
This article was first published in Aloe 36:34, (1999) and is now rewritten in part to accommodate some new collections and a conclusion modified accordingly.
The problem
von Poelnitz described Haworthia incurvula (Feddes Repertorium 31:85, 1932), from specimens ostensibly from Grahamstown, sent to him by Mrs. E. Ferguson. The important elements of the description are “leaves about 40, barely 20mm long, up to 12mm wide, broad ovate-oblong, without teeth, hairs or spines, on the face almost flat and inconspicuously turgid towards the apex, on the back convex and towards the apex with a rounded keel, very rarely inconspicuously double keeled, pale green and traversed by greenish longitudinal lines, which are somewhat anastomosing, some of which reach the pellucid tip, which is by no means abruptly pellucid”. von Poellnitz (Fed.Repert. 41:203, 1937) acknowledges that this is the plant illustrated in Flowering Plants of Southern Africa 9:t356 (1929), which is there identified there as H. cymbiformis var. planifolia (Haw.) Baker. Again (Fed. Repert.44:233, 1938) he acknowledges the illustration given by J.R. Brown (Desert Plant Life 8:45, 1936). Here he gives “… the exact locality is Pluto’s Vale in Albany district, between rocks and bushes.” citing R.A. Dyer as the source (Botanical Survey of South Africa, Memoir 17:98, 1937). von Poellnitz adds here that.. “in vigorous cultivated plants the leaves are less incurved or almost not at all”. He was uncertain what its affinities were and placed it in the section Muticae. However, it should be noted that von Poellnitz placed the synonymous H. planifolia (Obtusatae) and H. cymbiformis (Planifoliae) in different sections too. This was despite his confession and obvious in his writing, that he did not know the difference between them.
Smith (SA Journal of Botany 16:1, 1950) transferred the species H. incurvula to the section Obtusatae on the contention that it was continuous with H. cymbiformis which he said was also in the close Pluto’s Vale area. I wrote in my Haworthia Handbook (p124, 1976) “Smith’s field book untypically bears no reference to other collections of this kind in that specific area, and the writer’s limited field experience does not bear this out either. There is however, an apparent if obscure relationship with H. translucens subsp. tenera which bears further investigation. Forms very similar if not identical to H. cymbiformis var. incurvula also occur at Andrieskraal north of Humansdorp, which is a long way to the west.” In my New Haworthia Handbook I could add nothing new, and neither could I add anything substantial in an article I wrote for Cactus & Co 2:11 (1998). Col. C.L.Scott maintained H. incurvula as discrete, writing that “it has been gathered at a number of localities along Pluto’s Vale near Grahamstown”. His riposte is “H. incurvula is a distinct species, and is readily separated from H. cymbiformis“. It must be noted that he does not separate H. cymbiformis and H. planifolia either.
Pluto’s Vale is also the origin of H. tenera von Poellnitz (Fed.Repert. 31:86, 1932). The collector was Miss Blackbeard. The site is incorrectly given as “Plinters Vale” and apparently, by courtesy Albany Museum staff, the origin is “Cloete’s Vale”. This is a smaller more proliferating species with more, slender and less densely spined leaves. However, I made a collection from Pluto’s Vale myself circa 1972. In fact two collections (KG47/72a &b), within 75 meters of one another. In one the plants were spinescent, and in the other glabrous. The plants were too small, with the leaves too spinescent and too slender to be incurvula, but the potential for connection was there and that is the predictive stance that was taken. In my New Haworthia Handbook (1983), the point was also made of look-alikes in the Humansdorp area for both tenera and incurvula.
At this point I must state that in my revision (1999) I depart from the name translucens and replace it with gracilis. My thinking subsequent to this change and prior to current writing is, that the name gracilis (from Hellspoort, NW Grahamstown) is in fact a variant of tenera (or vice versa). Scott’s solution is not greatly different except that he applies the name tenera more widely to include elements in the Humansdorp area; and he limits the name gracilis (his use of the name translucens has no bearing on this discussion) to its point of origin.
The hypothesis I now pursue is my own, which is that the element incurvula does not constitute a species and that it can be explained in the context of two other systems (species) as I first suggested in 1976. These are H. gracilis (H. translucens sensu Bayer 1976, 1983: and including the variety H. gracilis var. tenera) and H. cymbiformis (hence H. cymbiformis var. incurvula). Lest anyone again question what I mean by species as do Crouch et al (Aloe 36:8, 1999) in their allusion to “very seldom have some of these authors suggested what might constitute a species”; let me spell out what is implicit in all my classification. Thus – “A species is a system of living organisms which are genetically and morphologically continuous in space and time”. Unfortunately Crouch et al pursue their allusion with considerable vigour and extraordinary circumlocution and tautology. They write of the “amateurish attempts of interested lay persons and taxonomists alike to force an absolute and linear and dichotomous order on the genus”. Unfortunately it is exactly this flawed reasoning and logic that force me to wonder if it is even worth pursuing knowledge anymore (C&SJ.US 70:183, 1998). Our classification system is essentially a dichotomous one and the very concept “genus”/”species” is itself a dichotomy. I will try to explain new findings without attempting to invent a new classification system, although we undoubtedly need one.
New records
While re-writing my Handbook and formally revising Haworthia, I was required to cite all herbarium specimens. There are 15 records for incurvula alone and they are all from Pluto’s Vale. None give any detail outside of that geographical information statement. Thus there is no evidence to support Scott’s contention of.. “a number of localities along Pluto’s Vale”. J.D. Venter has a collection (JDV93/60-81 [81 a-d]) by Scott’s colleague, the indefatigable Dr. Reddi, which states “middle of Pluto’s Vale”. This collection was said to have been made north of the road.
There is of course a whole range of specimens of H. cymbiformis from various places in the Eastern Cape. These should really be discussed, analyzed morphometrically and illustrated. These would show why the elements planifolia and cymbiformis can be treated as synonymous. This synonymy is largely due to the origination of the names, and suggests simply that two such morphological elements could have been from a single local system. The herbarium evidence would indicate why the concept “incurvula” is reasonably included there among the many varieties of H. cymbiformis described at one time or another. These are ramosa, reddii, transiens, obtusa (recognised by myself), and further names such as lepida, angustata, brevifolia, alta, agavoides, incrassata, calochlora, longifolia, compacta, exulata, setulifera, obesa and olivacea (which are not).
There appear to be several populations from the greater Grahamstown area in which the plants are smallish and with bluntish incurved leaves (E. Fort Brown, Blaaukranz, Vaalvlei, Fernkloof, Kaffirdrift). There are also populations in which the plants are larger with acuminate, flatter leaves in a more spreading rosette (Committees, Howiesonspoort, Kapp-Fish confluence) and populations with obtuse turgid leaves (Alicedale, Swartwaterpoort).
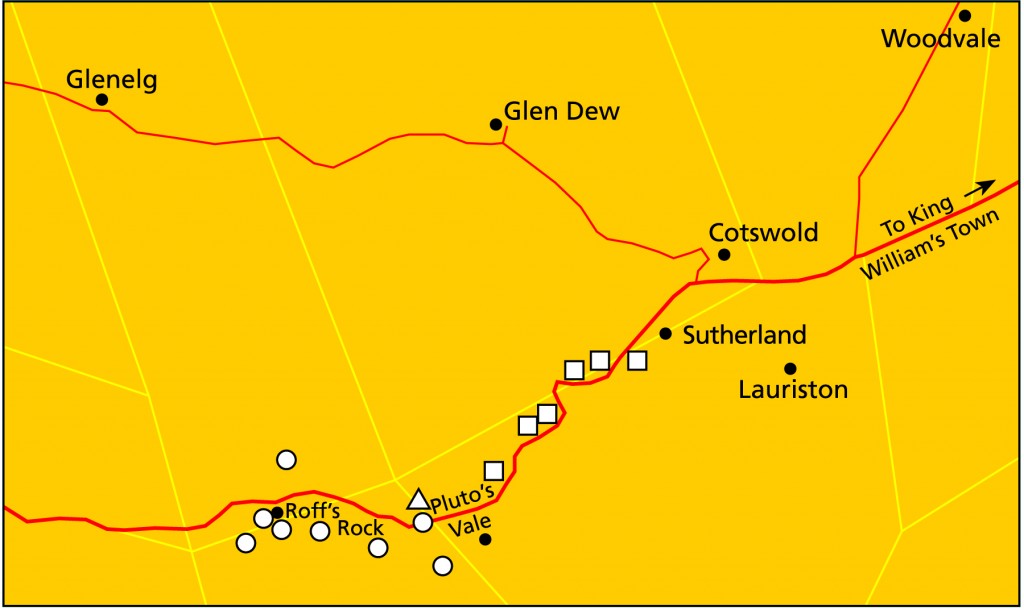
Here we are expressly concerned with Pluto’s Vale and the plants which occur there. Curiously incurvula is at its closest and most accessible to collection, near the top of the steep descent into the valley, Pluto’s Vale. There is a gate there into the field and a dwelling at a point here called Roff’s Rock. In the tradition of plant collecting in South Africa, as exemplified by the discovery of the first Lithop species by Burchell in the 1800s, collections are invariably alongside a road, or within very short walking distance of some obvious stopping point. The Roff’s Rock site is just such a place and incurvula is less than 50 meters from the roadside. There is a also a vast population of Haworthia attenuata (var. britteniana), and also the plant which the collectors probably actually and repeatedly came to view ie. Encephalartos trispinosus. Incurvula also occurs mid-way down the steepest descent of Pluto’s Vale, north of the road and immediately adjacent to it. Tenera is also known from the farm Glenelg to the north, to the east near Committees and also to the south-east at Hunt’s Drift. H. cymbiformis is a separate element in those same areas and I did cite specimens of incurvula from south of Hunt’s Drift too (Smith5741, Smith6508). I have not seen these populations to confirm that they are, as living specimens, in fact comparable.
In November 1998, I specifically visited Pluto’s Vale again to explore the area a little more thoroughly and to re-locate my glabrous H. tenera. My wife, Daphne visited the area again during July 1999 because of the failure of the previous visit to locate that form. We visited the site again in September 1999 together with Steven Hammer and with Tony Dold of the Schonland Herbarium. A sketch map is provided of the area to demonstrate what we found where. The first visit was collectively quite an extensive search, but by no means adequate to establish the whole truth.
Tenera was found to be common in the Vale itself i.e. east of Roff’s Rock (MBB6832 [82 a, b], MBB6833 [83], MBB6834 [84], MBB6835 [85], MBB6840 [86]). Venter collected at the exit from the valley (JDV90/36 [87 a-c]) and also at Hunt’s Drift (JDV91/103 [88 a, b]).
Incurvula was abundant from the vicinity of Roff’s Rock and extends along that ridge south of the road to where the ridge ends at the base of the Vale (MBB6836 [89 a-f], MBB6837 [90 a-f], MBB6838 [91.1 a, b], MBB6839 [92 a-c]). It was also collected by Marx and Venter (JDV93/73 [93 a, b]) on one occasion slightly towards the south-west of Roff’s Rock, and we also collected it at the second visit (MBB6893 [94 a, b]) still further west. I could not locate the original site of my glabrous tenera (KG47/72a) north of the road and in the Vale. The normal spinose form was on all the south facing rocks. The vegetation seems to have been modified by extensive grazing of the two farms and management systems now evident (the one area is now a game farm and goats have been introduced on the other). However, on the most recent visit we did again find the glabrous form (MBB6891 [95 a, b]) in profusion almost directly above the incurvula population (MBB6892 [96 a, b]) north of the road. In the first version of this report (Bayer, 1999) I incorrectly cited JDV93/60 [81 a-d] as Dr. Reddi’s collection of incurvula from south-west of Roff’ Rock. Apparently it was actually collected north of the road in the middle of the Vale. I can now associate it with my collection [96 a, b] of Pluto’s Vale which is however, of smaller plants than Reddi’s.
The Marx/Venter [93 a, b] collection is of plants with longer (to 50mm) sub-erect leaves which are very lanceolate as opposed to the ovate-oblong shape and incurvature of the leaves in the original description. Traditionally and conceptually incurvula has leaves which are obtuse at the tip. The description does not circumscribe this, and this could have become a very important point. The Marx/Venter plants have such dramatically lanceolate leaves that they throw a slightly new aspect onto the scene. My first collection [89 a-f] between the given Marx/Venter site and Roff’s Rock, also is of plants with such lanceolate and pointed leaves. My wife turned up a similar population north-west of Roff’s Rock (MBB6884 [97 a-d]) in which the plants were smaller and proliferous on a rockier, steeper, south face. There is a stronger tendency to spination. The colour is towards the grey rather than the green. The leaves are more gracile and acuminate. There is no turgidity or swelling of the leaf as one finds in pilifera or in cymbiformis var. obtusa, and given as “inconspicuously so” in Von Poellnitz’ description. But this collection is not the same as the two glabrous ones which so obviously relate to tenera. It does perhaps relate better to Hellspoort gracilis, although it is fairly similar too to [89 a-f], [93 a, b] and [94 a, b].
There is a collection by Marx and Venter (JDV96/4 [98] also by myself MBB6603 [99 a-d]), from north-east of Grahamstown at Glen Craig. H. cooperi var. pilifera also occurs there but about 2km to the east. This collection is of largish plants, greener rather than the bluish/purplish coloration of the cooperi complex. The leaves of the plants are ovate-lanceolate and not turgid above the mid-point. There are many collections of look-alikes from as far afield as Adelaide, the Zuurberg and the Humansdorp area. There is also a very interesting record by R.A. Bayliss from north-east of Grahamstown that needs following-up. These collections impinge on the interplay of H. cooperi var. cooperi, H. bolusii var. blackbeardiana, H. decipiens var. pringlei and H. aristata, which is the subject of another manuscript.
Again: the four collections made from a point south-west of Roff’s Rock to approximately 2 kilometers along the same southern face of the ridge down into the Vale. The first collection was the one described above [89 a-f], of plants with the more lanceolate and acuminate leaves of the Marx/Venter collection, the second [90 a-f] was about 200 meters eastwards which included plants with acute and awned, and plants with obtuse, leaf-tips. One specimen of the third collection [91.2] is identical to a clone in a collection from Alicedale (MBB6847 [100]), a significant collection of H. cymbiformis var. obtusa which I will write about on another occasion. There are close similarities to a collection of mine of this same element from NW Grahamstown (the Bayliss site?, Thornkloof, MBB6895 [101]). That collection was made in pursuit of a R.D. Bayliss collection KG382/76. The progression at Pluto’s Vale to the foot of the slope, was to obtuse leaves and the cymbiformis-like appearance which is also evident in collections of mine from greater Humansdorp (eg. [15 a-e]). The plants were often single and occasionally with more than one rosette in a cluster.
An interesting point is that of the relationship between marginal spination and pointedness of the leaf-tip. One might expect, as indeed I did, that the two situations would be correlated. To a degree they are. Incurvula has leaves which are seldom spined but often blunt-tipped, in tenera the leaves are generally spined and as in glabrous tenera, always with a pronounced point to the leaves. In the case of H. cooperi var. pilifera from Slagtersnek [104 a-e], there are clones in which the leaves are very truncated as in Col. Scott’s joeyii (the var. dielsiana). There is no tip to the leaf at all, and yet the margins do have spines.
The bluntness of the leaf-tips cannot thus be taken to be diagnostic. Here we have pungent and blunt leaf-tips in a continuum of specimens from a relatively extensive area. The “Flowering Plants” illustration appears to have no tip to the leaf per se, whereas J.R. Brown writes “Occasionally a leaf may be found with a very short pellucid point.” His photograph shows leaves with a distinct short abrupt blunt mucro (point), but nothing like the tapered sharp extended point evident in many of the plants I saw. This variation alternative is also evident for H. cymbiformis, for H. cooperi and for H. gracilis within the interactive systems that they are. The phenomenon is also evident in the south-western Cape complex where for example we have H. magnifica which has leaves which are acute with long end-awn, and the var. atrofusca in which the leaves are blunt (obtuse). This is evident within populations.
Tenera was collected in the Vale from a point north of the road [86] from where my search for incurvula terminated. We found it on all of the five rock faces we searched from this point to the eastern end of the Vale. There our earlier expedition ended without real conclusion. It is proliferous and rosettes are usually in clusters. The two elements tenera and incurvula thus grow in close proximity. While in Aloe (1999) I wrote that there was no direct evidence of obvious introgression, or morphological or genetic continuity. My recent collections of small incurvula [96 a, b] and glabrous tenera [95 a, b] require a more committed opinion. The connection is thus through the glabrous populations of tenera directly to incurvula, and also via the three most western and northern pointed-leaved plants from glabrous tenera to incurvula.
In the case of my latest exploration I returned to the Andrieskraal area (SW Patensie) in the eastern Baviaanskloof. I wanted to get some idea of what G.G.Smith had recorded in the Mistkraalkloof area. I already had collected H. gracilis var. picturata at Andrieskraal itself [14 a-c,15 a-e]. This time I collected a little further east [16.1 a-d, 16.2] and these are smaller, non-proliferous plants. One clone was only 16mm in diameter with 35 leaves, while another was 25mm diameter with less than 25 leaves. These plants bear a surprisingly similarity to incurvula, and apart from the blunt leaf-tips, also to the glabrous tenera. Curiously I collected from another population [17 a-c] a little further to the east. This was a glabrous H. gracilis var. isabellae which is very similar too, to glabrous tenera.
In this particular paper I do not consider H. cooperi, which can elsewhere found to be continuous with both gracilis and cymbiformis. H. cooperi var. pilifera is present to the south-west of Pluto’s Vale at Glen Craig, to the north at Fort Brown, and at Committees to the north-east. It does not appear to be involved as such at Pluto’s Vale.
The conclusion
There is an ambience to the literature on Haworthia which makes it very difficult to write meaningfully and truthfully about the genus. This ambience reaches a climacteric in the recent article by Crouch et al cited earlier. The review of Haworthia Revisited by the botanist Paul Forster is also so much in the stereotyped and conventional mould of taxonomic botany, that there is little useful to direct scepticism and inquiry in a direction which can possibly advance understanding of classification. This is not true only for Haworthia. Speciation in Haworthia (if not also in many other plant genera, as I have said elsewhere) is chaotic. This can be construed in various ways and I hope it will one day seen to be in a proper sense of order rather than disorder. A search, very thorough by the standards one is generally led to observe, has resolved the relationship between incurvula and tenera to a greater degree than the first publication of this report. Yet, it cannot be stated categorically that incurvula belongs in H. cymbiformis nor that tenera belongs in H. gracilis. A pattern is only evident here for which I have now evidence and this repeats itself in several other areas. Exploration and understanding of these patterns is necessary but is not going to make description and classification in the traditional way, any easier.
One of my observations about natural hybridisation in Haworthia is that hybrids are always very conspicuous, and usually more so because of their close proximity to both parents. On several occasions I have collected hybrids in physical contact with one or other of the putative parents and seldom in the absence of both. The word “putative”, means by inference, but my conclusion is that the use of the word in relation to natural Haworthia hybrids is that it is too passive. The inference I make is that genetic interchange between populations is very limited and populations are able to develop and maintain their own integrity over very short distances. Thus we have glabrous tenera and small incurvula existing as discrete entities in close proximity (75 meters), with a potential third element representing possibly gracilis as the four western collections of plants with the pointed leaves. A parallel situation with three main role players is also evident in the Baviaanskloof and it has always been observance of the degree of co-occurrence of these role players which has led me to my classification. I do not think that these role players really represent “systems” and thus species in themselves, but they can be explained in terms of my classification and revision. I would suggest that my classification is valid as an hypothesis for further inquiry along these same lines.
There are still many outstanding doubts and a decision to “force” change in the dichotomous order will remain very inviting to the uninitiated. There is a very wide interplay between many “species” in what is a very widely spread and intricate system or systems. We are dealing with visual imagery and trying to create order in this. The fact is that we are really dealing with unquantified miniscule differences of size, colour and leaf shape, with a tendency to seek and attach importance to even more miniscule differences of spination, translucence etc. in individual plants. There is also a tendency to attach significance to sophisticated technology applied to limited samples. The problem with this is that it is difficult then for adequate sampling, repetitive testing and casual review.
As mentioned, a similar manuscript which deals with the interplay of the species gracilis, cymbiformis and cooperi, and another which deals with the interplay of aristata, bolusii var. blackbeardiana and cooperi. These show that taxonomic solutions which were thought to be easy to present for some small areas and for small samples are not. Such solutions also break down in other areas. It is very difficult indeed to draw all these “species” into a single discussion, but I feel confident that my classification and revision can be used for this purpose. It has been successfully used like this since 1976. The “group” species solutions as given by T. Salter for Oxalis and by O. Hillyard for Streptocarpus are not very satisfying. It is not a question of forcing the imposition of a linear system. This IS the system in general use. We need another one. I am impressed with a bifurcation diagram given in Glieck’s book “Chaos, the Making of a New Science”. This particular diagram suggests that, made three-dimensional, with asynchronous and asymmetric bifurcation, it could model speciation in plants. The model would be presumably be described by a non-linear differential equation. We need a new language and/or a new paradigm, if we are to name and communicate about plants in a harmonious way, understandable and acceptable way. I hope we will be able to understand that when it comes, because we are not doing very well in the current language we use. ♦



















































