
This problem of continuity is one I seem to have difficulty in conveying to my readers and listeners. The difference between one species and another is a discontinuity and, if we believe in evolution, it is the resultant of a break-up of continuity in its ancestral parent species. The “model” we have in our minds, is of progressive change from one recognisable entity to another by evolution. Geographic distribution and re-distribution are key elements in this process. But we do not seem accept this in the way we try to classify plants or interpret classifications. Apart from recognising that change could be gradual and therefore manifest continuity, the change may be from a complex variable system which contains different levels of continuity within itself, and not from a simply understood uniform ‘ancestor’.
The result is that in a genus like Haworthia, which is by no means exceptional, the differences between species i.e. the discontinuities between “species”, may be very difficult to either recognise or rationalise. It in fact becomes a statistical operation in which all the characters should be involved i.e. multiple variate analysis. If all the characters could be measured and quantified it is statistically possible to subject all the data so obtained by one of several statistical methods to measure “distance” and “significant difference” between groups of plants which we want to ascertain are species, varieties or even just hybrids. The process of “cladistics” is the use of a system to generate a branching “tree” of relationships base on characters which are also evaluated and loaded for chronological priority (primitive versus advanced). In using such a mathematical package, it is pretended that the classification becomes “objective” and hence replicable to satisfy the scientific requirement. In my estimation, the cladistic process assumes that a two-dimensional “tree” adequately represents the spatial and temporal changes of evolutionary processes, and it does not work.
Somebody might one day try to apply such methods to Haworthia and I say “Good luck to you”. My experience of characterisation and variation in the biological systems I have experience of, and including Haworthia, suggest to me that sensible, practical, experienced “eye-balling” will prove the better bet. Ultimately in Haworthia, I expect that technology and cladistic methods will be testable on the result of my classification. This is not a conceited and arrogant claim. It is a simple reflection on what classification actually is and what it is for. Much of botanical classification has been done by amateurs with no, or minimal, specific training and qualification for that field at all e.g. G.W. Reynolds, L.C. Leach, T.L. Salter, J. Lavranos, C.L. Scott, G.G. Smith, M.B. Bayer etc. Their classifications form the basis of many scientific observations, sometimes by scientists who have no conception of the significance, or insignificance of the names they use or what they may actually mean. The classifications may have little to recommend them except the fact that they appear to conform to the approved nomenclatural style.
Putting that all aside I want to just demonstrate by means of a series of illustrations, what occurs in Haworthia. I have described several situations before but what follows is another account of the continuity which occurs in the southern districts of Somerset East, Eastern Cape. The main area concerned is Kaboega at the north and western Zuurberg near where it fades away north of the Klein Winterhoek Mountains (see map Fig.1). One thing should be engraved on the mind. The following comparisons are made on field impressions and on multiple plant samples. This at least takes note of statistical requirements and considerations and it would be wrong to try and use the individual illustrations to arrive at some other conclusion.
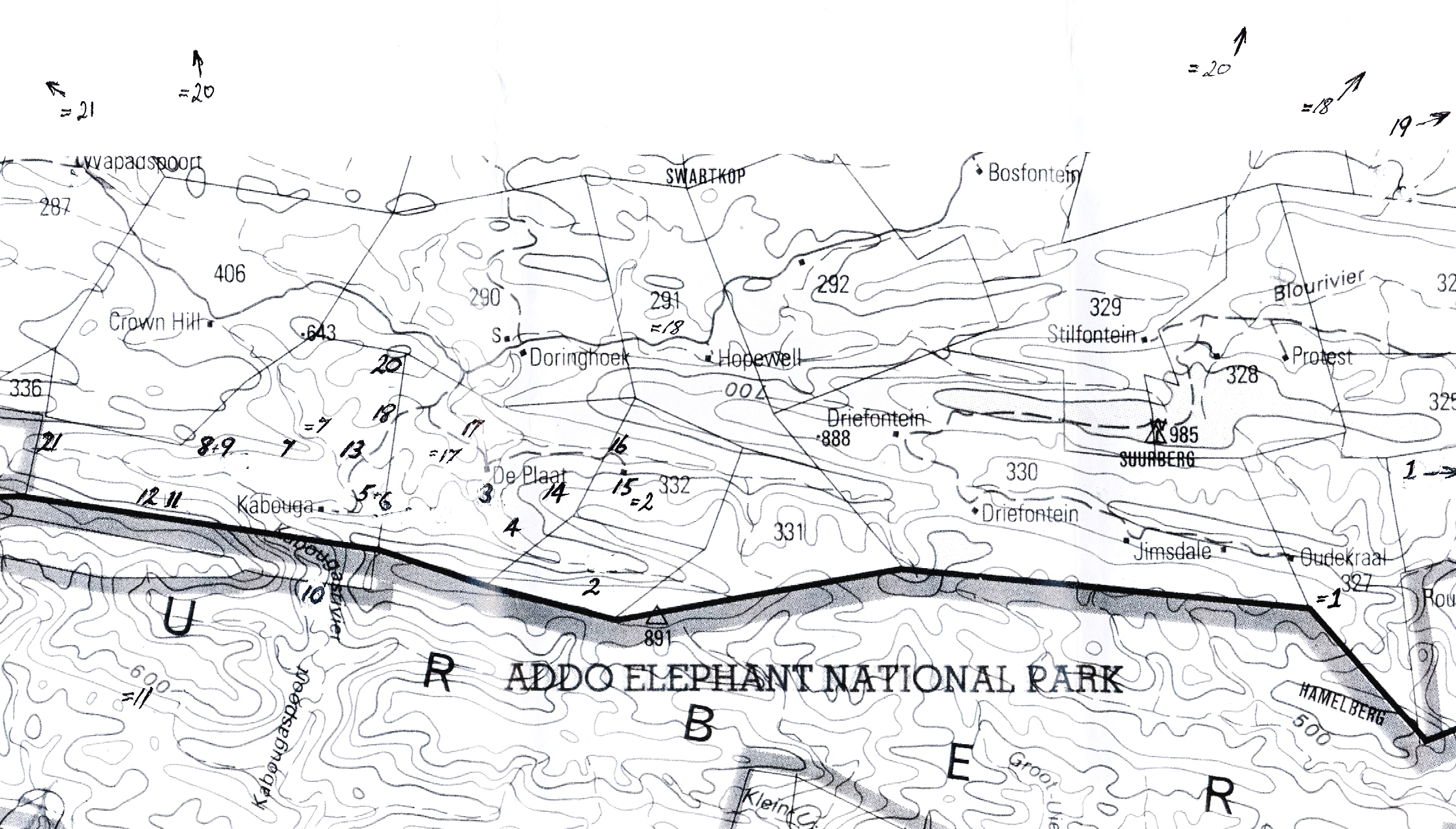
Determining a starting point is quite difficult because there simply is none unless a second problem is addressed. This is the one of typification in which names are fixed to a specimen of some kind. After this is done there is the question of how the name is applied to the body of the plants from which it comes. In this article, the question of the application of the names cooperi, pilifera, vittata and blackbeardiana is involved and so I reproduce the illustrations accompanying the origins of these names (Figs.2, 3, 4 & 5). It is my experience that in the field, it is possible to find resemblances to these illustrations in a large number of geographically disparate populations, and hence quite a problem to apply them in a consistent and sensible way. Only the name blackbeardiana can unequivocally be said to have been derived from any particular field population and this is at Bowe’s Farm near Imvani, south of Queenstown, and the single illustration by no means represents all the variants of the many populations which can be said to belong together here. Some of those variants invoke the use of the other names. How all these names are used is something of a personal choice and mine is laid out in the book Haworthia Revisited which I wrote in 1999.
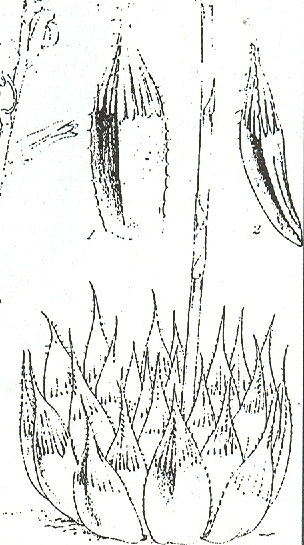
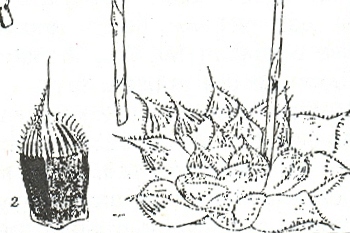
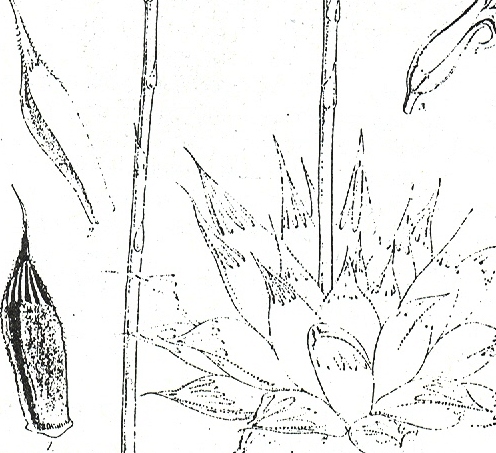
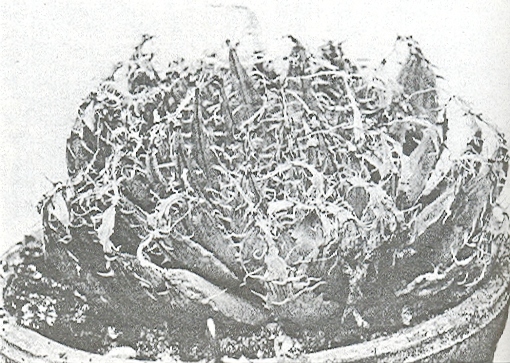
In this chapter I will take a collection (originally by G. Marx) from east of Alicedale and call it H. cooperi var. cooperi ([1]…denoting position on map Fig.6. MBB6845). In doing so I will have to point out again that there are further intrinsic difficulties in the requirements of nomenclature, which is that there is such an entity as a “typical” variety. My belief is that any plant which is not directly assignable to a variety of a species should simply be referred to as that species. Thus the Alicedale collection is H. cooperi var. cooperi and geographically in the distribution area of H. cooperi var. pilifera. The plant illustrated is of a single clone, but the plants in this particular population relate better to populations in the geographical area of the typical var. cooperi. The plants occurs here together with H. angustifolia and H. cymbiformis (in the same indeterminate varietal format as the H. cooperi, but for geographical reasons mainly, this population is referred to as H. cymbiformis var. obtusa. A similar form of H. cooperi depicted recurs on the Zuurberg and I saw it at Oudekraal when I explored there for H. angustifolia var. baylisii ca 1982. I have plants in cultivation from there which were collected by Ernst van Jaarsveld, and I made a new collection MBB6922 slightly west of there in 1999. I saw it again at Vyeboomfontein ([2]Fig.7 MBB6949) much further west and again just north of that at Klipfontein [2] MBB7049. I am unsure if the identification should be var. cooperi or var. pilifera but choose the latter.


Slightly further west at DePlaat, the problem starts with plants which are greener (particularly on the south slopes) and with longer more attenuate leaves. There is first the collection from north slopes ([3]Fig.8 JDV91/14) that I attribute to H. cooperi var. gracilis, and then mine from the south slopes ([4]Fig.9 MBB6935). Near the Kaboega farmhouse at Koksedam there is also a greenish element and two plants are illustrated ([5,6]Figs.10a & b MBB6915) to show that it is partly cooperoid and partly cymbiformoid. I propose calling these, with MBB6935, H. cooperi var. viridis because it is complicated (it could invoke the name gracilis of von Poellnitz, but I described a smaller green variant from near west as var. viridis, and this name seems more appropriate). I have, in Haworthia Update Vol.1, described that there is this greenish archetypal form which is quite widely spread and often in an indeterminate zone, between forms of H. cooperi and H. cymbiformis. On Kaboega these plants take three elemental directions:




1. Directly west to Spekboomberg ([7]Fig.11 MBB6914) and then to Wilgerfontein ([8,9]Fig.12a & b MBB6925) at. At Fig.11 the plants are consistent with Fig.9. At Fig.12a & b the plants resemble H. cymbiformis. Some clones are more similar to that species than others while odd clones are entirely indistinguishable from it. I find it difficult to determine what species it is, never mind add a varietal name, because of its obvious intergradation with H. cooperi variants and its geographical disassociation with H. cymbiformis. My suggestion is that this is where the case calls for the use of the species name, without the addition of the typical variety epithet. But even that is unsatisfactory here because I think the plants, despite their outside similarity with cymbiformis, are actually in the same genetic bag as cooperi var. viridis. It is worth stating that there is a transition from sandstones at DePlaat and eastwards, to Dwyka tillite westwards to Wilgerfontein.



2. Slightly to the south-west, entering the Kaboega gorge is JDV92/33 ([10]Fig.13). This is indeterminate and I would label it “graciloid”, although there is evidence of other variants in the gorge which do not agree with the concept of var. gracilis. That name itself presents problems as described in Update Vol.1. Fig.14 is in sandstones, and on the steep north facing slopes (also sandstone) of Wilgerfontein a little to the west and south of MBB6925, is MBB6904 ([11]Fig.14). I would suggest that this resembles H. cooperi var. gordoniana (subsequent to drafting this manuscript I did a limited exploration south of the Zuurberg, and found a fairly similar populations occurs at [=11] in the Addo National Park. But it is an astonishing ecotypic variant too, as 25 meters away is MBB6905 ([12]Fig.15) which is “isabelloid“. I have coined the name “puberula” for it. Bear in mind that these pictures represent only samples of many plants in small discrete habitats.



3. In a north-westerly direction from DePlaat is MBB6943([13]Fig.16), which is slightly spinose and indeterminate like the MBB6915 (Fig.10) collection. But it gets very curious. If we ascend the DePlaat hillside towards Klipfontein and [2]MBB7049, we find first MBB6910 ([14]Fig.18) almost indistinguishable from MBB6905 and close to MBB6904. A little further, and less than 250meters from MBB7049 is MBB7017 ([15]Fig.19), a small graciloid=puberula population in which the plants are very small and the leaves have surface spination. If we cross the valley and go now north of Klipfontein to the upper slopes, we find MBB6940 ([16]Fig.19). It is indistinguishable from MBB6904! If instead we stick to the low lying areas and move north of Koksedam, and northwest of DePlaat, there is first MBB6942 ([17]Fig.20) and then MBB6917 ([18]Fig.21). This latter is a relief because I have several collections from widely dispersed points [=18] to suggest these are my version of H. aristata. Not forgetting that the initialisation of that concept at Soutkloof near Addo, also involves H. cooperi. If we look at one collection from Commadagga ([19]Fig.22 MBB6897) we see a plant which is a variant of H. aristata (it could be cooperi var. pilifera!) that has look-alikes at Kirkwood in the arena of H. cooperi var. pilifera. But at Kaboega, the plot thickens further. Going still northwest from DePlaat and now north of MBB6943, is MBB6916 ([20]Fig.23). There is a problem again, and one has to go quite a way (about 20km) to find MBB6899 ([=20]Fig.24). The latter is quite a large plant in the mould of H. aristata. To find a better home for it among several other collections, including some dubious ones to the east and north at Ripon, and including Col Scott’s H. pringlei, I added it there while reducing Scott’s species to H. bolusii var. pringlei. This problem is elucidated elsewhere but I have not decided where best to place MBB6916 I think for practical and geographic reasons they could be placed under H. aristata, but they could equally be put under H. bolusii var. pringlei. I described H. decipiens var. virella to lessen tension on my use of the name pringlei and its relation to H. bolusii var. blackbeardiana and to H. cooperi var. dielsiana (includes Scott’s H. joeyi). While it does so in some areas of the classification, some tension does remain.










This link to the above variants is enhanced by MBB7012 ([21]Fig.25) from Buffelsnek, overlooking Lake Mentz and west of Wilgerfontein. This collection is also labelled H. cooperi ‘graciloid’ as I have done for MBB6904, but it could just as well be H. aristata or even bolusii var. pringlei (because of the nature of continuity offered by other populations to the northwest [=21]. To show just how complex the situation is, I illustrate JDV92/44 (Fig.26). This was initially a P.V. Bruyns collection from Inverbolo, far distant and northeast of Cathcart. I specifically went there to confirm (MBB6942) that this is H. bolusii var. blackbeardiana. I have said elsewhere that there is a problem of this question of continuity and that H. decipiens can be indistinguishable from H. bolusii var. blackbeardiana. To add insult to injury I illustrate MBB6730 (Fig.27). No! It is not H. aristata, it is H. mucronata var. habdomadis from Seweweekspoort in the Little Karoo, and from an entirely different set of populations. It is in fact extremely disconcerting to have to admit that the green plants of Figs. 9 or 10 can similarly shown to be indistinguishable from H. mucronata var. morrisiae, also from the Little Karoo.



In writing this, I hope that readers will now get to understand that in apparently attacking the work of aspirant Haworthia taxonomists, I am not trying to defend myself or my system. I simply recognise that these persons do not have the competences or the insight to follow and understand the difficult choices that have to be made any better than myself. The main point is, that I know that if we explore still further west of Kaboega (and there are collections which prove this), and then into the Klein Winterhoek mountains to the south-west, still further problems are going to unravel (us) as we roller-coaster between H. decipiens and H. cooperi all the way to Uniondale and beyond. These same problems bedevil most of the species systems in the southern and western Cape. Unless one understands and respects the intricate geographic relationships, the use of the names in the way I recognise them will perhaps not make sense and I truly see no other way in which a nomenclatural system and classification, which will work in the field, can be derived for Haworthia. ♦