25. Haworthia nortieri Smith, JS.Afr.Bot. 12:13(1946). Bayer :141(1976). Bayer :49(1982). Scott :88(1985). Type: Cape, Vanrhynsdorp, Smith 1676a (NBG): H. nortieri var. montana idem. 16:6(1950). Type: Cape, Clanwilliam, Smith 1678 (NBG): H. nortieri var. giftbergensis G.G.Sm. ibid. 16:7 (1950). Type: Cape, Vanrhynsdorp, Smith 7199 (NBG).
nortieri: for Dr. Nortier.
Rosette stemless, proliferous, 3-5cm φ. Leaves 25-45, soft sub-erect, ovate-lanceolate to obovate, pale to purplish green, with translucent spots on the leaves, small spines on margins and keel. Inflorescence slender, to 30cm. Flowers greyish-white, yellowish in tube.
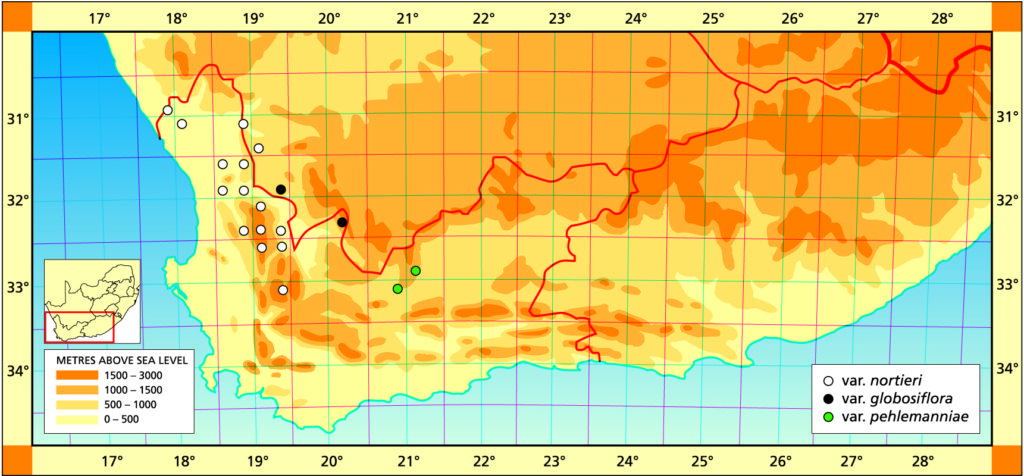
1982 – H. nortieri occurs in the area between Clanwilliam and Vanrhynsdorp, extending inland to the foot of Vanrhyns Pass and on top of the plateau of the northern Cedarberg mountains. The var. globosiflora occurs in the dryer Botterkloof area and is distinguished by the flower having a globose tube while the leaves are shorter and broader. However, the flowers are not always so inflated and plants in the Vanrhyns Pass area have normal flowers while being vegetatively similar to the var. globosiflora. This is the basis for rejecting species status for the last-named. H. nortieri occupies the mid‑western geographic locale for the genus. The distribution southwards is not known and there is a big gap between this species at Pakhuis Pass (near Clanwilliam) and a form of H. archeri var. dimorpha in the Elandskloof area southeast of Citrusdal. H. arachnoidea occurs to the north of Vanrhynsdorp and also to the north and far west of Vanrhyns Pass. It does not, however, seem to appear in the Botterkloof area. H. nortieri is distinguished largely by the opaque leaf surfaces with abrupt, ovoid, pellucid spots. The flowers, and particularly the buds, are greyish in colour but the colour inside the tube is variable.
1999 – The range of H. nortieri has been extended considerably. The reference to H. archeri var. dimorpha at Elandskloof is quite erroneous and simply arises from the difficulty in relating that collection to the nearest known species. On the otherhand, the Elandskloof plants are only known to me from herbarium record and from two living plants collected by Drs Muller-Doblies. These did not seem to unequivocally be H. nortieri. P.V. Bruyns has collected H. nortieri from as far north as the Groenriver, to south as far as Kromriver in the Cedarberg. The late Harry Hall also collected it in the northeastern Knersvlakte. The most southerly collection is from near Opdieberg, north of Ceres. These plants also resemble the var. globosiflora but the flowers are not globose. At both extremes the plants tend to resemble H. globosiflora vegetatively . The plants at Komkans tend to have globose florets, and this is also true of plants at Groenriver where the florets are short and squat. The decision to include H. pehlemanniae within this species is also on account of the flower which is identical to that of var. globosiflora. This is not only in shape, but in colour too. Although the flowers may be the usual white with greenish veins, brownish-green flowers have been observed in both elements. The reference in the original description to quadrantly as opposed to spirally arranged flowers defies plant growth principles. The distribution of the two elements is complementary and gives the species as a whole an extraordinary cosmopolitan character. The colour in the flowers of the typical variety can be remarkable and as if yellow paint had been daubed at the throat of the florets. Habitat ranges from the moist south slopes of the Cedarberg to the dry wastes of Namaqualand and the so-called Moordenaarskaroo.
a. var. nortieri The typical variety occurs over a very wide area in the Table Mountain sandstones from south-east of Citrusdal to south-west of Nieuwoudtville. In addition it extends out into the Namaqualand lowlands, into very arid conditions. It is thus a very variable taxon in its own right.
Distribution: 3017 (Hondeklipbaai): Groenriver (-DD), Bruyns 6728 (NBG). 3118(Vanrhynsdorp): Komkans (-AA), Bruyns 6146 (NBG); Klipdrif (-BB), Hall 3390 (NBG); Gifberg (-DA), Smith 7199 (BOL,NBG), Thomas in NBG626/69; Steenkampskop (-DB), Bruyns 6167 (NBG); Kobe Pass (-DB), Bruyns 6170 (NBG); Die Kom (-DC), Bayer in KG329/72 (NBG); Doornriver (-DC), Smith 1676 (NBG), Leighton (BOL); W. Doornriver (-DC), Bayer 3637 (NBG); SE. Klawer (-DC), Leipoldt 4146 (BOL), Herre in STE6695 (BOL); E. Doornriver (-DD), Smith 1676a (BOL), Smith 6212 (BOL, NBG), Esterhuysen 6008 (BOL). 3119 (Calvinia): Uitkomst (-AC), Barker 10753 (NBG); Vanrhyns Pass (-AC), Smith 6211 (NBG), Hall in NBG656/60 (NBG), Ross-Frames in NBG1200/26 (BOL). 3218 (Clanwilliam): S. Clanwilliam (-BD), Van Jaarsveld 8153 (NBG). 3219 (Wuppertal): Pakhuis Pass (-AA), Smith 1678 (NBG), ex hort Whitehill NBG68397; Waboomsriver (-AC), Henderson 2212 (NBG); Diamond Drift (-AC), Leipoldt 3107 (BOL); N. Dwarsrivier (-AC), Bruyns in Bayer 6505 (NBG); E. Dwarsriver (-AD), Bruyns in Bayer 6506 (NBG); Heksberge (-CA), Smith 6116 (NBG), Muller-Doblies 79/015 (NBG), E. Elandskloof (-CA), Esterhuysen 3987 (BOL); Sandfontein (-CB), Esterhuysen 27199a (BOL); Cedarberg (-CB), Wagener in NBG11/43 (NBG); Nuwerus (-CB), Bruyns (NBG). 3319(Worcester): NW. Ceres (-AB), Aslander 645 (NBG).
Haworthia nortieri var. nortieri JDV93/61 Pakhuis Pass. Ocuring as a solitary specimen, as seen here, or in clusters.
Haworthia nortieri var. nortieri PVB6728 Groenriver. In this collection the flowers are very short but not globose.
Haworthia nortieri var. nortieri JDV88/34 Klawer. A variety of the Western Cape sandstones – not penetrated by H. Arachnoidea.
Haworthia nortieri var. nortieri JDV88/34 Klawer. The flowers are normal for the subgenus, but usually have a bright yellow throat.
b. var. globosiflora (Smith) Bayer :119(1976). Bayer 49(1982). H. globosiflora Smith, JS.Afr.Bot. 16:11(1950). Scott :87(1985). Type: CAPE‑3119 (Calvinia): Doornbosch, N. Doorn River Bridge (‑CD), Smith 7198 (NBG).
globosiflora: rounded flowers.
This variety is not known from only the Doornbosch area south of Botterkloof, which is also relatively unexplored. It has also been collected from as far east as the Ouberg Pass, southwest of Sutherland. The illustration in Scott (:88, 1985) is not of this variety at all and is probably of H. decipiens.
Distribution: 3119 (Calvinia): Doornbosch, (‑CD), Smith 7198 (NBG); 50km N. Clanwilliam (-CD), Dyer 3750 (PRE); Botterkloof (-CD), Hall in NBG68414, Villet (BOL); Boontjiesrivier, Kansekraal (-CD), Leipoldt 4119 (BOL). 3220 (Sutherland): Ouberg Pass (-AC), Venter (NBG).
Haworthia nortieri var. globosiflora JDV93/62 Botterkloof northwest of Clanwilliam. Typically with globose flowers, there are identical forms with ordinary flowers. In the winter they often “relax” so far that they lie flat on the soil.
Haworthia nortieri var. globosiflora JDV88/40 Ouberg Pass.This variety is in more karoid dry vegetation.
c. var. pehlemanniae (Scott) Bayer comb. nov. H. pehlemanniae Scott, Cact.Succ.J.(U.S.) 54:70(1982). Scott :79(1085). Type: CAPE-3320 (Laingsburg): 5km W. of Laingsburg (-BB), Scott 7450 (PRE).
pehlemanniae: for Inge Pehlemann.
Since first collected, this variety has been found at several other localities in the close vicinity of Laingsburg, but also further north in the Moordenaarskaroo and north of Matjesfontein. It differs from the species in the absence of the translucent spots on the leaf, and the vegetative similarity to H. arachnoidea in the same area is deceptive. It does appear to favour shales in relatively exposed situations as opposed to H. arachnoidea which generally prefers cooler south slopes. The two taxa grow in very close association.
Distribution: 3221 (Merweville): Klipfontein, N. Laingsburg (-CC), Aslander 801 (NBG). 3320(Montagu): 5km SW. Laingsburg (-BB), Scott 7450 (PRE), Bayer 3906 (NBG); N. Laingsburg (-BB), Venter (NBG).
Haworthia nortieri var. pehlemanniae JDV91/47 north of Laingsburg. Southwest of Laingsburg it occurs in among small sandstone boulders, whereas elsewhere it is in shales.
Haworthia nortieri var. pehlemanniae JDV87/204 southwest of Laingsburg. Although its leaves lack the transclucent spots of var. globosiflora, the flowers are identical.
Haworthia nortieri var. pehlemanniae JDV91/47 north of Laingsburg. Probably overlooked because of its similarity to H. arachnoidea.
Barry Phipps, in an article reprinted in Haworthiad 20:61, writes that “the term species is a concept”. Donald Levins in The Origin, Expansion and Demise of Plant Species devotes a chapter to “The premise and species concepts”. There is no dearth of literature and the entire subject is indeed, as Levins suggests, a subject of “heated debate”. Levins also quotes from the literature, “… the idea of good species … an artifact of the procedures of taxonomy”, and “… our system of names appears to achieve a reality which it does not possess”. It is comforting for me to read his premise …”that the species is a dynamic entity that undergoes alterations in its gene pool, variation pattern and geographical distribution”, and his advice…”thus it is best to take a pluralistic approach to species’ passages in time, combining genetic and ecological perspectives”.
I myself am not sure if these and many similar statements really mean all that much. The range of living things is surely a reality of the human capacity to observe and rationalize to the limits of their experience and competence. They are phenomena spread in space and changing with time along with the rest of creation. We give them names in order to catalogue, describe and communicate about them. Those people, who make a point of collecting and studying study living objects, give those names and theorise and generalize about their relationships. We do not conceptualise these living things. They obviously are “kinds” and do surely have an objective reality. We may perhaps conceptualise a taxonomic system in which they are studied and classified according to some model or other. This has been an ongoing process since before Linnaeus and it is unfortunate that many writers seem to have become taxonomists with only the International Code of Nomenclature as reference. The result is a plethora of meaningless subjective names or names which can satisfy only the writer and readers who know as much or less. If we do take a pluralistic approach, recognise all the pitfalls, and base our observations on extensive observation, then surely the classification product will be satisfying to ourselves and have some value for others. This is what I have tried to achieve.
In seeking some kind of closure to my own long history as a pseudo-taxonomist and botanist writing about Haworthia, I am going to write about my observations on a number of populations which I attribute to the single species Haworthia nortieri. The newest trend seems to be to term these discrete, isolated, scattered populations as “metapopulations” and theorise about the interchange of genetic material among them and the effect this has on real differences; ultimately reflecting on evolutionary passage. Whatever is attempted or intended, an accurate account of one’s personal experience must surely be of value.
H. nortieri was first noted by Dr. Nortier of Clanwilliam and then collected at the old Doorn River Bridge 45km northwest of Clanwilliam in 1937 and described by G.G. Smith in 1946. Smith also collected a similar plant on the Pakhuis Pass also in 1937, and described this as the var. montana only in 1946. In 1948 he again collected a similar element on the Gifberg Mountain near VanRhynsdorp, which he described in 1950 as var. gifbergensis. In his collecting record, Smith records, also in 1948, the collection of a plant from Doornbosch northeast of Clanwilliam, and described that as H. globosiflora. In the description the collection is attributed to J. Thudichum.
History Thus in 1969 when I started the physical process of arranging the Compton Herbarium records, these four herbarium specimens virtually constituted the total record of the species. My views of the species and its taxonomic record in my 1982 synopsis and 1999 revision were as follows:-
“25. Haworthia nortieri Smith, JS.Afr.Bot. 12:13(1946). Bayer :141(1976). Bayer :49(1982). Scott :88(1985). Type: Cape, Vanrhynsdorp, Smith 1676a (NBG): H. nortieri var. montana idem. 16:6(1950). Type: Cape, Clanwilliam, Smith 1678 (NBG): H. nortieri var. gifbergensis idem. 16:7(1950). Type: Cape, Vanrhynsdorp, Smith 7199 (NBG).
nortieri: for Dr. Nortier.
Rosette stemless, proliferous, 3-5cm φ. Leaves 25-45, soft sub-erect, ovate-lanceolate to obovate, pale to purplish green, with translucent spots on the leaves, small spines on margins and keel. Inflorescence slender, to 30cm. Flowers greyish-white, yellowish in tube.
1982- “H. nortieri occurs in the area between Clanwilliam and Vanrhynsdorp, extending inland to the foot of VanRhyns Pass and on top of the plateau of the northern Cedarberg mountains. The var. globosiflora occurs in the dryer Botterkloof area and is distinguished by the flower having a globose tube while the leaves are shorter and broader. However, the flowers are not always so inflated and plants in the VanRhyns Pass area have normal flowers while being vegetatively similar to the var. globosiflora. This is the basis for rejecting species status for the last-named. H. nortieri occupies the mid‑western geographic locale for the genus. The distribution southwards is not known and there is a big gap between this species at Pakhuis Pass (near Clanwilliam) and a form of H. archeri var. dimorpha in the Elandskloof area southeast of Citrusdal. H. arachnoidea occurs to the north of VanRhynsdorp and also to the north and far west of VanRhyns Pass. It does not, however, seem to appear in the Botterkloof area. H. nortieri is distinguished largely by the opaque leaf surfaces with abrupt, ovoid, pellucid spots. The flowers, and particularly the buds, are greyish in colour but the colour inside the tube is variable.”
1999- “The range of H. nortieri has been extended considerably. The reference to H. archeri var. dimorpha at Elandskloof is quite erroneous and simply arises from the difficulty in relating that collection to the nearest known species. On the other hand, the Elandskloof plants are only known to me from herbarium record and from two living plants collected by Drs Muller-Doblies. These did not seem to unequivocally be H. nortieri. P.V. Bruyns has collected H. nortieri from as far north as the Groenriver, to south as far as Krommeriver in the Cedarberg. The late Harry Hall also collected it in the northeastern Knersvlakte. The most southerly collection is from near Opdieberg, north of Ceres. These plants also resemble the var. globosiflora but the flowers are not globose. At both extremes the plants tend to resemble H. globosiflora vegetatively. The var. nortieri is probably best considered as the more slender-leaved and less robust sandstone ecotype of the species. The plants at Komkans tend to have globose florets, and this is also true of plants at Groenriver where the florets are short and squat. The decision to include H. pehlemanniae within this species is also on account of the flower which is identical to that of var. globosiflora. This is not only in shape, but in colour too. Although the flowers may be the usual white with greenish veins, brownish-green flowers have been observed in both elements. The reference in the original description to quadrantly as opposed to spirally arranged flowers defies plant growth principles. The distribution of the two elements is complementary and gives the species as a whole an extraordinary cosmopolitan character. The colour in the flowers of the typical variety can be remarkable and as if yellow paint had been daubed at the throat of the florets. Habitat ranges from the moist south slopes of the Cedarberg to the dry wastes of Namaqualand and the so-called Moordenaarskaroo.
a. var. nortieri
The typical variety occurs over a very wide area in the Table Mountain sandstones from south-east of Citrusdal to south-west of Nieuwoudtville. In addition it extends out into the Namaqualand lowlands, into very arid conditions. It is thus a very variable taxon in its own right.
Distribution
3017(Hondeklipbaai): Groenriver (-DD), Bruyns 6728 (NBG). 3118(VanRhynsdorp): Komkans (-AA), Bruyns 6146 (NBG); Klipdrif (-BB), Hall 3390 (NBG); Gifberg (-DA), Smith 7199 (BOL,NBG), Thomas in NBG626/69; Steenkampskop (-DB), Bruyns 6167 (NBG); Kobe Pass (-DB), Bruyns 6170 (NBG); Die Kom (-DC), Bayer in KG329/72 (NBG); Doornriver (-DC), Smith 1676 (NBG), Leighton (BOL); W. Doornriver (-DC), Bayer 3637 (NBG); SE. Klawer (-DC), Leipoldt 4146 (BOL), Herre in STE6695 (BOL); E. Doornriver (-DD), Smith 1676a (BOL), Smith 6212 (BOL,NBG), Esterhuysen 6008 (BOL). 3119(Calvinia): Uitkomst (-AC), Barker 10753 (NBG); Vanrhyns Pass (-AC), Smith 6211 (NBG), Hall in NBG656/60 (NBG), Ross-Frames in NBG1200/26 (BOL). 3218(Clanwilliam): S. Clanwilliam (-BD), Van Jaarsveld 8153 (NBG). 3219(Wuppertal): Pakhuis Pass (-AA), Smith 1678 (NBG), ex hort Whitehill NBG68397; Waboomsriver (-AC), Henderson 2212 (NBG); Diamond Drift (-AC), Leipoldt 3107 (BOL); N. Dwarsrivier (-AC), Bruyns in Bayer 6505 (NBG); E. Dwarsriver (-AD), Bruyns in Bayer 6506 (NBG); Heksberge (-CA), Smith 6116 (NBG), Muller-Doblies 79/015 (NBG), E. Elandskloof (-CA), Esterhuysen 3987 (BOL); Sandfontein (-CB), Esterhuysen 27199a (BOL); Cedarberg (-CB), Wagener in NBG11/43 (NBG); Nuwerus (-CB), Bruyns (NBG). 3319(Worcester): NW. Ceres (-AB), Aslander 645 (NBG).
b. var. globosiflora (Smith) Bayer :119(1976). Bayer 49(1982). H. globosiflora Smith, JS.Afr.Bot. 16:11(1950). Scott :87(1985). Type: CAPE‑3119(Calvinia): Doornbosch, N. Doorn River Bridge (‑CD), Smith 7198 (NBG).
globosiflora: rounded flowers.
This variety is not known from only the Doornbosch area south of Botterkloof, which is also relatively unexplored. It has also been collected from as far east as the Ouberg Pass, southwest of Sutherland. The illustration in Scott (:88, 1985) is not of this variety at all and is probably of H. decipiens. Like the var. pehlemanniae, the florets are remarkably globose with a significantly shortened tube, and the two varieties cannot be separated on this score. The flowers of both may be uniformly greyish-green or brownish. Vegetatively, the leaves in the var. globosiflora are distinctively dotted with translucent spots which are absent in var. pehlemanniae.
Distribution
3119(Calvinia): Doornbosch, (‑CD), Smith 7198 (NBG); 50km N. Clanwilliam (-CD), Dyer 3750 (PRE); Botterkloof (-CD), Hall in NBG68414, Villet (BOL); Boontjiesrivier, Kansekraal (-CD), Leipoldt 4119 (BOL). 3220(Sutherland): Ouberg Pass (-AC), Venter 88/40 (NBG).
c. var pehlemanniae (Scott) Bayer comb.nov. H. pehlemanniae Scott, Cact.Succ.J.(U.S.) 54:70(1982). Scott :79(1085). Type: CAPE-3320(Laingsburg): 5km W. of Laingsburg (-BB), Scott 7450 (PRE).
pehlemanniae: for Inge Pehlemann.
Since first collected, this variety has been found at several other localities in the close vicinity of Laingsburg, but also further north in the Moordenaarskaroo and north of Matjesfontein. It differs from the species in the absence of the translucent spots on the leaf, and the vegetative similarity to H. arachnoidea in the same area is deceptive. However, the leaves have a slightly more greyish character than in H. arachnoidea. It does appear to favour shales in relatively exposed situations as opposed to H. arachnoidea which generally prefers cooler south slopes. The two taxa grow in very close association.
Distribution
3221(Merweville): Klipfontein, N. Laingsburg (-CC), Aslander 801 (NBG). 3320(Montagu): 5km SW. Laingsburg (-BB), Scott 7450 (PRE), Bayer 3906 (NBG); N. Laingsburg (-BB), Venter (NBG).
During the last few years several names have been published which in my opinion belong in the context of the above The first of these is H. agnis L. Battista, Alsterworthia International 2(2):8-9, the second H. devriesiiI. Breuer, Avonia21:3 (2003) and the third H. albispina M. Hayashi, Haworthia Study 9:2 (2002). These are all significant records and I do not question the fact that the populations are distinctive and important variants. I simply add that in terms of an objective view of an observer who wishes to understand these plants in the total context of the flora of the region and at another level in the context of the genus Haworthia and its three subgenera, they belong in the one species H. nortieri. For the collector I would suggest that someone make the formal taxonomic statement that these are varieties. The types are deposited who actually knows where, and being outside of the classic regional herbaria they have also not been assessed by the authors of the names, in terms of the total herbarium record. This disregard for rational herbarium practise does, and will continue to, make it difficult to technically formalise a classification.
In referring to the regional flora, I want to explain my viewpoint that botanical classification belongs in the realm of true biology and must be meaningful in terms of all life forms and the conditions which gave rise to their origins and which sustain them. Thus my experience was that the botanical community were defining a Cape Flora based only on the machia-fynbos vegetation peculiar to the predominent sandstone geology of the area. My view was that this was a misinterpretation and that whatever the origins of the fynbos was, there was in present time a Winter Rainfall Biome which included the karoid vegetation of the shales, tillites and igneous rock within and adjacent to the so-called Cape. Species systems, like H. nortieri, would only be understood is this massive impact of geology, skeletal soils and winter rainfall was properly acknowledged.
In a manuscript concerning H. pygmaea and a broader species concept for the genus, I have tried to explain that the reason that I have been drawn into the realm of formal taxonomy, is precisely because there are no reliable and infallible criteria or characters by which the species and the variants van be positively described and identified. The observer is totally dependent on imagery and by the experience garnered in the field and in cultivation. The classification thus can only be assessed in terms of comparable experience and motive.
I have avoided trying to rationalise the morphological differences between H. arachnoidea and H. nortieri because I have found them myself to be a bit obscure. Generally the colour of the leaves in arachnoidea is a darker uniform green as opposed to a pale and often maculate condition in nortieri. Where nortieri does seem to differ is that the mid- to end section of the upper leaf surface tends to be convex. This convexity of the upper leaf surface is reminiscent of the similar characteristic of the species of the southwestern Cape where a retuse truncated end-area often occurs, and may thus account for the truncation in the leaves that characterises some nortieri populations. I have observed this truncation as a response to growing conditions as well, notably in the specimen from the Blinkberg which Ernst van Jaarsveld grew in cultivation as compared with plants observed by myself in the field (see pictures). Similarly plants from Opdieberg collected and grown by Etwin Aslander seemed to have very truncated leaves under his growing conditions. Plants in the field at Arizona north of VanRhynsdorp were very retracted into the soil and the leaves also correspondingly truncated. Alternatively there are populations in which the leaves are attenuate, longer and pointed, bringing me back to an observation I would like to make that such morphological characters and differences are too often used to justify species differences when the variation is in fact an expression of the nature of the species. There are several populations in the greater Touws River area which are doubtful and a good deal of tough fieldwork will be necessary to get a better understanding of these disparate populations which could include both H. maculata as it occurs on and adjacent to the Audensberg at Worcester, as well as H. marumiana var. dimorpha east of Touws River
The variant agnis has been collected twice by Etwin Aslander both east of Nuwerus and at Middelpos near Bitterfontein. It has also been collected by Philip Desmet at Nuwerus. Both collectors observed the presence of H. arachnoidea at Nuwerus and Desmet submitted specimens of a putative hybrid. These plants are indeed small and Dr Ben Zonneveld (private communication) who has researched the mass of total nuclear DNA in many Haworthia specimens maintains that his figures suggest that it is a discrete species. I do not think that his argumentation or his methodology are substantial or convincing and that these kinds of peripheral data sets should be use in taxonomic decision making. In my opinion they could be used to describe variation within a species system more usefully than to support a rickety overburdened classification tree (cladogram). I should add that the Bruyns collection from Komkans, which is west of Nuwerus, is also of small plants and is actually a replication of a collection made by Rolf Rawe in 1969 also at Komkans. That collection was never formally deposited as a herbarium record but I remember it distinctly as there were many plants in the San Merino Nursery at Somerset West closely following the closure of the notable Bernard Carp nursery at Hout Bay.
The variant devriesii is indeed odd as it is from very far east for the species north of Prince Albert. I will show by the illustrations how it relates to H. nortieri by similar ecotypes from the karoid landscapes peripheral to the machia-fynbos of the sandstones where the species is better known. Like albispina, which is from Koup Station, it should simply be regarded as a variant of the existing var. pehlemanniae, although I do not believe that formalizing varieties really is very helpful in the botanical sense. The variation is, in my opinion, largely driven by the nature of skeletal substrate and is not conformable with any classification system that can account for the totality of variation in the species. Both the names as representing formal taxa, will be confounded by complete and true description of local variation within the actual populations the types were sampled from, and further so by samples from populations known to the authors of the names. Thus we have populations in among those cited in my revision as well as new records, particularly from northeast of Nieuwoudtville, and from N VanRhynsdorp, which need to be offset against these “new species”. There are also several new records of my own (Sneeuberg and Katbakkies) and by Ernst van Jaarsveld (Blinkberg, Cedarberg) and by Adam Harrower (Breekkranz, Cedarberg) that add to and extend the already wide variation of the sandstone components of the species. Bruyns (private communication) also reports the species from the Elandsberg in the Tanqua Karoo. Adding new names requires that one cast ones mind back to the situation prior to the revisions by either Col. Scott or myself and question whether these new names require the re-instatement of so many other names put into synonymy in those revisions. At the same time some note must be taken of the degree of exploration and how predictive any new name is likely to be.
Regarding flower characteristics and flowering time, my observation in other genera is that it often appears that variation between species samples is that the observed variation in selected characters, may within a species system exceed that observed between different species systems. I stress the word ‘observed’ because any opinion or judgement can only be made in terms of the sample and information available and utilized by the observer. Thus I concluded that the var. globosiflora belongs in the nortieri system by virtue of populations apparently in the same geographic and floristic range in which the flowers are neither globose nor normal for nortieri. The variant agnis for example has smaller globose flowers, the flowers of pehlemanniae cannot be distinguished from those of globosiflora, while in devriesii the curious conclusion is drawn that the less globose flower suggests an affinity with H. decipiens var cyanea. G.G. Smith was struck by the golden yellow colour in the flower throats of his original nortieri collection, and noted the canary yellow flowers of his var giftbergensis and the yellow-green in the case of var montana. My observation generally is that throat colour is quite variable and cannot be used to rationally separate out lesser taxonomic ranks in the sense of a formal biological and meaningful classification.
Flowering time is often used as in defining species systems. Here again the irony seems to be that variation in flowering time of nortieri is, just as the case of variation in flower morphology, characteristic of the system. Generally flowering time for Haworthia populations covers approximately a minimum six-week window. Different species systems can be found to be in flower virtually throughout the year. In H. cooperi var gordoniana this is late spring to early summer and I have observed that annual climatic conditions may shift this window by as much as six weeks. In the case of the greater H. pygmaea system flowering has been observed to vacillate between an early spring and a late spring flowering. In the Subgenus Robustipedunculares, H. minima populations can differ in a flowering time of late spring or mid-summer. I observe that in nortieri, there are populations in the drier northwest (including agnis) that flower early in spring while in the Cedarberg, flowering maybe as late as summer (December).
Gerhard Marx wrote a good account of albispina and H. arachnoidea var. scabrispina growing together at Koup Station (Aloe 43:6, 2006) in which he notes that the flowering time of the former precedes that of the latter. This is what I also observed in the case of the same two elements growing in close association both north and southwest of Laingsburg. This is also the case at Nuweplaas northeast of Nieuwoudtville where a globosiflora-like element is in close association with H. arachnoidea. It had set and shed seed, while the latter was still flowering.
To be realistic one needs to now consider that we are discussing only about 42 records for an area extending from Groenriver and Bitterfontein in the northwest, down to east of Citrusdal in the south and then eastwards to as far as Prince Albert. This is an enormous area and the actual sample size must be very small in relation to actual occurrence of the species. However, it seems unlikely now that the range is likely to be extended, and that we have a fairly good representation of what the species looks like over its whole range. Further exploration will prove very time consuming especially in the northeastern Cedarberg which is rugged country and largely inaccessible.
Records, collections and illustrations
1. PVB6728 Groenriver (left). Groenriver is in Namaqualandand and inland from Hondeklipbaai. It is northwest of any other known H. nortieri populations. I am not familiar with the area and have not seen the plants. H. arachnoidea is recorded from various places in N Namaqualand but I am also not knowledgeable about its true distribution in that area. There is also a collection PV76745 (right) also from Groenriver that seems to be H. arachnoidea. It would be nice to know more about both these collections.
1. PVB6728 H. nortieri. Groenriver.1b. PVB6745.1 H. nortieri. Groenriver.
2. EA1497 Middelpos, Bitterfontein. Inland from Hondeklipbaai and north of Nuwerus. These are small plants and I also have not seen the population. The flowers are small and tend to globose.
2. EA1497.1 H. nortieri ‘agnis’. Middelpos, Bitterfontein.2. EA1497.2 H. nortieri ‘agnis’. Middelpos, Bitterfontein
3. PVB6146 Komkans. West of Nuwerus. Low lying area and the plants are in among white quartz pebbles. This was probably where Rolf Rawe collected this as a new species which was distributed by San Marino Nursery in 1969.
3. PVB6146.1 H. nortieri ‘agnis’. Komkans.3. PVB6146.2 H.nortieri ‘agnis’. Komkans.3. PVB6146.5 H. nortieri ‘agnis’. Komkans.3. PVB6146.4 H. nortieri ‘agnis’. Komkans.3. PVB6146.3 H. nortieri ‘agnis’ Komkans.
4. EA1442 and EA1443 Kareeberg Nuwerus. This is a low range of hills south of the Khamiesberg. Ewin Aslander also informed me that H. arachnoidea was present with nortieri, and both his collections are of very small plants. JDV95/11 and 95/12 ex EA H. nortieri ‘agnis’. Kareeberg, Nuwerus
4. EA1442.1 H. nortieri ‘agnis’. Kareeberg, Nuwerus.4. EA1443.1 H. arachnoidea. Nuwerus. H. nortieri ‘agnis’. Nuwerus.4. JDV99-12.1 H. nortieri ‘agnis’. Nuwerus.4. JDV99-12.2 H. nortieri ‘agnis’. Numerus.4. JDV95.11.1 ex EA H. nortieri ‘agnis’. Kareeberg, Nuwerus.
5. IDC53 Nuwerus. This collection was sent to me as the putative hybrid H. nortieriXarachnoidea. These were two very small plants under 50mm diam. And differ very slightly from Etwin Aslander’s arachnoidea plants from the same area, except that they seem to be marked with a faint reticulation. I am not sure they are hybrids and in fact do not know if H. nortieri hybridizes with any other species.
5. IDC53.1 H. nortieriXarachnoidea. Nuwerus5. IDC53.2 H. nortieriXarachnoidea. Nuwerus
6. MBB7522. Arizona. This locality is north of VanRhynsdorp on the Knersvlakte on the farm Arizona which now belongs to Western Cape Nature Conservation Board. This is south of Nuwerus and the substrate is shale. The species is also present on the adjacent Kwaggaskop farm belonging to Buys Wiese the well-known nurseryman. The plants are very truncated as the vegetation is sparse and low. The plants were recorded by Annelise leRoux and Steven Hammer who told me of it, and we were taken there by very kind favour of Kobus Kritzinger of Western Cape Nature Conservation Board. *** I have confused the true identity of H. agnis in the above 4 populations and it apparently is not the small plants in the Nuwerus area that I garnered from the place name given in the description, but simply the larger version at Arizona. That is an even stronger contender for inclusion in H. nortieri.
6. 7522.1 H. nortieri. Arizona.6. 7522.2 H. nortieri. Arizona6. 7522.6 H. nortieri. Arizona.6. 7522.9 H. nortieri. Arizona.6. 7522.10 H. nortieri. Arizona.6. 7522.11 H. nortieri. Arizona.6. 7522.12 H. nortieri. Arizona.6. 7522.13 H. nortieri. Arizona.
7. MBB7578. Nuweplaas P.V. Bruyns reported H. nortieri northeast of Nieuwoudtville. We visited the area in early summer and first found H. arachnoidea and then H. nortieri. Nuweplaas is in the extreme southwestern Bushmanland and Aloe dichotoma is virtually at the extreme of its distribution here. The substrate is Dwyka Tillite and the vegetation is Succulent Karoo. There are species like Aloe krapholiana and Euphorbia cylindrical on the farm as well so there is an unusual element in the vegetation. The plants were extremely well-hidden and the plants must be able to tolerate very dry and hot conditions here.
7. 7578 rt, 7577 lt. nortieri cf arachnoidea. Nuweplaas.7. 7578.1 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.4 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.7 H. nortieri. Nuweplaas, Nieuwoudtville.7. 7578.10 H. nortieri. Nuweplaas, Nieuwoudtville.
8. PVB7167. Steenkampskop. This is the mountain between VanRhynsdorp and VanRhyns’ Pass. The plants are very similar to those at the foot of the pass and generally fairly similar to var. globosiflora, as indeed are those at Arizona and Nuweplaas. The flowers, however, are not globose.
8. PVB6167.1 H.nortieri. Steenkampskop, Vanrhynsdorp.8. PVB6167.2 H. nortieri. Steenkampskop, Vanrhynsdorp.8. PVB6167.3 H. nortieri. Steenkampskop, VanRhynsdorp.
9. MBB7523. Gifberg. This is the first higher altitude record southward from VanRhynsdorp at nearly 650m. The substrate is sandstone on the northwestern corner of this geological formation. The plants are smaller and with more lanceolate leaves than other records. The species has been recorded at Sandfontein a little to the southeast and also at the Koebee Pass to the east.
9. 7523.2 H. nortieri. Gifberg.9. 7523.3 H. nortieri. Gifberg.9. 7523.4 H. nortieri. Gifberg.9. 7523.8 H. nortieri. Gifberg.9. 7523.9 H. nortieri. Gifberg.9. 7523.10 H. nortieri. Gifberg.9. 7523.11 H. nortieri. Gifberg.
10. MBB7524. N. Clanwilliam. The var. montana was described from the Pakhuis Pass and it is not clear exactly where this is on the Pass. The eastern part of the Pass is dry Renosterveld and there is no confirmed record of the species from there to Doringbos at the northern foot of the Pass where var. globosiflora occurs. Derek Tribble described this locality on the western end of the pass, just north of Clanwilliam. At about 300m above sea level it is not really high in respect of the Cedarberg. The plants occur rather sparsely and are fairly solitary. Leaf shape varies from relatively abbreviated to more lanceolate and one plant particularly had leaves with linear translucent markings rather than rounded. It is generally difficult to make these observations under field conditions as the plants are well hidden and often withdrawn into the soil.
10. 7524.1 H. nortieri. N. Clanwilliam.10. 7524.3 H. nortieri. N. Clanwilliam.10. 7524.5 H. nortieri. N. Clanwilliam.10. 7524.6 H. nortieri. N Clanwilliam.10. 7524.7 H. nortieri. N Clanwilliam.10. 7524.8 H. nortieri. N. Clanwilliam.
11. MBB7579. EA1441 (top two). JDV99-11. Trawal Bridge. The species was first described from a collection made near the Bulkshoek Barage between Clawilliam and Klawer. I have seen it there, at the Doorn River Bridge and also on the northern rocky banks of the Olifant’s River near there. At the Trawal Bridge the plants are on a low lying very exposed and hot sandstone ridge. Note the flower photographed and how short the free parts of the petals are.
11. 7579.01 H. nortieri. Trawal Bridge.7579 H. nortieri. Trawal Bridge.11. EAs1441.1 H. nortieri. Trawal.11. EAs1441.2 H. nortieri. Trawal.11. EAs1441.3 H. nortieri. Trawal.11. EAs1441.4 H. nortieri. Trawal.
12. MBB7525. S. Clanwilliam. Whereas the other habitats have been out of or marginal to Fynbos Vegetation, this is true Fynbos at this site. It is west facing and very dry in summer. As the lichen growth shows, the winter conditions will be quite moist. There are several records south of this towards and southeast of Citrusdal which I have not been able to confirm. JDV99-11 H. nortieri. Trawal.
12. JDV99-11 H. nortieri. Trawal.
13. MBB6505 and 7595. N. Dwarsrivier. This is in the Central Cedarberg at about 800m. The plants are on large loose boulders lying in the valley floor. When I first saw the plants I was very impressed with the number of leaves in the rosettes and their attenuation. These impressions can be very unreliable. I still have a large specimen in cultivation but revisited the site recently to establish that the plants are within “the normal range of variation of the species”. I write this rather facetiously because it is actually very difficult to obtain a true impression and my long experience of plant descriptions and identifications suggests to me that taxonomy is characterised by such subjective judgements.
13. 6505.2 H. nortier. N Dawrsrivier, Cedarberg.13. 6505.3 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.4 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.5 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.6 H. nortieri. N Dwarsrivier, Cedarberg.
14. MBB6506. East Dwarsrivier. This collection first came to my notice as a plant said to have been collected by Clive McDowell at the Wolfberg Cracks in the Cedarberg. The plant was H. reticulata and hugely improbable. Fortunately I was able to vist the area. The plants are on a steep south facing riverine rockface and are H. nortieri. They are proliferous and the leaves are shorter and broader than is normally the case.
14. 6506.1 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.2 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.3 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.f1 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.f2 H. nortieri. Dwarsrivier, Cedarberg.
15. ADH2514. Breekkranz. This is a little further south. There is a hiking trail from east of Citrusdal towards Kromrivier. The collector was Adam Harrower and the single plant in cultivation seemed rather robust and thick-leaved. We visited the area and while not reaching the place along the Breekkranz trail indicated by Adam Harrower, we found the plants in the rocks around the Bakkamerfontein ruin. They were abundant and well hidden in rock crevices at about 800m.
15. ADH2514 H. nortieri. Breekkranz, Cedarberg.
16. MBB7580. Bakkamerfontein. The nortieri plants are just at the small distant upper notch on the skyline.
Blinkberg EvJ19550 is taken from just below and looking about 30km to the Sneeuberg (not Sneeukop) in the distance. Only four other populations were recorded in the scanned landscape! The view is just as impressive southwards where three populations would be covered. It is all Table Mountain sandstone – except the Blinkberg itself. Seen in that context, the pygmaea issue is just a small blob in a much more diverse landscape. Photos take at about 10:00am.
16. 7580.1 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.4 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.7 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.11 H. nortieri. Bakkamerfontein, Krommerivier.16. 7580.13 H. nortieri. Bakkamerfontein, Krommerivier.
17a. EvJ19550. Blinkberg. Ernst van Jaarsveld has been very helpful in recording interesting populations from inaccessible places. The plant he showed me was also very robust and similar to Adam Harrower’s Beekkranz specimen and I felt it was necessary to visit the site and the plants in habitat.
The nortieri plants are just at the small distant upper notch in the sky-line.EvJ19550.a Blinkberg.
The locality is nearly 30km southeast of Kromriver and moving out of the Cedarberg into a mountaineous area refrred to as the Swartruggens. The geology of the area is fascinating. The north/south road between ceres and Wupperthal runs along the intersection of table Mountain Sandstone to the west and the younger Bokkeveld Shale to the east. The layered rocks is tilted up in the west and down in the east . Because there are harder quartzitic bands in the Bokkeveld formation, the terrain is stepped and one climbs a series on near switchbacks to get to the steep upper more quartzitic Witteberg sandstone which caps the mountain. The Bokkevelf shale here is not rich in succulents and the Haworthia was in a lower band of Dry Mountain Renosterveld at about 850m. The plants were small, well hidden under grass and small bushes and unlike the specimen in cultivation. Again it is impossible to gain an overview of the variability, but I photographed three specimens to show that the leaves can vary from spotted with the translucent dots typical of H. nortieri, or none. The plants here are non-proliferous. The landscape scene among the pictures is a view towards the Central Cedarberg and the pointed peak in the middle distance is Cedarberg Sneeuberg at 2027m. It is equally impressive looking south. It is a rockbound landscape which is difficult to move through and it would require a most remarkable effort to get a better idea of the occurrence and distribution of the plants in that whole area. As it is, we have no records for a huge area extending from Blinkberg in the south to the Pakhuis pass in the northwest, and to the Ouberg Pass in the east near Sutherland. Mrs. J. Marais of Mount Ceder Resort (Blinkberg) reports the plants at a place east of Matjesrivier to the north. H. arachnoidea is not known in the area at all but it does occur as a small densely white spinose form at the SkitteryKloof to the southeast. Curiously the only other record of Haworthia for this area is of a plant like H. venosa var. granulata collected by Elsie Esterhuyzen on top of the Skurweberg Sneekop to the south of Blinkberg. I did climb that peak in 1973 in the hopes of finding the plant, but was unsuccessful.
17. EvJ19550.2 H. nortieri. Blinkberg.17. EvJ19550.3 H. nortieri. Blinkberg.17. EvJ19550.4 H. nortieri. Blinkberg.17. EvJ19550.5 H. nortieri. Blinkberg.EvJ19550 H. nortieri. Blinkberg.17. EvJ19550a H. nortieri. Blinkberg, Cedarberg.17. EvJ19550.7 H. nortieri. Blinkberg.17. EvJ19550.8 H. nortieri. Blinkberg.17. EvJ19550.9. H. nortieri. Blinkberg.EvJ19550.6 H. nortieri. Blinkberg.
17b. MBB7597 & 7599. Kunje, SE Citrusdal. The German team Muller-Doblies showed me a plant from this area. There are other herbarium records from the Waboomsberg further north, but also from the Elandskloof slightly to the west and from the Heksberg to the near south west. The plants are abundant on flat rock sheets where there are small pockets of soil and sparse vegetation. This gets extremely dry in summer while in winter it can be very wet. The plants can be proliferous and there were some very dense localized aggregations of individuals. Size varied quite considerably from small exposed specimens 30mm in diameter, to shade protected plants to 100mm diameter. Flowering seemed to have occurred in late November to December.
17b. Terrain south of Kunje.17b. 7597.1 H. nortieri. Kunje.17b. 7597.2 H. nortieri. Kunje.17b. 7597.3 H. nortieri. Kunje.17b. 7597.4 H. nortieri. Kunje.17b. 7597.5 H. nortieri. Kunje.17b. 7597.6 H. nortieri. Kunje.17b. 7597.7 H. nortieri. Kunje.17b. 7597.8 H. nortieri. Kunje.17b. 7597.9 H. nortieri. Kunje.17b. 7597.10 H. nortieri. Kunje.17b. 7597.11 H. nortieri. Kunje.17b. 7597.12 H. nortieri. Kunje.17b. 7599.1 H. nortieri. S Kunje.17b. 7599.2 H. nortieri. S Kunje.17b. 7599.4 H. nortieri. S Kunje.17b. 7599.5 H. nortieri. S Kunje.17b. 7599.6 H. nortieri. S Kunje.17b. 7599.7 H. nortieri. S Kunje.
18. MBB6158 Skurweberg, Sneeukop. We found these plants in grass in cracks in massive sandstone slabs at the foot of the higher peaks. We also saw them in similar but dryer conditions in the Witteberg formation east of the Katbakkies Pass and west of Skittery Kloof. The whole area seems to provide endless suitable looking habitat and it is surprising that there are not more records, except that people who move around these mountaineous areas are seldom focused on small succulents.
18. 6158.4 H. nortieri. Sneeukop, N Ceres.
19. JDV96-56. EA801. Nevertheless, it is really strange that we have to move nearly 30km south to OpdieBerg to where Etwin Aslander found H. nortieri again. Grown by Etwin, the plants were extraordinarily beautiful being highly colourful and well spotted and very reminiscent of the var. globosiflora except again for the flowers. Again the area is very awesomely rugged and even here one wonders what could have motivated Etwin to have even searched there.
19. JDV96-56.1 H. nortieri. Opdieberg, Ceres.
20. MBB6815 Audensberg and 7526 Die Nekkies (bottom left). I include these two collections in the discussion because they reflect the greater problem in Haworthia, which is to establish the relationship of the different species systems. H. maculata is essentially linked to H. herbacea and H. reticulata, in the Worcester vicinity and well south of OpdieBerg. The two collections illustrated here are from two different biogeographic zones on either side of the karoid zone occupied by H. herbacea. They could thus be as genetically distant from one another as they are from that species. The Die nekkies plant has a browner colouration than the Audensberg plants which can have the purplish colouration of H.nortieri. The flower seems convincing in that the Audensberg plant has the typical fish-tail buds of the southern Haworthia.
20. 6815.1 H. maculata. Audensberg.
21. EA1508 Hex River Pass. This is again a notable collection in an inauspicious looking area. It is in the eastern leg of the Cape fold-mountains in the eastern end of the Hex River valley. The plants have to be considered in the same light as the previous comment. H. arachnoidea is present to the southwest as a very spotted form which could suggest nortieri-like character as well as confounding the situation with regard to H. marumianai var dimorpha. There are two other questionable populations. One is at Sandhills to the west which is a weakly spinose form of H. arachnoidea and then one in the Rabiesberg to the southwest which suggests an affinity with H. herbacea. What this exposes is the paucity of information. There simply are not enough known populations to help any analysis of either the morphology (as non-decisive as this could be) or the biogeography of the species, which is the main ground on which I base my classification. The nearest known H. nortieri population is that at OpdieBerg in the west, Sneeuberg in the north and then var. pehlemanniae at Matjesfontein in the east.
21. EA1508.1 H. nortieri. Hex River Pass.
22. MBB7070. Laingsburg. My few illustrations of H. nortieri var pehlemanniae reflect how little is actually known about the real distribution of many of the species. I have no doubts about the continuity of this element with var. globosiflora in the northwest. It is known at the OubergPass near Sutherland in close proximity to H. arachnoidea. There is the unconfirmed Bruyns’ record from the Elandsberg, Etwin Aslander’s at Klipfontein in the Moordenaarskaroo north of Laingsburg, this collection from just north of Laingsburg, records from just north of Matjesfonein and the original location southwest of Laingsburg on the Agter-Witteberg road.
22. 7070.1 H. nortieri var pehlemanniae. N Laingsburg.
23. MBB7594 Koup. Hayashi must perhaps be commended for recognizing this as different from H. arachnoideai var scabrispina which grows adjacently in very karoid conditions. Both species of course also grow in proximity at Laingsburg. Superficially they could be taken to be the same when in fact the spination and colouration is really different as should be noticable from the illustrations where the origins of the name “albispina” is obvious. Gerhard Marx has written of these populations in Aloe 43:6 (2006) and also illustrates “albispina” from a little further west.
23. MBBsn. H. arachnoidea var scabrispina. Koup.
24. MBB7385. Prince Albert. In the company of Sue and Richard Dean I quite fortuitously found this population discovered by V. DeVries at Prince Albert. It is very far east for the species, but my contention is that species have range and variation. They need to be considered as systems not as single specimens which look a bit different from one another in some sort of directly or indirectly descriptive way. These plants do indeed look a little different, but to make it clear, only one plant is illustrated. The landscape is very similar to that at Arizona north of VanRhynsdorp. The plants have to be retracted into the soil and one cannot form a good opinion without destructive sampling.
24. 7385 H. nortieri var pehlemanniae ‘devriesii’. N Prince Albert.
Conclusion This account is by no means is absolute. I have only illustrated about a half of the known populations and wonder just how many there are which have not been recorded in anyway at all. Of course it can be imagined that in some way more information will better indicate taxonomic entities. My experience persuades me that just the opposite will occur. As my knowledge of the plants in the field has grown, so has my conviction that we are dealing with highly integrated and variable systems. Our taxonomic systems and our understanding of them, especially in the learned ranks where we would expect to find it, are simply inadequate. Our system of classification is based on a belief system that there are simple diagnostic keys which can be derived from simple morphological characters. I think that this is because classification has been built on a slowly growing but still inadequate sampling base. The International Code for Botanical Nomenclature dominates taxonomic thinking which means that nomenclature and the management of names, overrides the knowledge that these names should convey. More attention needs to be paid to explaining different plants as systems that relate to geographic factors and other similar systems.
In terms of the general taxonomic paradigm, it may be reasonable to suppose that the “nortieri” system is composed of discrete taxa. I seriously question that this is so. As the record grows, so does the pattern of increasing variability of otherwise recognizable systems. As regards var pehlemanniae (“albispina”, “devriesii” included) I am quite convinced that even growing these plants in mass from seed cannot produce anything which alters the fact that they belong in this single system which I have discussed in this article. Given the time, the opportunity and the youth, I would try to explore and illustrate these eastern populations in greater depth as well as try and build substantially on the known record. There are huge areas that offer suitable habitats, and similarly huge areas that do not. These areas are largely inaccessible by virtue of the limits of the road network and then the problem of gaining access via land ownership. ♦
How did this start? Is it possible to see anything with such a quick peek? Somewhere in my memory bank is my stated opinion that understanding H. arachnoidea would assist any botanist towards a better understanding of classification. This was long before I came to see that this is a lot more necessary than I thought then. It is not just botanists who seem dumbed down to the reality of a diversity that is necessary for response to change and largely denied by the nomenclatural code and how it is practiced. We have not understood change or what changes can occur. During perhaps only the last 15 to 20 years has it become apparent that changes are cataclysmic and frequent. I will skip the fact that the violence of catastrophe, or the drivers of catastrophe, may also induce change at even a cellular level.
I usually go into the field with a specific goal while also carrying a host of peripheral questions in my mind. While exploring the problem of H. schoemanii, I was thus also thinking of Conophytum. After photographing a species seen at Laingsburg, I mentioned to Steven Hammer that I had also seen it in the southeastern Tanqua Karoo (Bakoven). This was news to him and so in my wish to re-establish the reality of H. venosa subsp. granulata recorded at Patasriver (actually Patatsriver road and that is another story that will be told) as well as the Conophytum for Steven, I undertook an expedition to the Tanqua.
It was co-incidence and luck that brought us into contact with Dr. De Ville Wickens, who is an expert on the geology of the Gondwanaland connection to the Tanqua mountains and who now owns Bizaansgat to the northwest of the farms Droogekloof and Bakoven. Dr. Wickens facilitated our expedition by offering us accommodation at Bizaansgat and we were thus able to spend more time there than otherwise.
1. ADH3140 Haworthia nortieri Skitterykloof2.
Derek Tribble did alert me to a small Haworthia in the Skitterykloof about 20km to the west of Bizaansgat. I had myself seen the plant and concluded that it was H. arachnoidea. Fairly recently Adam Harrower gave me a small plant from the vicinity of Skitterykloof and I confess that I was a bit confounded because it did implicate H. nortieri (See ADH3140, figs 1 & 2) particularly because the leaves were so spotted. Why I say this is because the combined presences of the look-alikes H. arachnoidea and H. nortieri ‘Albispina’ at Koup and elsewhere, as well as their often superficial resemblances, now make me a little wary of snap decisions. I later concluded from the flower that Adam’s plant was in fact H. arachnoidea, and not because I am convinced I could separate the two on the basis of the flower only. So this report covertly deals with the reality of a massive variation within H. nortieri and also H. arachnoidea and hope that readers will generate their own open mindedness to realize that I am drawing conclusions here that could or should be better substantiated. I also hope that this does not mean that a whole new set of Latin names is required to try and do so.
Because my initial field work was to obtain an overview of the genus, I soon stopped paying much attention to H. arachnoidea, because it seemed to be so ubiquitous. A big mistake! Unfortunately digital cameras were then not available and space in the herbarium (contrary to what my learned friend has to say) was and still is at a premium. The consequence is that I do not have a proper physical or mental record of all the plants and populations that I might have seen. In fact I cannot even fully remember how or why I linked H. arachnoidea to H. decipiens the way I did in my Handbook.
What happened on this expedition is that at Droogekloof we were surprised to find a very small Haworthia that I take to be H. arachnoidea (see MBB7644a, figs 3a–j). It is important to note the differences in colour, spination and leaf-spotting and/or reticulation. These plants were single and scattered over a fairly wide area. They varied in size from as little as 20mm to an average of about 30mm diam and become extremely cryptic in the dry summer.
3a. MBB 7644a H. arachnoidea Droogekloof3b.3c.3d.3e. MBB 7644a H. arachnoidea Droogekloof3f.3h.3i.3j.3g.
We continued to Bakoven and found it again (see MBB7644, figs 4a-e). Fig. 4c. is a view from a mountain top at Bakoven looking NNW towards Droogekloof and then Bizaansgat in the distance. Fig. 4b illustrates the weak inflourescence with few flowers generally true for many of the populations we observed. We the retraced our steps and saw similar plants approximately midway back to the Droogekloof population (MBBsn see fig. 5). The next day at Bizaansgat we saw exactly the same thing at our first three stops and then came across an absolute field of them at a fourth. Again at several more stops on Bizaansgat (MBB7646 see figs 6a-h) and yet again on another farm 5km to the east.
4a. MBB7644 H. arachnoidea Bakoven4b.4c. View northwards from Bakoven4d. MBB7644 H. arachnoidea Bakoven4e.5. MBBsn H. arachnoidea Droogekloof6a. MBB7646 H. arachnoidea Bizaansgat6b.6c.6d.6e.6f.6g.6h.
The next week took us on the road, the Patatsriver road, from the southern Tanqua to Matjesfontein east of Laingsburg. We stopped on the advice of Gerhard Marx at a well known spot at Perdekraal where Lithops, Didymaotus and Tanquana occur. We saw the same small H. arachnoidea there (MBB7658 see figs 7a-c).
7a. MBB H. arachnoidea Perdekraal7b.7c.
From there we turned north to Bantamsfontein for H. venosa subsp. granulata and we were surprised to find again the same H. arachnoidea at every place we stopped on the farm (MBB 7660 see figs 8a-e).
8a. MBB7660 H. arachnoidea Bantamsfontein8b.8c.8d.8e
We travelled on in the direction of Matjiesfontein to Patatsriver and again found it ubiquitous (MBB7666 see fig 9).
9. MBB7666 H. arachnoidea Patatsrivier
We went on to Keurkloof just northwest of Matjesfontein, and found it on the two hillsides we searched (MBB7670 fig.10).
10. MBB7670 H. arachnoidea Keurkloof
From there we went to Bulhouer, which is north Matjesfontein, and found it at four places (MBB7671 & 7672 see figs. 11a-d and 12a-e).
11a. MBB7671 H. arachnoidea Bulhouer S11b.11c.11d.
It is very important to note that Emil Heunis recorded the form ‘scabrispina’ on this farm Bulhouer and this is what I was expecting to find. However, my four samples were observed at regular intervals of about 1km from south to north. The southern populations were indistinguishable from any we had seen from Perdekraal in the west, but the last population, (MBB7672 figs 12a-e), were as close to as must be possible. My contention is that we thus observed continuity from the west to the east.
12a. MBB7672 H. arachnoidea Bulhouer N12b. MBB7672 H. arachnoidea Bulhouer N12c.12d.12e.
On northwards to Dwarsindieweg some 15km further and we found the small forms (MBB7677 see figs 13a-b) en route as well as commonly at Josephkraal another 16km east (MBB7678 see fig. 14). We found it again the next day north of Dwarsindieweg (MBB7686 see figs 15a-d) on the high upper slopes of Klein Tafelkop (MBB7688 see 16 fig 54). Heading for home we stopped on a backroad on the way to the Patatsriver road at Witrantjies and found it yet again (see figs 17a-c). We had been very cautious all the time to be sure we were dealing with one thing and not two. On this last stop we were confronted with the first drama when it seemed fairly certain that we had also found H. nortieri and this turned out to be false. We did return later to climb a high mountain at Volstruisfontein west of Dwarsindieweg hoping to find H. marumiana. Instead we again found small H. arachnoidea (MBB7802).
13. A view from south Dwarsindieweg.13b. MBB7677 H. arachnoidea Dwarsindieweg S14. MBB7678 H. arachnoidea Josephkraal15a. MBB7686 H. arachnoidea Dwarsindieweg15b.15c.15d.16. MBB7688 H. arachnoidea Klein Tafelkop17a. MBB7693 H. arachnoidea Volstruisfontein18a. KG16/72 H. arachnoidea Ladismith18a. KG16/72 H. arachnoidea Ladismith18b.18c.
It was not flowering time. The pictures that should have accompanied this article will show that the arachnoidea we found were often still accompanied by an odd old inflorescence. A weak spindly thing with evidence of seldom more than three old capsules.
Generally the plants were very small and we very often came across seemingly mature plants that were down to 20mm in diameter, and seldom exceeding 35mm. Invariably we could find the odd bigger plant in any single population array. The spination was fairly variable in respect of size, spacing and arrangement. The leaves could be short and squat or narrow and elongate. Colour was usually on the bright green side and I do not know what the summer colour would be. Leaf markings were interesting because often there was a slight reticulate pattern to the leaves and translucent dots and stripes and margins could be observed. This seemed to diminish in a cline from west to east. We did conclude, because this is what we also wanted to observe, that there was some evidence for continuity with H. arachnoidea var. scabrispina. There was the one population, however, north of Bulhouer where the plants were indeed bigger (50-60mm diam) and there was a definite tendency to form overarching brown spination so characteristic of scabrispina. There were of course the odd variants elsewhere and one group of four plants east of Dwarsindieweg could have come from the very confounded interface of H. arachnoidea and H. mucronata that is covered in the Latin H. arachnoidea var. nigricans. Readers should recognize that there is no way in which the nomenclatural code can be used to account for this diversity and variation, and it is time that more space and recognition is given to this very common phenomenon in the general context of writing, speech and communication.
The pictures will include one of ‘candida’, an epithet conveniently supplied by M. Hayashi for a variant of H. arachnoidea that occurs very close to Ladismith (KG16/72 see figs 18a-c). The spines in these plants often are black at the tips and the same phenomenon can be observed in massive plants that occur near Worcester in the “typical” variety arachnoidea. But I would like to break away from this usage. The epithet arachnoidea is applied to every single plant of that species wherever it occurs. Varietal names of any kind may be attached and I especially recommend the form and the formalization that Gordon Rowley advocates too; this is the format example…H. arachnoidea ‘scabrispina’. For more precision a whole host of abbreviated prefixes could adorn the label e.g. cf ‘scabrispina’ meaning “compare with” or v ‘Scabrispina’ as a cultivar or var. scabrispina as a generalisation.
There are a set of varietal names in H. arachnoidea already, and the proper address for these small Tanqua plants discussed in this article is in terms of my own Revision H. arachnoidea v. namaquensis. I do not think that my dispensation is adequate or good enough for the scale of our present knowledge, but neither do I think formalization is going to do much more than arouse an egocentric mass of still more Latin epithets. So I strongly suggest that we as a community simply accept something like H. arachnoidea ‘tanqua’ or ‘minuta’ and drop any pretension that Latinization adds any lustre or glamour to plants that are already fascinating and attractive. One of my objections to what I find a really irksome process of formalization (typification, full citation of basionym and all that stuff), almost senseless, because unless a microchip is also implanted it is unlikely that there will be any chance of associating any plant out of habitat back to its origins.
I am not going to write any further about H. nortieri cf ‘pehlemanniae’ here because I need to know more as well as confirm something about the flowers and flowering times of both sets discussed here. There were the occasional dark coloured plants in the arachnoidea (and excuse me dropping the ‘H’ – it really is not necessary in this informal environment), but I doubt very much that anything further will support the suspicion that arachnoidea will now always be confusable with nortieri. It turns out that what I though might be H. nortieri because of the very grey colour tone of the leaves was in fact also H. arachnoidea. A specimen flowering in cultivation produced non-globose flowers.
Acknowledgement: Dr. De Ville Wickens, Bizaansgat; Mr. P Schietekat, Stellenbosch; Mr. M. Jan duToit, Remhoogte Boerdery; Mr. Pieter de Graaf, Patatsriver; Mr. C.K. Francois, Bulhouer; Mr. Wynand Theron, Dwarsindieweg; Mr. Johan Kriel, Citruspoort (Volstruisfontein); Messrs. Stephen and Gail Louw, Keurkloof. Hospitality and access to properties is deeply appreciated and valued. ♦
I have become increasingly concerned about the poor relations that exist between collectors and the authority of Nature Conservation. The argument that collectors threaten and despoil natural populations is very real and I do not dispute at all that Conservation authorities have a very valid complaint. They have a function to perform. On the other hand there is an interaction between human beings and nature in all its forms that should be fostered to the benefit of both sides.
Nurseries, traders and collectors are as much of the picture as are conservationists, institutions, researchers and landowners. It is unfortunate that there is no non-government party that lobbies for the rights and activities of the former group, but it is not my intention nor within my competence to argue all the aspects of the case.
I strongly believe that people have the right of access to nature in all its forms and the issue is one of individual responsibility and proper consideration of consequences. An appreciation of and sensitivity to nature should be reflected in whatever we do in our lives. My own collecting impulses led me to institutional employment where I could exercise my interest to what I thought were efforts more worthy than my personal interests. From that position I also did try to share and extend privileges to a wider circle. It is in this way that I became involved with Sheilam Nursery. It was not my wish or intention that my collection should have come to be housed there. However, Sheilam has succeeded over a period of nearly 40 years to maintain a fairly true record of my collections obtained as propagated material from the Karoo Garden at Worcester. My offer of permitted collections dating from my revision of Haworthia in 1966 to the Karoo Garden was rejected and for a while resided with Etwin Aslander at Brackenfell. It has since passed to Garth Schwegman at Sheilam who has taken a particular interest in the maintenance and propagation of that collection.
Hand pollination of haworthia has become the norm and Sheilam has made a massive effort to generate plants from this. Of course growing plants from seed is a lengthy process, but is richly rewarded with an astonishing array of variants and beautiful plants.
Need I say how important this activity can be for an understanding of the plants and the way this is expressed in a classification? A good example is of the hybridization found between H. lockwoodii and H. arachnoidea ‘scabrispina’ at the Floriskraal Dam east of Laingsburg (see figs 1-3). The hybrid is very reminiscent of plants that I have seen from between there and Prince Albert; and also southeast of the Rooinek Pass through the Witteberg, and again northeast of Calitzdorp. Roy Mottram underscored the issue of hybridization in Haworthia classification. I have not ignored the issue, but rather viewed it as an imponderable in the continuum between populations that seems to have pattern.
Figures 4 and 5 are seedlings of a collection of mine from north east of Nieuwoudtville of H. nortieri. They could be related to ‘globosiflora’ and it will be interesting to see the flowers. However, it may be foolish to draw any conclusion from whatever flower shape emerges in the broader nature of the variation already observed although not adequately documented, both in H. nortieri and in other species complexes. Plants with these stubby deeply mottled leaves can be the consequence of growing conditions just as much as of genetic variation intrinsic to a ’species’.
Figs 6-12 are seedlings of another collection of mine (MBB7608) from Melkhoutrivier south of the lower Breede River and north of the Potberg Mountain. This collection is reported in Chapter 7 of Update 4, I think very aptly entitled “The brutality of reality in Haworthia”. It is one of 12-15 populations in the greater area that support my suggestion of H. mirabilis as including H. maraisii, H. magnifica and H. heidelbergensis. I suspect that H. mutica is also involved to some degree or other, while it is not difficult to extend comparisons to as far afield as Riversdale and further to H. emelyae in the Little Karoo.
Collectors will no doubt always have a problem with plants that do not match the perceptions they attach to names. Kobus Venter has always expressed anathema for anything but random selection when collecting specimen plants. This is in some degree correct because the specimens are supposed to represent a population. However, the opinion on a population is largely based on field observation. Subsequent cultivation and observation do not greatly influence that, while oddities and unusual forms may help mould opinion. While Nature Conservation may object to the introduction of permitted collections eventually into the collecting arena, I see no ethical grounds for this. I insist that it is an undeniable right that people have right of access to a natural resource with the proviso that there is no pressure on sustainability. ♦
Some criticism about my supposedly having ignored the flowers in Haworthia comes at a very inopportune time. I set aside flowers for the reasons very obvious from the historical record but also because of the considerable problems of similarity in the appearance of the flowers in apparently quite different species. My priority was a geographic overview and a rational basis on which discussion and decision making could be based. It I just grossly unfortunate that other writers and critics seem to be wedded to a classification paradigm locked into the approach that prevailed 70 years and more ago. This in the total absence of a species definition other than the vague acceptance of a zoological one based on interbreeding capabilities. This ignoring the ease of hybridization among Haworthia variants in general.
While I have written an account of flower appearances in a small selection of populations, I also came across these few images I have of flowers in what I regard as the species H. nortieri. I have also added images of a single flower of H. maculata from a population high in the mountains at Worcester that could be seen as a southern extension of the H. nortieri set of populations. Note must be taken of my early contention that H. nortieri and H. globosiflora were the same species, based on my observation of the intermediate appearance of the flowers of a Vanrhyns Pass population. The H. maculata bud is typical of the species in the Southern Cape, whereas H. nortieri has rounded bud-tips.
The flower of the Trawal plant are dramatically different from that of, say, Sneeuberg. It is very understandable that differences like this lie at the base of all the argumentation and confusion that so despoils the naming and identification of Haworthia. A classification has been needed against which to explore and examine these differences. It seems to me totally unnecessary to try and construct another hierarchy of solely Latin names while so little is still unknown. ♦
6158 H. nortieri. Sneeuberg, N Ceres. 13. 6505.6 H. nortieri. N Dwarsrivier, Cedarberg.13. 6505.5 H. nortieri. N Dwarsrivier, Cedarberg.14. 6506.f2 H. nortieri. Dwarsrivier, Cedarberg.14. 6506.f1 H. nortieri. Dwarsrivier, Cedarberg.7579 H. nortieri. Trawal Bridge.EvJ19550 H. nortieri. Blinkberg. PVB6146.7 H. nortieri. Komkans. 3. PVB6146.4 H. nortieri ‘agnis’. Komkans.3. PVB6146.3 H. nortieri ‘agnis’ Komkans.4. EA1442.1 H. nortieri ‘agnis’. Kareeberg, Nuwerus. H. nortieri ‘agnis’. Nuwerus.
In Haworthia Update Vol. 9 dealing with H. maculata, I again draw attention to several populations that are problematic in an area very difficult to explore. The populations are MBB7865 at Keurkloof southeast of DeDoorns, EA1441 at the Hex River Pass, an unrecorded population south of Sandhills (about 3km east of Kanetvlei) reported by Ernst van Jaarsveld. In addition there are two populations apparently of H. arachnoidea at Tunnel Station east of Osplaas.
The illustrations that accompany this article are:
MBB6694 Vreesniet, Kanetvlei.
EA1441 Hex River Pass, east DeDoorns.
MBB7865 ex E. Van Jaarsveld, Keurkloof, DeDoorns.
This population MBB6694 is at Kanetvlei southeast of Sandhills, and I originally ascribed it to H. arachnoidea but this is not correct. To this I can add unexamined populations reported by P.V. Bruyns deep in the Langeberg at Keerom Dam and another of Ted Oliver in the mountains north of Nuy. MBB6694 is only 200-300m north of MBB7994 also in sandstone. The sandstone strata are by no means all the same and one can even find shale bands in what is essentially a sandstone formation. H. maculata flowers late winter to early spring and these other populations in early summer.
View westwards to the Kanetvlei
I have not examined the flowers in any number and remain doubtful that this information will clarify any deductions that can be drawn from the vegetative characters or the geographic positions of the populations. It appears to me that consideration must be given to a relationship with H. nortieri that in my assessment occurs from the far northwest to as far east as Prince Albert, i.e. H. nortieri (devriesii). What I have seen of the flower, and shown now is that it definitely does not belong in the context of the Southern Cape species where the bud-tips tend to be “fish-tailed”. The inflorescence are slender and few flowered.
There is some criticism of my emphasis on geographical distribution as a key indicator of species. It was even said, as though I would have denied the fact, that I could not identify plants unless I knew where they came from. There is an incredible amount of both ignorance and the obtuse behind these statements. One critic feels that I have ignored detail and even ignored flower characters in the development of my opinions. This same critic claims a high degree of agreement with me (80%) based apparently on the view that so much of my classification is correct (meets with his approval), and presumably that the 20 percent that he contributes to the system makes it 100% correct. This is wholly wrong. This critic is simply unable to bridge the historical chasm between his introduction to, or knowledge of, Haworthia and mine. He simply does not recognise the shift in method and why I switched from a “character-based” approach to a phytogeographic one. It is really very simple in that the character based system as used from the very beginning of classification, has not produced a solution to the identification and classification of Haworthia. It all rests on a very weak and shaky definition of species determined by breeding barriers and consequently that morphological differences necessarily exist. My approach was to show that breeding barriers (at least in plants) could be integral to species and that morphological differences could equally be nothing but variation within species. Therefore I arrived at a definition of species from the view that they were phenomena spread in space and changing with time. This is just a fundamental of plant species as they constitute the vegetation and the floras of the world. Virtually the first question following “What is that?”, is “where does it come from?”.
Why 6694 is so interesting is precisely because it occurs in an area which is so poorly known and represented in our knowledge of Haworthia. The reality in the subgenus Haworthia is that there are several areas of great species richness and there is real pattern in the distribution of the various elements as they are listed in the 80% agreement area. The 20% disagreement zone concerns truly trivial opinion and unsubstantiated statements, and almost ignores the real reasons for disagreement at all. The real reasons are the realities that superficial morphological difference does not mean species and I have posted a vast amount of material that demonstrates that. I have shown repeatedly that differences in single characters in what can be rationally said to be the same species, can be greater than between different species.
In the case of 6694 I did not pick any particular character to identify the plants as H. arachnoidea from probably as far back as 1975 and I can find no earlier record of this location. I do recall a specimen in the Compton herbarium made by W.F.Barker that I mentally allied with H.arachnoidea but it is not cited as such in my revision. The identification was based on little else other than the spiny-ness of the plants and the fact that the nearest known locality for that species was a very new one of mine a few kilometres to the southwest. At the time, H. nortieri was barely known and this was from far to the northwest at Clanwilliam. Since then H. nortieri has become much better known although still some distance north of Ceres, and with its globose-flowered variants at Laingsburg and eastwards. H. arachnoidea remains remarkably unknown from the Cedarberg and Koue Bokkeveld mountains but does appear in the upper Hex River Valley. Here it is odd that the leaves tend to have the translucent dots that one would expect to characterise H. nortieri. But otherwise the leaf coloration is the much darker green that is associated with H. arachnoidea. A twist is that at Kunje, southeast of Citrusdal, H. nortieri does have very dark green leaves and there is no doubt that confusion with H. arachnoidea is inevitable. I have very little to offer in this regard because this degree and level of confusion or doubt is intrinsic to the genus anyway.
Why I now decide to relate this Kanetvlei population to H. nortieri is not to be construed as a decision now to call white what previously was clearly and definitely black. It is taking all my knowledge and experience to suggest simply that this is a better reflection of the situation where there is not enough hard data to determine identity at a higher level of certainty than about 50%. What I do suggest that there are many situations in Haworthia as the case with H. maculata and H. pubescens only recently exposed. In this case the two very different elements can be very confidently be said to be the same in respect of distribution and linking populations AND characters. Throughout, particularly the subgenus Haworthia, we have situations of continuity and spatial (geographic trend) and nearly all my many publications deal with exactly this reality. What it suggests is that my critic should come to consider if he has some other motive between punting points of difference that prevent him from finding points of agreement, besides those that rest coldly on logic.
1. MBB6694 Vreesniet, Kanetvlei.
Flower profiles.
Flower faces.
Flower buds.
2. EA1441 Hex River Pass, east DeDoorns.
3. MBB7865 ex E. Van Jaarsveld, Keurkloof, DeDoorns.