
The writing of my grand finale was inspired by several things. One of these was another item of a mind-numbing foray into the classification of Haworthia. So I asked that deep thinker and observer, Gerhard Marx, for a devil’s advocate (abbrev. DA) point of view which he has done with the same competence he has as an artist. I have many times in my writing addressed the issue of a species definition and produced one too. Not surprisingly the first thing the DA does is dismiss my definition without producing one of his own. Simply being able to say that an indeterminate number of plants from some population are sufficiently different in respect of a character or two from other haworthia, is motivation enough for the generation of a new name?
The case of H. groenewaldii Breuer, described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald is the case in point. It presents the description of this supposed new species from Buffeljags east of Swellendam. The article is written in the first person (Breuer) who quotes extensively from Gerhard’s e-mails, and includes a piece by Jannie Groenewald under the heading “Description of the Vegetation type and distribution”. The overall impression is of an article that conforms to the style of a forgotten era and it is not possible or sensible to attempt a rational dismissal. Who is actually responsible for the article and how does one correct misleading statements without giving offence?
There is probably only one main issue that can be made level-headed sense of. This is the statement in the article…”another (it is not clear what the other was) striking feature was the fine but rough leaf texture…” Then quoted again…”the typical H. mutica grows only 20km to the west at Dankbaar”. But the Dankbaar plants also have the same shiny leaf surface. The word “typical” is generally overused in the article and I would counter claim that the Dankbaar plants are typically H. mutica and reinforce my argument for the similarity to the Buffeljags plants. But this is not the main issue, nor is it the least of many rectifications the article almost demands. In the new-age freedom of communication there are virtually no secrets anymore and this adds to my discomfort because the article only touches on some of the issues clouding Haworthia classification. The DA says I should meet this following argument in support of an H. groenewaldi’…” it is consistently smaller than H. mutica and reminds superficially actually of H. bruynsii or springbokvlakensis with its rounded leaves. Kobus Venter’s feeling was that it is closest to H. magnifica var atrofusca and I agree 100%.” These are subjective statements and I cannot agree about the size as I have seen many H. mutica populations that I do not think supported the opinion. The roundness of the leaves is also subjective. The element H. mirabilis ’atrofusca’ in any case has plants with variably rounded leaves, so which do you mean?. The name ‘mutica’ means leaves without a point and rounded and it is no co-incidence that Col. Scott initially identified H. springbokvlakensis as H. mutica until I pointed out to him that I thought H. mutica was actually the plants (note plural) in the Bredasdorp area. Scott plagiarized my words describing them alongside old wagon tracks that suggested their discovery in earlier times.
DA lists several more indeterminates…”all the unique morphological features like small size, rounded rough-textured and uniquely flecked leaves”. Of course there are not several more as I have already dismissed two and this leaves the third viz. the “uniquely flecked leaves”. Firstly not all the plants have flecked leaves and I have shown that these occur in H. mutica at Klipport too. The main issue is flowering time and I agree this is problematic. But this is actually so in respect of the whole Southern Cape Haworthia complex where it is quite evident that there are two flowering periods, summer and winter and that despite this difference there is hybridization. What happened in the past?
DA writes…”MBB 7801 flowers…with the typical thin tender peduncles and delicate flowers identical to that of H. maraisii and some H. magnifica and H. mirabilis. Normal H. mutica flowers in spring (Sept-Oct) and has more robust peduncles and flowers.” This word “typical” again, does not convey the truth. There are no floral characters by which one can separate the three “species” the sentence suggests. I have also seen small H. mutica (Crodini) with very small weak inflorescences and comment on this as a general variable. See fig. 149, a recent picture of peduncles of H. mutica from Grootvlakte, Riviersonderend. There is a population of H. mirabilis (I presume Breuer and Marx would use the name H. magnifica var. magnifica) from south Riversdale MBB6651. The plants are very small and cryptic and when I collected seed it was from small slender inflorescences with 3-4 seed capsules. In cultivation I raised a large number of plants that were all different and some were huge. The inflorescences were robust as well with 15 -20 capsules. So this reference to floral characters is simply a loose statement and I do not believe that a proper overview of the flowers in H. retusa and H. mirabilis will reveal any dramatic differences. I have taken good note of flowers and in particular the strange fish-tail bud-tip that characterizes the larger set of populations in the southern Cape.
DA states that I… ”made several blatant errors by ignoring the importance of flower features in his studies”. It this one supposed error or several? I have not ignored flowers at all and it is a blatant error to say so. I simply did not and still do not find them useful. It is fraudulent to suggest otherwise. That fish-tail bud is highly significant and I made a great effort to quantify the differences between H. reticulata and H. herbacea. That the attempt failed was only because the sample required to get to statistical difference became too large. Nevertheless I then tackled a good number of H. mirabilis populations that also have the same bud tip. I concluded that there was a difference between the populations south of the Langeberg and those north and then had to discard that as erroneous because the same problem of variability occurred as with the rosette of leaves. Even the general flowering times I gave for the area was quite wrong – flowering time ranged from November to late April. It is odd that the fish-tail bud tip does actually influence the tips of the upper tepals in that the margins tend to be flattened together. But there is a huge difference in that in H. herbacea and H. reticulata the upper tepals are held far out from the mid-axis (hence ‘subregularis’ or ‘cowboy-hat’) while in H. mirabilis the tepals may meet above the middle upper tepal.
More by the DA… ”So, only careful observation and comparison of diagnostic features, starting with flowers, is the solution.” This is a weak sop because it is something that I started to do and found quite unhelpful. The history is that the small aloid genera were virtually recognized by the fact that the flowers were small. The differences between the subgenera were dismissed by Dr L.A. Codd with the statement that “small differences” were being ignored. DA overlooks a single major binding factor between H. retusa and H. mirabilis viz. bud similarity. I have also discussed in detail the whole question of diagnostic features and went to the genus Oxalis to substantiate my observations that small detail is not helpful. It comes back to the issue of what is a species? I maintain that they are not these artificial elements circumscribed by morphological detail. They are systems set in a geographical framework. The species, whatever they are, have surely not arisen from independent sources. My field work is very extensive and I have shown over and over again that the populations are linked so that when one finds a single population like at Buffeljags, one asks how it fits. It is not any misconstrual by G.G. Smith that a sample from a population south of Heidelberg was likened to H. mutica. In his time it was not known where H. mutica even occurred and hence he made a mistake in describing H. otzenii. DA is defending a methodology that failed miserably and is not going to provide any solutions in the future.
Well, well, well one might say. The DA maintains that Bayer has lost his way, to support an argument that he had stopped “doing taxonomy”. DA is not strong on logic but it is intended that way and neither are the DA’s observations necessarily correct. DA writes this … “Bruce’s dilemma is that he spent a lifetime and hundreds of thousands of Rands doing his research and while he did indeed do an invaluable contribution, he is now faced with the fact that he can’t do that full and neat integration of it all because 1) there is more variety in the wild than what his dispensation recognizes and he feels overwhelmed by the idea of having to try and do a real truth-reflecting revision at this ripe age and 2) he made several blatant errors by ignoring the importance of flower features in his studies.” Point two I have already dismissed, but DA cannot be serious when he suggests that there is some kind of dilemma involving money and time spent. This is an issue that I will have to address when I meet the Angel of Death.
I do recognize that there is great ‘variety in the wild” quite contrary to DA’s claim that here is more variety than what my dispensation recognizes. The main gist of my writing has been this extraordinary variation. I am not overwhelmed by it at all and I maintain that my dispensation is the only way to rationally express it. To say it confuses me is not an overstatement because it is clarification that I have been seeking. Now Steven Hammer puts it very well… ”it’s almost as if you are being blamed for nature’s complexities” and I said the same thing in different words long ago. The Latin binomial system fails and I have argued and shown evidence for the need of a new and different model What is more I have proposed such a model too. Anyone who contemplates at any age, the “truth reflecting revision” the DA demands will be faced with a daunting task of citing the synonymy of all the species described or recognized by Breuer and Hayashi (let alone cite all the collecting records). That alone could occupy a lifetime. Not only that, but to also cover the usages and references by the many authors who have written about the plants would be a humungous task. I suspect that DA is implying that I have become senile with a “ripe old-age” comment. My response is that at least my senility was preceded by some degree of clarity in contrast to what I am forced to conclude about contemporary Haworthia taxonomists! I see no reason whatsoever to depart from a logical argument about species definition and the significance of geographical distribution from my species list of February 2009. Effectively this is a revision and it just does not have literature and specimen citations that meet the needs of “a full and neat integration”. Let me state categorically that there is not the slightest hope of such a pie-in-sky solution.
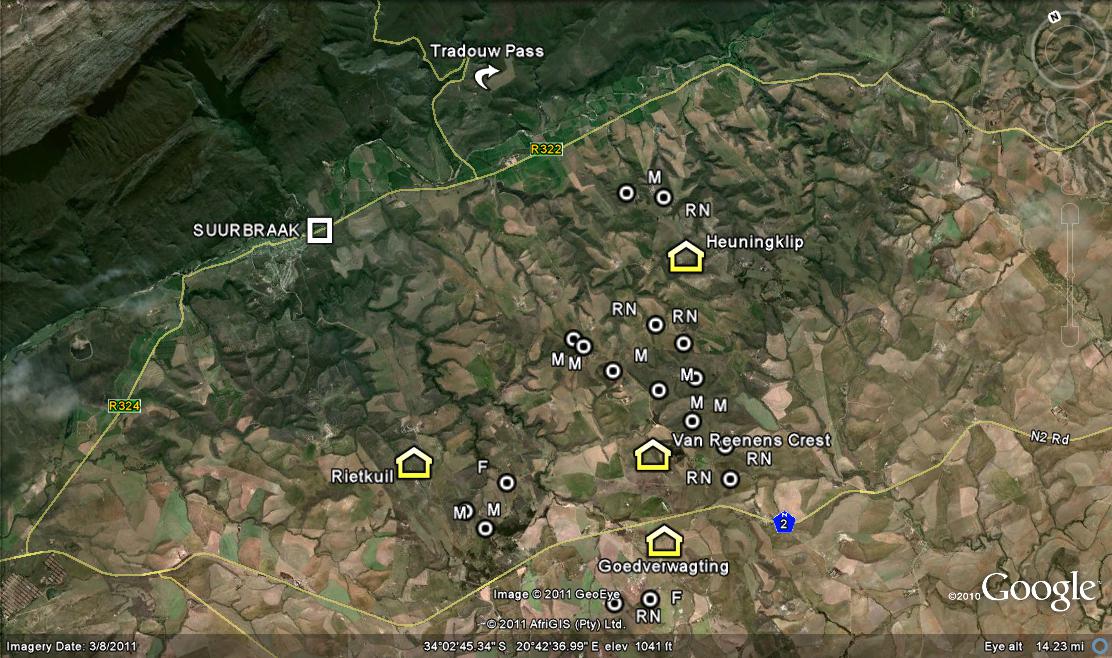
So we ask Lawrence Loucka’s question… ”How best to separate facts and knowledge from personalities and ego?” First present the facts. I want to add to the information that I presented in my article on “Haworthia retusa ‘nigra’ – A grand finale”. There I discuss and illustrate plants from the wider general area of Buffeljags and especially from the Tradouw area to the northeast. Covered are four populations from the farm Heuningklip, and three from the farm Rietkuil. Here I want to discuss and illustrate eight more populations from the farm Van Reenens Crest that lies between. The gist of my argument is that classification of Haworthia is far beyond this nonsensical and simplistic generation of new names in exactly the same way that a miasma of names was generated in the years before 1948.
It is not just chance that the area I have recently been exploring has been so neglected for so long. It is a high-lying and fairly featureless area and very grassy in comparison to the Karoid Broken Veld, Karoid Valley Bushveld remnants, and the rocky hills, streams and river beds of the areas north east and west. The vegetation is a grassy Renosterveld and the species diversity seems extraordinarily low. There is not much there to attract a succulent plant enthusiast and explorer compared to far more enticing and rewarding countryside elsewhere. My exploration there is a direct result of my having exhausted most other options available to me. An added consideration is that of prediction when long experience has proven that connections between populations exist and need to be found.
So it is not surprising that we found 8 new populations, 6 in the context of H. mirabilis as I circumscribe it, and 2 for H. retusa ‘nigra’ in the same vein of personal circumscription. These populations were all new to me and very exciting. Nevertheless I would not dream of seeing them as new species anymore than I did for H. mutica ‘Buffeljags’. The Van Reenen’s Crest populations fit a pattern and need to be considered in the context of the bracketing populations at Heuningklip and Rietkuil discussed in the preceding chapter. The collections and illustrations are mapped in an accompanying map and are identified as follows:-
The populations.
Set 1 – H. mirabilis.
The plants in the following six populations are collectively different from those at either Rietkuil or Heuningklip. Using the erroneous or absent logic that characterizes the description of “H. groenewaldii”, there is no doubt that at least one new Latin binomial should be generated. Far from being confused by the huge variation I have seen in Haworthia and guided by a systems view of species, I see these as being representative of one species. It would be useful if proponents of some other dispensation take these pictures individually and arrange them according to the characters they so diligently use to denote their many different species. The question of flower character I laugh off because I have tried to use it and know it will fail. I will come to flowering time again. Plant size is barely significant because in such clay soils the plants do not grow to the size that they will in cultivation. Generally the plants in these populations fall in the category 35mm to 800mm diam. But it is difficult to gauge age and the direct effects of niche favourability that will favour growth and size.
1. MBB7914 H. mirabilis, Van Reenens Crest See figs 1-19.
This was a small population as can be judged from fig.1 where the pressure burst and exposed white clay area barely exceeds 100sq m. The plants were only on the perimeter of the area. This is what makes it quite difficult to assess what the population structure may have been in the era before farming and cultivation. Very seldom indeed have I found plants anywhere in pristine arable or even semi-arable landscapes. Fig. 2 is a view looking across to Heuningklip and the locality of MBB7896 H. retusa ‘nigra’.



















2. MBB7915 H. mirabilis, Van Reenens Crest See figs 20-27.
Fig. 20 is a view looking south to a very large area that we did not explore because we chose instead a similar valley immediately beyond the one pictured. The MBB7915 locality was rather odd. In fact the whole exercise was in the respect that we kept being reminded how similar the situation was to Kiewietsvlakte where H. mirabilis and H. retusa play out a similar show. In this case there was no actual pressure burst. In this case there was less white clay and there were the characteristic platform-like ridges as if made by grazing animals. There were few plants and while rather scattered, were still confined to what I can only describe as low-biomass bearing area i.e. shallow gravelly soil that does not support dense vegetative cover.








3. MBB7916 H. mirabilis, Van Reenens Crest See figs 28-48.
Fig. 28 is a view of the white clay in an area very like that pictured in Fig. 20. Very heavily utilized by grazing animals, it is surprising that there were so many plants. The variability is quite evident from the pictures and some plants stand out as very different.






















4. MBB7917 H. mirabilis, Van Reenens Crest See figs 49-70.
This was a dramatic population because the plants were not quite where we expected them to be. Fig. 49 shows a more typical habitat in the distance. Here the plants were rather in a very brown gravelly situation and absent a short distance away where there was more white clay. As for all these populations there was a wide range of surface textures and also leaf shapes and endings. Often the leaves had a very distinct mucro (end-awn or point). Equally often the leaves were without any point and rounded and this was not true for all the leaves on any single plant. It is very problematic to make statements about the plants because when we were looking at Rietkuil plants we were reminded of a host of more easterly populations and plants. In this case even variants of H. emelyae came to mind.






















5. MBB7918 H. mirabilis, Van Reenens Crest See figs 71-87.
This population was unusual in that the plants were generally small and proliferous. Sometimes there were as many as 20 rosettes in a clump.

















6. MBB7919 H. mirabilis, Van Reenens Crest See figs 88-116.
One needs to refer to the map to see how the populations now relate to one another. These were again very variable as I have come to expect in the whole H. retusa/H. mirabilis collective – to coin yet another adjective to do what I do not think the formal system allows. Note carefully fig.100 in which a young bud is clearly visible.




























Set 2 – H. retusa ‘nigra’
We found just these two populations.
7. MBB7920 H. retusa ‘nigra’, Van Reenens Crest See figs 117-137.
These plants were in full flower and setting seed as one expects for the summer-flowering H. retusa elements. In fig. 128 a plant is shown with well-developed end-awn and fig. 134 a plant with rounded leaf tips. Fig. 137 is simply a view of the flower that does not even depict the more normal fish-tail bud tip – see next!





















8. MBB7921 H. retusa ‘nigra’, Van Reenens Crest See figs 138-148.
Fig. 138 is a plant that could have come from an H. retusa ‘retusa’ population from around Riversdale, while Fig. 146 could have come from H. retusa ‘turgida’ from somewhere around Heidelberg. Fig. 147 clearly shows what I mean by the fish-tail buds. Fig. 148 shows that there has been quite extensive exploitation of the white clay (kaolin). Fortunately (?) unlike the bentonite that is mined on large scale, the kaolin pockets are often small and restricted and apparently not commercially viable.











Conclusion
The plants in the above populations cannot be categorized without reference to the totality of populations and I will try and summarize my point of view. These populations occupy a position very central to the distribution of both H. retusa and H. mirabilis. I need to quote DA again who wrote … ”His (Bayer’s) emphasis is upon phylogeny and understanding the evolutionary processes at work in Haworthia and he loves getting involved with very complicated arguments and theories which (frustratingly or delightedly) to him nobody quite follows or is really interested to follow.”
DA is most fortunate to be able to dismiss the basis of classification and what all biologists involved in classification try to do. It is quite wounding to say that I enjoy this theorizing. I would be far happier if there was no contradiction and I could wander out in to the veld and be assured that there were smart neat boxes and simple ways of filling them from a manageable list of names and unequivocal identifying characters. My observation is that all this so-called “taxonomy” practiced by too many writers is so flawed because the very basis of biological variation is wholly ignored and forgotten. It does not take rocket science to imagine that science was supposed to set us free from superstition and blind belief and allow us to individually question the nature of creation.
Here we have a real problem that is overridden by a basic problem in formal classification. If I could start now afresh with a clean slate and knowing what I now do, I would still be handicapped by the restrictions of the nomenclatural code. This is that I do not believe that the name H. mirabilis as typified by a plant that comes from an outlying population truly represents the species. Perhaps this is a point not understood by the collector community, and in fact even by botanists in other disciplines. The “type” is only there to serve as a reference point for the NAME and may not be remotely typical of the species that this name is applied to. What is unfortunately true as with the crop of taxonomists who have muddied the classification of Haworthia in more ways than one, is that more often than not the application of names is not to species at all but to an odd assortment of plants that meet no criterion for a basic unit of life forms. In the case of the name and type of H. retusa there has been curiously no problem and the vast variation there is forgotten simply because of familiarity. The interesting thing is that the name H. mirabilis was not even in use before I resurrected it in about 1976, so anything that remotely departs from an unfamiliar norm must be “new”? For H. retusa ‘nigra’, I must have got the name ‘nigra’ from the old literature and reference to the Kransriviermond collection that Smith informally listed as H. mutica. The point is that in my acknowledged ignorance I attached the name to H. mutica because of the absence of an end-awn and a relatively rounded leaf tip to the plants seen. (Looking back to all my pictures I see that in some plants there is definitely an end-awn.) The dark-colour has resulted in a name that is not appropriate for the populations that can be considered to constitute the taxon. Neither is the population at a geographic end-point and as a probable hybrid entity representative. This statement also only scratches at the surface of a real problem of origins and changing faces of the plants with time.
Reflecting about relationship as a taxonomist is required to do, and with due respect to DA and the weird views of my preoccupation with relationships the invitation to him required; it seems to me that the unseen and unknown element, now only evident (H. retusa ‘nigra’) as a widespread and common element in this central area, is important. It may be the precursor to the real way in which H. retusa and H. mirabilis have developed in close synchrony and association in both easterly and westerly directions.
The really strange occurrences are evident from the map herewith (fig. 150). The two species do not grow in direct close association in the area under consideration. There are places where they do e.g. Skeiding about 20km eastwards; KomseRante, Riversdale; Soetmelksrivier, E Riversdale. Why is this? There is no observable difference in habitat or associated vegetation, and still there is pattern in the distribution. It appears to me that they are separated by flowering time (that does not preclude hybridization as field hybrids show) and in character by colour and a tendency to greater surface roughness of various kind and degree in H. mirabilis. The mystery deepens when H. floribunda appears in two places but again as sole occupant of habitat seemingly favourable to either H. retusa or H. mirabilis. Pattern is broken here because the evidence seems to be absorption into H. mirabilis in other places where the two species do closely meet. The conclusion has to be that I have come no nearer to a neat solution. The H. retusa ‘nigra’ populations cited above will attract no takers, but the H.mirabilis populations will. I create no new name because my personal view of Haworthia does not call for one.

Acknowledgement
I particularly want to thank Gerhard Marx for his considered points of view he has crafted as my Devils Advocate. He is an extraordinary grower and observer, and an artist of the highest order. He had driven me to more carefully and thoroughly explore and document what I see and think. I am extremely grateful to him. Lawrence Loucka has been a great help as an advisor, and so has Steven who brings something special to the whole field in the way he “communicates” with plants and about them. Kobus Venter is ever supportive. Access to Van Reenens Crest was through kindness of Trevenan Barry unsoured by the pressures of harvest time. ♦
Pingback: Haworthia mutica (groenewaldii) and its twisted leaves.Haworthia Updates
Great and instructive Chapter.
Thanks for all this Information.