2km. At the brickfield to the northwest as well as just northeast of the Brandvlei Dam wall it is evident to me that there is a transition between H. maculata and H.herbacea. I did report the known distribution of H. maculata in Update 9. While there is no suitable habitat between Die Nekkies hills at the Brandvlei Dam and the Audensburg or Kanetvlei, there is unexplored suitable habitat southwards to Moddergat and Hammansberg. There is no evidence of H. maculata eastwards to where H. reticulata is known about 15km east on Ribbokkop. Westwards no Haworthia is known although G.J. Payne did inform me that he had observed plants in the hills immediately southwest of the Dam at the now submerged hot spring in the Brandvlei prison area.
View 1 From Cilmor to the northeast.View 2 From Cilmor eastwards.
The submitted pictures include two views. View 1 is looking north of east across the Breede River to the Sandberg where H. pubescens occurs. Its full occurrence on those low hills is not known and this I will explore soon. View 2 is looking eastwards looking at a Dwyka Tillite hill across the river in the upper right. We found no Haworthia on that hill although both H. pumila and H. herbacea are present on the smaller rise to the right and behind it – also Dwyka. H.herbacea is very abundant on a Dwyka tillite hill about 10km to the south. The corresponding hill on the left is Ribbokkop where H. herbaea, H. reticulata and hybrids are present, and H. arachnoidea also occurs. The limits of H. mirabilis are the higher hills in the background viz. Rooiberg, Gemsbokberg and those are Witteberg sandstones. ♦
I need to point out that there is a still earlier article which covers Haworthia maculata (Haworthia maculata <–> Haworthia pubescens) that lays the basis for this discussion. In that article I note the position of the Sandberg to Cilmor and DeWetsberg and intended to include the Sandberg H. pubescens in that article. We could not get landowner contact and so that fell away. However, this problem was overcome and we first explored a Dwyka Tillite outcrop southeast of Sandberg. There is a vast accumulation of windblown sand on the first hill and we saw no Haworthia. There is a smaller hill further to the southeast that is also Dwyka and erosion exceeds wind deposition so smaller non-geophytes do quite well. We found both H. herbacea (see fig.1 MBB8014) and H. pumila there. From there we went to the southernmost point of the Sandberg. A misjudgment landed our vehicle in mud and the drama to get out limited the time we had to explore. We found a lone H. herbacea (fig. 2 MBB8012). Returning a week later we approached the Sandberg from the southwest, and almost immediately on reaching the top we found H. pubescent. Fig. 3 is a view towards Cilmor and DeWetsberg where the plants appear to be intermediate H. pubescens↔H. maculata. The picture is useful to get some idea of the role of geographic and geological considerations. The high mountains in the background are Table Mountain Sandstone and no Haworthia is known there. I am not certain that this is true and G J Payne did tell me that he had seen plants on the extreme lower right and south of the Brandvlei Dam. But also on the absolute distant and absolute left, is the Riviersonderend Mt. That is also TMS. The deep Wolfkloof Valley behind that is the locality for the much unexpected H. herbacea ‘lupula’. (These inverted single commas are not entirely necessary but I use them to underscore my informal use of names that have less reality. The var. lupula is real). The mountains ahead of that last line are Hammansberg on the left and the Moddergat to the right. Between there and DeWetsberg has not so far turned up Haworthia, but this is an exploration problem. Behind the DeWetsberg is also underexplored. H. herbacea does occur between DeWetsberg and behind the mountains on the low right just in the picture and also east of the brickfield out further right. H. maculata is only known in this area along the Nekkies north (further to the right) of the Brandvlei Dam just visible in the picture.
01. MBB8014 H. herbacea, E Sandberg.02. MBB8012 H. herbnacea, Klipkranz Camp.03 . View west of south from Sandberg.
Prior to this exploration H. pubescens was to me only known from the northern part of the Sandberg that lies south of a road going eastwards to Eilandia between Worcester and Robertson. Here H. herbacea does occur on the lower northwest warm slopes. H. pubescens seems to occur only on the upper two ridges and H. herbacea is not known to intermingle with it. This is all Witteberg Sandstone that as a formation overlies Bokkeveld Shale and underlies TMS.
Coming back to the southwest corner of the Sandberg where we found H. pubescens. The plants seem very similar to the species as it occurs to the north. They were very cryptic and often in shady rock retreats where they were really hard to see. It was mid- to late-morning when we were there and the plants were not going to be better exposed as the sun moved further west. Although there was very suitable well-drained habitat lower down on the shaded east slopes, there were no plants and I speculate that this may be because the plants may need the cooling effect of wind movement up on the ridge. The pictures tell the story of variability in respect of a whole range of leaf and rosette characters.
It is worth noting fig. 43 of the dead remains of a plant under a clump of restioid. It seems that seedling survival is closely coupled to early protection giving rise to the concept of nurse-plants. Plants are often very difficult to find because they are so hidden beneath accompanying vegetation. But they do need light and the dynamics of vegetation growth and densification must have quite a big impact on the ageing and survival of plants. It raises again the question of how long do the plants live? For plants like Aloe ferox and A. dichotoma I do have a real experience of a lower limit of about 35 years and a top limit in the several hundred. In the field, the plants seem comfortably ageless.
The really interesting part is this. While I was busy tediously cleaning a plant to photograph it, Daphne called to me to come and see a lighter green plant she had seen. Moving in that direction I saw a plant that registered as H. herbacea but with some hesitation and doubt (see fig.4). I then went to see what Daphne had observed. They seemed less obviously H. herbacea but that seemed to be a logical and conservative opinion (see figs. 5-11). That was until Daphne found two adjacent rosettes at the foot of a restioid clump that left me in no doubt that they were hybrid H. pubescens/maculate (see figs 9-12). These were in bud whereas H. pubescens plants showed no sign of impending flowering. Note the buds are less well developed than MBB8014 further east, even if possibly insignificant. Going back to the other plants we confirmed my doubts. They were a lighter colour and apparently softer texture that we would have expected in H. herbacea. These were the only plants we saw in a space bridging the occurrences of plants of H. pubescens.
04. MBB8013 H. herbaceaXherbacea, Sandberg. 05-11 MBB8013 H. herbaceaXherbacea, Sandberg
12-46 MBB8013 H. pubescens, SW Sandberg.
We did not explore the western slopes where habitat would have been more suitable for H. herbacea and I expect it does occur there. What puzzles me is that so frequently have I found very distinctive hybrids between species in close proximity and very seldom where the species are some distance from one another. I cannot say I have ever found a hybrid in the clear absence of both parents. The example of Astroworthia bicarinata at Lemoenkloof, east of Barrydale, may be an exception where only Astroloba corrugata (syn A. muricata, A. aspera) is present but H. pumila apparently not. Hybridization is thought to be an important element in the “evolution” of new species. I doubt this as it is quite evident that separation into two species is a pre-requirement. If new species have evolved in Haworthia by hybridization, how did they evolve as such in the first place? The answer to me lies in the continuities between populations. I observe, and have experienced of expected continuity between populations. While the Cilmor populations are thought to be H. pubescens ↔ maculata it cannot be said anymore that they are hybrid, or populations where the morph or drift to discrete elements has not reached a conclusion. The latter is more likely. As there is already apparent geographic continuity of H. maculata and H. herbacea, I was expecting some evidence of a similar relationship between H. herbacea and H. herbacea. So here it is. Hybridization as a factor in speciation in Haworthia does not seem to very likely. It confirms for me that there is a fractal “chaotic” order to species in Haworthia and the reality is that a view of many truly discrete species is a fabrication and a very ill-considered view.
Acknowledgement We are always greeted with such kindness and helpfulness that we might have expected this from the Sandberg landowners too. It came in no small measure. Driaan Griesel was most enthusiastic and interested and also helped us with extracting our vehicle from the mud on the one occasion, and then jump-starting it after a flat battery on the second. Our imposition did not so much as touch his view of the day.
[Ed.] Bruce made another visit to Southwest Sandberg on 9 December 2012 and includes the following flower pictures. He makes this comment; personal correspondence 27 December 2012.
I am actually not sure at all about flowering time now. I used to be quite sure of being able to collect seed of pubescens mid-Nov. But I observed at Humansdorp that gordoniana peak flowering could be out by 6 weeks. In any case the plants can produce successive spikes so one can get delayed flowering and added to that energy in the first or the second flower set. I know mirabilis at MacGregor can flower from Nov. thru to March while at Montagu mirabilis can flower as late as April/May. Retusa and geraldii are quite happy to produce flowers in either Spring or Summer and Kobus observed that splendens did that too. Maculata can flower from Sept. thru to late Dec. And each population does its own thing.
A whole bunch of pictures of H. maculata. I would have thought my Handbooks might have discouraged the “new species” guys who do not seem to recognise what the nature of the game is. There were about 450 names being bandied about that I reduced to about 140. It just seemed to spur these guys on to inventing several hundred more to recreate an unmanageable situation? Perhaps the science of botany itself lost the plot when it was decided that the practicality of genera was actually built around the need for management rather than a dubious science of evolution.

























































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}