In trying to achieve some sort of closure to my writing about Haworthia, I need to try and say a little more about the problems of communication between taxonomist/authority and non-authority. Mrs. Doreen Court as an author, botanist and taxonomist expressed some doubt about the correctness of regarding H. maughanii as a variety of H. truncata. Recently Stirling Baker, a grower and enthusiast, expressed similar views. This is a good example of the kind of problem that one encounters when the taxonomist has to make the decisions for an entire group based on his/her entire experience. It is in fact the sad burden and sorry lot of the taxonomist to have to field and bear valid comments of this nature, and opinions views and dissent, from those who are free of that responsibility. In my experience of co-operating with other taxonomists, I experience their frustration when I use names which have no authority. In fact in writing about species it is actually a problem to decide how to refer to collections when the conclusion will involve a change in how names are commonly used and will have been used in the text.
The question of actual knowledge is also relevant. The taxonomist has to organize and deal with a substantial body of information which often includes an array of herbarium specimens including those of doubtful origin and quality. He/she has to work with an array of images which may be quite conflicting whereas the casual observer may rely on a single image from either good or bad source. The taxonomist also has the problem of validity and chronological order of alternative names to deal with. Over and above these difficulties are the problems of definition and the reality of taxonomic ranks. Up to the present time, the evolutionary basis of speciation is pictured in the form of a two-dimensional branching tree. All the branching points are clearly indicated and the suggestion is that the end-points of the branches are the species with equal degrees of difference. It should be asked if this is a good model. There is evidence that taxonomists are questioning the reality of the species themselves, and certainly this should be extended to genera.
The tendency in South African botany is to jettison genera which are represented by only one species. This does have a lot of merit in the sense of the meaning of the word “genus”, but it only moves the problem of degree of difference to a different level. This problem of “level” is one of the biggest stumbling blocks to agreement in Haworthia. From my point of view the evidence is that the species differ enormously in their degree of differentiation and variability. There are vast complex systems of populations for which no simple nomenclatural solution is possible, as opposed to other simple systems which can be readily identified and referred to by a name. In the final chapters of this volume I discuss a series of populations from the lower Breede River drainage system. However, it is not the final word on Haworthia as it occurs there. I have since explored the area further and encountered other variants which I feel I almost dare not discuss in view of the further nomenclatural problems and identifications these populations raise.
I am extremely conscious of the fact that there is a vast area still to be explored, but I hold no optimistic ideas that such exploration is going to produce solutions. Other Haworthia taxonomists (and such an arrangement is fairly restricted to Haworthia) are content with the belief that an endless array of new names is as good a solution as any. Certainly this can be understood and followed by casual growers and observers where a name is useful in terms of what is commonly known and shared. My opinion is that this is acceptable within such closed communities, but quite useless in the process of building real and valuable information, and in trying to understand the world around us.
~ Bruce Bayer
Editor’s Note Update 3 brings up-to-date the results of Bruce Bayer’s field research, but as he is constantly in search of haworthias in habitat it will not be surprising if he eventually brings to light further populations, which have an impact on classification. We can all look forward to hearing about his further field work in due course.
It is by no means uncommon for Haworthia enthusiasts to visit South Africa for the purpose of seeing haworthias in habitat. Such visits do give an indication of the variability of plants, but it is probably only extensive field research which reveals the true extent of variability and the difficulties it poses for classification. Living in South Africa, Bruce is in a fortunate position to undertake such work. We cannot visit South Africa with a frequency which will allow us first hand experience of the full extent of variability and its implications for classification, but we can read Haworthia Revisited and Updates 1 – 3 and study the many photographs, excellent as herbarium specimens, in order to gain a better appreciation and understanding of the genus and its associated problems. The results of Bruce’s field work are now available for everyone to take into account when assessing the genus. This does not guarantee that everyone will come to the same taxonomic conclusions, but it does mean they will have the information at hand for consideration in arriving at their own conclusions.
Bruce mentions the different objective in classification perceived by different groups, from the professional taxonomist to traders and purchasers, all of whom want names to have meaning. Alas, there is no agreement on what that meaning should be. For one professional it may be a group of variable plants which he considers to be related – one name covers the lot. Another may see a number of different groups and give them different names, though there may be variability within a group. Sellers and purchasers on the other hand may want a name to indicate a group of plants which have very similar form so that a name will enable a purchaser to buy exactly what he wants – purchasers do not want a name to indicate a lottery. How these problems are solved is a matter for current debate. I am always interested in receiving contributions for publication in our journal Alsterworthia International.
It is, of course, possible to give plants cultivar names under the International Code of Nomenclature for Cultivated Plants, but this is another source of names which is not without its problems. At the present time Alsterworthia International is attempting to assemble information on hybrids and cultivars for publication as reference material. Further details will be announced when sufficient information has been assembled for publication.
~ Harry Mays
Acknowledgements I would like to re-iterate what I wrote in the acknowledgements for Update Vol. 2. Since completing the manuscripts I have been into the field many times and reflected on just how much good will, of so many people, is associated with the whole process. Landowner permission is the first requirement and this often means a lengthy inquiry process as one tries to establish who actually owns the property one wants to explore and who can be contacted and where. Then one has to justify oneself and also establish credibility. It simply is not sensible to even try and list these persons to whom one then is eventually indebted and so grateful to Steven Hammer has been kind enough to proofread Update 3, after also doing so for Update 2. I am grateful to have been able to stress his goodwill in this way and I trust that he will not suffer any backlash from an intolerant taxonomically minded community. My impressions seem to be constantly under review and my present opinion is that taxonomy is overlaid with pedagogic and pedantic activity which is assumed to be for science. It has to be asked if species do in fact have any objective reality and would it not be far better just to recognise that a system based on consensus would be desirable? While I seek closure on my contribution to the unhappy state of names for haworthias, I must say that I have some further writing to do.
The problems of species classification of Haworthia should now be well known to all enthusiasts of this interesting genus. I have proposed and maintained, with cosmetic changes, a nomenclatural system for it since 1975. It is a system with which I have managed an extensive collection and herbarium record, and I know it works within the limitations imposed by the evident fractal nature of “species” and their variability. In this paper I would like to expose these limitations with respect to the concept of two species viz. Haworthia mirabilis, and Haworthia maraisii, where there may be only one. In the original hardcopy publication of this article, the illustrations are all captioned H. maraisii when it would have been sensible to have used H. mirabilis.
In most discussions concerning the classification of Haworthia, participants have suggested that there are too many species and that some of them should be “lumped”. On the other hand, there have been several writers who, as prospective taxonomists and experts on the group, continue to expand the range of entities at the formal rank of species and varieties despite all the evidence and indications that this could be an endless path leading nowhere. My own inclinations have been to minimize the number of species and to use varietal rank in two ways as a communication medium. One is that I have tended to reject varieties of older authors which I did not think had a strong enough geographical base, but also recognizing that there is nevertheless some information inherent in such names. Often I felt these varieties simply expressed the variability in a way that was insignificant with respect to the species as a whole. Two, is that I have described some new varieties to provide names for morphological variation that I consider is new and previously unrecognized, accepting that these names should perhaps not be immortalized either. Thus my proviso has been that these new varietal names should not be taken too seriously. As far as the number of species is concerned, I know full well that there could be fewer species. I get caught up in the problem of identifying a “species” in a strict botanical definition of the word, as opposed to the need for “names” as a way of simple communication about the plants in the amateur fraternity. Because of the problem of similarity and continuity, the elimination of names becomes similar to that of falling dominoes and the question then arises of “where does it end?” My classification should not be confused with a system of names intended for horticulture or for trade. But neither do I think such a system should adulterate a formal botanical one.
Therefore, underpinning this presentation is a definition of a species as dynamic systems of living organisms morphologically, genetically and even behaviorally, continuous in space and time.
What is an Haworthia species? I do not accept that there are different kinds of species nor do I accept the term “biological species”. A species is a biological construct intended for life form and the classification of Haworthia has been presumably intended to identify those individual species that constitute a genus set. It seems evident that the commonest element of any species concept, expressed or unexpressed, is that of reproductive isolation i.e. members of different species cannot interbreed. In Haworthia this evidently does not apply because there is compatibility between individual plants of even different subgenera, what to say of intergeneric hybridization. If there are breeding barriers they are the obvious isolationary ones that are regarded by zoologists as prelude to speciation i.e. geographic isolation, behavioural (different flowering time) and genetic (polyploidy) factors. Genetic or other breeding complexities may, in my opinion, be characteristics within such sets as much as they help define them. In that respect I do consider that geographical (and geological) breeding and survival barriers are in evidence and are significant. My perception of the species is then of sets of individuals and populations which suggest morphological and genetic continuity in time and space
The problem of continuity is nothing more than the assessment of similarity and difference. This point needs examination, as it is basic to the question of characterization and classification. The species is the basic biological unit used to identify life forms. In the higher life forms (animals) it seems to be fairly straightforward and what could be problems are easily resolved e.g. the lion species is essentially African, but there is an Asian representative. This is similarly the situation of the elephant species. In one case are they classified as different species, in the other, two. Zoology recognizes and places strong emphasis on the capacity of animal life forms to interbreed and produce fertile offspring. In the lower life forms there are problems because interbreeding capacity does not seem to relate so closely to the life forms and fertile hybrids are produced from improbable parentage. However, it is unlikely that this is the criterion that is actually used to identify “species”. In my opinion reproductive isolation in a species definition has been a byproduct of evolutionary philosophy and that botanists have simply assumed that a species definition for animals also holds for plants.
Continuity is an aspect that I do consider in relation to occurrence with other species and in relation to geography. My observations have been that plants within populations, and thus the populations themselves, show trends. These trends are referred to in botany and zoology as clines and it does happen that the tails or two ends of such a cline may meet and thus produce a situation where a classification decision for one geographic area, may not be correct for another. Then there is the problem of skeletal soils and habitat difference and the effect this has on plant appearances – either generated simply by different soils and/or due to differential selection on the basis of genetic characters. It seems to me simply obvious that sometimes the quite apparent differences between two or more groups of plants is due to local differences in geology and has no general application to the same plants over a greater geographical range. Perhaps it is worth admitting Darwin’s observation to be correct: any local truth or integrity of species is greatly reduced over large areas, and perhaps extend the idea that it is also very greatly reduced by the amount of material examined.
Ideally similarity or difference should be measurable and this is manifested in numerical taxonomy whereby measurements of every conceivable feature are accumulated and processed statistically. It is not possible for me to generate such statistical data in any conventional way and I have to rely on perception derived from learning and experience of what constitutes “difference”. Collecting permits only allow three specimens and it would in any case be ridiculous to suggest that more plants than this should be gathered simply for statistical demands. This learning of mine has been done not only within the limits of Haworthia, but within animal (largely insect) and many other plant genera.
How can a Haworthia species be circumscribed? Description plays a critical role in the examination of similarity. The irony is that if one examines all the descriptions relating to the taxa (species and varieties) that I will cover in this article, it is patently obvious that descriptive detail is practically worthless. Detail barely enables one to describe a single specimen with confidence that it could be identified among others on the basis of its description, and this is even less likely if the growing conditions of that one plant are changed. It has been difficult even within the scope of this one study to photograph plants that are strictly comparable in terms of growing condition. We are also faced with the fact that individual plants within populations are highly variable and that the description of a single plant is far from the description of a population. Furthermore, the description of populations is also compounded by numbers and the difficulty of arriving at a norm by which it would be reasonably possible to identify individuals of any one population of few or many.
Objective? Recognizing the above limitations, this article is intended to examine whether or not H. mirabilis and H. maraisii, as I presented them in my revision (Haworthia Revisited, 1999), can be shown to be separable sets of individuals and populations. I wish to illustrate H. mirabilis and the interplay of this species with H. maraisii as I have experienced it in and around the Riviersonderend Mountains. I conclude that they are but one species and within that decision is nested the problem of what then of related species and where does it end? This is because the whole story can barely be told in one piece as both elements have extended distributions where they unfold other variance and interact with other “species”. Thus other names have also been used in this context and there is a plethora of such names associated with the problem of characterization and classification that need to be understood.
History and background information Although the name H. mirabilis has a long taxonomic history dating from the original description by Haworth in 1804, the name was curiously not associated with field plants until I did so in 1973 (Haworthia Handbook). In the interim several other species (and varieties) were described which originated out of what is obviously a single complex in the greater Bredasdorp/Caledon area. These species included names such as mundula, badia, triebneriana, eleven varieties of triebneriana, and emelyae var. beukmannii. Typification of the name is quite easy if the exact original locality is known. But it is difficult for the nomenclatural type to be typical of any taxon for the reason that in Haworthia, the plants grow in scattered and isolated populations. There is variability and difference of plants both within and between these populations. Often one can be completely certain from which population a plant may have come, and in other cases it is not even certain from which set of populations a plant may have come. Thus it was that the name mirabilis had not been associated with any natural source until I did so, and then I felt constrained to conclude that the population from which it most probably had come was that from which G.G. Smith had drawn his H. mundula. This particular population is very characteristic and individuals can be identified with a very high degree of certainty. The consequence is that we have a situation where the species name is based on plants that are not truly representative of the others in the set. Many of the other variants were described under the name H. triebneriana. Some of these can be identified with a high degree of certainty and others not. The name triebneriana seems to be associated with a population at Stormsvlei (it is certain that von Poellnitz wrongly used the name Strydomsvlei) and there are two other varieties that also had this place as source – they are H. triebneriana vars. depauperata and pulchra. This suggests that the var. triebneriana cannot be identified with certainty. The variety beukmannii was attached to the species H. emelyae, and this also indicates how indeterminate the species were in the absence of reasonable definition and capability.
A similar situation exists with regard to the name maraisii which was a species originally also collected at Stormsvlei. We only know this because it was originally reported in the description as having come from “near Swellendam”, and it was G.J. Payne who showed me the place at Stormsvlei. It was I who extended the use of the name to a wider range of populations, as I will discuss in this paper. I have said elsewhere (Haworthiad 17:83, 2003) that there was an unfortunate decision by W.T. Stearn to typify the name herbacea on the basis of an old illustration (Boerhaave, 1720) that was better associated with the name atrovirens. I unfortunately applied this name herbacea according to the descriptive content of the name (yellow-green in reference to spines) and used it for a species in the Worcester/Robertson area. The name atrovirens, the meaning of this name (dark green referring to leaves), and the accompanying original illustration could perhaps have been better applied to the species which von Poellnitz named H. maraisii and also to H. schuldtiana. The species name H. maraisii thus originates at Stormsvlei and I have used the name for a series of smallish dark-green plants occurring mostly north of the Riviersonderend Mountains in the Robertson area. I did this because I saw a similarity of the plants at Stormsvlei to these Robertson Karoo elements. von Poellnitz did not make this connection and applied the name H. schuldtiana (and of course several varieties) to most of the collections from north of the Riviersonderend just as he applied the name H. triebneriana (plus several varieties) to most of the collections south of the mountains. The problem of certain identification of names is again prominent and the schuldtiana varieties minor and subtuberculata are from the same locality north of McGregor. Again H. schuldtiana was described simply as “Robertson”. The collector was G.J. Payne and he pointed out to me the true origin west of McGregor.
There is a problem in the way the name H. maraisii has been typified and then applied by me. When collected it was reported by W.R.B. Marais to have come from “weathered slate crevices on hills facing the Karoo near Swellendam”. J.R. Brown reported having received a plant from W.E. Armstrong from Stormsvlei but this is perhaps from north of the Pass. G.J. Payne showed me the locality where Marais was purported to have actually collected the plant and I assumed Payne was correct as the plants there agreed closely with the illustration in the Berlin herbarium taken to be the type. This locality is south of the Stormsvlei Pass, and also south of the river, in shale outcrop among what is now a complex of buildings. The site is barely intact but while there were plants there in 1970, I was not been able to locate them on succeeding visits, although I have subsequently done so. In my Handbooks I cited a plant from north of McGregor as “a representative specimen” but indicated that I accepted the illustration in the Berlin herbarium as the type. Breuer, whose typifications I regard as untimely and unfortunate, makes a confusing statement in his first book World of Haworthias Vol.1. Here he cites my selection of the Berlin photograph as lectotypification of the name H. maraisii but cites the locality of my representative specimen from McGregor as the original locality of that species. He also maintained that as the Berlin specimen “is dated from 1937”, it is “therefore no original material”. In his World of Haworthias Vol.2 he has changed his mind about the date of the Berlin illustration and notes it as “undated” and apparently accepts the fact that this is the valid type. However, he uses a plant from his own collection IB5926 from Swellendam to also illustrate the species. The plant IB5926 looks very similar to clones in my own collection MBB6644 (and/or 6860, populations 10 and 11 in the text below) that comes from close to Swellendam and some way from Stormsvlei. The problem for me is that I associate the population at Swellendam (there are two similar populations) with a matrix of populations and individuals (Set A to follow) that I cannot classify with any degree of certainty. Thus Breuer may have unintentionally and inadvertently generated the end-point that I want to achieve with this paper, indicating that maraisii and mirabilis are the same thing by virtue of substituting a mirabilis-like plant for a maraisii-like one. It will transpire that my decision to use the name H. maraisii also for the complex north of the Riviersonderend Mountains viz. H. schuldtiana, is an identical misstep. This is because I revisited Stormsvlei while finalizing this manuscript and fortuitously found plants again very close to the original spot. While these plants do indeed appear to be very different to H. mirabilis as it occurs within the Stormsvlei Pass, it is now quite evident on the basis of experience and re-examination that the difference is due to the geological substrate on which the populations occur. The connection with populations north of the Riviersonderend Mountains, specifically to the one at McGregor, is still true but it means that I have used a name based on an ecotype of mirabilis viz. maraisii, when it would have been more appropriate to have used the name schuldtiana.
Considerations a. The classification of von Poellnitz (1940) The basic difference between the two species as recognized by von Poellnitz can be evaluated from a key which he produced and which was published in Fedde’s Rep.43:102 (1938). The main feature or character of his section Retusae in which he placed the two species was “leaves more or less erect, with minute teeth on the margin, more rarely smooth, at the top bent back sharply and resulting in an end-area which is somewhat translucent, with a few or more green stripes, smooth, tubercled or barely with minute teeth”. In this key von Poellnitz separated H. schuldtiana (with his H. paradoxa) on the basis of the end surfaces of the leaves with small teeth and tubercles. Species with the end-surfaces tubercled only or smooth included H. mirabilis together with H. triebneriana and H. willowmorensis, which I regard as synonymous. Also included here were H. maraisii, H. sublimpidula and H. whitesloaneana. I have explained that the former can be regarded as an ecotype of H. mirabilis. This could confound my use of the name as synonymous with H. schuldtiana where the latter two belong. He also includes in his key H. paradoxa and H. magnifica but these are two elements from east of the Breede River and it will become apparent that I exclude them from this discussion on the basis of geographical consideration only. In my Revision (1999) I have actually regarded H. paradoxa as a variety of H. mirabilis while H. magnifica is upheld as a distinct species despite the obvious affinity with the two species being discussed in this work. Although he also described H. badia in 1938, von Poellnitz did not include this species in his key to the Retusae where it unquestionably belonged.
In total von Poellnitz’ key separates 23 “species”. He published a key to nine varieties of H. triebneriana (two more were added in 1940) in Feddes’ Rep. 47:1 (1939). In the same journal 49:23 (1940) he published a key to 10 varieties of H. schuldtiana that included the former H. whitesloaneana and also what I upheld as H. maculata. My observations are that it is not possible to use the characters von Poellnitz used to separate the varieties as geographic entities by virtue of the inherent variability between plants even within single population.
b. The classification of Bayer (1999) I considered that H. mirabilis and H. maraisii constituted sets on the basis of geographical distribution. Thus in the southwest the plants are larger or more substantial, with erect leaves often spotted on the back or under surfaces, and with fairly conspicuous marginal spines i.e. H. mirabilis. In the northwest the plants are smaller and darker green, the leaf tips tend to retuse and the marginal spines are reduced i.e. H. maraisii. I did recognize a color difference in that H. mirabilis was generally reddish in color with translucence in the leaf end-area, as opposed to the dark-green non-translucence of H. maraisii. In essence, von Poellnitz’ key of 1940 suggests that the two elements I recognize were distinguishable on the basis of “end surfaces with small teeth and tubercles” in his H. schuldtiana (included in my maraisii) and “end surface with tubercles only or smooth” in H. mirabilis AND in H. maraisii (excluded from my broader use of the name). This is only partly true in that while such a tendency might be there, it is not an infallible distinction.
My opinions were simply generalisations derived from a body of populations, one south and one north of the Riviersonderend Mountains. The interplay of these two species, H. mirabilis and H. maraisii, is limited by the fact that the occurrence of mirabilis is at the extreme southwest of the distribution range of the subgenus Haworthia. At its eastern limits there may be some interaction with H. rossouwii in the south, and perhaps even with H. mutica. But otherwise the interaction is with H. maraisii (as I apply the name) and the flow of that species westward and southward with H. heidelbergensis and H. floribunda. It is not my intention to examine that whole complex issue here. What I will show is that across the Riviersonderend Mountains, H. mirabilis becomes inextricably interconnected with H. maraisii and perhaps even with H. maculata. Not only that, it will appear that at Stormsvlei, where H. maraisii, and H. mirabilis(triebneriana) vars. pulchra and depauperata originated, Haworthia maraisii and H. mirabilis can be assumed to be ecotypes of the same one species because of the proximity of the habitats and the synchronization of flowering time and seed set. The only reasonable inference seems to be that the plant differences, which are essentially minimal, are due to the dramatic differences in the skeletal soils and exposed rock they are associated with.
The overriding complexity is that the core of the H. maraisii/mirabilis relationship is expressed in a small series of populations that extends from Heidelberg in the east to Riviersonderend in the west. This includes the original site for H. maraisii at Stormsvlei. It should be borne in mind that I also initially regarded H. maraisii and H. magnifica as one and the same. While I uphold them as separate species in my revision I only do so to facilitate recognition of variants in such an east/west split. I cannot explain the situation presented by populations of plants down the Duiwenhoks river which have to some degree been discussed elsewhere (see Chapter 6). The interplay of H. maraisii with other taxa is a little more complex and I will deal with the issue by examining populations as sets, either in this presentation or separately (I have written a discussion on the H. heidelbergensis/H. magnifica interaction east of Heidelberg also Chapter 6, and also a discussion of the variation and distribution of H. floribunda¸ Chapter 1).
c. Skeletal soils, geology and ecotypification I have shown (Chapter 8, Update 2) that in the E. Cape geology plays a very significant role in the way the species vary and relate to one another. My original observations that this was true were made in the W Cape and Little Karoo and I have no doubt that this is very true for the species now being considered. It is probably true for the Cape winter rainfall biomes (I use the plural only because professional botanical wisdom maintains that there are more than one) that the floras are so species rich because of skeletal soils, geological and topographical diversity and strong rainfall differences (with reliable, consistent and effective winter rain). My study area in this case is dominated by rock formations of the Cape Supergroup although in the north the Karoo Supergroup makes its appearance. The Cape Supergroup comprised three groups viz. Table Mountain Sandstone, Bokkebeld Shales and Witteberg Sandstones and these comprise again about 27 formations. These rocks are invariably directly exposed and soils are often very skeletal. Rock is almost a prerequisite for the presence of Haworthia and it is quite evident to even the most casual observer that geology and geomorphology must in some way impact on the evolutionary processes and pressures within the genus whatever they are. But while geology is no doubt significant, I will only cite it here and there as a rough pointer. This is because although the rocks in the Cape supergroup vary from pure sandstone in the Table Mountain group to mudstones in the Bokkeveld group and again sandstones in the Witteberg group, there are many variants which are obscured at the general map scale of 1:50 000. There is apparently also no general rule that a particular vegetative form will repeat itself on identical rock formation or soil.
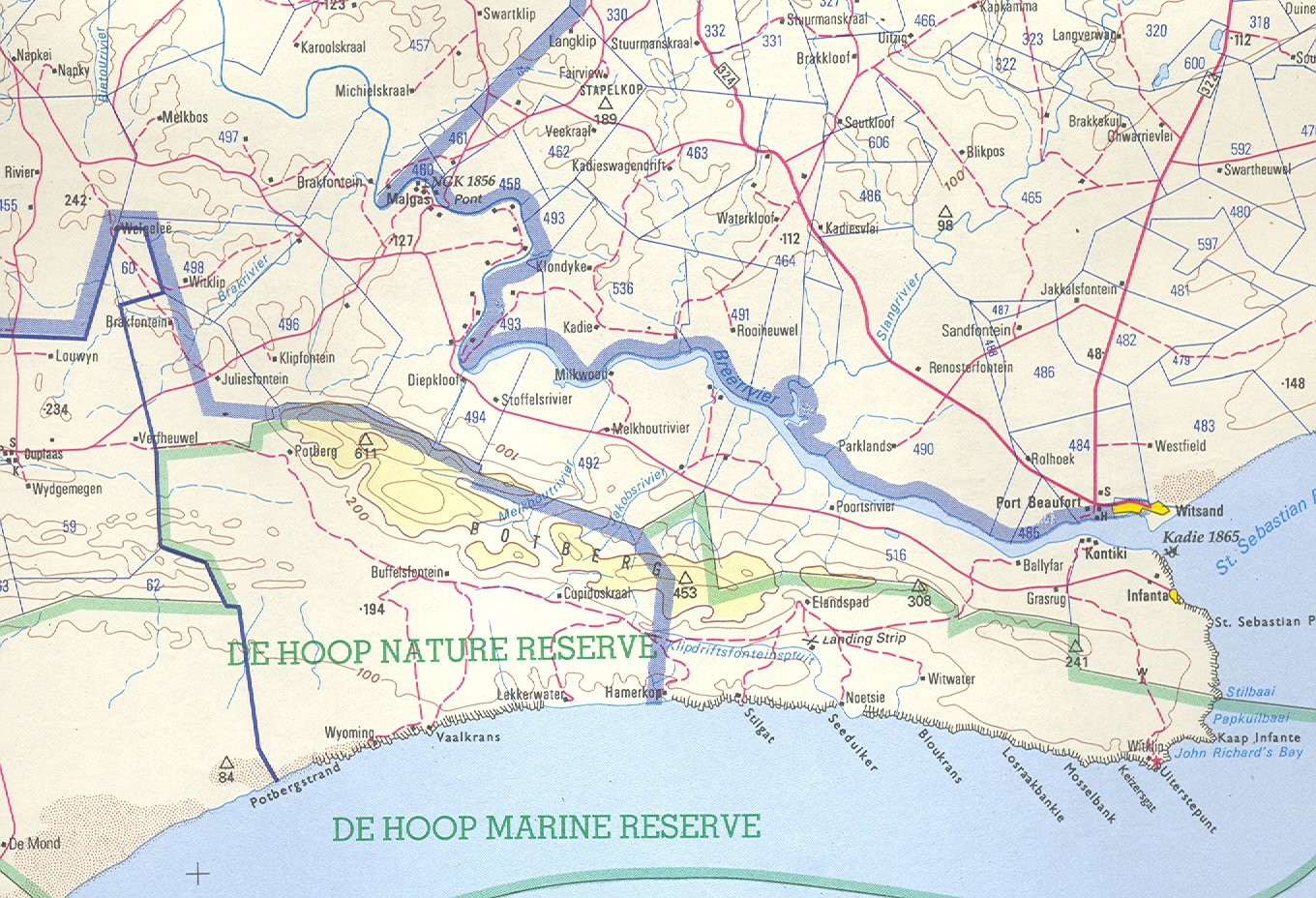
Further east in the Riversdale and Heidelberg areas, ferricrete inselbergs are significant Haworthia habitats. As can be seen from my map, the area between Swellendam and Bredasdorp where such inselbergs are abundant, there are almost no records for Haworthia. This could be due to poor exploration. On the other hand H. mutica does occur there and the void may be partly explained by virtue of the occurrences of a different species. To some extent the occurrence of an element which I refer to as H. heidelbergensis is used to fill a similar void in the south and southeast. On the accompanying distribution map, other species occurrences are thus also indicated.
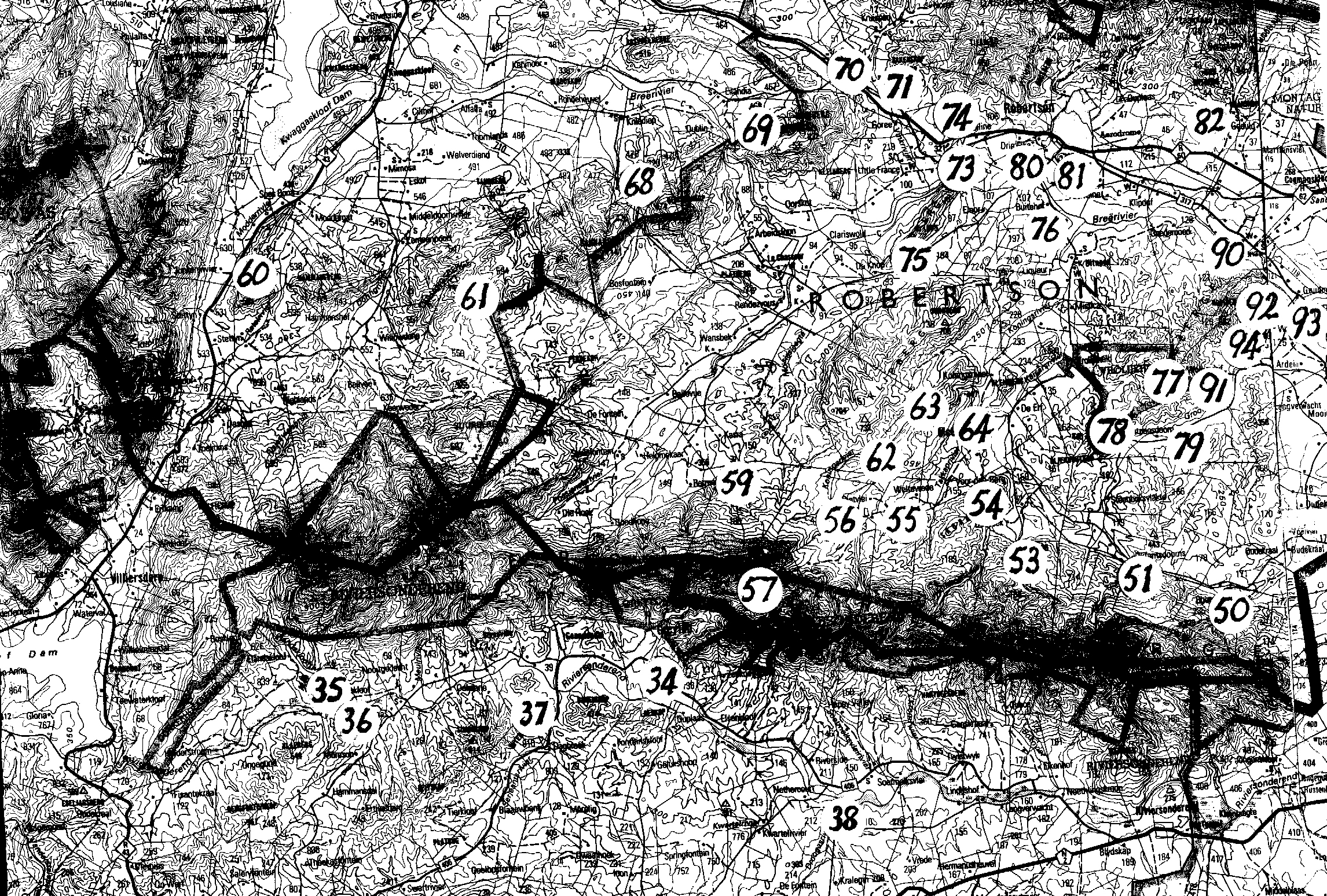
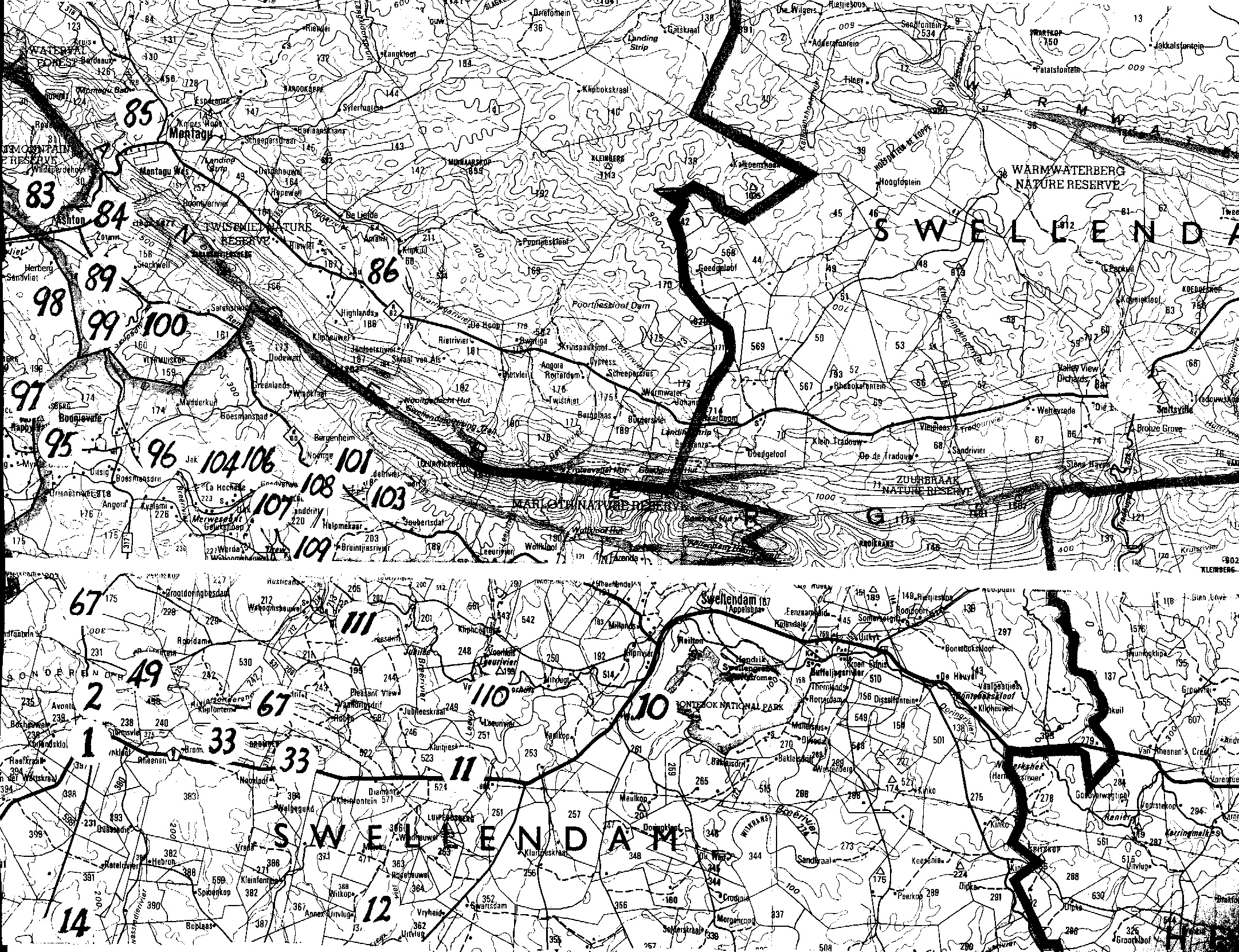
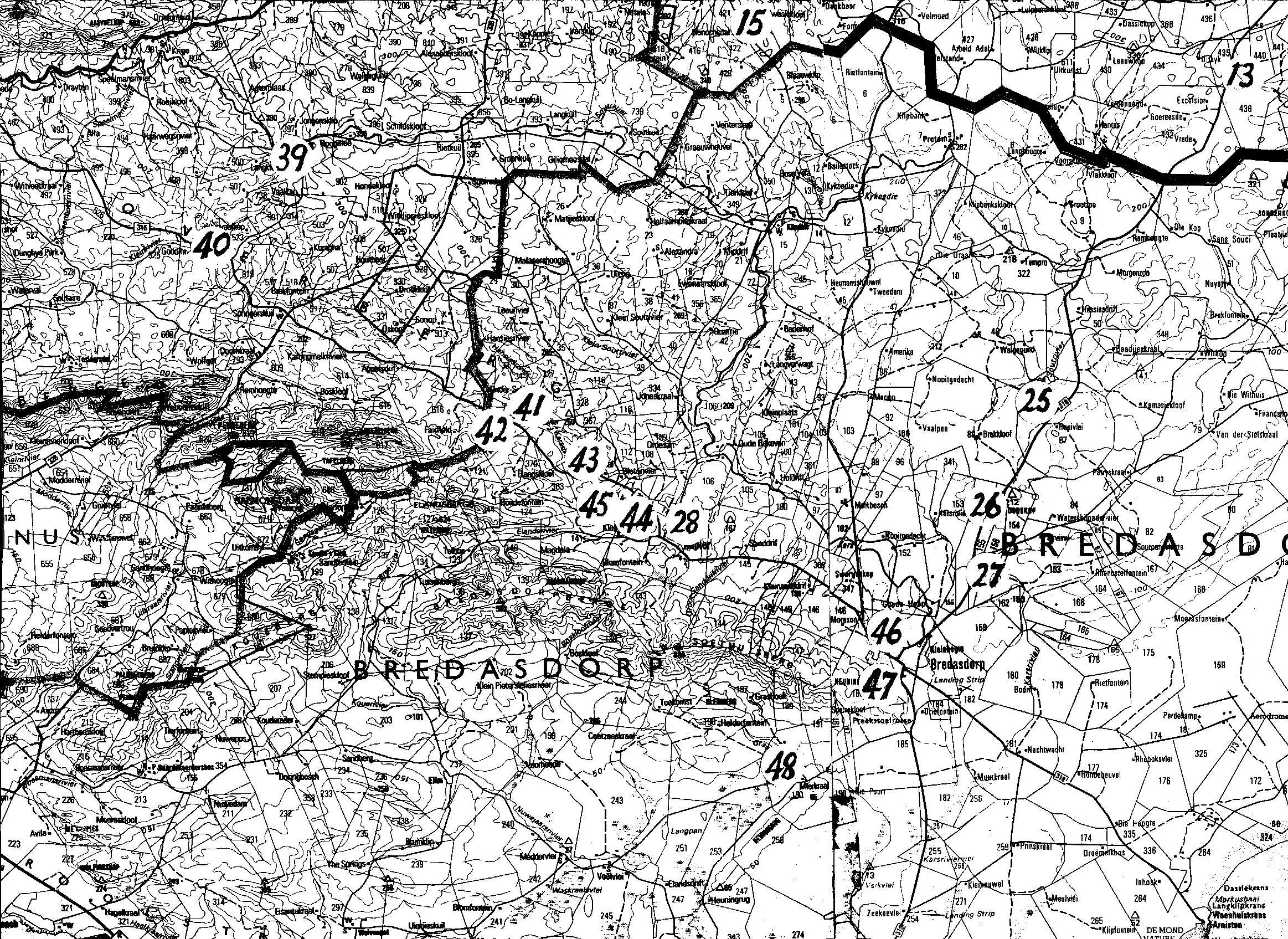
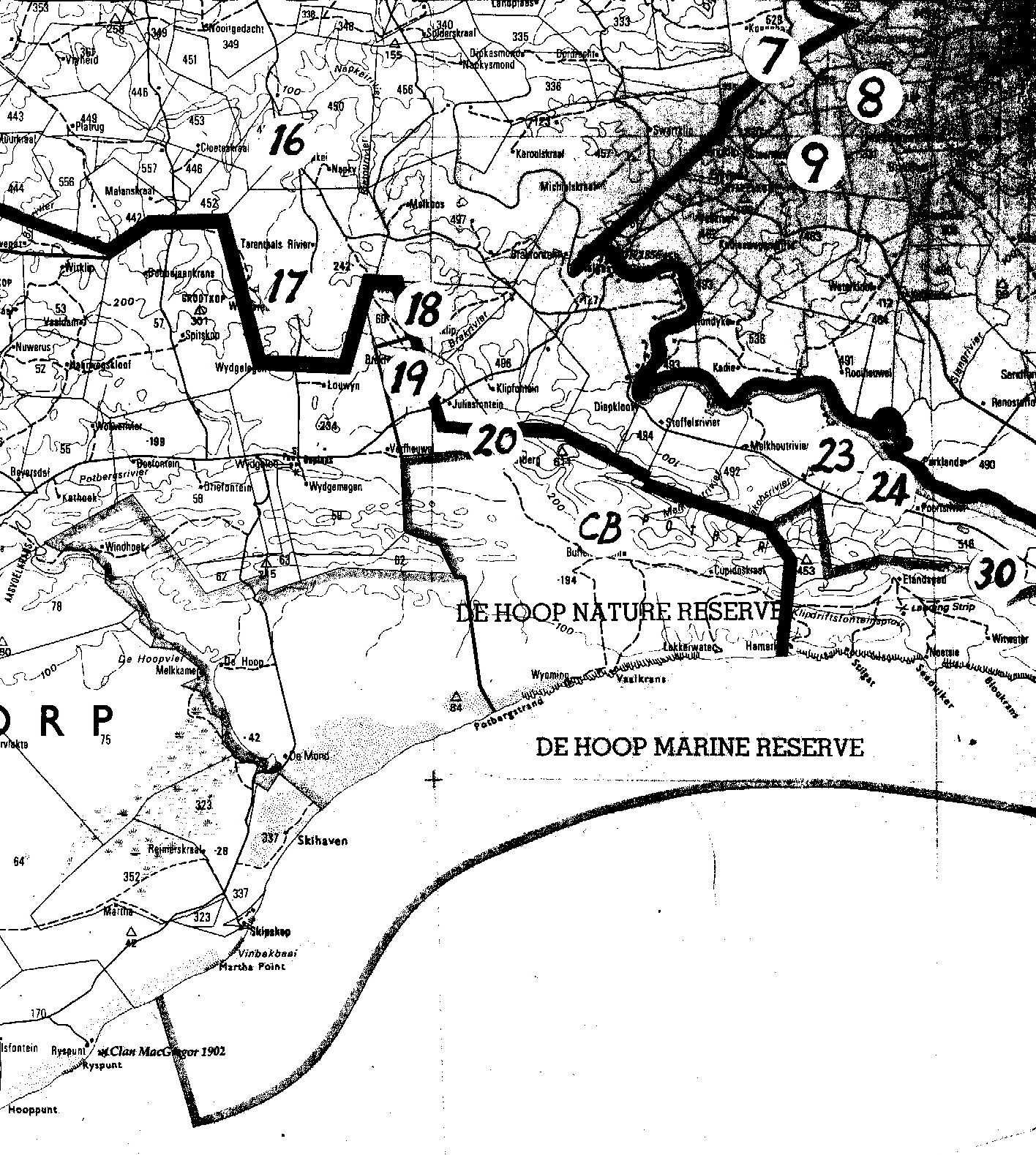
Map 1. Worcester to RobertsonMap 2. Ashton to Swellendam.Map 3. BredasdorpMap 4. E Bredasdorp.
The population sets examined The entire problem is summarized in the listing of six sets of populations (A…F). The location of these is presented on the accompanying map. Because of scale, a mapped site may indicate 2 populations and the sequential population number then omitted. The plants are illustrated and numbered in the same sequence that the populations are listed:-
A. Haworthia maraisii south and east of Riviersonderend Mountains These constitute a geographic set which occupies the southeastern range of the study area. A principle boundary is the Breede River but I have included six populations from further east of which four are off the map. Those four populations will show the integration of the sets considered here, with sets east of Swellendam. The plants are generally dark-green with reduced spination. In this area H. maraisii becomes inextricably linked with what I regard as H. heidelbergensis. That in turn has a complex interplay with H. floribunda in the southeast, with H. turgida in the east and then with H. magnifica still further east. There is a complication too in terms of a few populations along the south bank of the lower Breede River near Malgas and Infanta that is possibly an H. heidelbergensis or a H. floribunda linkage (but see set B). The distribution southwards to Bredasdorp (Rooivlei and Adoonskop) and then westwards to Napier may be in an integrated state with H. floribunda (i.e. neither one species nor the other). At Napier the appearance of the plants still does tend to H. maraisii, so that we have a population that appears to be this discrete species juxtaposed with H. mirabilis var. badia. They have the same flowering time and based on the westward changes in appearance of badia, it may actually be a product of “hybridization” or “introgression” of H. mirabilis and H. maraisii. Thus a ring-cline may exist, and what appears to be two distinct species could be due to the different routes taken in space and time to a point where they grow in close proximity and are interbreeding or have interbred. The southwestern forms of H. maraisii may be a composite of this species with H. mirabilis but also involving H. heidelbergensis and H. floribunda. H. rossouwii could also come into the reckoning, except that this is a spring-flowering species like H. mutica, as opposed to late-summer flowering as is the mirabilis/maraisii complex.
(Note:- the item numbers are used to indicate map position – if space does not permit, a site can be assumed to be in close proximity to the number given in squared brackets).
Stormsvlei MBB7257. This is from the original site shown to me by Payne where I thought the plants had been destroyed. I found about 70 plants on a much degraded site where a wattle tree had just been removed from the top of a small outcrop of Bokkeveld shale. The site is only about 50m away from the point shown to me by Payne but the shale ridge is so degraded that it is unlikely that more plants exist there. The plants were very exposed and burnt by the sun and it was only under the dense shade of a plant of Galenia africanathat I found three dark-green plants that I could comfortably equate with my original opinion that it closely resembled plants from north of McGregor. From the point of view of description and typification, it is important to note from the pictures that the leaf surfaces are with spines and thus do not match von Poellnitz’ description. Not all the plants were with retused leaves either. The species Galenia africana is a vegetation pioneer and an indicator of disturbance.
Stormsvlei MBB6954, JDV96/19. The plants illustrated are from north of the river in quarternary river boulders [1]. In situ they have strongly retused leaves and I use the word “retuse” under the impression that an original meaning of “retuse” was to indicate “bent back like a thumb”. The impression in the literature is that such retused leaves have a distinct end-area. This is not so. The leaves of many species in the southern Cape, including H. herbacea, H. reticulata, and H. rossouwii which are characterized by having rounded rosettes with incurving leaf-tips, are swollen convexly on the inner lower surface. This convexity can be emphasized to the extent that it manifests as an end-area. In addition to this variation, the leaf end may be either incurved or recurved. The original plants of H. maraisii from exposed vertical shale south of the river were strongly retuse and this character was to my knowledge maintained in the plants that were once in my possession at the Karoo garden. Plants from this site north of the river are retused in habitat but not always so in cultivation. Thus they become very similar in appearance to plants in the sandstone ridges within the pass viz. H. mirabilis var depauperata [2]
SW Heidelberg MBB6663, JDV97/35. The plants are very similar to collections from Swellendam (10, 11) and from Rondeheuwel (14) and Verdwaalskloof (15). The habitat geology is Enon conglomerate, as is 10, 11, 14 and 15 are on Bokkeveld shale.
Morning Star MBB7218, MBB7220, Witheuwel MBB7226, Somona MBB7229. It is difficult to clearly associate these two populations with the sets under discussion here. The problem is that while it seems obvious that H. magnifica and H. maraisii are conspecific, the truth is that east of Heidelberg H. magnifica is demonstrably continuous with H. heidelbergensis. I have expressed the opinion that 3 and 4 may also be influenced by H. floribunda.
Andrieskraal MBB7237, MBB7238, Skeiding MBB7239. These three populations are obviously related to H. magnifica and I should mention that H. magnifica var. atrofusca is a problem because its real position taxonomically may be dominant to the typical variety. Thus these Skeiding plants are closer to the var. atrofusca (with obtuse leaf-tips) than they are to the typical var. magnifica (with an end bristle). The geological substrate is ferricrete as at Spitzkop, Heidelberg (6). While I have stated the relationship to H. magnifica, it must be remembered that this species should be seen as an extension of the problem being discussed in this paper. (Spitzkop is also the name for the type locality of H. magnifica var. atrofusca at Riversdale).
Spitzkop (Heidelberg) JDV89/2. This population could also be considered to be maraisii-like but the leaf surfaces are inclined to be finely tubercled without the spine-like surface armature commoner in H. maraisii. But this is also not diagnostic, because there are magnifica-like populations where the individuals are with either finely tubercled surfaces or with very pronounced tubercles and/or surface spination. Descriptive detail is not adequate to make any distinction between the populations 5 and 6 and H. magnifica further east.
Koppies MBB6879. Here the plants show floribunda-like character and this is more evident in the next (8). H. rossouwii is present nearby.
S Oudekraal MBB6881. H. rossouwii is also present about 1km to the north. These plants are even more floribunda-like and I have included some of these populations in a resume of H. floribunda.
Stuurmanskraal J.Esterhuizen in JDV93/64. This population is geographically nearer to the Breede River and to Malgas and the plants have a similarity to that south and west of Malgas.
SW Swellendam MBB6644, MBB6860. The plants tend to have erect or sub-erect leaves. The leaf ends are generally not retused and the tips are often slender and floribunda-like.
W Swellendam MBB6861. These Swellendam populations mirror that at Heidelberg (3) and the plant form repeats at Rondeheuwel (14) and at Verdwaalskloof (15) to the west. I have always regarded the Rondeheuwel population as indeterminate between H. mirabilis and H. maraisii and considered that it might be hybrid. However, I have with time become increasingly skeptical that hybridization accounts for problem variability. My impression is that the sequence from Heidelberg through Swellendam to Rondeheuwel and Verdwaalskloof support the contention that there is genetic continuity that is not a product of hybridization. The similarity suggests to me that this may be archetypal for the populations in the five sets that I am dealing with here.
12. Witkop/Uitvlugt MBB7242, MBB7243. These two populations are on ferricrete inselbergs that are a conspicuous feature of the landscape between Stormsvlei and the Potberg in the southeast. Together with the next two populations the plants link Stormsvlei to the Potberg mélange that I think may relate to H. floribunda as well as to H. heidelbergensis as I already have alluded.
Die Kop MBB7244, MBB7245. (Not Die Kop Wydgelee MBB7500) These are south of the two former collections and are in the same mould. I treat them separately because the one collection is on a white-clay pressure burst and the plants are less scabrid than in the other collection that is on reddish pebbly soil. Both sites are on ferricrete.
Rondeheuwel MBB6862, JDV90/32, JDV96/43. My first impression of this population about 1972 was that it was hybrid between H. mirabilis and H. maraisii. Yet even then I was expecting some king of continuity between these two “species” other than hybridization.
Verdwaalskloof MBB7074. This site is near to Riviersonderend and is one of the strongest links between the series in Set C that could perhaps comprise a separate H. mirabilis.
Napky MBB7030, JDV96/18. This is near the origin of the von Poellnitz H. maraisii var. simplicior. It is now south and west of the Breede River and there is an element of floribunda in the plants viz. a more elongated leaf with flattening and rounding of the leaf-tip and the slight twist evident in that species (see 7 & 8, and also true for 3, 9 and 10). It should be noted that in my treatment of H. floribunda (Umdaus Press, in ms) I suggest that this may be a juvenile character perpetuated in that species.
Tarentaal MBB 6539. One of the few populations that transgresses the tendency to a geographic drift in similarity of populations. (Some of…)The plants in this particular population are very similar to robust forms of H. magnifica var. atrofusca as at Riversdale.
Witklipkop MBB6890. This and the following four populations are problematic and further populations occurring along the lower Breede River exacerbate the situation. They need to be included here but at the same time they are clearly connected to a different set of populations that I relate to H. heidelbergensis. The leaves are smaller, more numerous, more elongate and generally without an end-area.
Brakfontein MBB6886.
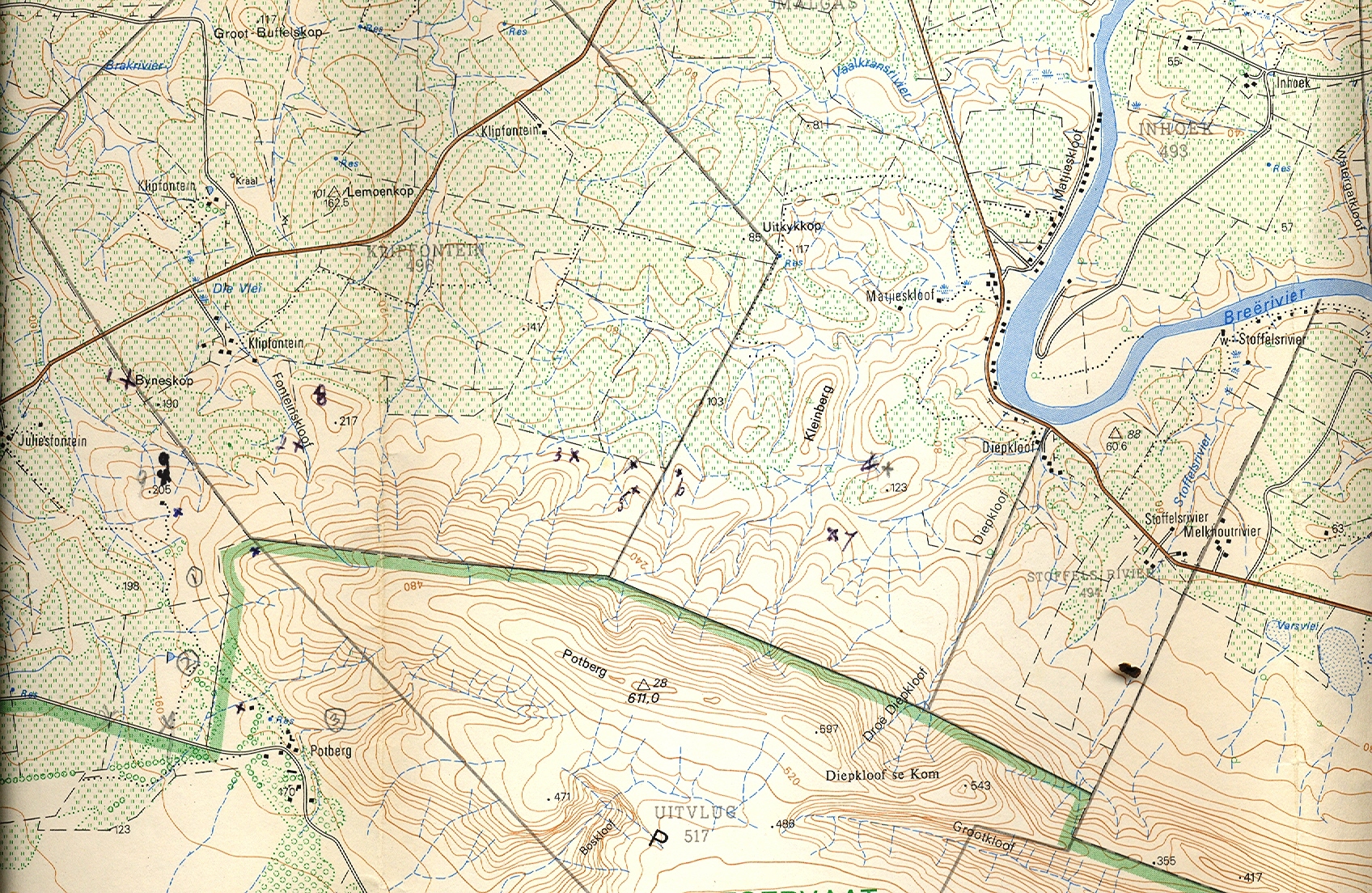
Juliesfontein MBB6882. In my Revision I cite an early collection of mine MBB1211 from here under H. maraisii. I also included a collection of P.V. Bruyns from Ziekenhuis (see 24) because the plants were very small and seemed similar. There are good herbarium specimens to show this. However, when I re-collected at Juliesfontein after the publication of my Revision in 1999, there were no plants at my remembered site and much larger plants nearby which I would have had difficulty putting with H. maraisii. The vast mass of material I have seen since has not made a taxonomic decision any easier. I can honestly say that recognizing a taxonomic difference between these two populations (20 and 24) would serve a very limited purpose.
N Potberg MBB6889 [20]. Chris Burgers (CB2506) first sampled this population and I also cited it under H. maraisii. This is not a comfortable solution.
NW Potberg MBB6544, JDV96/54 [20]. H. variegata occurs here too and may infuse populations in the area. I have one record of plants that seem to suggest this, but this would need to be covered in a discussion of H. variegata itself.
Jakobsrivier JDV92/85, JDV87/50. Needs to be considered with the following. They are very small plants with blunt and rounded leaf-tips suggestive of the Potberg collections 18 –22.
Ziekenhuis P.V. Bruyns in JDV87/5, Uitrus MBB7250. The first is a collection that P.V Bruyns made east of the river. These are very small plants with erect leaves much as I remembered the original Juliesfontein collection of mine. They flower in January as this whole complex generally does. The geology of the east bank of the river is Bokkeveld shale. At Uitrus it is rather uncertain as while there are exposed shales in the vicinity they are overlaid by pebbly deposits of recent origin. This may be very important in relation to the juxtaposition of this site (and 22) with Set B.
Rooivlei MBB6638. The following five populations are a continuance of the Napky (16) one where there seems to be a floribunda impact and the plants are also very dark green often conspicuously tubercled. The leaves are thick and blunt. An element also occurs here which is apparently H. heidelbergensis, and H. turgida is also present. The presence of a separate heidelbergensis-like population is confounding. Those plants flower a little earlier and should be discussed with plants from Kathoek, Beyersdal, Haarwegskloof. They cannot be excluded from a total concept of H. mirabilis that I am suggesting here, and as I am going to do for the same reason that this conspectus excludes populations from beyond Heidelberg.
Adoonskop MBB6640. These are like the Adoonskop and Napier variants (27) and easily put with a concept of H. maraisii if it were feasible to separate such an element from H. mirabilis in the overall relationship of the populations. The plants tend to have rounded leaf-tips as is the case in the following four populations.
NE Bredasdorp JDVsn – no specimens collected. Plants as above. This would have been an important record because of the proximity to Bredasdorp and to H. mirabilis as the var. sublineata. The site was also home to Aloe brevifolia and sadly the site is so degraded that neither Haworthia nor Aloe can now be found there. MBB7334 is a very recent collection, nearer o Bredasdorp and plants are very similar to (29)
Napier MBB7259, JDV85/15. There is a substrate difference in that at this site the soil is pebbly quartzitic whereas at the following site nearby (29) it is finely slabbed, coarse grained sandstone. MBB7259 is about 4km north and the substrate there is true micaceous shale.
Napier JDV85/96, MBB7247 [28]. Whereas plants at the former sites are blackish-green, here the plants are reddish-tinged.
A01 MBB7257.1 StormsvleiA02 JDV96-19.1 N StormsvleiA03 JDV97-35.1 SW HeidelbergA03 MBB6663.1 SW HeidelbergA04 MBB7218.1 Morning StarA04 MBB7220.1 DuiwenhoksA04 MBB7226.1 WitheuwelA04 MBB7229.1 SomonaA05 MBB7237.1 AndrieskraalA05 MBB7239 SkeidingA06 JDV89-2.2 Spitzkop, HeidelbergA07 MBB6879.1 KoppiesA08 MBB6881.1 S OudekraalA09 JDV93-64.1 StuurmanskraalA10 MBB6644.1 SW SwellendamA12 MBB7242.1 WitkopA12 MBB7243.1 UitvlugtA13 MBB7244.1 SE Die KopA14 JDV90-32.1 RondeheuwelA14 MBB6862.1 RondeheuwelA15 MBB7074.2 VerdwaalkloofA16 JDV96-18.1 NapkyA16 MBB7030.1 NapkyA17 MBB6539.1 TarentaalA18 MBB6890.1 WitklipkopA19 MBB6886.1 BrakfonteinA20 MBB6882.1 JuliesfonteinA21 MBB6889.1 N PotbergA22 JDV96-54.1 NW PotbergA22 MBB6544.1 NW PotbergA23 JDV92-85.1 JakobsriverA24 JDV87-5.1 ZiekenhuisA24 MBB7250.1 UitrusA25 MBB6638.1 RooivleiA27 MBB7334.2 H. maraisii. NE Bredasdorp.A28 JDV85-15.1 N NapierA28 MBB7259.1 Napier StationA29 JDV85-96.1 NE NapierA29 MBB7247.1 H. maraisii. NE Napier.
B. Haworthia mirabilis towards the Potberg This set is actually anomalous and consists of only three populations associated with Table Mountain Sandstone and calcareous limestone. At Karsriver, a short distance from 27 is where one finds H. rossouwii var. petrophila (which I had previously allied to H. variegata) in limestone. Further east is H. variegata var modesta and to the south of that H. rossouwii var. calcarea (previously as a variety of H. mirabilis). It is obvious that the classification is difficult and dubious. However, C. Burgers made another surprising collection (CB2018) from Buffelsfontein [CB] south of the Potberg. I cited it in my revision under H. mirabilis var. calcarea. I have had a problem with a specimen collected by Prof. H. Compton cited “from the Potberg sandy flats”. That specimen is of a retusa-like plant that I thought might perhaps be representing either H. mutica or H. mirabilis but I did not cite the specimen in my revision because I was so doubtful about it. I thought this could have been what Burgers found, but his collection was in rock crevices and not on the sandy flats. However, Adam Harrower made a collection (ADH594) from 20km further east, also in rock crevices and clearly the same as the Burger collection. On a very recent trip my wife and I found a similar plant (31 MBB7248) on the northeast end of the Potberg that I think may fit here. The problem is its proximity to 24 only 3km away.
Sandhoogte ADH594, MBB7251. The site is a most unprepossessing and unlikely looking one from a distance. The vegetation is dense waist-high fynbos, and Adam Harrower found the plants in the rocky north-facing border to a stream. They can be compared with H. turgida var. longibracteata (except they are less proliferous) and also with H. mirabilis var. sublineata. There is also a likeness to forms of H. rossouwii var. calcarea, albeit distant. More striking is a curious resemblance to H. magnifica var. splendens from near the Gouritz River Bridge.
Ballyfar MBB7248, MBB7249 [off map E of 22]. These two populations are only a few kilometers from 23 and 24, but they are on sandstone. What is more is that the plants are larger but have fewer leaves that are strongly retused as in the Compton specimen. What is striking about the plants is the presence of numerous surface spines and one is forced to consider the resemblance to a population of H. magnifica east of Riversdale with similar spination and general appearance, only proliferous. The site is about 3km due east of the Sandhoogte population and this suggests that the two populations are of the same species. There is an interesting dimension to the problem mirrored in the preceding (30). This is the occurrence of H. turgida at Diepkloof only 15km to the northwest and only five kilometers from Burgers 2018 at Buffelsfontein across the Potberg Mountain. A decisive factor may be flowering time, because H. turgida flowers in spring and the other populations considered here flower in mid-summer. But the main point is that the relationships are complex and varied.
B30 MBB7251.1 Sandhoogte De HoopB31 MBB7248.3 Ballyfar
C. Haworthia mirabilis and its variation south of the Riviersonderend Mountains This set also starts with Stormsvlei and is the south and westward set which is almost exclusively mirabilis. There is a possibility of a connection across the Riviersonderend Mountains closer to Villiersdorp where it may be suggested that H. maculata is implicated too, but this will instead be considered in Set 3. These are plants with a tendency to redden. They are usually larger than those in set 1. The height may be as much as 100cm and rosette diameter 110mm.
Stormsvlei Pass MBB6955. This population is in sandstone and immediately there is a significant difference to the plants immediately south (1) and north (2) of the Riviersonderend River a short distance to the south. There is no evidence of an end-area to the leaves. The leaves are longer and more slender and they show the redder colour one associates with H. mirabilis elsewhere. Some of the plants are very highly spotted on the back or under surfaces. This is a character that has been associated with H. mirabilis when in fact quite common to find plants with or without such marking in many of the populations in these sets.
E Stormsvlei Pass E. Aslander sn. MBB7252, MBB7253 and MBB7254. In exploring the southern slopes of the Bromberg I came across several populations. The geology here is rather complex as it is the close interface of sandstone and shale, with the pebbly patches associated with ferricrete (Gibbaeum austricolum is present). At one site (MBB7254) the plants were small and with narrow elongate leaves on a coarse-grained sandstone ridge, while immediately north on a pebbly band there were plants with fewer, shorter and broader leaves. There is no difficulty in allying these plants with 32.
Greyton JDV96/47. The continuity westwards is disrupted by lack of records and there does not seem to be suitable habitat between the Riviersonderend River and the mountains. A population at Lindashof recorded by G.G. Smith appears to have been destroyed in a road-making operation. This population at Greyton is of rather small plants with conspicuously finer and intense spination.
Uitkyk MBB7092, JDV95/97. This is in the extreme west for the set, at Elandskloof west of Genadendal. It is a very rocky south-facing site and the plants are robust and proliferous.
Leeukop JDV90/34, JDV85/98. This is the locality for the var. rubrodentata where the plants have long slender leaves with intensely red spines – hence the name.
Genadendal JDV90/33.
Skuitsberg JDV88/70, JDV96/101. Here there is quite an extensive population with a similar one also present on the adjoining farm Nethercourt. The plants are generally very robust and were described originally as H. emelyae var. beukmannii.
Jongensklip MBB7059. The interesting thing about this population is that it fills somewhat of a void between populations known respectively north and south of the Caledon/Bredasdorp wheatlands.
Goudini MBB6537. This population is the southwestern-most for the species. I have grown it from seed and the variability among the seedlings is quite striking, which confirms my opinion that the taxa described from north of Napier are not significant as discrete varieties.
Schietpad MBB4642. JDV94/90. See 43.
Fairfield MBB6643. See 43, except that the plants are very proliferous.
Mierkraal (Napier) MBB7091, S Mierkraal MBB7260. The plants now constitute a general form that appears in the preceding three populations and there are several such populations in the area between Mierkraal and Fairfield (42) and I do know that I have seen it in the Klipdale area to the northeast. The varieties H. triebneriana vars. multituberculata, napierensis (Schietpad, 41) and turgida, originated north of Napier and yet I would not consider the variability to be population based nor even so significant in terms of the variability of H. mirabilis var. triebneriana as I would designate the taxon. MBB7259 is a population south of Mierkraal, the plants tend to have more elongate and erect leaves. The site is geographically nearer to 40 (MBB6987) which is H. mirabilis var. badia.
NW Napier MBB6635. Although the geology is the Bokkeveld group, the rock here is very quartzitic and the vegetation alters from Cape Fynos to renosterveld over a very short distance. The site is that of H. mirabilis var. badia. The geology in this area is quite complex and there is also a small ferricrete inselberg to the east.
W Napier MBB6987. This population is a western continuation of the previous one and is also the var badia. However, there is a tendency for the leaves to be more erect and narrower. I do speculate that this var badia may have arisen from an interaction of H. mirabilis and H. maraisii. The four populations (28 & 29 of set A, 44 & 45) flower and seed at the same time, seedlings of both are highly variable and both have companion populations which suggest the deduction of continuity across a very strong geological but weak breeding barrier.
Bredasdorp N JDV86/8, MBBsn, MBB7261sn. This locality is immediately north of the town on the riverbank. The area was highly invaded by alien Acacia and the haworthia are no longer there. The plants are somewhat similar to those south of the town, but more robust and with more strongly retused leaves. This is a close approach to H. maraisii (26) and may parallel the situation at Napier (28 & 29) where this species and H. mirabilis var. badia (44 & 45) are almost sympatric. My MBB7261 are a few plants rescues from about 300m northwest of JDV86/8 from a site overgrown with bluegum. These plants had erect leaves.
Bredasdorp S MBB6639. The site is a north-facing sandstone mountain slope. The plants are highly variable and are named as H. mirabilis var. sublineata [47]. The leaves are long and slender. Some clones are proliferous and others not.
Mierkraal (Bredasdorp) MBB7090. H. mundula was described from this locality which is unusual in being recent sandstone and on the very southern fringe of the distribution of H. mirabilis. The plants are very proliferous and their leaves have a very pronounced end-area that is also smooth. The population is thus fairly different from others and this is unfortunate because it is in my opinion the source of the type and hence is the var. mirabilis. I first saw the population in 1969 and can testify to significant changes in the precise whereabouts of large proliferous mats in the past, as opposed to more scattered less proliferous clumps in the present.
C32 MBB6955.1 Stormsvlei PassC33 MBB7252.1 NoukloofC33 MBB7254.1 E StormsvleiC34 JDV96-47.1 S GreytonC35 JDV97-94.1 UitkykC35 MBB7092.1 UitkykC36 JDV85-98.1 S GreytonC37 JDV90-33.1 W GenadendalC38 JDV88-70 SkuitsbergC38 JDV96-101 SkuitsbergC39 MBB7059.1 JongensklipC40 MBB6537.1 GoudinieC41 JDV94-90 SchietpadC41 MBB4642 SchietpadC42 MBB6643.1 FairfieldC43 MBB7091.1 Mierkraal, NapierC44 MBB6635.5 NW NapierC45 MBB6987.1 W NapierC46 JDV86-8.1 N BredasdorpC46 MBB7261.1 NW BredasdorpC47 MBB6639.1 S BredasdorpC48 MBB7090.1 Mierkraal, Bredasdorp
D. Populations north of the Riviersonderend Mountains Stormsvlei is where the Riviersonderend Mountains begin to tail away to the lowland wheatfields. The Riversonderend River cuts through the mountain about 10km east of Stormsvlei, and the Stormsvlei Pass itself is a rift through the mountain tail that allows road access from north to south. The mountain to the west is Table Mountain Sandstone. Rainfall is high and the vegetation is Cape fynbos. It must be a formidable barrier to genetic flow and unsuitable habitat for succulent plants. There are very strong geological gradients coupled with precipitation differences north and south of the mountain. The geological gradients also have an east-west pattern as opposed to the essentially north-south pattern south of the mountain. Further to the west, folding and faulting have resulted in a huge wedge of more recent formations appearing between Worcester and Villiersdorp. McGregor is virtually at the southern end of a wedge of older Bokkeveld formations that run northwestward to Robertson. There it meets the great Worcester fault line and a massive igneous pluton. Rocks of the older Malmesbury Supergroup and rocks of the much younger Karoo Supergroup are juxtaposed with igneous intrusive rock. The plants dance to this pattern and this is reflected in the differences found in Haworthia populations. This set comprises the populations on the Riviersonderend Mountains itself and along the northern foothills. The plants are intermediate between sets C and D and include significant ecotypes, as for example the Boesmanspad plants that resemble plants in the H. turgida complex, the Moddergat plants associated with H. maculata and the Trappieskraalkloof plants which are the maraisii-like element.
NE Stormsvlei Pass (Brakfontein) MBB7087. This is on a dry north-facing slope where the vegetation is Fynbos and close to the transition to Renosterbos. The geology seems to be recent scree and sand. The plants have erect to sub-erect leaves, are quite narrow and have strong marginal spination. They thus seem to equate with plants within the Pass (32) in quartzitic sandstone of the Table Mountain Group and seem to differ from plants close-by to the northwest (67, MBB7086) that are on a shale/sandstone formation of the Bokkeveld Group.
Boesmansrivier MBB6711. The Boesmansrivier Valley is a very long and deep incision into the mountain running southwest to northeast. The plants are similar to Stormsvlei Pass (32) but the undersurfaces are often more strongly spotted and the faces of the leaves are frequently also very attractively spotted with translucent dots.
E Olifantsdoornkloof MBB6686. The habitat here is slightly drier and more exposed than the preceding and the plants have shorter squatter leaves.
Olifantsdoornkloof MBB6685 [51]. At this site there is again a deep incision into the mountain and the habitat is more sheltered and moister. The plants have longer leaves. von Poellnitz named H. triebeneriana var. diversicolor from this population and the name refers to his observation “plants therefore appear diseased and no plant agrees fully in colour with the other”. It is instructive to think what he would have thought if he had seen plants from the preceding populations extending through to the following 15 in this set.
Hoeksrivier MBB6575. What is surprising and different about this population, also in a rift into the mountain, is that the plants have the rosette shape of H. herbacea. Although the leaves have the swollen mid-section, the ends are curved inward so that the rosette is more rounded. These plants are particularly “diseased” as in 52.
Houtbaaiskloof MBB6760, JDV97/163, MBB7241. Houtbaaiskloof is a very deep incision into the mountain immediately south of McGregor, which meets a geological fault line north of the Hoekrivier. So I was able to make collections both north and south in this pseudo-gorge. This is where von Poellnitz’ H. whitesloaneana originated. The leaves revert to the tendency to have an end-area but the “diseased” appearance is reminiscent of 52.
Takkap MBB6723, JDV89/63. In this incision of the Takkap River into the mountain the plants become increasingly mottled and also slightly softer in texture coupled with increasing leaf numbers although they are still small in size.
Dwarswaterkloof MBB6865, JDV89/67. As with the Takkap plants (55) the plants are greener and quite proliferous. The Dwarwaterkloof is again a very long and deep incision into the higher peaks of the Riviersonderend Mountains. It is not surprising that the plants have continuity with the following populations.
Boesmanspad JDV84/67, JDV88/63, JDV89/67. The Boesmanspad refers to a footpath that traverses the Riviersonderend from McGregor to Greyton. There is a deep valley called Boesmanskloof that runs from Greyton in the south towards McGregor. It is a high rainfall area and it is quite surprising to find Haworthia in this gorge. Furthermore the plants are small and very turgid so that the comparison can be fairly drawn with that of plants of H. turgida var. turgida from similar wet and rocky situations in the Langeberg mountains e.g. Southey’s Pass or Garcia near Riversdale. The flowering time and flower itself in the context of distribution and flow in similarity suggests that this must be an ecotype related to populations both north and south of the mountain range.
Koeniesriver MBB6714 [56]. These are in a more exposed situation than the previous two and with the following (59) are again more reminiscent of plants from the Stormsvlei Pass itself (32).
Rietvleikloof MBB6720. The topography of this mountain incision is interesting because it is practically the last of a north/south series. The next incision is a major faultline that runs southwest/northeast. This forms a valley largely in the lee of high mountain (Jonaskop).
Moddergat JDV96/34, MBB7270. This population needs to be carefully considered. It oversteps the distribution of H. herbacea and the var. livida that I described from just east at Lemoenpoort. I also described H. herbacea var. lupula from an improbable habitat deep in the mountains east of Villiersdorp. I associate the Moddergat plants with H. maculata to the north at Brandvlei and this has still further associations in the Langeberg Mountains. However, the flowering time and appearance of the plants do suggest that a connection with H. mirabilis has to be considered even if improbable. I have indicated already that this area is geologically diverse.
Trappieskraalkloof MBB1210, Droeriviersberge MBB7263, MBB7264, Haumanskloof MBB7265. The first I considered from memory to be a very significant population that could better be placed in Set E. The plants are small, greyish in colour. The rosette is somewhat globular with short retused end-areas and very similar to 65. I initially made this collection MBB1210 in my first few years at Worcester, and it is only at the close of this work that I have revisited the area. The area is difficult to access and is very dry. The terrain is extremely rocky and rugged. Dywka Tillite interfaces with Witteberg Quartzite and the vegetation can change dramatically over a distance of a few meters. It is not clear how the latter three populations relate to the traditional perceptions of H. maraisii and H. mirabilis as discrete species, and nor to their relation with H. maculata, which is not far to the west at Moddergat (see 60 JDV96/34. MBB7270). It is evident that they are connective between sets D and E.
Whipstock MBB6864. Again one has to take geology into account as I discussed for the set. At Moddergat and Trappieskraalkloof one is confronted with more recent geological phenomena. Whipstock is at the southern end of a similar quartzitic formation and the plants follow the trend indicated in the early items of this set. The plants are somewhat mottled and an end-area is often apparent.
Klipbergdam MBB6681, MBB6682, JDV97/48. These plants demonstrate continuity between the early members of this set and the following set.
W McGregor MBB6673, JDV96/49. This is in the McGregor Nature Reserve and is the source of the original H. schuldtiana as was conveyed to me by G.J. Payne.
N McGregor MBB6646 [64]. This population is the origin of the names H. schuldtiana vars. minor, subtuberculata and unilineata. Obviously the population seems to be unusually variable because of the varieties that it has given rise to. But this is not so and simply reflects the weak taxonomic precepts of the time which seem to be so difficult to dispel in the present.
SE McGregor MBB6687 [64]. The plant I illustrate appears to have very pronounced end-areas to the leaves. But I have not explored the area thoroughly as the plant fits conformably with the populations I am listing.
NW Stormsvlei Pass MBB7086, & Vaandrigdrift MBB6534, JDV96/25. I have often observed plants northwest of Stormsvlei Pass and simply accepted that they belonged to the discrete species I had conceptualized viz H. maraisii. They now easily fit the conceptual model that the elements around Stormsvlei and the Pass are actually one and the same species. I have not collected at the latter two sites further to the east. Plants I observed there would have fitted those illustrated in 61, 64, 65 and 66 as well as with many in set E. This would correspond to a concept of the element H. maraisii as I recognized it in my Revision.
D49 MBB7087.1 NE StormsvleiD50 MBB6711.1 BoesmansrivierD51 MBB6686.1 E OlifantsdoornkloofD52 MBB6685.1 OlifantsdoornkloofD53 MBB6575.2 HoeksrivierD54 JDV97-153.1 HoutbaaiskloofD54 MBB7241.1 N HoutbaaiskloofD55 MBB6723.3 TakkapD56 MBB6865.1 DwarswaterD57 JDV84-67.1 BoesmanspadD58 MBB6714.1 KoeniesriverD59 MBB6720.1 RietvleikloofD60 JDV96-34.1 ModdergatD60 MBB7270.1 ModdergatD61 MBB7263.1 DroerivierbergD61 MBB7266.2 DroerivierbergD62 MBB6864.1 WhipstockD63 JDV97-38.1 KlipbergdamD64 JDV97-49.1 W McGregorD65 MBB6646.1 N McGregorD66 MBB6687.1 SE McGregorD67 MBB7086.1 N Stormsvlei Pass
E. Haworthia maraisii north of Riviersonderend Mountains This is the Worcester/Robertson Karoo set that I have simplified by excluding H. maraisii var. notabilis. The reason I have done so is because of a manuscript of mine dealing with the epithet intermedia which is with the Editor of Haworthiad. That manuscript deals with the impact of the granitic pluton at Robertson and associated dolomite on Haworthia although I have there not really discussed the geology as such. At Klaasvoogds where there is also a dramatic shift from ancient to very recent rocks and replication of the problem of a var. notabilis that is not dealt with in this paper. This set illustrates the populations that I used to formulate the concept of a species H. maraisii. It also includes the “escape” into the Little Karoo and the possible eventual link with H. emelyae east of Barrydale. The plants are characteristically very dark coloured and perhaps smaller in comparison to sets A, B and C. However, I think I can demonstrate reversion to a mirabilis-like plant and also similarity to H. heidelbergensis. I also include two populations that suggest a more far-reaching link to H. emelyae north and east of Garcia Pass, north of Riversdale.
Bosfontein JDV97/58. These are plants in a continuum from Trappieskraalkloof (61) on the same geological formation viz. Witteberg Group rocks.
Dublin F.J. Stayner in KG400/61, JDV94/18. The significance of these plants is that they were particularly dark-green, very tuberculate and the leaves were erect. Although I saw plants at the site with Frank Stayner I have not been able to find them at the site since. Plants do occur again a short distance to the east, but they do not have the same very erect and narrow leaves of Stayner’s collection. A plant is illustrated in my revision where it is under the number JDV94/18 and attributed to Roy Littlewood who was instrumental in first drawing attention to it.
Rooiberg MBB6956. This is north of the Breede River from the previous locality listed. The Rooiberg is quite a substantial massif and houses what can be regarded as discrete populations. Included in these is the very curious H. herbacea var. flaccida. I only observed five plants of that and suspect that it is of hybrid origin with the putative parents being H. maraisii and H. reticulata rather than H. herbacea.
Agtervink MBB6647. The following eight populations illustrate the geographic core of my concept of an H. maraisii.
N Agtervink MBB7176 [72].
Sandberg MBB7174, JDV98/18.
Nekkies JDV85/89, JDV96/24, EA083, EA1408. This population is very close to the Robertson pluton and I did associate this site with transition to the var. notabilis. I doubt if that this is actually true for this local area. Etwin Aslander made a collection (EA1408) from very close to Wolfkloof where the notabilis complex occurs, and this maraisii variant could be compared to those in 80 north of Ashton and even with 32 in Stormsvlei Pass.
S Sandberg MBB6670, JDV97/38.
Koningsriver MBB6672.
Vrolikheid MBB6968, W Kanonkop MBB6967, E Witkranz MBB7077. These and the next three populations are to the west of an area that should contain plants of the interface I would like to illustrate. However, it is a very dry area and not visitor friendly.
Steenbokshoogte MBB6966, Klein Spitzkop MBB6969.
Grootrivier MBB6677.
Breede River Bridge (Robertson) MBB6648. I noted this population in my Revision with the comment that note can be made of forms near Robertson which are rather similar to H. pubescens. The plants tend to have the same globular rosette shape and also the pubescence. However, the flower and flowering time is typical of H. maraisii as I defined it.
Muiskraalkop MBB6961, JDV87/208. This is the type locality for von Poellnitz’ H. schuldtiana var. robertsonensis as indicated to me by Payne. However, von Poellnitz does cite more than one locality for each in his description. The plants do have more slender erect leaves than most in this set but the distinction gets lost the more material that is considered.
Rietvlei MBB7055. This population is an odd one that I used in developing a justification for treating the element notabilis as a variant of H. maraisii. I was looking at the plants on a south slope under bush whereas it transpires that the bulk of the population is higher on the ridge in Enon conglomerate. Plants further north on Peninsula sandstone (Kranskop) are quite different and this difference compounds to the northwest where the plants are on Malmesbury shale.
Ashton MBB6668. This site is also Enon conglomerate and the plants are quite large and the leaves are rather recurved than with an end-area. The surfaces also tend to be smooth. Hence they do appear somewhat mirabilis-like cf. 32.
Cogmanskloof MBB6875 . These plants can also have leaves quite strongly recurved and there is a close similarity with the following.
Baden, Montagu JDV97/55, EA068.
E Montagu PVB7588. Malherbe of Sheilam stated that he had seen “small black plants” in the Koo valley north of the Langeberg. I have not been able to confirm this other than seeing them immediately northwest of Montagu. P.V. Bruyns recorded the species about 20km to the northeast (Dobbelaarskloof) and also at this site due east of Montagu. The plants have highly retused leaves but do not deviate substantially from others in this set.
Barrydale MBB6667, JDV97/39, MBB7214. J. Dekenah knew these plants at Barrydale and it is surprising that some varietal name or other did not evidence this. It is difficult to interpret outside of these sets and a complication is a very odd population south of Southey’s pass (MBB6666) that I have perforce regarded as H. magnifica var. magnifica. It is an embarrassment to me because it has resemblances to H. retusa and to H. mirabilis as much as to H. magnifica. In fact plants from there do not closely resemble H. magnifica in the form that it is manifest in 5 and 6 of set A. It (87) is important because of the following item.
Kleindoringrivier G. Marx in MBB6741. This population is midway between Barrydale and Muiskraal where H. emelyae var. major occurs, and it appears to be a connecting link. The flower and flowering time links it to the west and to these sets.
Loerkop MBB6979. This is southeast of Ashton and the plants have retused end-areas.
Viljoensdrift MBB6518. This is at the foot of the massif Elandsberg and seems to be in the continuum with 74, 75 and 76.
Middelplaas MBB7079. This population is similarly in some kind of juxtaposition between this set and the next and I interpret it like that in view of unlisted populations I have seen in this area.
E71 MBB6647.1 AgtervinkE72 MBB7176.1 N AgtervinkE73 JDV98-18 SandbergE73 MBB7174 SandbergE74 EA083.1 TierbergE74 EA1408.1 NW RobertsonE74 JDV85-89.1 Die NekkiesE74 JDV96-24.1 Die NekkiesE75 JDV97-38.1 SW SandbergE76 MBB6672.1 KoningsrivierE77 MBB6967.1 W KanonkopE77 MBB6968.1 VrolikheidE77 MBB7077.1 E WitkranzE78 MBB6966.1 SteenbokshoogteE79 MBB6677.1 GrootrivierE80 MBB6648.1 Breede River BridgeE81 JDV87-208.1 MuiskraalkopE81 MBB6961.1 MuiskraalkopE82 MBB7055.1 RietvleiE83 MBB6883.1 N AshtonE84 MBB6875.1 CogmanskloofE85 EA068.1 MontaguE86 PVB7588.1 E MontaguE87 JDV97-32.1 BarrydaleE88 MBB6741.1 KleindoringrivierE89 MBB6979.1 LoerkopE90 MBB6518.1 ViljoensdriftE91 MBB7079.1 Middelplaas
F. The interplay of Haworthia maraisii and H. heidelbergensis This set includes the var. meiringii at Bonnievale and a set of populations that suggest a closure with Set A and the emergence again of H. heidelbergensis. Many of the collections listed here seem to be entirely new to the literature of the genus. The tendency in this group, as a whole, is to erect leaves. They are also small with a maximum height of 50-70cm and a rosette diameter of 20-40cm. The geology of the area does not seem to be excessively complex or vary substantially from the previous set except that the topography is more uniform. However, the plants seem unable to hold the profile of the previous set and there is a drift to H. heidelbergensis.
W Goudmyn MBB6516. I start this series in the mid-west of the area because it has the more conventional forms of H. maraisii to both the north and the south. This population is west of Olifantskrans and extends on a long sharp ridge to the summit of the Elandsberg to the west. The plants are very small and have many small slender erect leaves.
Olifantskranz, E Goudmyn MBB6676, MBB7081. I have four collections from this general area and the plants are continuous with the previous set. The leaves are a little more substantial and often very spotted. The continuity extends both east- and northeastwards.
Olive Grove/Wakkerstroom MBB7081, MBB7084. This is the eastward extension and the plants are approaching H. maraisii var. meiringii at 95. The leaves are slender and erect and relatively opaque.
W Bonnievale MBB6509. There is also a record from north of this point which are less spinose and translucent than this particular population although the distance separating them in less than 100m. Here specifically, I find plants which I would compare favourably with H. heidelbergensis var. toonensis at Matjestoon east of Swellendam. The continuity is in fact with H. maraisii var. meiringii below.
E Bonnievale MBB7085, Edendale MBB7269. These plants are very proliferous and spinose. The rosettes are globular. The var. meiringii was actually based on the population to the near east of this one (MBB7269) where the vegetative similarity to H. herbacea was even more significant.
Bobbejaanskloof MBB7082, N Goudmyn MBB7083. Northeast of Olifantskranz (93) the continuity extends, not as expected to the conventional H. maraisii as at Loerkop, but to plants here with more slender erect leaves suggesting H. heidelbergensis.
Zandvliet MBB6964, MBB6965. Here there are two distinct populations that I include in the var meiringii together with the previous 6 populations. Plants do vary from individuals with stubby nearly terete leaves to plants with the conventional leaved wider than thick and narrowing to the apex.
SE Loerkop MBB6981. This is a very substantial population and while it would fit in the mélange of the previous item, it starts to relate uncomfortably to the next.
Tonteldoos MBB6974, MBB6977. These and the following to 111 are a nondescript assemblage of plants that do not relate comfortably with any of the other sets and neither do they collectively suggest any kind of taxonomic unity.
Mardouw MBB6990. Bob Kent also has two collections from between this and the following sites.
Nooitgedacht MBB6988, MBB6989 [102]. The first population is also a very substantial one close to eucalypts as is the following. However, the second is in natural vegetation.
Langverwacht MBB6992. This is a remarkable population of small very proliferous plants with quite turgid stumpy leaves. The site is notable as a quartzitic inselberg marked by the occurrence of several large gum trees. The haworthia seem to be the only plants tolerating this alien vegetation as the ground is otherwise devoid of plant growth.
Jakkalskop MBB6971.
Witkop (Drew) MBB7036.
NW Sanddrift MBB7056, W Sanddrift MBB7057.
Sanddrift MBB6625, JDV89/8, JDV90/31. Although very similar to the next, this population tends to have leaves with a retuse and opaque end-area. It is also small and very proliferous but nevertheless suggests an affinity with H. maraisii as in Set D.
N Sanddrift MBB6626, JDV87/193. This particular population comprises small plants with spreading leaves that are quite conspicuously translucent. Thus there is no difficulty in suggesting that they belong with H. heidelbergensis as does the next.
Middelrivier MBB7037.
Leeurivier JDV87/4. This particular population has been linked to H. heidelbergensis var. scabra and the following recent collection seems to confirm that this is the continuity that should be sought.
Nuutbegin MBB7099. This population is now south of the Breede River again and the problem is that it does not seem to relate to the initial populations in set A, but rather to those south in the Potberg area that I have not included in this particular discussion. Those populations are dealt with in a review I wrote of the H. floribunda complex and could initiate an overview of H. heidelbergensis.
F92 MBB6516.1 W GoudmynF93 MBB6676.1 OlifantskransF93 MBB7081.1 E GoudmynF94 MBB7080.1 WakkerstroomF95 MBB6509.1 W Bonnievale096 MBB7085.1 E BonnievaleF97 MBB7082.2 BobbejaanskloofF97 MBB7083.1 N GoudmynF98 MBB6964.1 E ZandvlietF98 MBB6965.1 ZandvlietF99 MBB6981.1 SE LoerkopF100 MBB6974.1TonteldoosF100 MBB6977.1 TonteldoosF101 MBB6990.1 MardouwF102 MBB6988.1 NooitgedachtF102 MBB6989.1 SE NooitgedachtF103 MBB6992.1 LangverwachtF104 MBB6971.1 JakkalskopF105 MBB7036.1 Witkop, DrewF106 MBB7057.1 W SanddriftF107 JDV89/8 SanddriftF107 JDV90/31 SanddriftF107 JDV96/25 SE DrewF107 MBB6625.1 SanddriftF108 JDV87/193 N SanddriftF108 MBB6626.01 N SanddriftF109 MBB7037.1 MiddelrivierF110 JDV87/4 Leeurivier
Discussion It is wishful thinking to even hope that some taxonomic decision can flow from the above discussion that will match the traditional classification of Haworthia. I consider that I set out to describe H. maraisii in the same format as a series of 5 articles that I wrote for the English Cactus and Succulent Journal entitled rather grandiosely as “Natural variation and species delimitation in the genus Haworthia” (ca 1972). I never completed the article because I could see that the 20 or so populations that I had then sampled held insurmountable problems. Since then an obstacle for me has been the seeming intellectual intransigence of readers, expressed by other writers, who like myself seem to have been wishing for some magical formula whereby the genus would unfold and disclose the obvious and comfortable units that we all wish species were – and also by lack of full exploration. My uncomfortable role has been to try vainly to educate and instruct a small willful segment in both academic and non-academic rank, that this Holy Grail is based on not only false hopes, but on false premises.
Classification of Haworthia, and indeed I contend that this will apply to many other genera, simply will not be accomplished unless there is agreement on purpose and definition. My botanist acquaintances seem to insist that there are many definitions for the word “species”. My non-botanist acquaintances are justifiably unconcerned about definition at all while they have names that they can use. That these names as Latin binomials should have a particular meaning in the whole context of biology seems to be lost to both groups. Thus the nomenclatural system as expressed by the International Code serves no true biological purpose. It simply regulates and legislates for the methodology behind a nomenclatural system whether this is for true “species” or for elements recognized out of sheer lack of sampling and poor definition.
My observation derived from the material discussed in this manuscript is that I can no longer see a clear distinction between H. maraisii and H. mirabilis as the names have been transfixed by typification either by myself or by I. Breuer or by the way in which I have applied them. The problem is not simplified by the fact that at both Bredasdorp and Napier it appears that two elements are in close geographic proximity. This rather suggests that a ring-cline may exist. I suggested in my Revision, that a taxonomic solution could be possible for species (populations) together in one locality (or close proximity), but wrong elsewhere. This observation seems to apply here. Similarly I cannot see that the set of populations in E above (H. heidelbergensis?) can be circumscribed as a species in anyway that H. heidelbergensis can truly be circumscribed and seen to be discrete. More discomfort extends from the interplay of H. heidelbergensis, H. magnifica, even H. floribunda, and the complexity of that element in the east toward Riversdale and Albertinia. The relationship of this aforementioned complex to H. emelyae in the Little Karoo is compounded by the connection via Barrydale of the greater H. mirabilis set discussed in this work. There are thus other sets of species which need to be considered in the same way as has been done in this work
What I do suggest is that species are fractal and that there will never be clear focus on all life forms. There are species that are presently in sharp delimitation and species in transitional mode that are not. This is particularly evident within Haworthia where some species are quite obvious and discrete and others are not. The scale of the taxa generates the same problems as it does in the artificial rank of genera. This is that there may be one or more very distinct elements that are quite disproportionate in numbers and populations to amorphous elements. The decision in Haworthia should be reached by consensus among individuals who indeed have the wisdom, competence and experience to make sensible choices. The evidence for these qualities has so far been sadly lacking.
Conclusion The set of populations that I have examined here, suggests that the probability that they can be proven to constitute two separable elements viz. Haworthia mirabilis and H. maraisii, is remote. From a practical point of view it is possible to uphold the names in the traditional way followed in my Revision, but only if it is accepted that these names cannot be applied with certainty to the greater proportion of the populations. From a botanical point of view it is patently evident that we are dealing with one species, Haworthia mirabilis. There is a greater problem in how this one species known by the populations discussed in this paper relates to the populations geographically further east. The implications of such a relationship for classifications of Haworthia may be profound and it could impact on classification of many other genera too. The obvious solution is to decisively abandon the dream of a classic nomenclatural solution to the problem. We could simply recognize that variants, which exist within the scope of any species, can be accommodated by simple descriptive statement and referred to by collecting reference and locality.
Acknowledgement I have been so privileged to have made the acquaintance of many people who have facilitated my excursions. Practically every spot on the map is associated with several persons. I truly regret that it is simply impractical to name them all and I sincerely hope that my appreciation and gratitude will not be taken to be lacking by dint of omission. ♦
I have several times been rather taken aback when botanists have been among those who have derided the fact that I have not apparently looked to flowers as a source of characters for identification of Haworthia species. Others have intimated that there are diagnostic characters in the seeds and even in the capsule structure. The essence of this kind of complaint is that there are these definable units called ‘species’ and that there is some linear and dichotomous set of characters by which they can be separated. The perception remains alive for the technology of surface structure, pollen sculpturing, DNA and molecular structure, and expectations which flow from and for these real and presumed character sources.
My opinion is that these techniques or methodologies will not tell us much more for Haworthia than what can be deduced by common-sense scrutiny of the plants. They may be extremely exciting and enlightening in view of broader relationships and theories of origin and migration even of vegetation. But their value to the collector and grower will always be minimal.
Here I want to picture some flowers of the subgenus Haworthia and show that while there is indeed ‘character’, the magnitude and ranges are such that they cannot be of any assistance for separating those elements that can be practically recognised as ‘species’. It is more probable that it will rather fuel the argument for less species. In the subgenus Hexangulares, my observation has been that while the species are vegetatively readily distinguishable and in most cases, geographically quite discrete, the flowers are indistinguishable. Within‑species differences may exceed the differences between the flowers of different species. In the case of subgenus Haworthia, it can be observed that in the one case where the flower is really notably different i.e. H. nortieri var. globosiflora (including H. pehlemanniae) there is a gradation to the normal flower of H. nortieri var. nortieri. Thus, within species difference may exceed that between species.
The flowers of H. herbacea and H. reticulata are interesting because they are quite large and the tip of the flower bud is ‘fish-tailed’. The tips of the upper outer petals are pinched together. But this character is present in the whole of the southern Cape complex of about nine species and extends to Little Karoo species as well.
Fig. 1. Left. H. maculata, Audensburg, N. Worcester, MBB6815. Centre. H. nortieri var.nortieri, Sneeuberg, Koue Bokkeveld, MBB 6158. Right. H. maculata, Audensburg. N. Worcester, MBB6815.
Here are H. maculata and H. nortieri var. nortieri – the former from the Audensburg north of Worcester, and the latter from the Sneeuberg in the Koue Bokkeveld north of Ceres. The plants are generally very similar and the argument to maintain species identity is no more substantial or tenuous than for any others of the subgenus. The flowers generally are different. H. maculata has the fish-tailed bud of H. herbacea and H. reticulata. This is evident in the illustration. The impact on the open petals is quite marked in that the upper outer petals tend to have pinched ends and the flower appears a little more regular i.e. the petals are equally spaced on radii from a point centred on the mouth of the tube. Colour is notable in this case because most of the Audensberg H. maculata flowers are notable pinkish. In the H. nortieri illustrated here they are white (ignoring the usual darker colour of the mid-vein). But I have photographed the one plant of H. maculata in which the colour is also white and the flowers less regular. It is almost impossible to separate this flower from that of the H. nortieri illustrated. This observation is also restricted to these particular specimens. It is well known that H. nortieri flowers can be very colourful with a notable yellow tube (yellowish-green, golden yellow and canary yellow fide G.G. Smith 1950).
The pinched bud does not always produce a more regular flower, and in H. maraisii, H. magnifica and H. mirabilis quite the opposite may occur with the upper outer petals almost overlapping behind the inner upper petal.
It has become increasingly evident to me that the separation of H. arachnoidea and H. mucronata as two independent systems, and therefore species, is difficult. Therefore I illustrate five specimens from four populations in:
Fig.2. Left – H. arachnoidea var. nigricans, between Barrydale and Montagu, MBB6868. 2scd Left – H. arachnoidea var. setata, Prinspoort, NE Montagu, JDV92/93. 3rd Left – H. arachnoidea var. arachnoidea, Robertson. 4th-5th – H. arachnoidea var. arachnoidea!, DeDoorns.
The spike on the left is of a doubtful population of H. arachnoidea. The plants are spinose as H. arachnoidea is supposed to be, but generally less than is expected in that species, and its location between Barrydale and Montagu makes it suspect. The second spike is that of unmistakeable H. arachnoidea var. setata from Prinspoort where the variety nigricans and H. mucronata var. inconfluens are also in proximity. The third spike is that of H. arachnoidea var. arachnoidea from west of Robertson. The fourth and fifth are of a collection from near De Doorns in the Hex River Valley. There are four plants in the collection and they have translucent spots on the leaf rather similar to H. marumiana var. dimorpha. The inflorescences are quite lax and as these two shown occupy a place in the greenhouse alongside the Robertson H. arachnoidea, it is obvious that the sparse lax spike is not light influenced. The flowers are sparsely spaced on this lax spike and the tube and flower colour is notable greenish. There does not seem to be any notable consistency in these differences and in many collections this kind of difference appears. The plants and flowers obfuscate the separation of H. arachnoidea and H. marumiana. The problem also arises as to how to name it. It is not typically H. arachnoidea var. arachnoidea as the nomenclatural system requires, and neither is it any of the other named variants.
Fig.3. Left – H. mucronata var. inconfluens!, west of Montagu. JDV97/56, MBB6690. 2scd Left – do. 3rd Left – H. mucronata var. inconfluens!, north of Montagu. JDV96/56, MBB6689. 4th Left – H. arachnoidea var. nigricans!, Schoemanspoort north of Montagu. JDV92/43. 5th Left – H. mucronata var. inconfluens, Touwsberg, JDV97/149.
The two left spikes are of an H. arachnoidea/H. mucronata intermediate from near Montagu. The flowers are more compressed together on one spike than on the other. The collection is notable for the intense yellow colour in the tube. The third spike is from the same geographic continuum also near Montagu but slightly eastward, inclining more to H. mucronata; as do the collections and next two spikes from Schoemanspoort near Oudtshoorn and the Touwsberg north-east of Montagu. The collections are probably of the variety H. mucronata var. mucronata. But I have explained elsewhere that H. arachnoidea var. nigricans is a buffer to absorb these problem elements which are neither one species nor the other, and not constituting any geographically cohesive entity. Incidental is that the third spike’s origin is in JDV97/56, a collection which also has the number MBB6689 and which contains individuals which could be H. arachnoidea var. nigricans, H. mucronata var. mucronata, H. mucronata var. rycroftiana, or none of these! This helps explain why I object so to opinions which are based on the unrecognised contention that one specimen can represent a population, a variety or a species. It also shows that it is better to work with a single accession system and one collector’s number rather than generate a series of personal numbers.
Fig.4. Left – H. arachnoidea var. nigricans, between Barrydale and Montagu, MBB6882. 2scd Left – do. 3rd Left – H. decipiens var. pringlei, Waterford, Jansenville, MBB6855. 4th Left – H. arachnoidea var. scabrispina, SW Laingsburg, MBB6486. 5th Left – H. decipiens var. pringlei, SE Mt Stewart, MBB6582. 6th Left – H. cooperi var. gordoniana, N Jeffrey’s Bay, JDV96/92.
Is a montage of two spikes on the left of H. arachnoidea var. nigricans from between Barrydale and Montagu. Then there is a spike from a plant from Waterford near Jansenville (my new H. decipiens var. virella). The plants are on the broad interface of that variety with H. decipiens var. xiphiophylla and they are not typical of anything. The next two spikes are from plants of H. arachnoidea var. scabrispina from south-west of Laingsburg. The differences are self-evident viz. a densely compacted raceme and a sparsely flowered one. Colour also differs. The sixth and seventh spikes are of H. decipiens var. pringlei from Mt Stewart and of H. cooperi var. gordoniana from north of Jeffrey’s Bay (this latter is actually close to H. cooperi var. picturata). The differences across the figure are actually minuscule and when one has to compare several spikes from each collection these differences become quite untrackable.
Fig.5. Left – H. cymbiformis var. cymbiformis! Kaboega, Zuurberg, MBB6903. 2scd Left – H. decipiens var. decipiens, S Aberdeen, GM547. 3rd Left – do. 4th Left – H. cymbiformis var. incurvula, Pluto’s Vale, MBB6884. 5th Left – H. cooperi var. viridis, Perdepoort, Sapkamma, MBB6858. 6th Left – H. cooperi var. isabellae, Holriver, Patensie, PVB7128. 7th Left – H. cooperi var. isabellae, Milton, Hankey, MBB6801.
Consists of a specimen which on vegetative grounds is unquestionably H. cymbiformis (but which variety is it? a la H. arachnoidea above). In the context of its near neighbours on the Zuurberg, and their similarities, it is actually continuous with H. cooperi. The plant (and others in the population) is very bright green yet in every other vegetative aspect is typically cymbiformis. The inflorescence is a bit sparse while the flowers are less white and perhaps sturdier than any norm which can be stated for that species. However, this inflorescence and flowers do not enable any better differentiation of the series of populations which link H. cooperi, several of its varieties, and also H. aristata in that area. The second and third spikes are from a collection of H. decipiens var. decipiens from south of Aberdeen. They are very different from each other. The fourth spike is of H. cymbiformis var. incurvula from Pluto’s Vale (and I have found it possible to make a perfect match of such a flower with the very improbable H. pulchella var. globifera, which is patently from another system). The fifth spike is from H. cooperi var. viridis and the last two are of H. cooperi var. isabellae from Holriver, south of Patensie and Milton south-west of Hankey.
Fig.6. Left – H. bolusii var. blackbeardiana (batteniae of Scott), Josefsdal, Cradock, JDV91/102. 2scd Left – H. decipiens var. decipiens, E Willowmore, MBB in KG140/72. 3rd Left – H. decipiens var. decipiens, Springbokvlakte, JDV94/85. 4th Left – H. cooperi var. gordoniana to H. decipiens, Georgida, Uniondale, MBB6937. 5th Left – H. decipiens var. decipiens, Steytlerville, PD2153.
Lastly Figure 6. These are flowers of H. bolusii var. blackbeardiana from Cradock, H. decipiens var. decipiens from east of Willowmore (note the many florets per bract), and H. decipiens var. decipiens from Springbokvlakte. The fourth spike is of H. cooperi var. gordoniana where it intergrades to H. decipiens at Willowmore and Uniondale. The last is another also of H. decipiens var. decipiens from the Springbokvlakte.
If any one species complex (as I have identified and recognised them) is taken, the same problem of variation arises. The variation in the flowers is of the same degree and intensity perceivable in the size, leaf arrangement, number, colour, texture, spination and armature of the rosette and leaves.
The conclusion I would like to make is that it is not possible to generate a classification system on any other logic than that I have used viz. geographic arrangement and broad vegetative similarity. It is possible to generate a diagrammatic model to show a cladistic (branching) structure and model for species. But this will have to have three axes – two for geographic space, and one for time. This is realistic, and far more so than the pressure that is exerted to produce the artificial two-dimensional cladograms botanists seem to favour and which the nomenclatural system caters for. ♦
This essay was published in Alsterworthia International Special Issue No. 4.
My fascination with Haworthia has presented me with many problems in the way the genera in the Alooideae have been discussed, appraised and modified in and subsequent to G.D. Rowley’s analysis (1967). Parr (1971) coalesced Astroloba, Haworthia and Poellnitzia and I refuted this in 1972 when I also wrote a rebuttal of Rowley’s paper. My remarks did not deter Mrs. Obermeyer-Mauve (1973) following and accepting Parr, nor in adding Chortolirion to Haworthia. Rowley (1976) quite pragmatically discussed the Aloid genera, but in 1980 suggested the incorporation of Poellnitzia in Aloe. He implemented this proposal in 1981 and promoted it again in 1985. Smith and van Wyk (1991) published a cladistic analysis of the Alooideae which I felt was unacceptable because of the fallacious character states and sets that were used there. Despite that paper and at least four others (Smith 1991, 1994, 1995; Smith & van Wyk 1992) generally supporting the uni-specific status of Poellnitzia, Manning and Smith (2000) incorporated the genus in Astroloba.
My objection to this manipulation of the genera is that the supporting arguments have been incorrect and that it essentially has not addressed or considered what I perceived to be the stumbling block to arriving at a better delimitation of the genera in the sub-family. This is the relationship within the genus Haworthia where it is quite evident to me that it comprises three distinct sets of species (the subgenera of Bayer ex Uitewaal). The floral and morphological differences for those three sets are absolute, and I am sure will need to be seen so in any way in which Aloe is configured. This is because the floral similarities within those subgenera are so minimal. I consider these floral differences to be as dramatic for genus delimitation as any of the character states covered in Rowley’s (1967) analysis. It would alarm me if the result of a DNA study produced any other result. The sets also appear to me to “behaviourally” different and with this consideration the genera (even if unispecific) Poellnitzia, Chortolirion, Astroloba are of similar status. This is not to imply that I do not recognise the problems with many other oddities in the Alooideae which require re-evaluation of the generic arrangement.
Adam Harrower of NBI asked me to identify an Haworthia he had collected near the Potberg (cf. H. heidelbergensis). In view of his interest I asked him to lookout for H. limifolia on a trip he was to undertake to the eastern Transvaal. On his return he presented to me a plant he took to be that species. It was in fact Chortolirion angolense. However, he produced photographs of another species at which I exclaimed “This is a new genus”, but probably a Chortolirion. Here I recognise the irony of this reaction when the generic arrangement in Aloe is so questionable.
The plant that Harrower collected has thin slender spineless hyacynthoid leaves (fig.1) with very pronounced bulbous bases. There seem to be few accumulated dry bulb scales as in Chortolirion and the plants have not displayed deciduousness as does that species. The older leaves blades dehisce leaving fleshy bulbous base and these are spirally arranged in the lowest order of the Fabonacci series viz. 2, 3 or 5 (fig.2). The roots seem rather sparse and are the yellowish colour of Aloe. The flower intrigues me more (figs 3 & 4). The free terminal portions of the flower are not channeled but are flared in the style of the sub-genus Haworthia. The midribs of the inner petals remain exposed and the margins of the outer petals adhere close to the mid-rib of the inner. This is the case in the subgenus Hexangulares and Chortolirion, as well as in this “new genus”. The lower petals are more undershot than is the case in either the Hexangulares or subgenus Haworthia.
The geographical location is the high-lying escarpment between the ranges of the species H. koelmanniorum and H. limifolia of the Haworthia subgenus Hexangulares.
Why I have stated so categorically that this is a “new genus”, is largely because of the historical maltreatment of the genera of Alooideae and the failure of students and commentators of Haworthia to exhibit any rational species concept. Such a concept appears to be missing for the genera as well. What has troubled me in recent times is the emerging belief that molecular study will provide the basis for a real and irrefutable phylogenetic classification. We have now a paper Phylogenic relationships in Asphodelaceae Taxon 52:193 (2003) by Treutlein et al, which will enable one to see to what extent these expectations are being met.
What strikes me immediately about the paper is firstly a statement in the abstract, and secondly the unfortunate selection of study material. The abstract summaries the results of the work done and this is: “The current taxonomic system does not reflect the phylogenetic affinities and relationships among the succulent genera Aloe, Chortolirion, Gasteria, Haworthia and Poellnitzia.” I find this extraordinary, as it has never occurred to me that it did, and I would have expected this sentence worded in such a way to form a key question to be answered by the study rather than as a conclusion. Similarly Treutlein’s closing paragraphs of his discussion should have been used as key questions to be answered by the study and not have been derived from it. One sentence reads “complete sampling … needed”.
The selection of material I take to be somewhat irresponsible given the past history of classification of the group and the literature. I would have expected some kind of predictive approach to the selection of material. The inclusion of unknown hybrids such as H. ryderiana, H. kewensis, and H. icosiphylla can tell us nothing. Ignorance of the actual taxonomic position of H. geraldii is similarly curious. The material is virtually entirely ex hortus and given my experience with identification; even voucher specimens are hardly likely give such material much credibility. I do think more thought and consideration should have been given to the species used in relation to the problems they present to their classification.
Putting these considerations aside, I would comment as follows on the results as presented primarily in Treutlein et al’s in fig 4. It does tell me what I had supposed i.e. subgenus Haworthia is very different from the other two subgenera of Haworthia. BUT H. geraldii (i.e. retusa) is grouped with H. attenuata and H. glauca and close to Gasteria! It is a result which does not make sense. The true H. geraldii is H. retusa and that is arguably an ecotype of H. turgida as much as it is a discrete species. We could indeed just be seeing ‑ in Treutlein’s own words ‑ a “gene tree”. Of course genes that can be sampled in the different types of DNA analysis are only another data set, and are not necessarily responsible for the morphology that we can see. Nevertheless it is equally true that, say; observed floral morphology may be the product of interplay of far more genetic material than those analyses entertain.
The inclusion of the unknown hybrids just introduces unnecessary tension or clutter or inaccuracies into the cladograms. H. kewensis and H. ryderiana may have parentage in subgenus Hexangulares or even Aloe, to explain their position in the cladogram. Astrowothia bicarinata/skinneri is the hybrid H. pumilaXA. corrugata (muricata!) so it is not surprising that it comes out with A. corrugata.
What is significant is that Chortolirion comes out with the grass Aloes, thus showing that the Hexangulares flower may be homoplasious (i.e. having evolved more than once). It would surprise me for a structure as complex as the flower to be so. Please note that Treutlein is wrong in saying Uitewaal divided Haworthia into two groups and that the “former including the subg. Haworthia and subgenus Robustipedunculares“. Uitewaal divided the group Hexangulares into Gracilipedunculatae and Robustipedunculatae. The way this is has been repeated in the closing paragraph of Uitewaal’s paper has confounded Treutlein, who goes on to say “This division is strongly supported by…”. This is not true. He has no Robustipedunculares in his analysis apart from the DNA (cytoplasmic) in the hybrid Astroworthia. It is thus not surprising that comes out in the “heterogenous group” that includes Aloearistata, Gasteria, Poellnitzia, Astroloba and, in a slightly wider group, H. retusa (“geraldii“, where its position here is very dubious).
Dr. Manning (personal communication) feels that the results vindicate the treatment he and Smith gave Poellnitzia. I must state that he is quite correct on the point of bird pollination and it has been shown that sunbirds do pollinate (some) Microloma species. But I cannot agree that Poellnitzia is unequivocally Astroloba. In Treutlein’s fig.3, Poellnitzia could be with either Gasteria or Astroloba, with Aloearistata as a wild card nearer to Astroloba than is Poellnitzia (one needs to look at distribution and variability of Aloearistata to know that something is not kosher here). In fig.4 Poellnitzia collapses below Astroloba as does Aloe aristata and the cladogram shows no “bootstrap values” to substantiate an opinion. Would one argue that Aloe aristata then also belongs in Astroloba?
As there is no member of the Robustipedunculares in the analysis and one can only wonder what a better species representation would have done to the cladogram. It is important that Chorolirion is seen to group with the “grass” aloes. The flower is definitely Hexangulareoid whereas the bulbous structure is replicated in Aloe buetneri as well as in a few of the “grass” aloes. My opinion regarding the Harrower “new genus” is that it is in fact discrete and introduces an entirely new dimension to the discussion.
Something should also be said about cladograms. The binomial system is essentially built on the concept of dichotomous branching. The fact of the matter is that in any two-dimensional cladogram which is used to depict relationships, the one axis will represent time and the other two-dimensional space. If one considers that species, like any other phenomenon in creation, are spread in space (they change with time), a two-dimensional diagram is very restricting and will lead, for example, to statements such as “Poellnitzia is nested in Astroloba. To think that a statistical bootstrap value will give a true measure of that distance in a two-dimensional array may be wishful thinking.
After seeing this Treutlein paper I am happier with the way in which the existing classification meets my needs for identification and communication and I do not think the word “merely” used by Treutlein in this connection is appropriate. Whatever high-grounded attitudes now seem to prevail that DNA studies give a new and correct dimension to phylogeny as the intent of classification, my contention is that this has always been the aim even in the most simplistic morphological studies. Classification is a prime function in zoology as well and there has never been any doubt that phylogeny was the prime aim of the classification. The only reason it seems to be obscure in botany is the absence of good archaeological or fossil evidence and the problem of determining homologies in a muscle and nerve free organelle.
Dr. Manning kindly made this comment among others which I may not be doing justice to:
“Clearly there are just two alternatives IF one wants to define genera in phylogenetic terms: lump everything into Aloe or split Aloe up into several other genera, each corresponding to one of the monophyletic branches that are revealed by the analysis. The latter treatment is confounded by a) incomplete sampling within Aloe and b) probable lack of good characters by which these segregate genera could be recognised”.
I would have omitted “within Aloe“.
Treutlein et al make another statement … “The further the status quo departs from reality, the more difficult it will be to integrate practice and theory.” This seems to be a twist of words. I think it should read … “The further theory departs from practise the more difficult it will be to integrate the status quo with reality?” Again that is the problem that should have been addressed in the preliminary approach to the study. The status quo by definition cannot depart from anything.
To close, I need to say something about definitions. I have already made several statements about the absence of definition at species level, where I still find resistance among students of Haworthia (if not wider). It is simply a fact that classification is a process of specifying so that one can generalise about a set of some kind. In dealing with Haworthia, I experience the group as three sets, and as stated above, these sets relate to the present classification as I have already described. The current generic classification expresses the experience of collective experience of all previous writers and researchers in the field. It has an historic value and an embedded “truth” of some kind; or, if the classification process has credibility, it should have and it should be respected accordingly. The definition of genus quite obviously has to be linked to that of species – systems of species which can be shown to be related sets in respect of morphology (physiology), genetics and geographical considerations. I contend that this has always been an unspoken aim and intent of classification and it is wrong to suggest now that it is absent from existing classifications. In my experience it is/was always seen to be the ideal that classification should reflect phylogeny. In doing so it would also reflect morphological/physiological, genetic and behavioural characters that lead to understanding. This is what I wrote in Asklepios 77:6 (1999) … “understanding of what it really means to be human”. If we are pursuing knowledge for any other purpose it may be that we are on an ego-trip. To maintain that floral structure is very limiting in the way it has largely driven classification is correct to a degree. I think it is wrong to make the implication that floral structure is driven by less characters than, say, those nucleotide sequences of the Treutlein analysis.
Touching on the “behaviour” aspect ‑ I think Poellnitzia is unique. The Robustipedunculares and Astroloba possibly could not maintain their integrity in each others’ company viz. hybridisation between members where they co‑occur. They complement each other geographically. Hexangulares maintains integrity wherever and with whatever it grows except with its own members. Subgenus Haworthia similarly always maintains integrity except with own members and presents a degree of plasticity far greater than any complementary set. Poellnitzia maintains its own integrity and is extraordinarily invariable in the close company of Gasteria, Robustipedunculares, and subg. Haworthia. Hexangulares and Robustipedunculares occupy quite different territories and virtually do not meet at all.
Acknowledgement Dr. J.C. Manning, Compton Herbarium, Kirstenbosch responded in very kind manner to my criticisms in Bayer 2003, and was most helpful in obtaining hard-copy of the Treutlein publication, as well as in interpreting the DNA methodology for me for the purposes of this paper. In having written this, I do not dispute his need for a pure phylogenetic model. I am simply trying to put a case based on my practical experience and knowledge of the plants concerned, hoping that the two poles will eventually meet in a functional way. Paul Forster of the Queensland Herbarium and Steve Hammer of California were kind enough to comment on, and edit, drafts of this article.
Literature cited
Bayer, M.B. (1999). Is classification science or art. Asklepios 77:3. [link]
Bayer, M.B. (1972). Re-instatement of the genera Astroloba and Poellnitzia. Natl.Cact.Succ.J. 27:77. [link]
Bayer, M.B. (2003). Classification with purpose. Alsterworthia Special issue 3:5.
Manning, J.C. & Smith, G.F.S. (2000) Asphodelaceae: Alooideae. The genus Poellnitzia included in Astroloba. Bothalia 30:53.
Smith, G.F.S. & van Wyk, A.E. (1992). Pollen morphology of the monotypic genus Poellnitziarubriflora (Alooidea: Ashodelaceae). S.Afr.J.Bot. 58:90.
Treutlein, J., Smith, G.F.S., van Wyk, B.E. & Wink, W. (2003). Phylogenetic relationships in Asphodelaceae (Alooideae) inferred from chloroplast DNA sequences (rbcl, matK) and from genomic finger-printing (ISSR). Taxon 52:193.
Addendum: Since the above was written, I have seen a copy of a second paper by Treutlein et al (2003). They make three observations to which I must draw attention.
“An exception in the current classification was found with the sister species H. geraldii and H. gracilis var. tenera: genetically they belong to group ll, whereas morphologically they show affinities to the subgenus Haworthia (represented by group 1).”
“H. geraldii and H. gracilis var. tenera are sister species according to rbcL (fig.1A) and matK (fig.fig.1B). Contrary to their previous morphological classification (Bayer, 1999), they are clearly grouped in Haworthia subgenus Hexangulares by both molecular markers.”
“… the taxonomy of the genus Haworthia must be revisited. More species of both groups need to be examined to determine their phylogenetic relationships before taxonomic consequences should be drawn.”
It has been disconcerting for me to conclude from past experience that perhaps botanists in other disciplines are not fully aware just what it is that taxonomists do, and the constraints that taxonomists work under. Another observation is that there are botanists who move between disciplines and thus do not have a firm foot in the taxonomic camp. This seems to result in decisions that may be thought to be relevant and correct but yet are incongruent from the purely taxonomic viewpoint.
It is stated quite clearly in most of my writing, certainly so where I have been at pains to explain my requirement for “species”, that my classification is barely morphological. It is clearly so in terms of the subgenera but not for the “species”. In my revision of 1999, I particularly stressed the “geographic component”. Thus for Treutlein et al to suggest that my classification is primarily a morphological one has no relation to my writing at all. The writers have not addressed my classification in respect of my approach, which has so often been explained. I have pointedly tried to analyze Haworthia as a system of integrated populations that is almost impossible to separate into discrete entities on morphological rounds. The very absence of morphological criteria and character is what motivated me to explore Oxalis where there is such character (see Bayer in Herbertia 48, 1992) and so validate my decision-making criteria. Perhaps the authors have never themselves addressed complex living systems and neither do they appreciate that there are taxonomists who have perforce had to do so. I have said elsewhere, there is a lack of discrimination between good and bad taxonomy because there is no definition and no standard.
The question of the subgeneric relationships of H. geraldii and H. gracilis var. tenera are concerned is a fundamental issue. If either of these had been considered and reviewed as taxa, it would have been quite evident that the conclusion Treutlein et al reach with such felicity is impossible. The taxon “geraldii” has been discussed many a time. Further discussion gives credibility to the recognition of local variants as full species, which is wholly misplaced. In the case of H. gracilis var. tenera, the writers can be excused for overlooking the discussion of this taxon in my book Haworthia Update Vol.1 (2002) or in Ecotypes in Haworthia (Aloe ca 2002). In the latter paper I explain why the species “gracilis” and all its varieties are transferred to H. cooperi. This was affected in Haworthiad 16:62 (2002). It is further evident to any reader that the two taxa “tenera” and “gracilis” cannot be excised from the subgenus Haworthia by any stratagem at all. That the term “sister species” is used, wholly disregards anything that I have written in respect of “species” concept for Haworthia. No matter how the subgenus Haworthia is manipulated these two taxa are elements in quite different sub-domains. Writers in the technical literature may claim that the popular literature where I mostly write is outside of formal science. IF they do so, then they must surely ensure that the academic and intellectual rationalisation and conclusions are acceptable and true even at that low level.
The Treutlein et al. paper brings to an end an era of faint hope that truth and sensibility would prevail. It is not the taxonomy of Haworthia that must be re-visited but to common sense that we should return. Botanists who do involve themselves with Haworthia should recognise the appalling state of affairs that has been introduced in the guise of science.
This essay was published in Alsterworthia International in a special issue No.3, June 2003.
This is a two-part essay. The first is to discuss a problem in the small Madagascan Aloe species, and the second to discuss Poellnitzia. The latter is now be listed in the genera of Southern African plants as Astroloba rubriflora. I think it is necessary to point out that while this may now be perceived to be ‘authoritative’, the taxonomic treatments are in fact not so.
A. The small Madagascan aloes: In my pursuit of a communication framework for Haworthia, I have experienced many problems. Not least, is the response I get from botanists. From this experience I thus find the article in Taxon authored by no less than four botanists, really most instructive. The article (Taxon 44:513, 1995) deals with the differences within the genus Aloe and the re-instatement and transfers by P.V. Heath of some of the Madagascan species (A. haworthioides, A. boiteaui and A. parvula to Lemeea, and A. bakeri, A. bellatula, A. calcairophylla, A. descoingsii and A. rauhii to Guillauminia together with A. albiflora.) As will be shown below, the information that Heath’s ‘classification’ either provides or obscures is virtually nil, and the reversion restores a heterogeneity which also says very little. The Taxon 5‑page article starts the discussion with a textbook quotation from Jeffrey (1968): “There is no single criterion which by itself can be regarded as unfailing for recognising a genus. For practical purposes the genus should be regarded as an inclusive category whose species have more characteristics in common with each other than with species of other genera of the same family. There should be a clear morphological break between the members of the particular genus and the members of other related genera. Past and current practise are also important, since a major change in classification and naming can create communication problems and result in loss of information.” If one considers that Taxon is a prestigious scientific journal, this quote is surely unnecessary as the Asphodeloid genera have defied logic since Linnaean times.
Smith et al claim to have undertaken an investigation of Aloe to show that it is polymorphic. The basis of any genus is polymorphism. The evidence of the investigation is firstly, a table listing the ‘characters’ used by Lemee to distinguish Aloinella (= Heath’s Lemeea), against Aloe species which are suggested by Smith et al to have the same “characters”. Secondly, is a similar table juxtaposing Guillauminia with Aloe species which suggestively also have the same “characters”.
Why I have put “characters” in inverted commas is because it is well known in classification, that recognition of categories may be intuitive rather than rational. I claim that this is the case here as well, and that the analyses made by Smith et al are just fallacious. Even were the facts of “character” true, it would still be a fairly pointless exercise to try and simplify a multivariate state the way they do.
To illustrate this:- Take Lemeea in Table 1 in the Taxon article:
Rosette small and dense. Smith et al cite A. aristata and A. humilis as having such rosettes as well, but Haworthia also has species which have small dense rosettes. The fact is that these small dense rosettes can readily be seen to be different, and recognition of species and even genera can be made on the basis of seeing only those rosettes. So an informed observer can distinguish many of the species on the basis of the rosettes only. This is in contradiction to a situation where experienced herbarium workers have confused members of wholly different genera e.g. Aloe aristata and Haworthia, H. pumila (maxima) and H. limifolia, Aloe humilis and Haworthia, Haworthia venosa and Astroloba muricata (aspera/rugosa/corrugata).
Leaves narrowly linear. Smith et al cite Sect. Graminaloe. This is another misleading comparison. Any average observer in Aloe can recognise that section on the basis of its leaves. To suggest that one could not separate any of these Graminaloe from the small aloes in the group proposed by Heath on the basis of leaf character is an indictment of the skills of ordinary people.
Pedicels negligible. Chortolirion too, has a negligible pedicel as do several species of the sub‑genus Haworthia. But A. bowieae too is cited, and it could be claimed that, as Chamaealoe, it communicated something which is lost when it is treated as Aloe.
Filaments broad. No comment by Smith et al. Aloe haworthioides has the most remarkable filament structure and the broad upper part of the filaments can be mistaken for petals and an extended perianth tube. In fact it is quite ironic, and likely, that the same experienced amateurs who can see differences in the rosettes which Smith et al cannot, may mistake the filaments for petals. These filaments cannot emerge from the tube before the flower opens. To suggest that this condition in A. haworthioides is the same as the extended filaments in say Aloe ferox where both anthers and stigma are presented to pollinators outside of the tube, is misleading.
Anthers exserted. Many aloes do indeed have exserted filaments and it is hardly necessary to cite something as spectacularly different as the huge-stemmed A. ferox in a discussion of plants with small compact rosettes.
Ovary acute. This is cited as erroneous, but there is so little evidence anywhere for the shape of ovary, style and stigma in Aloe species, that this character is hardly worth noting.
Now take Fig.2 for Guillauminia of Bertrand:
Inflorescence lax. Smith et al would have to give a very precise definition of this term and say exactly how they would use it in this particular context. In Haworthia there are inflorescences which can be described as sparse and lax, and inflorescences which can be described as dense and firm (sometimes within the same species!). Hardly any of them are quite as firm as is the case in A. tenuior for example ‑ which has a variant on the Zuurberg having a lax inflorescence compared with its more robust relatives on the karoid plains where the inflorescence could by comparison be described as firm.
Perianth segments free. Smith et al cite A. myriacantha and A. albida. The actual situation is one which does not seem to have been explored in the genus Aloe, nor in related genera. The flower in the Aloaceae comprises three outer tepals and three inner. There are many different states within these genera. The outer tepals may be free or partially free or form a separate tube; they may be fused to greater or lesser degree with the inner petals to form a more convincing tube. More to follow.
Anthers exserted. This is a fairly meaningless statement in view of the degree to which anthers are exserted or not exserted in the genera as a whole.
The facts thus are that these two tables are just a fallacious assembly of observations used by Smith et al to negate an unsubstantiated opinion put forward by Heath. They do not comprise a series of facts which face the scrutiny suggested by Smith et al, and which are therefore just as weak as the rationale for Heath’s genera.
Tables 3 and 4 of the Taxon article, which supposedly tabulate the “characters” of the species which Heath placed in Lemmea and Guillauminia are technically no stronger and have no more substance than the previous two tables.
Smith et al conclude… “The species transferred by Heath to either genus do not have any features that might even remotely warrant their segregation from Aloe“. It is really regrettable that this kind of a statement is a feature of taxonomy. It can be liked to Shakespeare’s “Me thinkest thou dost protest too much!” It is obvious that there are characters which suggest difference of quite a major degree. If there were two more species that had those exserted flattened coloured filaments of A. haworthioides, they WOULD in all probability be taken to be a separate genus. The weakness of the protest is underscored by the next statement. Despite now claiming proof that the species can only be classified as Aloe, Smith et al go on to say “Should Heath’s views be followed, many more Aloe segregates deserve recognition”. It is a curious co-incidence that a paper by Forster in Bothalia 30:53 (2000) follows one in which Poellnitzia is included in Astroloba. Forster writes of the “characters” by which species of Lomatophyllum are easily distinguished. In fact there are two characters and these are the fleshy fruit and wingless seed. Just how much difference there is between the fleshy fruit of Lomatophyllum and the said non-fleshy fruit of Aloe is, is conjectural. The fruit of Aloe is initially also fleshy. Winged versus winglessness is a character set which has not been explored within Aloe. Thus even Lomatophyllum, as genus or subgenus, rests on flimsy ground. What is disturbing is this feature of scientific writing. Forster cites the cladistic analysis of the Alooidae by Smith and Vanwyk published in Taxon 40:557 (1991) giving it a credibility which can be proven to be totally misplaced. That analysis rests on a set of characters which are just as fallacious as similar allusions I make in this paper.
Actually therefore the statement by Smith et al is the only substance in the entire 5‑page article by the four botanists which is of note. The article has actually demonstrated that classification is not science (as this writer has claimed it to be), and neither is it art. It is simply the arrangement of groups of plants in an ill‑defined way where they are said to be species related together in genera. It is done in a way in which we can communicate with one another about them in the hopes that we can make general statements about specific things.
My complaint now is that the papers by both Heath and Smith et al fiddle with the classification and nomenclature, without adding one iota of information to the subject. The facts are:
A. haworthioides and A. albiflora do in fact have characters which make them most unusual in the genus Aloe. There is no Aloe remotely like A. haworthioides which has the extraordinarily protruding flattened filaments. These form a reddish tube nearly as large and conspicuous as that of the perianth itself, which is almost hidden by the large floral bract. In the case of A. albiflora, the florets are widely campanulate and sparsely arranged on the slender peduncle. Reynolds placed 12 species together in his Group 1 of Madagascan Aloes and stated most plainly that they were not closely allied. Now Heath has ignored three of those 12 species and made two groups out of the disparate remnant. This is the problem that Smith et al should have addressed. The curious thing about this is that there appears to be a similarity between A. bellatula and A. perrieri (ignored and abandoned in Aloe by Heath, and not even considered by Smith et al), which in terms of variability could be extended to A. albiflora and perhaps further to A. parvula. There is no such similarity between A. haworthioides, and A. bouteaui. A. descoingsii and A. calcairophylla do seem to have unique florets. There is nothing odd about the florets in A. rauhii or A. bakeri that could necessarily separate them from, A. versicolor and A. parallelifolia.
In my own observation of Aloe, which I have had to consider when trying to find solutions for Haworthia, I have been struck by the artificiality of the whole classification by Reynolds. Not for a moment do I suggest anybody else could have done it better, but it is in fact just an eye-balled solution. Looking at the Madagascan aloes, it seems to me that species like Aloe conifera and cryptoflora are probably synonymous. In the South African aloes, there is a problem with Aloe striatula. in which the floret is quite unique (the inversion of the arrangement of outer and inner tepals). This species is also oddly placed with species like A. ciliaris and A. tidmarshii in a Subsection and Series (categories which Reynolds abandoned for his second volume for the non-South African species).
In once trying to identify a small Graminaloe from McClear in the Transkei, I could not separate it from neither A. myriacantha nor A. albida. Reynolds in his first volume had a very restricted distribution for A. myriacantha, but in the second volume it is shown to have the second widest distribution after A. buetneri. Thus it is obvious to me that it is in fact synonymous with A. albida, and may include even A. minima and A. saundersiae (the question of actinomorphy vs zygomorphy intrudes here!).
The Smith et al comment, that if Heath was followed many more segregates would have to be re-instated or created, is irrelevant. It suggests that these authors could authoritatively identify such segregates, and from what they have done in Tables 1 and 2 only, I have to conclude that they could not. The issue is what benefit would there be. Heath’s hypothesis, which is a flattering term, suggests affinities which are not there. The Smith et al annulment is appropriate, but poor in rationalisation.
An important issue is that of ‘characters’. The details of floral morphology are really not understood in the Aloaceae. This statement is underscored by the historical lack of attention that has been paid to the differences in the flowers of Haworthia and its three sub-genera, also of Astroloba and of Poellnitzia. In fact a recent treatment of Poellnitzia places it in Astroloba, which is so incongruous one has to ask why recognise genera in the Aloaceae? Several attempts have been undertaken to unravel the relationships of the genera and produce a structured phylogenetic tree (cladistics). None of these have considered the diversity within Aloe, nor the actual structure of the florets.
The quote from Jeffrey that Smith et al use, is probably useful at undergraduate level to suggest some kind of ideal. Why they have used it in this article in the Aloaceae where the genus Gasteria is probably the only genus which has more than 10 indisputably related species, is curious. My observation will be lost on the reader unless I state that subgenus Haworthia could be allied with the Graminialoe, the subgenus Hexangulares with Chortolirion, the subgenus Robustipedunculares with Astroloba, the genus Poellnitzia (now Astroloba) with Gasteria. Jeffrey’s requirements are no doubt met to some degree by these genera, but the genus Aloe is by far the greater body of species, which remains relatively intact by default rather than as a rational considered portrayal of morphological or genetic difference. As I would plead for Haworthia – do not make changes unless they contribute substantially to knowledge and understanding. Any changes should facilitate communication and make that communication meaningful. It may in fact be very useful to recognise some of those segregates of Smith et al which we are left to guess at. There are substantial segregates such as the group of Leptaloe and the maculate aloes. These could profitably be separated out simply to direct better attention to the extent of diversity in the family and the genera and make them more manageable.
The problem in Aloe may primarily be, one could suspect, because the revisionary work has been done by an amateur. This is G.W. Reynolds. No matter how much effort and diligence and intellectual skill has been applied, there is no way that the work can represent a real contribution to the essence of classification of biological systems. Smith et al are professional botanists and their observations here are no better directed. Reynolds, in his preface to The aloes of South Africa writes “The beginner will learn … ” and goes on to the superficial comment found in every popular work about mans ignorance, and the refusal of nature to conform with mans wish for precision. But neither he nor other botanists really seem to make much progress beyond the beginner stage. The fact is that a classification is for people to use. The vast assemblage of species in the single genus Aloe need to be teased out for practical purposes and the weak rationalisation described above is a blow to any hope of clarity. The fact is that a species may be “seen” to be different but there may be no “characters” by which the degree of difference can be demonstrated.
B. Poellnitzia rubriflora: A problem similar to that of Lemeea and Guillauminia, occurs with the genus Poellnitzia. This species P. rubriflora (L.Bolus) Uitewaal is a very distinctive species of plant which cannot be confused with any other. So it is really very curious that it has such a chequered taxonomic history. At least, this would be curious if this systematics and of its related genera and species had not suffered such cavalier and unbelievable treatment by botanists and would be botanists. Smith and Manning (Bothalia 11:53, 2000) now place P. rubriflora in Astroloba. They say … “Vegetatively the species shows close affinity to some species of Astroloba and accords completely with the genus in its tubular actinomorphic flowers with included stamens.” This is in contrast to Rowley’s suggestion (in Smith, 1994) that Poellnitzia shared more characteristics with the genus Aloe than with Haworthia. Or with Parr (Bull.Afr.Succ.Plant Soc., 1971) who placed Poellnitzia in Haworthia.
The facts are that Poellnitzia is similar to all seven Astroloba species in that the plants have the leaves carried on extended stems in clear 5-farious tiers. In any Haworthia which have such extended stems, the leaves are trifarious or multifarious. Trifarious and even 5-farious arrangements do occur in some stemless species but just as dictated by spiral phyllotaxis. Poellnitzia is characterised from Astroloba on the basis of a very waxy epidermis.
The statement about the accordance of the flowers is a most extraordinary statement. In Poellnitzia the perianth tube is apparently entire as the margins of the outer petals are fused to the mid-rib of the inner petals for their greater length. The midribs of the inner petals are clearly visible between the ‘margins’ of the outer as they are in Astroloba, as they are in two of the three Haworthia subgenera and as they are in Aloe. The most obvious similarity of the Poellnitzia flower is probably to that of Gasteria, except that the curvature of the tube is down as it is in Haworthia subgenus Haworthia, and opposed to up as in Gasteria.
This statement raises the question of actinomorphy. The flowers definitely are not actinomorphic in Astroloba – this is a persistent myth. There are always two distinguishable upper outer tepals and the single outer lower tepal. This is the same as in the Robustipedunculares of Haworthia. The Poellnitzia flower is only actinomorphic in a superficial sense and I would not call it so in an analytical and scientific sense. Just the curvature of the tube alone, denies any bilateral symmetry except in one plane. I frankly do not understand why this perception is so strongly held, but then I do not understand why formal botany has always treated the Liliaceae in such a cavalier way. The tepal tips may be actinomorphic but the tube is generally simply asymmetric and there is no way that one can bisect it in any plane but the vertical one. In Haworthia, and in most species of Alooidiae that I have examined, the upper inner petal overlaps the upper margins of both the lower petals. There is no similarity in the flower of Astroloba and Poellnitzia at all in the context of the smaller Aloid genera. On the other hand if flower structure is so important as to suggest one genus, what does one do with Aloe striatula, which must have a flower unique in the Aloaceae, if not in the Order. There is one upper outer tepal and two lower outer! – the flower is by the way, zygomorphic too. It needs to be seen what the arrangement is of the inner petals is to each other – they are truly turbiniform imbricate.
The authors close with a statement of pollination saying that Poellnitzia is pollinated by the Lesser Double-collared Sunbird. It is well known that Haworthia and Astroloba, with their whitish flowers, are also visited by the same Sunbird. The thrust of the Smith/Manning note is that a specialised pollination strategy is not sufficient grounds for separation of Poellnitzia as a genus. Thus the paper directly contradicts Smith, who in a 13-page article (Taxon 41:437, 1992) postulated Poellnitzia to have a mite-dominated pollination syndrome. It seems to have been then overlooked that the mites are possibly even more difficult to move between flowers than the smaller pollen grains. It is just curious logic, because a specialised pollination strategy surely implies any degree of associated structural adaptations relating to separation of even families. Smith in both that paper, and in S.Afr.J.Bot. 58,90, 1992, claimed either that Poellnitzia had a unique floral morphology or that “strong macromorphological evidence” separated it from other Aloid genera. Obermeyer-Mauve (Bothalia 11:119, 1973) wrote a short paper on the small flowered Aloineae which is also quite erroneous (as evident in Bayer, Nat.Cact.Succ.J. 27:78, 1972, and refuted in Bayer, 1976). She tacitly supported a suggestion made by Parr (Bull.Afr.Succ.Plant Soc. 6:145, 1971) that Astroloba and Poellnitzia be included in Haworthia. She fallaciously argued for the inclusion of Chortolirion in Haworthia as well. Rowley (Cact.Succ.J.Gt.Br. 43,2, 1981) actually validated his proposal that Poellnitzia belonged in Aloe. Smith wrote another paper (Hazeltonia 2:74, 1994 – in which no less than 43 literature references, but excluding the text-book by Jeffrey, are cited) saying…”morphological differences … still readily serve to distinguish.” Poellnitzia, and that based on these it should form a distinct entity as a separate unispecific genus.
These articles by Smith et al, Smith and Manning, and Obermeyer-Mauve just demonstrate the ease with which the whole classification system can be disrupted, and the massive effort which is then required to restore a status quo. Neither protagonist is required to display any particular degree of expertise, and the botanist claims higher moral ground on the basis of science which has no more substance than that of religion. The actual unstated problem these botanists have is the embarrassment of monotypic (euphemistically now “unispecific”) genera. The word genus implies more than one anyway, and it would be more honest just to acknowledge the impracticality of unispecific genera, than parade a host of fallacious arguments to obscure them. The other even more unfortunate aspect is that all this fault-ridden argument passes over the heads of the unsuspecting community that is interested in the plants and wants to use the names. All that actually happens is that credibility becomes a very rare commodity in plant classification and nomenclature. If this is what professionals do, what can be expected from amateurs? The foundation is secured for the most uninformed and ignorant to parade as taxonomists – and they do.
The question of pollination comes into question because of the latest indignity. This is the transfer, described above, of the species to Astroloba by G.F. Smith and J.C. Manning published in Bothalia 30.1:53 (2000). There are many in congruencies in this half-page of text, and many more pages have to be filled to clarify this issue than should ever have been necessary.
The following claims are made in the Bothalia article:
“Vegetatively Poellnitzia shows…
a. – close affinity to some species of Astroloba“. Experienced and skilled botanists have confused Astroloba and Haworthia (at the Pretoria herbarium Astrolobamuricata was identified for Mrs. Bohnen as Haworthiavenosa), and Aloe and Haworthia (in the Compton Herbarium there is a description by a very experienced botanist, of a new species Haworthianatalensis, attached to a specimen of Aloearistata). P.V. Bruyns also brought me the first specimen of Haworthia pungens as an Astroloba. Many years ago I visited Miss Grace Britten at the Albany Museum and showed me a plant from the Longkloof. My reaction was “But that looks like Poellnitzia!” The discovery of Haworthiapungens demonstrates that there is this similarity of an Haworthia to Poellnitzia as well.
b. – and “accords completely with the genus in its tubular actinomorphic flower with included stamens”.
“Actinomorphic” and “zygomorphic” are terms used to distinguish structural differences in flowers. “Actinomorphic” means the flower is radially symmetrical and can be bisected into two equal symmetric halves in more than one plane. “Zygomrphic” means that the flower can only be so bisected in one plane – usually the vertical. I have made this point for Haworthia and Astroloba before. The flowers in these genera are equally zygomorphic. There is no basic structural difference in respect of symmetry. What then is the case in Poellnitzia? The fact is that Poellnitzia has the zygomorphic floral structure of most of the Alooideae. There are two upper outer petals and one lower outer petal. There is one upper inner petal and two lower inner petals. The margins of the outer petals are fused to the midribs of the inner petals, but the lines of fusion are clearly visible on the outside of the tube so-formed. The raceme is borne horizontally and the individual flowers are borne secundly erect. Smith in Bothalia 25.1:35 (1995) has properly described this, and states that the upper-third of the tube is decurved. This is absolutely correct and also contributes to the fact that the flower cannot then be actinomorphic! If the flower is properly examined it can be seen that it is in fact zygomorphic. Only the tip of the flower with the connivent petals, is actinomorphic… see point 3.
Curiously the zygomorphy of the white flowered Aloealbida is occasionally introduced as a red herring in the discussion of the classification of Haworthia. This zygomorphy is actually not in the least structurally different to that in the flower of many other Aloe species, including the four Reynold’s placed with A. albida in the Graminaloe and which he described as “regular” (actinomorphic!). The tube in A. albida is curved upward.
“A close relationship (between Poellnitzia and Astroloba) is supported by a preliminary survey of… aglycones in the roots of … Alooideae“.
How close? What is the position of Astroloba to the three Haworthia subgenera? Or to Chortolirion or Gasteria? This is not evidence as it is presented here.
…”careful examination (sic!) of the flowers, shows that the aestivation is in fact imbricate and the species thus differs from Astroloba only in the more or less horizontal racemes…”.
This is not what careful examination will show. Careful examination will show that in Astroloba the upper inner petal includes the two lower slightly revolute inner petals. For this reason alone the flower can never be actinomorphic. In Poellnitzia, the margins of the inner petals are separate for their entire length. Sectioning the tube at any point in its length will show asymmetry and the fact of zygomorphy as in Astroloba. The cross-section is that of an inverted triangle with the base shorter than the two sides. If the base of the flower is examined from the rear of the flower, the perigon can also seen to be slightly egg-shaped with the thin end of the “egg” below. The perigon is not round and thus cannot be actinomorphic. This is true for Astroloba, and also for Haworthia subgenus Robustipedunculares. To suggest that there is actinomorphic accord as described in point 1, is thus fallacious. Further, the stated similarity of tubular structure is absurd given the tubular nature of the flowers in the Alooidea generally.
Rowley (1976) refers to my claim that “the flower in Astroloba is positively not actinomorphic”, writing that to say so is.. “to imply that no Aloineae at all are 100% actinomorphic”. I implied and stated no such thing. I explained the position with regard to structural symmetry or asymmetry as above. The only implication is that which the facts of structure have for the truth and practicality of classification. The fact that Aloealbiflora is actinomorphic (the flower is not even really tubular) is a problem of its own as is Poellnitzia and many other aloid oddities.
“These floral adaptations are now recognised as part of the syndrome of sunbird pollination…”
The authors are getting to the point where they eventually will claim that Poellnitzia is best treated as a species of Astroloba adapted to pollination by sunbirds. The fact is that Smith and Manning fail to cite a long paper in Taxon 41:437 (1992) by Smith, where Smith virtually excludes birds as pollinators. A curious point too is that the Poellnitzia flower (like Microloma) in contrast to most Aloe, Gasteria and the general red-flowered, bird-pollinated species has secund (erect) flowers. The others have pendulous flowers. Is Aloe albida then just a anthophorine bee-pollinated Haworthia.
… “and the resemblance between the flowers of Poellnitzia and the bird-pollinated genus Microloma (Asclepiadaceae) is particularly striking”.
This is a remarkable statement. It is to my knowledge only Microloma tenuifolium which has a very bright shiny red flower, and it is not always this red. I would not personally in my wildest dreams have considered the flowers of these two genera from vastly different families as comparable even on the most careless examination. One could say that Microloma is best treated as belonging in the Alooidae because it is also adapted to pollination by sunbirds. But, worse. Is Microloma pollinated by birds? The Asclepiads have a complicated pollination mechanism generally and most are insect pollinated. The flowers of Microloma sagittatum are not even red and the tube is not comparable to that of Poellnitzia. They are pinkish and even partly white. Microloma massonii has a small greenish brown obscure flower.
“In the wild the species is visited by Lesser Double Collared sunbirds…”
This statement is followed by an unlikely description of how the birds insert their beaks and tongues into the flower. Smith gave a very comprehensive account of the possible pollination of Poellnitzia in his Taxon (1992) account where he concluded that bird pollination was improbable. In my own experience I have often seen Poellnitzia in flower in the wild. It flowers in March-April. These are hot dry months when the veld offers very little nutriment to anything and flowers of any kind are few. In response to Smith and Manning I have made a point of examining Poellnitzia in the field and my observations suggest that Smith should stay with his 1992 more careful thoughts. The only prospective pollinator or mite “phoretor” (carrier) was the large Camponotus ant as observed by Smith. The flowers were tended by individual ants which walked over the tops of the flowers and carefully “palpated” the flowers with their feelers. The ants would often pay particular attention to the base of the tube and we (my wife and I) saw that occasionally a single flower would bear a small redder abraded spot to which the ants returned. The would place their mandibles on this point while palpating the flower with their feelers. The tube was never penetrated and more significantly over the period of two weeks, we never found any nectar in the flower tubes. This is not like Haworthia which can literally ooze nectar.
There are two complications. One may favour bird pollination and the other ant/mite pollination. In my field observation (early April when it is still hot and it was very dry too) seed-set was intense. But it was irregular. There was a syndrome of racemes with complete seed-set of the many consecutively maturing flowers from base to apex. There were racemes where most of the lower flowers had set seed, with the upper approximate third not doing so. There were racemes with seed set on the lower flower points, a long gap of 8-10 flowers points which were sterile and the end ones still bearing flowers and buds. There were also a few racemes in full flower and bud. Something had thus interfered with seed set over a period of about two weeks. The other interesting point is that there is barely any opening at all between the connivent tips of both sets of petals. The stigma is borne 3-4mm below the tip of the tube, surrounded by the anthers. Pollen can occasionally be seen outside the barely open slits between the connivent petal tips. Smith reasonably suggests that mites are responsible for this. Are they also responsible for carrying pollen back into the concealed stigma? I brought flowers back with me and placed them in water. A few days later I observed some yellow mites on the outside of the flowers. They were very active and appeared to be seeking re-entrance into the flowers which they could not achieve. I had to pry apart the petal tips for the mites to enter. Yet in the flowers there were many mites. How did they get there? The ants are thought to need a nitrogen source (feeding on bird droppings for example). Do they use the mites as a food source and do they transport eggs or young mites between flowers? If this flower is adapted to a bird-pollination syndrome it is sending a strong message to birds that they are not welcome. One might as well say that the damage starlings and weaver birds do to Aloe flowers is strong evidence that the flower is adapted to bird pollination!
“The sugar composition of the nectar of Poellnitzia also refelects a shift from insect- to bird pollination…”
How does when deal with this rationale? One could use the same statement to separate the species from Astroloba. But I do not think when treating genera, one can exclude other genera from consideration in this manner and with this kind of argument.
“Adaptations for specialised pollination strategies alone are insufficient grounds for recognition of genera..”
The grounds for recognition of genera are as cited from Smith et al from Jeffrey. Smith and Manning have produced no facts by which Poellnitzia can be more closely related to Astroloba than to any of the other Aloid genera. All the facts have been distorted in order to reach an end, which is obviously to exterminate unispecific genera. Any structural character can, if so desired, be construed to have some or other adaptive significance. It is absurd to suggest that if these adaptations are directed at pollination, they are less significant for classification purposes. There is no connection of this kind whatsoever. A point is missed which I cannot adequately explain. Poellnitziarubriflora demonstrates a uniformity which could even suggest that it is vegetatively propagated. I have yet to see two clones which I think I could consistently separate. I have even collected what I thought was a smaller form, only to find it grow out to equate others in cultivation.
It is actually very difficult to refute argumentation of this class against the general background of the treatment of the Alooideae by systematists and would-be systematists. I say this because Rowley (1976) also contributes to the debate with an odd statement. “Very often one finds that minute floral differences are linked with supporting vegetative characters…”. He is in fact referring to my recognition of the subgenera of Haworthia on the basis of floral characters. He is actually quite wrong and is repeating the attitude which Dr. L.E. Codd expressed in private communication to me when questioned about the amalgamation of the smaller Aloid subgenera. Dr. Codd said… “We are ignoring the small differences”. This is not true. Because, firstly, these writers are confusing the small size of the flowers with the significance of the differences. Were the flowers large, the differences would be more unquestionably seen to be large. The irony is that here in Smith and Manning, we have the case that more obvious large differences are discounted. Secondly, when a judgement of similarity or dissimilarity is made, the observer in fact makes a decision and then seeks a rationalisation. What are currently seen as Haworthia are three subgenera which have historically been judged to be similar and the rationalisation has never been properly constructed or questioned. The floral differences have been minimised. Capsule and seed structure has been ignored, and no attention has been paid to anatomy or things like root morphology. I have not done so either. The reason for this is that it is already obvious to me that the rationale already exists for the classification that I adopt viz. 3 subgenera, and one that I would propose viz. 3 genera. It is thus pointless to pursue other goals when proper attention is not paid to evidence already presented and which alone constitute grounds for the recognition of genera. But I do not think genera are real taxa in the sense that I see species to be. I feel that a classification arrived at over a period of 200 years has an accrued value. It is pointless to make changes which are not properly weighed against that value.
Rowley ends by sensibly making the point that the question of genera is artificial. I am very sceptical of his sanguine hopes for some future ideal solution when the … “time will be right”. To assume that botanists using sophisticated technology (DNA, molecular, cellular etc.) or statistical methodology (cladistics and multivariate analysis) are going to produce better classifications will be unwise. Botanists have not effectively used the available simplest of evidence to classify or discuss the classification of the Alooideae, and amateurs are exacerbating the situation. Botanists are playing with technology, and with classification to display the results of this game. Amateur and impostor taxonomists are playing with classification to display their grasp of technology and knowledge. The motives are questionable, and the results prove it. ♦
Fig. 1. Poellnitzia rubriflora, Middelplaas, Bonnievale, March 2002.Fig. 2. Non-uniform seed-set.Fig. 3. Camponotis ant at flower.Fig. 4. Microloma sagittatum. Not a look-alike.
In 1943 Dr. J. Luckhoff, who was better known as a collector of stapeliads, wrote up some observational and collecting records of Mr. M. Otzen after whom Haworthia otzenii was named. Most of these records have been followed up at one time or another and we know what most of the plants and populations were that he recorded. However, there is one particular item that remains a mystery. One of Otzen’s records reads like this:
”About six miles north of the six or seven houses forming Infanta there is a large salt-pan on your right … Four miles further south follow a narrow track on your right, which ends on a bluff on the foreland called Cape Infanta. On the southern slope towards the sea a very small Haworthia, almost black, with longer, narrower leaves than H. retusa, very scarce, and very difficult to find.”
Otzen does say for another record north of Bredasdorp that the small black species growing there is “similar to the Cape Infanta one”. This Bredadorp record is referable to H. maraisii, and I infer that the plant at Cape Infanta must be this species too. Nevertheless this small black plant at Infanta has bothered me for years. I did look for it in 1972 on the only excursion I ever undertook to this fairly remote area, but I mis-read the Otzen note and searched in vain on the rocks south of Infanta. While writing a review of my collections, a plant Adam Harrower of Kirstenbosch had given me started to create some problems. It did not fit the pattern of “flow” that I had so far observed for haworthias. Harrower’s record was for the Potberg Mountain where I had already obtained plants that were odd in one way or another. When I finally asked Adam for the detail of his collection, it appeared that he had collected it only about 10km from Cape Infanta. The plant is similar to that of one collected by the late C. Burgers of the Cape Department of Nature Conservation, at Buffelsfontein 20km further west. Both of these localities are on the south side of the Potberg, the vegetation is fynbos and the geology is Table Mountain Sandstone. My assessment of the two collections was that they were somewhat similar to H. mirabilis var sublineata. This is not a comfortable conclusion because of the geographical intercession of H. rossouwii var petrophila just west of Bredasdorp, and of H. rossouwii var calcarea from near the DeHoop residential complex. There are also five other populations recorded from the northwest corner of the Potberg but which have only an indirect relevance to the thrust of this article, so I exclude them. In fact I also exclude them from a magnum opus that I am preparing because the problem is so extensive and complex.
The question of the identity of H. maraisii is something that I am trying to answer in a separate rather long manuscript. There I will discuss a whole suite of species and the end product will probably be too long for journal editors or readers. The crux of that presentation will be that species names do not actually convey the information that we expect them to, and that collectors are caught up in a confusion of names that have commercial, egocentric and any other meaning but that within the orderly one of an informative botanical science that we unthinkingly expect
What I am considering and trying to explain in that big work in preparation, is that there is no distinction that one can actually make between all the variants of H. mirabilis and of H. maraisii (and more) that I recognize in my revision. Down at Infanta H. maraisii tails away to a tiny dark plant through a series of populations that probably confound any perception that only one species is involved. The problem is compounded by the fact that juxtaposed on either side of the Potberg Mountain are two plant populations. One could be H. mirabilis, the other H. maraisii. This confounds my argument because in the context of my longer manuscript I show that these are one and the same.
Thus Otzen’s record takes on a slightly larger dimension and I wanted to explore further south and east of the juxtaposed populations to see if any connection could be found. This found my wife and me at Infanta where J.D. (Kobus) Venter later joined us. We did find new populations. In the one collection the plants are small and dark with very erect leaves and similar to those cited in my Revision (1999) under the name H. maraisii var. maraisii (in which the leaves are classically retuse, ‘bent-back like a thumb” to use an apparently obsolete meaning – but see figures). The collection cited in my Revision is one by P.V. Bruyns from the east bank of the Breede River opposite the farm Ziekenhuis (Fig.1 JDV87-50. Kobus Venter found the same thing on the west bank at Jakobsrivier (Figs.2a & b JDV92-85). My new collection (MBB7250) is similar and about 1km east of that. The geology at this locality is rather hard to fathom and seems to be recent (quarternary) riverine or marine deposit overlying Ecca shale. The vegetation is also rather anomalous and would probably fall under Renosterveld. It is the second (MBB7248) and third populations (MBB7249) that stunned us, because the plants revert to being larger in size, with fully retuse leaves having an end-area level to the ground. Furthermore the end-surfaces are spinose (Figs.3 MBB7248). Such spinescence occurs in H. emelyae var. major and also in a population of H. magnifica just east of Riversdale. It does, however, also occur to a lesser degree at both population- and single-plant-level in many other populations. As an aside, it is actually possible to suggest that here we have the convergent emergence of a species character independently in another distant population. The geology at these last two sites is sandstone and the vegetation Fynbos.
Fig. 1. JDV87-50 H. maraisii=heidelbergensis. Ziekenhuis, InfantaFig. 2a. JDV92-85 H. maraisii=heidelbergensis. Jacobsrivier, InfantaFig. 2b. JDV92-85.2 H. maraisii=heidelbergensis. Jacobsrivier, InfantaFig. 3. MBB7248.3 Ballyfar
When Kobus joined us the following day, Keith Spencer and his staff of the De Hoop Nature Reserve accompanied us to Cape Infanta. Keith was naturally interested to know what we were looking for as well as serving the supervisory role for the Reserve. Cape Infanta is a windy bluff along a rocky shore where the rocks rise almost directly from the sea to a height of about 30m. The geology is quite complex and is mapped as calcarenite commonly called Cape Limestone. The vegetation was very typically Fynbos although there were many mesems too. We did not find any haworthias after quite a thorough search and we gave up the attempt deciding rather to trace Adam Harrower’s record. We did this without too much difficulty though we wondered why Adam should have thought the locality (near Sandhoogte) was promising for any odd things he was looking for in his search for herbaceous plants for horticultural screening. The locality is a rocky (sandstone) ravine bank with Fynbos vegetation. Adam’s plants were quite common but not all that easy to find. Although I make the connection to H. mirabilis, I also feel that there is some justification for calling in H. rossouwii var. calcarea for comparison (Fig.4a ADH594, & b MBB7251). One of Keith’s rangers then maintained that he had in fact seen a similar plant at Cape Infanta and so we retraced our steps. We searched in a different direction but still failed. Reading Otzen’s notes, one is filled with a sense of depression because it is apparent that the collection may have had commercial undertones. It is known that Triebner was purchasing plants from collectors and Otzen’s one record reads…”Doubt whether there are any left after Venter, Werth, Geyer, Malherbe and I have been there.” Another reads, “three of us searching for more than an hour, found only three specimens – doubt that there are any more.”
The one common binding factor in these populations we saw is the fact that they were in flower and beginning to fruit. The southwestern Cape haworthias seem to fall into two groups on the basis of flowering time viz. a spring-flowering time, and a late summer- flowering time. I wrote an article for Aloe (40:10, 2003) in which I discussed ecotypes of haworthias on the Zuurberg Mountains. I have to conclude that this is probably what we are seeing here, coupled perhaps with a dispersal history that has carried plants (genetic material) from the south and west, and independently from the north and east. However, it is more complicated than that. The resemblance of the Harrower collection to H. turgida var. longibracteata is noteworthy. Harrower’s population juxtaposes our spinescent find and only 3km separates them as the crow flies. Burger’s collection at Buffelsfontein is juxtaposed by only 5km with H. turgida var. longibracteata at Diepkloof. We should have gone on to examine the Buffelsfontein population as my confidence in declaring it to be the same as that at Sandhoogte was only based on a herbarium specimen. Co-incidentally, there is also a specimen collected by Prof. Compton that has a close likeness to the Buffelsfontein plant. Compton’s record is remarkable for the statement that it was collected on “the Potberg sandy flats”. There is so much “sandy flat” surrounding the Potberg, that the re-discovery of this population will be purely fortuitous. We did in fact get a later opportunity to explore Buffelsfontein and find Burger’s population (Fig.5 a & b MBB7374) and my conclusion is that it is a continuity of MBB7248 and ADH594. In the light of Chapter 15, I can hazard a guess as to how this really fits in.
Our excursion did not therefore resolve any problems and instead seems to have added to the complexity of the situation. I have been writing for a long time about haworthias and have been finding it increasingly difficult to precisely and concisely explain the ramifications of all these new things that keep turning up. I have never thought that simply generating new names is going to add anything to our knowledge of the plants as a group. My feeling is that a good classification should communicate something about the plants. It is therefore exceedingly disheartening to read a quotation from the book by D.A. Levin The Origin, Expansion and Demise of Plant Species (Oxord University Press, ca 2000). He quotes authors Raven, Berlin and Breedlove, who say… “our system of names appears to achieve a reality which it does not possess”. Levin himself writes…“our taxonomic system communicates little about the organism being considered, although it appears to communicate a great deal”.
Levin discusses the species concept at some length and I doubt if I could debate the subject at his academic level. My contention is that he makes much the same mistake as all these people who discuss the matter – they do not define the word “species”. Levin’s introductory chapter is a discussion of premise and concept and there are many sentences that can be taken to represent fragments of a definition, but he ends it by saying that “the choice of concept has to do, in part, with the perspective that gives one satisfaction”. I find this alarming because to my mind it leaves the definition open and subject to interpretation and whim. We will never have even crude agreement of a classification if each of us follows the matter to one’s own satisfaction. To some extent this objection is overcome by his closing statement that: “it is important to understand the biological properties and relationships of population systems rather than get hung up on putting them into conceptual pigeonholes”. My only response to this is that we should try to understand these properties and relationships in the process of conceptualizing such pigeonholes.
This brings me to a closing argument. In Avonia 21:45 (2003) Breuer describes six new species and six new varieties of Haworthia. Only one of them is meaningful to me; the others all fall into my existing conceptual pigeonholes derived from a definition of species that have genetic, morphological, geographic and behavioral similarities. My dissatisfaction is that the descriptions have as their one visible goal, the need to satisfy or nourish the acquisitive urge of collectors while neglecting – even negating – any obligation that these descriptions should possess or add meaning. This is the same argument I would put forward for any name changes. If they do not add or contribute meaning, they should not be made. Names should convey more than a collector’s item that is in essence only a source of income to a commercial outlet and an expense to a collector. Another goal may simply be the egocentric one that builds on this false notion of “meaning” and “discovery” associated with names, which of course could be satisfying to the author too (and of course the collector commemorated in the name).
What is the exception in those 12 new taxa of Breuer’s? It is H. elizeae said to be from west of Swellendam (Figs.6a, b, c, d, & e MBB7255). The plant was it seems first collected by Derek Tribble in 1977. According to Breuer it was apparently also collected by E. Esterhuizen and by V. deVries and the latter is credited in the citation of the type (deposited of all places, in Tokyo). It is quite strange that the plant is otherwise unknown except that so many places are unexplored in the sense that anyone can barely know what is new or common on any site. This particular site is within a stone’s throw of Stormsvlei that is a common collecting ground for Haworthia. On our way home from Infanta I thought I should try to examine the population. The resemblance I could see while looking for information and meaning in the description and illustration, is one to H. rossouwii (Fig.7) which species has a very odd broken distribution. There is also some resemblance in it to H. emelyae var. multifolia too. Breuer’s description in Avonia gives no meaning at all, other than to vaguely suggest a western extension of H. turgida and an equally vague comparison of the flower, viz. “the floral characteristics look a bit more like those of H. triebneriana than those of turgida”.
Fig. 6a MBB7255 H. elizeae. Stormsvlei.Fig. 6b MBB7255 H. elizeae. Stormsvlei.Fig. 6c. MBB7255 H. elizeae. Stormsvlei.Fig. 6d. MBB7255 H. elizeae. Stormsvlei.Fig. 6e. MBB7255 H. elizeae. Stormsvlei.Fig. 6f. MBB7255 H. elizeae. StormsvleiFig. 7a. MBB7299.3 H. rossouwii. NW Soutkloof.Fig. 7b. MBB7299.2 H. rossouwii. N Soutkloof.
Given the locality as “25km west of Swellendam in stony outcrops”, we actually struggled to find the plants. Fortunately Derek Tribble provided a better description of the site. It is about 30km west of Swellendam and is not “stony outcrops”. It is a significant band of finely textured Table Mountain Sandstone abutted by Ecca shale. The population occupies a surprisingly small area comprising the rocky foreground in the two illustrations (Figs.8a & b). As the ridge continues east it rapidly becomes coarse-grained sandstone and is the home of H. mirabilis in the form that this species has in the nearby Stormsvlei Pass. (Using the name H. triebneriana for that population as Breuer apparently does, removes information from the system as will be evident when my review of that system is published.) H. elizeae is indeed different. It is a highly proliferous clustering plant and flowers in spring, or certainly earlier than H. mirabilis. It does not comfortably fit anywhere in my pigeonholes and I see the need to uphold the name in the way I uphold my own H. pubescens. I see the need to explore further, to see if the distribution can be extended in anyway. However, it seems very unlikely. I did not see any resemblance of the plants in the field to H. rossouwii as I had surmised from the illustration. But I do not think the possibility of a link to H. rossouwii should be totally excluded when anyone comes to trying to explain the question “how did it all happen?” Perhaps there is a closer resemblance to H. maraisii var. meiringii at Bonnievale where I drew a comparison with the spring-flowering H. herbacea. I also consider that H. maraisii var. notabilis (Fig.9 JDV98-1) at Robertson and Klaasvoogds could yield variants which parallel H. elizeae, and this is also possible true for H. mirabilis var. consanguinea from MacGregor. But these are all summer flowering.
Fig. 8b. Habitat of H. elizeae. Stormsvlei.Fig. 8a. Habitat of H. elizeae. Stormsvlei.Fig. 9. JDV98-1 H. maraisii var notabilis. Wolfkloof, Roberson.
The message I would like to convey in this article is my belief classification must have meaning beyond that of items on a price-list or in fact on any list. My feeling is that botanical science has deluded and corrupted us all by the pursuit of classification as a cataloguing process instead of a methodology based on solid definition and purpose. Not in every instance, of course, and not to the extent that all taxonomists are culpable, but there certainly is an abuse of the system. This is perhaps what Levin refers to when he quotes Ehrlich and Raven who write of “a good species…an artifact of the procedures of taxonomy”. It is not surprising that many readers have become exasperated with taxonomists and “namenklutter”.
Acknowledgement I state that my collecting was under accreditation of permit 45/2003 issued by the Western Cape Nature Conservation Board. I would also like to acknowledge the co-operation of Mr. Graham Lewis and Mr. Keith Spencer of De Hoop Nature Reserve for their assistance, Mr. Adam Harrower, Mr. John Douglas-Hamilton, Mrs. Christine Wallace and Mr. Hanrico Cilliers. Mr. Ingo Breuer kindly sent me a copy of Avonia 16. Messrs. Bob Kent, Steven Hammer, Elliot Adler and Paul Forster kindly commented on the text of my article where idiosyncrasies remain unfortunately my own.
Addendum Flowers of MBB7255 H. rossouwii var. elizeae, Bromberg taken on 1 December 2012. ♦
The systematics of Haworthia and the production of a practical and useable list of names that reflects a predictive classification for these plants, has kept me preoccupied for more than 40 years. Initially I simply produced a list of names published in 1976 as Haworthia Handbook. This was not a great work but there was a desperate need then for some kind of conspectus that established order where there was none. I rewrote the Handbook published in 1982 as The new Haworthia Handbook. This was followed by C.L. Scott’s The genus Haworthia, A taxonomic Revision (1985), which effectively snatched defeat from the jaws of victory. In 1996 I was asked to produce a synopsis of Haworthia for two projects, and Steven Hammer and Kobus Venter persuaded me to formerly revise the genus. It should be explained that the greatest obstacle to such a work was the very difficult question of the validity and priority of names as required by an international code for plant nomenclature that sets out to standardize and lay down norms for this process of typification. The problem in Haworthia is that the illustrations and specimens on which the process is based, were and are, so confounded. Either there were no specimens at all, or the basis of the names existed only as barely identifiable illustrations. An additional difficulty is that specimens from different species so often resemble each other, that unless locality data was available, there was often no certainty to what the name actually applied to. Thus most of the available types were ambiguous and subject to alternative interpretations. This is a self-evident truth. The overriding consideration was to give meaning to the names in terms of biology and to give an objective reality to the different kinds of Haworthias that are the basic units of biological diversity i.e. species. This latter objective seems very mundane and straightforward and it is extraordinary to now recognize what a minefield of discussion, dispute and argumentation is appearing in the literature concerning the subject.
With nomenclature being so confounded, I had in my Handbooks not attempted to formalize the application of names, but rather to cite specimens to make it quite clear on what my usage was based. The application of names to specimens preserved in herbaria is a base-line requirement in systematic botany, but consistently appears to be a difficult concept to persuade eager amateur botanists to adhere to. At the same time I tried very conscientiously to get as close to a realistic appraisal of the sources of all the names with respect to the fundamental types required by the code. With the decision to prepare a list of names for Urs Eggli’s Illustrated Lexicon of Succulent Plants, and almost simultaneously the decision to rewrite the Haworthia Handbooks, the formalization of names became inevitable. Enter disaster. I. Breuer in Germany had become inspired to undertake the same task. He presented me with a manuscript of his The World of Haworthia – Vol.1 with a request for an introduction and intellectual sponsorship. It was evident to me that he had no more capacity to do the work any more effectively than Col. Scott or myself and there was no time to negotiate a solution. I declined. Breuer with Dr. Detlev Metzing had already submitted a formal typification of Haworthia names to the botanical journal Taxon and the die was cast. Of course it was a logical step for them and it is recognized in taxonomic botany that one can approach a revision in two ways. The first is to establish the validity and priority of existing names and then proceed to the field to establish how they can be used. The second is to proceed to the field and then only decide how best the available names can be assigned. The advantage of the second method is that it grants one the grace of being able to better interpret ambiguous types and to generate some sense from poor original illustrations. It also allows one the opportunity of fully appreciating the history of usage, past and present; and then properly formulating the way in which these names can be used with predictive value into the future. All of this assumes that there is a solid understanding of biological diversity and the nature of the classification process with respect to “species”. It was evident that Breuer and Metzing were not competent to undertake their mission with respect to either option and definitely not for the first.
In the introduction to my second Handbook, I wrote, “The complexity of what species are, is beyond the object of this book…”. It is very difficult for me to explain my thoughts on the subject against the background of the extraordinary statements now being made in the literature with regard to species concepts. The advent of molecular biology and the analysis of DNA have suddenly hit taxonomic botany as a revealed truth, and the vision of a true evolutionary history of plants is seen as an attainable goal. Having been educated in the environment of animal science it never occurred to me that there was any other objective than this in classification. Hence the ideal that DNA study now encourages viz. a phylogenetic species concept, is thus in my estimation simply a very damp squib. The fact that botanists have never had a clear and unobstructed definition of the species which they have been identifying and naming so industriously for so long is only evident now from the convoluted discussions about the reality of the concept and the serious proposals to even abandon the binomial system of names. The juxtaposition of the words phylogenetic, species and concept is not my construction. It is one by botanists who have several other species concepts and yet no precise definition of the one word “species”.
My Revision Haworthia Revisited, drafted in 1996, was printed in 1999. The text for Eggli’s Handbook was drafted earlier in 1996 and only published in 2002. There are some minor discrepancies in these two works, but far the greater confusion has been the impact of the typifications by Breuer and Metzing and the subsequent publication by Breuer of a series of books and articles that add fuel to the fire. Other authors and their editors seem to have taken every advantage of the existing confusion to make matters worse and fan the fire into a runaway inferno. The inevitable result has been a relentless struggle on my part to maintain some kind of credibility on behalf of the plants I am interested in, and also for those people who I imagine present my reading public. It can be likened to sitting at the control panel of a rocket fired wildly at the moon by a sightless scientist, and then firing directional rockets by remote control and a faulty keyboard, to get the act together and the rocket on course.
What has emerged now in one of the house journals of the National Botanical Institute of South Africa, Strelitzia 14: 983-1017, is an annotated checklist of the plants of Southern Africa with the section for Haworthia compiled by Ingo Breuer, E.M.E. Steyn, G.F. Smith, and N.L. Meyer. There is no doubt very good reason why my participation was not sought considering that I have curated the NBI herbarium collection at the Compton Herbarium in Cape Town, effectively since 1970. The most telling one is perhaps, facetiously, the assumption that I was senile or dead. Surprisingly the list of names is remarkably true to my classification, in stark contrast to the surprising and odd opinions maintained by the principal author in his prior publications. It is not clear quite when the list was drafted, but the book appeared in 2003.
Superficially the Strelitzia list seems to be very authoritative and certainly is a vast improvement to a similar earlier listing. That was a ridiculous compound of both Col. Scott’s classification and my own. The reason for that mess is simply explained in the miasma of a classification activity that had, or has no standard and by which any ambitious individual can assume or be granted credibility as an expert merely by publishing new names, irrespective of whether they reflect real taxa. However, the new list does have problems and a large measure of incongruence. My classification is founded, within the limits of my intellect, on a thorough effort to get as close as was reasonably possible to “how it could all have happened”. Where the list seems to follow my classification, I must therefore accept it as sound. However, the introduction of “new species” described primarily by Hayashi, makes a mockery of the intended all-encompassing and predictive value of my classification as a whole. This is a wholly different set of problems added to those already generated by the Breuer and Metzing typifications and associated interpretations and applications. I am just very grateful, as every other Haworthia enthusiast should also be, that Breuer has been seemingly constrained by his co-authors, to abandon many of his other radical and misconceived opinions or at least conceal them for the purposes of the publication.
The prime changes I would make to the list are:
Haworthia pumila (H. maxima). I did introduce the use the name maxima in the manuscript version for the Eggli publication and I seriously doubt if any author would have ventured to promote this name in the absence of myself as the “fall-guy”. While the name pumila is inappropriate, it is the earliest name for this Haworthia and the late Dr Onno Weinands is credited with the formal typification of the name on the basis of the same illustration on which maxima is based. Further debate on this issue that has already consumed so much paper and ink seems ridiculous.
The introduction of the name integra exposes the flawed nature of the whole list when it comes to the key problem of how these names are USED. There is a massive interplay of what I have recognized as two distinct species. These are H. arachnoidea and H. mucronata. I have written about this problem at length in many places and I cannot go on and on presenting the same arguments over and over again. The essence is that one has to recognize that “species” in Haworthia are not separate elements in the way in which we want to segment them for any purpose. The type specimen that Breuer has triumphantly unearthed is a pickled one found in the Berlin herbarium. It is quite evident to me that it has the darkened leaf-tips of plants that I know very well from Lemoenpoort near Barrydale, across to the area between Riversdale and Ladismith, which Dekenah patrolled so thoroughly. These glabrous elements generated the type material for the original concept of H. venteri and the entire ramification that culminated in my decision (among others) to regard the name integra as ambiguous and to exclude it as insufficiently known. Were I to reconsider this decision in the light of this new type, I would concede that it relates to H. arachnoidea var. nigricans but as nothing more than another problem variant. Here again, fussing over this minor problem when it is difficult enough to separate the major components in the puzzle is just incongruous. I see no need to refute my variety rycroftiana or to recognize Esterhuizen’s standeri. I make no pretensions that my system has any greater virtue than is possible in the face of the real nature of field variability and my capacity to sustain some kind of consistency across the genus as a whole. The name H. mucronata var. rooibergensis is similarly upheld in the list when I know very well that in this interplay of the two species aforementioned it is not possible to uphold any such small distinctions – what to say of the problem extended eastward to the “species” there.
One cannot pass this issue by without back reference to the uses of names. There is no doubt that the way in which Breuer would use the name aristata would reflect the confusion generated by Col. Scott when he had several species names such as mucronata, aristata, mclarenii and unicolor in different sections for the same one complex in the vicinity of Barrydale. I was inspired to think that the name aristata could more effectively be used for a dubious group of populations in the H. cooperi complex of the eastern areas. The list is in fact unsatisfactory because it excludes the changes that I was constrained to make when I wrote Haworthia Update in 2001 to describe and explain the results of my further exploration. These authors have been confused over elementary things like the zygomorphic structure of the flowers in the minor genera of the family, the spiral arrangement of leaves in H. viscosa, or the fact that the illustration of rossouwii cannot be equated with a depauperate specimen of serrata, and I have my doubts that they are able to apply the names they have listed, with any greater sensibility. The last example is replicated many times in Breuer’s books where there is incongruence between type material and added images of other plants that are taken to represent the same entity.
What is now left is the really curious array of one-off names that are paraded as “species”. They are in total conflict with my entire conceptual view of Haworthia and it is quite senseless to include my name anywhere in the list other than with the words “sensu Breuer et al” appended if this is the paradigm for which the list is required. I use the name geraldii as the prime example. If classification is the expression of a purely personal opinion, and many leading writers amazingly tout this nonsensical point of view, then of course one can generate new names in any way one chooses. The fact is that I generated a system of names with a view to explaining a complex system that includes ALL haworthias and relates to my experience with many other genera, both plant and animal. My system was intended to serve science and to efface my own predilections and opinions as far as it is possible to do so. Geraldii is a name that owes its origin to Col. Scott and it relates to one single population where it is not even clear that the plants are non-vegetative in origin. Similarly the odd agnis refers to a single population of the greater complex nortieri – a local ecotype. All the names generated by Hayashi fall into this same category of names generated purely by personal attraction to a particular variant and for self edification (viz. H. hayashi Hayashi), which can actually quite easily be accommodated in a descriptive statement of a far more predictive system. The hidden part of the floating iceberg, which this checklist is, is a barrage of new names being generated by Breuer, among others, before the print has cooled. This kind of classification has eventually to sink from its own weight and will do so when any real and honest attempt is made to apply it to the great range of ecotypic variants and continuities that are such an obstacle to our sensibilities. My few loyal friends suggest to me that I should just ignore these nomenclatural and descriptive incongruities and leave it to history in the belief that “truth will out”. I appreciate the sense in this remark, but the reality is that I am in the present and would prefer to experience commonality of sense now too. I plead that my system should stand in its entirety with no changes until these can be soundly supported by good technology, good statistics, and good appreciation and observation of the actual natural system. A last requirement is that botanical classification finds a way through proper definition, purpose and communal agreement to arrive at a standard of names for all living systems. ♦
Abstract Classification of plants is often controversial and it is common to suggest that further research and application of technology will resolve problems. This paper overviews six publications in peer reviewed botanical journals with respect to the classification of Asphodelaceae: Alooideae and particularly Haworthia. It demonstrates that there is a gulf between the results produced by researchers using sophisticated technology and the practical, ordinary observations of the layman. This may be because researchers are not familiarizing themselves with grassroots information and observation, which is observation of the plants in the field, in cultivation, in the herbarium and in the literature. Their results may thus be contrary to the experience of the layman who may in fact be better informed.
Introduction The classification of Haworthia has quite a turbulent history. This is generally attributable to the fact that it is a difficult genus in terms of the vegetative plasticity of individual plants and the many populations that have a shared history in time and space in geologically and geomorphologically diverse environments. Confounding these environmental variables are the concepts and techniques of the taxonomists that have been involved in naming plants within the genus. The popularity of Haworthia with succulent plant hobbyists has meant that many of these workers have by-and-large not been professional or qualified taxonomists. Therefore it is of considerable interest to note some significant publications in respect of the broader subject viz. the Alooideae, written by professionals and recently published in peer-reviewed botanical journals. This could confirm whether or not the problem lies with the difficulty of the subject and/or skills of the respective researchers or taxonomists.
Discussion The first publication of six that I review here is by Vosa and Bennet (1990) with respect to Gasteria. The general conclusion was reached that the cytological differences are minimal and that there may be only one species. In this work the nomenclature and terminology is inconsistent and the data are also presented in an inconsistent way. Furthermore, the material used is not representative of the genus (Bayer, 2000). The paper very effectively demonstrates the need for a sound organization (classification) of materials before work commences. There was no revision in place prior to the work by Vosa and Bennet and there is no statement to say how their material was identified nor by whom. They concluded…”The slight variability is probably not more than that found between the individuals of a normal natural population of a single species”. A correct conclusion would be that the technique employed produced no meaningful understanding of the genus.
A second paper is that by Smith & Van Wyk (1991). This is a cladistic study of the genera in the Alooideae. It is concluded that it is a monophyletic group and suggests that some clarity has been brought to the relationships of the genera. It can, however, be shown that the characters used and the evaluated states of those characters, are erroneous and that the results are consequently misleading. The following is a brief synopsis of the characters and coding (plesiomorphic – derived or advanced character state – 1; and apomorphic – residual or primitive character state – 0) applied by Smith & Van Wyk in their cladistic analysis:
Habit (caulescence) – all genera are scored 1 for this character except the outgroup Kniphofia. According to G.W. Reynolds (1960), Aloe has 130 species in southern Africa of which 30 are acaulescent, 56 caulescent and 47 may be neither of the two states. In my observation of 68 Haworthia species, 58 are acaulescent and 16 are semi-caulescent. Among the former are species like H. pumila and H. herbacea that can form quite distinctive, if prostrate, stems. In H. cymbiformis, which is generally acaulescent, there is the surprising var. ramosa, which is caulescent. It is part of an extensive population north of Wooldridge where the plants are otherwise acaulescent. If the condition is examined in the other genera such as Gasteria and Chortolirion, it seems that the coding used by Smith and van Wyk is simplistic and incorrect.
Leaf arrangement (congestion on stem) – Aloe and Lomatophyllum are scored 1 as each genus is said to have species in which the “leaves are comparatively widely spaced” (others are scored 0). Gasteria rawlinsonii has leaves which must be regarded as “widely spaced”. It can be said that in Haworthia, because the leaves are spirally arranged, it is differences in leaf congestion that result in trifarious, quinquefarious and multifarious leaf arrangements. Thus there is a range of spacing and in fact H. coarctata and H. reinwartdtii can be separated by the difference in the congestion of leaves on the stems of these two species.
Leaf consistency (succulence) – all genera are again scored 1. It is obvious that this is again a simplification. Even Uitewaal (1947) suggested that leaf consistency could provide a basis for separation of Haworthia into the three groups he proposed. In the Hexangulares there are species with the added state of having fibrous leaves. The consistency of the leaves of H. graminifolia cannot be equated with that of the case of, say, H. mucronata.
Leaf tuberculation – Astroloba, Gasteria and Haworthia are scored 1, while A. aristata is noted to be tuberculate as a “rare exception” in Aloe. My observation in Haworthia is that 26 species have tuberculate leaves and 42 have smooth non-tuberculate leaves. It is common for H. marginata to have both tuberculate and smooth leaves and this is less rarely the case in other species of the Robustipedunculares. In H. limifolia, the var. gigantea has tubercled leaves, the vars. ubomboensis and glaucophylla have entirely smooth leaves, while vars. limifolia and arcana are characterized by transverse ridges or semiconfluent tubercles. The state in A. aristata adds a complication as the “tubercles’ may be homologous or comparable with the surface spination, translucent or simple white markings on the leaves of many species in many of the genera.
Outline of leaf cross-section – all genera are again scored 1, as of course, against the outgroup (Kniphofia). Reynolds (1950) states that in A. cooperi, “leaves are keeled resembling a “V” in cross-section, and mentions this for A. variegata, A. sladeniana and A. dinteri too. It is quite misleading to suggest that leaf outline in the Alooid genera is crescentiform/cymbiform and synapomorphic for all the taxa in the Alooideae. Note the confusion of the terms outline as described by “cymbiform” and cross-section. How can one consider the shape of the leaves (cross-section or outline) of, say H. viscosa as even similar to the leaves of for e.g. H. cymbiformis var. obtusa? In total the two authors code sixteen such characters and it is beyond the scope of this paper to tediously discuss each of them. The question of the character “floral symmetry” does, however, perhaps deserve attention and that will be discussed in the fourth paper.
The third paper is by Smith et al. (1995). It concerns the reversal of the very curious re-instatement of the genera Aloinella (as Lemeea) and Guillauminia affected by Heath (1993, 1994). Heath’s argumentation was weak indeed, but the counter argument by Smith et al. is equally poor. The balance is only in favour of Smith et al. because there are no substantial criteria that suggest that different genera are involved in the first place. (Bayer, 2003a). My complaint now is that the papers by both Heath and Smith et al. fiddle with the classification and nomenclature, without adding one iota of information to the subject. The facts are:
A. haworthioides and A. albiflora do in fact have characters which make them most unusual in the genus Aloe. There is no Aloe remotely like A. haworthioides that has the extraordinarily protruding flattened filaments. These form a reddish tube nearly as large and conspicuous as that of the perianth itself, which is almost hidden by the large floral bract. In the case of A. albiflora, the florets are widely campanulate and sparsely arranged on the slender peduncle. Reynolds placed 12 species together in his Group 1 of Madagascan Aloes and stated most plainly that they were not closely allied. Now Heath has ignored three of those 12 species and made two groups out of the disparate remnant. This is the problem that Smith et al. should have addressed. The curious thing about this is that there appears to be a similarity between A. bellatula and A. perrieri (ignored and abandoned in Aloe by Heath, and not even considered by Smith et al.), which in terms of variability could be extended to A. albiflora and perhaps further to A. parvula. There is no such similarity between A. haworthioides, and A. bouteaui. A. descoingsii and A. calcairophylla do seem to have unique florets. There is nothing odd about the florets in A. rauhii or A. bakeri that could necessarily separate them from, A. versicolor and A. parallelifolia.
The fourth paper concerns the genus Poellnitzia and its transfer to Astroloba by Manning and Smith (2000). Again the argument is weak, particularly in respect of the statement… “accords completely with the genus in its tubular actinomorphic flower with included stamens”. The flowers in neither genus are actinomorphic and it was, and is, the elongation of the tube as well as the connivent petal tips that precluded anyone from suggesting that Poellnitzia was an Astroloba in the first instance (Bayer, 2003a)
The next paper is by Smith et al. (2000) and it concerns nectar sugars particularly with respect to Haworthia. Smith et al. cite my paper (Bayer, 1972) where the main message was that the genera in this group would never be resolved while the main elements within the genus Haworthia were not recognized to be discrete. Uitewaal’s (1947) attempt to subdivide Haworthia was a much labored effort. He was clearly a victim of his time in trying to establish a hierarchical classification and it is a distortion of the facts to say that he divided Haworthia into two main groups. There is no difficulty whatsoever in recognizing that he identified and recognized THREE, and not the two that Smith et al. claim. Uitewaal recognized two groups; Triangulares and Hexangulares based primarily on the shape of the flower base, but he then split Hexangulares into Robustipedunculatae and Gracilipedunculatae, primarily on the robustness of the peduncle. As an interested observer of Haworthia, I added the additional facts from geographical distributions and capsules to recognize Uitewaal’s groups as three sub-genera. It is interesting that no significance seems to be attached by Smith et al. to the fact that the nectar sugars in Astroloba are sucrose dominated whereas in Poellnitzia the nectar sucrose is less than 5%. These authors also find that… “The correlation of nectar in Astroloba and members of the H. subg. Robustipedunculares with the sucrose-rich Hexangulares type is surprising. Because of floral and other morphological features they expected that the sugar composition belonging to these two units would be nearer to the sucrose-low Aloe type nectar…”. This seems to be an odd statement when there is very little to support their expectation in the sense of the classification history of the groups and the statements concerning the two genera (Bayer, 1972).
The paper concerning a DNA study by Treutlein et al. (2003a) is also a very curious one and the result is summarized in this statement… “The current taxonomic system does not reflect the phylogenetic affinities and relationships among the succulent genera Aloe, Chortolirion, Gasteria, Haworthia and Poellnitzia.” That is quite obvious simply from the papers noted to this point and, had this been considered at the start of the study, surely this would have been the question to be addressed. The selection of material I take to be somewhat irresponsible given the past history of classification of the group and the literature. The inclusion of unknown hybrids such as H. X ryderiana, H. X kewensis, and H. X icosiphylla can tell us nothing. Likewise the failure to deposit vouchers in a recognised herbarium is problematic. Ignorance of the taxonomic position of H. geraldii is similarly curious and had the authors (and reviewers) known, as well they could have, that this taxon is simply a local variant in the H. retusa/turgida complex, and they would have been alerted to the anomalous and also startling result which finds them placing it in a different sub-genus. It can be noted that B.J.M. Zonneveld’s (geneticist, Leiden Univ. – pers.comm.) observations on total mass of nuclear DNA suggests that plants in cultivation in Europe are polyploids while field collected samples from South Africa fall in the normal diploid range for H. turgida or H. retusa as opposed to a higher value for species of the Hexangulares.
The authors repeat the mistake of stating Uitewaal’s contention that there were two main groups in Haworthia and mistakenly state…”two main units (Triangulares and Hexangulares), the former including the subgenus Haworthia and Subgenus Robustipedunculares.” Uitewaal divided the group Hexangulares into Gracilipedunculatae and Robustipedunculatae, not the group Triangulares. The way this has been repeated in the closing paragraph of Uitewaal’s paper has confounded Treutlein et al., who go on to say, “This division (into two groups) is strongly supported by…” their results. This is not true. Treutlein et al. have no representatives from the Robustipedunculares in their analysis apart from the DNA (cytoplasmic) in the hybrid XAstroworthia. It is thus not surprising that this “species” comes out in the “heterogenous group” that includes Aloe aristata, Gasteria, Poellnitzia, Astroloba and H. retusa (“geraldii”, with the position here of the latter extremely dubious if not totally erroneous). The suggestion that Haworthia could be split into two genera as a result of this study is misplaced because of the omission of members of the Robustipedunculares and ignorance of the actual situation reflected in the history of the groups where there are three groups that have to be considered.
Treutlein et al. are also the authors of another paper (2003b) and there three observations are made which I must contest.
1. …”An exception in the current classification was found with the sister species H. geraldiiand H. gracilis var. tenera: genetically they belong to group ll, whereas morphologically they show affinities to the subgenus Haworthia (represented by group 1).”
2. … “H. geraldii and H. gracilis var. tenera are sister species according to rbcL (fig.1A) and matK (fig.1B). Contrary to their previous morphological classification (Bayer, 1999), they are clearly grouped in Haworthia subgenus Hexangulares by both molecular markers.”
3. … ” the taxonomy of the genus Haworthia must be revisited. More species of both groups need to be examined to determine their phylogenetic relationships before taxonomic consequences should be drawn.
Where Treutlein et al., did not cite my Haworthia Revisited (Bayer, 1999) in their first paper, they do in this second one. It is difficult to know why, when they have not grasped that it is not a “morphological classification” as they claim, and thus could not have referred to it at all. My revision is probably unusual in that such pains are taken to explain a species concept and to state specifically that the geographical component is the over-riding consideration in the classification although it is also fortuitously a morphological one.
The question of the subgeneric relationships of H. geraldii and H.gracilis var. tenera is a fundamental issue. If either of these had been considered and reviewed as the taxa that the literature suggests (Bayer, 1974, Bayer 1999, Bayer, 2002a), it would have been quite evident that the conclusion Treutlein et al. reach with such facility is impossible. Certainly the taxon “geraldii” has been discussed many times. Further discussion gives credibility to the recognition of local variants as full species that is wholly misplaced. In the case of H. gracilis var. tenera, the writers can be excused for overlooking the discussion of this taxon in my book Haworthia Update (Bayer, 2002a) or in “Ecotypes in Haworthia” (Bayer, 2003b). In the latter paper I explain why the species “gracilis” and all its varieties are transferred to H. cooperi. This was effected in Haworthiad (Bayer, 2002b). It is obvious to any reader that the two taxa “tenera” and “gracilis” cannot be excised from the subgenus Haworthia by any stratagem at all, but this should have been apparent regardless of my two papers. That the term “sister species” (for ‘geraldii’ and ‘tenera’) is used, wholly disregards anything that I have written in respect of a “species” concept for Haworthia. No matter how the subgenus Haworthia is manipulated these two taxa are elements in quite different sub-domains of two species and in the subgenus Haworthia, not Hexangulares as Treutlein et al. show in their phylograms. It is unfortuante that researchers can be so unfamiliar with the classification of the elements they work with. Lee (2004) argues that such molecular diagnoses need to be based on an appropriate taxonomic framework in turn based on all appropriate biological information. Otherwise they are premature and likely to cause problems rather than solve them.
The observation that the classification of Haworthia should be revisited is quite unnecessary in the light of what I stated in my revision (1999). These authors have not understood what the basis of the revision was. It is based on a conceptualization of species as systems and recognition of three sub-genera. Thus where Treutlein et al. (2003b) state that…”The present study unequivocally shows that molecular evidence conflicts with the current treatment of Haworthia as a single genus” is therefore misleading. Similarly the statement that their data set supports a dichotomy of Haworthia is only true because of the omission of representatives of one of the sub-genera. Their failure to have familiarized themselves with the literature that they cite is evident.
One has to recognize that the classification in place is the product of a very long history and is reasonably sound and practical. There is also a very extensive herbarium record to support it. It is professional botanists that are not properly familiarizing themselves with the fields they engage. They are not asking logical questions about the relationships of the plants of interest, nor are they selecting authentic or representative samples. Their work in peer-reviewed journals does not pass scrutiny and one is left to consider that the selected peers are not competent to undertake the task expected of them in a proper review process. It should not be necessary for writers in the popular literature to be discredited by results and conclusions derived from frail, poorly conceived and badly planned research projects and publications at the higher academic and intellectual level. Neither should a functional classification system be shaken and threatened by weak research of this kind.
Fig. 1. JDV89.5 H. retusa fa ‘geraldii’ Riversdale.Fig. 2. MBB7236 H. turgida. NW Heidelberg.Fig. 3. Flowers of sg Haworthia.Fig. 4. Flowers of sg Hexangulares.Fig. 5. MBB6833 H. cooperi var tenera. Plutosvale.Fig. 6. MBB6892.1 H. cymbiformis var incurvula Plutosvale.
Acknowledgement I must thank the reviewers of this paper and the editor of this journal for instructive and necessary comment. Steven Hammer similarly provided incisive comment. Dr. Paul Forster has been particularly supportive in assisting me with significant literature as well as his views as a contemporary plant taxonomist and systematist. I must state that I am a parataxonomist (as opposed to a professional botanical systematist). The opinion that I should present a full phylogenetic analysis of my own, rather than direct my criticism of other efforts, should also take cognizance of the fact that all the evidence I can present is that within the extant genera and the three subgenera of Haworthia. It should also be stated that, the nature of variation may frequently defy any conventional Linnanean or Hennigian paradigms (Brummitt, 2002).
Literature Cited
Bayer, M.B. 1972. Re-instatement of the genera Astroloba and Poellnitzia (Liliaceae- Aloineae). Natl.Cact.Succ.J. 27:77-79.
Bayer, M.B. 2000. Observations – cytology as a character source in Gasteria. In Thoughts on Haworthia :96-99. Addendum 9. (ISBN No. 0-620-25323-1) Spiderwalk Services, Cape Town.
Bayer, M.B. 2002a. The case of Haworthia incurvula. Haworthia Update 1:25-33. Umdaus Press, Pretoria. (Revised from Aloe 36:34, 1969).
Bayer, M.B. 2002b. New names and combinations in Haworthia. Haworthiad 16:62.
Bayer, M.B. 2003a. Classification with purpose. In Alsterworthia International, special issue No.3.
Bayer, M.B. 2003b. Ecotypes in Haworthia. Aloe 40:10-15. [link]
Brummitt, R.K. 2002. How to chop up a tree. Taxon 51:31-41.
Manning, J.C. & G.F. Smith. 2000. The genus Poellnitzia included in Astroloba. Bothalia 30:53.
Reynolds, G.W. 1950. The Aloes of South Africa. Johannesburg.
Smith, G.F. & B.E. Van Wyk. 1991. Generic relationships in the Alooideae (Asphodelaceae). Taxon 40:557-581.
Smith, G.F., B.E. Van Wyk, M. Mossmer. & A.Viljoen. 1995. The taxonomy of Aloinella, Guillauminia and Lemeea (Aloaceae). Taxon 44:513-517.
Smith, G.F., B.E. Van Wyk, E.M.E. Steyn & I. Breuer. 2000. Infrageneric classification of Haworthia (Aloaceae): perspectives from nectar sugar analysis. Proceedings of XV1 AETFAT Congress, Belgium. Syst.Geogr.Pl. 71:391-397 (2001).
Treutlein, J., G.F. Smith, B.E. Van Wyk. & W. Wink. 2003a. Phylogenetic relationships in Asphodelaceae (Alooideae) inferred from chloroplast DNA sequences (rbcl, matK) and from genomic finger-printing (ISSR). Taxon 52:193-207.
Treutlein,J., G.F. Smith, B.E. Wyk. & W. Wink. 2003b. Evidence for the polyphyly of Haworthia (Asphodelaceae Subfamily Alooideae; Asparagales) Inferred from nucleotide sequences of rbcL, matK, ITS1 and genomic fingerprinting with ISSR-PCR. Plant. Biol. 5:513-521.
Uitewaal, A.J.A. 1947. A first attempt to subdivide the genus Haworthia, based on floral characters. Desert Plant Life 19:132-136.
Vosa, C. & A. Bennet. 1990. Chromosome studies in the Southern African Flora. Caryologia 43:235-247.
Let me not detract one iota from an exceptional and remarkable compilation of descriptions. There is no need for me to go through the book in detail because in respect of it being a compilation, it is outstanding.
What concerns me is what it portends and what it holds for the future of Haworthia. The book follows a Vol.1, which was reviewed in my book Thoughts on Haworthia (Spiderwalk, 1999). It is quite clear to me that Ingo Breuer is in a sense re-inventing the wheel. There is very little information in the book new to me and most of the work was available to Smith in 1947 and certainly to me in 1976. A prime problem I thus have with the work is that it is so firmly rooted in the past. It is thus a threat to the present and holds no promise for the future. The work does present problems, and portends disaster.
Some of the implicit interpretations in this work are possibly just as dubious as some of mine and often more so. I see little purpose in generating change for so small a result. I find the foreword by Prof. G.F. Smith disturbing. It demonstrates some of the malaise I describe in other chapters. Smith’s foreword ranks with a similar foreword to Breuer’s Vol. 1. by Prof. Ihlenfeldt which I commented on in Thoughts on Haworthia. I have said from extensive experience with many genera, that Haworthia does not offer any challenges which are not extant in other genera. It is my indictment of taxonomic botany that taxonomists are not better acquainted with natural diversity and the complex reticulate relationships of species.
To suggest that this book is going to help the reader in anyway towards understanding the actual species of Haworthia as Prof. Smith does, is unfounded. Classification is integral to language, organisation of thoughts and any communication. It is necessary for science and not necessarily science in itself as taxonomists seem to take science to be. To say that Breuer’s work comes as a refreshing change is to beg the question “From what?” In the context of the literature of Haworthia, the work by Breuer takes us back sixty years, and Smith is supporting this backward step. As I will point out, the work is fraught with potential disaster. To further say that this work allows the reader to make his or her own decisions on the application of “some of the controversial names” and further “assist in maintaining an acceptable level of stability” is just mis-statement and a poor assessment of the situation. I will set out to show why.
The purpose of a classification is for a single reasonably competent mind to review a group in such a way as to propose a system which unifies nomenclature and circumvents the need for individual judgements. If we each have our own interpretation of names, what purpose does classification then serve? It is like each of us having our own dictionary and own language. Unless authors driven by the communal need set out to respect nomenclatural stability, there will be no such thing. Names of plants are concerned with species. I have said in many places that if we use names, especially Latin binomials, we need to know what a species in fact is. This is something which not many taxonomists, nor aspirants to the task, seem to be aware of. Any botanist surely should know that the only real interpretation of an assemblage of plant specimens is to examine them in their actual field context. This compilation of descriptions by Breuer, however well intentioned, is self-evidently worthless in that respect and all the illustrations nearly so. This is particularly so in respect of the juxtaposed illustrations where Breuer has attempted interpretation.
The name “asperula” can be taken as an example. I discarded it for a very good reason. von Poellnitz used the name for at least four different elements and Scott for five. I am well aware that Hayashi and Esterhuizen (private communication) are, or were, of the opinion that it should be in the place of my concept of H. magnifica. What Breuer has done is to juxtapose J.R. Browns illustration and concept of what is almost certainly H. pygmaea as Aloe asperula with Salm-Dyck’s neotype and illustration. For him to have made this visual match is in my opinion a bad one even in the context of Haworthia. In my experience of Haworthia, I would say without question that Salm-Dyck’s icon is probably nearest to scabrid forms of H. retusa and, as Hayashi and Esterhuizen actually implied, specifically to a population from east of Riversdale.
Regarding H. emelyae, H. correcta and H. bayeri; it is not absolutely clear if Breuer has done a volte-face. He did intend correcta to supplant bayeri, and now it appears rather that picta may supplant emelyae. The origin of H. emelyae is quite clear from the exchange between Smith and Ferguson. I wrote this up in 1979 (part 4 of the Nat.Cact. Succ. Jl.). The plants were not in fact collected by Mrs. Ferguson, but by a Mrs. Le Roux, from near VanWyksdorp. The photos by Fourcade that Breuer uses to illustrate emelyae are quite misleading. One is H. reticulata and the other is H. turgida (Gamka River – and probably the lower Gamka which is the Gouritz River). Breuer has again demonstrated this curious inability, exhibited also by Col. Scott, to make visual comparisons. I liken it to the inability of some people to read maps. If one looks properly at the type illustrations of emelyae and correcta, one can conclude that they are more similar to each other, than are the Fourcade to emelyae. Breuer has simply assumed that Fourcade’s identifications were correct.
One of the reasons I regarded emelyae and bayeri as possibly the same species, was because I had two small collections of bayeri from De Rust and from south of Oudtshoorn. These were so scabrid that the difference between them and the Uniondale plants, was as great or greater than the difference between plants at VanWyksdorp and the predicted variation of emelyae between Rooiberg and Muiskraal (collections by Dekenah, and in recent years my own) and Springfontein (multifolia) and also its var. comptoniana from Georgida.
Breuer has a photograph by G.F. Wagner on page 570N, entitled “H. mirabilis sensu Bayer (mundula)”. I do not know how he arrives at this caption. It demonstrates a complete ignorance of all my writing and the problem I have stated of look-alikes, and demonstrates also the problem of identification which Breuer seems to assume does not exist. The illustration can superficially be taken to resemble mundula I will agree, but it is quite obvious that the plant cannot be and I certainly have never used the name “mundula” for this H. magnifica population at Barrydale. One needs to look at the illustration of mirabilis in Curtis’ Bot Mag. – the neotype – and see how well this compares with the Barrydale illustration in Breuer’s book. Then one must look under at H. emelyae var. beukmannii and see the various illustrations there from which Breuer also draws his epitype for mirabilis. Unfortunately Smith’s published illustration of his mundula is a very poor rendering of the element as it occurs in nature. Additionally there is a poor comparison of this with the real mundula photographed by Esterhuizen – of one of the less-lined yellowish clones. It can be noted that as a general rule, mirabilis tends to colour red to yellow under stress whereas maraisii (and magnifica) become darker and duskier.
There is actually a remarkable story of change attached to mundula which I should relate in another place. The plants are not now as they were in 1969 when I first saw them, and of course they might not be anything like what Otzen might have seen. It does raise an interesting point about the taxonomic process – is it designed to produce a classification with which we deal with the living phenomena we see, or do we really need a system which allows us to organise a very limited sample of dead and dried specimens with precision?
I class these Barrydale plants as either magnifica or maraisii although in my work it is clear that I have problems with separating maraisii from magnifica. Thus originally I specifically refer to these Barrydale plants as magnifica (see Haworthia Handbook, 1976 p100, under asperula). This is one of the problems I have in trying to discuss Haworthia with anyone. There is this reticulate relationship and one can find odd plants in populations which more closely resemble other ‘species’ than that to which they belong.
The problem with mirabilis as opposed to mundula is one which I refer to as a weakness in nomenclature. Breuer has thus designated an epitype for H. mirabilis from Skuitsberg whereas I select the Curtis Ker-Gawler illustration and state the similarity to Smith’s mundula. Therefore a local variant becomes H. mirabilis var. mirabilis, and the whole general body of mirabilis has to be treated as the var. triebneriana. In private correspondence Breuer makes it obvious that this aspect of nomenclature has eluded him, by asking me what can have happened to H. mirabilis if I now typify it with a specimen named mundula! An unpublished manuscript of Breuer’s in my possession suggests that he wanted to uphold the var. beukmannii of mirabilis also from Skuitsberg. This is done in his book. It does not make sense and any change at species level creates a host of complications at subspecies or varietal level (apart from the added complication of rank priority). Also concerning my species mirabilis in a similar way are the two illustrations used by Breuer to depict the var. rubrodentata. This particular variety is actually represented by a few specific slender leaved clones on a particular steep north facing clay slope indicated to me by the original collector G.J. Payne in 1970. There are at least four other localities west of Greyton where mirabilis occurs and Breuer’s IB5129, coming from any of these, is not comparable with the itemised Triebner 1143.
Three last examples of this incapacity to apply visual imagery with any credibility are:-
1. In the case of H. rossouwii where Brown’s 1953 illustration is obviously not comparable with the von Poellnitz lectotype. This particular picture is virtually identical with Breuer’s own illustration of H. emelyae var. major IB5539.
2. The illustration of H. stiemiei can be shown to be ambiguous and controversial and this will emerge from a manuscript of mine dealing with four intergrading elements from the Zuurberg north of Kirkwood. Breuer’s own depiction of this species could be fruitfully compared with elements as far afield as Wolfkloof, Robertson; and many places in the Baviaanskloof, as well as the Elandsriver Valley. While it no doubt could be that Breuer’s illustration may represent the same element that the type of stiemiei does, the actual resemblance of the two, in the sense of the book, is minimal.
3. What is disturbing is the fact that Breuer wants to apply the name denticulata (Bavaainskloof) here, and the visually and chronologically comparable aristata to Barrydale (my mucronata). I am comfortable with the former because I consider this is where aristata and denticulata BOTH COULD actually belong. In fact I am not comfortable at all with the imagery with which Breuer relates his aristata to the lectotype or to Triebner’s plant. I agree there is reasonable merit, but only in the absence of any prior attempt to better interpret the element. Scott’s views and imagery are/were never any better than Breuer’s. Breuer is remiss for having learnt nothing from history.
It occurs to me to now relate an example of the potential confusion that exists in Haworthia by explaining an example of what Breuer might well do. One can argue on the basis of both type illustration and description that bijliana of v. Poelln is actually either Rooivlei (Bredasdorp) H. heidelbergensis, OR H. serrata from northwest Bredasdorp. In terms of Haworthia these would both be valid arguments and I think this should be pre‑empted. In an unpublished manuscript I uphold H. rossouwii (which is a much later name than H. bijliana), over serrata. But it could be argued that bijliana also is this element.
In Feddes Repert. (1936) v. Poelln. cites the species bijliana from Klawer and Eenriet, and in 1937 cites it again as from Springbok. In this last publication he cites a specimen also from Springbok, another from Bonnievale, another from Ezeljacht Oudtshoorn, and also sinks H. fergusoniae here which in fact was supposed to have come from ‘near’ Grahamstown. Thus Breuer has:
H. bijliana v. Poelln. in Feddes Repert. 27:134 (1929); Bredasdorp, Mrs. van der Bijl. Type van der Bijl s.n. in (B). Not preserved. Lectotype designated Breuer 1999, unpublished photo. icon. (B) (my copy of the description does not have the word “doubtful” after Bredasdorp as Breuer adds in his book).
Funnily the original illustration of bijliana is in my opinion very like rossouwii, and fergusoniae like that of tenera ‑ whereas later treatment of the names puts them as arachnoidea. Breuer’s treatment thus in many instances demonstrates the inherent self-evident fact that types in Haworthia are, and always will be ambiguous. Types cannot be used to confidently identify and name plants outside of the geographic origins of those types. The point Breuer makes about ambiguity on XIII is actually just some of the juristic jargon which is supposes to lend credibility to nomenclature. It is a fact of life that a type in Haworthia is ambiguous unless it is very clearly illustrated and the locality properly stated. Thus in the case of emelyae it is self-evident from Breuer’s own use of the illustrations, that the lectotype was highly ambiguous.
Without making a point of finding any trivial errors:
On page 784 Breuer has Olifantsdorn for Olifantsdoorn, page 784 Breuer has Conwitz i.s.o. Gouritz and on page 786 Stromsvley for Stormsvlei (this is an interesting error because v.Poellnitz cites a Strydomsvlei when it may have been Stormsvlei (as it happens this is also in connection with H. triebneriana, and potentially damaging because Stormsvlei is the origin of the vars. depauperata and pulchra. All these fall within the broader context of mirabilis and mundula of either Bayer or Breuer).
There is merit in trying to arrive at another interpretation of H. helmiae and I think Breuer in this one case is possibly right. For some reason Scott was adamant that Mrs. Helm had told him she had collected this species at Schoemanspoort, which would have made my interpretation reasonable, but Scott often bent the truth to meet his needs If one looks in any way at the four illustrations that Breuer present for helmiae, one must be struck again at the incongruence of the match. Breuer’s lectotype could hardly be more ambiguous when held against the Fourcade photograph. The point should be made that Breuer seems to attach an omniscience to the Brown and Fourcade illustrations which is wholly at variance with the facts – as noted by myself in 1976.
The re-introduction of the name integra for my rycroftiana is probably acceptable on the basis of the new evidence (if true). I regarded ‘integra‘ as an unknown in 1982 with inadequate evidence to support it. Scott salvaged the name for what in my opinion is arachnoidea var. nigricans. Almost any specimen of any of the similar ‘soft’ Haworthia species from diverse populations, preserved in alcohol, could be taken to be a mucronata variant if so desired or required. This particular one with the dark tips evident to the leaves could in my opinion be from outside the Gamka Valley from a place like Ockertskraal or Muurkees. The species mucronata is any event so variable that the suggestion that integra be upheld as a discrete species from mucronata, is of very doubtful value. As it is I have enough evidence to show that arachnoidea and mucronata can often not be separated.
I think it would be useful to know if Breuer’s IB numbers refer to plants which he had any real familiarity with in terms of their field context. I am aware that Breuer is first in the queue for plants collected and propagated by myself which come to be distributed in the trade. This creates a problem because propagation has always been largely by vegetative means – and mistakes occur in transfer. Thus the trade, and collections, tend to be dominated by proliferous clones and they may be misnamed and wrongly accessioned as to origin. The variability of the population is wholly obscured. Evidence of Breuer’s capacity to match visual images, suggests to me that he would not be able to find any pattern unaided by modern interpretation. Hence the reference to re-invention of a wheel.
There is a problem with a H. gigas – the plants from Amalienstein are familiar to me and they are not quite the same rigid plants with brown spination that is so characteristic of the Laingsburg plants. Treating them as synonymous with my H. arachnoidea var. scabrispina removes the geographic significance of the taxon. It is not in the least significant that they look-alike. Attaching nomenclatural significance to this similarity is wholly unnecessary and misleading.
There will be a problem with aristata as Breuer and I interpret them so differently. There is in fact a problem with gracilis as well which Breuer may not be aware of. However, the dickering over nomenclature here is not going to resolve the problem of what anyone does to understand the relationship between what I have called gracilis, cooperi, blackbeardiana and cymbiformis, and which extends to pringlei, decipiens, xiphiophylla and even mucronata. Using names in any other way is not going to solve any problem and will simply add to the confusion when the problem has to be faced. There is no way that one can actually separate Breuer’s aristata (my mucronata var. mucronata) and his mucronata (my mucronata var. inconfluens) as species. This ONE element is confounded with H. arachnoidea in many places and how anyone is going to find names to accommodate the associated variation in any new arrangement will be fascinating to see.
I have written several manuscripts which deal with specific issues in this context and these are included in this volume of essays. These will show the difficulties which Breuer’s work presents, and will show that Prof. Smith’s use in the foreword of the term “golden age” is for a work which actually helps to usher in a blackout. I agree wholeheartedly with Prof. Smith that Breuer’s work as a librarian is beyond reproach, but for helping to reorganise Haworthia it has very doubtful value. ♦
At last a reasonably rational review which is written with some insight and understanding. It makes points which lead to communication and hence discussion. The then Editor of Haworthiad in relaying his feelings and that of his readers has criticised me many times for what is said to be my intolerance of the views of others. I want to set this record straight. I am intolerant of anything which stultifies and paralyses the understanding of Haworthia and distorts communication about it. I had written to Paul Forster before I heard he was preparing this review, and then I was thrown into a bit of a quandary. I do have enormous respect for people and it is not my wish to hurt anyone’s feelings. Paul is no doubt at all an excellent botanist and a human being of a high order. Unfortunately, I do not think his review is particularly good as it is couched in the paradigm of the orthodox and the conventional. I hoped for more, as I sense the complacence and almost smug security of the professional herbarium botanist who’s highly descriptive revisions will be held in awe by, and never ever tested in the crucible of, popular interest. There are many very positive things in the review, and in concentrating on the negative I really feel that progress is possible.
Firstly, I stated that herbarium space was a problem. It IS a problem. Whether Paul thought there was space or not is irrelevant. For many years random additions to the herbaria have been frowned on. Also for many years there was resistance to discarding the meaningless collections that occupied so much space in the cupboards. Paul should know how a herbarium is arranged, and that an addition to the full cupboard of a fern genus has a ripple affect all the way to the end of the collection. Space in the fern cupboard does not mean space in the Haworthia cupboard. The Bolus herbarium will only accept material regarded as necessary for bona fide research and the same is true for Compton Herbarium (NBG), where researchers are genuinely concerned about ‘their’ own space. Three of the only collectors I know (and who were persuaded and encouraged by me) to deposit specimens at all are Howard Gie, Emil Heunis and J.D. Venter. They were discouraged from doing so because of the real pressure for space that existed at the Compton and Bolus Herbaria – and still does. Gone are the days when collectors were welcomed with open arms and seen as a source of valuable new material. It is only in 1996, that space for Haworthia was created in the new herbarium building at Kirstenbosch. This is not because of a spacious new building which squeezed Haworthia into possibly less space than it had before, but because the curators allowed the transfer of about four boxes of G.G. Smith duplicates to Pretoria. I hate to think what problems these have created for Pretoria when I reflect on the lack of space I saw there in 1996, and available in terms of the total arrangement of the herbarium. I contributed many specimens to NBG and I share their concern that it is just not possible to provide the space necessary to reflect the diversity of the flora of the Cape. I must have driven them to distraction with my collections of Oxalis and Asparagus, which I think are packed in boxes in some remote and isolated room somewhere in the user unfriendly building. It is also not just a question of space. The management of new material from the collector is a mammoth task. Specimens must be accessioned, treated, mounted and labelled. Those I submitted more than two years ago, are still not available for me to see again!
Why does Paul cite DMC3625 Kubusie Drift, near Kingwilliamstown if there is no specimen? Because it is in fact meaningless and he intends this as an abject lesson to collectors who want a classification, but not the trouble required to support one. He later states that if there are no specimens (of intergrades) then they do not exist from a scientific data point of view. I would say, please leave out the red‑herring word ‘scientific’. It is beginning to convey to me the ultimate in insensibility. There are these intergrades and I really puzzled about how to cite them. I even described taxa to accommodate them e.g. H. decipiens var. minor, and H. gracilis var. viridis. Paul also writes of intergrades which he knows exists because he has seen them – but he also has made no specimens? His casual statement is thus good enough while mine is not? He does have the grace to say that new specimens have further blurred the distinctions between previously recognised taxa but he does not say which ones. Did he make specimens to support this statement? Paul does miss an important criticism ‑ the incorrect citation of specimens e.g. under H. cooperi var. pilifera, where Brigadoon is cited as Grahamstown and also King Williams Town; Peninsula is cited as Grahamstown, when it should have been King Williams Town. These are important errors because knowing this geography would perhaps have allowed one to connect the Kubusie Drift collection with other collections from nearby sites. Thus DMC3625 (if it existed) could in terms of my revision be only H. cooperi var. pilifera or H. cymbiformis var. setulifera. Also I find that I have several times erroneously cited the same specimens under different names. This arises from duplicates seen at different times and place, and ones which confused me often because of their intermediacy. However, this is what I hope the next revision will sort out ‑ thorough field work and assignment of each collection, in a more considered way, to each taxon. Part of my own frustration is that persons are fiddling about with nomenclature and guesswork when it is things like this that need attention and questioning. A herbarium specimen does not give the required information and ‘feel’ and sometimes a single dried specimen, and my memory, is all I had. Let me also say in mitigation – I have never been employed to do work on Haworthia and have never, in employ, been encouraged to do so. I do not have full-time access to the herbarium, and have never had. This is a serious problem when the herbarium record is the core of revisionary work and I might have made fewer mistakes had it been easier for me. I would also make fewer mistakes if half my mind was not held back so by the negative history of the literature on Haworthia.
There is also a serious and common problem with DMC numbers. This particular collector is well known to me and we have exchanged views on collection done without a permit, in numbers which no permit would concede, in a way which has no scientific principles at its base, and which does not ensure a herbarium record. 3600 meaningless numbers! It just is a fact of life that collectors do not make specimens and it plagues me more than it does him. In the normal course one should not expect them to do so, but for one he gets out into the field in the way that DMC does, it is another matter.
A bigger problem with herbarium material is that any haworthiophile values the plants he/she gets. To reduce them to squashed dried specimens is painful in the extreme. Apart from the pain of separation, it requires really tedious effort, more to make the specimens recognisable, and then the equipment and space to do so. Permit requirements, and a conservation ethic, severely limit what one can collect (my permit allows three offsets, and my ethic none!). As the plants require some observation, these are generally taken into cultivation with attempts to propagate and distribute material as well as produce a specimen from the product. The dried specimens seldom display the variation of the parent population and one usually finds a vegetatively propagated clone on the sheet. Miss Winsome Barker (past Curator of the Compton Herbarium) encouraged the use of colour photography to add to the usefulness of herbarium sheets. If there is any strength in my classification at all, it is because of what is in the herbarium like this. It disturbs me that so much of what I have seen, is not adequately documented in this way. Most of the herbarium records are of single specimens. While my permit allows three, I generally like to see at least six grown in cultivation to get an idea of what any population may be like. I have grown plants from field collected seed, and to record some of this variability in terms of essential herbarium record is a monumental job – an impossibility. But I have tried to do it and am still trying. I cannot add an apology for the fact that I like the plants living rather than dead.
Paul says I have all this ‘information’ about vegetation and habitat ‑ indeed I do. So much so, that I cannot fool myself into thinking that one can make specific statements about habitat preferences. His comment makes me think back to the comment of Gordon Rowley’s about the soil pH data given in Col. Scott’s book. Was this pH data really any good at all, and if it was, did it have any practical value whatsoever? (The story of the mima-like mounds of the winter rainfall Cape would be a useful reference in this respect.) It should be clear to the expert who knows something of the South African flora and geography that no statement he can make will be clear to the novice, and that quite probably it will not be clear to any other expert either. I am still wrestling with this problem fantasy that vegetation can be classified with the same facility that discrete species can. I believe this is one of the great misconceptions and myths of botany, which like so many things in science, is never as simple as it seems. Paul is said to be a cycad researcher. I doubt if he could explain the distribution and habitats of the E. Cape cycads in terms of the recorded and described vegetation of the area, or in terms of their classification. This is a considerably easier task than it will be for Haworthia. Paul should know that Haworthia is just one of many genera which are virtually unresolvable in terms of classic taxonomy, or describable by vegetation and habitat. I have named some before and I can add to the list with Hermannia, Rhus, Tetragonia, Lycium, Pteronia, Zygophyllum as examples I am familiar with. Vegetation study will be more meaningful when we have good classifications for these genera. I have recently seen a Haworthia change facies over a distance of 50 meters, because of local habitat differences. The only differences I can record is the fact that one plant form was on exposed vertical slabs of rock, and the other in sloping terrain with the same rock substrate, but with more soil and grass. One would need to be a geologist, geomorphologist, and soil scientist, in addition to be able to identify and quantify the associated plant species to do what Paul idealises. Perhaps this is the prerogative of a reviewer. Nothing is impossible and it should have been done.
Paul makes the excellent point that the only person who can say if this new classification is any good is I. So I say “Thank you, I actually think, and know, that it is very good indeed”! This should be true for the author of any revision. I am sorry to say that in my experience, many revisions which have been accepted and published by “scientific” journals (or as academic theses) have proved to be pretty useless. I should add that this is not only for new material not seen by the authors, and despite meeting all the requirements Paul has for my ‘synopsis’. He goes on to say that “While it may be easy (probably not) for Bruce to attempt to fit new and morphologically distinctive collections into his system, it is well nigh impossible for anyone else for the lack of data presented”. The unnecessary words ‘probably not’ points to the wrong place. The subgenera in Haworthia are morphologically distinctive and that is practically where it stops. My complaints have been directed at the fact that persons unlimited are creating new names and fiddling with old ones because they are so unaware of this problem. My ‘data’ includes geographic information and no one should even attempt to fit a grossly ‘new and morphologically distinctive’ collection into my system without familiarity with that system. Nobody without knowledge of the herbarium collections can know what is new and distinctive. If they do not know this they should not be fiddling with Haworthia. The problem is not caused by my presentation and its shortcomings – it IS a fact of life for many more genera than just Haworthia. Writers are playing with MY classification, that have not seen the material in the herbarium and neither do they have the knowledge of those specimens in the field. They may have seen material that I have not seen, but still the responsibility is on them to first know what forms the body of an existing classification and its predictive ness, before embarking on a new mission to repeat the exercise.
Paul skates about a bit with the questions of descriptions and measurements. He complains that dimensions (‘size dimensions’!) are not provided. In an age when people do not read, and look at pictures instead, I decided that this kind of data could as well be deduced – at the level that it is practical and needed – from the many illustrations. Of course a lot of importance is attached to these details Paul calls for, because they give the impression of quantification and hence ‘materialistic ‘science’. Intellectuals have to have something to keep them busy or create the pretensions of business. It would have made the book a lot thicker (and a lot more expensive). I also know very well that with all that detail my credibility in the academic environment would be assured. Of course living organisms are measurable and there is also a statistical science (biometry) that goes with it. This science depends on randomness and replication, deals in mostly normal data and probabilities. It is my knowledge of this science which supports my understanding of Haworthia, and part of the reason why my work is sound. Nobody is going to get anywhere in Haworthia neither with three specimens, nor very far with the specific mensuration that Paul calls for. I do know the group. Contrary to Paul’s statement (sorry #, Paul), it is because I know the group (and many others) that I also know that this variability cannot be easily rationalised and summarised for meaningful usage. Haworthia is not like, say, Hoya where a new species (see Marsdenia as a new genus) can be recognised on the basis of a single flowering specimen. In a collection, often the decision on whether one or two clones are involved is to look and see if their flowers are different! The variability of the plants is what it is, and I do not understand why readers seem to think that there must be some character state lurking somewhere that is going to provide an ultimate solution. It is all well and good saying, as Paul does, that every time he plants seed he is struck by the differences. I challenge him to document this frivolous observation and to explain to me what exactly has struck him and what it’s more general value is. This approach of his is the same as the weaknesses he rightly perceives in my revision. How do we get out of the condition? In biometry (the statistics of biological systems?) we were taught that you do not attempt to seek significance when it is obvious what the variability in the sample tells your common sense. It is a laughing issue in agricultural science that researchers seek significance in statistical test after statistical test until, on the basis of the very probability they are testing, they find the significance they seek. Botany has apparently not reached this stage yet.
What I have said about voluminous and detailed descriptions is that they are in practise almost useless. I can say confidently that Paul’s comments about the woeful and incomparable descriptions (I think they are masterful in their brevity, contrary to Paul’s use of the word!) is a true statement about Haworthia. It cannot be wished away or blamed on me. I would really like to give him just the currently deposited collections of what appear to be two species viz. H. cymbiformis and H. cooperi. He must then write a description (aided by as much living material as he wants) which allows the reader to identify them with any degree of certainty at all, and in so doing also exclude elements such as H. gracilis, H. bolusii and H. aristata. If his then comprehensive description adds anything to the synopses of my revision, I will indeed be surprised. I know what goes on in taxonomic accounts which Paul so reverently cites, and I am not impressed by the vast amount of detail ‑ look how it has been used in cladistic studies in the Aloeaceae at genus level! If Paul knew Haworthia well enough to write this review, he would have also been able to say that Esterhuizen and Battista’s taxon (H. mucronata var. rooibergensis) bore no relation to reality at all. Of what value to Paul (and Col. Scott’s book would have been adequate for this particular exercise) was the detail these authors supplied? How could he possibly have used it if I had supplied the statistically true comparable values for just H. mucronata alone? I had already written a response to Esterhuizen and Battista which will be published elsewhere, and this response does deal again with this issue of detail. I must say that I am familiar enough with Halda’s work to suggest a blacklisting by the International Association of Plant Taxonomists. We could both be there together!
Yes, I am aware that I have not given, say, the difference in flowering time between bayeri and emelyae. It is only about six weeks and I think the actual flowering dates vary from east to west. However, it is a lot more complicated than that and I do not know what I can truthfully say. G.G. Smith recorded flowering times of many collections for several years and these varied by as much as 5 months. In H. magnifica var. splendens, the west of Albertinia populations flower in November, those east, in September and perhaps again in November. In the field I have observed a population of H. cooperi var. gordoniana (which has affinities with cymbiformis var. transiens! – Jeffries Bay MBB6792) which in one year flowered six weeks later than in the preceding year. Flowering time is as erratic and variable as the populations are. A good record should also be bounded by statistical limits. I separated maraisii and mirabilis on flowering time (Apr./May vs Feb./Mar), but it does not always work. The McGregor complex may flower from Nov. to May. In cultivation there are nearly always odd plants flowering out of synchrony and my field work has not been that of the professional who can set goals to observe phenological events. Yes, I did indeed do a lot of work on the flowers, and I cannot use it because I could find so little pattern. Like in the asclepiads, differences in the flowers within populations often exceeded the differences between populations. Again, underlying Paul’s complaint is the fact that there is no Holy Grail that is going to unlock the conundrum of Haworthia.
No, the book without pictures is not user friendly. It was not expected to be and I said as much. If the publisher could have accommodated another 450 (is it 450 or more?) photographs and the book remain affordable, I am sure they would have done so. The question of intergrades is also thin‑ice stuff. Does the nomenclatural system allow these things? In my collection book I quite happily use the chemistry equilibrium equation to indicate how I would classify a collection and many of the herbarium collections are so indicated. The object of my classification was to try and identify divisions rather than obscure them, which is what most classifications do. My next exercise (which I wish was a competent somebody else’s) is to show that intergrades are not just two-way, but perhaps three- or even four-way.
As to H. pumila, many (may I repeat, many) professional plant taxonomists have entered the debate and indeed there is no vacuum here. It is a space filled with vacillation and doubt, conflicting opinion and retroactive wisdom. Nomenclature is not my strong point, and perhaps for the very reason that any good practicable workable solution can be contested and upturned (and will be so contested and upturned) for no other reason that there is a juristic element which caters for it. That the International Association of Plant Taxonomists has not resolved the particular issue of H. pumila is their problem. I find it extraordinary that Paul does not see the irony of the situation. IF I have made a mistake, the following is what I will concede – the author citation of H. pumila should read (L.)Scott, and not (Duv.)Scott or (L.)Bayer. I deliberately took the onus on myself for any flack that my continued use of the name would draw, but I am very happy to give the credit back to a gentleman and a friend. This name pumila is the first epithet for Haworthia in the Linnaean system and if the ICBN does not allow its use I will happily continue to condemn that august body, all its pettifogging recommendations, and the Committee that empowers it or is empowered by it. Paul is just another of a very long list of professional taxonomists who can blithely say “I am not sure what the outcome should be at this point in time”. When will this momentous decision be taken and by whom? He cannot thus be any more comfortable with nomenclature than I am, given that he is a professional. Paul is only going to be sure what the correct solution is when he is so informed by the high priests of nomenclature. So much for science. The fact that a committee has to resolve these things is, in my opinion, an indictment of the entire process. It is not just any committee of qualified taxonomists he can do this wondrous work as I painfully learned from a group discussion with four Doctored intellectuals. As for the “serious” oversight concerning the rule (Prof. Cronquist asked if nomenclaturists knew what they were doing, and I think not) of rank priority, forget it. I do not like the rule (or much else about the ICBN) and I do not see the necessity for its inflexibility. There seem to be more instability nurtured by the code than precluded by it. I prefer the names I have used and I also think they are more meaningful. Let the bandits do what they want to. It is perfectly clear what my names were derived from and what they mean. I think carping about the “rules” is just a nit-picking exercise derived from a failed historical attempt to rigidly control nomenclature. It does, and will continue to cause, more problems than it solves. As for “bandits”? Who stole the familiar name Astroloba muricata away from Groen and Roberts-Reinecke and substituted a new name corrugata under the guise of the strongest words associated with the Code i.e. to avoid confusion? This is what I refer to as trivial botany and I hope the IAPT will mint a medallion for this kind of thing.
A fact is that there is not one of my names (or Scott’s) for which there is any doubt as to origin, usage, or typification – except that it was not done in the style and letter-correct formality of the ICBN. The fact that Breuer and Metzing could do what they did (and add to this, all the words that the former author generated for the non-existent enigma of H. arachnoidea) is further adverse commentary on the ICBN.
Paul does dangle the predictable carrot…”It can be expected that with further exploration…new data… may radically change classification”. What does he think happened between 1982 and 1996 when the book was drafted? What is more important is what has happened since 1996? I have been almost horrified by the threatened return to the taxonomy of the early 1940’s, and in this respect Paul was kind enough to mention “the changing of a few names” by Breuer. The dearth of any signs of intelligent life in new ventures into Haworthia compelled me back into the arena by default. I have done some very thorough recent exploration and can say that my classification is even better than I had hoped it would be. Some very interesting new collections (and to date there are well over 400 new numbers in my accession book) can all be explained in terms of my classification if they can be explained at all. This is why I know my classification is so good. I hope to present the information somehow, in someway, as supplementary publications and the first has just appeared in Aloe 36:34 (1999).
There is a reference to refereeing of the book prior to publication. This is not fair to Peter Bruyns and I unfortunately cannot give a full explanation why I think this is so. We have worked together for more than 25 years. While his contribution was useful, this usefulness was not very forthcoming and it is not a door I would beg at. His big contribution to the debate about Haworthia is his forthright comment “Lump them!” It may well be the sensible thing to do and I have many times considered it seriously. Regrettably the opinion is based on an extraordinary ignorance of Haworthia, despite our long association.
Finally let me say this. For the full eighteen years that I was Head of the Karoo Botanic Garden (before, and even after) I sought the taxonomist Messiah that would lead us out of the unhappiness or confusion of Haworthia. Nobody has ever even nibbled at the bait. Botanists just do not want to mess with “difficult” groups, and I have seen them really mess with simpler problems. See what Prof. E.A.C.L.E. Schelpe said and did about Gasteria, or about publishing a really marvellous work on Astroloba by Pandora Roberts. If I have presented a revision of Haworthia, it is simply because there has not been anybody more competent, and this may be an unpleasant truth. I have never wished to say “Trust me, I am the expert”. I consider it most unfortunate that I am, and I would rather be doing something else. After reading Paul’s review I can confidently say that my classification is a lot better than Paul is guessing. As I digest and work over these latest collections of mine my confidence grows. If good sense prevails there will be very little in the way of name change or circumscription and we will all be able to enjoy the plants a lot more. If somebody competent does appear who wishes to build on what I have done and take over where I want to leave off, they will have my unstinted blessing, assistance and gratitude. ♦
I read a review of the book Rock of Ages authored by Stephen Jay Gould, by D. Bristow-Bovey (Sunday Independent, 27th May, 2001). Because the review left me feeling paltry and empty as a scientist, and defrauded, ignored or forgotten as a seeker, I acquired the book to find out what Gould actually had to say. Gould is a seminal figure in the popular literature of biology where I browse, and I look to him to interpret science and its progress. But this book with a subtitle “Science and religion in the fullness of life” seems to fail the subject, and me, completely.
I would have thought that the appearance of books like Aldous Huxley’s The Doors of Perception (describing the effects of drugs on awareness), Fritjof Capra’s The Turning Point, Bentov’s Stalking the Wild Pendulum, Gary Zukav’s The Seat of the Soul, Shirley McClean’s Dancing in the Light, Lyall Watsons books, as well as so many, many others; would have at the least indicated that mankind was on the threshold of a new age. There is a vast literature on the subject of new age philosophy and predictions of change. Where can science be in all this?
Gould’s book is a rude awakening to the facts of our collective spiritual dormancy. Far from providing any comfort, it coolly suggests that this intellectual and intelligent business of science has work to do, and people with spiritual inclinations can go and play somewhere else: i.e. religion is for believers, and science is for thinkers. Gould, in my opinion, has written in the intellectual climate of the age of Darwin as he to some degree honestly admits. In that age intellectuals were awakening to the harsh reality that the religious beliefs of the day, did not tally with their observations of the world. Gould holds that this problem can be resolved simply by recognising that there are two sides to the story. Two magisteria, the one being religion, and the other science. One being that of religion, where imagination can run riot without the strictures of hard evidence. The other, the dispassionate, disciplined, considered, calculated and wondrous field of science, where he himself is a great scientist and writer. He does not say what other magisteria there are in which one would then have to accommodate human aspirations or in which one could seek human happiness.
Gould seeks to exonerate himself from predicted criticism. With a respect to him at least equal to that he accords religion, I think his argument can be shown to lead nowhere.
In his Preamble, Gould states what science is, viz… “what is the universe made of and why does it work this way”. He is immediately fuzzy with the same fuzziness which he deplores when he later says.. “I have difficulty keeping a straight face and a peaceful pen”. He assigns to religion the task only of “ultimate meaning”, which is surely no different than the “why” of science. The difference is just one of belief.
He suggests that science deals with measurable things and that religion does not. The truth is that science just does not have a measure of the elements of the human mind such as pleasure, pain, hate, love, greed, contentment, humour, charity, happiness, wonder, truth and humility. For that reason then, all the things which drive emotions and feelings do not exist for the limited scientist. He perhaps overlooks the fact that in the broader sense, science actually has no absolute measure of space or time either. Creation is nothing but phenomena distributed in space and changing with time, and religion is there to fulfill a void that Gould’s version of science is too limited to deal with. As a scientist and intellectual, Gould opts out of his responsibility and leaves us all to the mercies of laymen. What has happened, and this is very well stated by Schumacher (in his book “A guide for the Perplexed”), is that science has simply departed from its original root and foundation. Where it was given birth to free mankind from the religious dogma, pedantry, ritual, ceremony and blind belief of organised religion, it has actually become another religion which simply denies the very things to which religion owes its origins. It has its own pedantic and blind priesthood, it’s same speculation and falsehood. It is just as accountable for the misery pain and suffering in this creation as anything else. Science has set us free from nothing. Even its claims of technological advance in so many fields have done nothing more for the human spirit than religion.
Freemasonry probably can be found at the roots of intellectual awakening, and I understand that the fathers of science like Bacon and Newton were freemasons. Was it not in their creed to study natural phenomena and extract meaning and purpose from such study? Was it not the beginning of an attempt to escape from the restriction and persecution of religious doctrine, creed and dogma of the day? The intrinsic binding injunction or requirement of the mason was the practise of truth and justice and a belief in God.
Schumacher placed great emphasis on ancient wisdom – but what is ancient wisdom? Is it all that old traditional pagan, heathen myth, legend and belief which underlay the evolution of the human spirit? Was it just the glue of a backward society without the wonder of science and intellectual achievement? Or was it the true, real product of an innate driving force which compels man to seek meaning and purpose? Thus we may have a scientist whose profession it is to develop technology, or study the biology of Seekatruthicus religiosus (this is a fictitious name for a non-existent mite). This same scientist may have an unquestioned fixation and unsubstantiated belief in life after death, promised by virtue of birth in a family which has organised his circumcision, christening, and later confirmation. Such a scientist may or may not consider and face the reality of death and what it will hold for him. Almost certainly he will not even think about, let alone follow the promises and practises of the religion he professes to follow by either conscious or unconscious choice. Another scientist may follow the same route and while examining the mites under his microscope, ask “What is the meaning and purpose of what I see?” “Does it tally with what the circumstance and experience of my life reveal to me?” Why Gould should suggest that such questions are invalid in the domain of science, may just be because of his personal belief that science should not address such questions. This is a limitation which Gould may place on science as his personal prerogative. Whatever other people also do, it is not a limitation of science or of scientific method. It is a personal limitation which the individual is free to examine and then retain or discard. We should all go through this process.
The problem that arose from Darwin was that man began to believe that religion too was a simple product of evolution, and that spirituality as descent of man from angel was unthinkable.
Gould places a restrictive meaning on the word “science” and maintains the restricted meaning of the word “religion”. Science, from the latin “scíre” meaning to know, and with a dictionary meaning (among others) of…”a body of organised knowledge”. Religion from “re” return and “ligare” to bind, meaning to bind back. This is no different from the meaning of the word “yoga” from the Sanskrit meaning to yoke (back to God). If a scientist is serious about what he thinks, he sets out to obtain the data, the knowledge and the experience which can help him answer his questions. Schumacher would simply say that it is a question of where and how he seeks happiness. Science and religion do not stand opposed as Gould suggests. Anyone can follow scientific method to understand how to bind back to God – however improbable that may sound or difficult it may prove to be. Enchantment with the outward physical world is no excuse for turning away from this human imperative. Religion is by definition “belief”, but that does not necessitate ignorance, nor invalidate any intellectual attempt to convert that belief to knowledge.
If Gould would read and study the literature (and the Bible is one of the lesser works in all that is actually available), and if he would penetrate the wisdom of the Rubaiyat which he freely quotes, he would see that these writers (and there are so many of them) are describing their travails, their hopes, their despair, their triumphs, their experiences, their methods. But by reading these works one does not acquire that experience or that knowledge. There are methods by which one obtains spiritual knowledge – as opposed to the outward practises of ceremony and ritual which accompany religion. There are actually many methods as any aspirant seeker will soon discover. The fact that so many of them, and so obviously, lead to scientifically and intellectually unacceptable conclusions and practises, may discourage one from further effort. This is again a choice one makes. To either persist or abandon the quest. Failure does not mean that it is not possible to get a result. The first rocket fired did not get to the moon.
Perhaps we can say.. “Science is method. It is not a magisterium of anything”. If it is a magisterium, then there is only one other magisterium and that is nescience”. We have “knowing” and “not knowing”. What religion does is present material which can be scrutinised. Hypotheses can be derived for examination by scientific METHOD. Scriptures are largely derived from experience and all can be used to rationally hypothesise the existence of God. Science is now required to properly examine and test such hypotheses if the scientist feels compelled to make public statements about their relevance and impose his non-belief and agnosticism on others. Gould is perhaps concerned that Gnosticism is directed at him and sullies his personal magisterium of science. The converse could also be true. His agnosticism may be sullying the field of religion and denying the emergence of greater truth, however much he professes otherwise. Religion takes it as a self-evident truth that God exists, because this is fundamental to human consciousness. This is ancient wisdom – it resides within each one of us. Perhaps it is just good fortune that our circumstances of birth and environment (Gould’s “accidental ontogeny”) allow this spark to flicker into flame. But elegant prose does not bring enlightenment nor lead us to an awakening of our true nature. Perhaps God IS consciousness, or better still, consciousness IS God. There are many levels (“mansions”?) of consciousness – this is axiomatic from the ordinary observation of people, let alone from a considered view of deep sleep, dream, hypnosis, fantasy, vision, direct perception and mystic transport. If one considers the Bible, which Gould is happy to quote and use so extensively, one can get a lot more from it than he apparently has done.
In respect of some of the quotes. Gould closes his book with a small part-quotation from St John, which he pointedly acknowledges also has another meaning. So he makes the same mistake that he would condemn in a “bible-thumper”. He quotes only the piece “In the beginning was the word”, to score intellectual points. Gould states that he “knows that the phrase bears another meaning in its original context”. Does he indeed “know” what that meaning is? If he did, he might find himself nailed to a wooden cross. It surprises me always that scientists are not “religious”, when it is so obvious what they need to do and can do, with such scriptural statements. They need to develop a testable hypothesis. St John (as written in the gospel) further stated that this “word” was with God and the word was God. Still further, he states that all that was created was created by “word”. What is unscientific about speculating that there is then a creative force behind space and time, and that this creative force is any different for “science” or for religion? There cannot be two truths. Gould’s insinuation that the “word” is just a written and spoken instrument of debate and discussion is going to take no-one further along the spiritual path than that same discussion and debate. Like so many preachers he has lost the gist and meaning of those words of St John and prostituted them for another end.
St. John elsewhere (4:8) also states that “God is love”. In other words we begin to develop that hypothesis. God = love = word. So the word is not a book or writing or language. Christ is reputed to have said “I am in my Father and ye in me and I in you” (John 14:20). He further goes on to state quite explicitly… “He that hath seen me hath seen my Father”. What does Gould even begin to imagine that Christ is talking about? What should a scientist do? Dismiss this as fantasy? He is of course free to think and do what he likes, and the faculty to take either option is this one which is denied any other living species. Is he free to espouse his view without review or scrutiny? Is this not what preachers demand from us too? In Corinthians (1,3:16) it is also clearly stated “Know ye not that ye are the temple of God”.
The Bible was written nearly 2 000 years ago and we can be excused for thinking that it is just imaginative prose and has no literal meaning. We can also forget that there were no dictionaries and no way in which the language and idiom of the day could also simultaneously used to satisfy the intellectual 2000 years into the future. But we are now being scientists either testing if a hypothesis can be extracted from this ancient writing and then testing that hypothesis. Perhaps we have evidence that man is devolving. If Gould just acknowledges that he does not wish to pursue the matter any further, that is entirely his prerogative. But to impose his feelings upon a reading public in the guise of a text dealing with the fullness of life is not correct because it shuts a door on that fullness. Will we say in another 2000 years time, “He did come again, and again, and again”. What use will that be to any of us today?
The Bible is the source of a great deal more wisdom which actually leads on to describe precisely how one comes to “know” that this body is the “temple” of God. Christ is reputed to have said “If therefore thine eye be single, thy whole body shall be full of light” (Matthew 6:22). Is it just chance that this statement is repeated in the Adi Granth (M4, p1323). Is it just chance that bells and lights figure in the tradition, ritual and ceremony of so many religions? What can be meant? The scientist, the real scientist, asks this question. The fool may shun it, but we have the beginnings of a worthwhile hypothesis that may be worth developing further. God is word, God is love, God is consciousness, He is knowable. How do we take this further?
The Bible does contain all the necessary answers, but it cannot take one to that experience that it so plainly describes. Christianity is also not the only religion after Judaism, nor is the bible remotely the only book of relevance that Gould’s béte noires would like to believe. There are many other texts, including the Rubaiyat, which state the same “truths” or principles that the Bible does. Again, the individual (whether he wishes to be seen as a scientist, agnostic, or atheist) is free to ignore all this and make of it what he wants. Gould’s claim that he is an agnostic based on his opinion that “truly one cannot know” is nothing that I would be proud to acclaim. The ancient Greeks said “Man know thyself”. Were they just guessing at something? Who said “Be still, and know that I am God”?
Gould states that we should “Acknowledge the personal character of these human struggles about morals and meaning and stop looking for definite answers in nature’s construction”. He does go on very meaningfully, but he obfuscates the point. The human body is nature’s chief construction. This is where we are to find what we seek (“seek and you shall find”) and it is the only place where this can be done. In the Adi Granth it is said “True devotees seek within, all others wander in delusion”. The Bible reads “The kingdom of God is within you” (Luke 17:21). “Man is made in the image of God”. Science as a process of organising knowledge should enable and empower us, not drive us from the door.
Gould once made a statement which impressed itself on my mind roughly as follows.. “When I see what is done with those things I know something about, I am very sceptical about what is done with things I know little about”. Instead of being disturbed by the weak arguments of syncretism, there should be absolutely nothing to stop Gould from properly applying his scientific mind to the question of the existence of God. I have personally experienced the failures and weaknesses of the scientific product, and have made many mistakes myself. These mistakes are nothing to do with science, unlike the woes of mankind which often have everything to do with religion. Man was not made for religion and neither was he made to be led up the garden path by mechanistic science. He was made for the purpose of realising God? What prevents us from doing this? Who knows? Gould quotes “blessed are they that have not seen and yet have believed”. Gould’s interpretation may only demonstrate blindness brought on perhaps by undoubted brilliance of intellect and erudition. It is a statement extending the view that “the meek shall inherit the earth”. Does it not state that even those who do not have the intellect or the knowledge or the privilege of high birth or rank or education, can find their way to God and fullness of life?
Gould writes…”A sceptical attitude towards appeals based only on authority combined with a demand for direct evidence, represents the first demand of proper scientific procedure.” This is indeed what is required if one wants to understand the true origins and aim of religions, and achieve the same one goal. This is true knowledge and enlightenment. There are thus only two magisteria. Man has his roots and origins in one, but insists in living in the other where he constantly pursues the illusion of “fullness”. Thus the great deception is that we confound our magisteria of “science” and “nescience” by misconstruction. Consequently, “Born in ignorance, we live in ignorance and we die in ignorance”. Knowledge of our physical universe may be very profitable to have, but in what sense?
This is precisely why I labour the point about classification being science. We have to write in a functional paradigm. In the “Scientific American Book of the Cosmos”, Gould contributes the chapter…”The evolution of life on earth” and he makes this dramatic statement.. “We will (NOT) complete Darwin’s revolution until we can find, grasp and accept another way of drawing life’s history”. (By chance, the omission of the word ‘not’ is a typographic error in the text!) He is saying in more eloquent language in a wider context, the same thing that I wrote in “The case of Haworthia incurvula” (Aloe 36:34, 1999) “we need a new language”. He speculates that we may be limited by “socially imposed conceptual locks rather than inherent restrictions of our neurology”. I speculate that we have intellectually imposed conceptual locks and that we indeed need to examine what Andrei Linde wrote, also in “Book of the Cosmos” …”The Self-reproducing Inflationary Universe”. He wrote that the universe may be a huge growing fractal and that if this model is correct…”then physics (science?) alone cannot provide a complete explanation for all properties of our allotment of the universe”. Further…”Does this mean that understanding…will require…a deep investigation of our own nature, perhaps even including the nature of our consciousness?” Personally I have come to the conclusion that this is a truth which we should all take as self-evident, and that we need to change our conceptual locks – if we want to understand Haworthia fully. ♦
This article appeared largely as follows in Haworthiad, and is reproduced here on account of Charles Craib’s comments included in the following chapter.
I often wonder why I have written and still continue to write about Haworthia. The plants have had a special fascination for me since childhood, but it is not that I really enjoy these plants more than I do many others. The interest for me lay in the problem of identification and naming and I was continually asking where a particular plant seen illustrated or growing came from and what was it and why did the names seem to differ. As an entomologist I came to question all these names and their meaning, and to wonder about the classification. After all, it is the names that we use as individuals or as groups of people to grow, collect and communicate about the plants we interest us. So classification and names are just as basic and fundamental to us as a group of hobbyists as they are to botanists pursuing academic and intellectual truths. The history of Haworthia was clouded with conflict before I started writing and the pattern has continued despite what history should have taught us. I have personally made my best effort to generate a stable and sensible set of names for a community that I would like to be part of. This community I wanted to encompass was that of the ordinary collector, the more dedicated collector, the horticulturist, the commercial grower, herbarium and field botanists, and conservationists.
It has been immensely frustrating to see my ideal so thwarted and to find it so difficult to communicate what I consider to be simple ideas to all the sectors of the community I want to share with. Now at the closing of my life and what career there has been in Haworthia I feel the need to make some final effort. A motivating factor has been the recent publication in the German journal Avonia, of twelve new species and varieties of Haworthia by Ingo Breuer. If I ask what significance this has for the community I perceive out there, I cannot answer the question except to suggest that it may be the commercial value – I can see no other sense in it. The field collection has been done by South Africans who are not accredited by Nature Conservation to do the collecting, neither by any institution that supports their activities as far as I have been able to establish. Is it a product of bona fide research and does it justify the tacit support given by individual botanists who are involved in the writing of forewords and introductions? Regarding the classification involved, I throw up my hands in despair because none of these new taxa suggest to me the taxonomic significance that the descriptions or the describer may attach to them. It is quite probable that the collectors have some individual motivation and are not working through local institutions that could thwart their private goals.
Few members of my imaginary community seem to share my concerns. As a writer, editors are the main vehicles for publication and I have expected them to function as a filter to separate the reasonable from the unreasonable. The kind of reaction I receive from them is that everyone is entitled to an opinion and that the reader can make what he/she wills of any classification put before him or her. An editor has also said to me “I think the “problem” is that most people who like plants that I am addressing with the Journal are just hobbyists. Some are more serious than others. They want a good name, pretty plants, field trip notes, good books to pore over, but not necessarily devour. They don’t want to be confused, to the contrary, most want a hard and fast true answer… as if there is only one or ever will be”. However, I cannot agree. My conclusion from so many years of frustration and intellectual isolation, is that the problems lies in how we have all been led to regard botanical names and their meaning. Readers are already confused and editors are co-responsible. This is what I would like to address in this article.
After describing Haworthia limifolia var. gigantea in 1962, I wrote “Haworthia as a problem genus” in 1970 and that article was published in this Journal. In effect what I said then was that there would never be order unless the species were to be seen as morphologically and geographically distinct entities. I also said that new species and varieties were not discussed adequately in terms of distribution or variability. Among my comments was it would take detailed and intensive coverage of the area before a sample could be taken as representative.
Since I wrote that article there has been a vast volume of water under the bridge and many new writers have come and gone or are still busy gnawing away at the carcase. The taxonomic situation in Haworthia is as confused now as it was in 1970 and before. Why? I think the answer is simple, and sadly so. The roots of taxonomy are in a foundation of shifting sand because a botanical name does not mean the same thing to everyone. As one editor put so succinctly…” There is no doubt that SPECIES should have a meaning, preferably one that is universally accepted so that everyone is on the same level”. Albeit that this conclusion was only reached after a long struggle to get this editor to distinguish between a species description and the definition of the word “species”.
This is a point that I have been struggling to make for a long time and it has amazed me that botanists talk about different kinds of species, biological species, phylogenetic species, chemical species and many more kinds of kinds. The word “species” has always meant to me “life-form”. Most people will agree that it is the basic unit of biology and yet there is this curious and incongruous use of the phrase “biological species” which suggests that there are species that are not of biology. Can one progress through a university curriculum and postgraduate study to a doctorate and still have no idea of what a species is or might be? My observation is that this is fact.
If botanists have not been able to establish a definition and agree on species definition, it is patently obvious that we have all the ingredients for an unholy mess. If there be any doubt about this, I would refer readers to all the literature on Haworthia, including my own. One botanist remarked that I was being silly insisting on a definition when the subject was one of continuous debate! Classification is largely a non-science. It is simply a descriptive activity ensconced in a web of legislative rules, articles and recommendations of a formal nomenclatural code. Anybody can become and be fully accepted as a taxonomist with absolutely no credentials, on the presentation of a description, a Latin synopsis and the citation of a type. If in addition they can quote the clauses and articles of the International Code and throw in some Latin phrases here and there, their stature is elevated. There is a whole field of literature which deals purely with the technicalities of the code and has no relation whatsoever to the plant species for which it was fabricated. It is a vast playing field. The tragedy is that there is no protection for the passive observer or interested party who should feel secure in the knowledge that there is logic and reason in the whole process of naming and communicating about plants.
Currently there are two trends in present time to remedy this unfortunate situation. Both are aimed at taking the subject to abstruse intellectual heights that will positively close the door and make it impossible for the uninitiated to participate. The one trend is to rely on the so-called science of cladistics and the other is to enter the realm of the sub-microscopic and analyze the genetic material on which the (still undefined) species rest. Neither trend is to address the underlying problem.
In my opinion cladistics should be seen clearly for what it is. It is multivariate analysis or character sorting where character states are chosen and modified as subjectively as in conventional methodology to produce a two-dimensional “tree”. This tree is presumed to be the representation of the evolutionary processes in the plants being discussed. This is the element of time. But in this model the element of space (the branch ends) is restricted to one dimension. DNA methodology is primarily based on examining the sequence of nucleotide bases (amino acids) on a piece of the total protein strands that constitute the chromosomes or cellular protein that drives genetics. It is said that these amino-acid sites represent characters and thus looking at, say, 1000 such sites (“characters”) is immensely superior to the guesses we make when we look at say flower colour or bract shape which we could choose to identify a “species”. The argument loses sight of the fact that these 1000 sites on a single gene sequence are but a fraction of the genome DNA (Sahtouris writes that the genes probably account for less than 1,5% of any single genome), whereas flower colour or bract shape may be the end-products of the interaction of many genes or of the entire genome. The-end model is also still presented in the same two-dimensional one of cladistics.
I have suggested that species follow a pattern that many natural systems do. My suggestion is that species are fractal. This means that they possess or exhibit variability that cannot be explained or plotted in linear or smooth curvilinear fashion. But I also define the word “species” – a dynamic system of living organisms in a group or groups of individuals that are morphologically, genetically and behaviorally continuous in space and time. This definition is not proposed as a fairy wand as one critic suggested. It is anything but. It does not make taxonomic decisions easier. It means that there is now a definition and standard in place against which one can question and evaluate species descriptions and taxonomic decisions. We have to see individual plants as members of systems and act on the fact that a name tied to a single dead herbarium specimen is less important than the meaning of the name referring to living forms. More incisively then, it makes it incumbent on the taxonomists to view species as systems in a greater whole of biology. There is a still greater objective and that is to see species as these life forms that give meaning to a creation science otherwise demands that we examine only mechanistically. It would mean that the classification process could be placed back on a proper scientific foundation and exclude individuals who pursue new names for atavistic pleasure, for egocentric and commercial goals, or for particular communities.
My experience has not been limited to Haworthia. I have to say this because inevitably when I meet another taxonomist the attitude is assumed that I am working in a strange and unique field; condescendingly, that I am grubbing around in my own tiny space quite ignorant of the deep thoughts and knowledge of the inner sanctums of science. Generally the genus Haworthia is seen to be taxonomically intractable and largely so simply because it fascinates amateur collectors (and amateur taxonomists). I wish I could demolish this myth for once and for all. It is only intractable because botanists have been so self-satisfied and mollycoddled by lack of peer review in their backroom havens of herbaria that they have seldom really had to face the realities of natural variation. They have worked secure in the knowledge that there is no definition and that under these circumstances one opinion is a good as another. In the general forum of human behavior, someone who comes in from outside with a plucked flower in the hand is almost seen as demonstrating an above natural interest in flora and a flair for botany. With a bit of early encouragement the individual could presumably have been a great botanist. Haworthia is difficult perhaps because it is so well collected and so widely grown.
For a journal editor to say that readers are not interested in the problems of nomenclature and do not want to be confused is a denial and renunciation of responsibility. It is to say that readers are asleep and they should not be awakened; that they have been educated to an untruth and they should be left there. Furthermore, this is any case what we feed them and we have a vested interest in maintaining untruth. There is no general close interest in plants that extends to proper nomenclature. Very few people in the vast mass of society actually have a Latin binomial anywhere in their vocabulary. As an oddball who has grown up and lived with such names, it has been a constant war to have them seen as keys to knowledge. Instead of respecting this fact, it is common for people to proudly deny knowing the scientific proper names for living things.
Classification and nomenclature has become a laughing stock in society and a subject to be avoided. Unfortunately someone has to take it seriously and give it meaning. I have tried to do this for Haworthia for sincere and honest people who deserve better than the untruth and misinformation that is laid before them on a daily basis. My community has dwindled away to nothing. Where I was myself a lost sheep, I now feel like a shepherd with no flock! ♦
This essay is inspired by the chapter, of “Taxonomy and the sociology of botanical knowledge” written by Charles Craib in his book “Grass Aloes in the South African Veld” (Umdaus, 2005). That particular chapter was written purportedly to explain why he prefers the taxonomic revision of Reynolds (1950) to that of Glen and Hardy (2000). Craib states that Reynolds provides “… a better reflection of the infrageneric taxonomy of the group”, and also better suited to “elucidation of processes concerning the autecology of the SA grass aloes.”
Craib maintains that… “The classification used by Glen and Hardy takes its place in the knowledge production process as a more abstract model than that proposed by Reynolds”. However, Craib states that abstraction is a trend “…in the development of SA botanical knowledge and that such trends can be expected in the history of this development”. He maintains that the discipline of a “sociology of knowledge” is essential for understanding how classification systems work as they do, and also useful in accounting for the regular revision of plant genera and changes in plant names. I would like to ask if this argument is true for Haworthia and can it in any way explain the remarkable plurality of names that exist there and being added to exponentially.
In my opinion, Craib’s views are extremely relevant. Not necessarily because they clarify the problem so much as demonstrate that there is indeed a problem. Many of Craib’s assertions need to be questioned if the acquisition of knowledge is really the motive for writing and reading. I am truly seeking closure on the whole issue of why I ever wrote about Haworthia and still do.
Analysis: What does the title “Taxonomy and the sociology of botanical knowledge” actually mean? Taxonomy according to Collins Dictionary is… “the branch of biology concerned with the classification of organisms into groups based on similarities of structure, origin etc.”. Sociology is…”the study of the development, organization, functioning and classification of human societies”; and knowledge is…”information about a subject”. So the title appears to mean that the object of the chapter is to enlarge on the way in which the classification of plants generally is related to the functioning of society with respect to the grass aloes and the communication and sharing of knowledge about them.
The impression I get from Craib’s chapter is that there are two available classifications which present him with something of a dilemma in deciding which one to use. The particular problem in Aloe needs to be properly explained and understood if any contribution is to be made in that respect to the “knowledge production process” in botany generally, and this is indeed what Craib attempts to do. Reynolds’ revision of the Southern African Aloe was published in 1950. It was and still is a remarkable product by a non-botanist (Reynolds was an optician). The publication of this work initiated a remarkable and quite extraordinary popular interest in these plants. Aloes became a collector’s realm and Latin names were the key to the collectibles. A vast new body of information became available and some of this was reflected in new names (species) in the formal literature. Thus it was evident as early as 1970 that a new revision to unify and consolidate that information was necessary. At that time there was also a drive in South African botany for a new Flora to list and synthesize all plant species. In respect of Aloe, it was probably entirely fortuitous that Glen and Hardy were available and interested, to be assigned the task of revising the genus.
The essence of Craib’s thesis is that Glen and Hardy’s classification is a “more abstract model” than that of Reynolds’s and implies that the reasons are to be found in new technologies and processes which inspire the revisionary processes in botanical classification. What concerns Craib is that such botanical revisions invariably lead to name changes and re-arrangements of species. He is not alone in his contention that this process confounds the various segments of society outside and even within botany that use Latin names to generate, accumulate and share knowledge. In Haworthia confusion is rife.
However, it appears to me that Craib’s chapter is only symptomatic of the problems that beset and confound the literature of Haworthia and similar popular plant groups. The revisionary process in the hands of competent taxonomists is required to produce a clearer and better understanding of a genus and its complement of species. If it does not do so and an abstraction results, we need to reflect on the people and the processes. For science to triumph it is absolutely necessary for there to be organized and proper skepticism. If this is lacking the whole process of progress is reversed and we come no closer to knowing anything. This is the problem I perceive in Haworthia. It does not require a “discipline of sociology’ to explain botanical revisionary cycles as these are the natural product of exploration and the inquiring mind. A discipline is needed to examine the increasing abstraction and theorizing of revisions and the rifts that are widening between users of names and the people who generate, change and argue about those names. Not only this, but revisions can appear, as in Haworthia, that have no intellectual substance at all. How does this happen?
Thus any real contribution to the knowledge of Aloe (and other plant genera) in society (Craib’s “sociology of botanical knowledge”) requires a more realistic appraisal of botanical science and the role of individual persons and the aptitudes and competence that they bring to the task. It is people and personalia which trade in knowledge and it is the invidious fact that these are the factors that have to be evaluated and for which there is no provision in any code. It is painful to have to express an opinion on the basis of personal merits. In respect of Aloe, Glen is by observation, a theorist, a nomenclaturist and an historian, whereas Hardy was acknowledged worldwide as a collector, grower and field naturalist par excellence. It is pure fantasy to suggest that a combination of these skills of two different people would necessarily result in a happy union. A key issue, which Craib has not properly addressed, is ensconced in his use of the term “model”. This is when he suggests that Reynolds generated one model for the classification of Aloe, and that Glen and Hardy generated another. This is at the heart of the entire problem and what the sociology of botanical knowledge needs to address. It is where the argument already begins to unravel, because Glen and Hardy used Reynolds’ revision in generating their own version of the SAME model. It should not take any place in the “knowledge production process” unless it merits a place and substantially contributes more information. The simple assertion that revisions become necessary once names have been assigned to the different entities in a genus is not correct. Revisions become necessary when exploration and the publication of new information and names proliferate to the degree that a new statement is necessary to synthesize and re-organize all the available information.
If Glen and Hardy have now produced another classification, it is clearly not as a question of model that it can be queried, it is one of competence. Have any changes, apart from obvious and necessary new additions, been based on tangible new evidence and new information to justify them? Craib provides some instances where he obviously disagrees and it is quite certain that he is the best positioned and equipped to make any contribution towards the knowledge and truth of the matter. If one takes the question of Aloe bowiea and its inclusion in a section Leptaloe along with other “grass” aloes, it is evident that Craib disagrees. Rather than discredit and question this action he makes the cognitive choice that it simply does not serve his purpose. But what is the truth of the matter. Knowledge can essentially only be of something that is true. Aloe bowiea was at one time the unispecies genus Chamaealoe africana. Despite the argumentation and merit put forward by Smith and van Wyk(1993) it is not an absolute certainty that its transfer to the genus Aloe represents any fundamental truth. It is unlikely that Glen and Hardy had any better insights and information by which they took a further step and put the species in the section of grass aloes. Any “sociology of knowledge” has to acknowledge that while “species” may be real, genera and infrageneric groups may very often not be so. Thus the transfer of Chamaealoe africana may be justified in several ways, but the driving force may have been derived primarily from the assessment… “This resulted in the unsatisfactory situation we have today where 27 generic names are available for some 450 species which could easily be accommodated in seven genera”.
The classification model used by Reynolds and that used by Glen and Hardy are depressingly and disappointingly exactly the same. There is no difference. The model is the two-dimensional dichotomously branching tree which is universally used to depict evolution and the relationships of species at the various levels of the classification hierarchy. All Glen and Hardy have done is to add in the new names not known to Reynolds and to shuffle a few about on the basis of some new information (hopefully) or on the strength of their opinions. It is an unfortunate reality for us that Reynolds is unavailable to express his opinion of this new product, and no doubt fortunate for Reynolds.
The question being asked in the botanical literature is if this existing old model is in fact adequate and is the binomial nomenclatural system valid. The consensus seems to be that the system does work and besides that, new advances in scientific technology (molecular biology particularly) are in any case producing such dramatic new insights into the genetics and hence the evolution and phylogeny of species. Hence any debate on the fundamentals of any model can be put aside for another time and place. The problem does not go away. It can be categorically stated that Haworthia will never be satisfactorily understood and interpreted on the basis of the current model as used by either Reynolds or Glen and Hardy. My contention is that this will be found to be true for many other plant genera.
This is a reflection on the model used by both Reynolds and Glen and Hardy in which families genera and species should be accommodated in a formal near symmetrical and easily managed hierarchy. This is the “sociological” aspect which determines how names and classification should be understood. The further examples that Craib cites where Glen and Hardy depart from Reynolds, has less to do with abstraction than it does with the competence and integrity of the taxonomic decisions. Craib is in fact evidencing the better knowledge on which a classification can be based without committing or exposing himself to the scrutiny of the botanical community (professional and very amateur alike) and which can indeed be unpleasant, trivial and prejudiced. The South African National Botanical Institute accepted the Glen and Hardy revision of Aloe, and Craib tacitly denounces it.
Craib raises one further issue which is extremely important. This is that there are groups and individuals who have different needs in respect of classifications and that some of these groups require certainty in plant nomenclature and identification. The answer is not to have different options in terms of classifications that can be used. The answer is to generate one sound classification which is not abstract and which can be properly understood.
The basic unit of this classification is the species and this is what Latin binomials denote. Do we ask how present society (the community that has a non-professional interest in plants) understands these names and their arrangement (taxonomy)? Do we ask if taxonomists understand what it is they are doing and if what they are doing is meeting the needs of society? Do we ask if taxonomists and society have and share a common definition of the “species”? In the role that language plays in sociology, names are given to objects to facilitate communication and thus an association of names and objects is built. The foundation of plant taxonomy is that single specimens serve as the primary objects (the types) to which any name is attached. Unfortunately it happens that the association of those names is very often built around objects other than the primary specimen. That is one source of confusion. The other source of confusion is the association of the single prime specimen or type with others of the same kind. There is no single common perception of what constitutes a single kind. The “single kind’ is of course the “species”, and while definition and perceptions of this differ from one person to another, there can never be a true shared knowledge about how they are classified. If we consider consciousness and experience, we have to recognize that life is manifest in an array of living things at many different levels of organization from simple to complex. We recognize these as different life forms and we use the word “species” to denote them. It is remarkable that the system of plant names works at all given the fact that there is so much difference in the minds of people of the associations that exist between Latin name and its primary object. A Latin binomial is not a common noun and a name for a single thing. The root of the problem is that the association of a name with a type, and the association of that type with any living system of organisms, is confounding. It is ridiculous to expect the Latin binomial system to be descriptive of all botanical variation. It is equally ridiculous that interpretations of the meaning of the Latin binomial and its attachment to a hopelessly inadequate definition and diffuse usage of the word “species” remain a sociological problem. Taxonomists, whatever the individual idiosyncrasies may be, are striving towards a classification which truly reflects evolutionary processes. There cannot be alternatives, but we also need to realize that there is embedded knowledge that becomes associated with the use of a name. It is absolutely incumbent on persons who generate name changes, and on user groups, to mutually and cognitively ensure that names and changes serve a common “knowledge” process.
It is just an obvious fact that in society, interest in plant names can range from the trifling collection of oddities to the deep wish of individuals to understand creation itself. The ancient Greeks had a saying “Man know thyself”. To many this instruction has no meaning, but there is a word in language which indicates that to a few it is important. This is “teleology” – do natural phenomena have a predetermined purpose and are not just determined by mindless mechanical laws? Can we start to consider this fundamental question if we do not properly understand those phenomena we observe? People are engaged in taxonomy in a way which may only meet the needs of a segment of society, but which does not meet the needs of science nor can serve the interests of a cognitive thinking greater society.
Conclusion:
My observation is that the knowledge of taxonomy within society is woefully inadequate. Hence “sociology of knowledge” is unlikely to find any more science than art in classification. It has been said that in botany the measure of a classification is that of the nomenclatural juristics rather than that of the practical usefulness or even scientific accuracy and relevance. It seems unlikely that the botanical community will come any nearer to providing better practical classifications in the future than they have been able to do in the past. Few botanists can match the application, energy and enthusiasm that a genuinely interested amateur brings to his chosen subject. A revision can only be a reflection of the degree of sampling and the capacity of the observer to synthesize what he has seen and, axiomatically, the usage which can be obtained from that revision will be relative to the knowledge of the user. It is evident therefore that the gauge of the usefulness and value of a revision will lie with the knowledgeable user.
Such users may be few and far between and Crain happens to be one of them. If there were more like him botanical classification might in fact be found to be quite fraudulent. An additional part of this problem is the excessive fragmentation of the literature and the huge volume of literature that is being produced as a by-product of the need of scientists to validate themselves. This confounds the knowledge production process and non-professionals find themselves able to join the ranks of taxonomy without much fear of redress.
A new paradigm is needed by which the Latin binomial serves the needs of science and is appropriately understood and modified to also meet the needs of interest groups. What a specialist group needs to do is to cognitively recognize the ideals of science, the need for proper and good information and a stable reliable structure of meaningful names of equal value by which they can communicate. Societies, committee and journal editors need to similarly recognize their responsibilities in terms of the stated or unstated objectives of their activities. This is surely to ensure that a screening process is in place by which the interests of their members and subscribers is protected and served. Individuals need to be identified who genuinely have the credibility and skills to effectively and truthfully evaluate comment similar to Craib’s and my own, and identify and apply corrections where they are needed. These in turn need to be fed back to, and respected by, competent scientists. Finally we need to find a less tortuous and circumlocutory way of expressing what needs to be said.
Acknowledgement: I would like to express my appreciation to Charles Craib for his positive and constructive responses to my communication with him.
COMMENTS ON THE SOCIOLOGY OF KNOWING HAWORTHIA AND HAWORTHIA AS A PROBLEM GENUS – 34 YEARS ON.
By Chareles Craib
Bruce Bayer has written two very useful, interesting and insightful essays with the above titles. One of them “Comments on the Sociology of Knowing Haworthia” refers in part to the chapter “Taxonomy and the Sociology of Botanical Knowledge” in my book “Grass Aloes in the South African Veld” (Umdaus Press 2005: 10 – 12).
The purpose of these comments is to look at the kinds of analysis which the Sociology of Knowledge can provide with specific reference to some aspects that Bayer has written about. The techniques used for analysis examine the knowledge production process. They do not provide alternative types of knowledge nor do they replace existing paradigms. Changes to existing knowledge or paradigms always originate from the knowledge producing communities themselves in this case people involved with the disciplines and sub-disciplines of botany.
Bayer correctly points out that Glen and Hardy use the same paradigm in their revision of Aloe (2000) as Reynolds (1969). The model however which Glen and Hardy use (meaning the constituent elements and concepts in the classification) is presented at a level of abstraction different from that used by Reynolds. Reynolds taxonomy was essentially rooted in extensive fieldwork with numerous justifications provided for his classifications, based on observable characteristics of plants. Glen and Hardy rarely, if ever in grass aloe, provide justification for the nomenclatural changes they make. I cite a number of examples of this in the book (2005: 33 – 34, 39 – 40, 50 – 52, 57 – 61, 75 – 76, 85 – 86, 88 – 90, 101 – 102 and 123 – 126). Essentially the abstraction does not represent any advance from the classification that Reynolds presented and is probably less accurate as it has little to do with aloes in the veld.
Techniques used in the Sociology of Knowledge judge this revision as another attempt to classify aloe with a number of shortcomings in the way in which the knowledge was articulated. Thus this revision does take its place in the knowledge production process concerning Aloe. The use and value of the revision must be determined by the botanical community itself. In the book I essentially used the techniques of criticism, provided for in the sociology of knowledge, to look at what existing classification provided the best model for an autecological study of grass aloes.
Revisions of plant genera occur for several reasons. As Bayer points out (2006 ….) a proliferation of plants names may necessitate a new synthesis. Other reasons are that analysts with different concepts of species, genus and family validly publish new species. All that is currently required for the naming of a new species is a description that has been validly published. The new species cannot be proved to exist empirically. This process will continue to have but a semblance of accuracy until such time as botanists are able to formulate a universally accepted code of what a validly published species concept comprises of. This is very significant since species are the primary units of understanding before grouping into families and genera. This problem has been extensively recognised and discussed by Bayer to some degree or another in most of his many publications.
The Sociology of Knowledge has one primary concern here and that is the characteristic that plant classifications probably represent an art rather than a science. This implies that unless paradigms change there will be any number of revisions of many plant genera in the future. Users of classifications will need to decide which one best suits their purposes. This will be based on principles such as the internal logic of the proposed system and whether or not it represents an improvement from a previous model for understanding the genus in question.
The sociology of knowledge focuses on published information. The competencies of botanists are important to the botanical community when plant genera are revised. However, from the point of view of the analyst of new knowledge, trends in knowledge production processes need to be examined, particularly those which might lend to paradigm changes.
It would be helpful if there was one universally accepted classification of a genus at a given point in time. In terms of the currently accepted methods for producing botanical knowledge this is unlikely to be achieved for two important reasons. The concept of species has no defined universally accepted boundaries and any newly described plant owes its existence to the fact that it was validly published. The huge amount of leeway that this provides will ensure a proliferation of names and provide room for various interpretations of what “species” are. If the concept of species is already abstract (it is the primary unit of understanding) then so much more so the higher level concepts of genus and family.
It is incumbent on the botanical community to change the methods for producing their own knowledge. This will require a paradigm change in the knowledge production process. In this respect perceived inadequacies of the status quo will be required to motivate botanists to change the way in which plants are classified.
Experience with plants in the field is and will probably remain essential for any plant revision. This is not always recognised as the formal point of departure for revisions and is likely to account for the abstractions that are published, which bear little resemblance to the physical reality. None of this is controlled by formal requirements and as a result it is legitimate to publish any kind of revision. The sociology of knowledge in this instance is only concerned with the consistency with which the internal components of the model are articulated at the level of abstraction they are presented.
There are several questions which will need to be asked and answered before botanists will be able to provide alternative or improved paradigms for the presentation of knowledge. One of the most significant is that empirical techniques of science which can be proved and demonstrated are used uncritically to elucidate concepts such as genus. The legitimacy for the combination of these two fundamentally different knowledge types needs to be theorised at an appropriate level before it is accepted as legitimate. Another problem is that there is probably insufficient motivation for botanists to involve themselves in these sorts of significant theoretical debates. A solution may be an expanded range of courses offered to botanists at tertiary level. More people would enter the discipline of botany with an enhanced range of skills for facing these sorts of intellectual challenges.
In a very interesting book by Stephen Gould entitled “Rock of Ages”, in which he propounds his principle of NOMA – non-overlapping magisteria. This states that science and religion should not be confused nor mixed.
So this is not a confession of confusion – you do not confess to what is obvious. It is an admission, and an admission can be construed as an apology. But, as a rhetorical question, how can one apologize and expect forgiveness when one continues to walk the errant path?
I started to write about Haworthia to dispel confusion, and yet more than 40 years on, this confusion has not become any less. The conclusion I have come to (and I wish it was a closure) is that the prime source of confusion is simply the human condition. In mystic philosophy one can read… “Born in ignorance, we live in ignorance and we die in ignorance.”
I think that my interest in Haworthia stems from my conscious effort to dispel this primal confusion and find some of the order in my view of creation. The classification of plants suggested just one small piece of my world which was available to me, and Haworthia as one group which no one else could explain to me. What have I now learned and what contribution does this make to dispel confusion?
My courage to now say something more directly arises from a recent request by SANBI to write a synopsis of Haworthia for an E. Cape Flora. I feel that I have done that fairly successfully. The problem is now to produce a similar product for the SW Cape and this is considerably more difficult.
To explain Haworthia one has to first cross the hurdle of the present classification of the Asphodelaceae and the perceptions that Aloe, Haworthia and other genera are clear discrete groups that the classification suggests. Ignore completely the issue of whether the so-called species are clearly separable. Haworthia consists of three quite discrete groups which have no closer relation to each other than do any other of the extant Asphodeloid genera. They each have their own problems and idiosyncrasies but it is the subgenus Haworthia which concerns me most.
In 1975 I prepared a map to illustrate the relationship of the SW Cape species (primarily south of the Langeberg Mts.) and this was published in Excelsa (5, 1975). In this map I tried to illustrate the problem of continuity which simply makes it impossible to recognize clear closed groups of plants which we can with any truthfulness say are species. It is the reality for Haworthia, and my conviction is that the model which underlies the Latin binomial system is fundamentally flawed. It works because our ignorance clouds its shortcomings.
Map 1 Distribution of SW Cape species of Haworthia
Updating my map may show why (Map.1). It is perhaps possible to identify three main elements that are nevertheless extremely difficult to map. Very seldom do we find all three growing in close proximity and never do we find any of the components I suggest doing so either. We often struggle to suggest which of the three we are being faced with.
The names we can use for these three are:-
H. retusa – to include or cover mutica, turgida and pygmaea
H. mirabilis – to include or cover maraisii, magnifica, emelyae and heidelbergensis
H. floribunda – to include or cover parksiana, chloracantha, floribunda and variegata.
There are other elements viz:-
H. herbacea – to include or cover reticulata, maculata and pubescens
H. rossouwii –
But this is a simplification and not a solution.
What simply has to be recognized as a fact of life that this problem is not of my creation and I do not believe that the use of any technical sophistry, trick of classification or nomenclature will possibly dispel it. These “species” are as interconnected as the strands of a complex web as intricate as any a confused spider could construct.
Essentially one has a mid- to late-summer flowering group and a spring flowering group, but they are linked. H. pygmaea, in the retusa complex, actually springs from the mirabilis complex; while its counterpart mutica in the west, seems to spring from the retusa complex. Emelyae, in the mirabilis complex, seems to be linked back to rossouwii via multifolia. These are the kinds of realities one has to deal with.
The relationship of the three main elements I have mentioned differs from point to point on the map, and to describe these relationships means that one has to virtually deal with populations and groups of populations one-by-one. I have done that, particularly in the book Haworthia Update Vol. 1n and again in Vol. 2. But since the drafting of the last manuscript, I have been to the field again and found more material, which I will show now.
Essentially there are three areas which I will cover:- a. the southern Potberg and where the Breede River enters the sea south of Swellendam (Map 2); b. secondly Klipfontein at the northwest of the Potberg (Map 3); c. thirdly an area just west of that which I also call Die Kop, Wydgelee (Map 4). The localities are not indicated precisely on these maps but the detail is in archived and available information.
Two of three relevant populations are discussed in Chapter 15. These are ADH594 Sandhoogte, and MBB7248 Lower Breede River H. mirabilis (pilosa). I refer to CB2018 and I can now report and illustrate it myself. This is MBB7251 (Figs 2a & b) from Buffelsfontein within the DeHoop Nature reserve. In the field the plants are not distinguishable from ADH 594 and grow in exactly the same Fynbos habitat viz a very rocky and grassy streamside slope with a dry northern aspect. The illustrations are of plants cultivated and hence far greener and softer than in habitat. What is striking is the tendency to surface spination which is such a feature of MBB7248.
b. Northwestern Potberg.
Some of these populations are also covered in Chapter 15. These are MBB6890, MBB6886, MBB6882, MBB6889 and MBB6544 (figs A18…22). My comment that H. variegata occurs here too should be borne in mind. This is on the upper slopes of the extreme western end of the Potberg. There is another record of C Burger’s from further east and south of Diepkloof for which I coined the name H. variegata var hemicrypta. Some doubt will now be thrown on the wisdom of that decision. The mirabilis populations referred to above are on the farm Juliesfontein and my new records are now east of that towards Diepkloof along the Breede River.
1. MBB7487 H. floridunda. Byeneskop (figs 3a to g). No good opinion is ventured in Chapter 15 concerning the populations noted above. Those populations seem to fall in the context of an interaction between H. maraisii/heidelbergensis/floribunda and I warn of infusion of H. variegata. The Byeneskop is on the very eastern border of the farm Juliesfontein and the plants are surprisingly and confidently comparable with the typical form of H. floribunda. The leaves are generally fairly strap like and flattened to the characteristic twisted ends as evidenced in floribunda. A few plants (see fig.3f) do have shorter and stubbier suberect leaves which characterize so many collections that confound the recognition of the relevant species. This particular habitat can be described as Renosterveld on silcrete with dominance of white clay and much grass and other succulents. Similar plants were also seen about a kilometer eastwards on the verge of vegetation transitional to Fynbos.
2. MBB7492 Klipfontein East (figs 4a to d). This is again a typically H. floribunda-like population. Whereas the preceding tends to be slightly tuberculate with the rougher greyer texture and colour of H. maraisii which would suggest floribunda var dentata, these plants tend to have a smoother leaf surface. Pictures of the flower are included although of very little diagnostic significance. The habitat is a steep rocky sandstone hillside vegetated with low Fynbos species.
3. MBB7494 Klipfontein East (figs 5a to d). The plants suggest H. floribunda except that the colour of the leaves is greener and there is a tendency to translucence, or less opacity of the leaves. The white flecks in the leaves are quite conspicuous and are more often encountered in H. turgida. Their significance taxonomically is probably nil as these markings/inclusions may appear in plants as remote as H. angustifolia var baylissii on the Zuurberg, E. Cape. The habitat is again short grassy Fynbos but unusual because of the presence of finely textured white quartz rock.
4. MBB7495 Klipfontein East (figs 6a to c). While we have been looking at plants on a series of buttresses of the Potberg, these plants are now closely associated with large white quartz boulders. They are small and although very similar to the preceding, the leaves seem to have lost the characteristic rounded ends and the margins are relatively strongly toothed.
5. MBB7496 Kleinberg (figs.7a to g). This population is only about 1km east of the preceding but there is a transition to Renosterveld. The habitat is again silcretious with the typical white clay substrate. This seems to be a very erodable surface and plant cover is quite low. The plants are surprising because they depart quite radically from the floribunda-like mode of the preceding populations. They are now distinctly magnifica-like and particularly reminiscent of var atrofusca except much smaller. Curiously a population across a small valley southwards in sandstone and Fynbos is in keeping with the floribunda-like finds westwards.
6. MBB7499 Juliesfontein (figs 8a & b). We returned to Juliesfontein to look at a promising rocky slope between Byeneskop and our previous closest collection on Juliesfontein. These are almost directly north of H. variegata and as can be seen from the pictures they revert to the forms described in Chapter 15 for that area viz. there is a similarity to heidelbergensis. The more leaves are more frequently pointed rather than rounded and translucence and venation is more conspicuous.
c. Die Kop, Wydgelee.
The nearest plants dealt with in other chapters are MBB7539 at Tarentaal that I liken to magnifica var atrofusca and MBB6030 at Napky which is maraisoid with floribunda-like character (type locality for H. maraisii var simplicior with rotate leaf-tips). It is important to note that the Tarentaal plants are not uniformly larger that my identification suggests and also that H. mutica is also present at its most southeasterly locality known south of Napky. Tarentaal only just appears on the map and the mutica locality is just off the map to the upper right (northeast) corner. It should be noted that populations assigned to H. heidelbergensis occur to the west at Kathoek, Beyersdal, N. Windheuwel and terminating in the contentious var minor at Rooivlei north of Bredasdorp. I think if an overview was now made of all these populations I have assigned to heidelbergensis in different chapters and in different publications, it will have to be concluded that it may represent the pivotal element in the whole classification process. Where it was the least represented species in the period preceding publication of Haworthia Revisited it is now far better known and any hypothesis prepared to test relationships in these southwestern Cape species Haworthia will need to question this centrality. Die Kop is a set of 5-6 silcrete and ferricrete hilltops of varying size and height but otherwise not significantly different from a large number of such habitats extending from Stormsvlei in the northwest and terminating against the Potberg and Breede River in the southeast. Collections from several of these have already been reported on in Chapter 15 with the general observation that their identification is in the uncertain linking maraisii and mirabilis. I hope, in the minds of readers, that I have not excluded heidelbergensis from this milieu.
1. MBB7500 Die Kop East (figs 9a to v). I doubt if any collection has generated such reservations in my mind as this one has and many illustrations are used to express the problem. The plants are large and up to 80-100mm in diameter. They are closely spaced in a small grassy, non-rocky area which seems to be geomorphically stable. They are extremely variable and resemblances extend from badia to retusa to mutica. The leaves may be short and rounded or fairly long and attenuate, closely adpressed to the ground (recurved) or sub-erect. One notable feature is that the plants have a milky colouration to the leaves that Seven Hammer once remarked on as a feature of H. mutica. While variability is known to be great, it is seldom that it is as dramatic as this in a population of plants as they occur in the field.
2. MBB7502 Die Kop West (figs 10a to c). The plants here are smaller and more widely distributed and scattered over a larger area. The habitat is skeletal and rocky and the vegetation more herbaceous than at the eastern site. The similarity of the plants to those of 7500 is close but the variation is less extreme. There is less evidence of a mutica influence and a return to more or less the maraisii/floribunda/heidelbergensis archetype.
Fig. 9 7502 Die Kop W
3. MBB7503 Die Kop Mid (figs. 11a to i). The first impression I had was that the plants included more mirabiloid influence. Not as big as 7500 and also scattered on a skeletal slope. At one point there was a gravel patch with Gibbaeum austricola which is commonly found on such gravel patches associated with ferricrete inselbergs.
4. MBB7504 Die Kop North (figs 12a to k). The plants are smaller and there is a more evident floribunda influence. I did not think that these plants were significantly different from the many populations discussed in Chapter 15 between Tarentaal and Stormsvlei.
Conclusions: It should be very evident from all the information provided so far that more information is not producing a better picture of discrete elements. To the contrary and this reinforces the conclusion already mooted, that movement to a new view of “species” in the subgenus Haworthia is necessary, but is it possible?
Acknowledgement: Mr. Keith Spencer, DeHoop Nature Reserve. Messrs. Christine & Malcolm Wallace, Ziekenhuis. Mr. John Douglas-Hamilton, Ballyfar. Mr. Johan Groenewald, Klipfontein, Potberg. Messrs. Saartjie and Neil Neetling, Juliesfontein. Messrs. Adele & Vlooi DuToit, Die Kop & Frederickskraal. Messrs. Pet & Johannes Uys, Die Kop. Messrs. Sofia & Jurie Vermeulen, Tarentaal
Since the publication of Haworthia Revisited (Umdaus 1999) I have written Haworthia Update Vol.1 (Umdaus 2002), and Haworthia Update Vol.2 is now in press. It has some 700 pictures and several maps and focuses largely on the vexing question of how we classify and name the plants in the social environment we have created.
In one chapter I demonstrate how difficult it is to distinguish H. mirabilis and H. maraisii and admit that I can no longer see that distinction. In this article I want to illustrate a population which shows that my problem is not limited to those two names. This is about a population on the farm Klippoort that is at the extreme southeast of the Worcester/Robertson Karoo. It is south of the Riviersonderend River just before the confluence with the bigger Breede River. The Riviersonderend runs south of the range of mountains which effectively forms the northern boundary to the distribution of H. mirabilis. East of Stormsvlei, the river cuts through the tail of the mountain range and turns northeast to link up with the Breede River. The confluence lies just south of the Bonnievale/Drew/Mardouw area which is populated by a dense array of variants of H. maraisii and H. heidelbergensis that cannot confidently be regarded as two different species either.
I have given my record a new number MBB7513, because I do not think I ever made a formal record. I am not even certain when I first saw the plants there at Klipfontein but I do know that I was again there in 1996. I was familiar with plants in the nearby area which I had no doubt were H. maraisii and remembered the Klippoort plants as small with rather erect incurving leaves. In October 2005 Kobus Venter, my wife Daphne and I were in the area and saw a Drosanthemum which I knew was important to Dr H. Hartmann. To show her these plants in February 2006 I had to get permission from a new landowner and this led to a curious exchange with the original owners (Mr. and Mrs. Urschel) who had retained the main part of Klipfontein. To demonstrate the significance of the area and to fuel her interest, I thought I would show Mrs. Urschel H. maraisii on the part of the farm they had retained.
After much searching in the very dry conditions then prevailing, we found one single clump which was coming into flower. We decided we would try to get back to see if any seed would set and to collect and grow same to increase numbers and replant in a seemingly disturbed habitat. We returned in mid-March to find the plant had lost its flowers, had been disturbed by some animal or other as one head had been dug away (see Figs. 1 & 2) and was in bud again. Unfortunately my digital camera ran out of power and I could not get a better picture. We then searched again quite thoroughly, but now aided by the fact that flowers were appearing. We found many plants hidden in the moss and litter under shrubs and also in the dense lichen on the huge conglomerate boulders forming the rim of the very small rocky hill. Anxious to get seed and a good picture, we again visited the site in early May after it had rained. Most of the seed was gone and we virtually gave up hope of finding any and set about trying to get some pictures. To our surprise and despite the thoroughness of our previous searches, we now found many more plants and in two small spots they even formed large dense clusters. There were also mature green seed capsules and some ripe seed – and also plants in flower and in bud.
Fig. 1. MBB7513.3 H. maraisii. Klipfontein, Stormsvlei.Fig. 2. MBB7513.4 H. maraisii. Klipfontein, Stormsvlei.
The curious thing for me was that I had no doubt up to this moment that the plants were H. maraisii confirmed by the dark grey-green colouration of fig 1. So you may not imagine my surprise when the first cluster I saw was a very green plant with many spreading leaves and notable end-surface translucence viz. H. heidelbergensis (figs. 3a & b). I also had a memory of plants rather as I have written about near Robertson, which are pubescens-like with surface fuzz (see fig. 4a, b & c). The plants do indeed have surface fuzz and also the plants figured are solitary and with incurving leaves.
Fig. 3a. Klipfontein 3rd May 015Fig. 3b. Klipfontein 3rd May 016Fig. 4a. Klipfontein 3rd May 006Fig. 4b. Klipfontein 3rd May 014Fig. 4c. Klipfontein 3rd May 012
Fig. 5 illustrates a plant which has the more rigid and less translucent leaves of H. maraisii, and figs 6a, b and c are of similar solitary plants. Figs. 7a & b are clustering clones. The plants are all small with the rosettes seldom more than 30mm diam., although the spread of leaves of a clone as in 3b may be as much as 70mm. The flower stalks bore few flowers and one is illustrated in Fig. 8 although I have been able to make little use of flowers in drawing any conclusions.
Fig. 5. Klipfontein 3rd MayFig. 6a. Klipfontein 3rd May 008Fig. 6b. Klipfontein 3rd May 010Fig. 6c. Klipfontein 3rd May 013Fig. 7a. Klipfontein 3rd MayFig. 7b. Klipfontein 3rd MayFig. 8. Klipfontein 3rd May
The habitat is a curious mixture of plant species. There are milkwood trees which can grow large, as well as Acacia karoo the common sweet thorn and Lycium which is also a medium sized spiny shrub. These are often along watercourses and here the Riviersonderend is barely 50m distant. But there are also Renosterveld species such as renosterbos itself, and karoid plants. The large conglomerate boulders are pocked with missing tennisball size stones, so that the dense lichen cover has many niches for plants. Apart from miniatures like Schizobasis, Holothrix, and Drimia pusilla there are also miniaturized Crassula, Adromischus and many tiny seedlings of the large Tylecodon paniculata. My observation is that the vegetation of many such sites bears very little relation to vegetation maps and plant associations mapped at large scales.
The site is fairly disturbed as Bluegum and Black wattle are invasive along the river. The farm itself was controversially planted to Redriver Gum and Pinus pinaster and planted gums impinge so closely on that site that Haworthia can be found in the stone and moss among the closest planted trees. The greater area is very marginal for farming on very rocky soil marginal to the sandstones of the higher mountains so that forestry option seems a very unpromising and unfortunate one. A larger similar rocky area to the northeast has been, and is scheduled to be mined again, for manganese. There is nothing to suggest that the Haworthia ever occurred there. Looking north across the river to the Bokkeveld shales one again sees the virtual ecological desert of total cultivation. Very few habitats are apparent. It is often speculated that many plants (particularly specialists like Haworthia) must have perished due to this cultivation. But my impression, based on the known collecting record of the last 80 years as well as on my own experience of where the plants occur, is to think it actually very unlikely that plants did occur where they presently are not. I followed up some records made in late 1930 by messrs. Otzen, Luckhoff, Herre and Malherbe and am confident that plants can still be found at most of their sites. Similarly the localities recorded by Dekenah and Smith are similarly still extant although they must be seen as under threat. What does cause concern is that the lost habitats are largely small rocky sites with surface rock that are targeted by road builders. Road maintenance has become a major problem because the road reserves which used to form uncultivated strips of natural vegetation are now cleared fence to fence in the case of new roads. In the case of existing and minor roads there is a constant attack by road clearing gangs, fire and weedkiller spraying. In the agricultural areas there are huge expanses I see as ecological deserts. The rocky and skeletal habitats are under severe pressure from animals and the introduction of new sheep breeds with more aggressive feeding habitats is a severe threat to these small islands. Farming has also become more of an industry and business and production is the goal irrespective of conservation ideals. This particular habitat seems to be as safe as it can possibly be by virtue of its intrinsic nature and in respect of the above threats.
Conclusion The distinction between H. maraisii and H. heidelbergensis is too tenuous to enable positive identifications and this Klippoort population displays more obviously the problem of variability within populations.
Acknowledgement Messrs. Leanne & Pieter Urschel, Klippoort East. Messrs. Cindy and Jon Webber, Klippoort West.
Dr. Canio Vosa in Caryologia 57:4, 395-399 (2004) hypothesizes two superspecies for the subgenus Haworthia. He cites my published works including Haworthia Update Vol 1 (2002). It is not clear if he has considered all the species that I recognize, or considered that if there are superspecies there may ipso facto be ‘inferior species” too. It is also not entirely clear that he as followed my argumentation. He writes…” the morphological characters, as used for species definition of the taxa in question, do not give a clear indication of true discontinuity over their geographical range which in some case(s) is rather restricted”. As early as 1972 I pointed out that there were fewer species rather than more, and most of my works have featured some kind of statement on the problem of identifying any discontinuities, other than geographic that suggest a realistic concept of species as systems.
A problem I have also had to face is that there is a very strong user group for names in Haworthia in terms of collectors and growers and it is extremely difficult to balance the different needs and perceptions of these non-professional but knowledgeable people against the needs and perceptions of professional botanist and other scientists that have often proved to be highly flawed. Also to be taken into account is the ground truth and what I would regard as objective reality. This entire Vol.2 is dedicated to extending this ground truth to a point that I can claim that there is enough now known to make a truthful statement. This is the point of Chapter 24 of this Volume where I provide what I think constitutes a rational list of names under which all that I have seen can be catalogued with some degree of confidence. It is from that point that I believe scientists, like Dr. Vosa, should hypothesise and test.
In this last chapter, I myself am going to propound a partial superspecies concept. On request from The South African Institute for Biodiversity, I have already generated a species list for a proposed flora for the Eastern Cape. That list excludes other geographic areas and here I limit my discussion to principally those species concerned in the chapters of this book and the southwester Cape.
Where Dr Vosa asserts that…”the subgenus Haworthia constitutes a single polymorphic assemblage of species which can be subdivided, albeit with some difficulty in some cases, into two groups”, he is not making a ground-breaking statement and neither is it accurate. In all three subgenera it is often quite difficult to allot specimens to groups that would then constitute species. At least one of the chapters explains that this problem extends to the genera of the Asphodelaceae. The problem in the subgenus Haworthia is simply more acute and there are more than one polymorphic assemblages.
Dr. Vosa makes further assertions about the role of specimens, types and herbaria. These have to be examined because there are profound misconceptions which fuel the opinions of users in any of the user groups. The role of the type is not intended to designate a species. It is solely to substantiate a name and link that name to a tangible reference point that may be a herbarium specimen or an illustration. Choices in this regard are the functions of taxonomists and nomenclaturists which are governed by a very comprehensive International Code for Botanical Nomenclature (ICBN). A revisioner would look at all those types in the process of reviewing all the literature and additional specimens and then generate a synonymy to put together any names which he thinks can be attributed to the same species set. This is all confirmed in the list of specimens, and any published or formally deposited illustrations if necessary, which are then cited to formally convey the total concept of what that revisioner thinks constitutes the “species’. Unfortunately the necessities of nomenclature do not bring the stability and understanding of names to the point where confusion and misunderstanding is totally excluded. Examining all the taxonomic literature may even suggest that there is a preoccupation with nomenclature which displaces the purpose and function of classification. This is only partly relevant to the present discussion in view of the choice by Dr. Vosa of H. cymbiformis as the typical species for one of the superspecies.
Why I say this is because in my presentation for a Flora of the Eastern Cape, I have seen the necessity for a superspecies concept and my decision has been that H. cooperi is the superspecies and H. cymbiformis an independent species. There is a problem created by the priority requirements of the ICBN. H. cooperi can in the field, seen to be the dominant and most widely spread element whereas H. cymbiformis comprises a smaller part of this complex. Were they amalgamated in a superspecies, the epithet cymbiformis would have priority and implementing this would distort the perceptions of the relationship.
I fully agree with Dr Vosa that species such as H. bolusii and H. decipiens could be included in one superspecies with H. cooperi but I do feel that too much usefulness will be lost if this is implemented.. While there is no doubt that very strong cymbiformis look-alikes emerge from ecotypification of H. cooperi, there is a concomitant set of populations at a higher level which have an independent distribution. The evidence for converse ecotypification is less persuasive. Certainly to document this kind of evidence by way of herbarium record is totally impractical, as Dr Vosa points out. The object of this book and of its predecessor was primarily to overcome this obstacle by providing an extensive pictorial record.
Regarding the two superspecies conjectured by Dr Vosa. Here I have to flatly disagree on the candidature of the two lists. Quite plainly H. reticulata is bound to the excluded H. herbacea and cannot be included in the superspecies H. cymbiformis. I started my intensive field career trying to understand the relationship of the former two species and how they related in terms of superspecies thinking to their geographic counterparts. My observation was the curious one that they were geographically contiguous or complementary. At several places they grew virtually together and hybridized whereas certain populations could not be positively referred to either of the species. Wider experience suggests that this kind of situation is deeply imbedded in classification and identification of many plants. I have not considered the problems this causes as a sound rationalization for lumping. I would rather suggest a growth process or learning curve that user groups and individuals truly examine their own definitions and understanding of species and classification. This will require a paradigm shift in how name are used without any tinkering with the system in place.
Dr Vosa presents six colour illustrations which, in the absence of explanation, are presumed to substantiate the claim that they are indistinguishable as discrete species. It is difficult to refute such a claim against the concomitant one that his years of field experience and careful observation support this claim. Certainly to the uninitiated this could be true whereas any true aficionado would recognise the untruth of the statement. H. chloracantha also cannot possibly be included in this same superspecies H. cymbiformis. It is confounded with H. floribunda and this in turn is central to the recognition of H. retusa as a superspecies. Furthermore the connection has to be followed through geographic distribution and the historic confounding of the identities of H. chloracantha with H. angustifolia and I turn the relationship of that species with H. cymbiformis. Recognising superspecies will simply move the problem of continuity to another area or level of classification.
I would again regard Dr. Vosa’s superspecies H. retusa as a rational option but it very largely skirts the problems contained in and surrounding the suggestion. H. bayeri and H. springbokvlakensis do not enter into the picture. They are “inferior” species in that they do not have problematic continuities. My own independent view is expressed in a letter that I wrote to Steven Hammer…”Your comment ‘illusion of understanding’ also resonates in a comment in the literature where the author, D.A. Levin, quotes from Raven, Berlin and Breedlove’s, “The origin, expansion and demise of plant species” (Oxford University Press, ca 2000)…“our system of names appears to achieve a reality which it does not possess”. Levin himself writes…“our taxonomic system communicates little about the organism being considered, although it appears to communicate a great deal”. This is the conundrum that I get quite lost in. Species have a reality and a meaning to me quite different to what I sense in people I have talked to over such a long period..
Re pygmaea – I need to find a superspecies solution to the SW Cape …”
I presented an outline of a proposal which I now modify as follows:-
H. retusa – to include mutica, turgida.
H.pygmaea – to include dekenahii, argenteomaculosa, splendens, fusca, esterhuyzenii, acuminate and vincentii (it should be noted that these names refer to elements which have a fairly restricted distribution and are highly sought after collector items. I do not personally consider them species although I do not doubt the user pressure for a Latin binomial. For those who have followed my classification it is important to note that I have previously allied them with H. magnifica whereas I am now suggesting that they have their closer origin in H. turgida).
H. mirabilis – to include maraisii,magnifica, emelyae and heidelbergensis.
H. floribunda – to include parksiana (?), chloracantha, floribunda and variegata.
There are other elements viz:-
H. herbacea – to include or cover reticulata, maculata and pubescens
H. rossouwii –
The essential problem is that we have three species in the west viz. mutica, turgida and mirabilis, while in the east we have two viz. turgida, and pygmaea. In the intervening area H. turgida ad H. retusa are continuous with each other. The retusoid element expresses itself in the west as H. mutica whereas in the east it seems to be transferred to the mirabiloid set suggested above while H. turgida remains independent.
There is a problem in this summation in the very complex relationships that also exist between retusoid, and mirabiloid elements elsewhere. These extend to floribundoid and rossouwoid. H. turgida crosses the distribution area of H. mutica to befuddle the proposal that they are elements of a clearly definable H. retusa superspecies. Similarly there is a suggestion that H. mutica has some degree of continuity with H. mirabilissensu stricto. There is a huge problem to segregate H. floribunda which has close ties to the mirabiloids as well as to H. chloracantha, H. variegata, and thus also H. monticola and perhaps eventually H. zantneriana and H. angustifolia.
I do have preliminary ideas for extending the concept of superspecies to the entire subgenus Haworthia but am reluctant to do so while I am aware of the deficiency in terms of field work. More fieldwork is required to examine known problems in the area between Uniondale and Oudtshoorn and also in the mountains south of Prince Albert. These problems suggest that similar situations to those discussed particularly in other chapters, exist in many more areas. Comprehensive as the field work has been, it still does not provide clearer insight. Nevertheless the system of names I now suggest in Chapter 17 does provide a framework of communication that will work if users properly recognized the definitions and premises on which it is based and the reservations that accompany it.
One of the most significant and important user groups is conservation bodies and environmental impact assessors. They require definitive names and lists. No shuffling and reshuffling of names can produce such a list. The reality is that species are intrinsically highly variable and this variability simply has not been exposed as it has been in Haworthia. Hence conservationists have to realize that all pristine and semi-pristine vegetation is valuable and that conservation is not about species, but about habitats with attributes that promote diversity and minority species.
The recognition of supergroups could mean that user groups would have to deal with the problem of using Latin trinomials and higher. This is where the paradigm shift will be the most acute and yet it would be simply solved in common usage by the substitution of the superspecies name where the genus name is presently cited. My conclusion is that the recognition of superspecies is an essential process in synthesising and extending our understanding of the genus but that it would have no merit in terms of the communication value of Latin binomials. Far more would be gained from a proper analysis of the many sources of misunderstanding of taxonomy and an educational process instituted where users and practitioners acquire better insights into biological diversity and how it is recorded.
My closing statement is that scientists seeking a better understanding and classification of Haworthia would do well to try and properly follow what I have written. I am confident that this is the text on which proper and valid hypotheses can be formulated and then tested. I would further add that my field work and observations have been so extensive and so thorough that they could and should be used to test the validity of sophisticated technological inquiry.
Addendum The above manuscript was submitted to Dr. Vosa for comment and I deeply appreciate and value his response printed below. Dr. Vosa, as a pre-immanent geneticist, cannot possibly be expected to be aware of the confusion that exists in the minds of non-scientific user groups and I consider that he has, as he set out to do, indeed made a major contribution to demonstrating that the taxonomic emperor is unclothed…
“Dear Bruce,
Thank you very much for your e-mail message and welcome criticism of my Haworthia paper and also for the rather important attachment (Chapter 33). The purpose of my initial effort has been to make clear the problems facing who is trying to classify Haworthia in a plain understandable and possibly correct way. My paper was also conceived as a kind of provocation aimed chiefly to the “classical” conservative taxonomists. I have never taken into account the opinion of the many “user groups” interested as they seem in “splitting” and in maintaining as many species and varieties as possible for sheer lust of collecting or,, worst, “economic” reasons. I agree very much with all you say about my very faulty and, of course, incomplete “groupings” but I considered them in the light of my research experience relative as it is but in all good faith.
I consider your Chapter 33 as very well written and argued and as an important and basic contribute to that knowledge we all earnestly seek. The Chapter itself will be for me as a sort of main guideline. Above all, I think very appropriate and wise your statement “that there is a preoccupation with nomenclature which displaces the purpose and function of classification”.
I came to choose H. cymbiformis as the typical species for one of my two “superspecies” in the light of my field experience in the Eastern Cape. However, taking into account the evidence, I think that your decision to consider H. cooperi as a superspecies as perfectly valid.
Bruce, I certainly understand fully you disagreement on the ‘candidatures’ as suggested in my two lists which were compiled and meditated using my, alas, fragmentary notes and material in hand. In any case, the reasoned purpose of my paper was to put forward the concept of superspecies as contributing to progress and as a discussion point for constructive criticism and useful argumentations and suggestions as yours.