Foreward
Much of the early work on Haworthia was based on fairly limited observations in nature. These were essentially point collections (often very close to roads) from which seemingly discrete elements were recognised as species. This is akin to a survey made from a plane flying at low altitude assuming that broader patterns are being more easily identified at this illusory distance. The obvious danger of such incomplete observations is that simplistic or incorrect conclusions might be drawn. Indeed, the classification often seems easy from such selective sampling.
When Bruce Bayer started his studies of the genus Haworthia circa 1970, a new era of more methodical collection and thorough recording of the material resulted. Due to the enormity of the task, Bruce’s fieldwork focused on the Worcester/Robertson Karoo, where he built up an excellent understanding of the natural variation of and complex interaction between the recognised species of the area. His resulting model was then extrapolated to lesser-known areas further east. This work culminated in the landmark publications The New Haworthia Handbook in 1982 and Haworthia Revisited in 1999, where a classification framework for understanding the genus was proposed. This framework has been extensively field-tested by a number of people, including myself, and found to be workable.
Bruce remained doubtful about some of his solutions which were based on insufficient collection data, and some new collections which were difficult to place within the framework. During the course of the past few years he has studied further some of the problem areas, the results of which are presented in this update. New insights were gained which are used to propose an improved framework, in the form of a hypothesis, for the major Eastern Cape species complexes. As Bruce points out, this is not the only possible solution as there seem to be several alternatives, each with some reservation, when attempting to explain the observed natural variation. Some problem areas in the Eastern Cape and Little Karoo warrant further detailed investigation. Nevertheless, this work presents a valuable extension of our knowledge of Haworthia.
In his foreword to The New Haworthia Handbook in 1982, Prof. H. B. Rycroft said “the wide range of species and their endless variability keep the professional taxonomist fully occupied and sometimes confused”. This account shows how local continuities exist between adjacent species, only to be contradicted by discontinuities elsewhere between the same species. It is a pleasure for me to introduce you to this fascinating story.
Kobus Venter
Still Bay
December 2001
Foreword by the Publisher This collection of essays deals with some of the more intractable problems in the classification of Haworthia, and demonstrates just how ambiguous types can be. It also demonstrates the scale and nature of the problem in a far clearer way than Bayer’s revision. Bayer himself has said that his revision is still probably premature, and that a lot more needs to be known about Haworthia. This up-to-date account shows where some of the difficulties lie and that it is difficult to determine discontinuities in the species of the genus. Bayer pointed out that there seemed little sense in the emphasis on, and pre-occupation with, types and names, when the elements to which they are being applied are simply misunderstood or even unknown.
The process of the classification of plants is ongoing and it is correct that work should be published before the final solution is reached, as such solutions are probably unattainable. It is therefore appropriate that Haworthia Revisited should be followed by the present series of essays. It is the intention to continue publishing further collections of essays, at present four sets are envisaged. When the first four are complete, they will be bound into “collectors” editions and made available to purchasers of the original books.
Contents 1.Introduction. 2.Haworthia mucronata and its new variety. 3.Haworthia gracilis, H. cymbiformis and H. cooperi in the greater Baviaanskloof area. 4.The case of Haworthia incurvula – an up-date. 5.Haworthia cooperi and H. bolusii var. blackbeardiana. 6.The Haworthias of Kaboega. 7.Summation.
Introduction When I undertook to re-write my New Haworthia Handbook, and present it as an authoritative revision, I did so with some apprehension. However, I did so because in the intervening years my classification had proved personally satisfying. Also I saw little no evidence of the “adequatio” for the task anywhere else, either in circumstance, or willingness, or field experience, to do so. I am an aspirant mystic with none of the transport experience which should have given me the vision I sought. The hermetic and intuitive approach is all I have really been able to bring to my work other than a passion for truthfulness and understanding.
Now, three years after the initial drafting, Haworthia Revisited is published. The initial shock and ‘post-natal’ depression is over and I can look at it more dispassionately. My self-doubts had driven me back into the field and through many bouts of self-examination. There are no doubt many minor mistakes in the book, one of which is the appearance in my preface, of the word “difference” instead of the word “deference”. The latter was intended to convey the sentiment “courteous regard for others”.
After drafting the book, I tried to divorce myself from the manuscript and even from the plants themselves. However, nagging doubts and uncertainties remained with me, and these papers are the product of some intensive field “work”. I use the word in inverted commas for two reasons. Firstly the word “work” in this regard is very loosely used. Wandering around in the field, stumbling on plants here and there and anxiously uprooting them by hands sticky with excitement at “new’ things, does not constitute “work”. Secondly, it is in fact the most rewarding and stimulating way of coming into contact with a most extraordinary and marvelous natural environment. The actual work entails the making of the tangible physical record that places a specimen on the scientific map. I also wrote in my revision (p.12) “A revision.. stands and falls.. on .. the dry and uninteresting herbarium record”. The work therefore also entails deliberating about and examining the herbarium record with a good knowledge of its content. The implications that any new collection may have for that record and the way in which it is arranged, have to be considered and evaluated. That constitutes the “work”, and helps define an important aspect of the “adequatio” required for it to be proven as such.
I have not been able to reduce all the new collections I have made since 1996 to such a record. The plants are in cultivation and are being propagated by seed and offset, and are being recorded in this written record and by photographs. There is still a great deal to be done. Unlike T.L. Salter, who closed his revision of Oxalis with words which suggested that he had had enough, or like G.G. Smith who retired invalidated by the criticism he felt directed towards him, I hope I can continue to “work” on Haworthia.
This book consists of five essays discussing specific problems in the classification and identification of Haworthia. The two subjects are inseparable and unfortunately it is simply a case in which the species cannot be identified unless they are known. My life’s experience suggests to me that this is a sad truth for many things. Thus I rely very heavily on photographs to provide a comprehensive picture description of all the items discussed. The relevant photograph is numbered and this number preceded by a hyphen, usually follows the first reference to a collection number. Each essay has its own lesson and its own conclusion. A final sixth essay attempts to summarise and suggest a solution to a difficult problem.
Acknowledgement I would particularly like to thank the following who have been so supportive and helpful during the exploration and writing of these papers:
W. Schwegmann, J. D. Venter, D. M. Cumming, E. J. van Jaarsveld, P. V. Bruyns, T. Dold, D. Weekes, J. G. Marx, E. Heunis, E. Aslander, D. Clark, L. Loucka and C. Marais.
Farm owners have also been particularly helpful and I am very grateful indeed for access to such properties. In this case I am indebted to Mr and Mrs. N. Sparg, Mr. and Mrs. Killian, Mr. P. Moolman and Mr. J. Truter.
Lastly I am very indebted to Ian and Sandy Ritchie of Kaboega for their hospitality and kindness, and for sharing their own Eden and love of nature so generously with Daphne and myself. I must also record my gratitude and respect to the owners of Kaboega, and note the contribution they are making towards conservation – Mr. and Mrs. Andre Bezuidenhout.
I am grateful to Umdaus Press, and particularly to Mr. J. A. Retief, for their encouragement, support and effort to bring this work to fruition.
Acknowledgement is hereby given to Cape Nature Conservation and to the Eastern and Northern Cape Provincial Nature Conservation Authorities for the permits which were issued for the necessary but limited collecting required for this work. In the Western, Eastern and Northern Cape Provinces of the Republic of South Africa, all species of the genus Haworthia are classified as “protected flora” in terms of the Nature and Environmental Conservation Ordinance, 1974 (Ordinance 19 of 1974), as it is applicable to each of the afore-mentioned Provinces. Material for this book was gathered and collected on terms of the spirit and letter of this Ordinance. Nothing in the book should be construed as an encouragement to acquire these plants from their natural habitat or to suggest they are better located anywhere but in that situation. ♦
M B Bayer, 16 Hope Str., 8001 Cape Town.
View from the upper slopes of the Zuurberg at Wilgerfontein, with Darlington Dam just visible in the distance.
I will be disappointed if anyone had concluded I had any fixed ideas on the classification of these three species and their relationship. It has a problem which has long been on my mind. What happened recently (Nov.1998) is that I was offered the use of a time-share apartment at Jeffrey’s Bay, near the mouth of the Gamtoos River. I used this opportunity to spend six days in the field testing my hypothesis concerning the species Haworthia cymbiformis, Haworthia cooperi and Haworthia gracilis, and this is what I would like to record. Subsequent to that trip (Mar.1999) I planned and executed an excursion through the Baviaanskloof to Grahamstown and Stutterheim in March 1999, and repeated the exploration in Sept. and Oct, 1999.
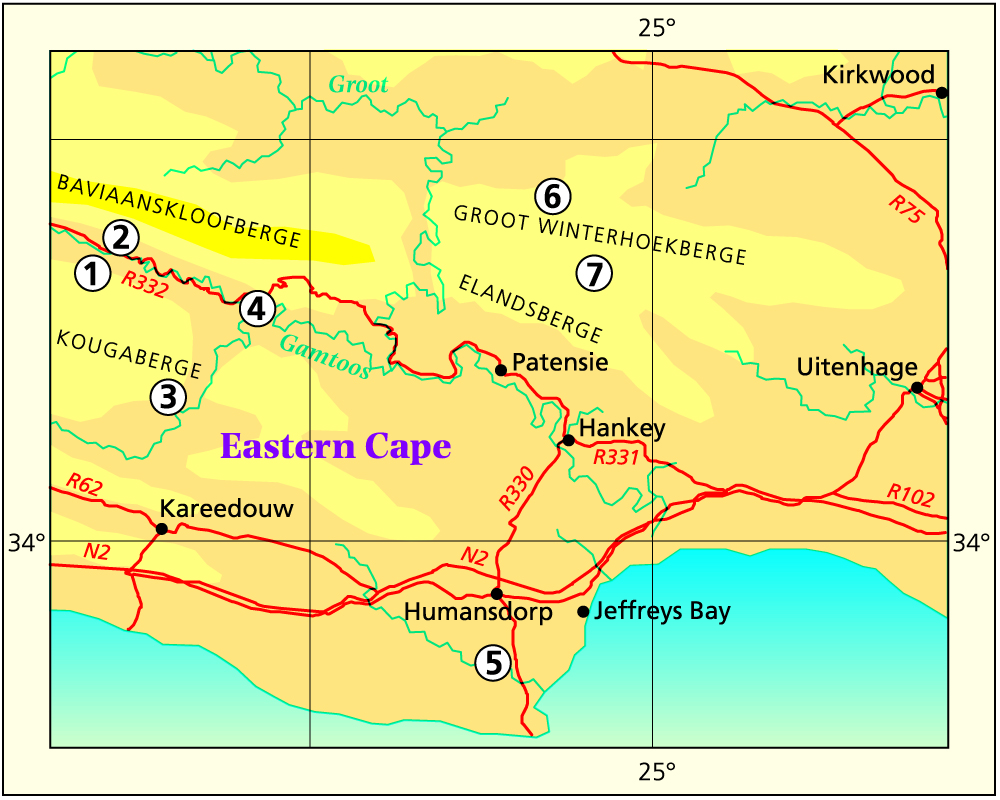
Baviaanskloof and environs: 1-Engelandkop, 2-Rooikloof, 3-Moordenaarskloof, 4-Rooihoek, 5-Ripon, 6-Ouplaas, 7-Houtkloof
First I must point out again that there is a problem with plant names in that they do not necessarily reflect a set of objects which can be seen and easily recognised. The naming and typification process is partly to blame. Typification is simply the process of ensuring that a name is based on some substantial recognisable item, preferably a specimen rather than an illustration or photograph. (A specimen is preferred because it lends itself to some other analysis e.g. a tissue sample). The type is used to secure and permanently attach a name to for reference purposes. A type specimen and hence a name need not be, and often is not, typical of the taxon which the specimen represents. This does create difficulties particularly when varieties are at issue. In Haworthia there are an inordinate number of populations of uncertain pedigree. This is not because of some obscure evolutionary process, and nor is it just because there is an absence of characters by which to characterise and circumscribe each population. It is what we should expect. Everything is in a state of change and Haworthia is simply following the same unsteady path. I have shown that in Oxalis, in which there are an embarrassment of riches where characters are concerned, that these are of surprising little help in circumscribing the species. I was interested to read this sentence in a book about chameleons, “Without collecting data many chameleons are difficult to identify.” One would expect chameleons to be less difficult to identify than plants. Darwin stated that geographic distribution is the doorway to understanding species, and this is all we have in Haworthia which I can see to make sense.
We simply use ordinary visual observation and sensibility to make judgements about similarities. The less objects we have to examine, the easier it is to assess similarities among them. The problems that then arise are no different from those that develop even with complicated and convoluted discussions and explanations that arise from sophisticated techniques and analysis of characters. In my experience it is very often the direct visual judgement which is used to test the worth of technology rather than the converse. These comments are made because readers seem to think that plant names are as centrally defined as their own unexpressed concept they have of species being groups of things with well-defined limits and readily compartmentalised. Thus they imagine that there is a solution and it is just a question of how it is found. These comments are made because several times in the literature, wild statements have been made about the use of some character or technique or other throwing great light in the darkness. It just does not happen that easily and simply. The more sophisticated the technique, the smaller the sample and the less ‘peer’ review and replication. In statistics it is also pointed out that you do not undertake sophisticated statistical analyses when the figures already show the obvious. Yet how many times do analysts not persevere with test after test until a result does appear significant – itself a matter of probability. Technique may become and end in itself and the purpose for which it is devised lost to view.
There is a nomenclatural code for plant names which regulates about everything except what the binomial stands for. What should be pointed out with some vigour, is that there is a tendency to think that species can be described from single specimens in the paradigm (this is a most useful word because it suggests a pattern, a habit and a model of a time period) of Salm Dyck, Von Poellnitz, G.G. Smith and company. The single plant (and its description) and specimen that is used to typify a name, does not constitute a circumscription of the species. These people described specimens – NOT species. Their work was thus in some measure comparable with stamp collecting. It has little to do with present time description, organisation and understanding of the living variable and changing systems which species are. The different elements in Haworthia named as ‘species’ have been determined by early writers on the basis of vegetative characters such as colour, rosette shape, leaf number and shape, translucence and spination. Nothing much has changed except the scope and extent of the problem. A difficulty with characters is stated in Vavilov’s Law of Homology which states that a set of characters in a taxon will also be present in related taxa. Thus if one finds awned and obtuse rounded leaf tips in the genus, it is possible and indeed probable, that this character set will also occur in lesser ranks. If a set of characters is present in one species, they will possibly and probably occur in related species. This article is to demonstrate the difficulty of classifying plants on this simple basis and in an era when so much more material is there to classify.
It is necessary to discard the name translucens as I have used it in my earlier books. Originally the name really is associated with what is obviously H. herbacea, and I only used it to follow a tradition of misuse. The misfortune of this is that the name gracilis then occupies the nomenclatural hierarchy, but not the same geographic position in my understanding of the species. The consequence is that an outlying variant (Hellspoort, Grahamstown) will become the species name (i.e. gracilis) and a name has to be unearthed for the core of the species in the Gamtoos valley viz. H. gracilis var. isabellae. This in my new book replaces H. translucens var. translucens. Similarly H. cooperi var. cooperi is not central to the plot as is the var. pilifera, and I unearthed the latter name in my 1999 revision for this reason, as also the name gordoniana. I opted in 1985 to use the rank ‘subspecies’ under H. translucens for reasons which should be obvious – a weaker geographical relationship of the known variants of the species associated with a broader distributional phenomenon.
This following discussion is done in terms of Darwin’s statement about the importance of geographic distribution, and the evidence I have in my mind for a wide range of species. The essential problem with the species considered in this article is that we have a set of plant populations from Grahamstown, replicated to a degree by a set of populations in the Humansdorp area. In neither area is the relationship in the set well understood, and this does not imply that they can be so understood either. I set out to deal only with recent limited observations in the Humansdorp area but like a stone thrown into a pond, the ripples soon spread.
The link between the Grahamstown and Humansdorp areas is a curious one. It can be seen in the vegetation types of Acocks, which is a broad view of species associations. Valley Bushveld occurs discontinuously from Natal through to Swellendam. Alexandra Thickets are coastal, east of Port Elizabeth. Knysna forest, west of Humansdorp. Dry fynbos dominates the immediate Humansdorp area. Karoid vegetation dominates north of the Groot Winterhoek Mountains. It is at species level that really interesting facts emerge that impact on our analysis of Haworthia. What happens in the genera Aloe, Gasteria, Astroloba etc.? I leave this as a largely unanswered question because unless the reader can respond, he or she is unlikely to appreciate that it may have implications for understanding Haworthia. Astroloba does not occur in the Humansdorp area. There is instead the curious Haworthiapungens. The Baviaanskloof is home to several endemic Gasterias as well as to Aloe pictifolia. Haworthiaviscosa is ubiquitous. H. nigra with an otherwise fairly similar distribution is not. The distribution of the genus Encephalartos (cycads) is particularly interesting because of its known antiquity and the similarity of present day species to evidence in the fossil record. Aloe must also have an ancient history as evidenced by presence of species of this genus on Madagascar – a continental drift pre-history. It would in fact be instructive for would-be authors to read what R.A. Dyer wrote about the classification of cycads, before trotting out any banal aphorism about ‘active evolution’. There is an interesting break (interval) between the cycad species, east of the Sundays and Bushmans River valleys which separate Grahamstown and Humansdorp. This break can be observed in the distributions of other genera and species and evaluated both for compliance and non-compliance. Haworthia has both conditions e.g. cymbiformis and pilifera happily cross the interval. Angustifolia and coarctata only just do so. Marumiana, and nigra do so in the inland areas. Attenuata does so. Viscosa is almost ubiquitous. Fasciata does not and neither do sordida, longiana nor xyphiophylla. No species jump the Knysna forest interval and the genus as a whole jumps the Transkei interval northward only via limifolia and koelmanniorum (+mcmurtryi). These east-west breaks are interesting because they thus indicate speciation pressures. They may be helpful in deducing possible or probable relationships in a two-directional way i.e. distribution and variation in one set of plant variants provides a tool for the examination of other sets. In the case of H. gracilis, one can argue simply that it complies with the interval, in which case the var. tenera east of Grahamstown is discrete from the Uitenhage, Baviaanskloof look-alikes. Alternatively one can hypothesise non-compliance and that continuity will be found between Grahamstown and Uitenhage. Hellspoort (the site of the var. gracilis) is somewhat supportive of this view, but their is still a wide gap. Perhaps this is only in the known distribution record.
Known collections and observations were the basis of my 1985 classification hypothesis. Many of these were either dry herbarium specimens perhaps supported by photographs of single plants grown in cultivation. This hypothesis for the species in question here was arrived at from specimens from the immediate Grahamstown area and from the lower and eastern Baviaanskloof (Humansdorp) area. For the 1999 revision I have broadened the range of varieties, and relevant to this discussion are:
Haworthia cooperi
var. cooperi
var. leightonii
var. pilifera
var. gordoniana
and included Scott’s two new species as varieties:-
H. joeyii (= var. dielsiana)
H. venusta (var. venusta)
Haworthia gracilis
var. gracilis
var. tenera
var. isabellae
Haworthia cymbiformis
var. cymbiformis
var. incurvula
var. transiens
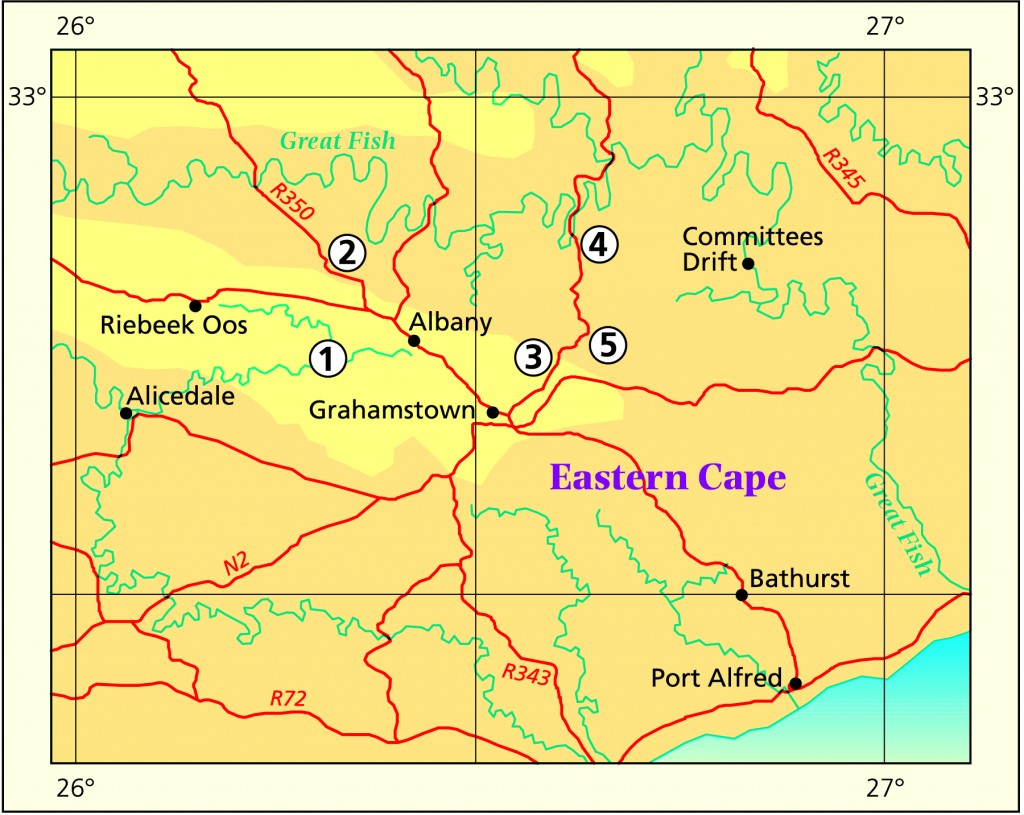
In this discussion I will largely avoid the issue of both H. bolusii var. blackbeardiana, H. cooperi var. cooperi and H. decipiens although they are closely involved. I will deal primarily with the Humansdorp (Hankey-Patensie); and to lesser degree with the immediate Grahamstown (Plutosvale) area.
I will use the term look-alikes because it is frequently used in the fynbos vegetation to refer to plant species from different genera, which are superficially so similar that close examination and full flowering material is often needed to establish their real identities. In Haworthia this detail is simply not available and thus there are specimens from different species which are geographically and thus probably genetically aeons apart, but which are visually identical.
Grahamstown (Albany) H. cymbiformis – is well represented and discrete. Yellow-green plants with spreading broader, flatter, ovate leaves. Clump forming plants on steep rocky faces. The var. incurvula – is known from many collections but only one locality (Plutosvale). It has look-alikes in the Patensie area particularly. Its relationship as a variety of cymbiformis really follows G.G. Smith who made the combination on the basis of a continuity which he claims to have seen. I recall (!) that its flowers are more reminiscent of tenera and this article may expose this as a possibility. Pale semi-opaque green, small clump-forming, with ovate, obtuse incurved leaves.
H. cooperi – is well represented by the var. pilifera. Blue-green plants tending to purplish, with incurved thicker, shorter ovate leaves, often spined. Solitary plants withdrawn into the soil. (The var. cooperi is not discussed in this appraisal although the concept presents a substantial problem on which I am preparing a manuscript).
H. gracilis – is known only from Hellspoort north-east of Grahamstown, with a second population of larger more cooperi-like plants from nearer Plutosvale (MBB6603-99, Glen Craig – a Gerhard Marx collection). Gracilis does have look-alikes in the Kirkwood area (Paardepoort, and a greater Humansdorp area). Paler grey-green plants tending to clump, with more leaves. The leaves are incurved narrow attentuate, and with or without spines. My opinion, based on my most recent collection from Hellspoort is that the var. tenera is actually the element concerned and is thus a superfluous name for gracilis. This var. tenera – is very well known from several populations north-east and east of Grahamstown and these include a glabrous population. It has look-alikes in the Uitenhage and Hankey areas of Humansdorp. Grey-green, small solitary or clump-forming, with many narrow, spined incurved leaves.
(Note:- the original description of gracilis seems to be of a moderately spined species, and von Poellnitz concept was apparently drawn from several disparate collections. The conception of the species from an illustration in Desert Plant Life, and indeed my own collection from Hellspoort in early 1970s, is of a moderately spineless plant. The recent collection of mine has forms which are relatively spineless as opposed to forms which are indeed similar to the smaller, densely spined, tenera. Thus we again have the problem where a name is neither central to the geographic element nor typical. The name tenera could be dispensed with and replaced with var. gracilis – this is evident from my discussion).
Humansdorp H. cymbiformis – represented by the typical variety in the small Baakens River valley and discrete. It may occur north-east of the Baakens River in the Rocklands area of Port Elizabeth. There is also the population at Hell’s Gate north-west of Uitenhage which I am regarding now as the var. transiens (It needs to be re-examined – hastening to add ‘by someone competent to do so’). This var. transiens – is only recently known to be very well represented in the upper Longkloof and in the Baviaanskloof. Its relationship with cymbiformis may not be as close as varietal rank, and the linking population at Hellsgate east of Uitenhage may be misunderstood. Leaves are incurved, pale-green and less opaque than in cymbiformis. Some of the variants resemble plants of the var. incurvula from Plutosvale.
H. cooperi as the var. pilifera – well represented and largely discrete. However, the typical variety with the necrosing-end area seems to have a western limit just outside Port Elizabeth and also near Uitenhage (the furthest west being at Perseverance, JDV90/40). It is then replaced by a variant which is often smaller, having the same blue-green colour darkening to purplish, but the leaf tips remain slender acuminate even in light conditions. This is what I have regarded as the var. gordoniana which in its typical form is not well represented.
H. gracilis as the var. isabellae – is well represented but apparently variable. The leaves tend to be spreading, pale greyish-green and spined. Some plants resemble gracilis from either Hellspoort (less-spined) or from Plutosvale (densely spined – tenera).
General It is known to me that in the area from Kirkwood, westward to the Little Karoo (north of the Baviaanskloof), there is an interaction between gracilis and decipiens. H. cooperi (not designating any variety) also occurs in that area. There is a problem in the Addo area, where another element (aristata – a separate weak hypothesis) may intrude. It has also been suggested, and this has always been in my own mind, that unicolor (mucronata) from the western Little Karoo may be continuous with either cymbiformis, bolusii or cooperi. It is continuous with arachnoidea, lockwoodii and with decipiens, and I am treating it as a separate issue because there are also other reasons for its exclusion here. The discussion could descend to the point where one says simply that there are not many species in the sub-genus Haworthia.
Results
Grahamstown I first looked at Hellspoort early in 1970 to collect and form an impression of H. gracilis. This was of a largish plant with leaves exceeding 3cm long, fairly spineless and weakly clustering. I also saw tenera in Plutosvale and at Hunts Drift. The former, small, densely spined and strongly clustering. At a later occasion I collected tenera again in Plutosvale as the conventional spiny element, and close by as a glabrous element of the same size. On that occasion I neither looked for nor saw incurvula or cymbiformis in the area. I revisited Hellspoort with J.D. Venter and G.D. Marx in 1996 and Venter made a re-collection again in 1998 (JDV89/42-117). My 1996 collection (MBB6614-118) reminded me forcefully of Plutosvale tenera, the plants were smaller than my previous collection and also spinier. Venter’s 1998 collection (all slightly different points within Hellspoort) was of the conventional gracilis that I visualise from the Von Poellnitz illustration, and from my early collection. These were quite large plants. During my visit of March 1999, I observed and recorded systematically from the bottom of Plutosvale, to the top (Roff’s Rock). On the north side of the kloof from the Cotswold farmhouse to the middle of the kloof which is Plutosvale, there was tenera both large clusters and relatively solitary plants. I could not locate or identify the site where I had collected the glabrous form before. The area has been degraded by the grazing of goats and it is possible the plants are now gone. From there we explored the south side of Plutosvale as the road climbed out of the kloof. Roffs Rock is very accessible and this no doubt accounts for the large number of herbarium records for incurvula. We found if from virtually the top of Plutosvale, to the lower and eastern point across from where we had observed tenera at our western-most search point. It is possible that the collection G.D. Marx in JDV93/73-93 was from further to the south-west than our starting point at Roffs Rock, at the upper point of Plutosvale. Nevertheless we found the same very pointed leaved plants of that collection, tending to be clump-forming. As we moved eastward, so the plants tended have leaves which were less pointed and more incurved. The colour was yellowish-green rather than the greyer or bluer-green of the Baviaanskloof incurvula look-alikes. The leaves seemed to lack the more swollen upper and end margins that may characterise cymbiformis.
My impression from this repeat visit to the Grahamstown area, is that the elements gracilis and tenera may be better considered as the same taxon. This may lay the basis for a better understanding, or be more compatible, with the range of look-alikes in the west around Humansdorp and Uitenhage.
The incurvula problem is now largely resolved and although it may still be necessary to find other links between incurvula and cymbiformis which Smith claimed and which is not supported by herbarium evidence. To the contrary, the collections of the gracilis-gordoniana plants from Glen Craig and Roffs Rock (viz. -99, -89, -93)), as well as the interplay of species in question in the Baviaanskloof, suggest that incurvula may equally be interpreted as a variant of H. gracilis or even H. cooperi. There is a further possibility based on new observations in respect of H. bolusii var. blackbeardiana. It may be possible to restructure the classification to relate the var. bolusii with H. semiviva; and to consider the element blackbeardiana as a variety of cooperi, (as nomenclaturally correct but geographically weak) as the species, with pilifera and gordoniana as principle variants. Nomenclatural considerations in ‘the strict terms of the code’, might confuse the issue, but I see no reason why reasoned and logical argument could not be used to dismiss names which are confusing because of their geographical connotation.
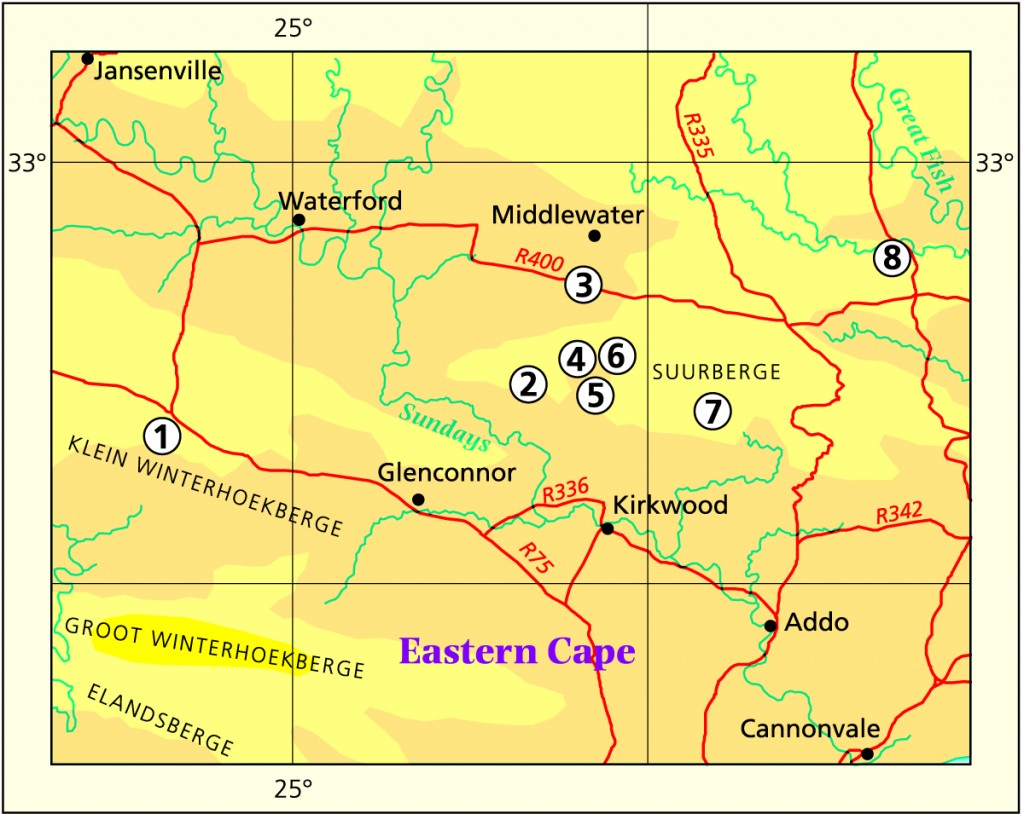
Humansdorp I looked at populations at Jeffrey’s Bay, Zuurbron between Humansdorp and Hankey, two at Draaihoek to the south-west of Patensie, three near Andrieskraal, one further west in the Groot River Poort, and three still further west in the Baviaanskloof, one at Patensie, one at Houtkloof in the Elandriver valley, one at Hankey, one north of Hankey, two south-west of Hankey, and one at the Gamtoos Bridge. Some of these populations were known to me before, and many of them are already represented in the Herbarium. I will also refer to other collections (mostly multiple plants) in my discussion which I have either not seen in the field at all, or in the more distant past. These are collections by credible field botanists and naturalists such as P.V. Bruyns, G.D. Marx, E.J. van Jaarsveld, T. Dold and D. Clarke. This thus reviews some 60 populations, but does exclude some significant ones in the Kleinwinterhoek mountains, where other considerations apply.
MBB6792-20 Jeffrey’s Bay. Fynbos. Very short vegetation on level sandstone outcrop. Early bud stage. Small single plants to 20mm diam. Leaves incurved at tips, no marginal spines and bluish-green, darkening to purplish hue (3) – gordoniana-picturata.
MBB6933-21 E. Humansdorp. Grassy fynbos. Rocky sandstone knoll. Small single plants, bluish-green with coarser spination than MBB6793 (2) – gordoniana.
MBB6553-22 and MBB6793-22 Zuurbron. Valley Bushveld. Scrub vegetation on level alluvial stream bank with loose sandstone rocks. Flowering and seeding. Small single plants to 30mm diam. Leaves thicker, leaves incurved at tips, many small closely spaced marginal spines, bluish- green (2) – gordoniana (possibly close to type locality).
MBB6784-23 SW Patensie. Valley bushveld, west sloping conglomerate. Seeding. Small single plants to 30mm diam. Leaves incurved at tips, forms with and without marginal spines, and blue-green (1). Smaller plants of this order were collected by Tony Dold at Drinkwaterskloof in the Baviaanskloof near Geelhoutboskloof – gordoniana.
MBB6786-24 SW Patensie. Valley bushveld, steeper south facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaves incurved at tips, mostly plants with marginal spines, blue-green (2) – gordoniana-isabellae.
MBB6827-27 Rooihoek. East of Geelhoutboskloof. Steep east facing slope. Plants single, greenish, very small – picturata-transiens.
EvJ14680-28 Vetmaakvlakte, S Rooihoek – isabellae (as for a clone of -41, resembling H. aristata).
MBB6789-29 Groot River Poort (Komdomo). Valley bushveld, very steep west-facing shale cliff. Flowering. Small clustering plants to 30mm diam. Leaves with spreading tips, slightly obtuse, no marginal spines, pale-green (5) – transiens-picturata.
MBB6830-30 E Komdomo. Riverine margins. Large clustering plants on steep soil-covered rock- gracilis.
J.N.Reddi in JDV93/86-31 2km NW Andrieskraal, is a collection which may be the same as -30 – gracilis.
EvJ15927-32.1 N. Groot River Poort. Steep south-facing cliffs. Clustering or single. Fairly similar to -29 – transiens-isabellae, the clone 32.2 to decipiens var. minor?
MBB6790-33 Andrieskraal. Valley bushveld, very steep south-facing conglomerate. Flowering and seeding. Small clustering plants to 30mm diam. Leaf tips moderately incurved, slightly obtuse, no marginal spines, pale yellow green (4+Y) – picturata.
MBB6791-34 Andrieskraal. Valley bushveld, steep east-facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaf tips incurved, slightly obtuse, no marginal spines. Pale yellow green (4+y) – picturata.
MBB6930-35.1 E. Andrieskraal, Nuwelande. Steep south facing conglomerate. Small plants down to 16mm diam. As above (4+y). – picturata. JDV90/55-35.2 from nearby westward, is also small plant with very pronounced windows.
MBB6928-36 Nuwelande. Upper steep conglomerate cliff. Plants nearly spineless and the leaves more slender and terete. Greenish (4). These are similar to JDV94/115-37 from the Paul Sauer Dam wall and also from the site for Gasteria glomerata where it was collected by E.van Jaarsveld (EvJ11076 in JDV90/118-38.1&2). A plant from that collection (-38.3) also resembles the Ripon plants (MBB6932-39) which have broader more deltoid, and more spinose leaves – isabellae.
J.G. Marx298 in JDV94/30-40, also Paul Sauer Dam, include both spined and spineless plants which also appear to be (4) – isabellae.
MBB6799-41.1 also (-9&-10) NW Patensie. Valley bushveld. South facing conglomerate slope. Seeding. More robust single plants to 45mm diam. Leaf tips incurved, with and without marginal spines. Pale green and blue-green (1-3) – gracilis-isabellae. One clone in this quite variable collection, strongly resembled a large specimen of H. aristata (-125,-142,-143).
MBB6801-42 SW Hankey. Early flowering. Valley bushveld. Vertical conglomerate. Small clustering plants to 20mm diam. Leaf tips incurved, with marginal spines. Pale blue-green (2-3) – isabellae.
MBB6802-43 SW Hankey. Valley bushveld. Steep east facing conglomerate. Early flowering. Plants mostly single to 30mm diam. Leaf tips tending to spread, with marginal spines. Pale blue-green (2-3) – isabellae.
PVB7128-44 Holriver, far S Patensie. Steep north-facing cliffs. Similar to above – isabellae.
D.Clark1050-45 Kouenek, Geelhoutboskloof. No detail, similar to above. – isabellae.
MBB6773-46 N Kareedouw. Fynbos. Conglomerate. South facing. Flowering late Dec. Plants single small, leaves incurved, very spiny and spines on leaf surfaces too. Blue-green (2) – isabellae.
MBB6771-47 Moordenaarskloof (N. Kareedouw). Steep south facing slope. Plants in small clusters or single. Relatively spineless. Leaves incurving. Green (4) – picturata.
MBB6805-48 NE Hankey. Valley bushveld. Gradual west facing slope. Seeding. Plants single to 50mm diam., leaf tips incurved, with and without marginal spines. Blue-green and pale-green and (1-4) – gracilis-isabellae.
MBB6804-49 N Hankey. Valley bushveld. Dense vegetation on alluvial stream bank. Seeding. Plants single to 50mm diam., leaf tips incurved, without marginal spines. Pale-green and blue-green (1-4) – gracilis-isabellae.
JDV97/3-50 E Hankey. Valley Bushveld. Plants similar to above – (1-2) – gracilis-isabellae.
MBB6808-51.1 Gamtoos Bridge. Valley bushveld. Steep west facing conglomerate. Seeding. Clustering plants to 50mm diam. Leaf tips spreading, with marginal spines. Blue-green (2) – isabellae. There are very similar plants at Longmore, east of Loerie (MBB6555-51.2).
MBB6798-52 Houtkloof (Upper Elandsriver valley). Fynbos. Rocky valley in sandstones. Neither flower nor seed (flowered in cultivation in Jan.). Plants single to 35mm diam. Leaf tips incurved, very spiny. Blue-green (2) – isabellae.
C. Marais in JDV96/95-53 Forest Glade and JDV92/136-54 Oaklands, both Elandsriver, are similar but more coarsely spined and paler green (4) – isabellae.
MBB1404a in JDV86/13-55 is an old collection of a tenera-like plant from the lower Elandriver valley (Groendal Dam) – cf. tenera.
D.M. Cumming6831 is from nearby and is more typically isabellae-like).
PVB7040 in JDV97/8-57 and PVB7944 in JDV97/7-58 Ouplaas (E. Cockscomb) are very similar to the above from the Elandsriver, but the plants are more robust, with much wider and more obtuse leaves. Particularly interesting because similar plants also occur in the Kleinwinterhoek eg JDV93/41-59 N Campherpoort, JDV87/180-60 S Campherpoort, JDV91/136-61 N Wolvefontein, JDV92/140-62.1 and JDV94/45-62.2 both Wolven (all decipiens var. minor). There is also the collection from the northern Groot River Poort (EvJ15927 – 32.2) linking these, and perhaps also the Cockscomb and Elandsriver plants (ie. isabellae) with transiens.
JDV97/6-63 W Braamrivier. Dry Fynbos. Steep W. facing cliff. Clump-forming, short obtuse incurving leaves. Pale green (5) – transiens. EvJ15342-64 Dieprivier, NE Kareedouw; EvJ17548-65 Horee, Saptou, SE Uniondale; PVB7077 in JDV97/5-66.1 Oshoek, E of Uniondale; MBB6729-66.2 S Uniondale, and Reddi in JDV93/54-67 Kabeljouwsriver, near Jeffrey’s Bay (a doubtful collection), are also this element – transiens.
The latter is similar to -29 Groot River Poort (transiens-picturata), ie. small clustering plants to 30mm diam. Leaves with spreading tips, obtuse, no or very few marginal spines, pale-green. I could not locate this population and it would prove that gracilis, transiens and cooperi co-exist. E. Aslander1247-68 made a collection from the place reported by Reddi, and these plants are unquestionably gordoniana-like.
PVB7093 in JDV97/1-69 Skrikrivier, north of -63 is similar to that collection, but leaves more elongate acuminate and some lightly spined. Flowered late Jan. Very reminiscent of gracilis var. gracilis and particularly a brighter green variant from Paardepoort (MBB6600-70, var. viridis in the new revision) in the Kleinwinterhoek mountains to the north-east. PVB5402 in JDV97/20-71.1 Palmietrivier and MBB6589-71.2 Dorschfontein, both E. Steytlerville, are similar to this latter collection but better related to decipiens var. minor. A problem arises here in trying to limit the scope of the paper because there are similar collections from north of Glenconnor (JDV91/17-72.1), and south of Lake Mentz (JDV91/116-72.2, which grows with gordoniana JDV91/115-73).
MBB6810-74 (also -11&-12) Joubertina. Fynbos. South facing, rocky sandstone slope. Neither flower nor seed. Plants single to 50mm diam. Leaf tips incurved, without marginal spines. Blue-green (1) – gordoniana.
MBB6811 & JDV90/80-75 (also -13) Uniondale Pass. Renosterveld. Rocky defile, steep sandstone. Early bud stage. Plants single to 25mm diam. Leaf tips incurving, with and without marginal spines. Blue-green (1) – gordoniana. PVB7079-76 Saptou, (Upper Longkloof), PVB7062-77 Redcliffe (NE Willowmore), JDV91/80-78 Engelandsekloof (Baviaanskloof), JDV94/95-79 Nuwekloof (W. Baviaanskloof), are all in this class. A collection G. Marx194 in JDV91/81-80 from Apieskloof (Baviaanskloof) is unusually pale green but otherwise also seems to compare with these gordoniana-like plants).
In completing this review of collections I would like to mention five which seem to touch on this issue, but which are even more relevant to the H. cooperi var. cooperi and H. bolusii var. blackbeardiana issue. These are G. Marx in JDV91/14-145 De Plaat (NW Kirkwood), PVB5002 in JDV92/33-147 Kaboega Gorge (Suurberg), JDV96/89-116 Gladhurst (S Adelaide), J.G. Marx in JDV96/4-98 and MBB6603-99 NE Grahamstown (Glen Craig), and JDV93/73-93 upper Plutosvale. These are all plants with elongate acuminate leaves about as wide as thick, and almost or completely without spines. They could be confused with H. cooperi var. leightonii, or with clones of H. bolusii var. blackbeardiana – and of course with H. gracilis var. gracilis.
Discussion In trying to circumscribe each collection I have been aware of my deficient descriptive skills. Can one rationalise such similar things one observes in writing in a way and which can lead others to identifications? I have elsewhere pointed out major weaknesses in the capacity of persons to compare what is written with what is seen, and also to compare plants and illustrations. Apart from the problem of variation in the populations, there is a problem of different habitat conditions (light, soil, temperature and moisture) which will cause phenotypic differences. It should be possible to define these if the plants could all be grown under the same conditions. However, local variation and sample size also becomes a problem. Colour must be critical. There are four main classes in this set of species under discussion – the opaque yellow-green of cymbiformis, the darker blue-green of cooperi which can develop reddish veins and a purplish colour under stress, the mid pale (or light) grey-green of translucens and the subdued pale green of transiens. Under stress the latter becomes very pale. In the descriptions above I have categorised colour on a scale of 1-5 from blue-green through to pale green, and added Y to indicate a more opaque yellow colour. In my book I recognise a variety (i.e. MBB6791-34 picturata from Andrieskraal) in which the transition from opacity to translucence is abrupt. This collecting trip does not strongly support such a geographic entity although their may be populations in that area which do. Certainly in the var. transiens, some plants are also very conspicuously patterned. Flowering time in early summer for all these collections appears to be very much the same. Curiously the Jeffrey’s Bay collection was still at bud stage when most of the other collections were seeding already. When I visited this population six weeks earlier in the previous year, the plants were already in flower.
The off-setting, clustering character is also worth commenting on. As in the case of H. turgida and H. retusa (remembering also an expressed contention of mine that the latter can be regarded as a variant of the former), clustering is usually associated with very steep rocky situations as opposed to plants on level sites being solitary. In this particular study the slopes varied from level through moderate to vertical. Clustering was most pronounced on vertical sites and at Andrieskraal where vertical and sloping sites were nearly adjacent, the plants were clustering on the vertical and solitary on the sloping. Clustering may be accompanied by the capacity of the plant stems to elongate. It is significant that populations around Hankey and Patensie which seemed to be var. gracilis, were invariably on moderate sloping or level ground (NOTE. I have previously always identified these collections as gordoniana), and using the name gracilis now is derived from the product of the papers in the compendium. My later papers suggest that transferring these elements to H. cooperi is worth consideration).
The above collections confirm my observation that Haworthia in the Baviaanskloof are very difficult to classify and circumscribe. It does seem on the basis of collections by Bruyns, Van Jaarsveld, and Venter that transiens is a very substantial element to the west, which may not have a strong connection with cymbiformis. Gordoniana is apparently a much stronger element than I had previously supposed and I am comfortable with the decision to now recognise it as a valid variety of cooperi. The above collections in the Longkloof, and some other recorded collections, seem to show convincingly that it effectively co-occurs with transiens. In the area just west of Patensie, three collections described above seem to indicate that gordoniana intergrades directly with transiens and this transition gives rise to plants which resemble incurvula and which I identify as H. gracilis var. picturata. Similarly south-west of Patensie gordoniana (-24) seems to alter to isabellae. South west of Hankey this latter variety (-42) resembles tenera. The resemblance to tenera is also apparent in the Groendal area of Uitenhage. The Elandsriver collection (-52), and a similar one by Clifton Marais in the same valley (-53), appear to be intermediate in appearance between tenera and isabellae. Isabellae as a separate entity is also represented by the Gamtoos Bridge collection (-51), and an old collection by W.R. Branch from the Krom River estuary at Ripon (-39).
A major difficulty is the Jeffrey’s Bay population (-20). These small solitary plants seem, by virtue of their growth habit and the near co-occurrence with isabellae (perhaps transiens too, if Reddi’s collection -31, was confirmed), to be the local equivalent of cooperi var. pilifera. However, if they had been observed at Grahamstown, they would be taken to be gracilis. In cultivation they have come to bear a stronger resemblance to the -47 which is closer to picturata than to gordoniana. The Zuurbron population (-22) passes for the equivalent of pilifera with some difficulty, and this does transpose to isabellae via Draaihoek, SW of Patensie (-24). The gracilis-like plants around Hankey (-48,-49,-50) and Patensie (-41) too, often present an isabellae facies. Thus we are faced with the possibility of a strange inversion. The upper Longkloof has cooperi as the var. gordoniana, but in the lower Longkloof similar plants may be derived from gracilis. A similar problem is presented by the similarity of gordoniana-like plants to H. bolusii var. blackbeardiana at several different places in the wider Eastern Cape.
The collection -46 is particularly interesting (I am indebted to Ernst van Jaarsveld for its discovery). This is a very small isabellae which has the same colour and (spininess) hairiness of the leaf surfaces as has Scott’s venusta. This may be very significant in providing supporting for an hypothesis that prior to inundation of the continental shelf (12-18000yrs ago) there was less of a vegetation interval east and west of the Sundays and Bushmans Rivers. I have suggested that this is reflected in the relationship and distribution of H. coarctata and H. reinwardtii, and will suggest elsewhere that H. fasciata may also be evidence of such a situation. However, this is very speculative. The surface spination is also evident in some collections from north and south-west of Jansenville (-129,-130,-131,-132 and -133 which I have identified with either decipiens vars. minor or pringlei), although the spination is sparse.
Conclusions These specific observations suggest that my classification hypothesis (the classification as presented in my two handbooks and in my recent revision) is a sound communication medium for the two areas in question, without implying thereby that it cannot be improved. What is problematic is that in the greater Baviaanskloof area, the three elements never seem to occur together. I repeat what I have written in respect of H. cooperi and H. bolusii var. blackbeardiana, and in my handbooks and revision. It is also stated indirectly in the preamble to this paper. “The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain good will.” In this particular article, the key question would have been:-
“Do three elements we can identify as separate living systems representing the species cooperi, cymbiformis and gracilis grow in the same close geographic proximity?”
Evidence available to me and in the herbarium record, is that they do not. There are a few known instances where two of these species are in such proximity – but not all three. What is curious is that it is rare for any two species of the subgenus Haworthia to be in immediate proximity to the extent that they share any specific habitat.
Observations seem to confirm my opinion that there is a connection between the three species in their occurrence east and west of the Sundays and Bushmans Rivers. What does concern me is the relation between cymbiformis and transiens, and the implications these observations have for incurvula, gracilis and cymbiformis around Grahamstown. The set of plants around Grahamstown favours cooperi, cymbiformis and tenera, while the mirror set of the Humansdorp area, favours gordoniana, transiens and isabellae. The relationship of incurvula needs to be explored. In the eventual integration of all these collections, sight should not be lost of the complex situation around Fort Beaufort, where the same set of species and variants seems to be present. Nor should it be forgotten that in the Kleinwinterhoek mountains H. decipiens becomes involved. The situation north of the Groot River is complex and there are substantial records from the Perdepoort and from Willowmore and Steytlerville which indicate a close interaction between decipiens and gracilis as well as gordoniana.
There are a good many other collections from the upper Baviaanskloof, but mostly these are represented only by herbarium records and I would like to see this material in its living state.
A final thrust of this article is to emphasise that Haworthia is not going to be understood by botanists or enthusiasts sitting on some other continent. This is particularly if they depend on the convolutions of the Code of Nomenclature, literature and limited collections for their information and the opinions they feel compelled to express. Less so if they do not and cannot, read and write the English language sufficiently well to communicate properly. Still less so if they have no knowledge and feel of South African geography and vegetation, and are operating in an outdated paradigm where specimens represent entire species. Explanations which only account for a few plants in cultivation are not going to be very satisfactory. The fact must be faced that classification is difficult and more information makes it more difficult. This is not a problem peculiar to Haworthia. It only seems so because the genus has received so much attention. The question now is:
“How is this information recorded and presented in a way that can be accessed, understood and appreciated?”
My conclusion is that we now need to resort to computerisation and digital imagery, recognising that dealing with single plants is not the same as dealing with populations. It is not easy to assess populations objectively as the sample sizes become unmanageable. We should progressively assess and re-assess populations and build up good herbarium documentation. In this way a proper revisionary process is set in motion, which limits Micky Mouse decisions about hypotheses, classification and nomenclature. ♦
1 a. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.1 b. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.2 a. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 b. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 c. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.3 a. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 b. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 c. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 d. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.4 a. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.4 b. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.5 a. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.5 b. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.6 a. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.6 b. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.7. MBB6825 H. cymbiformis var. transiens to H. gracilis var. picturata, lower Geelhoutboskloof.8. MBB6827 H. gracilis var. picturata to H. cymbiformis var. transiens, east of Geelhoutboskloof.9. EvJ14680 H. gracilis var. isabellae, Vetmaakvlakte, south of Rooihoek.10 a. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 b. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 c. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 d. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 e. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 f. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).11 a. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 b. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 c. MBB6830 H. gracilis var. gracilis, east of Komdomo.12. JDV93/86 H. gracilis var. gracilis, 2km northwest of Andrieskraal.13 a. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 b. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 c. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 d. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 e. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 f. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.14 a. MBB6790 H. gracilis var. picturata, Andreskraal.14 b. MBB6790 H. gracilis var. picturata, Andreskraal.14 c. MBB6790 H. gracilis var. picturata, Andreskraal.15 a. MBB6791 H. gracilis var. picturata, Andrieskraal.15 b. MBB6791 H. gracilis var. picturata, Andrieskraal.15 c. MBB6791 H. gracilis var. picturata, Andrieskraal.15 d. MBB6791 H. gracilis var. picturata, Andrieskraal.15 e. MBB6791 H. gracilis var. picturata, Andrieskraal.16. 1 a. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 b. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 c. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 d. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 2 JDV90/55 H. gracilis var. picturata, also east of Andrieskraal.17 a. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 b. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 c. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.18. JDV94/115 H. gracilis var. isabellae, Paul Sauer Dam Wall.19.1. EvJ13809 H. gracilis var. isabellae, upper Paul Sauer Dam.19.2. EvJ11076 in JDV90/118 H. gracilis var. isabellae (smooth), upper Paul Sauer Dam.19.3. EvJ11076 in JDV90/118 H. gracilis var. isabellae (= MBB6932, Ripon), upper Paul Sauer Dam.20. MBB6932 H. gracilis var. isabellae, Ripon, Krom River.21 a. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 a. MBB6799 H. gracilis var. isabellae, northwest of Patensie.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 c. MBB6799 H. gracilis var. isabellae, northwest of Patensie.22.2. MBB6799 H. gracilis var. gracilis to H. gracilis var. isabellae, northwest of Patensie – a clone resembling H. aristata.23 a. MBB6801 H. gracilis var. isabellae, southwest of Hankey.23 b. MBB6801 H. gracilis var. isabellae, southwest of Hankey.24 a. MBB6802 h. gracilis var. isabellae, southwest of Hankey.24 b. MBB6802 h. gracilis var. isabellae, southwest of Hankey.25 a. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.25 b. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.26. D.Clark1050 H. gracilis var. isabellae, Kouenek, Geelhoutboskloof.27 a. MBB6773 H. gracilis var. isabellae, north of Kareedouw.27 b. MBB6773 H. gracilis var. isabellae, north of Kareedouw.28 a. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 b. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 c. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 d. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.29 a. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 b. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 c. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.30 a. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 b. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 c. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.31 a. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.31 b. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.32.1 a. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.1 b. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.2 a. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.32.2 b. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.33 a. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.33 b. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.34 a. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.34 b. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.35 a. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 b. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 c. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.36 a. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.36 b. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.37. D.M.Cumming6831 H. gracilils var. isabellae, south of Groendal Dam.38 a. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 b. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 c. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.39. PVB7043 in JDV97/7 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.40 a. JDV93/41 H. decipiens var. minor, north of Campherpoort.40 b. JDV93/41 H. decipiens var. minor, north of Campherpoort.41. JDV87/180 H. decipiens var. minor, south of Campherpoort.42 a. JDV91/136 H. decipiens var. minor, north of Wolvefontein.42 b. JDV91/136 H. decipiens var. minor, north of Wolvefontein.43.1. JDV92/140 H. decipiens var. minor, Wolven near Wolvefontein.43.2. JDV94/45 H. decipiens var. minor, Wolven near Wolvefontein.44 a. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 b. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 c. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.45 a. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 b. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 c. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.46. EvJ15748 H. cymbiformis var. transiens, Horee southeast of Uniondale.47.1 a. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 b. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 c. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 d. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 e. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.2. MBB6729 H. cymbiformis var. transiens, De Vlugt, Prince Alfred’s Pass, south of Uniondale.48. J.N.Reddi in JDV93/54 H. cymbiformis var. transiens, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.50 b. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].50 c. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].51 a. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 b. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 c. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.52.1 a. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.1 b. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.2 a. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 b. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 c. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.53.1. JDV91/17 H. gracilis var. viridis, north of Glen Connor.53.2. JDV91/116 H. graciis var. viridis, south of Lake Mentz.54 a. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.54 b. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.55 a. MBB6810 H. cooperi var. gordoniana, Joubertina.55 b. MBB6810 H. cooperi var. gordoniana, Joubertina.56 a. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.56 b. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.57. PVB7079 H. cooperi var. gordoniana, Saptou upper Longkloof.58. PVB7062 H. cooperi var. gordoniana, Redcliffe northeast of Willowmore.59. JDV91/80 H. cooperi var. gordoniana, Engelandkop, Baviaanskloof.60 a. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.60 b. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.61 a. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.61 b. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.
A new variety, Haworthiamucronatavar.rooibergensis is described in Haworthiad 13:5 (1999). It raises many questions, and leaves as many unanswered.
What do the authors, Esterhuizen and Battista, mean by Haworthiamucronata? A very curious picture emerges. There are two sources which must be considered recent and hopefully authoritative. These are C.L. Scott, and M.B. Bayer. Bayer does not use the name mucronata and therefore Scott must be presumed to be the authority followed. But Scott regards H. habdomadis as a separate species whereas Esterhuizen and Battista treat it as a variety of H. mucronata. The two authors also say that their new variety, where it occurs east of Vanwyksdorp (and presumably also their mention of its occurrence south of Calitzdorp), is on the southern boundary of the H. mucronata complex. Who do they follow? If they are using Scott (or Von Poellnitz for that matter) they seem to have mistaken the given distribution. Scott’s distribution map gives five points for H. mucronata which must by south of Vanwyksdorp, and one of these is even west of Mossel Bay. There are also two points north and east of Queenstown.
Curiously, Von Poellnitz is considered to have taken the two Haworth species viz. altilinea and mucronata as synonymous. Scott uses the first for what I called H. cooperi (var. pilifera in my revision), where he also recognises H. pilifera with practically the same distribution. He places his two species altilinea and pilifera in different sections. He does cite the direct synonym H. altilinea var. mucronata = H. mucronata var. mucronata, but Von Poellnitz does not. Scott cites Feddes Repert. 45:169(1938) as his source. I have a handwritten translation in which H. altilinea var. typica is distinguished from H. altilinea var. mucronata. The distributions for these two elements is extraordinary viz. typica from Stockenstroom, Seymour, Redhouse, Grahamstown, Cathcart, Zwartkops, Prince Albert, Hankey, Longmore (Loerie), and mucronata from Adelaide, Albany, (Van)Wyksdorp and Wolwefontein (Jansenville). Effectively H altilinea var. typica was left in taxonomic limbo, and I am not sure if my new revision has followed all these devious ramifications. I can only hope it does. Triebner and Von Poellnitz transfer the varieties of altilinea to mucronata in Feddes Repert. 49:23, 1940 but seem to omit both var. mucronata and var. altilinea.
I can now only try to explain the incongruity of the situation in terms of what I know. Firstly, I know that both Von Poellnitz and Scott were confusing H. mucronata and H. bolusii (at least as far as the variety blackbeardiana is concerned). This confusion is itself indicative of the validity of a new variety which “at fist sight looked very much like H. bolusii“! Secondly I know that Scott also recognised H. aristata, H. mcclarenii and was also noting plants from west of Barrydale (mistaking them for my H. serrata), which I currently think belong with his concept of H. mucronata. This is where I think H. habdomadis also belongs. One has to know that the variety H. bijliana var. joubertii (which I believe came from immediately north of Ladismith per communication with A.J. Joubert) comes into contention. This is in relation to the comparison Esterhuizen and Battista make with H. altilinea var. brevisetata. That variety did in fact originate at Vanwyksdorp, but Von Poellnitz also cites H. altilinea var. mucronata from there. He later cites the var. brevisetata from also Albany and Port Elizabeth. This is in Feddes Repertorium 49:29 (1940) where he also cites H. altilinea var. inermis from Halesowen, Queenstown, Ladismith and Hankey. A better comparison could have been made with this concept of inermis or with joubertii of Von Poellnitz, rather than with brevisetata. I personally think either choice is irrelevant given either:-
1. the various localities cited by Von Poellnitz, or
2. the variation that occurs, for one example, just west if Ladismith where the element habdomadis ‘moves’ from sandstone to shale.
In describing the variety in the detail that they do, and in stating that it “at first sight looked very much like H. bolusii“, the authors invite this comment. I personally very much doubt if any descriptive detail has been given and could be given, which could be used by anyone, to identify a plant of this variety with any certainty from variants of the species H. bolusii, H. cooperi, H. gracilis or H. decipiens, let alone from the many variants of H. mucronata itself. These species are themselves interlinked and I have stated that they are only real as systems in their geographical context.
A curious aside is that, I cannot find the transfer of the name var. brevisetata from H. altilinea to H. mucronata. Von Poellnitz made all the other changes in Feddes Repert.49:29 (1940), and Scott cites this reference for the change to H. mucronata var. brevisetata. I cannot find this in my photocopy of Von Poellnitz’ paper, and Breuer (in his outstanding compilation of the literature) also loses the epithet, as a “Nomen dubium” under H. altilinea. I think he also has lost the var. typica. My statement in my Handbook (1983).. “Von Poellnitz in the same year reduced this variety to synonymy with H. mucronata“, is fallacious and I cannot explain it.
There is a further problem with the citation by Esterhuizen and Battista which reads “North of Uniondale plants are found which may also be H. mucronata.” I consider that this is the same citation for H. decipiens which they give earlier in the paper.. “this species..is growing about 20km north west of Uniondale”. They also relate this to their new variety as “showing (some of the same) characteristics”. The problem is that this is all a vast understatement of its true nature. In my newest book which constitutes a formal revision, I have re-described the situation of the species arachnoidea and mucronata. Here I wholly alter the order I had in my previous works and try to accommodate my respected friend Col. Scott as well. The Uniondale references above are not resolved. There is already a massive interaction or interplay between H. arachnoidea and H. mucronata. The same is evident for H. decipiens and H. mucronata north of the Kleinswartberg, between Calitzdorp and Oudtshoorn, and for H. mucronata and H. lockwoodii south of the Rooiberg Pass and east of the Anysberg. The Uniondale area is grossly under-explored but is known to be touched by the interplay of H. cymbiformis, H. gracilis, H. decipiens, H. bolusii and H. cooperi. This is all fairly evident in Hayashi’s speculative analysis of Haworthia in the same edition of Haworthiad, and it is also evident from the localities cited by von Poellnitz. Unfortunately that work, although its intrinsic message is excellent, also has severe limitations. I have my doubts as to whether Esterhuizen and Battista are aware of any of the above ramifications relating to their new taxon.
Since I first drafted my revision in 1996, I have recognised the reversal in the fortunes predicted by Bruyns in Kew Magazine (4:148, 1987) where he wrote of Haworthia taxonomy being set back 40-50 years. Consequently I have felt the need to do further exploring and recording to expand my observations and possibly verify the predictiveness of even my latest work. I can here confirm that there is a better solution in at least one area and that relates to H. bolusii var. blackbeardiana. I am preparing a separate extensive explanation of that solution. The illustrations for this present article also contribute to that new solution.
The photographs submitted, show the variation which extends continuously from Aliwal North (H. bolusii var. blackbeardiana, JDV97/62-1), southwards to Queenstown (JDV92/22-2), to Stutterheim (JDV94/60-3), across westwards to Somerset East (H. decipiens var. pringlei, MBB6561-4,-108, H. aristata MBB6852-5,-126), to Jansenville and Willowmore (H. decipiens var. pringlei MBB6582-6,-131, MBB6583-7,-132, H. bolusii var. bolusii! MBB6856-8). Then the illustrations show the continuity from H. cooperi var. gordoniana (it could be H. gracilis var. isabellae) from Hankey and Patensie (MBB6799-9,-10,-41) through Joubertina (MBB6810-11,-12,-74) going westward to Uniondale (JDV90/80-13,-75). From here I switch to H. decipiens var. cyanea in the Beaufort West and Merweville districts (JDV90/105-14, MBB6885-15), H. decipiens morphing to H. mucronata west of Prince Albert (H. decipiens var. decipiens! JDV93/71-16,-17), and finally just two of many H. mucronata variants (JDV86/84-18, JDV91/74-19). ♦
62. JDV97/62 H. bolusii var. blackbeardiana, southeast of Aliwal North.63. JDV97/22 H. bolusii var. blackbeardiana, Fincham’s Neck, Queenstown.64. JDV94/60 H. bolusii var. blackbeardiana, Stonefell, northeast of Stutterheim.65. MBB6561 H. decipiens var. pringlei, Patryshoogte, east of Somerset East.66. MBB6852 H. aristata, Stonefountain, south of Somerset East.67. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.68. MBB6583 H. decipiens var. pringlei, Waaipoort, southeast of Mt. Stewart.69. MBB6856 H. bolusii var. bolusii (!), northeast of Fullarton.70. MBB6799 H. cooperi var. gordoniana, northwest of Patensie.71. MBB6799 H. cooperi var. gordoniana, northwest of Patensie.72. MBB6810 H. cooperi var. gordoniana, north of Joubertina.73. MBB6810 H. cooperi var. gordoniana, north of Joubertina.74. JDV90/80 H. cooperi var. gordoniana, south of Uniondale.75. JDV90/105 H. decipiens var. cyanea, east of Merweville.76. MBB6885 H. decipiens var. cyanea, Trakaskuilen, south of Beaufort West.77. JDV93/71 H. decipiens, Gamkadam, west of Prince Albert.78. JDV93/71 H. decipiens, an H. mucronata var. inconfluens look-alke, also Gamkadam.79. JDV86/84 H. mucronata var. inconfluens, east of Calitzdorp.80. JDV91/74 H. mucronata var. inconfluens, northwest of Seweweekspoort, Ladismith.
This article was first published inAloe 36:34, (1999) and is now rewritten in part to accommodate some new collections and a conclusion modified accordingly.
The problem von Poelnitz described Haworthia incurvula (Feddes Repertorium 31:85, 1932), from specimens ostensibly from Grahamstown, sent to him by Mrs. E. Ferguson. The important elements of the description are “leaves about 40, barely 20mm long, up to 12mm wide, broad ovate-oblong, without teeth, hairs or spines, on the face almost flat and inconspicuously turgid towards the apex, on the back convex and towards the apex with a rounded keel, very rarely inconspicuously double keeled, pale green and traversed by greenish longitudinal lines, which are somewhat anastomosing, some of which reach the pellucid tip, which is by no means abruptly pellucid”. von Poellnitz (Fed.Repert. 41:203, 1937) acknowledges that this is the plant illustrated in Flowering Plants of Southern Africa 9:t356 (1929), which is there identified there as H. cymbiformis var. planifolia (Haw.) Baker. Again (Fed. Repert.44:233, 1938) he acknowledges the illustration given by J.R. Brown (Desert Plant Life 8:45, 1936). Here he gives “… the exact locality is Pluto’s Vale in Albany district, between rocks and bushes.” citing R.A. Dyer as the source (Botanical Survey of South Africa, Memoir 17:98, 1937). von Poellnitz adds here that.. “in vigorous cultivated plants the leaves are less incurved or almost not at all”. He was uncertain what its affinities were and placed it in the section Muticae. However, it should be noted that von Poellnitz placed the synonymous H. planifolia (Obtusatae) and H. cymbiformis (Planifoliae) in different sections too. This was despite his confession and obvious in his writing, that he did not know the difference between them.
Smith (SA Journal of Botany 16:1, 1950) transferred the species H. incurvula to the section Obtusatae on the contention that it was continuous with H. cymbiformis which he said was also in the close Pluto’s Vale area. I wrote in my Haworthia Handbook (p124, 1976) “Smith’s field book untypically bears no reference to other collections of this kind in that specific area, and the writer’s limited field experience does not bear this out either. There is however, an apparent if obscure relationship with H. translucens subsp. tenera which bears further investigation. Forms very similar if not identical to H. cymbiformis var. incurvula also occur at Andrieskraal north of Humansdorp, which is a long way to the west.” In my New Haworthia Handbook I could add nothing new, and neither could I add anything substantial in an article I wrote for Cactus & Co 2:11 (1998). Col. C.L.Scott maintained H. incurvula as discrete, writing that “it has been gathered at a number of localities along Pluto’s Vale near Grahamstown”. His riposte is “H. incurvula is a distinct species, and is readily separated from H. cymbiformis“. It must be noted that he does not separate H. cymbiformis and H. planifolia either.
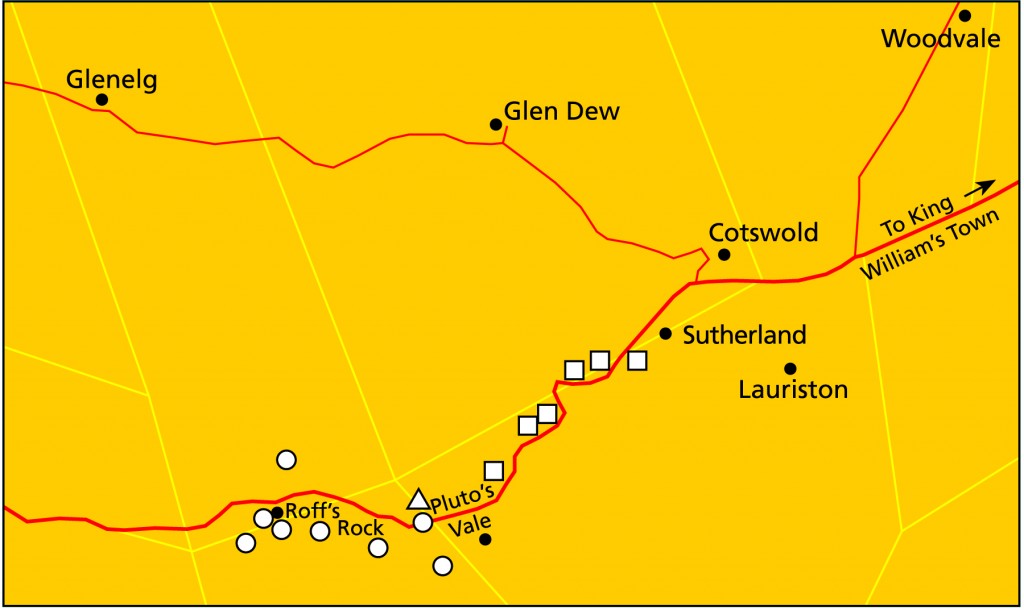
Pluto’s Vale area. The squares represent the sites for “tenera”, the circles for “incurvula” and the triangle marks the site of glabrous “tenera”. “tenera” has also been collected at Glenelg to the north, and H. cymbiformis at Bosky Dell just off the map to the northwest. The origin of H. cymbiformis var. lepida is still further north and west.
Pluto’s Vale is also the origin of H. tenera von Poellnitz (Fed.Repert. 31:86, 1932). The collector was Miss Blackbeard. The site is incorrectly given as “Plinters Vale” and apparently, by courtesy Albany Museum staff, the origin is “Cloete’s Vale”. This is a smaller more proliferating species with more, slender and less densely spined leaves. However, I made a collection from Pluto’s Vale myself circa 1972. In fact two collections (KG47/72a &b), within 75 meters of one another. In one the plants were spinescent, and in the other glabrous. The plants were too small, with the leaves too spinescent and too slender to be incurvula, but the potential for connection was there and that is the predictive stance that was taken. In my New Haworthia Handbook (1983), the point was also made of look-alikes in the Humansdorp area for both tenera and incurvula.
At this point I must state that in my revision (1999) I depart from the name translucens and replace it with gracilis. My thinking subsequent to this change and prior to current writing is, that the name gracilis (from Hellspoort, NW Grahamstown) is in fact a variant of tenera (or vice versa). Scott’s solution is not greatly different except that he applies the name tenera more widely to include elements in the Humansdorp area; and he limits the name gracilis (his use of the name translucens has no bearing on this discussion) to its point of origin.
The hypothesis I now pursue is my own, which is that the element incurvula does not constitute a species and that it can be explained in the context of two other systems (species) as I first suggested in 1976. These are H. gracilis (H. translucens sensu Bayer 1976, 1983: and including the variety H. gracilis var. tenera) and H. cymbiformis (hence H. cymbiformis var. incurvula). Lest anyone again question what I mean by species as do Crouch et al (Aloe 36:8, 1999) in their allusion to “very seldom have some of these authors suggested what might constitute a species”; let me spell out what is implicit in all my classification. Thus – “A species is a system of living organisms which are genetically and morphologically continuous in space and time”. Unfortunately Crouch et al pursue their allusion with considerable vigour and extraordinary circumlocution and tautology. They write of the “amateurish attempts of interested lay persons and taxonomists alike to force an absolute and linear and dichotomous order on the genus”. Unfortunately it is exactly this flawed reasoning and logic that force me to wonder if it is even worth pursuing knowledge anymore (C&SJ.US 70:183, 1998). Our classification system is essentially a dichotomous one and the very concept “genus”/”species” is itself a dichotomy. I will try to explain new findings without attempting to invent a new classification system, although we undoubtedly need one.
New records While re-writing my Handbook and formally revising Haworthia, I was required to cite all herbarium specimens. There are 15 records for incurvula alone and they are all from Pluto’s Vale. None give any detail outside of that geographical information statement. Thus there is no evidence to support Scott’s contention of.. “a number of localities along Pluto’s Vale”. J.D. Venter has a collection (JDV93/60-81 [81 a-d]) by Scott’s colleague, the indefatigable Dr. Reddi, which states “middle of Pluto’s Vale”. This collection was said to have been made north of the road.
There is of course a whole range of specimens of H. cymbiformis from various places in the Eastern Cape. These should really be discussed, analyzed morphometrically and illustrated. These would show why the elements planifolia and cymbiformis can be treated as synonymous. This synonymy is largely due to the origination of the names, and suggests simply that two such morphological elements could have been from a single local system. The herbarium evidence would indicate why the concept “incurvula” is reasonably included there among the many varieties of H. cymbiformis described at one time or another. These are ramosa, reddii, transiens, obtusa (recognised by myself), and further names such as lepida, angustata, brevifolia, alta, agavoides, incrassata, calochlora, longifolia, compacta, exulata, setulifera, obesa and olivacea (which are not).
There appear to be several populations from the greater Grahamstown area in which the plants are smallish and with bluntish incurved leaves (E. Fort Brown, Blaaukranz, Vaalvlei, Fernkloof, Kaffirdrift). There are also populations in which the plants are larger with acuminate, flatter leaves in a more spreading rosette (Committees, Howiesonspoort, Kapp-Fish confluence) and populations with obtuse turgid leaves (Alicedale, Swartwaterpoort).
Here we are expressly concerned with Pluto’s Vale and the plants which occur there. Curiously incurvula is at its closest and most accessible to collection, near the top of the steep descent into the valley, Pluto’s Vale. There is a gate there into the field and a dwelling at a point here called Roff’s Rock. In the tradition of plant collecting in South Africa, as exemplified by the discovery of the first Lithop species by Burchell in the 1800s, collections are invariably alongside a road, or within very short walking distance of some obvious stopping point. The Roff’s Rock site is just such a place and incurvula is less than 50 meters from the roadside. There is a also a vast population of Haworthia attenuata (var. britteniana), and also the plant which the collectors probably actually and repeatedly came to view ie. Encephalartos trispinosus. Incurvula also occurs mid-way down the steepest descent of Pluto’s Vale, north of the road and immediately adjacent to it. Tenera is also known from the farm Glenelg to the north, to the east near Committees and also to the south-east at Hunt’s Drift. H. cymbiformis is a separate element in those same areas and I did cite specimens of incurvula from south of Hunt’s Drift too (Smith5741, Smith6508). I have not seen these populations to confirm that they are, as living specimens, in fact comparable.
In November 1998, I specifically visited Pluto’s Vale again to explore the area a little more thoroughly and to re-locate my glabrous H. tenera. My wife, Daphne visited the area again during July 1999 because of the failure of the previous visit to locate that form. We visited the site again in September 1999 together with Steven Hammer and with Tony Dold of the Schonland Herbarium. A sketch map is provided of the area to demonstrate what we found where. The first visit was collectively quite an extensive search, but by no means adequate to establish the whole truth.
Tenera was found to be common in the Vale itself i.e. east of Roff’s Rock (MBB6832 [82 a, b], MBB6833 [83], MBB6834 [84], MBB6835 [85], MBB6840 [86]). Venter collected at the exit from the valley (JDV90/36 [87 a-c]) and also at Hunt’s Drift (JDV91/103 [88 a, b]).
Incurvula was abundant from the vicinity of Roff’s Rock and extends along that ridge south of the road to where the ridge ends at the base of the Vale (MBB6836 [89 a-f], MBB6837 [90 a-f], MBB6838 [91.1 a, b], MBB6839 [92 a-c]). It was also collected by Marx and Venter (JDV93/73 [93 a, b]) on one occasion slightly towards the south-west of Roff’s Rock, and we also collected it at the second visit (MBB6893 [94 a, b]) still further west. I could not locate the original site of my glabrous tenera (KG47/72a) north of the road and in the Vale. The normal spinose form was on all the south facing rocks. The vegetation seems to have been modified by extensive grazing of the two farms and management systems now evident (the one area is now a game farm and goats have been introduced on the other). However, on the most recent visit we did again find the glabrous form (MBB6891 [95 a, b]) in profusion almost directly above the incurvula population (MBB6892 [96 a, b]) north of the road. In the first version of this report (Bayer, 1999) I incorrectly cited JDV93/60 [81 a-d] as Dr. Reddi’s collection of incurvula from south-west of Roff’ Rock. Apparently it was actually collected north of the road in the middle of the Vale. I can now associate it with my collection [96 a, b] of Pluto’s Vale which is however, of smaller plants than Reddi’s.
The Marx/Venter [93 a, b] collection is of plants with longer (to 50mm) sub-erect leaves which are very lanceolate as opposed to the ovate-oblong shape and incurvature of the leaves in the original description. Traditionally and conceptually incurvula has leaves which are obtuse at the tip. The description does not circumscribe this, and this could have become a very important point. The Marx/Venter plants have such dramatically lanceolate leaves that they throw a slightly new aspect onto the scene. My first collection [89 a-f] between the given Marx/Venter site and Roff’s Rock, also is of plants with such lanceolate and pointed leaves. My wife turned up a similar population north-west of Roff’s Rock (MBB6884 [97 a-d]) in which the plants were smaller and proliferous on a rockier, steeper, south face. There is a stronger tendency to spination. The colour is towards the grey rather than the green. The leaves are more gracile and acuminate. There is no turgidity or swelling of the leaf as one finds in pilifera or in cymbiformis var. obtusa, and given as “inconspicuously so” in Von Poellnitz’ description. But this collection is not the same as the two glabrous ones which so obviously relate to tenera. It does perhaps relate better to Hellspoort gracilis, although it is fairly similar too to [89 a-f], [93 a, b] and [94 a, b].
There is a collection by Marx and Venter (JDV96/4 [98] also by myself MBB6603 [99 a-d]), from north-east of Grahamstown at Glen Craig. H. cooperi var. pilifera also occurs there but about 2km to the east. This collection is of largish plants, greener rather than the bluish/purplish coloration of the cooperi complex. The leaves of the plants are ovate-lanceolate and not turgid above the mid-point. There are many collections of look-alikes from as far afield as Adelaide, the Zuurberg and the Humansdorp area. There is also a very interesting record by R.A. Bayliss from north-east of Grahamstown that needs following-up. These collections impinge on the interplay of H. cooperi var. cooperi, H. bolusii var. blackbeardiana, H. decipiens var. pringlei and H. aristata, which is the subject of another manuscript.
Again: the four collections made from a point south-west of Roff’s Rock to approximately 2 kilometers along the same southern face of the ridge down into the Vale. The first collection was the one described above [89 a-f], of plants with the more lanceolate and acuminate leaves of the Marx/Venter collection, the second [90 a-f] was about 200 meters eastwards which included plants with acute and awned, and plants with obtuse, leaf-tips. One specimen of the third collection [91.2] is identical to a clone in a collection from Alicedale (MBB6847 [100]), a significant collection of H. cymbiformis var. obtusa which I will write about on another occasion. There are close similarities to a collection of mine of this same element from NW Grahamstown (the Bayliss site?, Thornkloof, MBB6895 [101]). That collection was made in pursuit of a R.D. Bayliss collection KG382/76. The progression at Pluto’s Vale to the foot of the slope, was to obtuse leaves and the cymbiformis-like appearance which is also evident in collections of mine from greater Humansdorp (eg. [15 a-e]). The plants were often single and occasionally with more than one rosette in a cluster.
An interesting point is that of the relationship between marginal spination and pointedness of the leaf-tip. One might expect, as indeed I did, that the two situations would be correlated. To a degree they are. Incurvula has leaves which are seldom spined but often blunt-tipped, in tenera the leaves are generally spined and as in glabrous tenera, always with a pronounced point to the leaves. In the case of H. cooperi var. pilifera from Slagtersnek [104 a-e], there are clones in which the leaves are very truncated as in Col. Scott’s joeyii (the var. dielsiana). There is no tip to the leaf at all, and yet the margins do have spines.
The bluntness of the leaf-tips cannot thus be taken to be diagnostic. Here we have pungent and blunt leaf-tips in a continuum of specimens from a relatively extensive area. The “Flowering Plants” illustration appears to have no tip to the leaf per se, whereas J.R. Brown writes “Occasionally a leaf may be found with a very short pellucid point.” His photograph shows leaves with a distinct short abrupt blunt mucro (point), but nothing like the tapered sharp extended point evident in many of the plants I saw. This variation alternative is also evident for H. cymbiformis, for H. cooperi and for H. gracilis within the interactive systems that they are. The phenomenon is also evident in the south-western Cape complex where for example we have H. magnifica which has leaves which are acute with long end-awn, and the var. atrofusca in which the leaves are blunt (obtuse). This is evident within populations.
Tenera was collected in the Vale from a point north of the road [86] from where my search for incurvula terminated. We found it on all of the five rock faces we searched from this point to the eastern end of the Vale. There our earlier expedition ended without real conclusion. It is proliferous and rosettes are usually in clusters. The two elements tenera and incurvula thus grow in close proximity. While in Aloe (1999) I wrote that there was no direct evidence of obvious introgression, or morphological or genetic continuity. My recent collections of small incurvula[96 a, b] and glabrous tenera[95 a, b] require a more committed opinion. The connection is thus through the glabrous populations of tenera directly to incurvula, and also via the three most western and northern pointed-leaved plants from glabrous tenera to incurvula.
In the case of my latest exploration I returned to the Andrieskraal area (SW Patensie) in the eastern Baviaanskloof. I wanted to get some idea of what G.G.Smith had recorded in the Mistkraalkloof area. I already had collected H. gracilis var. picturata at Andrieskraal itself [14 a-c,15 a-e]. This time I collected a little further east [16.1 a-d, 16.2] and these are smaller, non-proliferous plants. One clone was only 16mm in diameter with 35 leaves, while another was 25mm diameter with less than 25 leaves. These plants bear a surprisingly similarity to incurvula, and apart from the blunt leaf-tips, also to the glabrous tenera. Curiously I collected from another population [17 a-c] a little further to the east. This was a glabrous H. gracilis var. isabellae which is very similar too, to glabrous tenera.
In this particular paper I do not consider H. cooperi, which can elsewhere found to be continuous with both gracilis and cymbiformis. H. cooperi var. pilifera is present to the south-west of Pluto’s Vale at Glen Craig, to the north at Fort Brown, and at Committees to the north-east. It does not appear to be involved as such at Pluto’s Vale.
The conclusion There is an ambience to the literature on Haworthia which makes it very difficult to write meaningfully and truthfully about the genus. This ambience reaches a climacteric in the recent article by Crouch et al cited earlier. The review of Haworthia Revisited by the botanist Paul Forster is also so much in the stereotyped and conventional mould of taxonomic botany, that there is little useful to direct scepticism and inquiry in a direction which can possibly advance understanding of classification. This is not true only for Haworthia. Speciation in Haworthia (if not also in many other plant genera, as I have said elsewhere) is chaotic. This can be construed in various ways and I hope it will one day seen to be in a proper sense of order rather than disorder. A search, very thorough by the standards one is generally led to observe, has resolved the relationship between incurvula and tenera to a greater degree than the first publication of this report. Yet, it cannot be stated categorically that incurvula belongs in H. cymbiformis nor that tenera belongs in H. gracilis. A pattern is only evident here for which I have now evidence and this repeats itself in several other areas. Exploration and understanding of these patterns is necessary but is not going to make description and classification in the traditional way, any easier.
One of my observations about natural hybridisation in Haworthia is that hybrids are always very conspicuous, and usually more so because of their close proximity to both parents. On several occasions I have collected hybrids in physical contact with one or other of the putative parents and seldom in the absence of both. The word “putative”, means by inference, but my conclusion is that the use of the word in relation to natural Haworthia hybrids is that it is too passive. The inference I make is that genetic interchange between populations is very limited and populations are able to develop and maintain their own integrity over very short distances. Thus we have glabrous tenera and small incurvula existing as discrete entities in close proximity (75 meters), with a potential third element representing possibly gracilis as the four western collections of plants with the pointed leaves. A parallel situation with three main role players is also evident in the Baviaanskloof and it has always been observance of the degree of co-occurrence of these role players which has led me to my classification. I do not think that these role players really represent “systems” and thus species in themselves, but they can be explained in terms of my classification and revision. I would suggest that my classification is valid as an hypothesis for further inquiry along these same lines.
There are still many outstanding doubts and a decision to “force” change in the dichotomous order will remain very inviting to the uninitiated. There is a very wide interplay between many “species” in what is a very widely spread and intricate system or systems. We are dealing with visual imagery and trying to create order in this. The fact is that we are really dealing with unquantified miniscule differences of size, colour and leaf shape, with a tendency to seek and attach importance to even more miniscule differences of spination, translucence etc. in individual plants. There is also a tendency to attach significance to sophisticated technology applied to limited samples. The problem with this is that it is difficult then for adequate sampling, repetitive testing and casual review.
As mentioned, a similar manuscript which deals with the interplay of the species gracilis, cymbiformis and cooperi, and another which deals with the interplay of aristata, bolusii var. blackbeardiana and cooperi. These show that taxonomic solutions which were thought to be easy to present for some small areas and for small samples are not. Such solutions also break down in other areas. It is very difficult indeed to draw all these “species” into a single discussion, but I feel confident that my classification and revision can be used for this purpose. It has been successfully used like this since 1976. The “group” species solutions as given by T. Salter for Oxalis and by O. Hillyard for Streptocarpus are not very satisfying. It is not a question of forcing the imposition of a linear system. This IS the system in general use. We need another one. I am impressed with a bifurcation diagram given in Glieck’s book “Chaos, the Making of a New Science”. This particular diagram suggests that, made three-dimensional, with asynchronous and asymmetric bifurcation, it could model speciation in plants. The model would be presumably be described by a non-linear differential equation. We need a new language and/or a new paradigm, if we are to name and communicate about plants in a harmonious way, understandable and acceptable way. I hope we will be able to understand that when it comes, because we are not doing very well in the current language we use. ♦
81 a. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 b. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 c. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.81 d. J.N.Reddi in JDV93/60 “middle of Pluto’s Vale”.82 a. MBB6832 H. gracilis var. tenera, east of Roff’s Rock.82 b. MBB6832 H. gracilis var. tenera, east of Roff’s Rock.83. MBB6833 H. gracilis var. tenera, east of Roff’s Rock.84. MBB6834 H. gracilis var. tenera, east of Roff’s Rock.85. MBB6835 H. gracilis var. tenera, east of Roff’s Rock.86. MBB6840 H. gracilis var. tenera, east of Roff’s Rock.87 a. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.87 b. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.87 c. JDV90/36 H. gracilis var. tenera, east of Pluto’s Vale.88 a. JDV91/103 H. gracilis var. tenera, Hunt’s Drift.88 b. JDV91/103 H. gracilis var. tenera, Hunt’s Drift.89 a. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 b. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 c. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 d. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 e. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.89 f. MBB6836 H. cymbiformis var. incurvula, southwest of Roff’s Rock.90 a. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 b. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 c. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 d. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 e. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.90 f. MBB6837 H. cymbiformis var. incurvua, south of Roff’s Rock.91.1 a. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock.91.1 b. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock.91.2. MBB6838 H. cymbiformis var. incurvula, southeast of Roff’s Rock cf.[100, 105a-f].92 a. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.92 b. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.92 c. MBB6839 H. cymbiformis var. incurvula, far east of Roff’s Rock.93 a. JDV93/73 H. gracilis var. gracilis, southwest of Roff’s Rock, Pluto’s Vale.93 b. JDV93/73 H. gracilis var. gracilis, southwest of Roff’s Rock, Pluto’s Vale.94 a. MBB6893 H. cymbiformis var. incurvula, 400m west of Roff’s Rock.94 b. MBB6893 H. cymbiformis var. incurvula, 400m west of Roff’s Rock.95 a. MBB6891 H. gracilis var. tenera, smooth form at foot of Pluto’s Vale.95 b. MBB6891 H. gracilis var. tenera, smooth form at foot of Pluto’s Vale.96 a. MBB6891 H. cymbiformis var. incurvula, foot of Pluto’s Vale, north f the road.96 b. MBB6891 H. cymbiformis var. incurvula, foot of Pluto’s Vale, north f the road.97 a. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 b. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 c. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.97 d. MBB6884 H. gracilis var. gracilis, northwest of Roff’s Rock.98. Marx & Venter in JDV96/4 H. gracilis var. gracilis, northeast of Grahamstown.99 a. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 b. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 c. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.99 d. MBB6603 H. gracilis var. gracilis, northeast of Grahamstown at Glen Craig.100. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.101. MBB6895 H. cymbiformis var. obtusa, Thornkloof, Grahamstown.
One of the greatest difficulties in Haworthia is that of trying to recognise discrete species. This translates into confusion which can be attributed to writers. The initial source of confusion is without doubt the nature of the plants themselves, and this is not a problem confined to Haworthia. The species are often not easily recognisable and discrete entities. I abhor the statement that the genus is in a state of active evolution, but this does at least seem to convey a message that readers understand, even if it is somewhat hackneyed. My observations on Haworthia are based on a definition of species as a system of living organisms which are continuous in time and space. In my New Haworthia Handbook, I suggested that a primary problem lay in separating H. bolusii and H. cooperi, and for the purposes of that work I largely discounted the secondary problems. My first concern was to identify core areas and names as working postulates. This did not mean I was unaware of lesser problems contained within the recognition of those two species. The purpose of this paper is to present my current understanding of the problem.
In my opinion the publication of Scott’s book and whatever merit it had, broke the foundation for understanding. As Bruyns said (Kew Magazine, 1986), it set taxonomy in Haworthia back by 40-50 years. This is because Scott did not attempt to examine what I had done, and was doing, in any objective and cognitive way. Thus there was no progression from a common hypothesis, or from a common concept of ‘species’. In my opinion his work was written in the paradigm of Reynold’s work on Aloe, or of Von Poellnitz and G.G. Smith, and was thus anachronistic. My work was in the intellectual climate of the day, was influenced by researchers of the time and was based on a definition of ‘species’. Scott’s thoughts and actions were both understandable and excusable. His classification reflects the same problems, but differently to mine. Where I recognised H. bolusii he recognised H. bolusii. Where I recognised the var. blackbeardiana he recognised H. cooperi and H. batteniae. Where I recognised cooperi he recognised H. pilifera and H. altilinea. Except for his contention that these latter are separate species, there is really concordance. We recognised different markers along the same continuum.
Essentially I considered that there were two species with two major facies, thus:-
1. H. bolusii – var. bolusii the smaller very densely spined typical variety which seems to diffuse into the species H. semiviva in the west of its distribution range.
– var. blackbeardiana a larger less densely spined element to the south and east which seems to diffuse into H. cooperi. Scott used the name cooperi for this element. There is merit in this, but he had to describe another species, batteniae, because of the inherent tension in the solution (I must point out here that Scott’s problem may have been the inadequate herbarium record that he used for his work, plus the fact that he made few specimens. Thus there is no way of assessing his decisions. Had he had to physically file specimens as I have done, he might have had great difficulty with batteniae and his cooperi, and also in separating his altilinea and pilifera). I regarded bolusii as a more spinose species than cooperi, with thinner and wider leaves. Cooperi then generally less spinose, squatter and with thicker leaves.
2. H. cooperi – var. cooperi itself, with relatively erect slender leaves, and including all the forms with more truncated abrupt leaves.
– var. leightonii – the coastal form with even more attenuate leaves than the typical form, and also very proliferous.
The solutions offered by either myself (1982) in respect of my cooperi and blackbeardiana, or in the case of Scott (1985), altilinea and pilifera, cooperi and batteniae, do have problems. These arise from the nature of the terrain in the area between Graaff Reinet to Queenstown and down to Fort Beaufort. Rugged broken terrain difficult to explore. Like the Baviaanskloof it offers many different classes of habitat, and thus potential ecotypes. The relationship between the varieties is complex and compounded by continuities with other species e.g. gracilis, cymbiformis, and decipiens. It should also always be remembered that any decision is a product of the collecting record and must emphasise that this is the context in which this article is written. Initially I did think that the typical variety of H. cooperi may possibly prove to be well-defined geographically. It has not proved indisputably so, but my classification nevertheless does allow good expression of the continuities which occur. I used the name to cover forms with erect slender leaves as well as forms with relatively blunt truncated leaves. Such plants do occur within the same populations and this is evident in the J.G. Marx collection from Fort Brown. The corresponding Scott names were altilinea and pilifera. To make my system more workable, in my new book (1999) I recognised H. bolusii and its var. blackbeardiana as before, but in H. cooperi, several more varieties, thus:-
– var. dielsiana – a more truncated version of pilifera, the leaves often without end-awns. (Scott’s “joeyii” is synonymous).
– var. truncata – a eastern lighter coloured, proliferous and smaller version of dielsiana.
– var. leightonii – the pinkish, proliferous, slender leaved form growing on granites at Kaisers Beach (Scott maintained this as a separate species).
– var. gordoniana – the erect, slender leaved form in the Hankey and Patensie area, which is very like the typical variety cooperi. In truth var. gordoniana from the type locality is a small very compact plant with short incurved and finely spined leaves.
– var. venusta – a very localised hairy variant from the coast near Alexandria.
I hoped this would explain the variations and resolve the tensions in the solution in an economical way. There is no doubt that Scott’s argument of cooperi sensu Bayer, and blackbeardiana sensu Bayer being the same (and he used only the name cooperi in this context), is correct in the nomenclatural sense. The Thomas River specimen he cites in his book as representative (although Scott does cite the Kew specimen as type, which I have not seen), would have been attributed by Bayer (myself, using the Saunder’s Refugium illustration as type) to my concept of H. cooperi, as also the doubtful gracilis-like elements for the Adelaide and Kingwilliamstown citations. I would have named the Cradock specimen H. bolusii var. blackbeardiana. To bridge the difficulties inherent in his solution, Scott had to later recognise three species viz. batteniae, pringlei and joeyii, with the potential for many more. I have said elsewhere, that a classification which grows with the description of new species, is indicative of a weak system. As in the case of bolusii, the Scott solution is not as economical as mine. His concept of an altilinea and pilifera is essentially the image of my problem in recognising a cooperi/blackbeardiana interface. It would have helped had I initially recognised pilifera as a variety. However, Scott’s interpretation in the sense of the geographical relationships of his various species is problematic throughout his revision, where specimens are not cited and full use has not also been made of the available herbarium record.
I was predicting that an element, namely H. cooperi var. cooperi occurred which could be defined in a geographic context. The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain goodwill. This article thus is written to expand on the problem and indicate where the difficulties lie. These are not difficulties that I was oblivious of, or tried to obfuscate. I have many times said that we can find solutions which work in one area, but they may not work in another. This will again be evident in this paper.
In my revision (Haworthia Revisited, 1999) I made the combination H. decipiens var. pringlei and resurrect the old species name H. aristata. These two elements, and an explanation for the tensions which these two names create and try to address, will be touched on here but will follow in more detail in a later article. As in this immediate case, new material has been seen and collected since the revision was drafted, and a better explanation can now be made.
For this particular issue, the key questions to the hypotheses of Bayer and Scott are difficult to formulate because they are confounded by the different use of names. Therefore I frame the questions in terms of my own classification like this:-
Does an element we can identify as cooperi grow in the same close geographic context as blackbeardiana? (Bearing in mind the assumption that the hypothesis regarding continuity between bolusii and blackbeardiana in fact is valid. Also noting that to maintain the classification hypothesis that there is a difference at the rank of species, the answer must be ‘yes’).
Does the element we can identify as cooperi grow in the same close geographic context as pilifera? (Bearing in mind that to maintain the classification hypothesis of a difference only at the rank of variety, the answer must be ‘no’).
Results The answer to question 1:- has never been a strong point of my classification because I saw cooperi in a broader sense to include pilifera (and altilinea of Scott). The recognition of varieties now strengthens the classification in one respect but weakens it in another. It strengthens the classification in respect of recognising the extent of the variability within my concept of H. cooperi. It weakens it in suggesting both a strong geographical separation of the varieties and a weaker relation with H. bolusii var. blackbeardiana. In Nov.1996, I went with Peter Bruyns to the Eastern Cape and we spent some time in the greater Somerset East area. In Dec.1996 I travelled with Kobus Venter. In Dec.1997, Dec.1998, in Sept. (with Tony Dold, Dez Weekes and Steven Hammer) and again in Oct.1999, I travelled with my wife to these areas. We made many significant collections.
The first of these concerns Scott’s species, H. joeyii and H. pringlei. In the case of joeyii my contention that it is continuous with pilifera barely needs discussion, and I do not think the presence or absence of an end-awn is necessarily diagnostic for such an element – hence synonymy with var. dielsiana. We found three discrete populations within a small radius around Eastpoort (MBB6558-102, 6559 and 6560-103) which support this observation. I do not think there are strong grounds for separating it from pilifera, although I have now done so. At Bedford (there are plants with the similar abrupt leaf-tip as well as plants without. I currently have four batches of seedlings which I want to examine for variation. One from Slagtersnek, south-east of Somerset East (MBB6778-104) is particularly variable. I even have a collection of H. cymbiformis (MBB6847-100,-105) which has the same truncated, awnless leaftips as Scott’s joeyii. Regarding Scott’s pringlei, my conclusion that it is related to decipiens is possibly geographically and otherwise incorrect. (It should be noted that no specimen of pringlei has to date been deposited in the Pretoria herbarium. My understanding of the species is from the description, from two plants sent to J.D. Venter by J.N. Reddi (JDV93/46-106) who collected the plants for Scott, and from two plants given to G. Marx (JDV93/52-107) also by Dr Reddi). It is a problem of ‘look-alikes’ and I will deal with that a little later as a separate issue.
Further south and east of Eastpoort, at Patryshoogte (MBB6561-108 also -4), we found a different population not to my knowledge co-occurrent with a cooperi variety, which must be comparable with Scott’s species i.e. pringlei. It has narrower and more elongate leaves than blackbeardiana generally has, and the keel may be more pronounced. Nevertheless I am sure it has a connection with H. bolusii var. blackbeardiana. In terms of a much wider knowledge of other populations, I decided to link pringlei to decipiens, to emphasise the evident and probable continuity between that species and H. bolusii. This is particularly so in the area north and east of Jansenville.
To the south-west at Somerset East (MBB6776-109, Glen Avon), we found a population of bluer green plants with rather more attenuate leaves than in the vars pilifera and dielsiana. These plants are very like an Adelaide (Koonap Bridge, MBB6563-110) collection, these both satisfy my concept of the var. cooperi. It was not much further to the south (-104), and also not far south of Eastpoort, where we found forms of the vars pilifera and dielsiana in the same population. Thus it seemed that a hypothesis maintaining the species cooperi and H. bolusii var. blackbeardiana, was well supported, but not so the contention that pilifera and dielsiana were good varieties.
The most significant discovery up to Dec.’97 was of two different elements growing together towards the south, near Ripon Station. By together, I mean as populations occupying different niches in the same close proximity – not growing as a common medley of individuals. One population was clearly the typical blunt-leaved pilifera (MBB6557-111) which included dielsiana, but the other (MBB6556-112) was with more slender and erect leaves viz pringlei, and possibly the leaves being broad and spinose enough to even represent blackbeardiana. Thus the added complexity is that it is not possible to separate those two elements. After two years in cultivation as field collected clones and as seedlings, the difference from pilifera (as -111) is maintained. The collected clones grown in Kobus Venter’s collection convincingly demonstrate that the one population is identifiable with pringlei/alias blackbeardiana and the other with cooperi var. pilifera. Thus the answer to the first key question is “Yes, they do grow together in the same geographical context, because there is that obvious connection of blackbeardiana with pringlei. It seems probable that both these elements could, and perhaps should, be interpreted as varieties of H. cooperi. It is a complex problem which cannot be separated from consideration of other elements such as H. gracilis and H. aristata, and these are the issues I am addressing in these papers.
There is not much further evidence that cooperi var. cooperi is a strong, discrete and valid element. There is a collection by J.G. Marx of plants in the Hogsback area (in JDV91/82-113, Woburn) which may be identified as this variety, as can the two collections already mentioned viz. -109 Glen Avon, and -110 Koonap Bridge (originally a Marx collection). However, there are collections generally (one is D.M. Cumming8489-114 of plants south of the Waterdown Dam, Upper Chilton), where the plants can be confused with H cooperi var. gordoniana (in the context of the problem in the Baviaanskloof), whereas geographically they should be blackbeardiana. I also made a collection at Waterdown Dam itself (MBB6569-115), of a plant which is very like a spineless smooth gracilis. These plants tend to be much more blue in colour than the southern look-alikes. There is also JDV96/89-116 Gladhurst, Adelaide; and JDV96/4-98 and MBB6603-99 from Glen Craig, north-east Grahamstown, which pose a similar conundrum. These are gracilis-like and while exploring that problem, the issue became very much more prominent, and is even expressed in the discussion regarding H. cymbiformis var. incurvula. Kobus Venter and I had both re-collected H. gracilis var. gracilis at Hellspoort (JDV89/42-117, MBB6614-118), although we had not concerned ourselves with a search over the whole of the valley. What is now becoming more evident is that there is an archetype which is gracilis-like. Thus Hellspoort needs to be examined as closely as Pluto’s vale.
A later collection of mine (Oct.’99, MBB6927-119, W Ripon) influences the picture dramatically and may further prove a statement of mine true. This is that there instances where there are no real boundaries between species. My collection is further west of -111 (pilifera/dielsiana) and -112 (pringlei/blackbeardiana). Here in -119, the plants include individuals which could be either identified as the vars cooperi or pilifera, with the former collection equally well representing pringlei/blackbeardiana.
The answer to question 2:- has not really been a problem. What we found was what we expected to find and more. South of Adelaide one finds populations of plants which satisfy the tendency towards slender leaves ie. var. cooperi (-109 Glen Avon or -110 Paardefontein, Koonap Bridge); and populations which satisfy the tendency to short obtuse leaves ie. var. pilifera eg. MBB6564-120 Chancery Hall or MBB6591-121 The Tower, S. Fort Beaufort. The same applies just east of Somerset East where in the broader geographical context we can identify var. cooperi (-109, Glen Avon), var. dielsiana (when mostly or wholly without end-awn, MBB6565-122, W. Somerset East, -111 W. Ripon Statio, and vars dielsiana and pilifera as geographically complementary and even in admixture (-104, Slagtersnek). Thus “No, the varieties cooperi and pilifera do not grow together in the same close geographical context as discrete entities.” In fact they are often represented as forms in the same collections as in -119 (far west of Ripon) where both varieties are present as single plants as well as a longer leaved form representing blackbeardiana as decipiens var. pringlei.
The answers given to these two questions thus do still not solve the problem as there remain populations which can not confidently be ascribed to either of the names here used. There are populations which may thus be assigned to cooperi or to blackbeardiana or pringlei. An example is the Marx collection from north-east of Grahamstown (W Fort Brown, in JDV91/85-123) which is ascribable to pilifera although some of the clones are inseparable from blackbeardiana. JDV98/39-124 from Brakkloof, northwest of Grahamstown poses a similar problem with the leaves of the plants tending to lengthen in cultivation. The ultimate difference then between cooperi and blackbeardiana becomes a very subtle one of degree of spination and leaf bulk. The issue is further complicated by the fact that four other elements are involved in the total geographical context, namely H. cymbiformis, H. gracilis, H. decipiens, H. aristata and H. arachnoidea var. xiphiophylla.
My most recent field work, was in following up information and collections concerning H. gracilis. Tony Dold of Rhodes University gave specimens of an Haworthia from the Annsvilla area to Gerhard Marx (MBB6851-125). He thought they may have some bearing on the existence and reality of my interpretation of H. aristata, and this is indeed the case. The plants are small, the darkish blue-green of the cooperi/blackbeardiana elements, and with more, smaller, and quite spinose leaves. Annsvilla is close to three localities for that species cited in my revision viz. Verdun, Stonefountain (re-collected in MBB6852-126 also -4) and Kommadagga. Following Dold’s find, recollection in the vicinity of each of those sites, produced plants which can be interpreted and identified in the light of the Ripon collections. While they equate in some way to H. bolusii var. blackbeardiana, they also do to H. cooperi and to H. decipiens var. pringlei. An (unfortunately doubtful) Kommadagga collection (EvJ sn.-127) is very similar to the long-spined gordoniana-like plants from the eastern Joubertina -74, Uniondale -75, Longkloof -76 and Baviaanskloof (Geelhoutboskloof -78, Nuwekloof -79). The leaves of the plants are rather flatter and broader, and so closer to blackbeardiana. In Sept.’99 Dold and Hammer found a small plant with shorter and squatter leaves than my concept of aristata, south-east of Kommadagga (in MBB6897-128). We also saw plants from the area east of Jansenville which seem to link the greater H. bolusii (i.e. var. blackbeardiana) complex with either H. decipiens or H. arachnoidea var. xiphiophylla. Included in that broad statement is an explanation for the specimens which I have cited as H. decipiens var. pringlei in my revision. These include recent collections from north of Jansenville (MBB6580-129), north-west (MBB6581-130), and south-east of Mt Stewart (MBB6582-131 also -6, 6583-132, also -7). When I first saw a specimen of pringlei my first reaction was recall of plants I had collected at Mt Stewart, and of plants said to have been from Jansenville. Thus I created H. decipiens var. pringlei, expecting to find this east-west continuity between blackbeardiana and decipiens. I did not think that the connection between Middleton (pringlei) and Jansenville would be found to be as evident and as strong as it is. There was already some evidence that there is south to north-east connection between Jansenville and, say, Cradock. This is of a connection between H. decipiens and H. bolusii.
A complicating issue is that MBB6587-133 (further SE Mt Stewart than -132) is a population which is H decipiens var. minor. A small plant with a tendency for incurving leaves but with the widely spaced large spines I associate with the more classical view of H. decipiens. The next collection was MBB6589-71.2 (a little south-east of -133) and it throws us into the H. gracilis context of smaller plants, clustering on cool south-facing rock faces. There are many collections which continue this trend to gracilis, and in fact I suspect to xiphiophylla too, and that will be dealt with as a separate issue. The field-work also revealed populations (J.G. Marx in MBB6845-134, east of Alicedale, and MBB6847-105, also Alicedale) which link cooperi to cymbiformis. This also can be dealt with together with two collections from near Kagasmond (MBB6562-135, south of Adelaide and JDV96/89-116, Gladhurst) as a slightly different issue.
Regarding H. aristata, I resurrected this name for several collections from the Eastern Cape, and somewhat justified by the Dold collection at Annsvilla (-125). However, it seems quite certain that the issue is clouded. Re-collections at the Soutkloof (JDV96/90-136, Addo, Dead Man’s Gulch) were more like pilifera, but there is evidence of deviation. Here D.M. Cumming (DMC3870-137 collected pilifera-like plants, whereas Venter, Marais and Bayer (-136) collected a variant – there was a more typical bluish-green pilifera and a plant tending to the opaque yellow-green of xiphiophylla. I returned there in Oct.1999 because I still had not seen anything like the plants I had seen there on an earlier visit, or like W.R. Branch’s original (WRB459 in JDV87/53-138), and the other cited collections. On this occasion I did find a small population (MBB6920-139) of this kind, and these confirm a relation to the more northern collections mentioned above. At the nearby village of Addo itself, there is the fairly normal pilifera (JDV86/117-140). The Annsvilla collection (Dold and Marx -125) and the recent collection of my own from Stonefountain (-126 also -5) seem also to be in the context of the Ripon collection (-112) of pringlei. These are, however, collections of much smaller plants than from Ripon. Thus it confirms for me that the Haworth names aristata and denticulata could easily have had their origins in plants from this area between Ripon and the Zuurberg, or further south to Soutkloof. My collections MBB6916-141, MBB6917-142 and MBB6901-143, from Kaboega and Hopewell are of the same order.
Discussion and conclusions There is still a very large area unexplored. There are several like collections from the greater area Kirkwood to Uniondale which I have generally ascribed to H. cooperi var. gordoniana. None of these are the short-leaved pilifera type. The teasing probability is that it is in fact blackbeardiana, pringlei, gracilis or aristata (as variants of H. cooperi) which are the main role players. They differentiate (clearly?) in the east to pilifera or cymbiformis and in the west into decipiens. In the south they pass to xiphiophylla, cymbiformis and varieties of gracilis. North-westwards it is to bolusii. My opinion now is that we have an archetype which is in the mould of H. gracilis and this is the root of all the elements I have named here. Curiously Tony Dold has recently sent me specimens from Chalumna (T.Dold3961 in MBB6921-144) which suggest that an aristata-like element is also associated with H. cooperi var. leightonii. Other Chalumna collections (MBB1621 and G.G.Smith 514) bear a very close resemblance to plants (-99, -116) which I have said are gracilis-like.
The above statements all have to be seen in the overall statement about continuity. It barely seems practical (nor legally possible) to sample and analyze plants on the scale that will be necessary, using whatever technique, to get a more definitive answer. The best answer will be continual exploration of the simple kind reported here, which explains occurrences on a smaller and smaller scale. There are herbarium records which need to be corroborated in the field but these do not suggest to me a better solution. Ultimately each recorded population will have to be assigned unequivocally to a taxon (or taxa!) and this is the next necessary step in Haworthia classification. Another revision based on less exacting field observations and a lesser record, will simply exacerbate the regression in time which Scott’s work precipitated, and the consequences which we now suffer. My recent collections are only preserved as living collections and photographically. Specimens need to be made.
I am very conscious of other tensions in my classification and I think it is imperative that we stay with one nomenclatural arrangement and hypothesis to resolve these. The confusion which arises from nomenclatural changes which are nothing but pretentious and cosmetic, is not worth any price. Any difficulties in respect of nomenclature can be resolved by a process of explanation and conservation without formal name change. These changes can be made when it is expeditious to do so and when changes can really offer a better explanation of the genus and hence better communication.
What this discussion should demonstrate is the problem of really observing and discussing variability in Haworthia. There are many possible arrangements of names which can be presented as conclusions in themselves – but done like this they simply cloud and destroy any hope of a broader understanding and good communication. Haworthia is now so much in the public domain, that I would suggest to editors that they move in the direction of encouraging authors to adopt a conserved nomenclature. I regret to be so straightforward and blunt. I see nothing but further confusion if persons feels that they can contribute to an understanding of the situation in the field without:
Consensus on the issue of species definition.
Consensus on the issue of a set of names.
Familiarity with herbarium records and what these represent in terms of fact and fixed reference points.
Familiarity with the written record.
Familiarity with South African geography and the ability to interpret populations in that context.
♦
102 a. MBB6558 H. cooperi var. dielsiana, north of Eastport.102 b. MBB6558 H. cooperi var. dielsiana, north of Eastport.103 a. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 b. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 c. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 d. MBB6560 H. cooperi var. dielsiana, south of Eastport.104 a. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 b. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 c. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 d. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 e. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.105 a. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 b. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 c. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 d. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 e. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 f. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.106 a. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.106 b. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.107 a. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.107 b. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.108 a. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 b. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 c. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.109 a. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 b. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 c. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 d. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 e. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.110 a. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.110 b. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.111 a. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 b. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 c. MBB6557 H. cooperi var. pilifera, west of Ripon station.112 a. MBB656 H. decipiens var. pringlei, west of Ripon station.112 b. MBB656 H. decipiens var. pringlei, west of Ripon station.112 c. MBB6556 H. decipiens var. pringlei, west of Ripon station.112 d. MBB6556 H. decipiens var. pringlei, west of Ripon station.113 a. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.113 b. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.114 a. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.114 b. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.115. MBB659 H. bolusii var. blackbeardiana, Waterdown Dam.116 a. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 b. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 c. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 d. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 e. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.117 a. JDV89/42 H. gracilis var. gracilis, Helspoort.117 b. JDV89/42 H. gracilis var. gracilis, Helspoort.117 c. JDV89/42 H. gracilis var. gracilis, Helspoort.118 a. MBB6614 H. gracilis var. gracilis, Helspoort.118 b. MBB6614 H. gracilis var. gracilis, Helspoort.119 a. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 b. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 c. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.120. MBB6564 H. cooperi var. pilifera, Chancey Hall, south of Adelaide.121 a. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.121 b. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.122 a. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 b. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 c. MBB6565 H. cooperi var. pilifera, west of Somerset East.123 a. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.123 b. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.124 a. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.124 b. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.125 a. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.125 b. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.126 a. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 b. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 c. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 d. MBB6852 H. aristata, Stonefontain, s. Somerset East.127. EvJ H. aristata, Aalwynspoort, east of Commadagga.128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.129 a. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 b. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 c. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 d. MBB6580 H. decipiens var. pringlei, north of Jansenville.130 a. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 b. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 c. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 d. MBB6581 H. decipiens var. pringlei, northwest of Janesville.131 a. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.131 b. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.132 a. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.132 b. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.133 a. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.133 b. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.134 a. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 b. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 c. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.135 a. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 b. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 c. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 d. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.136 a. JDV96/90 H. cf. aristata, soutkloof, Addo.136 b. JDV96/90 H. cf. aristata, soutkloof, Addo.136 c. JDV96/90 H. cf. aristata, soutkloof, Addo.137. D.M.Cumming3870 H. cf. pilifera, Soutkloof, Addo.138 a. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.138 b. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.139 a. MBB6920 H. aristata, Soutkloof, Addo.139 b. MBB6920 H. aristata, Soutkloof, Addo.140. JDV86/117 H. cooperi var. pilifera, Addo.
Introduction Kaboega (also spelt Kabouga) is now an assemblage of farms (De Plaat, Wilgerfontein, Vygeboomfontein, Klipfontein) nestled against the north slopes of the Zuurberg mountains, north of Kirkwood. It is only about 15km away from Kirkwood as the crow flies, but 150km away by road. Oudekraal is about 20km east and it is the source of Haworthia angustifolia var. baylissii and Gasteria baylissiana. There are several records of Haworthia for the Kirkwood area, and von Poellnitz named H. stiemiei (Regarded as insufficiently known and not recognised by Col. C.L. Scott or myself) from there. He also identified plants from Kaboega and Uyepoort, both described as “at Kirkwood”) as H. altilinea var. denticulata (Haw.) v. Poelln. These plants are all in the melange that I attribute to H. cooperi var. gordoniana (the subject of another long essay). The Kaboega farm lies on the Kaboega river which drains an area of about 1m ha and then flows through the long Kaboegapoort into the Sundays River just north-west of Kirkwood. The terrain is very broken with the sandstone Zuurbergs themselves dominating the southern boundary at about 850 to 950m above sea level. The lowest point on the farm is at about 300m and the northern lesser shale or dolerite peaks reach 550 to 650m. The vegetation on the sandstones is Dry Mountain Fynbos. North of this is Karoo Valley Bushveld. Thus Kaboega is at an ecotone of the karoid veld, Eastern Cape grassland and the Noorsveld (Euphorbia thicket) of the Jansenville area.
The name Kaboega means ‘the Big Hole’, referring to the deep gorge which the Kaboega River cuts through the Zuurberg. This river joins the Sundays River where it skirts the eastern end of the Kleinwinterhoek Mountains. Thus Kaboega Gorge is about 20km east of the Sundays River Gorge and 20km west of Oudekraal where the Witrivier also cuts through the Zuurberg flowing southwards.
The only known Haworthia collections from Kaboega prior to this report are two collection by Gerhard Marx (JDV91/14-145, JDV91/15-146) from the easterly part of the farm (DePlaat, north and south aspects), and a similar collection by Peter Bruyns (PVB5002 in JDV92/33-147) from Kaboegapoort itself. Discounting the strange (expected) internal variation in the latter, these three collections are fairly similar. Plants with the brighter yellow-green of H. cymbiformis, but with more terete and slender sub-erect leaves. The plants are quite robust and in cultivation reach about 80mm diam. with leaves up to 90mm long. Look-alikes are found in the gracilis, cooperi and cymbiformis var. transiens complex of the Baviaanspoort, and I have generally referred these all to H. cooperi var. gordoniana. However, that variety is actually quite a distinctive one from the Hankey Pass, north of Humansdorp and perhaps I should never have adopted it for general use in the way I have. Thus in my discussion about H. cooperi and H. bolusii var. blackbeardiana, I speculate that gracilis is an archetypal form which may lie at the root of the Eastern Cape species here being discussed.
The species H. aristata poses similar problem, and so does H. decipiens var. pringlei (Scott) Bayer as well as two collections which I and Bruyns made at Ripon station which is north-east of Kaboega. One of these latter collections is H. cooperi var. pilifera (-111) and the other is H. decipiens var. pringlei (-112). Largely because of that collection, I felt pressured into believing that the latter would best be coupled with H. bolusii var. blackbeardiana rather than with H. decipiens, and I was contemplating a major change of this kind. This would also have involved subsuming that element in an enlarged H. cooperi var. cooperi. There are, however, some other collections from the greater Darlington Dam (Lake Mentz) area to the west, which are relevant to this problem. These include older ones which suggested the link of pringlei with decipiens which I was thus also predicting, and new ones which confirm that this does in fact happen.
Because of the extent and complexity of the problem, this report deals specifically with the Kaboega area. In conjunction with it, a manuscript regarding H. cymbiformis var. incurvula, was written to give another indication of the nature and scope of the problem of classification of Haworthia. However, the chief reason for the visit to Kaboega was somewhat fortuitous. I was intending visiting Pluto’s Vale again, also the farm Thornkloof where Col. R. Bayliss had collected; the place Aalwynpoort to check on an Ernst van Jaarsveld collection and also hoping to cast light on a collection from the Bosberg at Somerset East. Peter Bruyns was hoping I would recollect a Stapelia aff. kougabergensis which he had seen on the Zuurberg, and there are also some other Haworthia records in that general area which need verification. What also materialised was a visit by Steven Hammer to South Africa, and contact with Tony Dold of the Schonland Herbarium at Grahamstown. The best of all was contact by my wife Daphne, with the gracious managers of the Kaboega farming enterprise, Sandy and Ian Ritchie. Through their kindness and hospitality we came to spend four days on the farm and briefly explored what it has to offer. One day there was spent with Steven Hammer, and with Tony Dold and Dez Weekes of Grahamstown. My wife and I returned for a second visit in Sept. ’99, when we also went to Oudekraal via a direct farm road from Kaboega to the east.
Results On our first visit we first explored the western side of the farm known as Wilgefontein. Tony and Dez went up the slopes of the Spekboomberg on the north side of the valley, and the remainder of the party climbed to near the top of the Zuurberg. We climbed straight to the grassveld where the grass was very long and thick. We soon found a solitary-growing greyish-green plant in flower (MBB6904-148), and then higher and on vertical rocks, a less translucent clump-forming plant with a velvety texture – also in flower (MBB6905-149). The plants looked rather different and we were quite excited about it being something apparently out of the ordinary. It is possible in the context of later collections, that these two collections are ecotypes. Looking at similar rocks about 200m to the south-east, we found what at first was obviously H. coarctata var. adelaidensis (MBB6907). This turned out to be a big population of plants which can, as such, be collectively regarded as intermediate between H. glauca and H. coarctata. This is a very significant collection because of the occurrence of glauca in its typical form at Zuurberg Pass, and nearer at Oudekraal, both to the east. Var. coarctata itself is not known nearer than at Patterson 70km eastwards and var. adelaidensis from east of Riebeek East which is still further away. (see also ‘Haworthia Revisited’ p179). What we did hope to see was H. angustifolia known at its western limit from Oudekraal. It must be on Kaboega and we just have not seen it yet.
Across the valley Dez Weekes had collected three specimens from a south facing steep cliff (MBB6903-150). These had long stems with bright green terminal rosettes and I have identified these as H. cymbiformis (with reservations! as I think this could again be a local ecotypic adaptation) without seeing the plants in habitat myself. This species is also not known from nearer than Hell’s Gate 50km to the south, but – it almost certainly has affinities with the plants collected by Marx and Bruyns. Steven, Tony and Dez had to leave after the first day, but Daphne and I continued the exploration the second day with a long climb up the hill east of DePlaat. We soon found Gerhard Marx’ (MBB6909-151) plants at the base of the mountain and continued eastwards and upwards. We came across a very extensive and dense mass of H. glauca (MBB6908). The plants were variously tubercled and lacked the distinctive grey colouration of the species. Any affinity with “coarcata” was less obvious than at Wilgefontein. Another interesting plant there was a dwarf form of Aloe tenuior. We crossed over to the steep north slopes and on the way down came across three plants (MBB6910-152) of what appeared to be similar to the velvety plant of the previous day (-149). It was in the same rocky situation. We looked further for it, but failed to find it again probably only for lack of concerted effort. The terrain was very difficult and we were getting a little hot and tired. We came across the Marx plants again. These were further down the hill and looking rather bleached in the sun.
149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.
On the way home we were travelling across some very stony ground covered with scattered bush and a low-growing Felicia, probably filifolia. Underneath these, in algae and moss, and with Euphorbia stellata and Tylecodon reticulatus were single plants of a cryptic small blue-green species (MBB6917-142). This I relate to collections from Stonefountain and Verdun cited in my revision under H. aristata, again collected as MBB6852-126 and Dold in MBB6851-125 (Modderfontein).
142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
The following day, accompanied by Sandy Ritchie, we ventured into the Kaboegapoort itself. We walked to the boundary with the Addo National Park. On the way we had seen inaccessible clumps of an Haworthia on a very steep cliff and we tried to reach these on the way back. We were lucky to be able to sample four clumps on the first of the rock faces where some plants had established themselves off the face (MBB6911-153). On the other cliff we could not reach anything. The plants were not cymbiformis but relate rather to the Marx plants except that they were clump-forming and bleached. A better comparison is with (-148). They were also in flower.
148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.
Peter Bruyns collection (-147) is of six clones from this poort and each of these plants is different. However, one clone (-147.1) resembles gracilis var. viridis from Perdepoort (-70) and also resembles a collection of Bruyns of cymbiformis var. transiens (-69) from the Langkloof. (This latter collection is involved in the issue of that species or gracilis var. picturata). Kobus Venter also has a collection of similar plants from south of Lake Mentz (-72.2). A second clone (-147.2) was an aberrant plant with very terete, abruptly mucronate leaves which were also unusually translucent. Another clone (-147.3) was of a plant identical to the big gracilis var. isabellae of the Krom River Estuary (Ripon, WRB1, recollected by myself -39), and comparable with a collection (MBB6855-154) from Waterford, east of Jansenville. That collection could be identifiable as H. arachnoidea var. xiphiophylla and perhaps hinting at a link of that element with H. decipiens var. minor. Two clones (147.4, 147,5) are the same as the DePlaat collections (-145, -146), resembling the Gladhurst (-116) and Glen Craig (-98,-99) forms of gracilis. Thus representing my view of an archetype.
147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.
On our last day, Ian Ritchie kindly took us on a drive to territory which had looked quite visitor friendly from the top of the Zuurberg. On closer contact they are anything but so. On the Spitzkop, which is on the northern boundary of the farm, we found aristata (-141) growing on bluish shale in a situation favoured by H. cooperi var. cooperi. It was a little bigger than our previous collection, but smaller than plants we subsequently collected northwards on the road between Riebeek East and Jansenville (Paddafontain, MBB6899-155), which I refer to pringlei). Driving to the east of Spekboomberg we saw plants (MBB6914-156) similar to the Marx plants of DePlaat (-145) in profusion, some of them without the softer translucence of the Marx collection. The leaves were occasionally much flatter and ovate and distinctly reminiscent of cymbiformis. Daphne and I walked down the hill from that point seeing the plants for most of the descent. Ian in the meantime drove back and further to the south-east and then found very similar plants on a steep slope also facing south-east (MBB6915-157). These plants can also occur in large clusters.
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.
Daphne and I drove to Somerset East to reconnoitre the Bosberg and on the way back saw aristata again about 10km north-east of Kaboega (-143). We had looked at the Paddafontein (-155) plants on the way out and they quite obviously can be linked to aristata too. Unlike the other collections of aristata, however, the Paddafontein plants have a large robust inflorescence with many flowers open (usual in decipiens) as opposed to say the Commadagga (-128), aristata, but with fewer and stumpier leaves) dwarfs with only 8-10 flowers per stalk and one open at a time (thus more pilifera-like).
128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.
During our second visit, we again went to the top of the mountain at Wilgefontein, after Ian had shown us H. glauca in the Kaboegapoort itself. This population was not typical of the species and also more like the De Plaat plants. We revisited the site of -148 and -149. Both were in flower and on this occasion we found the grassy ecotype within about 75m of the stone-face plants. The latter had flowered but seed-set was very poor, as opposed to the grass element which had well-formed capsules. From there we went to the Dez Weekes’ slopes via a different route and collected (MBB6925-158) plants ranging from the same greenish cymbiformis-like plants of -156 to specimens which could be nothing else but typical of true cymbiformis. We saw the same plants again at the dam to the north-west of the homestead, and again on the south slopes (-159) behind the previous De Plaat collection (-151). We completed the stay with a visit to Soutkloof where we saw again the true aristata (-139), and also to the office of the Addo National Park on top of the Zuurberg. Here we saw specimens of H. cooperi var. pilifera from that vicinity, and similar to a collection by Ernst Van Jaarsveld from Oudekraal (I had seen these plants when I collected H. angustifolia var. baylissii there many years ago). On our visit to Oudekraal we stopped to the west of my previous visit and probably also west of where Ernst had collected. We found the cooperi-like plants (MBB6922-160) again growing among rocks in dense grassland. The plants had very pronounced reddish-lines in the leaves and this was evident in all the Kaboega collections.
158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.
Curiously a post-graduate botany student busy with a study of succulent endemism brought in a number of plants for identification. This is P. Desmett, and among his collections, two are relevant. One is PD2310-161 from Boplaas. This is north-east of Kirkwood where the Kaboega meets the Sundays River. The plant is a small spinose specimen which could relate to the arachnoidea-like (as von Poellnitz compared it) stiemiei. It could alternatively, and because of its colour and translucent patterning of the leaves, be more probably compared with H. decipiens var. minor represented by several MBB collections from Sapkamma (MBB6618-162, MBB6619-163, MBB6620-164) to the west. The other is PD2309-165 from the southern end of Kaboegapoort. It is the apparently puberulous-like element resembling -148, and also -153. Kobus Venter also collected and aristata-like plant from the Sundays River Poort (-73) which I think compares very favourably indeed with a large number of collections from afar afield as Redcliffe (north-east of Willowmore), the Baviaanskloof, Uniondale, down the Longkloof to Humansdorp and Hankey/Patensie. These are all collections which I have identified as H. cooperi var. gordoniana, and considered in the context of another paper.
161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
Discussion It is apparent to me that there are can be only two elements (species) of the sub-genus Haworthia present on Kaboega. These are from either of the geographical elements cymbiformis, aristata, gracilis and cooperi, and they are directly continuous. In cultivation it is apparent and obvious to me, that aristata from Spitzkop (-141) is mirrored by the gracilis-like -152, which is continuous with the more gracilis-like -151. This latter element leads through several collections to the cymbiformis-like plants in -150, -158 and -159. Similarly a very cooperi-like element in -148 is the apparent ecotype of the very gracilis-like -149. But -148 (and -165) must also be compared with -152 and to -153, which take us back to the gracilis-like archetypes. More significantly these seem to be the elements which best relate to the collections from Oudekraal, and with what occurs still further east at Zuurberg. These collections are considered to be H. cooperi var. pilifera.
There is no doubt that the Spitzkop aristata (-141) must be compared with -155 at Paddafontein and thus connecting aristata to the greater Jansenville area, and to the western elements of H. decipiens var. pringlei. There is the Waterford collection north of Lake Mentz (MBB6855-154) which is problematic as it does not have the opacity nor darker blue-green colouration of pringlei. It is better compared with xiphiophylla (or perhaps this is H. decipiens var. minor) in 72.2 south of Lake Mentz. It also bears a remarkable likeness to the Krom River collection of gracilis var. isabelllae (-39) as indicated above.
Conclusions I conclude that at Kaboega we have a situation where cooperi is excluded by the fact that the archetypal gracilis is represented by an advanced version of gracilis from which aristata and cymbiformis are extended. This pattern of identifications and classification true for one area, are not true for another. Already fully aware of the complex interaction between species like H. bolusii, H. cooperi and H. decipiens, and fast becoming even more aware of the extension of this complexity to H. gracilis, H. cymbiformis, H. arachnoidea var. xiphiophylla and even H. marumiana, I have to express conclusions very guardedly. Any classification of Haworthia will undoubtedly have tensions within it. It has been long apparent to me that sophisticated technology is unlikely to prove of much value in dealing with the nuance of variation between populations. If it is, it has going to have to first take into account the kind of variation one sees at the scale covered by this article. My belief was, and is now confirmed, that this is indeed the scale at which observation is now required. It can still be a lot closer. We did not spend sufficient time at Kaboega to explore the area thoroughly, and neither have we yet made any permanent record of our observations other than this report and accompanying illustrations. The point may now have been passed at which casual generalisation from a memory bank of images is possible. Extensive photographic and herbarium records are going to be essential to create a physical record which can be studied and manipulated. There is a series of eight mountain ranges from near the coast, with the Zuurbergs being the last of these in the north-east. If I calculate how long it would take to explore that area on the scale of our limited survey of Kaboega, I reckon on at least three years of continual search.
At this point I realise that the expectations of “Haworthia Revisited” are not going to be met. There are already snivels and meuls because there is no “data” in my revision. My experience tells me that this is not because the average reader would in fact take any cognisance of such data – but it is part of the illogical and faulty paradigm of modern “science” (“materialism”, the Theosophist would say). My conviction is that I have in my revision presented there a very comprehensive picture of the genus. This can definitely meet the time-worn wishful thought of the platitudinous foreword that “this book will stimulate/encourage/direct/guide further research”. Classification of Haworthia is not simply sorting a few single specimens as they are represented in collections or on herbarium sheets. It is trying to understand a complex system of closely interrelated and similar looking elements, as populations, which do not fit a classic and static image of a genus and so-many discrete species. My contention is that this is not only the case for Haworthia, but for many other genera too. ♦
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.144 a. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. arista, Chalumna. (H. cooperi var. doldii – ed.)144 b. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. aristata, Chalumna. (now H. cooperi var. doldii – ed.)145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.146. J.G.Marx in JDV91/15 H. cooperi var. cooperi, De Plaat, south aspect, Zuurberg.147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
M. B. Bayer, Cape Town. Ian Ritchie, Somerset East. ♦
The five papers in this book represent analyses and discussion of a large number of collections revolving principally around the problems of the Eastern Cape Haworthia species. The paper regarding H. mucronata does provide some insight into the overall problem. The citations by Von Poellnitz for varieties of H. altilinea and H. mucronata indicate just how difficult it is to differentiate between species such as H. bolusii and H. mucronata, from their respective geographic locations. In the case of H. cymbiformis var. incurvula; then in the case of H. cooperi, H. bolusii var. blackbeardiana, H. aristata and H. decipiens var. pringlei; then H. cooperi, H. gracilis and H. cymbiformis; and finally in the case of Kaboega; it is evident that a better solution can be sought.
During the course of the analyses, it became evident to me that a gracilis-like element is present in every one of the cases. It is less evident in the case of H. mucronata, because I have not presented the data and also because I have not collected sufficiently either. Certainly H. mucronata is closely allied to cooperi, cymbiformis, blackbeardiana and to decipiens. However, it is also very closely intertwined in H. arachnoidea and a lot more field observations are required to determine what its archetypal kinships may be. They may be the same as for cooperi and cymbiformis. Regardless whether or not this is so, the question still arises:- If gracilis is the archetype for so many species which the evidence leads me to suggest it is, what then is this element I call gracilis?
I have always had problems of clearly separating variants of cooperi and variants of gracilis particularly in the Baviaanskloof. Examination of the Middleton, Baviaanskranz (pringlei) and Ripon collections demonstrates the tension that exists in the classification. The two former collections seem to constitute an element which can be differentiated from blackbeardiana and from cooperi (represented mainly by its variety pilifera), albeit with difficulty. That is, it has more slender leaves than either of those species. Thus I must point out that cooperi is problematic because in my 1999 revision, the main geographic component is assigned to its varieties. The typical form of cooperi occupies a no-man’s-land between blackbeardiana and pilifera. A similar situation arises with the discarding of the name translucens for gracilis. The elements which I had previously included as typical for translucens, became the var. isabellae of gracilis. Gracilis itself came to occupy a relative no-man’s-land and possibly only truly represented at its origin at Hellspoort. These problems are discussed in some detail in Haworthia Revisited.
The fact that none of the elements discussed in the preceding papers form clear discontinuities, suggests that I need to seek a better way of expressing the situation and the problem. The Kaboega situation is particularly difficult, and the fact is that we have recognisable cymbiformis, aristata and gracilis var. isabellae, connected by an array of forms which seem to be archetypal of gracilis. At Pluto’s Vale, we have gracilis var. tenera and cymbiformis connected by a small array of a similar archetypal gracilis. In the Baviaanskloof, we have variants of gracilis, and variants of cooperi which are linked through cymbiformis-like picturata to transiens. But there do not there seem to be three discrete species any more than at Kaboega, or at Pluto’s Vale. The same is true of the Ripon (Somerset East) area. There, there are only two elements. In terms of my 1999 revision, one is cooperi (MBB6557) and the other is either blackbeardiana or pringlei (MBB6556). It cannot be both. Furthermore, these two elements seem to be ecotypes of the same species, and proven by their co-occurrence in a third collection (MBB6927) from a differing habitat.
I have suggested that a more technically orientated botanical revision would result in a reduction of the number of species.These previous papers suggest that such species reductions may even be necessary at the present level if we are to maintain coherent communication. The reference to a more technically botanical solution immediately gives me problems, because I do not from my experience see the adequatio to undertake such a task. I here try to summarise, distill and synthesise the conclusions arrived at in my other papers in my own way. The questions I ask are:-
Is there are common solution to the classification of the variants of Haworthia over the geographic areas of the middle Fish River, the upper Sundays River and the Longkloof (Couga River) and Baviaanskloof (lower Groot River and Baviaanskloof rivers).
Can any classification solution be offered which provides a better explanation for the variation and the continuities we see in the field?”.
Results The problem relating to Ripon, makes it quite obvious that we have cooperi var. cooperi, cooperi var. pilifera, cooperi var. dielsiana, bolusii var. blackbeardiana, decipiens var. pringlei and aristata involved in a common discussion.The problem at Kaboega is that we have cymbiformis, aristata and possibly gracilis as emergent elements from a core which could be gracilis. Similarly at Pluto’s Vale, tenera and incurvula emerge from a gracilis core. In the Baviaanskloof, there is a single core which leads either to gracilis, cooperi or cymbiformis variants. In nearly each case I have mentioned two key collections.These are JDV96/89 from Gladhurst, south of Adelaide, and MBB6603 from Glen Craig, north-east of Grahamstown. To this can be added the collection by Gerhard Marx in MBB6845 just east of Alicedale. These are all difficult to reconcile with my 1999 classification concepts of cooperi or gracilis. If we re-examine gracilis, we find that it is only truly represented at Hellspoort, north-east of Grahamstown. Kobus Venter collected there (JDV89/42) and again in 1988. I collected there together with him (in MBB6614) in 1966. Looking at these plants in cultivation and in the light of the classification problem, it becomes obvious that pringlei via particularly the Ripon collection MBB6556, is comparable with gracilis from Hellspoort. With this consideration in mind, it becomes evident that if the concept of cooperi is enlarged to incorporate gracilis, a radically new and more practical solution is presented. The element which I have called gracilis var. gracilis becomes cooperi var. cooperi.
The implications are as follows:
H. cooperi becomes a substantial element which absorbs all the varieties of the obsolete gracilis. The variety dielsiana is abandoned as a variant within the variety pilifera. Cooperi also absorbs pringlei from decipiens in relation only to the collection from the type locality viz. Scott in PRE8970 and two others mentioned in my revision viz. MBB&PVB6561, Baviaanskrans, Patryshoogte, and MBB&PVB6556 from Ripon, both cited under bolusii var. blackbeardiana. It should be noted that no specimen of pringlei was in fact deposited and a letter promising such a specimen occupies the herbarium file. My perception and understanding of the element is from two plants sent to J.D.Venter by Dr Reddi, and also from two plants obtained from Gerhard (J.G.) Marx, all from the ostensible type locality.
Thus cooperi would virtually comprise a superspecies which excludes cymbiformis and its varieties with some difficulty. It would also be difficult to exclude bolusii. Cooperi would in fact absorb the variety picturata of gracilis and could absorb transiens of cymbiformis. A further implication is the variety viridis of gracilis. The evidence suggests that it belongs with cooperi, but that it is interwoven with decipiens and perhaps with xyphiophylla. In this respect, there is not enough data to build a clearer picture of what occurs in the field. There is a similar problem with aristata. There can be little doubt that it is the geographic complement to gracilis and pringlei, and while it is distinctive at Kaboega, there is also the evidence there that it is continuous with cooperi and cymbiformis. There is also the question of the remaining specimens which are cited in my revision under pringlei. These are from the area around Jansenville, where we already know (Lootskloof) that bolusii var. bolusii intrudes, and where we also have a series of greenish plants (Mt. Steward area principally) with numerous erect spinose leaves rather green in colour. They differ from decipiens in having smaller narrower and elongate leaves. These elements need to be further researched and perhaps re-named. Aristata can be linked with these as it does bear a striking resemblance to the darker and bluer forms of decipiens viz. var. cyanea from south and west of Beaufort West.
Conclusion The first question can be answered with some confidence. There is a solution which can be offered for the portions of the river drainage systems mentioned above. This is to merge the concepts of cooperi var. cooperi, gracilis var. gracilis and part of decipiens var. pringlei.
The second question cannot be answered because of the scope and extent of the problem in relation to the available data. It is quite obvious that small scale collecting and observation is needed. From what has been done, it is very evident that decipiens, bolusii, cymbiformis, arachnoidea and mucronata are very poorly represented in terms of their actual variability. This is particularly true of decipiens. If one considers its geographic central role between mucronata and cooperi, it is predictable that research will disclose some very interesting variants. There are vast areas (in relation to the scale of exploration done at Pluto’s Vale and at Kaboega) which can be regarded as wholly unknown. Particularly void are the areas between Oudtshoorn and Willowmore, and the between Willowmore and Graaff Reinet and southwards to the Cockscomb. The many ranges of hills between the Longkloof and the Zuurberg must hold many populations too, while the mountains to the general north of Somerset East must do the same.
I have done quite a degree of detailed exploration in the south-western Cape which has exposed some extraordinary populations. Some of the variability and endless difference borders on the bizarre, without detracting from the beauty and fascination of the genus. Thus I do not think there is a classification system in the classic sense of dichotomy that characterises taxonomic botany. The variability within Haworthia appears to be highly reticulate and ductile. Many different solutions can possibly be provided which will all be influenced by the material available, and the form in which it is seen by the viewer.
Ironically, my definition of species is that they are systems which are genetically and morphologically continuous in space and time. In my revision I wrote that “I was trying to identify nodes in a complex interlinked web”. This may suggests that the nodes are more important than the matrix in which they occur. I also wrote “… when a variety is selected (described) the balance of variability is loaded towards the typical”. What happened with the recognition by myself of pilifera after I had in my previous Handbooks placed it in synonymy with cooperi, is that it suggested cooperi to have much more substance. Similarly, recognising varieties of translucens and abandoning the name in favour of gracilis, created a similar problem there. Varieties occur as nodes in the matrix of the parent species, just as species do in the matrix of the genus. At some point the discontinuities (in either case) may be obvious and perhaps absolute. I suggest that in the sub-genus Haworthia these continuities are largely obscure. It seems to me that we need to examine the herbarium specimens in terms of a sampling record. It is by now quite substantial, and should thus be considered as a very largely representative of the matrix of the genus. I have become increasingly confident that my classification based on that record, is a good one. Furthermore, that it is has a sufficiently predictive element to accommodate the greater number of new records that will be made.
While writing these papers, I have been carrying some ideas in the back of my mind. These relate to the species in the southern and south-western Cape. I have many new records from there as well, and I am aware of several collections which are otherwise outside of my ken. What I have seen suggests to me that it may be possible to identify the background matrix of an archetype such as I in this work suggest for the eastern Cape (viz. cooperi). ♦