14. Haworthia heidelbergensis Smith, JS.Afr.Bot. 14:42(1948). Bayer :121(1976). Bayer :41(1982). Scott :131(1985). Esterhuizen, Aloe 29:64(1992). idem. 33:15(1996). Type: CAPE‑3420 (Bredasdorp): W. Heidelberg (‑BB), J. Dekenah 230 in Smith 6566 (NBG).
heidelbergensis: from Heidelberg, Cape.
Rosette stemless, proliferous, to 8cm φ. Leaves many, erect to recurved, generally with small marginal and keel spines, usually dark green with reddish hues, end-area of leaf slightly translucent. Inflorescence simple. Flowers 10-15, white with brownish veins.
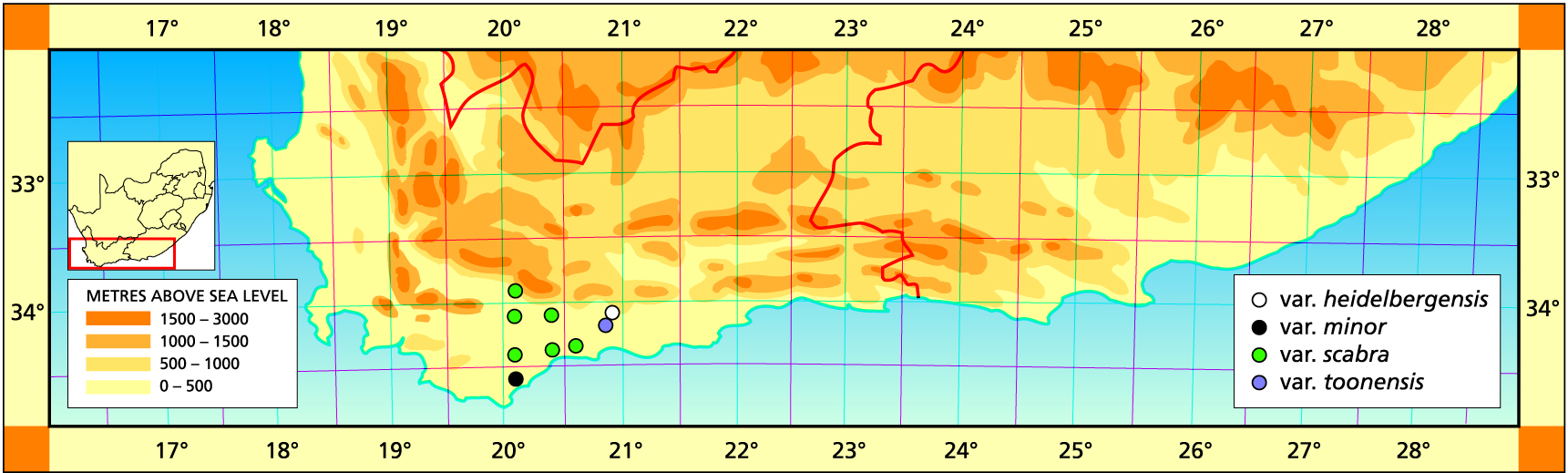
1982 – This is a small species occurring immediately east of Heidelberg. It grows deeply imbedded in moss and lichen and although the leaves have a distinct end area they are markedly acuminate. Known only from one locality, there is a related population at Matjestoon to the southwest in which the plants have longer recurving leaves, and another at Bredasdorp similar to the last. Together these three populations have no obvious affinities with any other species in the complex. Heidelberg is a very significant area for haworthias, as species such as H. turgida, H. magnifica, H. floribunda and H. retusa all occur nearby. H. heidelbergensis is not continuously variable with any of these. It is a winter‑growing species, invariably non‑proliferous and difficult in cultivation. The occurrence in moss and lichen suggests that it requires more shade than usual, as well as a distinct summer resting period.
1999 – This has proved to be the most surprising species among all the collections of the last ten years. Many collections have been made including at all the compass points immediately around Heidelberg, although those north are really considered hybrid with H. turgida. As Esterhuysen (1992) pointed out, the type locality is probably west of Heidelberg as recorded in Smith’s collecting records and on his type specimen. This is cited in the published description simply as ‘Swellendam District’. It extends further east towards Riversdale, westwards to Bonnievale and is common east and north of Bredasdorp. It has proved to be a major role player among the species in that area apart from its association with H. emelyae already dealt with. The populations around Heidelberg show evidence of direct introgression with H. turgida and cast new light on the relationship of that species with H. retusa. H. heidelbergensis is apparently the eastern counterpart of H. mirabilis and these two species never co-occur. H. serrata also cannot be excluded from discussion of this species (see H. serrata). At its westernmost limit at Bonnievale, it is confounded with H. maraisii var. meiringii. At the type locality it is rather atypical, an oxymoronic statement. Here the leaves are erect, but with a distinct end-area and there are not many leaves. Characteristic of the species is, however, the abundance of individuals at each site and their density in colonies usually on slightly southern slopes and in thick moss. The leaves may be smoothish and cloudily translucent or with translucence between veins on the upper leaf surfaces. The leaves tend to spread and recurve rather than being distinctly retused or with a swollen end-area as in the type. This is also one of the main feature separating H. mirabilis from H. maraisii, although it is by no means an absolute discriminant. The colour is also usually a reddish tinted greenish-brown. At Bredasdorp the population is greener than normal and the plants are like miniaturised H. mirabilis. Several varieties are distinguished to facilitate perception of this species.
a. var. heidelbergensis Resende in Feddes Repert.Spec.Nov. 48:114(1940) suggested a change in the rules of nomenclature so that if a name proved not to be the true stem form (in other words not central to the circumscription of the species) the type should be changed for one that was. This is a case where that rule would prove quite useful. A manuscript concerning the pivotal role this species plays in the Southern Cape has been submitted to ALOE. Despite the major role this species plays in relation to all the species in the Southern Cape, not one of the illustrations in Bayer (1980), ‘The story of Haworthia nitidula’ depicts this. Larger plants occurs around Heidelberg and mid-way to Riversdale. The actual locality of the collection is rather odd, as Dekenah pointed out the locality to the author as east of the town, whereas there are two collection of Smiths received from Dekenah, one east and one west. They are virtually identical and Esterhuizen (1992) confirms this.
Distribution: 3420 (Bredasdorp): W. Heidelberg (‑BB), J. Dekenah 230 in Smith 6566 (NBG); E. Heidelberg (-BB), Smith 5756 (NBG), Bayer 2550 (NBG); Koppie opposite sports grounds (-BB), Smith 5044 (NBG); S. Heidelberg (-BB), Smith 6989 (NBG).
Haworthia heidelbergensis var. heidelbergensis JDV93/54 G=Between Heidelberg and Riversdale. An extensive population where the plants are robust.
Haworthia heidelbergensis var. heidelbergensis JDV87/1 east of Heidelberg. The upper leaf surfaces are always a little translucent.
Haworthia heidelbergensis var. heidelbergensis JDV87/1 east of Heidelberg. Plants are smallish and grow in lichen and moss.
b. var. minor var.nov. Type: CAPE-3420( Bredasdorp): Rooivlei (-CA), Bayer in KG 36/70 (NBG, Holo.).
minor: smaller.
Differs in being small, to 3cm φ, and in being light yellowish-green and well spined. (A var. heidelbergensis foliis parvioribus flavo-virentibus pallidis et spinosis differt).
North of Bredasdorp the plants resemble miniaturised forms of H. mirabilis var. sublineata and have the same plain green colour. The leaf-tips are also notably translucent.
Distribution: 3420 (Bredasdorp): Rooivlei (-CA), Bayer in KG 36/70 (NBG), Smith 3903 (NBG).
Haworthia heidelbergensis var. minor JDV86/5 north of Bredasdorp. Leaves narrower and smaller than the typical variety.
c. var. scabra var.nov. Type: CAPE-3420 (Bredasdorp): Leeurivier (-AB), Bayer 1700 (NBG, Holo.).
scabra: rough.
Differs in being small, to 3cm φ and as tall. The colour is usually very dark green. The leaves erect or sub-erect, slightly scabrid especially along the margins and keel. (A var. heidelbergensis foliis recurvatioribus cum fenestra apicali distincta differt).
This is the commonest variety in terms of population distribution. The plants are quite small and very easily mistaken for erect-leaved forms of H. maraisii despite the general absence of ‘cloudy’ translucent windows in the leaves of the latter species. Although populations are often small, they are usually very dense.
Distribution: 3320 (Montagu): Drew (-CC), Fouche in PRE 34945; W. Bonnievale (-CC), Bayer 6509 (NBG), Lewis in NBG2457/33 (BOL). CAPE-3420 (Bredasdorp): Kliphoogte (-AA), Bayer 4677 (NBG); Leeurivier (-AB), Bayer 1700 (NBG); Brakfontein (-AC), Bayer 2547 (NBG); Beyersdal (-AD), Bayer 2556 (NBG); Haarwegskloof (-AD), Bayer 5101 (NBG); W. Kathoek (-AD), Bayer & Bruyns 6566 (NBG), Burgers 2581 (NBG); E. Kathoek (-AD), Bayer & Bruyns 6561 (NBG); N. Potberg (-BC), Bayer & Bruyns 6544 (NBG), Bayer & Bruyns 6545 (NBG).
Haworthia heidelbergensis var. scabra JDV86/3 north-west of Bredarsdorp. Occurs here in very dense clusters but very localized.
Haworthia heidelbergensis var. scabra JDV92/85 north of Infanta. The leaves are shorter and broader than typical.
Haworthia heidelbergensis var. scabra JDV87/4 east of Drew. Less translucent and the leaves are a little more rigid and scabrid than the other varieties.
Haworthia heidelbergensis var. scabra JDV87/4 east of Drew. This population was severely damaged by cultivation and has survived only in a few remaining small ridges.
d. var. toonensis var.nov. Type: CAPE-3420 (Bredasdorp): Matjestoon (-BB), Smith 6797 (NBG, Holo.).
toonensis: abbreviation for origin at Matjestoon.
Differs in the more recurved leaves with a distinct transparent end-area to the leaves.
This appeared as an article in ALOE 38:31 (2001). Unfortunately there was a problem with illustrations and captions and these are corrected here. A comment is also added as an addendum to respond to criticism by I. Breuer published in Alsterworthia 1:13(2002).
Introduction:
I described Haworthia serrata in 1973 (Jl S.AFr.Bot.39:249, see Figs.1) from Oudekraal, southwest of Heidelberg. I commented then on the wisdom of describing a new species when “the recognition, estimation of taxonomic rank and circumscription of elements in Haworthia…” was so problematic. The new species was said to resemble H. emelyae var. multifolia (Figs.2). In respect of its distribution, I said it was closest to H. heidelbergensis at Heidelberg (Figs.3 JDV87/1) and as at Matjestoon (Fig.4 JDV87/3), and also to H. sublimpidula at Swellendam (now known to be H. floribunda var. major (Fig.5 MBB6859, taxonomically with little connection to H. rossouwii). The implication was that it could have been taxonomically related to those elements in terms of geographic distribution. I was still puzzled by the relationships of H. serrata when I wrote (New Haworthia Handbook :55, 1982) that collections by C. Burgers from the coastal limestones might throw more light on the matter (Fig.6 MBB6985 H. mirabilis var. calcarea).
Fig. 01a. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.rossouwii 1 043Fig. 01b. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.Fig. 01c JDV87-47 H. rossouwii, Oudekraal, Heidelberg.Fig. 01d. JDV88-53 H. rossouwii. Oudekraal, Heidelberg.Fig. 02a. JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 02b JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 03a. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03b. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03c. MBBsn H. heidelbergensis. E Heidelberg.Fig. 04. JDV87-3 H. heidelbergensis var toonensis. Matjestoon, SW Heidelberg.Fig. 05. MBB6859 H. floribunda var major. S. Swellendam.
In his revision of Haworthia (1983), C.L. Scott recognised my species. He confused it, and illustrated it, with a photograph of H. mucronata var. mucronata sensu Bayer (1999), which he called H. mclarenii. This does exemplify the problem of look-alikes in the subgenus, and this curiously does have some connection to the actual problem.
When I wrote Haworthia Revisited (1999), I was still not sure what the real affinities of H. serrata were. I mentioned that my 1982 observations were vague because of the unusual elements which occurred in the coastal limestones. I described these as new: H. mirabilis var. calcarea from De Hoop Nature Reserve (see Fig.6), H. variegata var. petrophila (CBurgers2158) from Karsriver and H. heidelbergensis var. minor from N Bredasdorp (Figs.7 JDV86/5). In my 1999 classification, I placed H. magnifica var. paradoxa (Figs.8 JDV86/78 & 94/89) in H. mirabilis, suggesting that it may have a connection there via the var. calcarea and an element from the Potberg (CBurgers 2018) which I have not yet seen in the field myself (since I wrote the article I have in fact seen this population, MBB7356, and it is discussed in the Chapter re H. mirabilis). The connections of paradoxa must be viewed in its relationship to H. emelyae var. major, in the same way that serrata is compared with H. emelyae var. multifolia. I also suggested that H. serrata had a very close resemblance to some forms of H. mirabilis var. sublineata from south of Bredasdorp. This present paper addresses some of these observations again.
Fig. 17b. Fig. 06a. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 06b. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 07a. JDV86-5 H. heidelbergenis var minor. Rooivlei, Bredasdorp.Fig. 07b. JDV86-5 H. heidelbergensis. Rooivlei, N Bredasdorp.Fig. 07c. JDV86-5 H. heidelbergensis var minor. Rooivlei, Bredasdorp.Fig. 08a. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08b. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08c. JDV98-89 H. mirabilis var paradoxa.
New Records In June 1999, I revisited H. serrata at Oudekraalkop (MBB166) again, this time together with J.D.Venter. Despite rumours of over-collection, it was still very abundant, although localised. It was also still present at Koppies (Fig.9 JDV99/6) a short distance to the west, but in very low numbers at a badly burnt site. P.V. Bruyns (priv. comm.) has reported it at a more northerly site as well. This could be where J.N.O. Uys also showed it to me at a second site on Oudekraal when he first introduced me to this species. It struck me again how similar the plants were to H. mirabilis (particularly the var. sublineata, Figs.10 MBB6639, bearing in mind the vast variation that exists there) and to H. heidelbergensis var. minor. The visit in June ’99 was a general excursion to explore the reported presence of H. maraisii at Koppies (ex E. Esterhuizen), in relation to that species and heidelbergensis as I know it in that general area. Our finds and observations led us later in the year to explore the area north of the Potberg. The extraordinary finds there led me to further explore westward to Bredasdorp itself and to see for myself the var. calcarea in the De Hoop. Two visits for that purpose were fruitless, but thanks to Dennis DeKok, I did locate it in the company of Lawrence Loucka in July 2000 (MBB6985-fig.11). This latter excursion was particularly fruitful because, by great good fortune, we discovered H. serrata in unmistakable form north of Bredasdorp (Soutkloof, Fig.12a & b MBB6983), and Nooitgedacht, MBB6984). Both sites are west of Rooivlei where heidelbergensis var. minor occurs. We also collected variegata var. petrophila (Fig. 13 MBB6986) west of the Karsriver limeworks. For that variety I have C. Burger’s record (CB2158) from east of the limeworks, and a collection I made at Renosterfontein (Fig.14 MBB6632) on the east bank of the Karsriver in 1997. This latter collection, in particular, reminded me in both form and colour of H. emelyae var. multifolia (see fig.2).
Fig. 10a. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10b. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10c. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10d. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10e.Fig. 10f.Fig. 10g.Fig. 11a. MBB6985 H. rossouwii var calcarea. DeHoopFig. 11b.Fig. 12a. MBB6983 H. rossouwii. Soutkloof. Fig. 12b.Fig. 13a. MBB6986 H. petrophila KarsrivierFig. 13b.Fig. 14. MBB6632 H. rossouwii var petrophila. Renosterfontein, Karsriver.
The conclusion from seeing H. serrata north of Bredasdorp, and both var. calcarea and var. petrophila, is that this is the prime connection. It seems now more improbable that the var. petrophila is as close to H. variegata as I had supposed, and that the var. calcarea is closely connected to both H. serrata and to var. petrophila. The other observations remain valid viz. that there is a strong resemblance between serrata and mirabilis var. sublineata; but particularly that serrata may be connected to heidelbergensis via the var. minor. However, heidelbergensis in its general relationships is extremely complex. It is interfused with H. maraisii, it may be the direct (if complex) geographic extension of H. mirabilis in the context of the relationship between that species and maraisii, and it may have affinities and continuity with both H. variegata and H. floribunda. There is still another problem: H. variegata var. modesta at Kathoek (Figs.15 JDV97/24) and Potberg (Figs.16 MBB6542). My collection of this element at Kathoek (Figs.17 MBB2551) suggests that it is possibly a small very proliferous sand dwelling serrata. J.D. Venter’s collection from the same farm is a more robust element with rather longer, more erect leaves which suggests H. variegata. My Potberg collection (fig.16) is another small element which I relate to variegata although the leaves are shorter and more turgid. It must be borne in mind that H. variegata var. hemicrypta occurs north of the Potberg on the gravelly flat so that there is ecotypic variation in evidence. Further work will have to be done to elucidate what could be an unresolvable situation i.e. the Potberg is home to variants which link all the elements so far named. The evidence for this is already available, but insufficient to crystallise a solution.
Fig. 15a. JDV97-24 H. vaiegata var modesta. Kathoek.Fig. 15b.Fig. 16a. MBB6542 H. variegata var modesta. Potberg.Fig. 16b.Fig. 16c.Fig. 17a. MBB2551 H. variegata var modesta. Kathoek.Fig. 17b.
There will be difficulty in considering the extent and the degree of similarity of H. serrata to other populations. Some of these which include serrata-look-alikes are:
Fig. 18. JDV96-52 H. heidelbergensis var . N Kathoek.Fig. 19. JDV93-51 H. heidelbergensis. Melkboom.Fig. 20a. JDV87-4 H. heidelbergensis var scabra, Leeurivier, Drew.Fig. 20b.Fig. 20c.Fig. 20d.Fig. 21. JDV97-49 H. mirabilis var consanguinea. Dwarswaterkloof, MacGregor.Fig. 22. JDV88-83 H. mirabilis var consanguinea. Dwarswaterkloof, Macgregor.Fig. 23a. JDV96-38 H. maraiss var notabilis. Agtervink, Robertson.Fig. 23b.Fig. 23c.Fig. 23d.Fig. 24a. JDV98-1 H. maraisii var notabilis, Wolfkloof, Robertson.Fig. 24b.Fig. 25. JDV97-30 H. cf. heidelbergensis. N Ashton.Fig. 26. JDV86-1 H. mirabilis var triebneriana. Stormsvlei.
Some of these populations and their variants are dealt with in other chapters (Particular note must be made of H. elizeae which has to be considered in relation to H. rossouwii).
H. serrata was coming into bud in July a good five months after mirabilis had started to flower. This shows that my predictive thoughts about the relationship with calcarea and petrophila may not be particularly sound if flowering time is considered. Calcarea flowered a little before mirabilis var. sublineata in January, petrophila flowered in February while Roovlei heidelbergensis and Leeurivier (Drew) heidelbergensis flowered in December and March respectively. Flowering time for other collections needs to be observed, noting that this is not necessarily definitive either (for example while H. mirabilis var. badia is essentially late-summer flowering, single plants may flower in winter.
When I consider how much time I have spent in the area, it was really surprising to find serrata so far west. It means that something very interesting can still be found in the large triangle Swellendam/Bredasdorp/Riviersonderend, and that exploration will have to be done on a very small scale. The Karsriver and Renosterfontein petrophila are just smaller more spinose versions of Oudekraal, and the leaves are more spotted on the reverse. Calcarea was previously known from two clones which had rather shorter leaves and, perhaps, fewer leaves per rosette. This, the locality, and the spotting on the back of the leaves would have been the reason for opting for a relationship with mirabilis. We only found it with quite detailed directions because it was such a small population and so cryptic, consisting of three tiny locations under bush with a total of about 15 clumps. One location could have been the product of vegetative propagation with six to seven clumps. The second was similar with only two clumps and evidence of porcupine activity around the clumps. The third covered about 15sqm and there were another 15 or so plants as clumps or as single rosettes. These did vary and only one clump really resembles the two I have seen in cultivation viz. the Karoo Garden plant of C. Burgers, and a plant I received from Dennis DeKok. Burgers, however, appears to have made two collections from De Hoop; this needs to be confirmed. The plants seen by me in habitat do vary quite a bit and the range of variation equates that of both serrata ex Oudekraal (the Koppies population is reduced to about five plants) and north of Bredasdorp. The mean length of the leaves and leaf number per rosette may be shifted to the shorter and fewer, respectively.
The two Bredasdorp populations of H. serrata are remarkable for the similarity of the plants to serrata at the type locality. The habitat is a little different and at the westernmost of the populations, the plants cluster much more freely than the Heidelberg populations. At Koppies the plants are on a Witteberg substrate, whereas at Oudekraal and N Bredasdorp, it is Bokkeveld shale. We may have a slightly biased image of serrata with turned out leaves; the plants can be quite globose, and I did illustrate a plant like that in my original description when I drew the comparison with herbacea.
Where serrata really differs from mirabilis is by the greater number of leaves per rosette, the thinness of the leaves, narrowness at their bases, lack of a mid‑leaf swelling and by the incurving of the leaf tip; that character is fairly significant. H. mirabilis var. sublineata grown recently from seed has proved to be remarkably variable, but none of the plants have incurving leaves. Some of the clones are very similar to serrata, but the flowering time is earlier in the year and more in keeping with H. mirabilis in that respect.
Discussion H. variegata var. petrophila and H. mirabilis var. calcarea should be transferred to the species H. serrata. The same may be true for H. heidelbergensis var. minor. However, H. heidelbergensis has a complex distribution and there are other complexities in variation that preclude a definitive statement. It is quite probable in the light of the findings discussed in Haworthia Update Vol.1, that there is no discontinuity which satisfactorily and convincingly can be claimed to distinguish species. This is probably just another example of a more general problem which I have experienced with the identification and classification of many plant genera.
I am conscious that I have not dealt fully with the problem in omitting mention of co-occurrence (I mean sympatry – when two obviously different elements grow together at the same site or very closely adjacent). I see this as the best measure of discontinuity. Thus if two elements grow in close proximity, even if not sharing the same habitat, they are probably discrete. This does not preclude the possibility that they are simply ecotypes and this is taken into consideration. However, I have before pointed out that a taxonomic solution which can be deduced in this way for populations might not be true for the entire distribution of the species concerned. It is evident that serrata generally occurs alone. At Oudekraal and at Koppies, there is a floribunda/maraisii element in the vicinity. In the case of heidelbergensis var minor, both turgida and maraisii are present. I have always been struck by the similarity of mirabilis and serrata, and by the fact that they do not co-occur. Mirabilis, in the context of this paper, is presently not known east of Napier except at Bredasdorp and Mierkraal south of Bredasdorp. This is as the var. sublineata and var. mirabilis respectively, which differ from serrata in respect of flowering time, less leaves per rosette, and the non-incurving leaves. Curiously, it appears that mutica, at least in the Bredasdorp area, grows generally alone too, although at Haarwegskloof (Bredasdorp) it does grow with heidelbergensis. At Drew it is in the close company of both maraisii and heidelbergensis, and south of Stormsvlei is near to a maraisii/mirabilis intermediate. Thus even an element as apparently different as H. mutica may presumed to be, cannot be ignored.
However, another consideration now enters the picture with the discovery of serrata so far west. There are three poorly understood elements which were reported to have come from Bredasdorp and Napier respectively. The first of these was H. bijliana von Poelln. that does, however, seem to belong rather with H. arachnoidea than with anything else. G.G. Smith appears to have had a plant of this ilk which he received from Maj. Venter 9, from Caledon. The other two, however, are H. altilinea var. bicarinata Triebner and H. rossouwii v.Poelln. They were both described from the same collection viz. Napier at Bredasdorp, Rossouw in Triebner 1059. The former was never illustrated and the description is rather brief … “Backs of leaves always with two keels and with teeth. Leaves 3-5cm long, 6-8mm broad, with 4-6 dark lines in lighter part, with two keels at back, keels with up to 2mm long denticula”. This description is in the joint paper by Triebner and von Poellnitz, but authorship is attributed to Triebner alone. von Poellnitz later retained this variety when replacing the name altilinea with mucronata (note that in the context of the names which I use, Scott confused my serrata with mucronata as well). This is very curious because it is evident from the description of H. rossouwii, which is very much more detailed, that another plant from the same collection is in fact again described. The differences are essentially that von Poellnitz gives the leaves as 5-6cm long, and 8-10mm wide as opposed to the respective 3-5cm and 6-8mm by Triebner; and with three lines on the ‘end surface’ as opposed to four to six by Triebner. This says a great deal about the detailed descriptions which are routinely called for, and which in Haworthia seldom amount to much because of the problem of look-alikes, and the variability within and between populations.
I repeat the von Poellnitz description here to illustrate that it is a very good description that nevertheless omits some key information:
H. rossouwii von Poellnitz spec.nov. (Stemless rosette, spirally many-leaved. Leaves rather pale green, nearly oblong lanceolate, acuminate, on the face somewhat retused near the apex thus forming a triangular face; on the back, towards the apex, with sub-pellucid tubercle-like spots, denticulate on the margins and keels; the triangular face strongly acuminate, sub-translucent, not tubercled, but carrying some sub-pellucid small teeth, and traversed by a few longitudinal lines).
Stemless many-leaved rosette about 5-6cm in diameter. Leaves more or less erect, 5-6cm long, 8-10mm wide, about oblong-lanceolate, tapering gradually, pointed, slightly widened at the base, almost straight or a little bent to one side near the tip, rather light green, dull. On the face flat or usually somewhat convex, near the tip slightly bent backwards (retused) at an angle of 20 degrees; on the back, from the middle outward or near the tip obliquely keeled, or somewhat more rarely with two keels, near the tip with a few, often elongated, very often raised slightly pellucid spots, and frequently with a few leaf-coloured tubercles. The end-surface half-pellucid, 20mm long or somewhat longer, 7-9mm wide at the base, triangular, tapering very gradually, pointed at the tip often slightly curved inward, somewhat convex, smooth, rarely with a few indications of tubercles, but with some semi-translucent teeth (these teeth loosely arranged in a lengthwise middle line, now and then also in another line nearer the margin and very rarely scattered about in small numbers) with three long, occasionally interrupted, green lengthwise lines, of which only the middle one may reach almost to the apex, and usually with a few very short line; leaf margins and keel with pale teeth, or near the base often leaf-coloured, teeth, up to barely 2mm long; end-bristle pale, with small side bristles, 5-7mm long.
… On account of the teeth on the end surface stands near to H. schuldtiana and to H. paradoxa (I treat these respectively as H. maraisii and H. mirabilis var. paradoxa) but is easily distinguished from both. Its flowers are thus far unknown”.
The omissions are the flower of course, and there is no stipulation of the approximate number of leaves, and leaf thickness. The weakness is that it is a single plant description, evidenced by the fact that another plant from the same collection is accepted as a variant of a totally different complex.
In the past I have I regarded rossouwii speculatively as a synonym of H. mirabilis, due largely to the fact that it was recorded from the Bredasdorp area (and to the ambiguity of the type). Another reason was the similarity I observed between the illustration of H. rossouwii in Kakteenkunde with H. heidelbergensis var. minor and with H. mirabilis var sublineata. I opted for the latter (with which Scott was in accord) and the similarities are also recorded in the discussion above. I dismissed bicarinata as insufficiently known because I assumed that Triebner may have had some better reason to have equated his specimen with the Little Karoo H. mucronata. Possibly there has been a mistake in the Triebner numbers. It is this discovery of serrata so close to Bredasdorp, and equidistant from Napier, which revives and also casts new light on the problem. I conclude that my H. serrata is a later synonym of H. rossouwii of von Poellnitz.
There are still two other considerations. In my 1999 revision, the fact is that I placed paradoxa as a variety of mirabilis while discussing the possible connection of these elements via calcarea. In the light of this paper it is evident to me that paradoxa could also be linked to serrata rather than with mirabilis. The same is true for emelyae var. multifolia. I would not like to commit myself to this probable solution at this stage, as I think more evidence should be obtained. It is also improbable that, however attractive, the solution will not avoid impacting on H. emelyae, H. magnifica and H. heidelbergensis. It is also evident that the requirements of the ICBN are that should paradoxa indeed need to be incorporated into serrata, more name changes will result, as paradoxa will have priority over rossouwii. My opinion is that this is a fundamental weakness in the code, which inhibits speculative treatments, and also which results in smaller taxa becoming the typical main element of a species. My further opinion is that it is a weakness of the Code that the description of an element automatically creates an equivalent taxon. This may be acceptable and desirable where the elements are clearly separable and geographically defined, but not where the typical element is required to absorb any other variants which are not so defined. Thus H. paradoxa var. paradoxa could become the main component of a species where it is actually a geographical isolate. In a system which does not have an adequate general definition for ‘species’, this probably does not matter; whereas it might if there is a definition which suggests that the ‘species’ is an operating living system (see Bayer in Asklepios 77:3, 1999).
The formal statement of this paper, which would better be regarded as an hypothesis subject to test, is thus as follows:
H. rossouwiivon Poelln. in Kakteenk.7:75 (1938). Type: Napier near Bredasdorp, collected by Mr Rossouw, South Africa Police in Napier, Triebner 1059. Not preserved. Lectotype: designated by Breuer & Metzing 1998, icon in Kakteenk. non syn. H. mirabilis sensu Scott 1985, Bayer 1976, 1982, 1999.
H. altilinea var. bicarinata Triebn. in Triebner & von Poellnitz in Feddes Repert.Spec.Nov. 45:170 (1938). Type: Napier at Bredasdorp leg. Mr Rossouw (=Triebn.1059). Not preserved.
H. serrata Bayer, Jl S.Afr.Bot. 39:249(1973). Bayer :55(1976). Bayer :147(1999). non Scott :62(1985). Type: CAPE‑3420(Bredasdorp): Oudekraalkop, Heidelberg (‑BA), Bayer 166 (NBG).
H. rossouwii var. calcarea (Bayer) comb.nov. H. mirabilis var. calcarea Bayer in Haworthia Revisited :110(1999). Type: CAPE-3420(Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG, Holo.).
Material seen:- 3420(Bredasdorp): NNE. Buffelsfontein, Potberg (-BC), Burgers 2018 (NBG). De Hoop (-AD), C. Burgers 1648 (NBG), Bayer 6985.
H. rossouwii var. petrophila (Bayer) comb.nov. H. variegata var. petrophila Bayer in Haworthia Revisited:159(1999). Type: CAPE-3420(Bredasdorp): Renosterfontein (-AC), Burgers 2158 (NBG, Holo.).
The provisions of this solution are that the epithet paradoxa may be instated preferentially, and that emelyae var. multifolia and variegata var. modesta also be included.
A second provision is that the species H. bijliana v.Poelln. be rejected as insufficiently described and sourced, and that the type (the illustration in the Berlin herbarium) is ambiguous – as indeed are many Haworthia types and descriptions based on single plants and not correctly tied to geographical origin. This is demonstrated by the citation by VPoelln. (Feddes Rep. 1936) of this species viz. bijliana from Klawer and Eenriet and also Feddes Rep. in 1937 from Springbok – which cannot be anything other than H. arachnoidea. In this last publication he cites a specimen from Bonnievale, another from Ezeljacht Oudtshoorn and also sinks fergusoniae here which in fact was supposed to have come from ‘near’ Grahamstown.
H. bijliana Poelln. in Feddes Repert. 27:134 (1929); Bredasdorp, Mrs va der Bijl. Type van derBijl s.n. in (B). Not preserved. Lectotype desig. Breuer 1999, unpublished photo. icon. (B). Species rejecienda.
Acknowledgement I am grateful for the company on the field trip of Lawrence Loucka of Connecticut (USA), and that of my wife, Daphne. The assistance of Paul Botma of Cape Nature Conservation is acknowledged over and above the general acknowledgement to that Department. Mr. C.L. Neethling, Mr. Thys Swart and Mr. Pieter deKok were most kind and helpful in respect of access to sites. I am also grateful to Kobus Venter for support, advice, and constructive exchange of ideas. Not least to Steven Hammer, with comment from Bob Kent, who was so kind as to read this manuscript critically and make corrections. Residual errors will be mine.
Addendum Alsterworthia Vol 1:13(2002) “A comment on Bayer’s latest taxonomic contribution” by Ingo Breuer. [link] With reference to the comments in Alsterworthia 1:13(2002) regarding the identity of H. rossouwii and H. serrata:
Notes
Fig. 27 H. rossouwii. Vpoelln. type.Fig. 28a. MBB6984.1 H. rossouwii. Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.Fig. 28b.Fig. 29a. MBB6984.1 H. rossouwii. Colour photo originals of the preceding. Fig. 29b. Fig. 30. MBB6984.2 H. rossouwii. Nooitgedacht, Bredasdorp. A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second. Fig. 31. MBB6983 H. rossouwii. Soutkloof, N Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might. Fig. 33 Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Fig. 27 A copy of the von Poellnitz lectotype of H. rossouwii.
Figs 28a & b Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.
Figs 29a
& b. The colour originals of the preceding.
Fig.30 A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second.
Fig.31 A photograph of a plant of MBB6983 Soutkloof, north of Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might.
Figs 32a
& b. Scanned and manipulated copies of figs 28a & b.
Fig.33
Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Figs.7a, b & c. These are clones of JDV86/5 H. heidelbergensis var. minor, Rooivlei, NNE Bredasdorp. This is a small population, of also small plants, from northeast of the collection MBB6984, Nooitgedacht. The plants partly resemble H. rossouwii from there, and partly the var. petrophila from Karsriver. Classifying them as a variety of H. heidelbergensis is not necessarily correct. Considering habitat and substrate, one must consider that ecotypification is playing a role, and that was habitat available, there would be further populations which would confirm continuity with these known collections.
Regarding Figs.28a & b. A bit of play with light and focus and the keel and under-leaf spotting could be varied. I would like to suggest that they be considered them faded, less sharply focused, printed with less contrast and on a coarser grained paper. Figs.32a & 32b are in fact such scanned copies of Figs.29a & b. I do not think it even necessary to consider that the original rossouwii of Fig.1 endured a long journey in the post to a foreign land. The assertion by Breuer in Alsterworthia is that not only are the plants I figured as H. rossouwii different from Fig.1, but that this Fig.1 lectotype is “probably” of the maraisii-complex. This is at variance with…
1. the Triebner description of the same collection as a variety of H. mucronata;
2. with Col. Scott who regarded it as H. mirabilis. What to say of von Poellnitz’ own assessment – the man who described both species i.e. maraisii and rossouwii in the first instance.
3. with the facts of the pictures before you.
What should an observer make of maraisii growing very near to rossouwii at Oudekraal and near to rossouwii at Napier? Can we really see a closer resemblance of Figs 27 & 28 with those long attenuate, double-curved, thin leaves – to H. maraisii rather than to H. serrata. I truly do not see that such a judgement can be seriously considered in the light of Breuer’s interpretations. It is of course valid when one considers all the variants that I illustrated in my article above and in terms of my over all classification.
Generally these plants of the SW Cape tend to have the double curvature of the leaves as is evident in the VonPoellnitz lectotype, but one can find in the same population and/or taxon, clones with incurved, outcurved or erect leaves (see fig.30). Broadly speaking H. maraisii has thick recurved leaves. But this is a non-argument even if one (outside of a “lumping vs. splitting” red-herring) considers the complexity of the so-called maraisii-complex, and I am sure that Breuer does not. I frankly think that most observers cannot even dream how complex it is and that it is just braggadocio to write as though anyone can. Breuer implies that one can make the inference of similarity between rossouwii and maraisii from the illustration (Fig. 27), the description and its given locality. This is patently and self-evidently false as is evident from the fact that I used this same information to reasonably deduce the similarity of rossouwii and serrata.
I did not originally submit a copy of the von Poellnitz lectotype, perhaps because I felt that the response “you will see that rossouwii is quite a different plant from serrata“, is what I could have expected from an uninformed audience! The fact is that all the evidence, circumstantial and descriptive, show that they are the same. Including the lectotype would have just been “overkill” and I have never before felt required to write for a readership with less than common sense or basic knowledge, nor for obstructive readers.
It should not be necessary to say this, but the character of the flowers cannot be specified in the way that writers are inclined to suggest. They have this same enormous range of variability and it is just fraudulent to say that there are floral characters by which these taxa (the varieties of H. rossouwii which I now recognise) can be unmistakeably differentiated. It is the typological concept which drives people to think there is a single description which can encompass the description of a taxon. This is nonsense. It is a statistical problem and we have the difficulty of establishing the probabilities associated with “decision”. Breuer writes “… von Poellnitz’ description probably depends only on 1 or very few selected plants and no other records are reported for this taxon”. This is circa 1945 circumlocution, it is unnecessary and it is untrue. There was never a question of “selection”, and furthermore we have the Triebner description of the same collection as H. altilinea var. bicarinata. Why this sudden doubt about the interpretation of types. It is a shifting of goalposts.
There are few collections in flower now in April, but I could take this picture of three flowers from the one collection MBB6672 H. maraisii – Koningsriver, SW Robertson (Fig.33). It is not an exceptional case or an exceptional collection. But one can see the differences in thickness of peduncle and variation in the individual flowers. It can happen like this, albeit to lesser degree, on the same plant. Colour is also frequently quite variable between and often within collections. Many years ago I undertook to illustrate all the flowers in my maraisii collections. But they were so variable that I just gave it up. The variability in the flowers mirrors the variability of the rosette and the latter is just a little easier to deal with. One has the added problem with flowers, of the changes which take place during the opening and aging process. So then how does one define the flower, say, of H. mirabilis. It cannot even be done for the range of leaf rosettes, and this has now been shown in the literature many times.
May I also point out that one of my understandings of botany and plant classification is that there is a tendency to overreach ones talents. I have freely admitted that I am there because of the paucity of talent. I frankly do not think the environment is enriched by the particular article I now refer to in these added notes.
I can point out that there are writers who really persist in flogging dead horses. The result is phantasmagoric fanfaronades. The article concerning the name H. correcta (Alsterworthia 1:6, 2001) is not particularly accurate either and indeed it may be my own fault. Smith explained that Mrs. Blackburn did not in fact collect correcta ‑ it was a Mrs. le Roux and the plants came from near Vanwyksdorp. Later Mrs. Ferguson sent a plant from 26 miles east of the town. Thus from this information H. correcta has to be a later synonym for H. emelyae.
Regarding types:- the painful realisation was actually expressed by Schelpe in the distant past, and brought home to me many times. These old types are doubtful and ambiguous. I was faced with that option long ago – just forget them all, or try to reach a compromise with the understanding and information available at the time. H. rossouwii is a case where there is real new information which would have made me use the name rossouwii in 1976 and before, had it been available. But here, had I persisted with the name serrata for this new material MBB6983 and 6984, there is absolutely no doubt in my mind that I would have been criticised for failing to see the connection to rossouwii. Not that this would concern me unduly, for I think there is actually very little benefit in this mindless re-interpretation of types which takes us nowhere. Notwithstanding, and without compromising, anything so far written:- the really disconcerting thing about the whole matter is that without the locality data, it will be possible to generate an argument bolstered by illustrations, to show that rossouwii was in fact H. decipiens, H. cooperi or H. herbacea (among others), of the option less improbable than H. maraisii.
The closing paragraphs of the article are a rather hackneyed parody sung by most “wannabe” taxonomists. Of course it is right when Breuer says if we lump it means we would have to lump many other things. It is just as true to say, if we split we have to split many other things. That does not make sense either. This is ancient stuff of 60 years ago and it seems strange that it still provides so much fodder for writers, who have in fact no rational species concept to begin with. The Linnaean system was based very largely on floral characters and hence taxa at all levels are based on “related floral characters”. To suggest that this pertains in some special way to “series” is not correct. This sums up the article in Alsterworthia. ♦
This essay was published in Haworthiad 16:86, 2002.
I wrote an article about Haworthia rossouwii in Aloe 38:31(2001), in which I resurrected this old name to replace that of my own H. serrata. This was necessary because I had found this plant (because of its localisation and its abundance there, it is better to say ‘this species’) at two places near Bredasdorp as opposed to where I had described my species from near Heidelberg.
One needs to know something about the geography and geology of the Southern Cape (and the Overberg as a part of it is also known) to really follow all the ramifications of any discussion about Haworthia, including this one. In fact one needs to know a whole lot more, and I will also try to explain that and its implications for the collector and Haworthiophiles. This “whole lot more”, I will call the Corporate Mind because it includes so much – so remember CM! If I regard H. rossouwii as a species, I have to consider all the plants and all the places where they grow in order to determine the nature of this particular system of living things. As I explained in my article, there is a problem with the fact that little is actually known about the Haworthia of the Overberg. They occur in small populations scattered over a fairly wide area which has been heavily impacted on by agriculture. Thus about 90% or more of the Overberg is now wheatfield or pasture. Like Gasteria carinata, which is also a Southern Cape species, Haworthia is associated with rocky outcrops and thus also with the geographical erosion and drainage systems of the area. It is quite probable that cultivation has had relatively very little impact on Haworthia in terms of actual available and suitable habitat.
The problem is to get to all the small potential habitats and explore these at a scale which will help to resolve the problems of variability, relationships and classification. While writing about H. rossouwii, I was very conscious of this problem and the fact that my explanation is not complete. There are some collections by Chris Burgers which I have not seen, and there are many potential habitats which I have not explored either. The place Kathoek and H. variegata var. modesta which grows there is a particular case in point. Kathoek is at a point midway between the Breede River and Bredasdorp. It is just east of the De Hoop wetland/stream and on the northern edge of the Coastal limestones. In the west, also on limestones, is H. rossouwii var. petrophila which I first placed with H. variegata. In the south (also on coastal limestone) is H. rossouwii var. calcarea, which I originally had with H. mirabilis. To the north is H. variegata var. hemicrypta which is on Silcrete of a quite different geological origin. But H. variegata variants, as I interpret the populations, are also on and about the Potberg to the east which is Cape Sandstone.
My problem is “How do these populations actually relate to one another?” What is missing? What can I find to better explain their similarities and their differences? So at the back of my mind is the wish to explore the Coastal limestones between Bredasdorp and the Potberg and find out more. But it is not as easy as that. I also want to know more about H. mirabilis, H. maraisii, H. heidelbergensis, H. mutica, H. minima, H. marginata and H. pumila, which all also occur in the Overberg area and each have question marks attached. They each present problems that I would like to resolve for my own satisfaction, without being confused and befuddled by others who are toying with the same body of knowledge at different levels of expertise and ignorance. In this play we are using words, terms, names and conventions. For this play we need to have a commitment to truth and use the same language or else it is all just meaningless.
So we come to what took me to the Overberg in November 2001. Was it was just my curiosity about all those inviting rocky unexplored places? Is it to learn something more myself? Is it to impress anybody? Actually it was many things including a fascination with chameleons – largely my wife Daphne’s. In March 2000, Daphne and I had come across a most remarkable chameleon at Napier. It was on a small shrub among whitish dried grass growing on a calcareous outcrop strewn with white quartzitic pebbles. The tiny reptile was ivory white, with a stunning array of small orange tubercles on sides and back. One has to know the Cape Dwarf chameleon (Fig.1) and its cousin, the Robertson Dwarf, to know how beautiful and amazing these creatures can be. Another thing was H. mutica and my fascination with collections of this species and their histories of discovery and exploration which are stories of their own. Still another was the news that the Botanical Society had funded a researcher to explore the remnants of the vegetation of the wheat areas. What had he found? Apart from Haworthia, I had once been involved in a preliminary attempt to map and identify vegetation remnants for conservation purposes, and had vainly tried to draw attention to the curiosities of the Silcrete outcrops and remnants of vegetation among the wheatfields.
This researcher was an old acquaintance, Nick Helme, who kindly told me what Haworthias he had seen in his exploration. There was not much new, except for H. mirabilis east of Caledon, and this became our first stop. The site is south of the main area for the species which is along the Riviersonderend river north of Caledon. It does, however, occur in recognisable form south-east of Caledon, also around Napier where it transmutes to some degree (inter alia to the var. badia), and then also at Bredasdorp. There it also transmutes to the var. sublineata (sandstone) and var. mirabilis (the old mundula – and there may be some other way out, but for me this represents the low cussedness of botanical nomenclature). What H. mirabilis does to the north and east of Riviersonderend actually fully explains the problem of classification of Haworthia, and who knows if anyone will ever write that explanation – and truthfully.
Nick Helme’s site is a deep valley which drains to the south. Typical of the area, are these small drainage areas which seldom have permanent water. There may be odd pools lasting into mid- or late summer. There are often exposed shale ridges which host a particular array of succulents and other species – and are often home to owls too. The first problem is access and where does one find the landowner after one has succeeded in finding the place? If one does find the dwelling, will it be the right one and will one find the landowner. Will it be a big aggressive StBernard and more aggressive and ugly Rottweiler reflecting the mindset and security fears of the owner and grossly offending my own, like once near Worcester – or will it be a lonely and over friendly sheepdog. November is a very bad time. This is when the wheat harvest is on. At the end of a bad season, when the crop is drying and it has to be cut and reaped the weather is playing its usual strange tricks, farmers cannot be expected to be approachable. But we did find a helpful soul. From the farmhouse, we drove twenty kilometres back to the main road to the roadside stall, where a family member, Herman DeKock, gave us the necessary permission to venture onto the property.
It does seem an easy business, but in fact it becomes very complicated. Nature Conservation regulations are strict, and land ownership, presence, trespass, stock theft, personality and who knows what else, influence the mindset when setting out an expedition of this kind at all. With those problems all out of the way, and only then, is one able to really enjoy being out in the field and communing with nature – which it is all about in the first place. So we found Nick’s site. We climbed through the fence (s) and tried not to disturb the forlorn sheared, miserably self-conscious sheep that clustered around the earthen dam. It was hot and dry and the most promising spots were quite a long way off. One has to plan one’s approach as the southern slopes are often quite densely vegetated and very uncomfortable to traverse. Snakes are also a consideration and especially in early summer. On this trip we never saw one, although Daphne surprised, and was surprised, by a Lynx drinking at a waterhole. So we did find the plants that Nick had said and took a photograph for the record (Fig.2). Just ordinary H. mirabilis var. triebneriana (cussedness – there should be no typical variety, just a broad general name mirabilis).
From here of course we went to the chameleon at Napier. What a laugh! We had difficulty finding it on the same day on the same bush, five minutes after first seeing it. So expecting to find any chameleon at all, and I have never seen a chameleon in the field in all my years of plant exploration, was quite the wildest dream. But with a dossier of 60 individuals recorded on our small-holding at Kuilsriver over a period of about a month, we felt entitled to hope (all contributing to CM). It is also a most wonderful excuse to be out in the country.
We headed for De Hoop. This is also quite a story. Where do we stay? The DeHoop Nature Reserve is a wonderful place with great accommodation, but it is not where we wanted to be. The staff does not know what it is we are looking for, what our CM data base contains and what our needs are. We want to see the area around the north of the wetland. This is half private land, and half state-owned. Besides there is a travellers Lodge on the edge of the reserve which we understood offered easy and official access to the reserve, presumably to near where we wanted to be. Not so. It is one thing dreaming about this exploration while sitting at home with the map, and quite another to be faced with the problems of roads, fences, ownership, dogs, snakes, flies, mosquitoes, permissions, heat, cold, wet, terrain, bush, visibility, time, distance, misinformation, local “knowledge”, incipient old-age, food, water and the spirit of inspiration and energy which drive the legs (CM). So the Lodge was an unfortunate mistake (we did look, and we did also look where we were told an Haworthia did grow – it was Gasteriacarinata), and we did end up at the office at DeHoop Reserve. You see over the years, one comes to realise that helpful and kind as people by nature are, an awful lot of time is spent chasing red-herrings and wild geese (CM). The help and information one gets is very often of a negative hindrance rather than a positive help. So my real gratitude to the manager there, Mr Ben Swanepoel, who without serious qualm, entrusted us with the key to the northern part of De Hoop where we needed to be.
Unfortunately the problems listed above rather limited our activity. We exhausted our energy climbing up and down rocky slopes in an area which should have occupied us for a week. It is an area which has to be explored by someone at sometime. We found naught, which should not in the least be interpreted to say there are no haworthias there. The terrain is formidable and also most promising. But CM called.
We moved to accommodation closer to our budget restraints and nearer to our goals. Driefontein, where the owner graciously made the necessary contacts for us. What a place – the cold vegetated coastal limestone behind, and featureless cultivated endlessness in front. I have an old record of a collection of mine of H. mutica from “Beyersdal”. Subsequent travels indicated that place, name and memory did not match. So we went to Beyersdal. We did find mutica and it was a different place and a new record (Figs.3a, b & c). Who can actually describe the wonder of plants like these? In tiny patches of rock in a sea of wheat and low-grade (by rainfall and vegetation standards) pasture. Also the quiet, the two owls, also Gasteriacarinata again, this time the Haworthias among them – unusual. The extraordinary colour of the plants. This time the bronze colours of badia. Nearby we also visited an old, by thirty years, locality for H. heidelbergensis. Still there and astonishingly in among white quartz in a small outcrop, but in two little groups occupying hardly two square meters in the approximately two hundred available (Fig. 4).
Then we went to Vanderstelkraal. We had met the owner, Mr Adam Albertyn, who very kindly let us wander over his farm. CM. How does one explore a place like that without living there? It is on the edge. The agriculturally useless coastal limestone at the back, and the productive massively utilised Bokkeveld fields at the front. If terrain had allowed, there would not be so much as a hedgerow to have conserved anything. We saw some very interesting sites and exhausted quite a number of things, like energy, time, passion for the quest, tolerance to bad weather, ostriches, tight-strung barbed-wire fences, no roads and also to the heaviness of the overwhelming destructive productive activities of man. We did not find Haworthia. But I omit a lot, including all the good farming. Mr Albertyn owns the eastern side of the De Hoop wetland where he has a 4X4 route – and there is also a sixty-year old legend of H. minima in that particular area which I would like to confirm. Not this trip though. We did try to the west later, where CM eventually prevailed against us – aided and abetted by overgrowth of alien acacias and the negative vibe they seem to generate (by the side, at Vermaaklikheid, Riversdal, H. mirabilis var. paradoxa grows/survives under Acaciacyclops).
Then we decided to climb the mountain behind Kathoek. Why? I cannot remember why I climbed there in the first place, or how I even did it. Then I found plants in a burnt out area, under the remnant blackened skeletons of Protea – natural fire cycle encouraged by farmers. It does produce a grazeable growth and also invasive aliens. The plants of H. variegata var. modesta were just discernible sticking out from loose sand. Kobus Venter later also visited Kathoek and found plants which were more robust. The plants have spreading variegated leaves (in cultivation?) and while I had no real problem associating them with Karsriver, viz. H. rossouwii var. petrophila, they seemed to have a greater affinity with variegata at the Potberg and Luiperdskop. But my collection and Kobus’ seemed at variance. So Kathoek was a must. From the road CM prevails and it looks quite an expensive option in terms of CM resources. Actually the climb was quite easy. We found the plants before we expected to. They did not look like what we expected them though. They were densely clustering (expected) and in rock crevices (Figs.5a, & b) and pans (Figs.6a & b). The rosettes were tiny and reminiscent of even H. reticulata (see Fig.7). At one place a dense cluster under and in a restioid grasslike plantlet, looked just like a yellowish-brown windblown fallen mass of dry leaf bracts (Fig.8). What impression did it make? Nothing new. It provoked the existing doubt. The population is very similar to petrophila and less so to variegata and hence this variety, with the exclusion of the Potberg population COULD BE transferred to H. rossouwii. But this does not help much, because the Potberg var. modesta is less like the northern Potberg’s two populations and the Luiperdskop population of H. variegata var. hemicrypta, and not more like petrophila! So the CM has to carry this anomaly together with the position of calcarea until whatever else to be found is found. These finds are not going to make things easier to resolve in the CM, nor to explain in a way which assuages the CM of anybody else.
All the while we were pondering other clues to the nature and relationship of things. Like the endemic Braunsiivanrensburgii; a curious specimen of Carissabispinosa so laden with large red fruits it resembled a Pyracantha; Euphorbiasilenifolia everywhere, and what a strange distribution this summer-deciduous geophyte has; and what about Asparagus; what is the relationship of A. capensis, A. mariae, A. suaveolens and A. stipulaceus? Where was Aloe brevifolia or A. saponaria? What was that pugniform Euphorbia at Driefontein or the bayeri-like one at Vanderstelkraal?
Unexpressed CM – we are using my son’s LDV as in my own car anything below well-maintained road is not negotiable. CM – the cellphone we arranged for failed. There is a call in my sub-conscious ‘vehicle needed back home’. We end the trip a day earlier and the message turns out to be true as there is a breakdown of one of the other delivery trucks.
On the way home we pick up some loose ends. We call at a spot north of Bredasdorp where in my lucky ignorance I once found mutica. In several subsequent visits I could not locate it, until one year Daphne and I together with Lawrence Loucka had a very thorough search of the small spot. We found about five plants in a spot about 1/2sq meter. At a later visit we found that the plants had seeded and there were many seedlings but crowded together with seedlings of Aloeferox. Generally Aloe and Haworthia do not mix. Aloe need more root room than Haworthia and Haworthia cannot tolerate competition from larger plants. So we thinned out both aloe and haworthia seedlings. This visit revealed the successful fruits of our activity (Fig.9). The two bigger adult plants were almost being pushed out of the ground by a burgeoning aloe seedling, but there were about 15 other vigorous and healthy plants to carry the group forward. CM – this was an interesting repeat observation of the small distances that seed and seedlings actually do commonly disperse. I have seen the same thing with hybrids where the hybrid is often within the multiple body of one of the parent species.
Mission ending, we made our last visit to Drew. This story is written more fully elsewhere. We had found the pollen source for a plant of H. mutica which I collected there more than thirty years ago and which Kobus and I had taken to the field again to see if seed would set. This surviving single plant is now a collector’s dream which I dub “Silver Widow”. No one had succeeded in finding plants around where a huge centre-point pivot installation now graces the landscape. We were sure the plants were gone, and taking the plant back there was a kind of forlorn gesture of regret that more had not been done to conserve the population – and that so much had been done to eradicate the plants (two hundred plants had been collected and sold to Triebner circa 1935). If anything can be done or could have been done at all to have saved its family? But it worked and we got seed. Where did the pollen come from The pollinator is mainly a solitary bee with a fairly limited flying range. The pollen parent had to be somewhere near. Daphne and I had made several excursions to add to the many different attempts that are known to have been made to find them. Finally I decided that we just had to look again and the plants had to be where we had failed to find them before. So what happened? We left the farm house and walked straight to the spot (getting ‘stung’ by the electric stock fence on the way) less than 300m away and probably about 150m from where we had left Silver Widow to be pollinated. There were about 35 plants (Fig.10) in an area of about 3sq m in flower under Eriocephalusafricanus. So now we returned to successfully collect seed.
The curious thing in CM and all this discussion about big things like Continental Drift, and Biomes, and Vegetation, and Habitats, and Ecotypes, is that H. mutica is not particularly concerned. Here at Drew it grows in an ancient river-boulder situation in what we can call renosterveld under Eriocephalus. At Beyersdal it is on the edge of a stream-bed on exposed Bokkeveld shale with Gasteriacarinata. At Hasiesdrift it is on a gravelly bank (Bokkeveld shale) with dense Aloeferox. Near Napky it is on silcrete with grass. West of Swellendam, it is on whitish-clay, probably associated with silcrete and growing within a restioid species and with Gibbaeum esterhuyseniae. I have also seen it on a fairly bare hillside with a scattering of quartzitic pebbles where the parent shale is too dense and shallow for cultivation or to support denser vegetation. The places hardly have anything in common other than their more general geographic containment, climate, and low biomass potential. How, ever, did they get to their widely-spaced, isolated island sites?
So the tale ends with the reflection on the criticism that I have not treated H. scabra and H. starkiana in the same way I have H. retusa and H. mutica. I have been “inconsistent”. Readers can research and educate their own individual CMs (corporate minds) to the point that they can form opinions which are consistent to the point and purpose of any communication. Aren’t we lucky that the Robertson Dwarf chameleon is a small animal (Fig.11 – by Scott Russel).
Acknowledgement: This fun work is really just about people. Daphne, who kept me company. Janet Albertyn at Nachtwacht, Herman Dekock of Jongensklip, Thys DeVilliers at Boskloof, Tom Ambrose at the cement-block operation at Napier, Rory Allardice at Buchu Rest-camp, Ben Swanepoel at De Hoop Cape Nature Conservation, Annette VanEeden of Driefontein, Mr. Dawid Beyer of Beyersdal, Adam Albertyn of Vanderstelkraal, Theo Blom of Soutspanvlakte, Jan Joubert of the adjacent farm, and Beth and John Humby of Sanddrift. Their good-will and kindness is unforgettable.
Fig.1 Cape dwarf Chameleon – Bradypodion pumilum. Markings, colour, tubercles and spines are individual characteristics. It is said that even the genus is in doubt!Fig.2 Haworthia mirabilis var. triebneriana, MBB7059 Jongensklip, Caledon. Note the reddish spines.Fig.3a Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. This plant is exposed to full afternoon sun for at least a few hours each day. Note the mucronate (pointed leaves).Fig.3b Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A more general specimen with rounded leaf-tips.Fig.3c Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A particularly compact specimen. In this population, the waxy sheen of the species seemed less conspicuous.Fig.4 Haworthia heidelbergensis var. minor, MBB7063 Beyersdal, Bredasdorp. Varietal names are just a geographical convention, as indeed even the species which I recognise, seem to be.Fig.5a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster under a small rock overhang.Fig.5b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster in an exposed crevice and visible as a dark mossy fill.Fig.6a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill.Fig.6b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill – a closer view.Fig.7 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Very reminiscent of H. reticulata say south-west of Robertson.Fig.8 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Barely recognisable to the undirected eye.Fig.9 Haworthia mutica var. mutica, MBB6982 Hasiesdrift, Bredasdorp. Nine seedlings in a tiny island of plants, in a bigger island of Aloe ferox and surviving vegetation on the edge of a wheatfield.Fig.10 Haworthia mutica var. mutica, MBB7064 Sanddrift, Drew. This lovely plant is nearly 120mm diam. and the largest of about 50 plants. They survive, and have survived in this small heavily impacted area for so long. There is no where for them to go when options must have been very limited to start with.Fig.11 Robertson dwarf – Bradypodium gutturale, photo by Scott Russel. Zoologists cannot determine if the Robertson dwarf is different from the Karoo dwarf, or from the two subspecies of the Namaqua dwarf. They use the same fuzzy logic that botanists do, plagued by low sampling and difficult application of statistical principles and probabilities.
Since the publication of Haworthia Revisited (Umdaus 1999) I have written Haworthia Update Vol.1 (Umdaus 2002), and Haworthia Update Vol.2 is now in press. It has some 700 pictures and several maps and focuses largely on the vexing question of how we classify and name the plants in the social environment we have created.
In one chapter I demonstrate how difficult it is to distinguish H. mirabilis and H. maraisii and admit that I can no longer see that distinction. In this article I want to illustrate a population which shows that my problem is not limited to those two names. This is about a population on the farm Klippoort that is at the extreme southeast of the Worcester/Robertson Karoo. It is south of the Riviersonderend River just before the confluence with the bigger Breede River. The Riviersonderend runs south of the range of mountains which effectively forms the northern boundary to the distribution of H. mirabilis. East of Stormsvlei, the river cuts through the tail of the mountain range and turns northeast to link up with the Breede River. The confluence lies just south of the Bonnievale/Drew/Mardouw area which is populated by a dense array of variants of H. maraisii and H. heidelbergensis that cannot confidently be regarded as two different species either.
I have given my record a new number MBB7513, because I do not think I ever made a formal record. I am not even certain when I first saw the plants there at Klipfontein but I do know that I was again there in 1996. I was familiar with plants in the nearby area which I had no doubt were H. maraisii and remembered the Klippoort plants as small with rather erect incurving leaves. In October 2005 Kobus Venter, my wife Daphne and I were in the area and saw a Drosanthemum which I knew was important to Dr H. Hartmann. To show her these plants in February 2006 I had to get permission from a new landowner and this led to a curious exchange with the original owners (Mr. and Mrs. Urschel) who had retained the main part of Klipfontein. To demonstrate the significance of the area and to fuel her interest, I thought I would show Mrs. Urschel H. maraisii on the part of the farm they had retained.
After much searching in the very dry conditions then prevailing, we found one single clump which was coming into flower. We decided we would try to get back to see if any seed would set and to collect and grow same to increase numbers and replant in a seemingly disturbed habitat. We returned in mid-March to find the plant had lost its flowers, had been disturbed by some animal or other as one head had been dug away (see Figs. 1 & 2) and was in bud again. Unfortunately my digital camera ran out of power and I could not get a better picture. We then searched again quite thoroughly, but now aided by the fact that flowers were appearing. We found many plants hidden in the moss and litter under shrubs and also in the dense lichen on the huge conglomerate boulders forming the rim of the very small rocky hill. Anxious to get seed and a good picture, we again visited the site in early May after it had rained. Most of the seed was gone and we virtually gave up hope of finding any and set about trying to get some pictures. To our surprise and despite the thoroughness of our previous searches, we now found many more plants and in two small spots they even formed large dense clusters. There were also mature green seed capsules and some ripe seed – and also plants in flower and in bud.
Fig. 1. MBB7513.3 H. maraisii. Klipfontein, Stormsvlei.Fig. 2. MBB7513.4 H. maraisii. Klipfontein, Stormsvlei.
The curious thing for me was that I had no doubt up to this moment that the plants were H. maraisii confirmed by the dark grey-green colouration of fig 1. So you may not imagine my surprise when the first cluster I saw was a very green plant with many spreading leaves and notable end-surface translucence viz. H. heidelbergensis (figs. 3a & b). I also had a memory of plants rather as I have written about near Robertson, which are pubescens-like with surface fuzz (see fig. 4a, b & c). The plants do indeed have surface fuzz and also the plants figured are solitary and with incurving leaves.
Fig. 3a. Klipfontein 3rd May 015Fig. 3b. Klipfontein 3rd May 016Fig. 4a. Klipfontein 3rd May 006Fig. 4b. Klipfontein 3rd May 014Fig. 4c. Klipfontein 3rd May 012
Fig. 5 illustrates a plant which has the more rigid and less translucent leaves of H. maraisii, and figs 6a, b and c are of similar solitary plants. Figs. 7a & b are clustering clones. The plants are all small with the rosettes seldom more than 30mm diam., although the spread of leaves of a clone as in 3b may be as much as 70mm. The flower stalks bore few flowers and one is illustrated in Fig. 8 although I have been able to make little use of flowers in drawing any conclusions.
Fig. 5. Klipfontein 3rd MayFig. 6a. Klipfontein 3rd May 008Fig. 6b. Klipfontein 3rd May 010Fig. 6c. Klipfontein 3rd May 013Fig. 7a. Klipfontein 3rd MayFig. 7b. Klipfontein 3rd MayFig. 8. Klipfontein 3rd May
The habitat is a curious mixture of plant species. There are milkwood trees which can grow large, as well as Acacia karoo the common sweet thorn and Lycium which is also a medium sized spiny shrub. These are often along watercourses and here the Riviersonderend is barely 50m distant. But there are also Renosterveld species such as renosterbos itself, and karoid plants. The large conglomerate boulders are pocked with missing tennisball size stones, so that the dense lichen cover has many niches for plants. Apart from miniatures like Schizobasis, Holothrix, and Drimia pusilla there are also miniaturized Crassula, Adromischus and many tiny seedlings of the large Tylecodon paniculata. My observation is that the vegetation of many such sites bears very little relation to vegetation maps and plant associations mapped at large scales.
The site is fairly disturbed as Bluegum and Black wattle are invasive along the river. The farm itself was controversially planted to Redriver Gum and Pinus pinaster and planted gums impinge so closely on that site that Haworthia can be found in the stone and moss among the closest planted trees. The greater area is very marginal for farming on very rocky soil marginal to the sandstones of the higher mountains so that forestry option seems a very unpromising and unfortunate one. A larger similar rocky area to the northeast has been, and is scheduled to be mined again, for manganese. There is nothing to suggest that the Haworthia ever occurred there. Looking north across the river to the Bokkeveld shales one again sees the virtual ecological desert of total cultivation. Very few habitats are apparent. It is often speculated that many plants (particularly specialists like Haworthia) must have perished due to this cultivation. But my impression, based on the known collecting record of the last 80 years as well as on my own experience of where the plants occur, is to think it actually very unlikely that plants did occur where they presently are not. I followed up some records made in late 1930 by messrs. Otzen, Luckhoff, Herre and Malherbe and am confident that plants can still be found at most of their sites. Similarly the localities recorded by Dekenah and Smith are similarly still extant although they must be seen as under threat. What does cause concern is that the lost habitats are largely small rocky sites with surface rock that are targeted by road builders. Road maintenance has become a major problem because the road reserves which used to form uncultivated strips of natural vegetation are now cleared fence to fence in the case of new roads. In the case of existing and minor roads there is a constant attack by road clearing gangs, fire and weedkiller spraying. In the agricultural areas there are huge expanses I see as ecological deserts. The rocky and skeletal habitats are under severe pressure from animals and the introduction of new sheep breeds with more aggressive feeding habitats is a severe threat to these small islands. Farming has also become more of an industry and business and production is the goal irrespective of conservation ideals. This particular habitat seems to be as safe as it can possibly be by virtue of its intrinsic nature and in respect of the above threats.
Conclusion The distinction between H. maraisii and H. heidelbergensis is too tenuous to enable positive identifications and this Klippoort population displays more obviously the problem of variability within populations.
Acknowledgement Messrs. Leanne & Pieter Urschel, Klippoort East. Messrs. Cindy and Jon Webber, Klippoort West.
In an earlier article I described how Haworthia floribunda (at B on map) transmutes eastwards to H. variegata (at A on map) between the localities Klipfontein and Kleinberg, which are north of the Potberg Mountain. This is despite the fact that they both occur in close proximity at the northwestern end of the mountain. Difficulties now arise in the immediate vicinity to the west (at point W) and this extends northwestwards (to points C and D on the map). While we can confidently ascribe names to floribunda and variegata at those particular sites, the plants to the west and northwest are confounding. They fall into a no-man’s-land of these two species with H. mirabilis, H. maraisii, H. heidelbergensis and even H. mutica thrown in.
Localities and their names can also be confusing, so they are listed as follows:
A – the north and western point of the Potberg mountain on the De Hoop Nature Reserve and adjoining both Klipfontein and Juliesfontein farms (Juliesfontein farm has been renamed Poteberg Farm)
B – Byeneskop, a small hill on the western boundary of the farm Klipfontein
C – the farm Brakfontein
D – a hilltop north of Brakfontein named Witklip Kop
W – the two farms at Die Kop, Wydgelee, at the entrance to the De Hoop reserve
Figs 1 to 4 are from immediately west of A and show characteristics of floribunda in respect of the leaf tips. Figs 5 & 6 are of plants also from close by and the influence of H. maraisii is more evident. Generally the leaves are much shorter and stouter than one would expect in H. floribunda and the small size and numerous leaves suggest H. heidelbergensis. Both pointed leaves and leaves with the flattened and rounded tips may occur on the same plant (see figs 7 & 8).
Figs 9 to 13 are of two collections on the same ridge to the south of the Brakfontein homestead and are generally similar to the plants in figs. 1 to 8.
9. MBB6886 H. heidelbergensis – maraisii Brakfontein10. MBB6887 H. heidelbergensis – maraisii Brakfontein11. MBB6887 H. heidelbergensis – maraisii Brakfontein12. MBB6887 H. heidelbergensis – maraisii Brakfontein13. MBB6887 H. heidelbergensis – maraisii Brakfontein
Figs 14 to 22 are plants from a small hill to the north of Brakfontein. There were few plants there as the hill is severely grazed by sheep. There is quite a strong resemblance to H. mirabilis in that the leaves are firmly sub-erect and slightly more robust. The hill is very typical of the ferricrete/silcrete/gravel inselbergs which occur throughout the southern Cape. They are characterized as hard, conglomerated, solid rock that caps the hills, and these are underlain by Bokkeveld shale that has decomposed to kaolin. Exposed Bokkeveld shale is the dominant parent material for the soils of the area and these are arable, whereas the rocky inselbergs are not. The consequence is that habitats suitable for Haworthia are the relatively un-weathered skeletal soils which can only support sparse vegetation. Thus distribution of the plants is island-like and no doubt this plays a significant role in the way the plants relate to each other. ♦
14. MBB6890 H. heidelbergensis – maraisii N Brakfontein15. MBB6890 H. heidelbergensis – marasii N Brakfontein16. MBB6890 H. heidelbergensis – marasii N Brakfontein17. MBB6890 floribundoid leaf18. MBB6890 floribundoid leaf19. MBB6890 more pointed leaf20. MBB6890 more pointed leaf21. MBB6890 more pointed leaf22. MBB6890 more pointed leaf