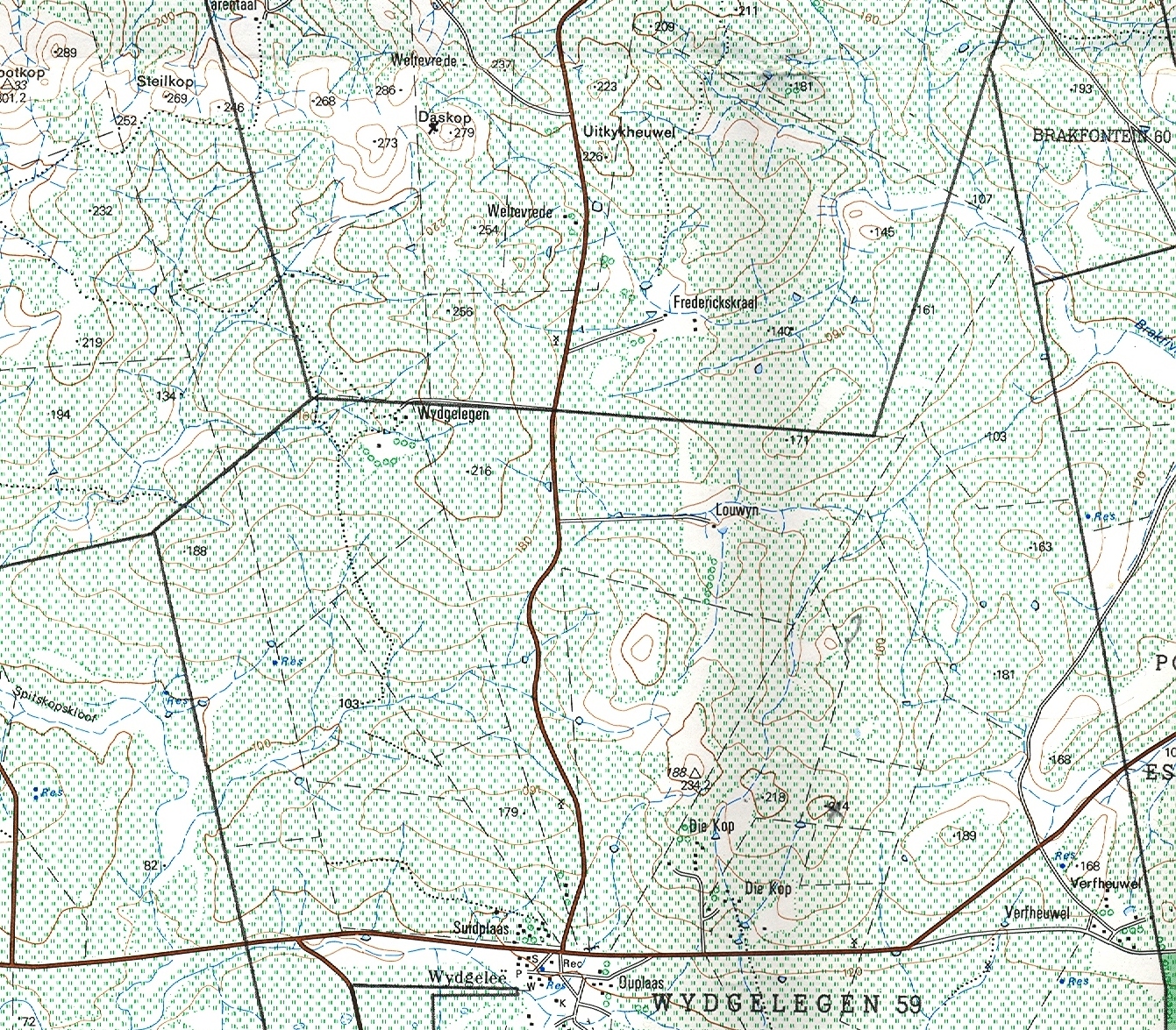
Haworthia mirabilis was described by Haworth in 1804 from plants said to have been brought to England by the collector Francis Masson. It could thus have originated from practically anywhere in the southern and south-western Cape. Haworth referred to this species as the ‘rough cushion cushion’, and it was characterised by having retuse-deltoid leaves with ciliate-spinose margins and keels, the leaves being smooth on the face and the back surfaces almost tubercled and indistinctly reticulate. There is no extant type specimen and only the brief original description, and illustrations in Curtis’ Botanical Magazine (t 1354, 1811) and in Salm-Dyck’s Aloe Monograph (s9, t 1, 1836-49) serve to identify and typify the species.
Previously published Cact S.Jl 52.1 1980
M. B. Bayer, Karoo Botanic Garden, National Botanic Gardens of South Africa
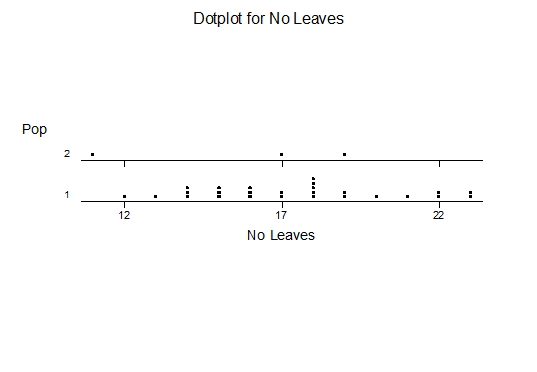
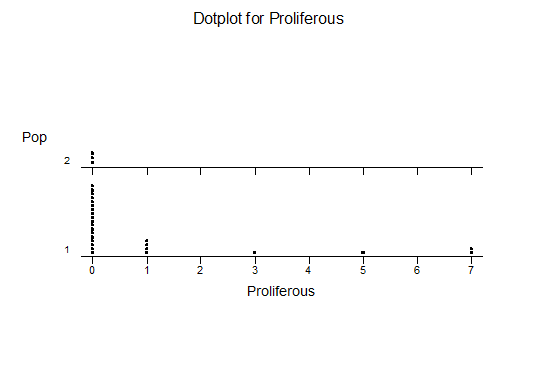
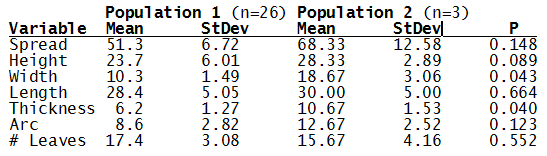
Haworthia nitidula was described by von Poellnitz in Desert Plant Life (11:192, 1939) from plants collected by Major H. Venter. Venter was very generous with his localities and like most of his others, this no. 15 was also simply cited… “in the environs of Worcester, Swellendam, Caledon and Bredasdorp”. This is an ideal example for demonstrating the problems of identification of haworthias and especially in the section Retusae Haworth. J.R.Brown illustrated H. nitidula in the Cactus and Succulent Journal (18:89, 1946) but apart from this and the original illustration accompanying the description, the plant has not figured in the succulent literature. In G.G.Smith’s records, the species and nine varieties are distinguished. The interesting thing is that these varieties are drawn from three geographically separated species. The object of this article is to present Smith’s photographs and notes of his “varieties” and discuss their actual position as they relate to distribution.
21. Haworthia mirabilis Haw., Syn.Pl Succ. :95(1812). Bayer :136(75). Bayer :47(1982). Bayer, Excelsa 7:37(1977). pp. Scott :116(1985). Aloe mirabilis Haw., Trans.Linn.Soc. 7:9(1804). Ker-G., Curtis’ Bot.Mag. t1354(1811). Type: Cape, Masson. Not preserved. Neotype (designated here): Icon t1354, Curtis’ Bot.Mag.: H. mundula Smith, JS.Afr.Bot. 12:8(1946). H. mirabilis ssp. mundula (Smith) Bayer :139(1976). Bayer :47(1982). Type: CAPE‑3419 (Caledon): Mierkraal, Bredasdorp (‑DB), M. Otzen 10 in Smith 5479 (NBG).
mirabilis: wonderful.
Rosette stemless, proliferous, to 7cm φ. Leaves 10-15, retused, 3-4cm X 1,5cm, markedly retused, acute above, face translucent and lined, dark green, with marginal spines turning reddish in the sun. Inflorescence slender. Flowers narrow, elongate, biarcuate bud, upper lobes pinched at tips.
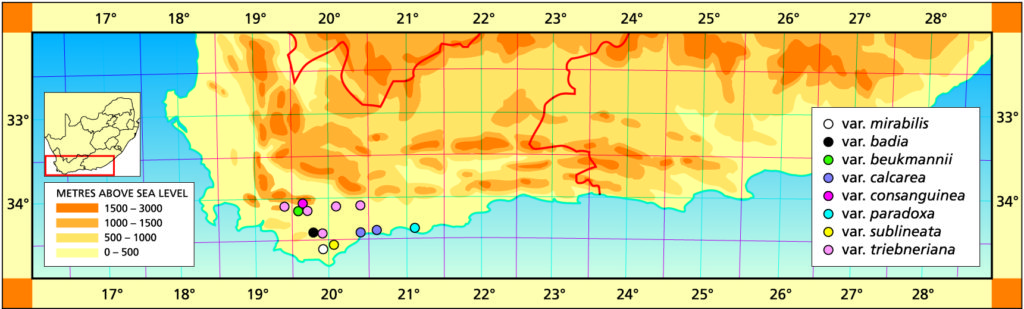
1982 – A full account of H. mirabilis is given in Excelsa (Bayer, 1977). It is intimately associated with H. magnifica but the nature of the relationship is not fully understood. The species grow near together at Stormsvlei and an intermediate population occurs a few miles to the south. The species also approach one another near the Potberg at the mouth of the Breede River, at Bredasdorp and, unconfirmed, in the area east of Drew. The initial distinction was that H. mirabilis (as H. triebnriana) was smooth on the leaf faces while H. magnifica was not. H. mirabilis is generally a larger plant, and with more translucence in the leaves than H. magnifica. The flower is a little larger and generally brown‑veined. Several names such as H. willowmorensis, H. triebneriana, and H. rossouwii are placed under H. mirabilis on rather speculative grounds. The synonymy thus indicates a very variable species and this is in fact so. There are several places, notably at Bredasdorp and Napier, where ecological differences have resulted in forms very different from one another. At Napier the differences are of such an order that the subspecies badia is recognised. At Greyton, a form high in the mountains, with the typical flower of H. mirabilis, occurs. For the present it is regarded as a variant of H. mirabilis. There are also variants in the limestone formations of the coastal belt. The subspecies mundula is a very proliferous variant south‑west of Bredasdorp. It is possible that H. magnifica var. paradoxa belongs with H. mirabilis (see H. magnifica) but the coastal area has not been fully explored so that a good decision is not yet possible.
1999 – The typification of this species is obvious although Haworth in 1812 does not cite the Botanical Magazine illustration. He does cite the preceding t.1353, Aloe recurva under Haworthia recurva, so this is quite an extraordinary oversight. There is an illustration in the Kew library apparently labelled H. mirabilis which Scott regards as illustrating this species. The illustration is almost identical to t.1353 and is of that species viz. H. recurva Haw. rather than of H. mirabilis. The illustration is of a trifarious plant and, despite the annotation ‘mirabilis, ‘there can be no question of any confusion between this and the description of H. mirabilis which reads ‘quinquefarius’. The other points are that 1. Ker-Gawler obtained his plants from Haworth and acknowledges this source; 2. Haworth refers to its singularity and ‘productions of art’; 3. Haworth places it between H. margaritifera and H. translucens(?).
This has been an old problem in Haworthia – the proliferation of names without the first establishment of the application of the old. The recognition of the correct application of the species name requires some additional modification to the structure of the species. While citing the wrongly labelled Kew illustration, Scott also omits mention of the Ker-Gawler illustration. In addition to this he appears to have identified several collection of H. maraisii as H. mirabilis which is also apparent from his distribution map. These two species do not co-occur and yet they seem to be discrete. Two populations are known which appear to be interactions of the two elements.
a.var. mirabilis. It is clear that H. mirabilis had to have some application and this has previously been found in terms of a host of varieties. In retrospect it is apparent that the only variety that satisfies the requirements of the species as originally described is H. mundula of Smith, transferred only to H. mirabilis as a subspecies in 1976. It is a problem of the nomenclatural system that the species may be described from a non-representative element, as has happened here where only one deviant population is concerned. The specimen nominated as the epitype by Breuer and Metzing is again unfortunate because this is the variant beukmannii.
Distribution: CAPE‑3419 (Caledon): Mierkraal, Bredasdorp (‑DB), M. Otzen 10 in Smith 5479 (NBG), Smith 3951, 5364, 5612 (NBG).
Inadequately located: ex hort, Malherbe in NBG303/60.
Haworthia mirabilis var mirabilis JDV96/33 southwest of Bredasdorp. G.G.Smith had made no attempt to relate his collections to Haworth’s original species. In fact very few authors have made the attempt to relate names to a concept of specie as natural phenomena.
Haworthia mirabilis var mirabilis JDV96/33 southwest of Bredasdorp. Very proliferous and mat-forming at the single locality known.
Haworthia mirabilis var mirabilis JDV92/84 southwest of Bredasdorp. The plants colour deeply in the dry summer, usually reddish.
b.var. badia (V.Poelln.) Bayer comb.nov. H. badia V.Poelln., Kakteenkunde 7:76(1938). Haworthia mirabilis subsp. badia (V.Poelln.) Bayer :101(1976). Bayer, Excelsa 7:42(1977). Bayer :47(1982). Type: Cape, Napier, G.J. Payne in Triebn. 1058. Not preserved. Lectotype (B&M): icon, Kakteenk. en Kakteenfr. :76(1938).
badia: reddish brown.
This is a robust, slowly proliferous variety growing in sandstone derived depauperate clay, in among pebbles and low-growing fynbos. The leaves are quite attenuate and develop a very deep shiny brown colour in the sun. Unfortunately most of its rather unique habitat has been destroyed by quarrying activity and invasion by alien vegetation.
Distribution: 3419 (Napier): Napier (‑BD), I. Williams 616 (NBG), Payne in PRE 34870, Smith 3239, 3269, 5207, 5480 (NBG), Bayer in KG628/69, Rossouw in NBG2095/37 (BOL); 3km Napier to Caledon (-BD), Scott 2207 (PRE).
Haworthia mirabilis var. badia JDV88/66 west of Napier. Usually a chestnut brown in the field.
Haworthia mirabilis var. badia JDV88/66 west of Napier. Often protected under low shrublets. Can be surprisingly difficult to see.
Haworthia mirabilis var. badia JDV88/66 west of Napier. The shiny end area of the leaf is conspicuous and characteristic.
c.var. beukmannii (V.Poelln.) Bayer comb.nov. H. emelyae var. beukmannii V.Poelln., Feddes Repert.Spec.Nov. 49:29(1940). Type: Cape, Caledon, C. Beukman. Not preserved. Lectotype (B&M): icon (B). Epitype (designated here): CAPE‑3419 (Caledon): Skuitsberg (‑BA), Smith 3969 (NBG).
beukmannii: for C. Beukman.
This is a very robust form with strongly retused leaves and spined margins.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. The plants are large despite growing in restricted shale cracks.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. The leaf tips are generally retused.
Haworthia mirabilis var. beukmannii JDV96/45 Skuitsberg. Caledon. Occasionally proliferous.
Haworthia mirabilis var. beukmannii JDV88/70 Skuitsberg. It has been suggested that, historically, this was the possible origin of the species.
d.var. calcarea var.nov. Type: CAPE-3420 (Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG, Holo.).
calcarea: pertaining to lime.
Differs from the species in having short erect leaves, with a short retused end-area. It is proliferous and the rosettes, at least in cultivation, tend to be raised as opposed to remaining flattish to the ground. (A var. mirabilis foliis brevioribus erectis viridibus sordidus differt).
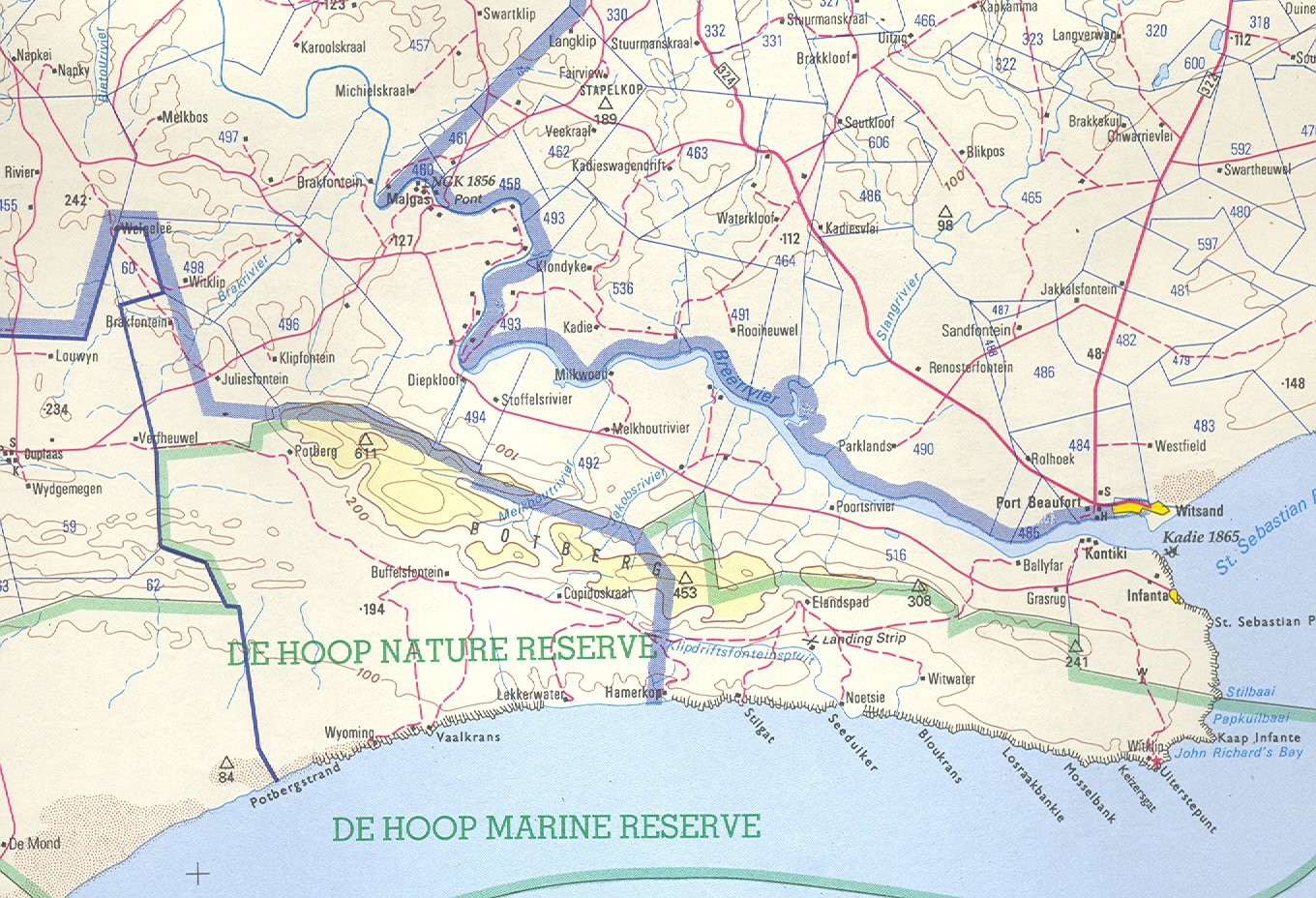
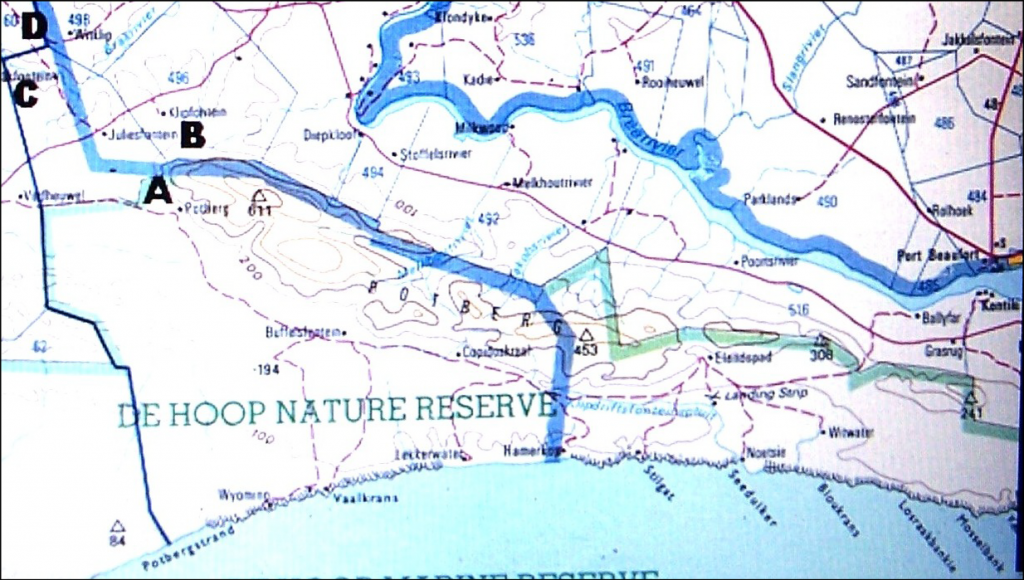
First collected by C. Burgers in the De Hoop Nature reserve where it grows on low-lying limestone rocks. There are two other collections from the Potberg area which may have some connection with this variety, or with the species. One is by Prof. Compton in which the plants appear to have been in sand and very obscure. At first sight they appear to be of H. mutica, but they have very pointed leaves. The other collection is also by C. Burgers and appear to be of plants with more slender and longer leaves from the lower slopes of the Potberg. There is also a collection from the Potberg once made by A. Mitchell of a very small plant, like the variety consanguinea described below, but which is H. variegata var. modesta. H. heidelbergensis is also known from this area.
Distribution: 3420 (Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG); NNE. Buffelsfontein (-BC), Burgers 2018 (NBG).
Haworthia mirabilis var. calcarea JDV84/65 De Hoop. An unusual occurance for the species in limestone.
A small proliferous, relatively soft-leaved form comparable with the small proliferous mountain form of H. turgida. (A var. mirabilis rosulis parvioribus prolificantibus facile et foliis virellis differt).
It is difficult to deal with these montane forms which seem to occur in the sandstone ranges from the Potberg, the Riviersonderend, the Langeberg, the Swartberg and the Cedarberg mountains. This particular element seems to be associated with the lower-lying varieties in the shales, in the same way that H. turgida transposes to retusa-like forms. Although looking very similar to the sandstone forms of H. turgida, it has the very narrow elongate buds of H. mirabilis, and also the leaves have the brownish-red coloration associated with H. mirabilis. The P.V. Bruyns collection cited below is significantly different from the two others as the leaves are quite slender.
Inadequately located: Near McGregor, Esterhuysen 5218, 5219 (BOL).
Haworthia mirabilis var. consanguinea JDV88/63 Dwarswaterkloof southwest of McGregor. This form is also in sandstone and the leaves are slender and slightly cylindrical.
Haworthia mirabilis var. consanguinea JDV84/76 Die Galg northeast of Greyton. This form is very similar to H. turgida except that it has the brownish-red tints of H. mirabilis, and also a similar flower.
f.var. paradoxa (V.Poelln.) Bayer comb.nov. H. paradoxa V.Poelln., Feddes Repert.Spec.Nov. 33:240(1933). idem. Desert.Pl.Life 99:90(1937). H. maraisii var. paradoxa (V.Poelln.) Bayer :143(1976). H. magnifica var. paradoxa (V.Poelln.) Bayer, Nat.Cact.Succ.J 32:18(1977). Bayer :45(1982). pp. H. asperula Haw. sensu Scott :119(1985). Type: Cape, Vermaaklikheid, Mrs Ferguson in Stellenbosch 6692. Not preserved. Neotype (B&M): Riversdale, Ferguson (BOL).
paradoxa: paradoxical.
Von Poellnitz did not explain his epithet and simply related it to his H. schuldtiana (H. maraisii). He was struck by the spines on the leaf face which are unusual. A greater paradox is the relation of this element to H. emelyae var. major which also has a very spined leaf surface. It does not follow that such spines are a dichotomously allocated character. Their occurrence in H. magnifica does not necessarily suggest that var. paradoxa is better placed there on these and geographical grounds, than with H. mirabilis. The var. paradoxa also seems to be associated with limestones and there is no other known link with H. mirabilis east of the Breede River. The solution which I offer is the somewhat dubious connection of the species through H. heidelbergensis, and possibly also through the two populations ascribed to var. calcarea.
Distribution: 3421 (Riversdale): Vermaaklikheid (‑AC), J. Dekenah 8 (NBG), Kramer 433 (PRE), Fourcade 277 (NBG), Smith 3272, 5388, 6109 (NBG).
Inadequately located: Ferguson 1 (BOL).
Haworthia mirabilis var. paradoxa JDV94/89 south of Vermaaaklikheid. Proliferous and growing on coastal limestones.
Haworthia mirabilis var. paradoxa JDV94/89 south of Vermaaaklikheid. Very similar both vegetatively and florally to H. emelyae var. major.
g.var. sublineata (V.Poelln.) Bayer comb.nov. H. triebneriana var. sublineata V.Poelln. in Feddes Repert.Spec.Nov. 44:135(1938). Type: Cape, Bredasdorp, G.J. Payne in Triebn. 1106. Not preserved. Neotype (designated here): CAPE-3420 (Bredasdorp): S. Bredasdorp (-CA), Smith 3966 (NBG).
sublineata: almost lined.
This is also a sandstone variant from south of Bredasdorp. The leaves are relatively long and slender. A similar, well-lined but more robust form used to be abundant immediately north of the town too along the river bank. It should be expected to occur further to the west.
Distribution: 3420 (Bredasdorp): S. Bredasdorp (-CA), Smith 3966 (NBG), Stayner in KG209/60; N. Bredasdorp (-CA), Smith 3252, 3266, 3830, 3976 (NBG).
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. A more robust form used to occur to the north of the town as well.
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. A lighter green colour and with long attenuate leaves.
Haworthia mirabilis var. sublineata JDV88/67 south of Bredasdorp. Very similar forms also occur in the sandstones north of Riviersonderend mountains.
h.var. triebneriana (V.Poelln.) Bayer comb.nov. H. triebneriana V.Poelln., Cactus J 5:33(1936). idem. Feddes Repert.Spec.Nov. 41:214(1937). idem. Cactus J 6:36(1937). idem. Desert.Pl.Life 99:101(1937). idem. Feddes Repert.Spec.Nov. 47:8(1939): H. mirabilis Haw. pp. Bayer :136(1976). pp. Bayer :47(1982). pp. Bayer, Excelsa 7:42(1977). pp. Scott :69(1985). Type: Cape, Strydomsvlei, Mrs Helm in Triebn. 841. Not preserved. Lectotype (Bayer, 1976): icon. (B): H. willowmorensis V.Poelln., Feddes Repert Spec.Nov. 41:216(1937). non Scott, Aloe 11:42(1973). Type: Cape, Willowmore, Mrs Helm in Triebn. 840. Not preserved. Lectotype (B&M): icon (B): H. triebneriana var. depauperata idid. 43:94(1938). ibid. Desert.Pl.Life 9:101(1937). Type: Cape, Stormsvlei near Robertson, Payne in Triebn. 990. Not preserved. Lectotype (Bayer, 1976): icon. (B): H. triebneriana var. multituberculata idem. Feddes Repert Spec.Nov. 44:135. idem. 47:10(1939). Type: Cape, NW. Napier, G.J. Payne in Triebn. 1111. Not preserved. Lectotype (Bayer, 1976): icon, (B): H. triebneriana var. rubrodentata Triebn. et V.Poelln. ibid. 47:10(1939). Type: Cape, between Villiersdorp and Greyton, G.J. Payne in Triebn. 1143. Not preserved. Lecotype (B&M): icon (B). Epitype (designated here): CAPE-3419(Caledon): near Genadendal (-BA), Bayer in KG692/69 (NBG): H. triebneriana var. napierensis ibid. Type: Cape, Napier, G.J. Payne in Triebn. 1145. Not preserved. Neotype (designated here): CAPE-3419(Caledon): Skietpad (-BD), Bayer 4642 (NBG): H. triebneriana var. turgida Triebn. ibid. 47:11(1939). Type: Cape, N. of Napier, G.J. Payne in Triebn. 1107. Not preserved: H. triebneriana var. subtuberculata V.Poelln. idem. 47:10(1939). Type: Cape, S. Caledon, G.J. Payne in Triebn. 1114. Not preserved: H. triebneriana var. pulchra V.Poelln. ibid. 49:29(1940). Type: Cape, N. Stormsvlei, Stellenbosch 19. Not preserved. Neotype (designated here): CAPE-3420(Bredasdorp): Stormsvlei Pass, Beukmann in Smith 5609 (NBG): H. rossouwii V.Poelln., Kakteenkunde 7:75(1938). Type: Cape, Napier, Rossouw in Triebn. 1059. Not preserved. Lecotype (B&M): icon (B). H. nitidula V.Poelln., Desert.Pl.Life 11:192(1939), Bayer, Cact.Succ.J(U.S.) 52:10(1980). Type: Cape, Worcester, Swellendam etc., Venter 15. Not preserved. Lectotype (B&M): icon (B). Epitype (designated here): CAPE-3419(Caledon): near Greyton, Bayer in KG31/70 (NBG).
triebneriana; in honour of W. Triebner.
In recognising the typical variety as the previously named ssp. mundula, it becomes necessary to find a name for the main body of the species, and this is now taken from the first name that can be fairly definitely associated with the species. This is despite the improbable locality cited for H. triebneriana and considering the very poor records associated with Triebner’s contributions. Where the typical variety of the species is restricted to the single locality southwest of Bredasdorp, this generalised variety is widespread in the area from Caledon, east to Swellendam, to Napier and back to Caledon. At Swellendam there is an additional population to the one south of Stormsvlei where this species is confounded with H. maraisii.
Distribution: 3419 (Caledon): Uitkyk (-AB), Smith 5562 (NBG), Bayer in KG682/69 (NBG); N. Uitkyk (-AB), Bayer in KG28/70 (NBG); Dagbreek (-BA), Bayer 2453 (NBG); near Greyton (-BA), Smith 3245, 3260, 3265, 3419, 3811, 3904, 3905, 3968, 5481, 5482, 5643 (NBG), Bayer in KG 30/70, in KG31/70 (NBG); Near Genadendal (-BA), Bayer in KG692/69 (NBG); Skietpad (-BD), Bayer 4642 in KG510/70 (NBG); Mierkraal, Napier (-BD), Bayer in KG681/69 (NBG). 342O(Bredasdorp): Stormsvlei (-AA), Smith 3254, 3457, 3971, 5609, Bayer in KG26/70 (NBG), De Kok 296 (NBG); Near Breede River (-AB), Tomlinson 13680 (PRE).
Inadequately located: ex Triebner, Smith 4947 (NBG); Venter in NBG6290/39; Caledon, Venter 13, 15 (BOL).
Haworthia mirabilis var. treberiana JDV96/47 southeast of Greyton. Large plants which are often subjected to fire.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad, Napier. The typical variety is the residue of plants of the species not named as varieties. May be robust or relatively small.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad. This was the locality for the ‘var. napierensis’ V.Poelin.
Haworthia mirabilis var. treberiana JDV94/90 Schietpad. Some plants are somewhat tuberculate.
Haworthia mirabilis var. treberiana JDV90/34 west of Genadendal. Occasionally the marginal spines may be conspicuously reddish.
This appeared as an article in ALOE 38:31 (2001). Unfortunately there was a problem with illustrations and captions and these are corrected here. A comment is also added as an addendum to respond to criticism by I. Breuer published in Alsterworthia 1:13(2002).
Introduction:
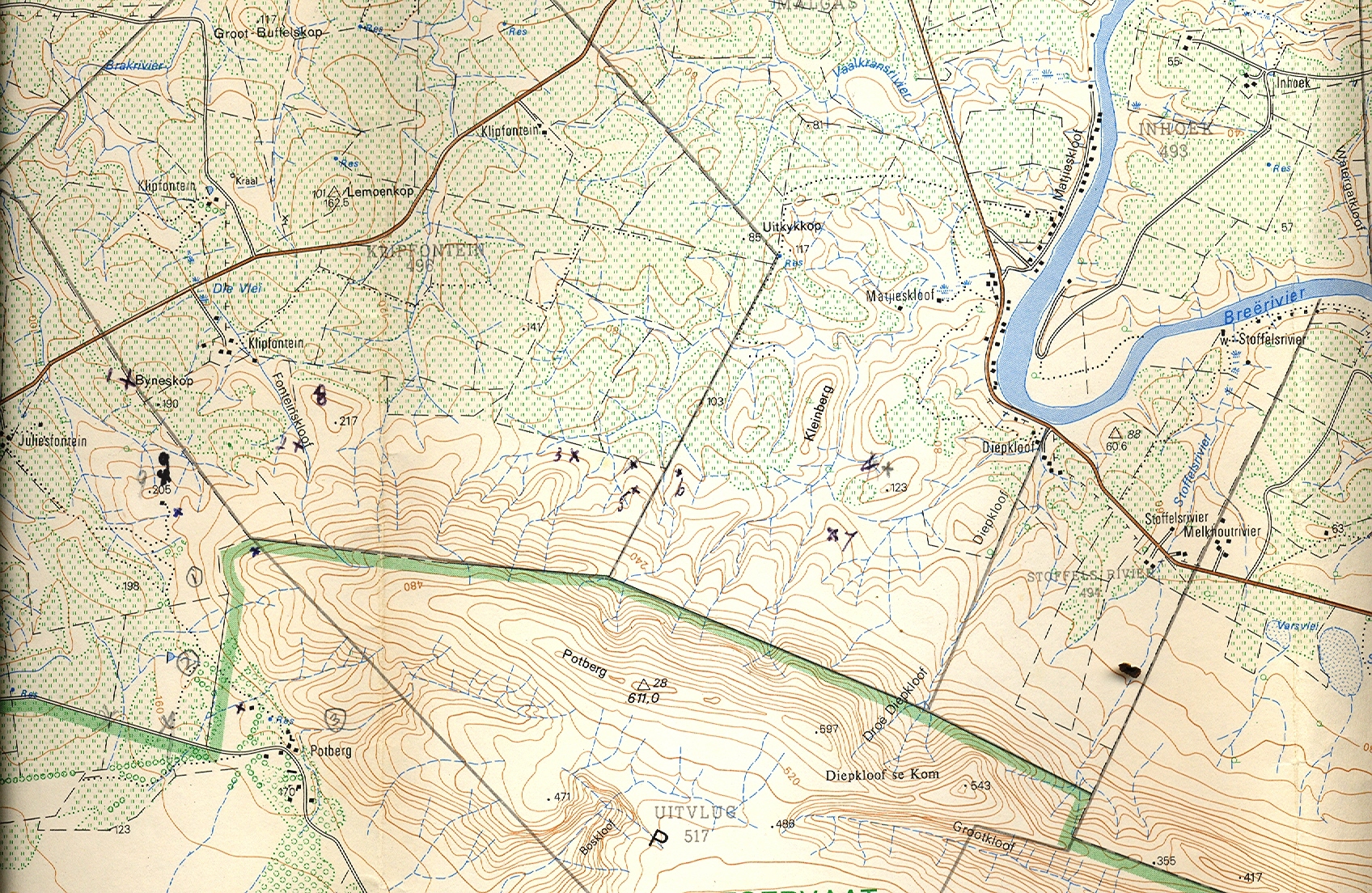
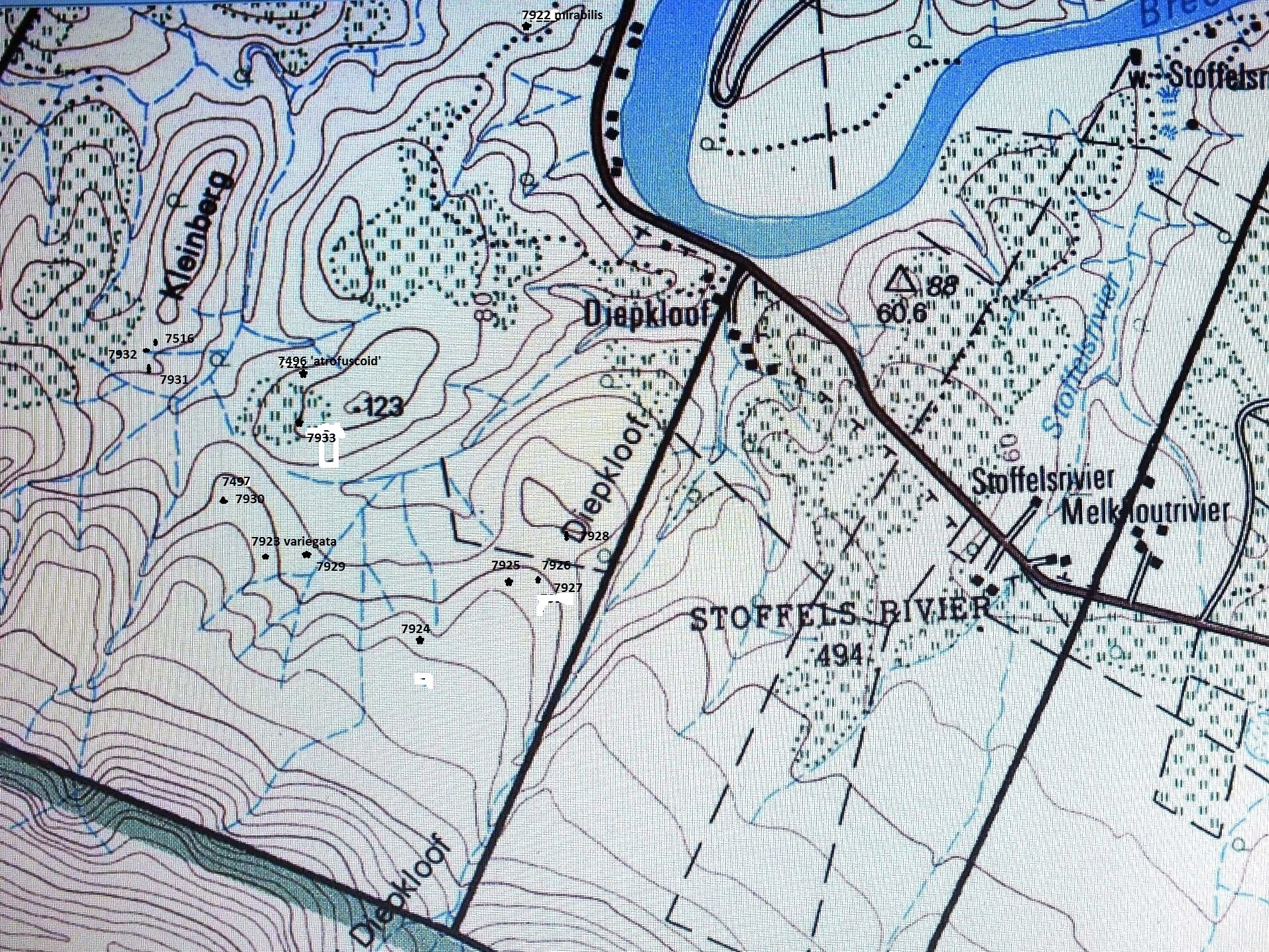
I described Haworthia serrata in 1973 (Jl S.AFr.Bot.39:249, see Figs.1) from Oudekraal, southwest of Heidelberg. I commented then on the wisdom of describing a new species when “the recognition, estimation of taxonomic rank and circumscription of elements in Haworthia…” was so problematic. The new species was said to resemble H. emelyae var. multifolia (Figs.2). In respect of its distribution, I said it was closest to H. heidelbergensis at Heidelberg (Figs.3 JDV87/1) and as at Matjestoon (Fig.4 JDV87/3), and also to H. sublimpidula at Swellendam (now known to be H. floribunda var. major (Fig.5 MBB6859, taxonomically with little connection to H. rossouwii). The implication was that it could have been taxonomically related to those elements in terms of geographic distribution. I was still puzzled by the relationships of H. serrata when I wrote (New Haworthia Handbook :55, 1982) that collections by C. Burgers from the coastal limestones might throw more light on the matter (Fig.6 MBB6985 H. mirabilis var. calcarea).
Fig. 01a. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.rossouwii 1 043Fig. 01b. JDV87-47 H. rossouwii. Oudekraal, SW Heidelberg.Fig. 01c JDV87-47 H. rossouwii, Oudekraal, Heidelberg.Fig. 01d. JDV88-53 H. rossouwii. Oudekraal, Heidelberg.Fig. 02a. JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 02b JDV87-162 H. emelyae var multifolia. Springfontein, Karoo.Fig. 03a. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03b. JDV87-1 H. heidelbergensis. E Heidelberg.Fig. 03c. MBBsn H. heidelbergensis. E Heidelberg.Fig. 04. JDV87-3 H. heidelbergensis var toonensis. Matjestoon, SW Heidelberg.Fig. 05. MBB6859 H. floribunda var major. S. Swellendam.
In his revision of Haworthia (1983), C.L. Scott recognised my species. He confused it, and illustrated it, with a photograph of H. mucronata var. mucronata sensu Bayer (1999), which he called H. mclarenii. This does exemplify the problem of look-alikes in the subgenus, and this curiously does have some connection to the actual problem.
When I wrote Haworthia Revisited (1999), I was still not sure what the real affinities of H. serrata were. I mentioned that my 1982 observations were vague because of the unusual elements which occurred in the coastal limestones. I described these as new: H. mirabilis var. calcarea from De Hoop Nature Reserve (see Fig.6), H. variegata var. petrophila (CBurgers2158) from Karsriver and H. heidelbergensis var. minor from N Bredasdorp (Figs.7 JDV86/5). In my 1999 classification, I placed H. magnifica var. paradoxa (Figs.8 JDV86/78 & 94/89) in H. mirabilis, suggesting that it may have a connection there via the var. calcarea and an element from the Potberg (CBurgers 2018) which I have not yet seen in the field myself (since I wrote the article I have in fact seen this population, MBB7356, and it is discussed in the Chapter re H. mirabilis). The connections of paradoxa must be viewed in its relationship to H. emelyae var. major, in the same way that serrata is compared with H. emelyae var. multifolia. I also suggested that H. serrata had a very close resemblance to some forms of H. mirabilis var. sublineata from south of Bredasdorp. This present paper addresses some of these observations again.
Fig. 17b. Fig. 06a. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 06b. MBB6985 H. rossouwii var calcarea. DeHoop.Fig. 07a. JDV86-5 H. heidelbergenis var minor. Rooivlei, Bredasdorp.Fig. 07b. JDV86-5 H. heidelbergensis. Rooivlei, N Bredasdorp.Fig. 07c. JDV86-5 H. heidelbergensis var minor. Rooivlei, Bredasdorp.Fig. 08a. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08b. JDV86-78 H. mirabilis var paradoxa. Vermaaklikheid.Fig. 08c. JDV98-89 H. mirabilis var paradoxa.
New Records In June 1999, I revisited H. serrata at Oudekraalkop (MBB166) again, this time together with J.D.Venter. Despite rumours of over-collection, it was still very abundant, although localised. It was also still present at Koppies (Fig.9 JDV99/6) a short distance to the west, but in very low numbers at a badly burnt site. P.V. Bruyns (priv. comm.) has reported it at a more northerly site as well. This could be where J.N.O. Uys also showed it to me at a second site on Oudekraal when he first introduced me to this species. It struck me again how similar the plants were to H. mirabilis (particularly the var. sublineata, Figs.10 MBB6639, bearing in mind the vast variation that exists there) and to H. heidelbergensis var. minor. The visit in June ’99 was a general excursion to explore the reported presence of H. maraisii at Koppies (ex E. Esterhuizen), in relation to that species and heidelbergensis as I know it in that general area. Our finds and observations led us later in the year to explore the area north of the Potberg. The extraordinary finds there led me to further explore westward to Bredasdorp itself and to see for myself the var. calcarea in the De Hoop. Two visits for that purpose were fruitless, but thanks to Dennis DeKok, I did locate it in the company of Lawrence Loucka in July 2000 (MBB6985-fig.11). This latter excursion was particularly fruitful because, by great good fortune, we discovered H. serrata in unmistakable form north of Bredasdorp (Soutkloof, Fig.12a & b MBB6983), and Nooitgedacht, MBB6984). Both sites are west of Rooivlei where heidelbergensis var. minor occurs. We also collected variegata var. petrophila (Fig. 13 MBB6986) west of the Karsriver limeworks. For that variety I have C. Burger’s record (CB2158) from east of the limeworks, and a collection I made at Renosterfontein (Fig.14 MBB6632) on the east bank of the Karsriver in 1997. This latter collection, in particular, reminded me in both form and colour of H. emelyae var. multifolia (see fig.2).
Fig. 10a. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10b. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10c. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10d. MBB6639 H. mirabilis var sublineata. S Bredasdorp.Fig. 10e.Fig. 10f.Fig. 10g.Fig. 11a. MBB6985 H. rossouwii var calcarea. DeHoopFig. 11b.Fig. 12a. MBB6983 H. rossouwii. Soutkloof. Fig. 12b.Fig. 13a. MBB6986 H. petrophila KarsrivierFig. 13b.Fig. 14. MBB6632 H. rossouwii var petrophila. Renosterfontein, Karsriver.
The conclusion from seeing H. serrata north of Bredasdorp, and both var. calcarea and var. petrophila, is that this is the prime connection. It seems now more improbable that the var. petrophila is as close to H. variegata as I had supposed, and that the var. calcarea is closely connected to both H. serrata and to var. petrophila. The other observations remain valid viz. that there is a strong resemblance between serrata and mirabilis var. sublineata; but particularly that serrata may be connected to heidelbergensis via the var. minor. However, heidelbergensis in its general relationships is extremely complex. It is interfused with H. maraisii, it may be the direct (if complex) geographic extension of H. mirabilis in the context of the relationship between that species and maraisii, and it may have affinities and continuity with both H. variegata and H. floribunda. There is still another problem: H. variegata var. modesta at Kathoek (Figs.15 JDV97/24) and Potberg (Figs.16 MBB6542). My collection of this element at Kathoek (Figs.17 MBB2551) suggests that it is possibly a small very proliferous sand dwelling serrata. J.D. Venter’s collection from the same farm is a more robust element with rather longer, more erect leaves which suggests H. variegata. My Potberg collection (fig.16) is another small element which I relate to variegata although the leaves are shorter and more turgid. It must be borne in mind that H. variegata var. hemicrypta occurs north of the Potberg on the gravelly flat so that there is ecotypic variation in evidence. Further work will have to be done to elucidate what could be an unresolvable situation i.e. the Potberg is home to variants which link all the elements so far named. The evidence for this is already available, but insufficient to crystallise a solution.
Fig. 15a. JDV97-24 H. vaiegata var modesta. Kathoek.Fig. 15b.Fig. 16a. MBB6542 H. variegata var modesta. Potberg.Fig. 16b.Fig. 16c.Fig. 17a. MBB2551 H. variegata var modesta. Kathoek.Fig. 17b.
There will be difficulty in considering the extent and the degree of similarity of H. serrata to other populations. Some of these which include serrata-look-alikes are:
Fig. 18. JDV96-52 H. heidelbergensis var . N Kathoek.Fig. 19. JDV93-51 H. heidelbergensis. Melkboom.Fig. 20a. JDV87-4 H. heidelbergensis var scabra, Leeurivier, Drew.Fig. 20b.Fig. 20c.Fig. 20d.Fig. 21. JDV97-49 H. mirabilis var consanguinea. Dwarswaterkloof, MacGregor.Fig. 22. JDV88-83 H. mirabilis var consanguinea. Dwarswaterkloof, Macgregor.Fig. 23a. JDV96-38 H. maraiss var notabilis. Agtervink, Robertson.Fig. 23b.Fig. 23c.Fig. 23d.Fig. 24a. JDV98-1 H. maraisii var notabilis, Wolfkloof, Robertson.Fig. 24b.Fig. 25. JDV97-30 H. cf. heidelbergensis. N Ashton.Fig. 26. JDV86-1 H. mirabilis var triebneriana. Stormsvlei.
Some of these populations and their variants are dealt with in other chapters (Particular note must be made of H. elizeae which has to be considered in relation to H. rossouwii).
H. serrata was coming into bud in July a good five months after mirabilis had started to flower. This shows that my predictive thoughts about the relationship with calcarea and petrophila may not be particularly sound if flowering time is considered. Calcarea flowered a little before mirabilis var. sublineata in January, petrophila flowered in February while Roovlei heidelbergensis and Leeurivier (Drew) heidelbergensis flowered in December and March respectively. Flowering time for other collections needs to be observed, noting that this is not necessarily definitive either (for example while H. mirabilis var. badia is essentially late-summer flowering, single plants may flower in winter.
When I consider how much time I have spent in the area, it was really surprising to find serrata so far west. It means that something very interesting can still be found in the large triangle Swellendam/Bredasdorp/Riviersonderend, and that exploration will have to be done on a very small scale. The Karsriver and Renosterfontein petrophila are just smaller more spinose versions of Oudekraal, and the leaves are more spotted on the reverse. Calcarea was previously known from two clones which had rather shorter leaves and, perhaps, fewer leaves per rosette. This, the locality, and the spotting on the back of the leaves would have been the reason for opting for a relationship with mirabilis. We only found it with quite detailed directions because it was such a small population and so cryptic, consisting of three tiny locations under bush with a total of about 15 clumps. One location could have been the product of vegetative propagation with six to seven clumps. The second was similar with only two clumps and evidence of porcupine activity around the clumps. The third covered about 15sqm and there were another 15 or so plants as clumps or as single rosettes. These did vary and only one clump really resembles the two I have seen in cultivation viz. the Karoo Garden plant of C. Burgers, and a plant I received from Dennis DeKok. Burgers, however, appears to have made two collections from De Hoop; this needs to be confirmed. The plants seen by me in habitat do vary quite a bit and the range of variation equates that of both serrata ex Oudekraal (the Koppies population is reduced to about five plants) and north of Bredasdorp. The mean length of the leaves and leaf number per rosette may be shifted to the shorter and fewer, respectively.
The two Bredasdorp populations of H. serrata are remarkable for the similarity of the plants to serrata at the type locality. The habitat is a little different and at the westernmost of the populations, the plants cluster much more freely than the Heidelberg populations. At Koppies the plants are on a Witteberg substrate, whereas at Oudekraal and N Bredasdorp, it is Bokkeveld shale. We may have a slightly biased image of serrata with turned out leaves; the plants can be quite globose, and I did illustrate a plant like that in my original description when I drew the comparison with herbacea.
Where serrata really differs from mirabilis is by the greater number of leaves per rosette, the thinness of the leaves, narrowness at their bases, lack of a mid‑leaf swelling and by the incurving of the leaf tip; that character is fairly significant. H. mirabilis var. sublineata grown recently from seed has proved to be remarkably variable, but none of the plants have incurving leaves. Some of the clones are very similar to serrata, but the flowering time is earlier in the year and more in keeping with H. mirabilis in that respect.
Discussion H. variegata var. petrophila and H. mirabilis var. calcarea should be transferred to the species H. serrata. The same may be true for H. heidelbergensis var. minor. However, H. heidelbergensis has a complex distribution and there are other complexities in variation that preclude a definitive statement. It is quite probable in the light of the findings discussed in Haworthia Update Vol.1, that there is no discontinuity which satisfactorily and convincingly can be claimed to distinguish species. This is probably just another example of a more general problem which I have experienced with the identification and classification of many plant genera.
I am conscious that I have not dealt fully with the problem in omitting mention of co-occurrence (I mean sympatry – when two obviously different elements grow together at the same site or very closely adjacent). I see this as the best measure of discontinuity. Thus if two elements grow in close proximity, even if not sharing the same habitat, they are probably discrete. This does not preclude the possibility that they are simply ecotypes and this is taken into consideration. However, I have before pointed out that a taxonomic solution which can be deduced in this way for populations might not be true for the entire distribution of the species concerned. It is evident that serrata generally occurs alone. At Oudekraal and at Koppies, there is a floribunda/maraisii element in the vicinity. In the case of heidelbergensis var minor, both turgida and maraisii are present. I have always been struck by the similarity of mirabilis and serrata, and by the fact that they do not co-occur. Mirabilis, in the context of this paper, is presently not known east of Napier except at Bredasdorp and Mierkraal south of Bredasdorp. This is as the var. sublineata and var. mirabilis respectively, which differ from serrata in respect of flowering time, less leaves per rosette, and the non-incurving leaves. Curiously, it appears that mutica, at least in the Bredasdorp area, grows generally alone too, although at Haarwegskloof (Bredasdorp) it does grow with heidelbergensis. At Drew it is in the close company of both maraisii and heidelbergensis, and south of Stormsvlei is near to a maraisii/mirabilis intermediate. Thus even an element as apparently different as H. mutica may presumed to be, cannot be ignored.
However, another consideration now enters the picture with the discovery of serrata so far west. There are three poorly understood elements which were reported to have come from Bredasdorp and Napier respectively. The first of these was H. bijliana von Poelln. that does, however, seem to belong rather with H. arachnoidea than with anything else. G.G. Smith appears to have had a plant of this ilk which he received from Maj. Venter 9, from Caledon. The other two, however, are H. altilinea var. bicarinata Triebner and H. rossouwii v.Poelln. They were both described from the same collection viz. Napier at Bredasdorp, Rossouw in Triebner 1059. The former was never illustrated and the description is rather brief … “Backs of leaves always with two keels and with teeth. Leaves 3-5cm long, 6-8mm broad, with 4-6 dark lines in lighter part, with two keels at back, keels with up to 2mm long denticula”. This description is in the joint paper by Triebner and von Poellnitz, but authorship is attributed to Triebner alone. von Poellnitz later retained this variety when replacing the name altilinea with mucronata (note that in the context of the names which I use, Scott confused my serrata with mucronata as well). This is very curious because it is evident from the description of H. rossouwii, which is very much more detailed, that another plant from the same collection is in fact again described. The differences are essentially that von Poellnitz gives the leaves as 5-6cm long, and 8-10mm wide as opposed to the respective 3-5cm and 6-8mm by Triebner; and with three lines on the ‘end surface’ as opposed to four to six by Triebner. This says a great deal about the detailed descriptions which are routinely called for, and which in Haworthia seldom amount to much because of the problem of look-alikes, and the variability within and between populations.
I repeat the von Poellnitz description here to illustrate that it is a very good description that nevertheless omits some key information:
H. rossouwii von Poellnitz spec.nov. (Stemless rosette, spirally many-leaved. Leaves rather pale green, nearly oblong lanceolate, acuminate, on the face somewhat retused near the apex thus forming a triangular face; on the back, towards the apex, with sub-pellucid tubercle-like spots, denticulate on the margins and keels; the triangular face strongly acuminate, sub-translucent, not tubercled, but carrying some sub-pellucid small teeth, and traversed by a few longitudinal lines).
Stemless many-leaved rosette about 5-6cm in diameter. Leaves more or less erect, 5-6cm long, 8-10mm wide, about oblong-lanceolate, tapering gradually, pointed, slightly widened at the base, almost straight or a little bent to one side near the tip, rather light green, dull. On the face flat or usually somewhat convex, near the tip slightly bent backwards (retused) at an angle of 20 degrees; on the back, from the middle outward or near the tip obliquely keeled, or somewhat more rarely with two keels, near the tip with a few, often elongated, very often raised slightly pellucid spots, and frequently with a few leaf-coloured tubercles. The end-surface half-pellucid, 20mm long or somewhat longer, 7-9mm wide at the base, triangular, tapering very gradually, pointed at the tip often slightly curved inward, somewhat convex, smooth, rarely with a few indications of tubercles, but with some semi-translucent teeth (these teeth loosely arranged in a lengthwise middle line, now and then also in another line nearer the margin and very rarely scattered about in small numbers) with three long, occasionally interrupted, green lengthwise lines, of which only the middle one may reach almost to the apex, and usually with a few very short line; leaf margins and keel with pale teeth, or near the base often leaf-coloured, teeth, up to barely 2mm long; end-bristle pale, with small side bristles, 5-7mm long.
… On account of the teeth on the end surface stands near to H. schuldtiana and to H. paradoxa (I treat these respectively as H. maraisii and H. mirabilis var. paradoxa) but is easily distinguished from both. Its flowers are thus far unknown”.
The omissions are the flower of course, and there is no stipulation of the approximate number of leaves, and leaf thickness. The weakness is that it is a single plant description, evidenced by the fact that another plant from the same collection is accepted as a variant of a totally different complex.
In the past I have I regarded rossouwii speculatively as a synonym of H. mirabilis, due largely to the fact that it was recorded from the Bredasdorp area (and to the ambiguity of the type). Another reason was the similarity I observed between the illustration of H. rossouwii in Kakteenkunde with H. heidelbergensis var. minor and with H. mirabilis var sublineata. I opted for the latter (with which Scott was in accord) and the similarities are also recorded in the discussion above. I dismissed bicarinata as insufficiently known because I assumed that Triebner may have had some better reason to have equated his specimen with the Little Karoo H. mucronata. Possibly there has been a mistake in the Triebner numbers. It is this discovery of serrata so close to Bredasdorp, and equidistant from Napier, which revives and also casts new light on the problem. I conclude that my H. serrata is a later synonym of H. rossouwii of von Poellnitz.
There are still two other considerations. In my 1999 revision, the fact is that I placed paradoxa as a variety of mirabilis while discussing the possible connection of these elements via calcarea. In the light of this paper it is evident to me that paradoxa could also be linked to serrata rather than with mirabilis. The same is true for emelyae var. multifolia. I would not like to commit myself to this probable solution at this stage, as I think more evidence should be obtained. It is also improbable that, however attractive, the solution will not avoid impacting on H. emelyae, H. magnifica and H. heidelbergensis. It is also evident that the requirements of the ICBN are that should paradoxa indeed need to be incorporated into serrata, more name changes will result, as paradoxa will have priority over rossouwii. My opinion is that this is a fundamental weakness in the code, which inhibits speculative treatments, and also which results in smaller taxa becoming the typical main element of a species. My further opinion is that it is a weakness of the Code that the description of an element automatically creates an equivalent taxon. This may be acceptable and desirable where the elements are clearly separable and geographically defined, but not where the typical element is required to absorb any other variants which are not so defined. Thus H. paradoxa var. paradoxa could become the main component of a species where it is actually a geographical isolate. In a system which does not have an adequate general definition for ‘species’, this probably does not matter; whereas it might if there is a definition which suggests that the ‘species’ is an operating living system (see Bayer in Asklepios 77:3, 1999).
The formal statement of this paper, which would better be regarded as an hypothesis subject to test, is thus as follows:
H. rossouwiivon Poelln. in Kakteenk.7:75 (1938). Type: Napier near Bredasdorp, collected by Mr Rossouw, South Africa Police in Napier, Triebner 1059. Not preserved. Lectotype: designated by Breuer & Metzing 1998, icon in Kakteenk. non syn. H. mirabilis sensu Scott 1985, Bayer 1976, 1982, 1999.
H. altilinea var. bicarinata Triebn. in Triebner & von Poellnitz in Feddes Repert.Spec.Nov. 45:170 (1938). Type: Napier at Bredasdorp leg. Mr Rossouw (=Triebn.1059). Not preserved.
H. serrata Bayer, Jl S.Afr.Bot. 39:249(1973). Bayer :55(1976). Bayer :147(1999). non Scott :62(1985). Type: CAPE‑3420(Bredasdorp): Oudekraalkop, Heidelberg (‑BA), Bayer 166 (NBG).
H. rossouwii var. calcarea (Bayer) comb.nov. H. mirabilis var. calcarea Bayer in Haworthia Revisited :110(1999). Type: CAPE-3420(Bredasdorp): De Hoop (-AD), C. Burgers 1648 (NBG, Holo.).
Material seen:- 3420(Bredasdorp): NNE. Buffelsfontein, Potberg (-BC), Burgers 2018 (NBG). De Hoop (-AD), C. Burgers 1648 (NBG), Bayer 6985.
H. rossouwii var. petrophila (Bayer) comb.nov. H. variegata var. petrophila Bayer in Haworthia Revisited:159(1999). Type: CAPE-3420(Bredasdorp): Renosterfontein (-AC), Burgers 2158 (NBG, Holo.).
The provisions of this solution are that the epithet paradoxa may be instated preferentially, and that emelyae var. multifolia and variegata var. modesta also be included.
A second provision is that the species H. bijliana v.Poelln. be rejected as insufficiently described and sourced, and that the type (the illustration in the Berlin herbarium) is ambiguous – as indeed are many Haworthia types and descriptions based on single plants and not correctly tied to geographical origin. This is demonstrated by the citation by VPoelln. (Feddes Rep. 1936) of this species viz. bijliana from Klawer and Eenriet and also Feddes Rep. in 1937 from Springbok – which cannot be anything other than H. arachnoidea. In this last publication he cites a specimen from Bonnievale, another from Ezeljacht Oudtshoorn and also sinks fergusoniae here which in fact was supposed to have come from ‘near’ Grahamstown.
H. bijliana Poelln. in Feddes Repert. 27:134 (1929); Bredasdorp, Mrs va der Bijl. Type van derBijl s.n. in (B). Not preserved. Lectotype desig. Breuer 1999, unpublished photo. icon. (B). Species rejecienda.
Acknowledgement I am grateful for the company on the field trip of Lawrence Loucka of Connecticut (USA), and that of my wife, Daphne. The assistance of Paul Botma of Cape Nature Conservation is acknowledged over and above the general acknowledgement to that Department. Mr. C.L. Neethling, Mr. Thys Swart and Mr. Pieter deKok were most kind and helpful in respect of access to sites. I am also grateful to Kobus Venter for support, advice, and constructive exchange of ideas. Not least to Steven Hammer, with comment from Bob Kent, who was so kind as to read this manuscript critically and make corrections. Residual errors will be mine.
Addendum Alsterworthia Vol 1:13(2002) “A comment on Bayer’s latest taxonomic contribution” by Ingo Breuer. [link] With reference to the comments in Alsterworthia 1:13(2002) regarding the identity of H. rossouwii and H. serrata:
Notes
Fig. 27 H. rossouwii. Vpoelln. type.Fig. 28a. MBB6984.1 H. rossouwii. Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.Fig. 28b.Fig. 29a. MBB6984.1 H. rossouwii. Colour photo originals of the preceding. Fig. 29b. Fig. 30. MBB6984.2 H. rossouwii. Nooitgedacht, Bredasdorp. A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second. Fig. 31. MBB6983 H. rossouwii. Soutkloof, N Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might. Fig. 33 Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Fig. 27 A copy of the von Poellnitz lectotype of H. rossouwii.
Figs 28a & b Black and white profiles from two angles, of the same one plant actively growing, undessicated and turgid. The plant is a clone from Bayer 6984 Nooitgedacht, NW Bredasdorp. This is correctly H. rossouwii, and correctly H. serrata.
Figs 29a
& b. The colour originals of the preceding.
Fig.30 A second plant of MBB6984. This is a more vigorous grower and it shows the differences that obviously confound the uninitiated. There were not many plants at Nooitgedacht. I only collected from two clones and this is a photograph of the second.
Fig.31 A photograph of a plant of MBB6983 Soutkloof, north of Bredasdorp. Showing a plant in which the convexity of the mid-upper surface (and concavity of the lower surface) is virtually absent. I could have, from this sample (MBB6983), produced pictures to match Figs.27 to 29. So no one should claim that Fig.30 is a different species as someone now well might.
Figs 32a
& b. Scanned and manipulated copies of figs 28a & b.
Fig.33
Flowers of MBB6672 H. maraisii Koningsrivier, SW Robertson.
Figs.7a, b & c. These are clones of JDV86/5 H. heidelbergensis var. minor, Rooivlei, NNE Bredasdorp. This is a small population, of also small plants, from northeast of the collection MBB6984, Nooitgedacht. The plants partly resemble H. rossouwii from there, and partly the var. petrophila from Karsriver. Classifying them as a variety of H. heidelbergensis is not necessarily correct. Considering habitat and substrate, one must consider that ecotypification is playing a role, and that was habitat available, there would be further populations which would confirm continuity with these known collections.
Regarding Figs.28a & b. A bit of play with light and focus and the keel and under-leaf spotting could be varied. I would like to suggest that they be considered them faded, less sharply focused, printed with less contrast and on a coarser grained paper. Figs.32a & 32b are in fact such scanned copies of Figs.29a & b. I do not think it even necessary to consider that the original rossouwii of Fig.1 endured a long journey in the post to a foreign land. The assertion by Breuer in Alsterworthia is that not only are the plants I figured as H. rossouwii different from Fig.1, but that this Fig.1 lectotype is “probably” of the maraisii-complex. This is at variance with…
1. the Triebner description of the same collection as a variety of H. mucronata;
2. with Col. Scott who regarded it as H. mirabilis. What to say of von Poellnitz’ own assessment – the man who described both species i.e. maraisii and rossouwii in the first instance.
3. with the facts of the pictures before you.
What should an observer make of maraisii growing very near to rossouwii at Oudekraal and near to rossouwii at Napier? Can we really see a closer resemblance of Figs 27 & 28 with those long attenuate, double-curved, thin leaves – to H. maraisii rather than to H. serrata. I truly do not see that such a judgement can be seriously considered in the light of Breuer’s interpretations. It is of course valid when one considers all the variants that I illustrated in my article above and in terms of my over all classification.
Generally these plants of the SW Cape tend to have the double curvature of the leaves as is evident in the VonPoellnitz lectotype, but one can find in the same population and/or taxon, clones with incurved, outcurved or erect leaves (see fig.30). Broadly speaking H. maraisii has thick recurved leaves. But this is a non-argument even if one (outside of a “lumping vs. splitting” red-herring) considers the complexity of the so-called maraisii-complex, and I am sure that Breuer does not. I frankly think that most observers cannot even dream how complex it is and that it is just braggadocio to write as though anyone can. Breuer implies that one can make the inference of similarity between rossouwii and maraisii from the illustration (Fig. 27), the description and its given locality. This is patently and self-evidently false as is evident from the fact that I used this same information to reasonably deduce the similarity of rossouwii and serrata.
I did not originally submit a copy of the von Poellnitz lectotype, perhaps because I felt that the response “you will see that rossouwii is quite a different plant from serrata“, is what I could have expected from an uninformed audience! The fact is that all the evidence, circumstantial and descriptive, show that they are the same. Including the lectotype would have just been “overkill” and I have never before felt required to write for a readership with less than common sense or basic knowledge, nor for obstructive readers.
It should not be necessary to say this, but the character of the flowers cannot be specified in the way that writers are inclined to suggest. They have this same enormous range of variability and it is just fraudulent to say that there are floral characters by which these taxa (the varieties of H. rossouwii which I now recognise) can be unmistakeably differentiated. It is the typological concept which drives people to think there is a single description which can encompass the description of a taxon. This is nonsense. It is a statistical problem and we have the difficulty of establishing the probabilities associated with “decision”. Breuer writes “… von Poellnitz’ description probably depends only on 1 or very few selected plants and no other records are reported for this taxon”. This is circa 1945 circumlocution, it is unnecessary and it is untrue. There was never a question of “selection”, and furthermore we have the Triebner description of the same collection as H. altilinea var. bicarinata. Why this sudden doubt about the interpretation of types. It is a shifting of goalposts.
There are few collections in flower now in April, but I could take this picture of three flowers from the one collection MBB6672 H. maraisii – Koningsriver, SW Robertson (Fig.33). It is not an exceptional case or an exceptional collection. But one can see the differences in thickness of peduncle and variation in the individual flowers. It can happen like this, albeit to lesser degree, on the same plant. Colour is also frequently quite variable between and often within collections. Many years ago I undertook to illustrate all the flowers in my maraisii collections. But they were so variable that I just gave it up. The variability in the flowers mirrors the variability of the rosette and the latter is just a little easier to deal with. One has the added problem with flowers, of the changes which take place during the opening and aging process. So then how does one define the flower, say, of H. mirabilis. It cannot even be done for the range of leaf rosettes, and this has now been shown in the literature many times.
May I also point out that one of my understandings of botany and plant classification is that there is a tendency to overreach ones talents. I have freely admitted that I am there because of the paucity of talent. I frankly do not think the environment is enriched by the particular article I now refer to in these added notes.
I can point out that there are writers who really persist in flogging dead horses. The result is phantasmagoric fanfaronades. The article concerning the name H. correcta (Alsterworthia 1:6, 2001) is not particularly accurate either and indeed it may be my own fault. Smith explained that Mrs. Blackburn did not in fact collect correcta ‑ it was a Mrs. le Roux and the plants came from near Vanwyksdorp. Later Mrs. Ferguson sent a plant from 26 miles east of the town. Thus from this information H. correcta has to be a later synonym for H. emelyae.
Regarding types:- the painful realisation was actually expressed by Schelpe in the distant past, and brought home to me many times. These old types are doubtful and ambiguous. I was faced with that option long ago – just forget them all, or try to reach a compromise with the understanding and information available at the time. H. rossouwii is a case where there is real new information which would have made me use the name rossouwii in 1976 and before, had it been available. But here, had I persisted with the name serrata for this new material MBB6983 and 6984, there is absolutely no doubt in my mind that I would have been criticised for failing to see the connection to rossouwii. Not that this would concern me unduly, for I think there is actually very little benefit in this mindless re-interpretation of types which takes us nowhere. Notwithstanding, and without compromising, anything so far written:- the really disconcerting thing about the whole matter is that without the locality data, it will be possible to generate an argument bolstered by illustrations, to show that rossouwii was in fact H. decipiens, H. cooperi or H. herbacea (among others), of the option less improbable than H. maraisii.
The closing paragraphs of the article are a rather hackneyed parody sung by most “wannabe” taxonomists. Of course it is right when Breuer says if we lump it means we would have to lump many other things. It is just as true to say, if we split we have to split many other things. That does not make sense either. This is ancient stuff of 60 years ago and it seems strange that it still provides so much fodder for writers, who have in fact no rational species concept to begin with. The Linnaean system was based very largely on floral characters and hence taxa at all levels are based on “related floral characters”. To suggest that this pertains in some special way to “series” is not correct. This sums up the article in Alsterworthia. ♦
This essay was published in Haworthiad 16:86, 2002.
I wrote an article about Haworthia rossouwii in Aloe 38:31(2001), in which I resurrected this old name to replace that of my own H. serrata. This was necessary because I had found this plant (because of its localisation and its abundance there, it is better to say ‘this species’) at two places near Bredasdorp as opposed to where I had described my species from near Heidelberg.
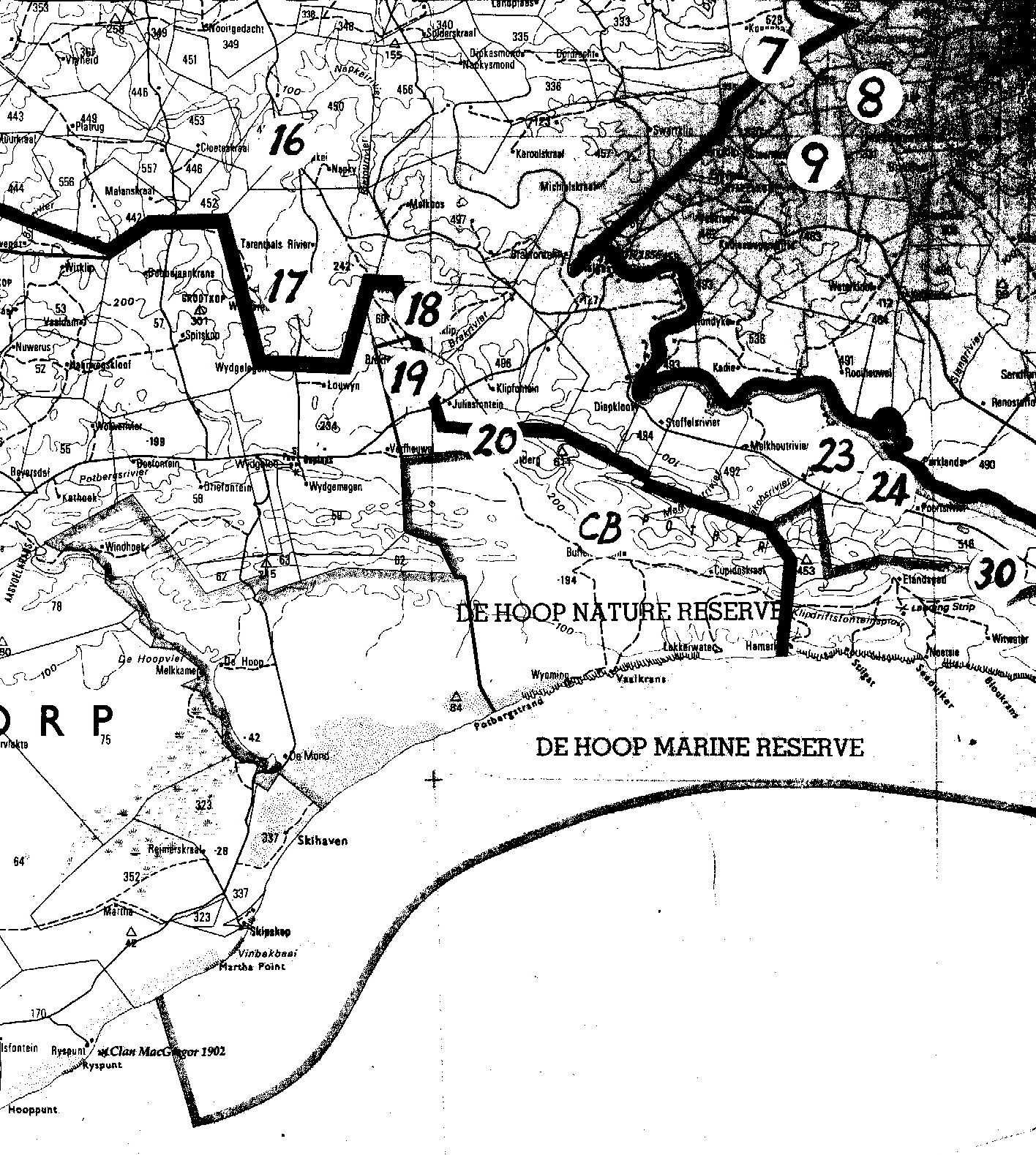
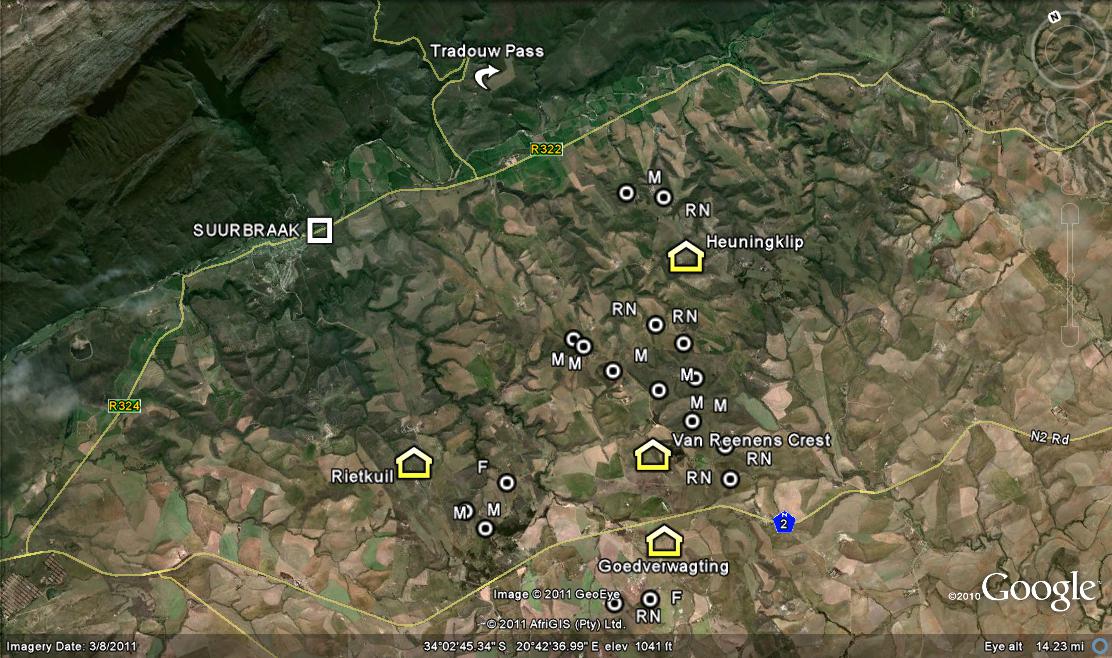
One needs to know something about the geography and geology of the Southern Cape (and the Overberg as a part of it is also known) to really follow all the ramifications of any discussion about Haworthia, including this one. In fact one needs to know a whole lot more, and I will also try to explain that and its implications for the collector and Haworthiophiles. This “whole lot more”, I will call the Corporate Mind because it includes so much – so remember CM! If I regard H. rossouwii as a species, I have to consider all the plants and all the places where they grow in order to determine the nature of this particular system of living things. As I explained in my article, there is a problem with the fact that little is actually known about the Haworthia of the Overberg. They occur in small populations scattered over a fairly wide area which has been heavily impacted on by agriculture. Thus about 90% or more of the Overberg is now wheatfield or pasture. Like Gasteria carinata, which is also a Southern Cape species, Haworthia is associated with rocky outcrops and thus also with the geographical erosion and drainage systems of the area. It is quite probable that cultivation has had relatively very little impact on Haworthia in terms of actual available and suitable habitat.
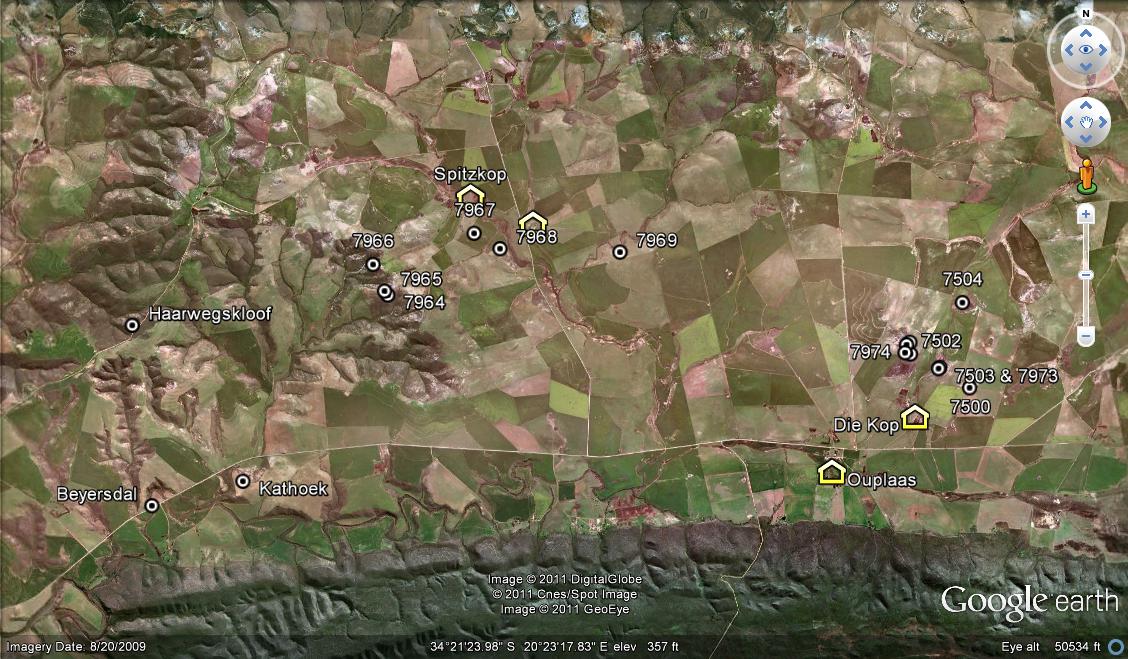
The problem is to get to all the small potential habitats and explore these at a scale which will help to resolve the problems of variability, relationships and classification. While writing about H. rossouwii, I was very conscious of this problem and the fact that my explanation is not complete. There are some collections by Chris Burgers which I have not seen, and there are many potential habitats which I have not explored either. The place Kathoek and H. variegata var. modesta which grows there is a particular case in point. Kathoek is at a point midway between the Breede River and Bredasdorp. It is just east of the De Hoop wetland/stream and on the northern edge of the Coastal limestones. In the west, also on limestones, is H. rossouwii var. petrophila which I first placed with H. variegata. In the south (also on coastal limestone) is H. rossouwii var. calcarea, which I originally had with H. mirabilis. To the north is H. variegata var. hemicrypta which is on Silcrete of a quite different geological origin. But H. variegata variants, as I interpret the populations, are also on and about the Potberg to the east which is Cape Sandstone.
My problem is “How do these populations actually relate to one another?” What is missing? What can I find to better explain their similarities and their differences? So at the back of my mind is the wish to explore the Coastal limestones between Bredasdorp and the Potberg and find out more. But it is not as easy as that. I also want to know more about H. mirabilis, H. maraisii, H. heidelbergensis, H. mutica, H. minima, H. marginata and H. pumila, which all also occur in the Overberg area and each have question marks attached. They each present problems that I would like to resolve for my own satisfaction, without being confused and befuddled by others who are toying with the same body of knowledge at different levels of expertise and ignorance. In this play we are using words, terms, names and conventions. For this play we need to have a commitment to truth and use the same language or else it is all just meaningless.
So we come to what took me to the Overberg in November 2001. Was it was just my curiosity about all those inviting rocky unexplored places? Is it to learn something more myself? Is it to impress anybody? Actually it was many things including a fascination with chameleons – largely my wife Daphne’s. In March 2000, Daphne and I had come across a most remarkable chameleon at Napier. It was on a small shrub among whitish dried grass growing on a calcareous outcrop strewn with white quartzitic pebbles. The tiny reptile was ivory white, with a stunning array of small orange tubercles on sides and back. One has to know the Cape Dwarf chameleon (Fig.1) and its cousin, the Robertson Dwarf, to know how beautiful and amazing these creatures can be. Another thing was H. mutica and my fascination with collections of this species and their histories of discovery and exploration which are stories of their own. Still another was the news that the Botanical Society had funded a researcher to explore the remnants of the vegetation of the wheat areas. What had he found? Apart from Haworthia, I had once been involved in a preliminary attempt to map and identify vegetation remnants for conservation purposes, and had vainly tried to draw attention to the curiosities of the Silcrete outcrops and remnants of vegetation among the wheatfields.
This researcher was an old acquaintance, Nick Helme, who kindly told me what Haworthias he had seen in his exploration. There was not much new, except for H. mirabilis east of Caledon, and this became our first stop. The site is south of the main area for the species which is along the Riviersonderend river north of Caledon. It does, however, occur in recognisable form south-east of Caledon, also around Napier where it transmutes to some degree (inter alia to the var. badia), and then also at Bredasdorp. There it also transmutes to the var. sublineata (sandstone) and var. mirabilis (the old mundula – and there may be some other way out, but for me this represents the low cussedness of botanical nomenclature). What H. mirabilis does to the north and east of Riviersonderend actually fully explains the problem of classification of Haworthia, and who knows if anyone will ever write that explanation – and truthfully.
Nick Helme’s site is a deep valley which drains to the south. Typical of the area, are these small drainage areas which seldom have permanent water. There may be odd pools lasting into mid- or late summer. There are often exposed shale ridges which host a particular array of succulents and other species – and are often home to owls too. The first problem is access and where does one find the landowner after one has succeeded in finding the place? If one does find the dwelling, will it be the right one and will one find the landowner. Will it be a big aggressive StBernard and more aggressive and ugly Rottweiler reflecting the mindset and security fears of the owner and grossly offending my own, like once near Worcester – or will it be a lonely and over friendly sheepdog. November is a very bad time. This is when the wheat harvest is on. At the end of a bad season, when the crop is drying and it has to be cut and reaped the weather is playing its usual strange tricks, farmers cannot be expected to be approachable. But we did find a helpful soul. From the farmhouse, we drove twenty kilometres back to the main road to the roadside stall, where a family member, Herman DeKock, gave us the necessary permission to venture onto the property.
It does seem an easy business, but in fact it becomes very complicated. Nature Conservation regulations are strict, and land ownership, presence, trespass, stock theft, personality and who knows what else, influence the mindset when setting out an expedition of this kind at all. With those problems all out of the way, and only then, is one able to really enjoy being out in the field and communing with nature – which it is all about in the first place. So we found Nick’s site. We climbed through the fence (s) and tried not to disturb the forlorn sheared, miserably self-conscious sheep that clustered around the earthen dam. It was hot and dry and the most promising spots were quite a long way off. One has to plan one’s approach as the southern slopes are often quite densely vegetated and very uncomfortable to traverse. Snakes are also a consideration and especially in early summer. On this trip we never saw one, although Daphne surprised, and was surprised, by a Lynx drinking at a waterhole. So we did find the plants that Nick had said and took a photograph for the record (Fig.2). Just ordinary H. mirabilis var. triebneriana (cussedness – there should be no typical variety, just a broad general name mirabilis).
From here of course we went to the chameleon at Napier. What a laugh! We had difficulty finding it on the same day on the same bush, five minutes after first seeing it. So expecting to find any chameleon at all, and I have never seen a chameleon in the field in all my years of plant exploration, was quite the wildest dream. But with a dossier of 60 individuals recorded on our small-holding at Kuilsriver over a period of about a month, we felt entitled to hope (all contributing to CM). It is also a most wonderful excuse to be out in the country.
We headed for De Hoop. This is also quite a story. Where do we stay? The DeHoop Nature Reserve is a wonderful place with great accommodation, but it is not where we wanted to be. The staff does not know what it is we are looking for, what our CM data base contains and what our needs are. We want to see the area around the north of the wetland. This is half private land, and half state-owned. Besides there is a travellers Lodge on the edge of the reserve which we understood offered easy and official access to the reserve, presumably to near where we wanted to be. Not so. It is one thing dreaming about this exploration while sitting at home with the map, and quite another to be faced with the problems of roads, fences, ownership, dogs, snakes, flies, mosquitoes, permissions, heat, cold, wet, terrain, bush, visibility, time, distance, misinformation, local “knowledge”, incipient old-age, food, water and the spirit of inspiration and energy which drive the legs (CM). So the Lodge was an unfortunate mistake (we did look, and we did also look where we were told an Haworthia did grow – it was Gasteriacarinata), and we did end up at the office at DeHoop Reserve. You see over the years, one comes to realise that helpful and kind as people by nature are, an awful lot of time is spent chasing red-herrings and wild geese (CM). The help and information one gets is very often of a negative hindrance rather than a positive help. So my real gratitude to the manager there, Mr Ben Swanepoel, who without serious qualm, entrusted us with the key to the northern part of De Hoop where we needed to be.
Unfortunately the problems listed above rather limited our activity. We exhausted our energy climbing up and down rocky slopes in an area which should have occupied us for a week. It is an area which has to be explored by someone at sometime. We found naught, which should not in the least be interpreted to say there are no haworthias there. The terrain is formidable and also most promising. But CM called.
We moved to accommodation closer to our budget restraints and nearer to our goals. Driefontein, where the owner graciously made the necessary contacts for us. What a place – the cold vegetated coastal limestone behind, and featureless cultivated endlessness in front. I have an old record of a collection of mine of H. mutica from “Beyersdal”. Subsequent travels indicated that place, name and memory did not match. So we went to Beyersdal. We did find mutica and it was a different place and a new record (Figs.3a, b & c). Who can actually describe the wonder of plants like these? In tiny patches of rock in a sea of wheat and low-grade (by rainfall and vegetation standards) pasture. Also the quiet, the two owls, also Gasteriacarinata again, this time the Haworthias among them – unusual. The extraordinary colour of the plants. This time the bronze colours of badia. Nearby we also visited an old, by thirty years, locality for H. heidelbergensis. Still there and astonishingly in among white quartz in a small outcrop, but in two little groups occupying hardly two square meters in the approximately two hundred available (Fig. 4).
Then we went to Vanderstelkraal. We had met the owner, Mr Adam Albertyn, who very kindly let us wander over his farm. CM. How does one explore a place like that without living there? It is on the edge. The agriculturally useless coastal limestone at the back, and the productive massively utilised Bokkeveld fields at the front. If terrain had allowed, there would not be so much as a hedgerow to have conserved anything. We saw some very interesting sites and exhausted quite a number of things, like energy, time, passion for the quest, tolerance to bad weather, ostriches, tight-strung barbed-wire fences, no roads and also to the heaviness of the overwhelming destructive productive activities of man. We did not find Haworthia. But I omit a lot, including all the good farming. Mr Albertyn owns the eastern side of the De Hoop wetland where he has a 4X4 route – and there is also a sixty-year old legend of H. minima in that particular area which I would like to confirm. Not this trip though. We did try to the west later, where CM eventually prevailed against us – aided and abetted by overgrowth of alien acacias and the negative vibe they seem to generate (by the side, at Vermaaklikheid, Riversdal, H. mirabilis var. paradoxa grows/survives under Acaciacyclops).
Then we decided to climb the mountain behind Kathoek. Why? I cannot remember why I climbed there in the first place, or how I even did it. Then I found plants in a burnt out area, under the remnant blackened skeletons of Protea – natural fire cycle encouraged by farmers. It does produce a grazeable growth and also invasive aliens. The plants of H. variegata var. modesta were just discernible sticking out from loose sand. Kobus Venter later also visited Kathoek and found plants which were more robust. The plants have spreading variegated leaves (in cultivation?) and while I had no real problem associating them with Karsriver, viz. H. rossouwii var. petrophila, they seemed to have a greater affinity with variegata at the Potberg and Luiperdskop. But my collection and Kobus’ seemed at variance. So Kathoek was a must. From the road CM prevails and it looks quite an expensive option in terms of CM resources. Actually the climb was quite easy. We found the plants before we expected to. They did not look like what we expected them though. They were densely clustering (expected) and in rock crevices (Figs.5a, & b) and pans (Figs.6a & b). The rosettes were tiny and reminiscent of even H. reticulata (see Fig.7). At one place a dense cluster under and in a restioid grasslike plantlet, looked just like a yellowish-brown windblown fallen mass of dry leaf bracts (Fig.8). What impression did it make? Nothing new. It provoked the existing doubt. The population is very similar to petrophila and less so to variegata and hence this variety, with the exclusion of the Potberg population COULD BE transferred to H. rossouwii. But this does not help much, because the Potberg var. modesta is less like the northern Potberg’s two populations and the Luiperdskop population of H. variegata var. hemicrypta, and not more like petrophila! So the CM has to carry this anomaly together with the position of calcarea until whatever else to be found is found. These finds are not going to make things easier to resolve in the CM, nor to explain in a way which assuages the CM of anybody else.
All the while we were pondering other clues to the nature and relationship of things. Like the endemic Braunsiivanrensburgii; a curious specimen of Carissabispinosa so laden with large red fruits it resembled a Pyracantha; Euphorbiasilenifolia everywhere, and what a strange distribution this summer-deciduous geophyte has; and what about Asparagus; what is the relationship of A. capensis, A. mariae, A. suaveolens and A. stipulaceus? Where was Aloe brevifolia or A. saponaria? What was that pugniform Euphorbia at Driefontein or the bayeri-like one at Vanderstelkraal?
Unexpressed CM – we are using my son’s LDV as in my own car anything below well-maintained road is not negotiable. CM – the cellphone we arranged for failed. There is a call in my sub-conscious ‘vehicle needed back home’. We end the trip a day earlier and the message turns out to be true as there is a breakdown of one of the other delivery trucks.
On the way home we pick up some loose ends. We call at a spot north of Bredasdorp where in my lucky ignorance I once found mutica. In several subsequent visits I could not locate it, until one year Daphne and I together with Lawrence Loucka had a very thorough search of the small spot. We found about five plants in a spot about 1/2sq meter. At a later visit we found that the plants had seeded and there were many seedlings but crowded together with seedlings of Aloeferox. Generally Aloe and Haworthia do not mix. Aloe need more root room than Haworthia and Haworthia cannot tolerate competition from larger plants. So we thinned out both aloe and haworthia seedlings. This visit revealed the successful fruits of our activity (Fig.9). The two bigger adult plants were almost being pushed out of the ground by a burgeoning aloe seedling, but there were about 15 other vigorous and healthy plants to carry the group forward. CM – this was an interesting repeat observation of the small distances that seed and seedlings actually do commonly disperse. I have seen the same thing with hybrids where the hybrid is often within the multiple body of one of the parent species.
Mission ending, we made our last visit to Drew. This story is written more fully elsewhere. We had found the pollen source for a plant of H. mutica which I collected there more than thirty years ago and which Kobus and I had taken to the field again to see if seed would set. This surviving single plant is now a collector’s dream which I dub “Silver Widow”. No one had succeeded in finding plants around where a huge centre-point pivot installation now graces the landscape. We were sure the plants were gone, and taking the plant back there was a kind of forlorn gesture of regret that more had not been done to conserve the population – and that so much had been done to eradicate the plants (two hundred plants had been collected and sold to Triebner circa 1935). If anything can be done or could have been done at all to have saved its family? But it worked and we got seed. Where did the pollen come from The pollinator is mainly a solitary bee with a fairly limited flying range. The pollen parent had to be somewhere near. Daphne and I had made several excursions to add to the many different attempts that are known to have been made to find them. Finally I decided that we just had to look again and the plants had to be where we had failed to find them before. So what happened? We left the farm house and walked straight to the spot (getting ‘stung’ by the electric stock fence on the way) less than 300m away and probably about 150m from where we had left Silver Widow to be pollinated. There were about 35 plants (Fig.10) in an area of about 3sq m in flower under Eriocephalusafricanus. So now we returned to successfully collect seed.
The curious thing in CM and all this discussion about big things like Continental Drift, and Biomes, and Vegetation, and Habitats, and Ecotypes, is that H. mutica is not particularly concerned. Here at Drew it grows in an ancient river-boulder situation in what we can call renosterveld under Eriocephalus. At Beyersdal it is on the edge of a stream-bed on exposed Bokkeveld shale with Gasteriacarinata. At Hasiesdrift it is on a gravelly bank (Bokkeveld shale) with dense Aloeferox. Near Napky it is on silcrete with grass. West of Swellendam, it is on whitish-clay, probably associated with silcrete and growing within a restioid species and with Gibbaeum esterhuyseniae. I have also seen it on a fairly bare hillside with a scattering of quartzitic pebbles where the parent shale is too dense and shallow for cultivation or to support denser vegetation. The places hardly have anything in common other than their more general geographic containment, climate, and low biomass potential. How, ever, did they get to their widely-spaced, isolated island sites?
So the tale ends with the reflection on the criticism that I have not treated H. scabra and H. starkiana in the same way I have H. retusa and H. mutica. I have been “inconsistent”. Readers can research and educate their own individual CMs (corporate minds) to the point that they can form opinions which are consistent to the point and purpose of any communication. Aren’t we lucky that the Robertson Dwarf chameleon is a small animal (Fig.11 – by Scott Russel).
Acknowledgement: This fun work is really just about people. Daphne, who kept me company. Janet Albertyn at Nachtwacht, Herman Dekock of Jongensklip, Thys DeVilliers at Boskloof, Tom Ambrose at the cement-block operation at Napier, Rory Allardice at Buchu Rest-camp, Ben Swanepoel at De Hoop Cape Nature Conservation, Annette VanEeden of Driefontein, Mr. Dawid Beyer of Beyersdal, Adam Albertyn of Vanderstelkraal, Theo Blom of Soutspanvlakte, Jan Joubert of the adjacent farm, and Beth and John Humby of Sanddrift. Their good-will and kindness is unforgettable.
Fig.1 Cape dwarf Chameleon – Bradypodion pumilum. Markings, colour, tubercles and spines are individual characteristics. It is said that even the genus is in doubt!Fig.2 Haworthia mirabilis var. triebneriana, MBB7059 Jongensklip, Caledon. Note the reddish spines.Fig.3a Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. This plant is exposed to full afternoon sun for at least a few hours each day. Note the mucronate (pointed leaves).Fig.3b Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A more general specimen with rounded leaf-tips.Fig.3c Haworthia mutica var. mutica, MBB7060 Beyersdal, Bredasdorp. A particularly compact specimen. In this population, the waxy sheen of the species seemed less conspicuous.Fig.4 Haworthia heidelbergensis var. minor, MBB7063 Beyersdal, Bredasdorp. Varietal names are just a geographical convention, as indeed even the species which I recognise, seem to be.Fig.5a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster under a small rock overhang.Fig.5b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. A cluster in an exposed crevice and visible as a dark mossy fill.Fig.6a Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill.Fig.6b Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. In a shallow rock-pan on the crown of the hill – a closer view.Fig.7 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Very reminiscent of H. reticulata say south-west of Robertson.Fig.8 Haworthia variegata var. modesta, MBB7061 Kathoek, Bredasdorp. Barely recognisable to the undirected eye.Fig.9 Haworthia mutica var. mutica, MBB6982 Hasiesdrift, Bredasdorp. Nine seedlings in a tiny island of plants, in a bigger island of Aloe ferox and surviving vegetation on the edge of a wheatfield.Fig.10 Haworthia mutica var. mutica, MBB7064 Sanddrift, Drew. This lovely plant is nearly 120mm diam. and the largest of about 50 plants. They survive, and have survived in this small heavily impacted area for so long. There is no where for them to go when options must have been very limited to start with.Fig.11 Robertson dwarf – Bradypodium gutturale, photo by Scott Russel. Zoologists cannot determine if the Robertson dwarf is different from the Karoo dwarf, or from the two subspecies of the Namaqua dwarf. They use the same fuzzy logic that botanists do, plagued by low sampling and difficult application of statistical principles and probabilities.
The problems of species classification of Haworthia should now be well known to all enthusiasts of this interesting genus. I have proposed and maintained, with cosmetic changes, a nomenclatural system for it since 1975. It is a system with which I have managed an extensive collection and herbarium record, and I know it works within the limitations imposed by the evident fractal nature of “species” and their variability. In this paper I would like to expose these limitations with respect to the concept of two species viz. Haworthia mirabilis, and Haworthia maraisii, where there may be only one. In the original hardcopy publication of this article, the illustrations are all captioned H. maraisii when it would have been sensible to have used H. mirabilis.
In most discussions concerning the classification of Haworthia, participants have suggested that there are too many species and that some of them should be “lumped”. On the other hand, there have been several writers who, as prospective taxonomists and experts on the group, continue to expand the range of entities at the formal rank of species and varieties despite all the evidence and indications that this could be an endless path leading nowhere. My own inclinations have been to minimize the number of species and to use varietal rank in two ways as a communication medium. One is that I have tended to reject varieties of older authors which I did not think had a strong enough geographical base, but also recognizing that there is nevertheless some information inherent in such names. Often I felt these varieties simply expressed the variability in a way that was insignificant with respect to the species as a whole. Two, is that I have described some new varieties to provide names for morphological variation that I consider is new and previously unrecognized, accepting that these names should perhaps not be immortalized either. Thus my proviso has been that these new varietal names should not be taken too seriously. As far as the number of species is concerned, I know full well that there could be fewer species. I get caught up in the problem of identifying a “species” in a strict botanical definition of the word, as opposed to the need for “names” as a way of simple communication about the plants in the amateur fraternity. Because of the problem of similarity and continuity, the elimination of names becomes similar to that of falling dominoes and the question then arises of “where does it end?” My classification should not be confused with a system of names intended for horticulture or for trade. But neither do I think such a system should adulterate a formal botanical one.
Therefore, underpinning this presentation is a definition of a species as dynamic systems of living organisms morphologically, genetically and even behaviorally, continuous in space and time.
What is an Haworthia species? I do not accept that there are different kinds of species nor do I accept the term “biological species”. A species is a biological construct intended for life form and the classification of Haworthia has been presumably intended to identify those individual species that constitute a genus set. It seems evident that the commonest element of any species concept, expressed or unexpressed, is that of reproductive isolation i.e. members of different species cannot interbreed. In Haworthia this evidently does not apply because there is compatibility between individual plants of even different subgenera, what to say of intergeneric hybridization. If there are breeding barriers they are the obvious isolationary ones that are regarded by zoologists as prelude to speciation i.e. geographic isolation, behavioural (different flowering time) and genetic (polyploidy) factors. Genetic or other breeding complexities may, in my opinion, be characteristics within such sets as much as they help define them. In that respect I do consider that geographical (and geological) breeding and survival barriers are in evidence and are significant. My perception of the species is then of sets of individuals and populations which suggest morphological and genetic continuity in time and space
The problem of continuity is nothing more than the assessment of similarity and difference. This point needs examination, as it is basic to the question of characterization and classification. The species is the basic biological unit used to identify life forms. In the higher life forms (animals) it seems to be fairly straightforward and what could be problems are easily resolved e.g. the lion species is essentially African, but there is an Asian representative. This is similarly the situation of the elephant species. In one case are they classified as different species, in the other, two. Zoology recognizes and places strong emphasis on the capacity of animal life forms to interbreed and produce fertile offspring. In the lower life forms there are problems because interbreeding capacity does not seem to relate so closely to the life forms and fertile hybrids are produced from improbable parentage. However, it is unlikely that this is the criterion that is actually used to identify “species”. In my opinion reproductive isolation in a species definition has been a byproduct of evolutionary philosophy and that botanists have simply assumed that a species definition for animals also holds for plants.
Continuity is an aspect that I do consider in relation to occurrence with other species and in relation to geography. My observations have been that plants within populations, and thus the populations themselves, show trends. These trends are referred to in botany and zoology as clines and it does happen that the tails or two ends of such a cline may meet and thus produce a situation where a classification decision for one geographic area, may not be correct for another. Then there is the problem of skeletal soils and habitat difference and the effect this has on plant appearances – either generated simply by different soils and/or due to differential selection on the basis of genetic characters. It seems to me simply obvious that sometimes the quite apparent differences between two or more groups of plants is due to local differences in geology and has no general application to the same plants over a greater geographical range. Perhaps it is worth admitting Darwin’s observation to be correct: any local truth or integrity of species is greatly reduced over large areas, and perhaps extend the idea that it is also very greatly reduced by the amount of material examined.
Ideally similarity or difference should be measurable and this is manifested in numerical taxonomy whereby measurements of every conceivable feature are accumulated and processed statistically. It is not possible for me to generate such statistical data in any conventional way and I have to rely on perception derived from learning and experience of what constitutes “difference”. Collecting permits only allow three specimens and it would in any case be ridiculous to suggest that more plants than this should be gathered simply for statistical demands. This learning of mine has been done not only within the limits of Haworthia, but within animal (largely insect) and many other plant genera.
How can a Haworthia species be circumscribed? Description plays a critical role in the examination of similarity. The irony is that if one examines all the descriptions relating to the taxa (species and varieties) that I will cover in this article, it is patently obvious that descriptive detail is practically worthless. Detail barely enables one to describe a single specimen with confidence that it could be identified among others on the basis of its description, and this is even less likely if the growing conditions of that one plant are changed. It has been difficult even within the scope of this one study to photograph plants that are strictly comparable in terms of growing condition. We are also faced with the fact that individual plants within populations are highly variable and that the description of a single plant is far from the description of a population. Furthermore, the description of populations is also compounded by numbers and the difficulty of arriving at a norm by which it would be reasonably possible to identify individuals of any one population of few or many.
Objective? Recognizing the above limitations, this article is intended to examine whether or not H. mirabilis and H. maraisii, as I presented them in my revision (Haworthia Revisited, 1999), can be shown to be separable sets of individuals and populations. I wish to illustrate H. mirabilis and the interplay of this species with H. maraisii as I have experienced it in and around the Riviersonderend Mountains. I conclude that they are but one species and within that decision is nested the problem of what then of related species and where does it end? This is because the whole story can barely be told in one piece as both elements have extended distributions where they unfold other variance and interact with other “species”. Thus other names have also been used in this context and there is a plethora of such names associated with the problem of characterization and classification that need to be understood.
History and background information Although the name H. mirabilis has a long taxonomic history dating from the original description by Haworth in 1804, the name was curiously not associated with field plants until I did so in 1973 (Haworthia Handbook). In the interim several other species (and varieties) were described which originated out of what is obviously a single complex in the greater Bredasdorp/Caledon area. These species included names such as mundula, badia, triebneriana, eleven varieties of triebneriana, and emelyae var. beukmannii. Typification of the name is quite easy if the exact original locality is known. But it is difficult for the nomenclatural type to be typical of any taxon for the reason that in Haworthia, the plants grow in scattered and isolated populations. There is variability and difference of plants both within and between these populations. Often one can be completely certain from which population a plant may have come, and in other cases it is not even certain from which set of populations a plant may have come. Thus it was that the name mirabilis had not been associated with any natural source until I did so, and then I felt constrained to conclude that the population from which it most probably had come was that from which G.G. Smith had drawn his H. mundula. This particular population is very characteristic and individuals can be identified with a very high degree of certainty. The consequence is that we have a situation where the species name is based on plants that are not truly representative of the others in the set. Many of the other variants were described under the name H. triebneriana. Some of these can be identified with a high degree of certainty and others not. The name triebneriana seems to be associated with a population at Stormsvlei (it is certain that von Poellnitz wrongly used the name Strydomsvlei) and there are two other varieties that also had this place as source – they are H. triebneriana vars. depauperata and pulchra. This suggests that the var. triebneriana cannot be identified with certainty. The variety beukmannii was attached to the species H. emelyae, and this also indicates how indeterminate the species were in the absence of reasonable definition and capability.
A similar situation exists with regard to the name maraisii which was a species originally also collected at Stormsvlei. We only know this because it was originally reported in the description as having come from “near Swellendam”, and it was G.J. Payne who showed me the place at Stormsvlei. It was I who extended the use of the name to a wider range of populations, as I will discuss in this paper. I have said elsewhere (Haworthiad 17:83, 2003) that there was an unfortunate decision by W.T. Stearn to typify the name herbacea on the basis of an old illustration (Boerhaave, 1720) that was better associated with the name atrovirens. I unfortunately applied this name herbacea according to the descriptive content of the name (yellow-green in reference to spines) and used it for a species in the Worcester/Robertson area. The name atrovirens, the meaning of this name (dark green referring to leaves), and the accompanying original illustration could perhaps have been better applied to the species which von Poellnitz named H. maraisii and also to H. schuldtiana. The species name H. maraisii thus originates at Stormsvlei and I have used the name for a series of smallish dark-green plants occurring mostly north of the Riviersonderend Mountains in the Robertson area. I did this because I saw a similarity of the plants at Stormsvlei to these Robertson Karoo elements. von Poellnitz did not make this connection and applied the name H. schuldtiana (and of course several varieties) to most of the collections from north of the Riviersonderend just as he applied the name H. triebneriana (plus several varieties) to most of the collections south of the mountains. The problem of certain identification of names is again prominent and the schuldtiana varieties minor and subtuberculata are from the same locality north of McGregor. Again H. schuldtiana was described simply as “Robertson”. The collector was G.J. Payne and he pointed out to me the true origin west of McGregor.
There is a problem in the way the name H. maraisii has been typified and then applied by me. When collected it was reported by W.R.B. Marais to have come from “weathered slate crevices on hills facing the Karoo near Swellendam”. J.R. Brown reported having received a plant from W.E. Armstrong from Stormsvlei but this is perhaps from north of the Pass. G.J. Payne showed me the locality where Marais was purported to have actually collected the plant and I assumed Payne was correct as the plants there agreed closely with the illustration in the Berlin herbarium taken to be the type. This locality is south of the Stormsvlei Pass, and also south of the river, in shale outcrop among what is now a complex of buildings. The site is barely intact but while there were plants there in 1970, I was not been able to locate them on succeeding visits, although I have subsequently done so. In my Handbooks I cited a plant from north of McGregor as “a representative specimen” but indicated that I accepted the illustration in the Berlin herbarium as the type. Breuer, whose typifications I regard as untimely and unfortunate, makes a confusing statement in his first book World of Haworthias Vol.1. Here he cites my selection of the Berlin photograph as lectotypification of the name H. maraisii but cites the locality of my representative specimen from McGregor as the original locality of that species. He also maintained that as the Berlin specimen “is dated from 1937”, it is “therefore no original material”. In his World of Haworthias Vol.2 he has changed his mind about the date of the Berlin illustration and notes it as “undated” and apparently accepts the fact that this is the valid type. However, he uses a plant from his own collection IB5926 from Swellendam to also illustrate the species. The plant IB5926 looks very similar to clones in my own collection MBB6644 (and/or 6860, populations 10 and 11 in the text below) that comes from close to Swellendam and some way from Stormsvlei. The problem for me is that I associate the population at Swellendam (there are two similar populations) with a matrix of populations and individuals (Set A to follow) that I cannot classify with any degree of certainty. Thus Breuer may have unintentionally and inadvertently generated the end-point that I want to achieve with this paper, indicating that maraisii and mirabilis are the same thing by virtue of substituting a mirabilis-like plant for a maraisii-like one. It will transpire that my decision to use the name H. maraisii also for the complex north of the Riviersonderend Mountains viz. H. schuldtiana, is an identical misstep. This is because I revisited Stormsvlei while finalizing this manuscript and fortuitously found plants again very close to the original spot. While these plants do indeed appear to be very different to H. mirabilis as it occurs within the Stormsvlei Pass, it is now quite evident on the basis of experience and re-examination that the difference is due to the geological substrate on which the populations occur. The connection with populations north of the Riviersonderend Mountains, specifically to the one at McGregor, is still true but it means that I have used a name based on an ecotype of mirabilis viz. maraisii, when it would have been more appropriate to have used the name schuldtiana.
Considerations a. The classification of von Poellnitz (1940) The basic difference between the two species as recognized by von Poellnitz can be evaluated from a key which he produced and which was published in Fedde’s Rep.43:102 (1938). The main feature or character of his section Retusae in which he placed the two species was “leaves more or less erect, with minute teeth on the margin, more rarely smooth, at the top bent back sharply and resulting in an end-area which is somewhat translucent, with a few or more green stripes, smooth, tubercled or barely with minute teeth”. In this key von Poellnitz separated H. schuldtiana (with his H. paradoxa) on the basis of the end surfaces of the leaves with small teeth and tubercles. Species with the end-surfaces tubercled only or smooth included H. mirabilis together with H. triebneriana and H. willowmorensis, which I regard as synonymous. Also included here were H. maraisii, H. sublimpidula and H. whitesloaneana. I have explained that the former can be regarded as an ecotype of H. mirabilis. This could confound my use of the name as synonymous with H. schuldtiana where the latter two belong. He also includes in his key H. paradoxa and H. magnifica but these are two elements from east of the Breede River and it will become apparent that I exclude them from this discussion on the basis of geographical consideration only. In my Revision (1999) I have actually regarded H. paradoxa as a variety of H. mirabilis while H. magnifica is upheld as a distinct species despite the obvious affinity with the two species being discussed in this work. Although he also described H. badia in 1938, von Poellnitz did not include this species in his key to the Retusae where it unquestionably belonged.
In total von Poellnitz’ key separates 23 “species”. He published a key to nine varieties of H. triebneriana (two more were added in 1940) in Feddes’ Rep. 47:1 (1939). In the same journal 49:23 (1940) he published a key to 10 varieties of H. schuldtiana that included the former H. whitesloaneana and also what I upheld as H. maculata. My observations are that it is not possible to use the characters von Poellnitz used to separate the varieties as geographic entities by virtue of the inherent variability between plants even within single population.
b. The classification of Bayer (1999) I considered that H. mirabilis and H. maraisii constituted sets on the basis of geographical distribution. Thus in the southwest the plants are larger or more substantial, with erect leaves often spotted on the back or under surfaces, and with fairly conspicuous marginal spines i.e. H. mirabilis. In the northwest the plants are smaller and darker green, the leaf tips tend to retuse and the marginal spines are reduced i.e. H. maraisii. I did recognize a color difference in that H. mirabilis was generally reddish in color with translucence in the leaf end-area, as opposed to the dark-green non-translucence of H. maraisii. In essence, von Poellnitz’ key of 1940 suggests that the two elements I recognize were distinguishable on the basis of “end surfaces with small teeth and tubercles” in his H. schuldtiana (included in my maraisii) and “end surface with tubercles only or smooth” in H. mirabilis AND in H. maraisii (excluded from my broader use of the name). This is only partly true in that while such a tendency might be there, it is not an infallible distinction.
My opinions were simply generalisations derived from a body of populations, one south and one north of the Riviersonderend Mountains. The interplay of these two species, H. mirabilis and H. maraisii, is limited by the fact that the occurrence of mirabilis is at the extreme southwest of the distribution range of the subgenus Haworthia. At its eastern limits there may be some interaction with H. rossouwii in the south, and perhaps even with H. mutica. But otherwise the interaction is with H. maraisii (as I apply the name) and the flow of that species westward and southward with H. heidelbergensis and H. floribunda. It is not my intention to examine that whole complex issue here. What I will show is that across the Riviersonderend Mountains, H. mirabilis becomes inextricably interconnected with H. maraisii and perhaps even with H. maculata. Not only that, it will appear that at Stormsvlei, where H. maraisii, and H. mirabilis(triebneriana) vars. pulchra and depauperata originated, Haworthia maraisii and H. mirabilis can be assumed to be ecotypes of the same one species because of the proximity of the habitats and the synchronization of flowering time and seed set. The only reasonable inference seems to be that the plant differences, which are essentially minimal, are due to the dramatic differences in the skeletal soils and exposed rock they are associated with.
The overriding complexity is that the core of the H. maraisii/mirabilis relationship is expressed in a small series of populations that extends from Heidelberg in the east to Riviersonderend in the west. This includes the original site for H. maraisii at Stormsvlei. It should be borne in mind that I also initially regarded H. maraisii and H. magnifica as one and the same. While I uphold them as separate species in my revision I only do so to facilitate recognition of variants in such an east/west split. I cannot explain the situation presented by populations of plants down the Duiwenhoks river which have to some degree been discussed elsewhere (see Chapter 6). The interplay of H. maraisii with other taxa is a little more complex and I will deal with the issue by examining populations as sets, either in this presentation or separately (I have written a discussion on the H. heidelbergensis/H. magnifica interaction east of Heidelberg also Chapter 6, and also a discussion of the variation and distribution of H. floribunda¸ Chapter 1).
c. Skeletal soils, geology and ecotypification I have shown (Chapter 8, Update 2) that in the E. Cape geology plays a very significant role in the way the species vary and relate to one another. My original observations that this was true were made in the W Cape and Little Karoo and I have no doubt that this is very true for the species now being considered. It is probably true for the Cape winter rainfall biomes (I use the plural only because professional botanical wisdom maintains that there are more than one) that the floras are so species rich because of skeletal soils, geological and topographical diversity and strong rainfall differences (with reliable, consistent and effective winter rain). My study area in this case is dominated by rock formations of the Cape Supergroup although in the north the Karoo Supergroup makes its appearance. The Cape Supergroup comprised three groups viz. Table Mountain Sandstone, Bokkebeld Shales and Witteberg Sandstones and these comprise again about 27 formations. These rocks are invariably directly exposed and soils are often very skeletal. Rock is almost a prerequisite for the presence of Haworthia and it is quite evident to even the most casual observer that geology and geomorphology must in some way impact on the evolutionary processes and pressures within the genus whatever they are. But while geology is no doubt significant, I will only cite it here and there as a rough pointer. This is because although the rocks in the Cape supergroup vary from pure sandstone in the Table Mountain group to mudstones in the Bokkeveld group and again sandstones in the Witteberg group, there are many variants which are obscured at the general map scale of 1:50 000. There is apparently also no general rule that a particular vegetative form will repeat itself on identical rock formation or soil.
Further east in the Riversdale and Heidelberg areas, ferricrete inselbergs are significant Haworthia habitats. As can be seen from my map, the area between Swellendam and Bredasdorp where such inselbergs are abundant, there are almost no records for Haworthia. This could be due to poor exploration. On the other hand H. mutica does occur there and the void may be partly explained by virtue of the occurrences of a different species. To some extent the occurrence of an element which I refer to as H. heidelbergensis is used to fill a similar void in the south and southeast. On the accompanying distribution map, other species occurrences are thus also indicated.
Map 1. Worcester to RobertsonMap 2. Ashton to Swellendam.Map 3. BredasdorpMap 4. E Bredasdorp.
The population sets examined The entire problem is summarized in the listing of six sets of populations (A…F). The location of these is presented on the accompanying map. Because of scale, a mapped site may indicate 2 populations and the sequential population number then omitted. The plants are illustrated and numbered in the same sequence that the populations are listed:-
A. Haworthia maraisii south and east of Riviersonderend Mountains These constitute a geographic set which occupies the southeastern range of the study area. A principle boundary is the Breede River but I have included six populations from further east of which four are off the map. Those four populations will show the integration of the sets considered here, with sets east of Swellendam. The plants are generally dark-green with reduced spination. In this area H. maraisii becomes inextricably linked with what I regard as H. heidelbergensis. That in turn has a complex interplay with H. floribunda in the southeast, with H. turgida in the east and then with H. magnifica still further east. There is a complication too in terms of a few populations along the south bank of the lower Breede River near Malgas and Infanta that is possibly an H. heidelbergensis or a H. floribunda linkage (but see set B). The distribution southwards to Bredasdorp (Rooivlei and Adoonskop) and then westwards to Napier may be in an integrated state with H. floribunda (i.e. neither one species nor the other). At Napier the appearance of the plants still does tend to H. maraisii, so that we have a population that appears to be this discrete species juxtaposed with H. mirabilis var. badia. They have the same flowering time and based on the westward changes in appearance of badia, it may actually be a product of “hybridization” or “introgression” of H. mirabilis and H. maraisii. Thus a ring-cline may exist, and what appears to be two distinct species could be due to the different routes taken in space and time to a point where they grow in close proximity and are interbreeding or have interbred. The southwestern forms of H. maraisii may be a composite of this species with H. mirabilis but also involving H. heidelbergensis and H. floribunda. H. rossouwii could also come into the reckoning, except that this is a spring-flowering species like H. mutica, as opposed to late-summer flowering as is the mirabilis/maraisii complex.
(Note:- the item numbers are used to indicate map position – if space does not permit, a site can be assumed to be in close proximity to the number given in squared brackets).
Stormsvlei MBB7257. This is from the original site shown to me by Payne where I thought the plants had been destroyed. I found about 70 plants on a much degraded site where a wattle tree had just been removed from the top of a small outcrop of Bokkeveld shale. The site is only about 50m away from the point shown to me by Payne but the shale ridge is so degraded that it is unlikely that more plants exist there. The plants were very exposed and burnt by the sun and it was only under the dense shade of a plant of Galenia africanathat I found three dark-green plants that I could comfortably equate with my original opinion that it closely resembled plants from north of McGregor. From the point of view of description and typification, it is important to note from the pictures that the leaf surfaces are with spines and thus do not match von Poellnitz’ description. Not all the plants were with retused leaves either. The species Galenia africana is a vegetation pioneer and an indicator of disturbance.
Stormsvlei MBB6954, JDV96/19. The plants illustrated are from north of the river in quarternary river boulders [1]. In situ they have strongly retused leaves and I use the word “retuse” under the impression that an original meaning of “retuse” was to indicate “bent back like a thumb”. The impression in the literature is that such retused leaves have a distinct end-area. This is not so. The leaves of many species in the southern Cape, including H. herbacea, H. reticulata, and H. rossouwii which are characterized by having rounded rosettes with incurving leaf-tips, are swollen convexly on the inner lower surface. This convexity can be emphasized to the extent that it manifests as an end-area. In addition to this variation, the leaf end may be either incurved or recurved. The original plants of H. maraisii from exposed vertical shale south of the river were strongly retuse and this character was to my knowledge maintained in the plants that were once in my possession at the Karoo garden. Plants from this site north of the river are retused in habitat but not always so in cultivation. Thus they become very similar in appearance to plants in the sandstone ridges within the pass viz. H. mirabilis var depauperata [2]
SW Heidelberg MBB6663, JDV97/35. The plants are very similar to collections from Swellendam (10, 11) and from Rondeheuwel (14) and Verdwaalskloof (15). The habitat geology is Enon conglomerate, as is 10, 11, 14 and 15 are on Bokkeveld shale.
Morning Star MBB7218, MBB7220, Witheuwel MBB7226, Somona MBB7229. It is difficult to clearly associate these two populations with the sets under discussion here. The problem is that while it seems obvious that H. magnifica and H. maraisii are conspecific, the truth is that east of Heidelberg H. magnifica is demonstrably continuous with H. heidelbergensis. I have expressed the opinion that 3 and 4 may also be influenced by H. floribunda.
Andrieskraal MBB7237, MBB7238, Skeiding MBB7239. These three populations are obviously related to H. magnifica and I should mention that H. magnifica var. atrofusca is a problem because its real position taxonomically may be dominant to the typical variety. Thus these Skeiding plants are closer to the var. atrofusca (with obtuse leaf-tips) than they are to the typical var. magnifica (with an end bristle). The geological substrate is ferricrete as at Spitzkop, Heidelberg (6). While I have stated the relationship to H. magnifica, it must be remembered that this species should be seen as an extension of the problem being discussed in this paper. (Spitzkop is also the name for the type locality of H. magnifica var. atrofusca at Riversdale).
Spitzkop (Heidelberg) JDV89/2. This population could also be considered to be maraisii-like but the leaf surfaces are inclined to be finely tubercled without the spine-like surface armature commoner in H. maraisii. But this is also not diagnostic, because there are magnifica-like populations where the individuals are with either finely tubercled surfaces or with very pronounced tubercles and/or surface spination. Descriptive detail is not adequate to make any distinction between the populations 5 and 6 and H. magnifica further east.
Koppies MBB6879. Here the plants show floribunda-like character and this is more evident in the next (8). H. rossouwii is present nearby.
S Oudekraal MBB6881. H. rossouwii is also present about 1km to the north. These plants are even more floribunda-like and I have included some of these populations in a resume of H. floribunda.
Stuurmanskraal J.Esterhuizen in JDV93/64. This population is geographically nearer to the Breede River and to Malgas and the plants have a similarity to that south and west of Malgas.
SW Swellendam MBB6644, MBB6860. The plants tend to have erect or sub-erect leaves. The leaf ends are generally not retused and the tips are often slender and floribunda-like.
W Swellendam MBB6861. These Swellendam populations mirror that at Heidelberg (3) and the plant form repeats at Rondeheuwel (14) and at Verdwaalskloof (15) to the west. I have always regarded the Rondeheuwel population as indeterminate between H. mirabilis and H. maraisii and considered that it might be hybrid. However, I have with time become increasingly skeptical that hybridization accounts for problem variability. My impression is that the sequence from Heidelberg through Swellendam to Rondeheuwel and Verdwaalskloof support the contention that there is genetic continuity that is not a product of hybridization. The similarity suggests to me that this may be archetypal for the populations in the five sets that I am dealing with here.
12. Witkop/Uitvlugt MBB7242, MBB7243. These two populations are on ferricrete inselbergs that are a conspicuous feature of the landscape between Stormsvlei and the Potberg in the southeast. Together with the next two populations the plants link Stormsvlei to the Potberg mélange that I think may relate to H. floribunda as well as to H. heidelbergensis as I already have alluded.
Die Kop MBB7244, MBB7245. (Not Die Kop Wydgelee MBB7500) These are south of the two former collections and are in the same mould. I treat them separately because the one collection is on a white-clay pressure burst and the plants are less scabrid than in the other collection that is on reddish pebbly soil. Both sites are on ferricrete.
Rondeheuwel MBB6862, JDV90/32, JDV96/43. My first impression of this population about 1972 was that it was hybrid between H. mirabilis and H. maraisii. Yet even then I was expecting some king of continuity between these two “species” other than hybridization.
Verdwaalskloof MBB7074. This site is near to Riviersonderend and is one of the strongest links between the series in Set C that could perhaps comprise a separate H. mirabilis.
Napky MBB7030, JDV96/18. This is near the origin of the von Poellnitz H. maraisii var. simplicior. It is now south and west of the Breede River and there is an element of floribunda in the plants viz. a more elongated leaf with flattening and rounding of the leaf-tip and the slight twist evident in that species (see 7 & 8, and also true for 3, 9 and 10). It should be noted that in my treatment of H. floribunda (Umdaus Press, in ms) I suggest that this may be a juvenile character perpetuated in that species.
Tarentaal MBB 6539. One of the few populations that transgresses the tendency to a geographic drift in similarity of populations. (Some of…)The plants in this particular population are very similar to robust forms of H. magnifica var. atrofusca as at Riversdale.
Witklipkop MBB6890. This and the following four populations are problematic and further populations occurring along the lower Breede River exacerbate the situation. They need to be included here but at the same time they are clearly connected to a different set of populations that I relate to H. heidelbergensis. The leaves are smaller, more numerous, more elongate and generally without an end-area.
Brakfontein MBB6886.
Juliesfontein MBB6882. In my Revision I cite an early collection of mine MBB1211 from here under H. maraisii. I also included a collection of P.V. Bruyns from Ziekenhuis (see 24) because the plants were very small and seemed similar. There are good herbarium specimens to show this. However, when I re-collected at Juliesfontein after the publication of my Revision in 1999, there were no plants at my remembered site and much larger plants nearby which I would have had difficulty putting with H. maraisii. The vast mass of material I have seen since has not made a taxonomic decision any easier. I can honestly say that recognizing a taxonomic difference between these two populations (20 and 24) would serve a very limited purpose.
N Potberg MBB6889 [20]. Chris Burgers (CB2506) first sampled this population and I also cited it under H. maraisii. This is not a comfortable solution.
NW Potberg MBB6544, JDV96/54 [20]. H. variegata occurs here too and may infuse populations in the area. I have one record of plants that seem to suggest this, but this would need to be covered in a discussion of H. variegata itself.
Jakobsrivier JDV92/85, JDV87/50. Needs to be considered with the following. They are very small plants with blunt and rounded leaf-tips suggestive of the Potberg collections 18 –22.
Ziekenhuis P.V. Bruyns in JDV87/5, Uitrus MBB7250. The first is a collection that P.V Bruyns made east of the river. These are very small plants with erect leaves much as I remembered the original Juliesfontein collection of mine. They flower in January as this whole complex generally does. The geology of the east bank of the river is Bokkeveld shale. At Uitrus it is rather uncertain as while there are exposed shales in the vicinity they are overlaid by pebbly deposits of recent origin. This may be very important in relation to the juxtaposition of this site (and 22) with Set B.
Rooivlei MBB6638. The following five populations are a continuance of the Napky (16) one where there seems to be a floribunda impact and the plants are also very dark green often conspicuously tubercled. The leaves are thick and blunt. An element also occurs here which is apparently H. heidelbergensis, and H. turgida is also present. The presence of a separate heidelbergensis-like population is confounding. Those plants flower a little earlier and should be discussed with plants from Kathoek, Beyersdal, Haarwegskloof. They cannot be excluded from a total concept of H. mirabilis that I am suggesting here, and as I am going to do for the same reason that this conspectus excludes populations from beyond Heidelberg.
Adoonskop MBB6640. These are like the Adoonskop and Napier variants (27) and easily put with a concept of H. maraisii if it were feasible to separate such an element from H. mirabilis in the overall relationship of the populations. The plants tend to have rounded leaf-tips as is the case in the following four populations.
NE Bredasdorp JDVsn – no specimens collected. Plants as above. This would have been an important record because of the proximity to Bredasdorp and to H. mirabilis as the var. sublineata. The site was also home to Aloe brevifolia and sadly the site is so degraded that neither Haworthia nor Aloe can now be found there. MBB7334 is a very recent collection, nearer o Bredasdorp and plants are very similar to (29)
Napier MBB7259, JDV85/15. There is a substrate difference in that at this site the soil is pebbly quartzitic whereas at the following site nearby (29) it is finely slabbed, coarse grained sandstone. MBB7259 is about 4km north and the substrate there is true micaceous shale.
Napier JDV85/96, MBB7247 [28]. Whereas plants at the former sites are blackish-green, here the plants are reddish-tinged.
A01 MBB7257.1 StormsvleiA02 JDV96-19.1 N StormsvleiA03 JDV97-35.1 SW HeidelbergA03 MBB6663.1 SW HeidelbergA04 MBB7218.1 Morning StarA04 MBB7220.1 DuiwenhoksA04 MBB7226.1 WitheuwelA04 MBB7229.1 SomonaA05 MBB7237.1 AndrieskraalA05 MBB7239 SkeidingA06 JDV89-2.2 Spitzkop, HeidelbergA07 MBB6879.1 KoppiesA08 MBB6881.1 S OudekraalA09 JDV93-64.1 StuurmanskraalA10 MBB6644.1 SW SwellendamA12 MBB7242.1 WitkopA12 MBB7243.1 UitvlugtA13 MBB7244.1 SE Die KopA14 JDV90-32.1 RondeheuwelA14 MBB6862.1 RondeheuwelA15 MBB7074.2 VerdwaalkloofA16 JDV96-18.1 NapkyA16 MBB7030.1 NapkyA17 MBB6539.1 TarentaalA18 MBB6890.1 WitklipkopA19 MBB6886.1 BrakfonteinA20 MBB6882.1 JuliesfonteinA21 MBB6889.1 N PotbergA22 JDV96-54.1 NW PotbergA22 MBB6544.1 NW PotbergA23 JDV92-85.1 JakobsriverA24 JDV87-5.1 ZiekenhuisA24 MBB7250.1 UitrusA25 MBB6638.1 RooivleiA27 MBB7334.2 H. maraisii. NE Bredasdorp.A28 JDV85-15.1 N NapierA28 MBB7259.1 Napier StationA29 JDV85-96.1 NE NapierA29 MBB7247.1 H. maraisii. NE Napier.
B. Haworthia mirabilis towards the Potberg This set is actually anomalous and consists of only three populations associated with Table Mountain Sandstone and calcareous limestone. At Karsriver, a short distance from 27 is where one finds H. rossouwii var. petrophila (which I had previously allied to H. variegata) in limestone. Further east is H. variegata var modesta and to the south of that H. rossouwii var. calcarea (previously as a variety of H. mirabilis). It is obvious that the classification is difficult and dubious. However, C. Burgers made another surprising collection (CB2018) from Buffelsfontein [CB] south of the Potberg. I cited it in my revision under H. mirabilis var. calcarea. I have had a problem with a specimen collected by Prof. H. Compton cited “from the Potberg sandy flats”. That specimen is of a retusa-like plant that I thought might perhaps be representing either H. mutica or H. mirabilis but I did not cite the specimen in my revision because I was so doubtful about it. I thought this could have been what Burgers found, but his collection was in rock crevices and not on the sandy flats. However, Adam Harrower made a collection (ADH594) from 20km further east, also in rock crevices and clearly the same as the Burger collection. On a very recent trip my wife and I found a similar plant (31 MBB7248) on the northeast end of the Potberg that I think may fit here. The problem is its proximity to 24 only 3km away.
Sandhoogte ADH594, MBB7251. The site is a most unprepossessing and unlikely looking one from a distance. The vegetation is dense waist-high fynbos, and Adam Harrower found the plants in the rocky north-facing border to a stream. They can be compared with H. turgida var. longibracteata (except they are less proliferous) and also with H. mirabilis var. sublineata. There is also a likeness to forms of H. rossouwii var. calcarea, albeit distant. More striking is a curious resemblance to H. magnifica var. splendens from near the Gouritz River Bridge.
Ballyfar MBB7248, MBB7249 [off map E of 22]. These two populations are only a few kilometers from 23 and 24, but they are on sandstone. What is more is that the plants are larger but have fewer leaves that are strongly retused as in the Compton specimen. What is striking about the plants is the presence of numerous surface spines and one is forced to consider the resemblance to a population of H. magnifica east of Riversdale with similar spination and general appearance, only proliferous. The site is about 3km due east of the Sandhoogte population and this suggests that the two populations are of the same species. There is an interesting dimension to the problem mirrored in the preceding (30). This is the occurrence of H. turgida at Diepkloof only 15km to the northwest and only five kilometers from Burgers 2018 at Buffelsfontein across the Potberg Mountain. A decisive factor may be flowering time, because H. turgida flowers in spring and the other populations considered here flower in mid-summer. But the main point is that the relationships are complex and varied.
B30 MBB7251.1 Sandhoogte De HoopB31 MBB7248.3 Ballyfar
C. Haworthia mirabilis and its variation south of the Riviersonderend Mountains This set also starts with Stormsvlei and is the south and westward set which is almost exclusively mirabilis. There is a possibility of a connection across the Riviersonderend Mountains closer to Villiersdorp where it may be suggested that H. maculata is implicated too, but this will instead be considered in Set 3. These are plants with a tendency to redden. They are usually larger than those in set 1. The height may be as much as 100cm and rosette diameter 110mm.
Stormsvlei Pass MBB6955. This population is in sandstone and immediately there is a significant difference to the plants immediately south (1) and north (2) of the Riviersonderend River a short distance to the south. There is no evidence of an end-area to the leaves. The leaves are longer and more slender and they show the redder colour one associates with H. mirabilis elsewhere. Some of the plants are very highly spotted on the back or under surfaces. This is a character that has been associated with H. mirabilis when in fact quite common to find plants with or without such marking in many of the populations in these sets.
E Stormsvlei Pass E. Aslander sn. MBB7252, MBB7253 and MBB7254. In exploring the southern slopes of the Bromberg I came across several populations. The geology here is rather complex as it is the close interface of sandstone and shale, with the pebbly patches associated with ferricrete (Gibbaeum austricolum is present). At one site (MBB7254) the plants were small and with narrow elongate leaves on a coarse-grained sandstone ridge, while immediately north on a pebbly band there were plants with fewer, shorter and broader leaves. There is no difficulty in allying these plants with 32.
Greyton JDV96/47. The continuity westwards is disrupted by lack of records and there does not seem to be suitable habitat between the Riviersonderend River and the mountains. A population at Lindashof recorded by G.G. Smith appears to have been destroyed in a road-making operation. This population at Greyton is of rather small plants with conspicuously finer and intense spination.
Uitkyk MBB7092, JDV95/97. This is in the extreme west for the set, at Elandskloof west of Genadendal. It is a very rocky south-facing site and the plants are robust and proliferous.
Leeukop JDV90/34, JDV85/98. This is the locality for the var. rubrodentata where the plants have long slender leaves with intensely red spines – hence the name.
Genadendal JDV90/33.
Skuitsberg JDV88/70, JDV96/101. Here there is quite an extensive population with a similar one also present on the adjoining farm Nethercourt. The plants are generally very robust and were described originally as H. emelyae var. beukmannii.
Jongensklip MBB7059. The interesting thing about this population is that it fills somewhat of a void between populations known respectively north and south of the Caledon/Bredasdorp wheatlands.
Goudini MBB6537. This population is the southwestern-most for the species. I have grown it from seed and the variability among the seedlings is quite striking, which confirms my opinion that the taxa described from north of Napier are not significant as discrete varieties.
Schietpad MBB4642. JDV94/90. See 43.
Fairfield MBB6643. See 43, except that the plants are very proliferous.
Mierkraal (Napier) MBB7091, S Mierkraal MBB7260. The plants now constitute a general form that appears in the preceding three populations and there are several such populations in the area between Mierkraal and Fairfield (42) and I do know that I have seen it in the Klipdale area to the northeast. The varieties H. triebneriana vars. multituberculata, napierensis (Schietpad, 41) and turgida, originated north of Napier and yet I would not consider the variability to be population based nor even so significant in terms of the variability of H. mirabilis var. triebneriana as I would designate the taxon. MBB7259 is a population south of Mierkraal, the plants tend to have more elongate and erect leaves. The site is geographically nearer to 40 (MBB6987) which is H. mirabilis var. badia.
NW Napier MBB6635. Although the geology is the Bokkeveld group, the rock here is very quartzitic and the vegetation alters from Cape Fynos to renosterveld over a very short distance. The site is that of H. mirabilis var. badia. The geology in this area is quite complex and there is also a small ferricrete inselberg to the east.
W Napier MBB6987. This population is a western continuation of the previous one and is also the var badia. However, there is a tendency for the leaves to be more erect and narrower. I do speculate that this var badia may have arisen from an interaction of H. mirabilis and H. maraisii. The four populations (28 & 29 of set A, 44 & 45) flower and seed at the same time, seedlings of both are highly variable and both have companion populations which suggest the deduction of continuity across a very strong geological but weak breeding barrier.
Bredasdorp N JDV86/8, MBBsn, MBB7261sn. This locality is immediately north of the town on the riverbank. The area was highly invaded by alien Acacia and the haworthia are no longer there. The plants are somewhat similar to those south of the town, but more robust and with more strongly retused leaves. This is a close approach to H. maraisii (26) and may parallel the situation at Napier (28 & 29) where this species and H. mirabilis var. badia (44 & 45) are almost sympatric. My MBB7261 are a few plants rescues from about 300m northwest of JDV86/8 from a site overgrown with bluegum. These plants had erect leaves.
Bredasdorp S MBB6639. The site is a north-facing sandstone mountain slope. The plants are highly variable and are named as H. mirabilis var. sublineata [47]. The leaves are long and slender. Some clones are proliferous and others not.
Mierkraal (Bredasdorp) MBB7090. H. mundula was described from this locality which is unusual in being recent sandstone and on the very southern fringe of the distribution of H. mirabilis. The plants are very proliferous and their leaves have a very pronounced end-area that is also smooth. The population is thus fairly different from others and this is unfortunate because it is in my opinion the source of the type and hence is the var. mirabilis. I first saw the population in 1969 and can testify to significant changes in the precise whereabouts of large proliferous mats in the past, as opposed to more scattered less proliferous clumps in the present.
C32 MBB6955.1 Stormsvlei PassC33 MBB7252.1 NoukloofC33 MBB7254.1 E StormsvleiC34 JDV96-47.1 S GreytonC35 JDV97-94.1 UitkykC35 MBB7092.1 UitkykC36 JDV85-98.1 S GreytonC37 JDV90-33.1 W GenadendalC38 JDV88-70 SkuitsbergC38 JDV96-101 SkuitsbergC39 MBB7059.1 JongensklipC40 MBB6537.1 GoudinieC41 JDV94-90 SchietpadC41 MBB4642 SchietpadC42 MBB6643.1 FairfieldC43 MBB7091.1 Mierkraal, NapierC44 MBB6635.5 NW NapierC45 MBB6987.1 W NapierC46 JDV86-8.1 N BredasdorpC46 MBB7261.1 NW BredasdorpC47 MBB6639.1 S BredasdorpC48 MBB7090.1 Mierkraal, Bredasdorp
D. Populations north of the Riviersonderend Mountains Stormsvlei is where the Riviersonderend Mountains begin to tail away to the lowland wheatfields. The Riversonderend River cuts through the mountain about 10km east of Stormsvlei, and the Stormsvlei Pass itself is a rift through the mountain tail that allows road access from north to south. The mountain to the west is Table Mountain Sandstone. Rainfall is high and the vegetation is Cape fynbos. It must be a formidable barrier to genetic flow and unsuitable habitat for succulent plants. There are very strong geological gradients coupled with precipitation differences north and south of the mountain. The geological gradients also have an east-west pattern as opposed to the essentially north-south pattern south of the mountain. Further to the west, folding and faulting have resulted in a huge wedge of more recent formations appearing between Worcester and Villiersdorp. McGregor is virtually at the southern end of a wedge of older Bokkeveld formations that run northwestward to Robertson. There it meets the great Worcester fault line and a massive igneous pluton. Rocks of the older Malmesbury Supergroup and rocks of the much younger Karoo Supergroup are juxtaposed with igneous intrusive rock. The plants dance to this pattern and this is reflected in the differences found in Haworthia populations. This set comprises the populations on the Riviersonderend Mountains itself and along the northern foothills. The plants are intermediate between sets C and D and include significant ecotypes, as for example the Boesmanspad plants that resemble plants in the H. turgida complex, the Moddergat plants associated with H. maculata and the Trappieskraalkloof plants which are the maraisii-like element.
NE Stormsvlei Pass (Brakfontein) MBB7087. This is on a dry north-facing slope where the vegetation is Fynbos and close to the transition to Renosterbos. The geology seems to be recent scree and sand. The plants have erect to sub-erect leaves, are quite narrow and have strong marginal spination. They thus seem to equate with plants within the Pass (32) in quartzitic sandstone of the Table Mountain Group and seem to differ from plants close-by to the northwest (67, MBB7086) that are on a shale/sandstone formation of the Bokkeveld Group.
Boesmansrivier MBB6711. The Boesmansrivier Valley is a very long and deep incision into the mountain running southwest to northeast. The plants are similar to Stormsvlei Pass (32) but the undersurfaces are often more strongly spotted and the faces of the leaves are frequently also very attractively spotted with translucent dots.
E Olifantsdoornkloof MBB6686. The habitat here is slightly drier and more exposed than the preceding and the plants have shorter squatter leaves.
Olifantsdoornkloof MBB6685 [51]. At this site there is again a deep incision into the mountain and the habitat is more sheltered and moister. The plants have longer leaves. von Poellnitz named H. triebeneriana var. diversicolor from this population and the name refers to his observation “plants therefore appear diseased and no plant agrees fully in colour with the other”. It is instructive to think what he would have thought if he had seen plants from the preceding populations extending through to the following 15 in this set.
Hoeksrivier MBB6575. What is surprising and different about this population, also in a rift into the mountain, is that the plants have the rosette shape of H. herbacea. Although the leaves have the swollen mid-section, the ends are curved inward so that the rosette is more rounded. These plants are particularly “diseased” as in 52.
Houtbaaiskloof MBB6760, JDV97/163, MBB7241. Houtbaaiskloof is a very deep incision into the mountain immediately south of McGregor, which meets a geological fault line north of the Hoekrivier. So I was able to make collections both north and south in this pseudo-gorge. This is where von Poellnitz’ H. whitesloaneana originated. The leaves revert to the tendency to have an end-area but the “diseased” appearance is reminiscent of 52.
Takkap MBB6723, JDV89/63. In this incision of the Takkap River into the mountain the plants become increasingly mottled and also slightly softer in texture coupled with increasing leaf numbers although they are still small in size.
Dwarswaterkloof MBB6865, JDV89/67. As with the Takkap plants (55) the plants are greener and quite proliferous. The Dwarwaterkloof is again a very long and deep incision into the higher peaks of the Riviersonderend Mountains. It is not surprising that the plants have continuity with the following populations.
Boesmanspad JDV84/67, JDV88/63, JDV89/67. The Boesmanspad refers to a footpath that traverses the Riviersonderend from McGregor to Greyton. There is a deep valley called Boesmanskloof that runs from Greyton in the south towards McGregor. It is a high rainfall area and it is quite surprising to find Haworthia in this gorge. Furthermore the plants are small and very turgid so that the comparison can be fairly drawn with that of plants of H. turgida var. turgida from similar wet and rocky situations in the Langeberg mountains e.g. Southey’s Pass or Garcia near Riversdale. The flowering time and flower itself in the context of distribution and flow in similarity suggests that this must be an ecotype related to populations both north and south of the mountain range.
Koeniesriver MBB6714 [56]. These are in a more exposed situation than the previous two and with the following (59) are again more reminiscent of plants from the Stormsvlei Pass itself (32).
Rietvleikloof MBB6720. The topography of this mountain incision is interesting because it is practically the last of a north/south series. The next incision is a major faultline that runs southwest/northeast. This forms a valley largely in the lee of high mountain (Jonaskop).
Moddergat JDV96/34, MBB7270. This population needs to be carefully considered. It oversteps the distribution of H. herbacea and the var. livida that I described from just east at Lemoenpoort. I also described H. herbacea var. lupula from an improbable habitat deep in the mountains east of Villiersdorp. I associate the Moddergat plants with H. maculata to the north at Brandvlei and this has still further associations in the Langeberg Mountains. However, the flowering time and appearance of the plants do suggest that a connection with H. mirabilis has to be considered even if improbable. I have indicated already that this area is geologically diverse.
Trappieskraalkloof MBB1210, Droeriviersberge MBB7263, MBB7264, Haumanskloof MBB7265. The first I considered from memory to be a very significant population that could better be placed in Set E. The plants are small, greyish in colour. The rosette is somewhat globular with short retused end-areas and very similar to 65. I initially made this collection MBB1210 in my first few years at Worcester, and it is only at the close of this work that I have revisited the area. The area is difficult to access and is very dry. The terrain is extremely rocky and rugged. Dywka Tillite interfaces with Witteberg Quartzite and the vegetation can change dramatically over a distance of a few meters. It is not clear how the latter three populations relate to the traditional perceptions of H. maraisii and H. mirabilis as discrete species, and nor to their relation with H. maculata, which is not far to the west at Moddergat (see 60 JDV96/34. MBB7270). It is evident that they are connective between sets D and E.
Whipstock MBB6864. Again one has to take geology into account as I discussed for the set. At Moddergat and Trappieskraalkloof one is confronted with more recent geological phenomena. Whipstock is at the southern end of a similar quartzitic formation and the plants follow the trend indicated in the early items of this set. The plants are somewhat mottled and an end-area is often apparent.
Klipbergdam MBB6681, MBB6682, JDV97/48. These plants demonstrate continuity between the early members of this set and the following set.
W McGregor MBB6673, JDV96/49. This is in the McGregor Nature Reserve and is the source of the original H. schuldtiana as was conveyed to me by G.J. Payne.
N McGregor MBB6646 [64]. This population is the origin of the names H. schuldtiana vars. minor, subtuberculata and unilineata. Obviously the population seems to be unusually variable because of the varieties that it has given rise to. But this is not so and simply reflects the weak taxonomic precepts of the time which seem to be so difficult to dispel in the present.
SE McGregor MBB6687 [64]. The plant I illustrate appears to have very pronounced end-areas to the leaves. But I have not explored the area thoroughly as the plant fits conformably with the populations I am listing.
NW Stormsvlei Pass MBB7086, & Vaandrigdrift MBB6534, JDV96/25. I have often observed plants northwest of Stormsvlei Pass and simply accepted that they belonged to the discrete species I had conceptualized viz H. maraisii. They now easily fit the conceptual model that the elements around Stormsvlei and the Pass are actually one and the same species. I have not collected at the latter two sites further to the east. Plants I observed there would have fitted those illustrated in 61, 64, 65 and 66 as well as with many in set E. This would correspond to a concept of the element H. maraisii as I recognized it in my Revision.
D49 MBB7087.1 NE StormsvleiD50 MBB6711.1 BoesmansrivierD51 MBB6686.1 E OlifantsdoornkloofD52 MBB6685.1 OlifantsdoornkloofD53 MBB6575.2 HoeksrivierD54 JDV97-153.1 HoutbaaiskloofD54 MBB7241.1 N HoutbaaiskloofD55 MBB6723.3 TakkapD56 MBB6865.1 DwarswaterD57 JDV84-67.1 BoesmanspadD58 MBB6714.1 KoeniesriverD59 MBB6720.1 RietvleikloofD60 JDV96-34.1 ModdergatD60 MBB7270.1 ModdergatD61 MBB7263.1 DroerivierbergD61 MBB7266.2 DroerivierbergD62 MBB6864.1 WhipstockD63 JDV97-38.1 KlipbergdamD64 JDV97-49.1 W McGregorD65 MBB6646.1 N McGregorD66 MBB6687.1 SE McGregorD67 MBB7086.1 N Stormsvlei Pass
E. Haworthia maraisii north of Riviersonderend Mountains This is the Worcester/Robertson Karoo set that I have simplified by excluding H. maraisii var. notabilis. The reason I have done so is because of a manuscript of mine dealing with the epithet intermedia which is with the Editor of Haworthiad. That manuscript deals with the impact of the granitic pluton at Robertson and associated dolomite on Haworthia although I have there not really discussed the geology as such. At Klaasvoogds where there is also a dramatic shift from ancient to very recent rocks and replication of the problem of a var. notabilis that is not dealt with in this paper. This set illustrates the populations that I used to formulate the concept of a species H. maraisii. It also includes the “escape” into the Little Karoo and the possible eventual link with H. emelyae east of Barrydale. The plants are characteristically very dark coloured and perhaps smaller in comparison to sets A, B and C. However, I think I can demonstrate reversion to a mirabilis-like plant and also similarity to H. heidelbergensis. I also include two populations that suggest a more far-reaching link to H. emelyae north and east of Garcia Pass, north of Riversdale.
Bosfontein JDV97/58. These are plants in a continuum from Trappieskraalkloof (61) on the same geological formation viz. Witteberg Group rocks.
Dublin F.J. Stayner in KG400/61, JDV94/18. The significance of these plants is that they were particularly dark-green, very tuberculate and the leaves were erect. Although I saw plants at the site with Frank Stayner I have not been able to find them at the site since. Plants do occur again a short distance to the east, but they do not have the same very erect and narrow leaves of Stayner’s collection. A plant is illustrated in my revision where it is under the number JDV94/18 and attributed to Roy Littlewood who was instrumental in first drawing attention to it.
Rooiberg MBB6956. This is north of the Breede River from the previous locality listed. The Rooiberg is quite a substantial massif and houses what can be regarded as discrete populations. Included in these is the very curious H. herbacea var. flaccida. I only observed five plants of that and suspect that it is of hybrid origin with the putative parents being H. maraisii and H. reticulata rather than H. herbacea.
Agtervink MBB6647. The following eight populations illustrate the geographic core of my concept of an H. maraisii.
N Agtervink MBB7176 [72].
Sandberg MBB7174, JDV98/18.
Nekkies JDV85/89, JDV96/24, EA083, EA1408. This population is very close to the Robertson pluton and I did associate this site with transition to the var. notabilis. I doubt if that this is actually true for this local area. Etwin Aslander made a collection (EA1408) from very close to Wolfkloof where the notabilis complex occurs, and this maraisii variant could be compared to those in 80 north of Ashton and even with 32 in Stormsvlei Pass.
S Sandberg MBB6670, JDV97/38.
Koningsriver MBB6672.
Vrolikheid MBB6968, W Kanonkop MBB6967, E Witkranz MBB7077. These and the next three populations are to the west of an area that should contain plants of the interface I would like to illustrate. However, it is a very dry area and not visitor friendly.
Steenbokshoogte MBB6966, Klein Spitzkop MBB6969.
Grootrivier MBB6677.
Breede River Bridge (Robertson) MBB6648. I noted this population in my Revision with the comment that note can be made of forms near Robertson which are rather similar to H. pubescens. The plants tend to have the same globular rosette shape and also the pubescence. However, the flower and flowering time is typical of H. maraisii as I defined it.
Muiskraalkop MBB6961, JDV87/208. This is the type locality for von Poellnitz’ H. schuldtiana var. robertsonensis as indicated to me by Payne. However, von Poellnitz does cite more than one locality for each in his description. The plants do have more slender erect leaves than most in this set but the distinction gets lost the more material that is considered.
Rietvlei MBB7055. This population is an odd one that I used in developing a justification for treating the element notabilis as a variant of H. maraisii. I was looking at the plants on a south slope under bush whereas it transpires that the bulk of the population is higher on the ridge in Enon conglomerate. Plants further north on Peninsula sandstone (Kranskop) are quite different and this difference compounds to the northwest where the plants are on Malmesbury shale.
Ashton MBB6668. This site is also Enon conglomerate and the plants are quite large and the leaves are rather recurved than with an end-area. The surfaces also tend to be smooth. Hence they do appear somewhat mirabilis-like cf. 32.
Cogmanskloof MBB6875 . These plants can also have leaves quite strongly recurved and there is a close similarity with the following.
Baden, Montagu JDV97/55, EA068.
E Montagu PVB7588. Malherbe of Sheilam stated that he had seen “small black plants” in the Koo valley north of the Langeberg. I have not been able to confirm this other than seeing them immediately northwest of Montagu. P.V. Bruyns recorded the species about 20km to the northeast (Dobbelaarskloof) and also at this site due east of Montagu. The plants have highly retused leaves but do not deviate substantially from others in this set.
Barrydale MBB6667, JDV97/39, MBB7214. J. Dekenah knew these plants at Barrydale and it is surprising that some varietal name or other did not evidence this. It is difficult to interpret outside of these sets and a complication is a very odd population south of Southey’s pass (MBB6666) that I have perforce regarded as H. magnifica var. magnifica. It is an embarrassment to me because it has resemblances to H. retusa and to H. mirabilis as much as to H. magnifica. In fact plants from there do not closely resemble H. magnifica in the form that it is manifest in 5 and 6 of set A. It (87) is important because of the following item.
Kleindoringrivier G. Marx in MBB6741. This population is midway between Barrydale and Muiskraal where H. emelyae var. major occurs, and it appears to be a connecting link. The flower and flowering time links it to the west and to these sets.
Loerkop MBB6979. This is southeast of Ashton and the plants have retused end-areas.
Viljoensdrift MBB6518. This is at the foot of the massif Elandsberg and seems to be in the continuum with 74, 75 and 76.
Middelplaas MBB7079. This population is similarly in some kind of juxtaposition between this set and the next and I interpret it like that in view of unlisted populations I have seen in this area.
E71 MBB6647.1 AgtervinkE72 MBB7176.1 N AgtervinkE73 JDV98-18 SandbergE73 MBB7174 SandbergE74 EA083.1 TierbergE74 EA1408.1 NW RobertsonE74 JDV85-89.1 Die NekkiesE74 JDV96-24.1 Die NekkiesE75 JDV97-38.1 SW SandbergE76 MBB6672.1 KoningsrivierE77 MBB6967.1 W KanonkopE77 MBB6968.1 VrolikheidE77 MBB7077.1 E WitkranzE78 MBB6966.1 SteenbokshoogteE79 MBB6677.1 GrootrivierE80 MBB6648.1 Breede River BridgeE81 JDV87-208.1 MuiskraalkopE81 MBB6961.1 MuiskraalkopE82 MBB7055.1 RietvleiE83 MBB6883.1 N AshtonE84 MBB6875.1 CogmanskloofE85 EA068.1 MontaguE86 PVB7588.1 E MontaguE87 JDV97-32.1 BarrydaleE88 MBB6741.1 KleindoringrivierE89 MBB6979.1 LoerkopE90 MBB6518.1 ViljoensdriftE91 MBB7079.1 Middelplaas
F. The interplay of Haworthia maraisii and H. heidelbergensis This set includes the var. meiringii at Bonnievale and a set of populations that suggest a closure with Set A and the emergence again of H. heidelbergensis. Many of the collections listed here seem to be entirely new to the literature of the genus. The tendency in this group, as a whole, is to erect leaves. They are also small with a maximum height of 50-70cm and a rosette diameter of 20-40cm. The geology of the area does not seem to be excessively complex or vary substantially from the previous set except that the topography is more uniform. However, the plants seem unable to hold the profile of the previous set and there is a drift to H. heidelbergensis.
W Goudmyn MBB6516. I start this series in the mid-west of the area because it has the more conventional forms of H. maraisii to both the north and the south. This population is west of Olifantskrans and extends on a long sharp ridge to the summit of the Elandsberg to the west. The plants are very small and have many small slender erect leaves.
Olifantskranz, E Goudmyn MBB6676, MBB7081. I have four collections from this general area and the plants are continuous with the previous set. The leaves are a little more substantial and often very spotted. The continuity extends both east- and northeastwards.
Olive Grove/Wakkerstroom MBB7081, MBB7084. This is the eastward extension and the plants are approaching H. maraisii var. meiringii at 95. The leaves are slender and erect and relatively opaque.
W Bonnievale MBB6509. There is also a record from north of this point which are less spinose and translucent than this particular population although the distance separating them in less than 100m. Here specifically, I find plants which I would compare favourably with H. heidelbergensis var. toonensis at Matjestoon east of Swellendam. The continuity is in fact with H. maraisii var. meiringii below.
E Bonnievale MBB7085, Edendale MBB7269. These plants are very proliferous and spinose. The rosettes are globular. The var. meiringii was actually based on the population to the near east of this one (MBB7269) where the vegetative similarity to H. herbacea was even more significant.
Bobbejaanskloof MBB7082, N Goudmyn MBB7083. Northeast of Olifantskranz (93) the continuity extends, not as expected to the conventional H. maraisii as at Loerkop, but to plants here with more slender erect leaves suggesting H. heidelbergensis.
Zandvliet MBB6964, MBB6965. Here there are two distinct populations that I include in the var meiringii together with the previous 6 populations. Plants do vary from individuals with stubby nearly terete leaves to plants with the conventional leaved wider than thick and narrowing to the apex.
SE Loerkop MBB6981. This is a very substantial population and while it would fit in the mélange of the previous item, it starts to relate uncomfortably to the next.
Tonteldoos MBB6974, MBB6977. These and the following to 111 are a nondescript assemblage of plants that do not relate comfortably with any of the other sets and neither do they collectively suggest any kind of taxonomic unity.
Mardouw MBB6990. Bob Kent also has two collections from between this and the following sites.
Nooitgedacht MBB6988, MBB6989 [102]. The first population is also a very substantial one close to eucalypts as is the following. However, the second is in natural vegetation.
Langverwacht MBB6992. This is a remarkable population of small very proliferous plants with quite turgid stumpy leaves. The site is notable as a quartzitic inselberg marked by the occurrence of several large gum trees. The haworthia seem to be the only plants tolerating this alien vegetation as the ground is otherwise devoid of plant growth.
Jakkalskop MBB6971.
Witkop (Drew) MBB7036.
NW Sanddrift MBB7056, W Sanddrift MBB7057.
Sanddrift MBB6625, JDV89/8, JDV90/31. Although very similar to the next, this population tends to have leaves with a retuse and opaque end-area. It is also small and very proliferous but nevertheless suggests an affinity with H. maraisii as in Set D.
N Sanddrift MBB6626, JDV87/193. This particular population comprises small plants with spreading leaves that are quite conspicuously translucent. Thus there is no difficulty in suggesting that they belong with H. heidelbergensis as does the next.
Middelrivier MBB7037.
Leeurivier JDV87/4. This particular population has been linked to H. heidelbergensis var. scabra and the following recent collection seems to confirm that this is the continuity that should be sought.
Nuutbegin MBB7099. This population is now south of the Breede River again and the problem is that it does not seem to relate to the initial populations in set A, but rather to those south in the Potberg area that I have not included in this particular discussion. Those populations are dealt with in a review I wrote of the H. floribunda complex and could initiate an overview of H. heidelbergensis.
F92 MBB6516.1 W GoudmynF93 MBB6676.1 OlifantskransF93 MBB7081.1 E GoudmynF94 MBB7080.1 WakkerstroomF95 MBB6509.1 W Bonnievale096 MBB7085.1 E BonnievaleF97 MBB7082.2 BobbejaanskloofF97 MBB7083.1 N GoudmynF98 MBB6964.1 E ZandvlietF98 MBB6965.1 ZandvlietF99 MBB6981.1 SE LoerkopF100 MBB6974.1TonteldoosF100 MBB6977.1 TonteldoosF101 MBB6990.1 MardouwF102 MBB6988.1 NooitgedachtF102 MBB6989.1 SE NooitgedachtF103 MBB6992.1 LangverwachtF104 MBB6971.1 JakkalskopF105 MBB7036.1 Witkop, DrewF106 MBB7057.1 W SanddriftF107 JDV89/8 SanddriftF107 JDV90/31 SanddriftF107 JDV96/25 SE DrewF107 MBB6625.1 SanddriftF108 JDV87/193 N SanddriftF108 MBB6626.01 N SanddriftF109 MBB7037.1 MiddelrivierF110 JDV87/4 Leeurivier
Discussion It is wishful thinking to even hope that some taxonomic decision can flow from the above discussion that will match the traditional classification of Haworthia. I consider that I set out to describe H. maraisii in the same format as a series of 5 articles that I wrote for the English Cactus and Succulent Journal entitled rather grandiosely as “Natural variation and species delimitation in the genus Haworthia” (ca 1972). I never completed the article because I could see that the 20 or so populations that I had then sampled held insurmountable problems. Since then an obstacle for me has been the seeming intellectual intransigence of readers, expressed by other writers, who like myself seem to have been wishing for some magical formula whereby the genus would unfold and disclose the obvious and comfortable units that we all wish species were – and also by lack of full exploration. My uncomfortable role has been to try vainly to educate and instruct a small willful segment in both academic and non-academic rank, that this Holy Grail is based on not only false hopes, but on false premises.
Classification of Haworthia, and indeed I contend that this will apply to many other genera, simply will not be accomplished unless there is agreement on purpose and definition. My botanist acquaintances seem to insist that there are many definitions for the word “species”. My non-botanist acquaintances are justifiably unconcerned about definition at all while they have names that they can use. That these names as Latin binomials should have a particular meaning in the whole context of biology seems to be lost to both groups. Thus the nomenclatural system as expressed by the International Code serves no true biological purpose. It simply regulates and legislates for the methodology behind a nomenclatural system whether this is for true “species” or for elements recognized out of sheer lack of sampling and poor definition.
My observation derived from the material discussed in this manuscript is that I can no longer see a clear distinction between H. maraisii and H. mirabilis as the names have been transfixed by typification either by myself or by I. Breuer or by the way in which I have applied them. The problem is not simplified by the fact that at both Bredasdorp and Napier it appears that two elements are in close geographic proximity. This rather suggests that a ring-cline may exist. I suggested in my Revision, that a taxonomic solution could be possible for species (populations) together in one locality (or close proximity), but wrong elsewhere. This observation seems to apply here. Similarly I cannot see that the set of populations in E above (H. heidelbergensis?) can be circumscribed as a species in anyway that H. heidelbergensis can truly be circumscribed and seen to be discrete. More discomfort extends from the interplay of H. heidelbergensis, H. magnifica, even H. floribunda, and the complexity of that element in the east toward Riversdale and Albertinia. The relationship of this aforementioned complex to H. emelyae in the Little Karoo is compounded by the connection via Barrydale of the greater H. mirabilis set discussed in this work. There are thus other sets of species which need to be considered in the same way as has been done in this work
What I do suggest is that species are fractal and that there will never be clear focus on all life forms. There are species that are presently in sharp delimitation and species in transitional mode that are not. This is particularly evident within Haworthia where some species are quite obvious and discrete and others are not. The scale of the taxa generates the same problems as it does in the artificial rank of genera. This is that there may be one or more very distinct elements that are quite disproportionate in numbers and populations to amorphous elements. The decision in Haworthia should be reached by consensus among individuals who indeed have the wisdom, competence and experience to make sensible choices. The evidence for these qualities has so far been sadly lacking.
Conclusion The set of populations that I have examined here, suggests that the probability that they can be proven to constitute two separable elements viz. Haworthia mirabilis and H. maraisii, is remote. From a practical point of view it is possible to uphold the names in the traditional way followed in my Revision, but only if it is accepted that these names cannot be applied with certainty to the greater proportion of the populations. From a botanical point of view it is patently evident that we are dealing with one species, Haworthia mirabilis. There is a greater problem in how this one species known by the populations discussed in this paper relates to the populations geographically further east. The implications of such a relationship for classifications of Haworthia may be profound and it could impact on classification of many other genera too. The obvious solution is to decisively abandon the dream of a classic nomenclatural solution to the problem. We could simply recognize that variants, which exist within the scope of any species, can be accommodated by simple descriptive statement and referred to by collecting reference and locality.
Acknowledgement I have been so privileged to have made the acquaintance of many people who have facilitated my excursions. Practically every spot on the map is associated with several persons. I truly regret that it is simply impractical to name them all and I sincerely hope that my appreciation and gratitude will not be taken to be lacking by dint of omission. ♦
In a very interesting book by Stephen Gould entitled “Rock of Ages”, in which he propounds his principle of NOMA – non-overlapping magisteria. This states that science and religion should not be confused nor mixed.
So this is not a confession of confusion – you do not confess to what is obvious. It is an admission, and an admission can be construed as an apology. But, as a rhetorical question, how can one apologize and expect forgiveness when one continues to walk the errant path?
I started to write about Haworthia to dispel confusion, and yet more than 40 years on, this confusion has not become any less. The conclusion I have come to (and I wish it was a closure) is that the prime source of confusion is simply the human condition. In mystic philosophy one can read… “Born in ignorance, we live in ignorance and we die in ignorance.”
I think that my interest in Haworthia stems from my conscious effort to dispel this primal confusion and find some of the order in my view of creation. The classification of plants suggested just one small piece of my world which was available to me, and Haworthia as one group which no one else could explain to me. What have I now learned and what contribution does this make to dispel confusion?
My courage to now say something more directly arises from a recent request by SANBI to write a synopsis of Haworthia for an E. Cape Flora. I feel that I have done that fairly successfully. The problem is now to produce a similar product for the SW Cape and this is considerably more difficult.
To explain Haworthia one has to first cross the hurdle of the present classification of the Asphodelaceae and the perceptions that Aloe, Haworthia and other genera are clear discrete groups that the classification suggests. Ignore completely the issue of whether the so-called species are clearly separable. Haworthia consists of three quite discrete groups which have no closer relation to each other than do any other of the extant Asphodeloid genera. They each have their own problems and idiosyncrasies but it is the subgenus Haworthia which concerns me most.
In 1975 I prepared a map to illustrate the relationship of the SW Cape species (primarily south of the Langeberg Mts.) and this was published in Excelsa (5, 1975). In this map I tried to illustrate the problem of continuity which simply makes it impossible to recognize clear closed groups of plants which we can with any truthfulness say are species. It is the reality for Haworthia, and my conviction is that the model which underlies the Latin binomial system is fundamentally flawed. It works because our ignorance clouds its shortcomings.
Map 1 Distribution of SW Cape species of Haworthia
Updating my map may show why (Map.1). It is perhaps possible to identify three main elements that are nevertheless extremely difficult to map. Very seldom do we find all three growing in close proximity and never do we find any of the components I suggest doing so either. We often struggle to suggest which of the three we are being faced with.
The names we can use for these three are:-
H. retusa – to include or cover mutica, turgida and pygmaea
H. mirabilis – to include or cover maraisii, magnifica, emelyae and heidelbergensis
H. floribunda – to include or cover parksiana, chloracantha, floribunda and variegata.
There are other elements viz:-
H. herbacea – to include or cover reticulata, maculata and pubescens
H. rossouwii –
But this is a simplification and not a solution.
What simply has to be recognized as a fact of life that this problem is not of my creation and I do not believe that the use of any technical sophistry, trick of classification or nomenclature will possibly dispel it. These “species” are as interconnected as the strands of a complex web as intricate as any a confused spider could construct.
Essentially one has a mid- to late-summer flowering group and a spring flowering group, but they are linked. H. pygmaea, in the retusa complex, actually springs from the mirabilis complex; while its counterpart mutica in the west, seems to spring from the retusa complex. Emelyae, in the mirabilis complex, seems to be linked back to rossouwii via multifolia. These are the kinds of realities one has to deal with.
The relationship of the three main elements I have mentioned differs from point to point on the map, and to describe these relationships means that one has to virtually deal with populations and groups of populations one-by-one. I have done that, particularly in the book Haworthia Update Vol. 1n and again in Vol. 2. But since the drafting of the last manuscript, I have been to the field again and found more material, which I will show now.
Essentially there are three areas which I will cover:- a. the southern Potberg and where the Breede River enters the sea south of Swellendam (Map 2); b. secondly Klipfontein at the northwest of the Potberg (Map 3); c. thirdly an area just west of that which I also call Die Kop, Wydgelee (Map 4). The localities are not indicated precisely on these maps but the detail is in archived and available information.
Two of three relevant populations are discussed in Chapter 15. These are ADH594 Sandhoogte, and MBB7248 Lower Breede River H. mirabilis (pilosa). I refer to CB2018 and I can now report and illustrate it myself. This is MBB7251 (Figs 2a & b) from Buffelsfontein within the DeHoop Nature reserve. In the field the plants are not distinguishable from ADH 594 and grow in exactly the same Fynbos habitat viz a very rocky and grassy streamside slope with a dry northern aspect. The illustrations are of plants cultivated and hence far greener and softer than in habitat. What is striking is the tendency to surface spination which is such a feature of MBB7248.
b. Northwestern Potberg.
Some of these populations are also covered in Chapter 15. These are MBB6890, MBB6886, MBB6882, MBB6889 and MBB6544 (figs A18…22). My comment that H. variegata occurs here too should be borne in mind. This is on the upper slopes of the extreme western end of the Potberg. There is another record of C Burger’s from further east and south of Diepkloof for which I coined the name H. variegata var hemicrypta. Some doubt will now be thrown on the wisdom of that decision. The mirabilis populations referred to above are on the farm Juliesfontein and my new records are now east of that towards Diepkloof along the Breede River.
1. MBB7487 H. floridunda. Byeneskop (figs 3a to g). No good opinion is ventured in Chapter 15 concerning the populations noted above. Those populations seem to fall in the context of an interaction between H. maraisii/heidelbergensis/floribunda and I warn of infusion of H. variegata. The Byeneskop is on the very eastern border of the farm Juliesfontein and the plants are surprisingly and confidently comparable with the typical form of H. floribunda. The leaves are generally fairly strap like and flattened to the characteristic twisted ends as evidenced in floribunda. A few plants (see fig.3f) do have shorter and stubbier suberect leaves which characterize so many collections that confound the recognition of the relevant species. This particular habitat can be described as Renosterveld on silcrete with dominance of white clay and much grass and other succulents. Similar plants were also seen about a kilometer eastwards on the verge of vegetation transitional to Fynbos.
2. MBB7492 Klipfontein East (figs 4a to d). This is again a typically H. floribunda-like population. Whereas the preceding tends to be slightly tuberculate with the rougher greyer texture and colour of H. maraisii which would suggest floribunda var dentata, these plants tend to have a smoother leaf surface. Pictures of the flower are included although of very little diagnostic significance. The habitat is a steep rocky sandstone hillside vegetated with low Fynbos species.
3. MBB7494 Klipfontein East (figs 5a to d). The plants suggest H. floribunda except that the colour of the leaves is greener and there is a tendency to translucence, or less opacity of the leaves. The white flecks in the leaves are quite conspicuous and are more often encountered in H. turgida. Their significance taxonomically is probably nil as these markings/inclusions may appear in plants as remote as H. angustifolia var baylissii on the Zuurberg, E. Cape. The habitat is again short grassy Fynbos but unusual because of the presence of finely textured white quartz rock.
4. MBB7495 Klipfontein East (figs 6a to c). While we have been looking at plants on a series of buttresses of the Potberg, these plants are now closely associated with large white quartz boulders. They are small and although very similar to the preceding, the leaves seem to have lost the characteristic rounded ends and the margins are relatively strongly toothed.
5. MBB7496 Kleinberg (figs.7a to g). This population is only about 1km east of the preceding but there is a transition to Renosterveld. The habitat is again silcretious with the typical white clay substrate. This seems to be a very erodable surface and plant cover is quite low. The plants are surprising because they depart quite radically from the floribunda-like mode of the preceding populations. They are now distinctly magnifica-like and particularly reminiscent of var atrofusca except much smaller. Curiously a population across a small valley southwards in sandstone and Fynbos is in keeping with the floribunda-like finds westwards.
6. MBB7499 Juliesfontein (figs 8a & b). We returned to Juliesfontein to look at a promising rocky slope between Byeneskop and our previous closest collection on Juliesfontein. These are almost directly north of H. variegata and as can be seen from the pictures they revert to the forms described in Chapter 15 for that area viz. there is a similarity to heidelbergensis. The more leaves are more frequently pointed rather than rounded and translucence and venation is more conspicuous.
c. Die Kop, Wydgelee.
The nearest plants dealt with in other chapters are MBB7539 at Tarentaal that I liken to magnifica var atrofusca and MBB6030 at Napky which is maraisoid with floribunda-like character (type locality for H. maraisii var simplicior with rotate leaf-tips). It is important to note that the Tarentaal plants are not uniformly larger that my identification suggests and also that H. mutica is also present at its most southeasterly locality known south of Napky. Tarentaal only just appears on the map and the mutica locality is just off the map to the upper right (northeast) corner. It should be noted that populations assigned to H. heidelbergensis occur to the west at Kathoek, Beyersdal, N. Windheuwel and terminating in the contentious var minor at Rooivlei north of Bredasdorp. I think if an overview was now made of all these populations I have assigned to heidelbergensis in different chapters and in different publications, it will have to be concluded that it may represent the pivotal element in the whole classification process. Where it was the least represented species in the period preceding publication of Haworthia Revisited it is now far better known and any hypothesis prepared to test relationships in these southwestern Cape species Haworthia will need to question this centrality. Die Kop is a set of 5-6 silcrete and ferricrete hilltops of varying size and height but otherwise not significantly different from a large number of such habitats extending from Stormsvlei in the northwest and terminating against the Potberg and Breede River in the southeast. Collections from several of these have already been reported on in Chapter 15 with the general observation that their identification is in the uncertain linking maraisii and mirabilis. I hope, in the minds of readers, that I have not excluded heidelbergensis from this milieu.
1. MBB7500 Die Kop East (figs 9a to v). I doubt if any collection has generated such reservations in my mind as this one has and many illustrations are used to express the problem. The plants are large and up to 80-100mm in diameter. They are closely spaced in a small grassy, non-rocky area which seems to be geomorphically stable. They are extremely variable and resemblances extend from badia to retusa to mutica. The leaves may be short and rounded or fairly long and attenuate, closely adpressed to the ground (recurved) or sub-erect. One notable feature is that the plants have a milky colouration to the leaves that Seven Hammer once remarked on as a feature of H. mutica. While variability is known to be great, it is seldom that it is as dramatic as this in a population of plants as they occur in the field.
2. MBB7502 Die Kop West (figs 10a to c). The plants here are smaller and more widely distributed and scattered over a larger area. The habitat is skeletal and rocky and the vegetation more herbaceous than at the eastern site. The similarity of the plants to those of 7500 is close but the variation is less extreme. There is less evidence of a mutica influence and a return to more or less the maraisii/floribunda/heidelbergensis archetype.
Fig. 9 7502 Die Kop W
3. MBB7503 Die Kop Mid (figs. 11a to i). The first impression I had was that the plants included more mirabiloid influence. Not as big as 7500 and also scattered on a skeletal slope. At one point there was a gravel patch with Gibbaeum austricola which is commonly found on such gravel patches associated with ferricrete inselbergs.
4. MBB7504 Die Kop North (figs 12a to k). The plants are smaller and there is a more evident floribunda influence. I did not think that these plants were significantly different from the many populations discussed in Chapter 15 between Tarentaal and Stormsvlei.
Conclusions: It should be very evident from all the information provided so far that more information is not producing a better picture of discrete elements. To the contrary and this reinforces the conclusion already mooted, that movement to a new view of “species” in the subgenus Haworthia is necessary, but is it possible?
Acknowledgement: Mr. Keith Spencer, DeHoop Nature Reserve. Messrs. Christine & Malcolm Wallace, Ziekenhuis. Mr. John Douglas-Hamilton, Ballyfar. Mr. Johan Groenewald, Klipfontein, Potberg. Messrs. Saartjie and Neil Neetling, Juliesfontein. Messrs. Adele & Vlooi DuToit, Die Kop & Frederickskraal. Messrs. Pet & Johannes Uys, Die Kop. Messrs. Sofia & Jurie Vermeulen, Tarentaal
Statistical analysis of two populations of Haworthia mirabilis (v.Poelln.) M.B.Bayer
M.B.Bayer & L.M.Loucka
Introduction: In discussing H. rossouwii (Aloe 38:31, 2001), Bayer mentions the possible continuity with H. mirabilis var. sublineata. But any comment like this is complicated by the problems of variation, description and circumscription. We want to discuss the variation in the latter element and indicate further where the problems are in the delineating species and varieties. It seems that one of the assumptions of classical plant taxonomy is that of linear dichotomy, black and white, this species or that, and also that there is hierarchical and consistent in-group similarity to some unstipulated degree. Haworthia, and particularly the subgenus Haworthia, presents a problem to those interested in the genus in that the classification is confused and that identifications are difficult. Attempts been made to explain that the classification is confused by the perceptions associated with classical taxonomy, and that the sharp and precise discontinuities suggested by a ‘key’ to the taxa, simply do not occur in the subgenus Haworthia, in fact they do not occur in many other genera, and this simple truth seems to be difficult for some to accept.
Thus this article includes a report of a study done on a batch of seedlings of H. mirabilis var. sublineata. It shows that there is very little probability that one could quantify separation of this from other populations presumed to be the same species.
Outcomes in the biological world are influenced by genetics and environmental factors, but also by chance events. It is because of the probability of chance events that knowledge of statistics is essential to scientific research and our studies of the genus Haworthia.
Plants have parts that can be mechanically measured in terms of spread, height, width, length, thickness, arc; or counted as in the number of leaves or offsets. Other attributes like colour, and surface texture are more subjective. When we observe leaf structures in Haworthia our perception often tries to make sense and draw patterns from many subtle characteristics viz. this plant is species X because it has a pattern that matches our experience. But our brains can be easily fooled when we try to compartmentalize. When we subject our observations to measurement we begin to sort out the characteristics we use to describe one plant, population, or species from another. Viewing this collection in the leisure of the greenhouse, we can see that some leaves were thicker or longer than others, but are they “really”? That is, are they significantly thicker, or longer? When we count the number of leaves, or measure their length can we make some statements to compare and differentiate one population from another? Do the differences we measure translate into differences in overall size and shape of the leaf? Let’s investigate this further by formulating the following question:
Method: Seed of H. mirabilis var. sublineata was field collected at the type locality south of Bredasdorp in April 1997, accessioned as MBB6639. By Sept.2000 these seedlings were at flowering size and 71 plants were available for the study. A second study sample was taken from a population of H. mirabilis var. triebneriana from Fairfield, west of Napier (MBB6643). H. mirabilis is very widespread and this Napier population is apparently no more or less variable than many others known.
Population 1, MBB6639 – 3420CA South of Bredasdorp. April 1997. North facing steep rocky slope, Table Mountain Sandstone with grassy mountain fynbos.
Population 2, MBB6643 – 3419DD Fairfield 15km West of Napier. April 1997. North facing shale ridge, Bokkeveld Shale with grassy renosterveld.
Prior to this study, an attempt was made to reduce the size of the group of MBB6639 seedlings by discarding any single clones which were obviously similar. To do this Bayer tried to arrange the plants according to similarity. He could not find any pattern, and each plant appeared to be fairly unique. Five plants were removed in order to propagate them vegetatively, and for this study a further four clones were removed which were either obviously atypical or retarded. Four smaller apparently under grown clones were retained in the final study batch of 62.
A first attempt was again made to try and identify groups by visual comparison. This proved impossible to do. Attempting to reduce the size of the set by the identification of “identical” pairs also failed. No two plants were sufficiently similar to be regarded as near to identical. The simplest way forward was then to match the plants as closely as possible into pairs, repeating the process five times. After pairing, the pairs were broken and the plants returned to a random array. Pairing was then done again by picking up any one clone and finding the best match. Some re-sorting was done in order to accommodate the last of the clones as the pairing process neared an end. In some cases clones which could not be adequately paired were compared over all the others to find the best match. Although an attempt was made to score the “goodness” of pairs, this proved difficult to do and an inconsistent result was obtained. Some clones simply proved more difficult than others to match.
With five attempts, the total pairings was 310. Sixteen clones were unpaired. Of the pairings, 210 were unique, in 74 cases the same two clones were paired, in 18 cases the same clones were paired three times, in 8 cases four times and in two cases, all 5 times. This shows that there is very little consistent pattern in the group as a whole when trying to match or differentiate based simply on assessing visual cues.
Recognising now that no progress was being made, Bayer considered what ‘characters’ he was using in his visual assessment and then quantified his observations. Some of the characters used in the visual process could include:
a. Size b. Colour c. Leaf number d. Leaf width e. Leaf length f. Leaf thickness g. Leaf recurvature: i. compactness, ii. erect or reflexed h. Leaf marginal spination i. Leaf upper surface: i. translucence, ii. texture, iii. venation, iv. basal opacity, v. basal marking j. Leaf lower surface (as above) k. Proliferation (off-setting)
For statistical analysis it was decided to use the measured spread of the leaves from the tips of the most extended leaves, the height reached by the highest leaf tip measured from the base, the width, length, and thickness of the largest leaf at maximum, and to measure the arching of such leaves by the gap between a line drawn from base to tip and the lower leaf surface (i.e. recuravture). A subjective measurement was made of leaf surface roughness, size and number of spines, and degree of tubercles. The resulting data was analyzed by a Dotplot for each characteristic and a Two-sample T-test used to assess the null hypothesis.
Descriptive Statistics
The MBB6639 plants show the classic bell-shaped normal distribution for most characteristics with the exception of spread and proliferation (off set count). Perhaps the population culling of the runts and natural survival account for the truncated spread data. In time more of the clones may have offsets and thus show a different distribution. For the other characteristics we can see a central tendency and data points spread to the left and right. It’s this central tendency, or average (statistically called the mean) that we’ll study further.
Hypothesis Testing
Question: Are plants from the two populations in this study of the same or different sizes and shape?
Hypotheses: The difference in the average physical characteristics between the two populations is zero; i.e. the null hypothesis (Ho:). If the difference between the means of the samples is among those that would occur rarely by chance when the null hypothesis is true, then the null hypothesis is rejected and the investigator describes the results as statistically significant. That is, the difference between the average of each population characteristic must be great enough to conclude if the populations are the same or different.
Prediction: If the leaves of the two populations are the same size, then the spread, height, width, length, thickness, and arc of the populations should be the same.
Analysis:
The P value we subject our data to is 0.05, or a 5% chance that we would accept that the difference is the means of the two populations is zero when in fact it isn’t.
A null hypothesis is not accepted just because it is not rejected. Data that is not sufficient to show convincingly that a difference between means is not zero do not prove that the difference is zero. Such data may even suggest that the null hypothesis is false but not be strong enough to make a convincing case that the null hypothesis is false. For example, if the probability value P were 0.15, then one would not be ready to present one’s case that the null hypothesis is false to the (properly) skeptical scientific community. More convincing data would be needed to do that. However, there would be no basis to conclude that the null hypothesis is true. It may or may not be true, there just is not strong enough evidence to reject it.
What can we conclude? The two populations appear to be different for leaf width and thickness. For all other characteristics we just don’t have enough data to make any conclusive statement.
We have illustrated the individual clones for the reader to appreciate how different the individual plants may be, and also to consider that the appearances of the plants will vary according to growing medium, exposure to light, season and watering.
Discussion and Conclusion
Our observation is that a statistical approach simply does not penetrate the nuance of difference that visual observation can discern. The data we present is valuable only in that it illustrates the variation within the population MBB6639. The coefficients of variation of the characters used are so high that larger samples would be required to provide more reliable statistics. In this study we have used a second population 6643 which is obviously different. At Bredasdorp there are in fact two other populations. One of these is on a north-facing river bank in among river-eroded sandstone boulders, while across the river about 250M away is a smaller population in Bokkeveld shale. Plants from all three populations are relatively easy to identify. The riverine boulder plants have fewer stumpier and more reflexed leaves, while the shale plants have erect leaves. To what extent these differences are “real” cannot be proven without a replication of the seed collecting and propagating effort done for 6639. In the context of ALL the populations which may comprise the species H. mirabilis it is quite evident that this is impractical. ♦
My experience is that Latin names definitely mean different things to different people. I submitted this manuscript as a draft to various people and the response varied from one which was nil, to some sort of general accord. I am, however, no longer confident that botanists either do or will agree with my contention is that the real essence of Latin names should, in addition to their many other usages, be in the relation of plants to their origins, relationships, behavior and imagined future. A classification can only have the authority that experience and knowledge permit, and be really evaluated and understood by persons with the same sort of evidence before them. In coming to closure I have been exploring some more, and with my wife Daphne, made two finds which further convince me that we have to come to a classification by agreement. However, the requirement is that species are seen to be highly complex systems with none of the rigidity and inflexibility that nomenclatural rules imply, nor any of the egocentric authoritarianism that a history, of which I have been a part, suggests.
I have recently written two papers. One deals with the H. nortieri complex, and the other with H. pygmaea. In writing the latter I interwove the comments of observers whose opinions I value, and conclude with the proposal that the elements acuminata, splendens, dekenahii, argenteo-maculosa, fusca and vincentii all be included in the super-species H. pygmaea. Concomitantly I suggest that H. turgida and its variants be absorbed in H. retusa. Behind these two propositions, I was intending to re-enforce my view expressed in Haworthia Update Vol.3, that H. mirabilis too be re-structured to include magnifica, maraisii, heidelbergensis and any associated varieties. The reason of course been the problem of continuity, however that is understood; and I suggest it is best understood by familiarity with the plants in the field and some recognition of biogeographical factors and the role these may play in driving change, adaptation and evolution.
In Update Vol.3 I illustrate examples from many of the populations known to me, and also draw attention to populations in the lower Breede River valley which are significant. Chapter 13 is entitled “Haworthia is confusing” and Kobus Venter kindly used this material to make a presentation to the Succulent Congress at Calitzdorp in 2006. In addition I sent a draft of this recent manuscript to Bob Kent who replied that he was not sure what I meant by agreement. My missive to Bob included two collections made subsequent to Kobus’ presentation and that are a harsh reality check. These need to be added to the material touched on by Kobus. The plants I illustrate come from only a few populations that I know of and I believe that the only logical conclusion that one can draw from that greater body of known populations is that there is one super species, H. mirabilis – among others. My view now is that we need to take ALL the populations known at the very least to me and try to build a rational, consistent and coherent classification which might fit an imagined model of a product for botanical science. Therefore what I mean by “agreement” is virtually that readers have to submit to the uncomfortable situation that I may be right and that there is no alternative but to go along with what I have concluded. The reservation is that new material may necessitate modification and my opinion is that it will probably drive the classification towards a still more conservative position.
My most recent excursion has in fact been to try and resolve the dramatic juxtaposition of variants that we lower down the Breede River. In summation of these, we have Adam Harrower’s collection (Fig.1) from Sandhoogte and Chris Burger’s collection, and (Fig. 2) at Buffelsfontein. These are both south of the Potberg.
Fig. 1. MBB7251 H. mirabilis. Sandhoogte, De Hoop.Fig. 2. MBB7374 H. mirabilis. Buffelsfontein, De Hoop.
Then there are my collections from Stoffelsriver (Fig. 3), one nearer to Infanta (Fig. 4) and then two from Kleinberg (Figs 5 & 6). A curious addition that I have yet to see in the field, is a collection by Ismael Ebrahim (Fig. 7) of SA Biodiversity Institute from southwest of Vermaaklikheid. That collection resembles my Stoffelsriver collection and so provides the inevitable link to paradoxa.
Fig. 3 MBB7520.1 H. mirabilis. Stoffelsrivier.Fig. 4. MBB7248 H. mirabilis. Lower Breede River.Fig. 5. MBB7496.1 H. mirabilis (alias maraisii cf atrofusca. ESE Kleinberg, Matjieskloof.Fig. 6. MBB7516.1 H. mirabilis alias maraisii. Kleinberg.Fig. 7. Ismael Ebrahim sn. H. mirabilis alias paradoxa. Koensrust
At two localities near Melkhoutrivier (Figs 8 & 9), between Stoffelsrivier and the Infanta collection, we found plants which have to be related to those mentioned above. The plants are highly variable as I have now come to expect, and individuals can be likened to badia, acuminata and mutica. The surfaces have a curious sheen and may be incipiently spiny as the Sandhoogte and Buffelsfontein plants also are. The paradox is that we are also driven to the conclusion that the very small plants at Ziekenhuis (Fig. 10) are the same species. Collections, figures, 1 to 10 are a set within an incontestable biogeographic zone where geology is probably the most notable primary variable.
Fig. 8. MBB7608.1 H. mirabilis. Melkhoutrivier W.Fig. 9. MBB7609.1 H. mirabilis. Melkhoutrivier E.Fig. 10. MBB7615. H.mirabilis alias maraisii. Aalwee, Diepkloof.
What re-enforces this seemingly improbable juxtapositioning of such different things, apart from my lengthy dissertation on the matter in Update 3, is a second find we made southeast of the Bromberg (Figs 11 & 12). This is near the locality for what I suggest is H. rossouwii var elizeae, and also for several variants of what have in the past, been classified as maraisii and mirabilis or variants thereof. My new find is a remarkable array of large very dark-green (nearly black), plants which dramatically enforce the close association between mirabilis and maraisi as well as with heidelbergensis. These plants include the elements of both “species” as they might have been understood. What is striking is again the huge variation in the population that I have come to expect in this complex. The plants do lack the opaque dots on the under-leaf surfaces while the upper surfaces may be clearly windowed or opaque. They bring forcibly to my mind a sequence of populations from Heidelberg in the east to Verdwaalskloof near Riviersonderend in the west that I suggested forged the interface of mirabilis and maraisii. Coupled with this is the array of populations both north and south which enforce that continuity and that also lead on to heidelbergensis. So while collection 11 & 12 … may fall outside the compass of a Lower Breede biogeographic zone, it is unlikely that evidence can be found to suggest that it is not in a continuum of many other populations that link it with that zone. We are thus dealing with elements which are grossly different in imagery that constitute one system that can be said to be a “species”. (Note:- the pictures are of single plants and they convey neither the respective sizes of the plants, nor the gross variability within those populations. It is nevertheless true that generally one could probably assign a given specimen to each population – if required to do so).
Fig. 11. MBB7612.3 H. mirabilis. Diamant.Fig. 12. 7627.1 H. mirabilis. Diamant E.
My recommendations are:
1. deficiencies in respect of a species definition be admitted and rectified.
2. the nomenclatural code is summarily assigned a secondary role to a dispensation which is more flexibly attuned to the realities of a truly asymmetric species structure with more emphasis on reflecting field relationships, and to the competence of its users.
3. the illusion of reality that the ranks of genus, subspecies and variety provide should be admitted. Genera should be recognised for their historical and artificial value, but for species I would suggest that much more attention be given to the huge asymmetric and asynchronous variability that underlies capacity to change and adapt.
With the adoption of, say, H. mirabilis as a super species, variants can be indicated by the addition of any other epithet in inverted commas; thus H. mirabilis ”maraisii”, H. mirabilis “heidelbergensis”, H. mirabilis “Melkhoutrivier” in the knowledge that there is in fact no clear and infallible distinction, or pretension that minor ranks have any reality either.
In completing this manuscript and re-reading it, I could not help but keep referring to the book by Felipe Fernadez-Armesto entitled “Truth”. The cover blurb includes the words… ”We need a history of truth – though until now no-one has tried to write one.” There is also a note by Robert Winder which reads … ”A sharp and interesting work bound to enrage specialists in the fields he sprints through.” My own summation is that I have tried to write a truthful account of Haworthia. It will surely also enrage others who try to do the same, as well as others who may perceive the truth in some other fashion. To them I truly apologize. I do not have the answers and find my own writing quite as pompous and irritating as my readers may too. It does appear to me that we have as a society ”… lost faith in the reality of it and lost interest in our search for truth.” ♦
Clichés may often fall into the category of often repeated untruths that come to be believed. One I have used too often is … ”The problem is …”, without ever seemingly being to explain what it actually was. I studied Oxalis and it seemed that where there was an awful amount of detail to explain difference, all this detail simply obscured the fact of similarity. So what I did is “reverse engineer” the process and apply the principle to Haworthia where I have for so long tried fruitlessly to explain that we were all explaining and accepting that there was difference based on detail.
I made some progress in finding facts to support this contention in the approximate 40 populations of Haworthia in the Zuurberg that seem to suggest that plants looking as different as H. cooperi and H. cymbiformis may be the same species. However, this was not very convincing.
Recently I had reason to explore more populations in the arena of a larger problem in H. mirabilis where I may be considering as many as 400 populations or many more. Illustrated by the following sample pictures: … It is now my contention that different as all these single plants appear to be, they are in fact members of one species. The inference is drawn from observations of approximately 150 populations occurring in a geographically coherent pattern in the restricted area between Worcester and Riversdale and southward to the coast. The inference is strengthened by the observations of similar continual intergradation of variation in similar sets of populations throughout the distribution range of the genus.
I do think that this is evidence to put to rest the disputes about the classification of this group of plants. But I do think that there are implications for plant taxonomy in general perhaps starting simply in the Alooideae where there is such confoundment in the understanding of simply the genera. ♦
1. Haworthia mirabilis ‘paradoxa’. Vermaaklikheid2. H. mirabilis ‘paradoxa’. Vermaaklikheid (per Ismael Ebrahim)3. H. mirabilis ‘badia’. Napier4. H. mirabils ‘pilosa’. Lower Beede River.5. H. mirabilis ‘atrofusca’. W.Heidelberg.6. H. mirabilis ‘meiringii’. W Bonnievale
Steven Hammer, in his inimitable style, put a very fresh face on Haworthia in Cact. Succ. J. (Los Angeles) 80:140 (2008). He drew attention to the wonder of the plants in cultivation for the collector, contrasted to a reality of unglamorous scruffiness in the field as per the lens and pen of Bayer. It has fallen to my lot as a very unwilling taxonomist to reduce the fascination these plants have for me, and for so many others, to the mundane vortex of labels, their proliferation and continual amendment. The fact is whether on a label or on the tongue, a name is a part of any language we use to talk to each other; but we are not learning anything from a well-documented history and in Haworthia seem to remain lost in a maze.
The unhappy truth for Haworthia is that by the time von Poellnitz in Germany, G.G. Smith in South Africa, F. Resende in Portugal, A.J.A. Uitewaal in Holland, W. Triebner in Namibia, J.W. Dodson and J.R. Brown in USA had either left or abandoned the scene, there were any number of names that meant very little more than the Latin they were written in. J.R. Brown presented a talk, A brief review of the Genus Haworthia, to the Los Angeles Cactus and Succulent Society that was published in the Cactus and Succulent Journal of America 29:125-135 (1957). He noted the number of species and varietal names at 160 and 370 (!) respectively, arranged in 20 sections.
While J.R. Brown was winding down (his last note on Haworthia was published in 1970), I was busy trying to make sense of a two large files that seemed to form the body of a manuscript by G.G. Smith for which Mrs. M. Courtenay-Latimer had drafted a title… “A monograph of the genus Haworthia.” This manuscript comprised a collection of all current species descriptions arranged in the purported twenty sections of Berger and accompanied by many illustrations from the original publications, as well as by many of Smith’s own photographs and those of H.G. Fourcade. We know that Smith retired in a huff, but was there really good reason for his exit?
It is quite evident from the legacy of notes that the manuscript could not have been anywhere near complete. I drew attention to this in an article…”Reflections on G.G. Smith’s Haworthia collection” that was published in Excelsa 8:46-52 (1978). While stuck in the paradigm of 20 sections, he envisaged many more species and varieties and I cannot imagine how he could ever have reconciled this with the evidence of his compiled manuscript. I suspect he realized this too, hence the (self-directed?) huff.
It took me several years to absorb the specimens in the Compton Herbarium while at the same time taking Smith’s meticulous (in the sense of the literature from which it was extracted) collation of material to pieces. Putting it all back together as best I could, I eventually produced Haworthia Handbook (1976) published by National Botanic Gardens of South Africa. It was not a great work and neither was it very well-received at the time (though it has had a curiously perky afterlife once it went out of print). Charles Glass wrote to me that he was a bit disappointed (by its elliptic compressions?). Gordon Rowley was kind enough to point out many nomenclatural errors and a page of errata had to be inserted.
However, I was actually committed to helping Col. Scott write a formal revision as I had no aspirations to do so myself. He had been refused access to Smith’s collection, as the Curator of the Compton Herbarium had concurred with Mrs. Courtenay-Latimer (and perhaps Smith himself) that Col Scott would not do credit to a formal revision. It became quite problematic for me when I realized that this estimate of Scott’s botanical capability was uncomfortably close to true. This led me to re-write my handbook as The New Haworthia Handbook (1981), also published by National Botanic Gardens, hoping that Col. Scott would follow the ideas expressed in my optimistic foreword.
This did not happen and Scott’s…The Genus Haworthia – A Taxonomic Revision, appeared in 1983 published by Aloe Books, Johannesburg, as a closely and clumsily kept secret from me.
My honest opinion is that this book was a total disaster, taxonomically though not photographically, and it was most disheartening and confounding to consider that the flattering introduction was by Dr. L. E. Codd, then head of the national Division of Botany and of the Pretoria Herbarium. Not only that, Dr. Beth Gibbs Russell later telephoned me to ask whose version of Haworthia the herbarium and the institute should follow! I do not know what my response was other than thinking how unfair it was to place the onus on me to judge. Whatever I might have said, that did not stop the institute from publishing an extraordinary mish-mash of names for Haworthia in a national species list taken from Scott, myself and other sources. It was simultaneously no comfort to welcome the grand old man of Haworthia, J.W. Dodson, to the Karoo Garden. Bewildered by what he saw in the collection there, he could do no more than express his conviction that Col. Scott had produced a very workable revision!
As far as I was concerned, Haworthia was all emptied back into Smith’s “monograph,” and readers may imagine why I found this so disturbing. Around 1993-4 news of a new succulent lexicon to be compiled by Urs Eggli to replace the famous Jacobsen set was circulated. At the same time, by virtue of my contact with the Compton Herbarium, I was asked to write a synopsis of Haworthia for a list of plant species for the Cape Flora. These were the defining factors that led me to writing a formal revision. I cannot bring myself to detail the communications with other aspirants to taxonomic fame then and since, as these border on the bizarre and do nothing more than add to the disrepute of taxonomy in the minds of plant collectors who care about and are compelled to use Latin names.
When I eventually wrote my revision I had no illusion that it would be the last word on the subject. Indeed it seems in retrospect that it may well have been but the first! Not only has there been an explosion of new names and opinions, I have also been driven to examine and re-examine those areas of my understanding built not only on available field knowledge and exploration, but also on Smith’s vast legacy. It is necessary to note that the contributions of people like Jay Dodson and J.R. Brown played no small part either. The little book written by Brown, viz., Succulents for the Amateur, played such a significant part in cultivating my interest in plants that I have been very sensitive to the resonance of the devotion and interest Brown showed for Haworthia. This respect for others like F.R. Long and H.G. Fourcade, whom I never met, extended to people like Frank Stayner, G.J. Payne, Gordon King and so many others I met solely as a consequence of these plants. That the human relations aspect of their classification and identification took such a downturn in the wake of Smith’s criticism of, primarily, Resende in 1947 is a sad record in history. Relations never recovered and my involvement in and distress at the later half-repetitions of history is at the heart of this article.
Since 1995 my accession records have climbed from 6400 to 7700 and these numbers represent almost entirely new population records for Haworthia and I trust that I have not used my own numbers where a very few contributing collectors have helped me. I have written innumerable articles for Haworthia journals while Umdaus published Haworthia Update Vol 1 in 2002. This seems to have been a commercial failure but my belief is that editors and publishers have never understood the issues. I could not help because despite my excessive attempts to explain the problems causing strife and confusion in the literature, I really never exposed the heart of the matter. Fortunately for me Harry Mays of Alsterworthia kindly published Haworthia Updates Vols 2, 3 and 4 between 2003 and 2007.
In these volumes I tried to address the realities of variation and the confoundment that exists in my own and the communal mind, about what species are or might be as related to a perceived botanical and scientific process for naming plants. But if 40 delegates could be brought together as happened recently in an effort to create a united front just for the single genus Aloe, it does seem fairly senseless to be functioning so alone in Haworthia where a greater need exists. In my “unsparing” view of things I find it extraordinary that the symposium did not seem to address the wider problem of genera in the Alooideae. I am quite sure that if this were done first, it would facilitate the common understanding of lower ranks. In Haworthia there has been absolutely no common will to reach any kind of consensus and no reason has been found for that. My conviction has grown that Latin names, and the way the nomenclatural system is used, do nothing more than strain good will in that they implies a reality in a staged relationships of plants that is simply not true. If I were a more dedicated taxonomist I would have simply worked towards a functional generic arrangement: Haworthia for the glamour queens, Jayarbia (honoring J.R. Brown) for the Hexangulares, and Catevala for the Robustipedunculares.
I recently submitted a short list of names that I think should serve Haworthia to the Haworthia Society in England. As Gordon Rowley pointed out in a letter to me… “The problem is that Bruce Bayer changes his mind.” Well, I do want to change my mind about that list again. I would advise incorporating H. heidelbergensis, H. magnifica and H. maraisii and their variants all under H. mirabilis. Furthermore I would suggest abandoning the formal lower ranks of subspecies, variety or forma for appended third epithets in inverted commas. The list of species is thus reduced to a realistic 54.
But there is still a wealth of undiscovered things out there and the many existing names mean very little to me in understanding what I have seen and continue to find. So I give a brief overview of the product of very recent fieldwork where in less than 10 days my wife and I found 24 previously unrecorded Haworthia populations. These included two populations of H. floribunda, one of H. retusa ‘nigra’ (implementing a name change and a new way of expressing difference), five of H. rossouwii, one of H. minima, one of H. marginata, two of H. mirabilis ‘paradoxa’ and 12 of H. mirabilis. While each of these has a story attached, I am only going to touch on the latter.
I have written in detail about H. mirabilis in Haworthia Update 3:87 in a chapter that deals primarily with the populations in the western area of the distribution. I also wrote in detail of associated plants in Update Vol. 2:51-79 dealing with populations east of Heidelberg in the eastern area. So these new populations are relevant to the central zone and I will add several others that were found earlier but subsequent to the Update 3 publication, plus another by Marc Mougin (brought to my attention by Diederik Van den Abbeele). They all deal with an area extending eastwards from Stormsvlei and the Bromberg Mountain to the Karringmelks River southwest of Heidelberg and southwards to Koppies and Stuurmanskraal, where there are populations already covered in Updates.
Five of the populations can be unequivocally assigned to H. mirabilis ‘atrofusca’ and yet with a fair degree of variation. These are all just west of Heidelberg. The other seven that do nothing more than confound understanding of anything that can be construed as H. maraisii, H. magnifica, H. heidelbergensis or H. mirabilis, occur southwest of Heidelberg and extend westward to Diamant (east of the Bromberg). The only significant thing about the two groups is that where ‘atrofusca’ is evident in the populations, H. floribunda may be in the vicinity. Where H. floribunda is not present the seedlings resemble that species as an odd indication that perpetuated juvenile characters underlie the species floribunda. The variation within the populations is quite dramatic and it is indeed difficult to verbally report and sensibly convey this to a reader. The plants vary in size, texture, markings, leaf armature, color and translucence. So I submit pictures with the manuscript and offer the following comment as best I can:-
Fig . 1 Map of greater Swellendam area showing population points
A map is provided as Fig.1. Greater Swellendam area to show positions of Haworthia populations noted.
1. MBB7254 Bromberg. (Fig. 2). This really belongs with a large set of populations extending westwards and north along the Riviersonderend Mountains and into the Robertson Karoo. I start with it because it serves as a virtual ‘outgroup’ — and this is because the southern slopes of the Bromberg are different to the northern slopes. They are quartzite (silicon oxides) sandstones and the northern slopes felspathic (other silica salts). This difference in mother material manifests itself in the Haworthia populations and can be used to explain why H. maraisii v.Poelln. in the shales and siltstones at Stormsvlei are so different to H. mirabilis ‘depauperata’ VPoelln. in the quartzitic sandstones less than 1km north. The plants are relatively small and have the reddish color often associated with H. mirabilis south of the Riviersonderend Mountains.
Fig. 2 7254 H. mirabilis. Bromberg.
2. MBB7612 Diamant W (Figs 3 & 4). These are very big plants up to 110mm in diameter with a similarity to ‘maraisii’ by virtue of their blackish-green color and more retused (bent back) leaves. The classic difference of ‘maraisii’ and mirabilis as being separated by translucence absent in the leaves or present, is again flouted in this population as in so many others; and similarly leaf surface texture is variable. The significant thing is that the mountain they are on is an extension of the Bromberg (Riviersonderend Mountain tail) but it is the northern slope and the sandstones are felspathic.
Fig. 3 7612.3 H. mirabilis. Diamant W.Fig. 4 7612.4 H. mirabilis. Diamant W.
3. MBB7626 Diamant Mid (Figs 5 & 6). This population is less than a kilometer east of the receding on a more eastern and thus cooler aspect. The plants are a little smaller, the leaves generally more erect and are a little more reminiscent of mirabilis to the southwest in that spines are better developed on the margins and surfaces than in the case of classic ‘maraisii’ (I am trying to show that ultimately there is no infallible distinction).
Fig. 5 7626.1 H. mirabilis. Diamant Mid.Fig. 6 7626.2 H. mirabilis. Diamant Mid.
4. MBB7718 Diamant E (Figs 7, 8, 9 & 10). This is a huge population on a lower elevation than the preceding and the skeletal habitat is significantly different. It is on the interface of the Felspathic sandstone and shales and siltstones of an older formation while a strong vein of quartzite transects it. In addition there is evidence of an eroded away, and later, ferricrete deposit. The plants are also extremely variable, reflecting the range of divergent surfaces although generally much smaller than preceding. Leaf surfaces are mostly roughish.
Fig. 7 7718 H. mirabilis. Diamant E.Fig. 8 7718 H. mirabilis. Diamant E.Fig. 9 7718 H. mirabilis. Diamant E.Fig. 10 7718 H. mirabilis. Diamant E.
5. MBB7633 Luiperdsberg (Figs 11 & 12). A small population quite high up on a slope of felspathic sandstone. These are small plants and because of quite a grassy habitat the leaves are a little elongated generating the range of forms that I have generally assigned to a little understood or known range of plants and populations in H. ‘heidelbergensis’ Smith.
Fig. 11 7633 H. mirabilis. Luiperdskop.Fig. 12 7633 H. mirabilis. Luiperdskop.
6. MBB7719 Dagbreek (Figs 13, 14, 15 & 16). An established population in felspathic sandstone but marginal to shale and siltstone. Very grassy but with occasional rocky ridges so that the plants are concealed in grass tufts or wedged in crevices. The plants are moderately sized to 60mm diameter and differ in that many have a rather smooth surface with corresponding finer and more closely spaced marginal spines. Some plants develop a shiny opacity about the leaf tips that suggest surfaces perhaps comparable with the smoothness of the H. retusa complex.
Fig. 13 7719 H. mirabilis. Dagbreek.Fig. 14 7719 H. mirabilis. Dagbreek.Fig. 15 7719 H. mirabilis. Dagbreek.Fig. 16 7719 H. mirabilis. Dagbreek.
7. MBB7704 Bontebok Park (Figs.17, 18, 19, & 20). There is an earlier collection (6644) from the ferricrete/shale interface alongside the N2 highway near the northeastern corner of the park, which I considered belonged in a series of five populations that seemed to be neither H. ‘maraisii’ nor generally accepted H. mirabilis. It is on ferricretes but there is some problem how to relate this apparent river terrace gravel with the solid caps of the inselbergs to the east and west. There is nothing common in the general suite of species that could be described as Fynbos on the former and grassy Renosterveld on the latter. The plants are moderate in size; surfaces can be quite rough with a tendency to spination, and the marginal spines quite coarse. Leaves erect or spreading.
Fig. 17 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 18 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 19 7704 H. mirabilis ‘maraisii’. Bontebok Nat. Pk.Fig. 20 7704 H. mirabilis ‘maraisii’. Bontebok Nat.Pk.
8. MBB7705, 7706, 7713, 7714, 7715 Uitvlugt (Figs 21, 22, 23, 24, 25, & 26). These are five quite independent populations on the one farm that is unusual in that it seems to be on predominantly an earlier variant of Cenozoic ferricretes. The plants are generally related to H. ‘atrofusca’ with rounded leaftips and roughish surfaces. The actual habitats are all different in respect of surface detail and associated vegetation. The plants vary in size between the different populations from small (30mm diam) to large (70mm diam). In the one site where the soil is whitish bentonite, the plants are small and also have a curious similarity of color to that of the clay and the associated plant Gibbaeum haaglenii (austricola).
Fig. 21 7705 H. mirabilis ‘atrofusca’. Uitvlugt NW.Fig. 22 7706 H. mirabilis ‘atrofusca’. Uitvlugt.Fig. 23 7713 H. mirabilis ‘atrofusca’. Uitvlugt W.Fig. 24 7713 H. mirabilis cf ‘magnifica’. Uitvlugt W.Fig. 25 7714 H. mirabilis ‘atrofusca’. Uitvlugt E.Fig. 26 7715 H. mirabilis ‘atrofusca’. Uitvlugt N.
9. VA-RW07-27 (Figs 27, 28, 29 & 30). This population located by M Mougin, was reported to me by D. Van den Abbeele. It is much closer to Heidelberg and obviously in a ferricrete related habitat. The plants are all very ‘atrofusca‘-like with some evidence of the pointed leaf-tips that would have been ‘magnifica’. The surfaces have the slightly riffled texture that one associates with ‘atrofusca‘ rather than the tendency to roughish surfaces or almost spinescent tubercles associated with, but not peculiar only to ‘magnifica’.
Fig. 27 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 28 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. R Wellens.Fig. 29 VA-RW07-27 H. mirabilis cf ‘magnifica”. Basryhoogte. M.Mougin.Fig. 30 VA-RW07-27 H. mirabilis ‘atrofusca’. Basryhoogte. D. Van den Abbeele.
10. MBB7728, 7729 Lilliendal (Figs 31, 32, 33, 34 & 35). The significance of these two populations must be emphasized. The place is the Karringmelks River southwest of Heidelberg. This river drains into the Slangriver that flows then southwest into the Breede River. It is quite a different drainage basin to the Klip and Duiwenhoks rivers that drain the area north and west of Heidelberg and enters the sea south of Heidelberg. Matjestoon and the origin of H. ‘heidelbergensis var. toonensis’ is south of Lilliendal and on the same drainage system. The plants would at the time have made it difficult to imagine H. ‘heidelbergensis’ as a discrete system, and these plants force a direct comparison with H. ‘maraisii’, if that system could be generalized. The plants are small, dark-green, rough and with a degree of spination on the surface tubercles. Surfaces may be opaque or translucent. There is a striking resemblance of the seedlings to H. floribunda. The populations are about 1km apart. Both are on Bokkeveld but at the second population it is curious to observe that the plants are not on the ferricrete only meters away.
Fig. 31 7728 H. mirabilis. Lilliendal.Fig. 32 7728 H. mirabilis. Lilliendal.Fig. 33 7729 H. mirabilis. Lilliendal.Fig. 34 7729 H. mirabilis. Lilllendal.Fig. 35 7729 H. mirabilis. Lilliendal.
11. JDV87-3, MBB7165 Matjestoon(Figs 36, 37 & 38). This is one of the original populations known to G. G. Smith and it is an uncomfortable fit anywhere on account of the length and spiny-ness of the leaves. That the plants were placed in ‘heidelbergensis’ is a reflection of the doubtful and composite nature that the species gained by default. There is a semblance of similarity to H. rossouwii but what may be more notable is that at Dassieklip to the southeast there is a very aberrant population that is assigned to H. floribunda simply for want of a more likely category. I say this is notable because in my field observations of Haworthia there is a very strong and predictive geographic continuity between populations. This is evidenced in this general discussion.
Fig. 36 JDV87-3 ‘H. heidelbergensis var toonensis’. Matjestoon.Fig. 37 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.Fig. 38 7165 ‘H. heidelbergensis var toonensis’. Matjiestoon.
12. MBB7736 Koppies (Figs 39 & 40). These are small dark-green plants with strongly deflexed leaves and a little unlike an earlier record from 2km west (MBB6879 ex E. Esterhuizen) where the plants have more pronounced surface tubercles becoming quite robust and dark in cultivation. The habitat is among white quartz occasionally associated with ferricrete.
Fig. 39 7736 H. mirabilis. Koppies.Fig. 40 7736 H. mirabilis. Koppies.
13. MBB7723 Stuurmanskraal (Figs 41 & 42). These rank among what might be regarded as the general body of H. ‘maraisii’. Small, dark roughish plants with strong evidence of floribunda in the way the leaf-tips tend to be flat, twisted and rounded. The habitat is Bokkeveld shale.
Fig. 41 7723 H. mirabilis. Stuurmanskraal.Fig. 42 7723 H. mirabilis. Stuurmanskraal.
14. MBB7734 Stuurmanskraal N (Figs 43 & 44). This is a previously known population similar in its responses to the Koppies plants (MBB6879) that become quite robust and dark in cultivation. The habitat is white bentonitic clay that develops under the ferricrete sills from decomposure of underlying shale.
Fig. 43 7734 H. mirabilis. Stuurmanskraal.Fig. 44 7734 H. mirabilis. Stuurmanskraal.
15. MBB7731 Oudekraalkop (Figs 45, 46 & 47). Finding this population at all suggests that there may be many more evading the eye. The plants are small and dark and completely hidden among dense but short vegetation on a rocky scree-like area. As in other populations one is compelled to look at every plant and consider its departure from a hoped for norm for the plants there. Three illustrations hardly do justice to the variability and this true for all these populations discussed here.
Fig. 45 7731 H. mirabilis. Oudekraalkop.Fig. 46 7731 H. mirabilis. Oudekraalkop.Fig. 47 7731 H. mirabilis. Oudekraalkop.
Conclusions My despair over the years that I have been involved with Haworthia, is that the classification model and nomenclatural system we use, does not seem to allow me the freedom to express what I have seen and experienced. My argument has been that we are dealing with a biological system in which things that seem different are the same; and conversely things that seem the same are different. It is not a problem to apply a formal classification system except that a ranked or tiered system suggests quantum differences too, and this is simply not true. The greater problem is that most plant enthusiasts, and those who feel strongly about classification and names, confuse the reality of the sameness that suggests ‘species’ with the sameness that plants of different species may exhibit.
It is with a source of considerable curiosity that I now take note that a taxonomist is one who classifies by appearances and a systematist one who classifies by evolutionary pathways. In truth I never considered that classification was anything but an attempt to accomplish both aims so that meaningful names could be established. The issue is clouded in complexity when it is considered that neither taxonomist nor systematist may be sure when a species is a species. Probably hence the observation that species (binomials) carry a reality they do not possess. My proposal, supported by the observations in this article, is that we adopt a far more informal and flexible approach to the use of all the names we now have at our disposal. This suggestion is also supported by an experienced taxonomist/systematist/botanist who wrote: “The informal system that I prefer allows for endless forms that are identifiable by their locality and do not clog up the literature. Essentially it is a reversion to the polynomials of the pre-Linnaean era.”
My contention is that the only rational classification of the above 20 populations is within a greatly expanded concept of H. mirabilis that includes H. maraisii, H. magnifica, and H. heidelbergensis. This corroborates the observations reported in the chapters in Haworthia Updates and specifically Chapter 6, Update 2,1:50-79 “How to understand H. mutica var. nigra”, and Chapter 1, Update 3,1:1-87 “H. mirabilis and the interface with H. maraisii”.
Acknowledgements The population MBB7704 was shown to me by Ms. Tineke Kraaij of RSA National Parks Board. The concurring botanist is Dr John Manning, who has authored many botanical works and who is best known for his familiarity with the highly complex Cape bulb flora. Dr. Syd Rhamdani introduced me to the new direction that molecular biology is driving plant systematics. Gerhard Marx has been a great help in getting me to resign to the unrealities of popular taxonomy. Steven Hammer, considerably more successful than I have at adapting to the fact that our perceptions about many things are very different to that of others, helped me with the manuscript. Rhett Hiseman of Cape Nature, Jaap Viljoen, Henniie and Elma van Deventer of Koppies, the Odendaals of Dagbreek, Diamant and Dankbaar, Alfred and Andrea of Lilliendal, Dirk Papendorp of Uitvlugt, D. van As of Stuurmanskraal, Anna and Arno Steenkamp of Oudekraalskop (Anna’s Farm) are sincerely thanked for information and/or access to plants. ♦
I trust that it is clear by now that I consider H. magnifica, H, maraisii, and H. heidelbergensis to be essentially the same one species and emphasize that it is really a self-evident truth that species are complex systems and not simply a randomly occurring set of similar looking things. In a recent manuscript submitted to Haworthiad, I wrote about new finds elaborating this point of view. The essence of this chapter is to discuss exploration focused on clarifying the position in the eastern area between Riversdale and the very problematic H. pygmaea “squadron” that I have also discussed at length. Prior to the trip I had an ongoing communication with Gerhard Marx and we agreed that H. ‘splendens’ was in fact better fitted in H. mirabilis, the major obstacle for me being the fact that there was no field record nearer than Riversdale itself to substantiate such a view. I do, however, want to here also record two further populations east of the Breede River and other populations of H. mirabilis west of the Breede and south of anything previously noted.
1. MBB7442 H. mirabilis. Dankbaar. This population is of small plants but they are not substantially outside of anything noted in any of four compass directions although they do have their own look about them. I found it quite odd that Cameron McMaster had sent me pictures of a population of H. mirabilis from Fonteinskloof to the southwest of Stormsvlei that reflected the same co-similarities with H. mirabilis and H. marasii that I had recorded at Rondeheuwel south of Stormsvlei. The Dankbaar plants are small versions of this and of course tie up with both older and newer (MBB7704) records for the Bontebok National Park.
2. MBB7744 H. mirabilis. Bontebok Park. Having written that, we did in fact locate still another population and of course it looked different as the area where it occurs had been recently burned and being on a northwest aspect the plants were very exposed and even more cryptic than usual.
3. MBB7749 and MBB7751 H.mirabilis. Kadies Landing.: These two populations are in my opinion tremendously significant. While H. mirabilis has been reported from this the east bank of the Breede River, it was a very small form such as I recorded for the lower west bank (Ziekenhuis, Aalwee). The plants we located are down the river from Malgas but on the east bank. They are not any smaller than the general size of H. mrabilis anywhere in the central area of the range and highly similar to the plants especially in the Bontebok park quite some distance upstream from a very winding river course. There is thus now a very substantial connection and continuity from west to east and into ‘atrofusca’ that occupies the inland upper areas from west of Heidelberg to east of Riversdale.
4.MBB7753 H. mirabilis ‘toonensis’. Brakkekuil. It is important to note how I have named this as I have no intention of going through the formalities of nomenclature to change anything. I have been terrorized to my limits by “law” and I think that ethic and common sense can now prevail. ‘Toonensis’ is only known from Matjestoon southwest of Heidelberg and upstream along the Slang River. As I explained in the chapter on H. retusa, the Slang River has its own peculiarities and besides which it is quite unexplored. So it is no surprise to find this rather smaller version of H. mirabilis in a second population, differing from the general body by more and smaller more slender leaves. Naturally there is considerable variation within the population too. The differences relate to the problem of the Dassieklip and Vermaaklikheid areas where H. retusa and H. mirabilis ‘paradoxa’ and the problem should not be deferred to me as a bad taxonomist but as the reality that there is a major geological contributor to the problem of difference viz. shale vs calcrete. It is quite obvious in the series of pictures that one can see resemblances to ‘turgida’ and ‘paradoxa’.
5. MBB7761 H. mirabilis ‘splendens’? Plattekop Farm; MBB7762 do. W Platkop; MBB7763 do. E Plattekop Farm, MBB7765 do. NNW Platkop: MBB7769 do. Toringskop; MBB7770 do. W Soetmelksrivier. I am grouping these together and using the epither ‘splendens’ in an apparently loose and provocative fashion. This is to deliberately hew away at this mental sludge that isolates the really magnificent and incredible plants from their equally interesting but less glamorous close relatives. These populations are all on ferricrete inselbergs in virtually any of the three or four main geomorphological derivatives. The official geosciences description of the Riversdale area makes specific mention of the difference between these and the Soutpan (true ‘splendens’ habitat) by saying that the latter is a pan-area and the relatively thin ferricrete layer apparently developed in a marshy area and close to the surface (opposed to the inselbergs as deeper deposits in underground water systems in sandstone?). These populations are what we predicted and they are the direct link between the most easterly known only true H. mirabilis ’magnifica’ at the Frehse Reserve just south of Riversdale. The element ‘magnifica’ is a phantasm (available as an optional cultivar name as there are many plants in that parent population that do not fit the description?) based on an initial description and limited illustration and has been maintained by myself while trying to resolve the issues in my own mind. It has been kept alive for me by populations at KomseRante and Kruisrivier that I am about to further explore after writing this particular piece, and that will be a separate chapter.
I cannot say that I have resolved the entire complex puzzle but these populations validate the decision reached with Gerhard that it “feels” right to really include ‘splendens’ with H. mirabilis rather than with H. pygmaea. This is where the actual relation of H. retusa and ‘turgida’ takes on another dimension similar to the H. mutica situation to the west where it is confounded by H. mirabilis.
The most significant population is the last noted one viz MBB7770. Not only are the plants larger than in other populations they also manifest the general pattern in Haworthia of “flow”. They resemble true ‘splendens’ that much more than the other populations because they are also geographically closer. Another oddity is their occurrence in the pure ferricrete at the top of the inselberg that I discussed under H. retusa. The most extraordinary thing is that this population shares habitat with H. retusa and while I cannot say there are hybrids, it is evident from some of the plants that hybridization may well have occurred despite the big differences in flowering times. ♦
In a recent set of articles published by the Haworthia Society I wrote the following in connection with H. floribunda… “MBB7738 H. floribunda ‘major’. Swellendam: These plants were in fact small when first collected and in cultivation grew so large that I coined the name ‘major’ for them. They do still exist in a very small and disturbed area close to gum trees but curiously in moss free of leaf litter. I did also find them a little further away in a more grassy area where they are/were more typically small and dark coloured. I should note that I also recorded this ’dentata’-like version within the Bontebok Park close to where H. mirabilis occurs and I am still committed to again finding that population in the light of this new material”.
In connection with H. mirabilis, I wrote … ”The Dankbaar plants are small versions of this and of course tie up with both older and newer (MBB7704) records for the Bontebok National Park. 2. MBB7743 H. mirabilis. Bontebok Park: Having written that, we did in fact locate still another population and of course it looked different as the area where it occurs had been recently burned and being on a northwest aspect the plants were very exposed and even more cryptic than usual.”
Since those finds and comments, we have again been back to the Bontebok Park. It is of course a large area of largely recent geological origins and therefore mostly ferricrete and riverine gravels. The lower lying areas are seasonally flooded. Being unsuitable for cropping or grazing, the area was handed over to the conservation cause (thank goodness for non-arable land). H. venosa occurs on the exposed Bokkeveld Shale cliffs along the Breede River, and H. minima is present in small scattered populations. H. marginata has virtually disappeared from thePark.
It is quite difficult to identify suitable Haworthia habitats as even in the seasonally flooded areas there is much gravel and consequently good drainage. It was in such a site that I seemed to remember seeing the record for H. floribunda cited in my Revision. Our searching has been very limited as it is quite difficult to explore the park with the restraints of accompanying rangers however willing or enthusiastic and knowledgeable. This is because I personally need mental space and physical freedom to walk where my intuition takes me. On this occasion we first went to an old entrance to the Park from the town where we thought H. marginata was first recorded. I was very disappointed to find that the hillside there had suffered the same mindless engineering fate that hit the gravel hills around Worcester. These were ”strip-mined” to obtain road materials. Of course this may have been necessary, but not the mindless extent of the surface scraping when some consideration for simply good housekeeping would have limited the desecration of the landscape and demanded some degree of resortaion.
We moved some way west to avoid the disturbed area and eventually found H. mirabilis (see MBB7805 Figs 1) about 500m east of the MBB6513 record of 1996 when I was writing the Revision, and illustrated elsewhere. That record came from a more distinct interface with the riverine shale while the new record was in more sandstone gravels from an old river bank. The plants were rather smaller than in 7704 with quite slender leaves which were also fairly toothed. There was little evidence of the flattened rounded leaf tips that characterize H. floribunda. We drove around the park trying while I was wracking my memory banks for where I might have seen H. floribunda so many years ago and eventually was drawn to where Tineke Kraaij had on a previous visit shown us H. mirabilis on a burnt zone (see MBB7704 Figs 2). These are new images to the original published ones, because after 6 months the plants had largely lost the burnt look but also being in flower they were far more visible. The leaves were more elongate and pointed than suggested by the photographs we took on the first visit and the plants reminded me very strongly of what I named as H. floribunda var. dentata. I also add here images for H. mirabilis (see MBB7744 Figs 3) from another population approximately midway between the previous two. Here the area had also been burnt as part of the management plan for the Park, but the plants themselves do not seem to have been affected possibly because there was a lighter fuel load that at the 7704 locality at the time of burning. The plants were more “maraisoid” and similar to the very first record of mine for this species in the Park (MBB6513). That collection I often noted among a set of 4 or more populations extending from Riversdale westwards to Riviersonderend that confounded any distinction between H. magnifica, H. maraisii and H. mirabilis.
All things considered, it seems highly unlikely that H. floribunda could possibly co-exist with H. mirabilis other than in the present known configuration where we have H. floribunda var major close to and north of the N1 highway near the town and inland, whereas H. mirabilis is further south. This geographic and presumed taxonomic relationship seems to hold at Heidelberg and Riversdale with some anomaly at Albertinia down the Gouritz river. This is where H. mirabilis is absent (with consideration for the retusa/pygmaea conundrum) and where H. chlrorocantha or H. variegata may be confounded in H. floribunda (or vice versa!). ♦
If the name “H.enigma” applies to the plant (or plants) from east of Riversdale at Komserante, it is a name that I really do not advise to be taken seriously from a botanical point of view. It is useful at population level and to demonstrate the nature of classification difficulties but it is a minor problem in so far as those difficulties extend. The plants were first shown to me by J. Dekenah on the same day that he also showed me ”H. magnifica” in the Nature Reserve just south of Riversdale that is less than 3km away. My impression then was that it was the same element even if it did look a bit different. The plants are quite large (to 70mm diameter), fairly tubercled and often with lines in the upper retused area of the leaf face. While I originally classified “H. maraisii” under “H. magnifica”, I later separated them because it seemed so incongruous to include all the variants of the western “H. maraisii” with the few populations of “H. magnifica” then known. Also, as Essie Esterhuizen pointed out, “H atrofusca” as a variant of “H. magnifica”, seemed to be more dominant than had been realized. There were several other complications largely due to ignorance. Since my revision I have done so much more exploration and turned up so much new material that I have been forced to the conclusion that there is really one main element involved and that is H. mirabilis. This is where I believe the Komserante plants belong and the difference from the Nature Reserve population is due to a degree of infusion of H. retusa.
I revisited the site with Kobus Venter many years ago but did not look at a reported second population higher up the hill, taking it to be a little different based on plants I saw in Kobus’ collection. What was on my mind while we were recently exploring the area further east to examine the possible connection of H. mirabilis “magnifica” to “splendens” (and which we confirmed), was the fact I had never seen Kobus’ plants from Kruis Rivier northeast of Riversdale other than in Kobus’ collection. The plants I saw were also generally more robust than “H. magnifica” and more evenly tubercled. Kobus kindly took me to that Kruis River locality and much to my surprise the plants were in flower late October (see JDV92/65 Figs1). This is quite wrong for H. mirabilis, which is essentially a summer flowering species. I later went again to explore Komserante more thoroughly and to look at both the “magnifica” populations to which I believe the name “H. enigma” has been applied. The populations are in fact no more than 75m apart and cannot be considered to be genetically discrete at all (see MBB7778 Figs 2, and MBB7779 Figs 3). While it is true that the habitats are slightly different, this is reflected in the plants that at the upper slope of the hillside are vegetatively more robust and even clump forming, while those lower down in a bushier grassier habitat tend to be solitary and more withdrawn into the soil. These plants flower in summer and it is evident to me that there must have been some genetic exchange with H. retusa that grows approximately 200m away on the same hillside.
1. JDV92/65 H. mirabilis Kruisrivier.
2. MBB7778 H. mirabilis. Komserante.
3. MBB7779 H. mirabilis. Komserante.
Habitat preferences are very strict and it is curious to observe that there are four small buttresses forming the western slope of the low hillside. On the northern one we have the two H. mirabilis populations. On the second is H. retusa ’geraldii’ (see MBB7780 Figs 4) that is very clump forming, the third has no haworthias and on the fourth is a less clump forming H. retusa that is often referred to as ‘fouchei’ (see MBB7781 Figs 5) because the leaves are slightly longer and more upright than in ’geraldii’. This is not quite correct. Mr. Dekenah showed me what he took to be the true H. retusa ‘fouchei’, further south off the hillside and along the stream at Komserante. This habitat has since been cleared for farmland. The plants were very large and the leaves considerably more erect than is the case for the plants on the hillside. Curiously H. minima is present on the first and last buttresses. Flowering time for these H. retusa is spring but we did see vicarious flowering in summer so that definitely it is possible and probably a regular occurrence for out-of-season flowering to occur with the possibility of hybridization. I noted such hybrids between H. retusa and H. mirabilis in our exploration further east from Riversdale and I did think some of the clones in the Komserante plants evidenced this as well.
4. MBB7780 H. retusa. Komserante.
5. MBB7781 H. retusa cf. ‘foucherii’. Komserante.
The reality is that the Kruisriver and Komserante “magnifica” may flower at different times, but they are vegetatively very similar indeed. I consider that they belong in the same system as H. mirabilis and this conservative view of “species” as systems is the most sensible way to interpret and understand the plants. The approach can be taken further, as I tried to do in an earlier article concerning H. mutica ‘nigra’. In that article I dealt with a whole series of populations in and around Heidelberg, to show that there are three main elements viz. H. retusa (including H. turgida), H. mirabilis (including H. magnifica) and H. floribunda. I did not deal specifically with, nor resolve, the issue of H. mutica var nigra. I also noted populations along the lower Duiwenhoks river and suggested that H. floribunda is absorbed into H. mirabilis virtually south of the N2 east west highway, re-emerging on the northwestern corner of the Potberg in the south.
My most recent exploration was to check the flowering times of the original population of H. mutica var nigra at its origin at Kransriviermond and what I assumed to be the same thing on rather limited examination further north at Morning Star. Both these populations are along the Duiwenhoks river southeast of Heidelberg and in a continuum of four populations of H. retusa extending from just east of Heidelberg to still further east at an eastern Diepkloof locality (I think it has been referred to as Droekloof and Droerivier too, and the name ‘chromatica’ attached to the plants – see MBB7794 Figs 6). Less than 10km away is a population of H. retusa at Pienaarsriver (see MBB7776 figs in Chapter 3) that is very near to the whole array of populations of Kiewietskraal that I discussed in my writings about H. mutica var nigra. Two of those were distinctly H. retusa and so is the Pienaarsriver population that is different in that the plants are mostly quite dark in colour.
6. MBB7794 h. retusa ‘chromatica’. SE Heidelberg.
In those same writings I mentioned several populations of H. mirabilis along the Duiwenhoks river as well as populations of H. retusa var turgida. Here I am unashamedly relating “turgida” to “retusa” as I believe that when one calls all the populations now known to mind, as well as the variants they contain, this is the correct position of the plants in relation to a sensible and rational species concept. It is evident that H. floribunda is merged in H. mirabilis south of Heidelberg and the same thing occurs south of Swellendam. Thus I see no problem in now dealing with the population at Kransriviermond to which I applied the name H. mutica var nigra. I attached the varietal name to H. mutica because this is what G.G.Smith had done, and I believed that it was the link between H. retusa in the east and H. mutica in the west, when there were still huge gaps in the known distribution records for both species.
I have mentioned a population of ostensibly H. mutica var nigra at Morning Star, which is about 5km north of the Kransriviermond origin of the plants so named. This Morning Star population (see MBB7221 Figs 7) is very significant because while it includes virtually the same range of variants as the Kransriviermond plants, it flowers in summer as opposed to spring for Kransriviermond. Not only that, but it is 300meters away from a population of H. mirabilis (previously this would have been H. magnifica) that flowers and seeds at the same time. The habitat is marine plane ferricrete that overlies Bokkeveld shale but there are subtle differences between the two habitats.
7. MBB7221 H. retusa ‘nigra’. Morning Star.
The Kransriviermond plants (see MBB7804 Figs 8) are down in the Duiwenhoks valley in exposed Bokkeveld shale and I consider that the dark colour of the plants in the two populations is due to infusion of H. mirabilis. The rounded leaf tip would be derived from the “atrofuscoid” element of H. mirabilis while some plants also have the tuberculate roughness that that element often has.
8. MBB7804 H. retusa ‘nigra’. Kransriviermond.
Further south and west is the Slangrivier valley where there is also a suite of populations that includes smaller forms of H. mirabilis (H. heidelbergensis var. toonensis) as well a H. retusa var turgida also in an unusual form that supports the taxonomic position I now adopt.
The conclusion is that the Morning Star and Kransriviermond ‘nigra’ can be regarded as H. retusa within the same broader concept that includes ‘turgida’. The problem now arises from still another new discovery. This is from along the Buffeljags River (see MBB7801 Figs 9) about 10km southeast of Swellendam and 12km from the nearest obvious H. retusa at Goedverwagting further east; and from the less obvious H. retusa (that I have elsewhere noted as H. mirabilis) south of the Tradouw Pass. The nearest H. mutica are within about 15km to the west at Napky and Luiperdskop. The plants are the now customary “variable”. The leaf tips are very rounded. Some plain, some lined, some with curious submersed pinkish dots. The surfaces can be quite smoothish or else the surface cells terminate in minute spines that give the surfaces a glistening appearance – evident in H. mutica too. These plants flower in summer whereas both H. mutica and H. retusa are spring flowering. Note needs to be taken of the habitat. It is an extensive and old river gravel embankment and terrace, and therefore another variant on the interface of recent ferricrete deposits and the older Bokkeveld shales. The vegetation is grassy Karooid bushveld, unlike the dry fynbos to the northwest or the renosterveld (mainly cultivated) to the east.
Kobus Venter saw the photographs and suggested that the plants might be H. mirabilis’ and reminded him of ‘atrofusca’, perhaps also by virtue of the flowering time. This is of course what my “enigma” is all about. It is compounded in this case because these Buffeljags ‘muticas’ are rather odd in that the leaf ends tend to be flattened almost as though H. floribunda has also entered the fray. Kobus was much happier with the H. mutica connection when he saw the plants in the field.
The story here closes with H. mutica MBB7741 from Dankbaar about 12km west of the Breede River bridge at Swellendam. The plants here also have very glistening surfaces and thus seem to confirm the very original perception of G.G. Smith’s that the Duiwenhoks plants and the plants west of the Breede River were connected. It is certain that he never envisaged the complexity of the interrelationships between three main elements viz. H. mirabilis, H. floribunda and H. retusa.
Footnote. It is just assumed that perhaps the name “H. enigma” has been used for the Komserante H. mirabilis, because that population is indeed enigmatic as this article explains. Gerhard Marx informs that “enigma” applies to a H. mirabilis population further west where the more atrofusca variants occur. Since writing this chapter yet another similar population to the Kruis River and Komserante populations has been found about 15km SE Riversdale. ♦
This chapter is based on recent field exploration and embroiders around many aspects of Haworthia species discussed in earlier chapters. What should be striking is that new populations follow the very predictable geographic pattern that all my earlier exploration has exposed and in my estimation confirm in every way what I consider a sound and satisfactory taxonomic solution and help explain its limitations.
1. Haworthia minima.
Two new populations of this widespread species are recorded:
MBB7807 Swartrug, SE Heidelberg. This is a steepish east facing slope. The plants are small, the leaf length barely exceeding 50mm and with a tendency to form clusters in the shallow clay banks that rift the area. There are many plants in quite a small area and it curious to see them within very large clumps of Euphorbia tridentata. This Euphorbia itself has a strange geographic distribution being known at Riversdale, Calitzdorp and then at Cradock in the Eastern Cape. (see Figs 1)
MBB7813 NW Skeiding, E Tradouw Pass. This is a high rainfall area and we were expecting to find H. mirabilis in the lower pressure bursts and white clay that can be seen from the farmhouse far away. But instead we came across H. minima at the highest point of the hill among sparse grass. They are rather green in colour and it is interesting to note in the one plant pictured that it has non-tuberculate upper leaf surfaces and white cartilaginous margins and keel of the leaves. (see Figs 2)
2. Haworthia rossouwii.
MBB7803b Morning Star, S Heidelberg. Morning Star was known to J. Dekenah but the area has been very developed since those years. It was very surprising to find this small form in abundance at the top of a high hill in ferricrete pebbles and virtually no vegetation ground cover. However on this subsequent visit we crossed through an internal farm fence and found the plants in abundance about 150m further away and over quite a large area. We later looked at a lower promising hillside and small area nearer the Morning Star homestead and found the plants again. (see Figs 3)
Looking from Morning Star to Kransrivermond 097
3. Haworthia mirabilis.
This item reports on 6 populations, three of which are new. They confirm the continuity down the Duiwenhoks River and also southwards and westwards from east of Riversdale through to west of Robertson and Greyton and Napier to the south of that. Attention is drawn to south of Heidelberg where the ‘enigma’tic relationship of this species with H. retusa is suspected.
MBB7808 Swartrug, SE Heidelberg. This farm is well known as the source of ‘Haworthia chromatica’ that I dismissed as an element in the complex surrounding the e H. retusa (mutica) ‘nigra’. The names Droekloof and Doringkloof have been used while the map reference gives Diepkloof for a large area extending to west of the Duiwenhoks River. The plants are on an east facing boulder terrace in a stony low bush site with some grass. The plants are small and show the expected wide range of variation. They are fairly similar to the plants in the lichen patch just east of Heidelberg that I have always mistakenly taken to be the origins of H. heidelbergensis, but here are subtle differences that mark a population difference, but I will come back to this. (see Figs 4)
Looking from Swartrug towards Riversdale.
MBB7809 Koeisekop, SE Heidelberg. This is on the same farm and a similar ferricrete hillside southwards. There are several hilltops by the same name in the general area and I cannot even speculate on the origins of the name (“cowshead”). The plants are larger than in the previous population and very abundant. The similarity to populations in the Swellendam and Riviersonderend area is in my opinion striking. Odd clones are miniature images of H.mirabilis from the southwestern populations west of Napier. (see Figs 5)
MBB7220 Morning Star, S Heidelberg. I have discussed this population before and I repeat it here because I did not have many field photographs. It is only 300m from the Morning Star MBB7221 H. retusa ‘nigra’’ population and they flower and seed virtually in unison. Therefore they probably also reflect on the strange interaction between the two major systems. What I have surmised is that H. floribunda gets absorbed into H. mirabilis at the southern limits of its distribution and in these pictures there is clear evidence of the rounded and flattened leaf tips of that species. Therefore we may actually here have a three way species interaction. Reinforced by the presence of three different things at Kransriviermond, covered later under 4. H.retusa ‘turgida’. (see Figs 6a and b)
MBB7221 Morning Star
MBB7811Kransriviermond, S Heidelberg. Populations further down the Duiwenhoks river have been reported on and this one fall within that mould and the same one as the previous Swartrug and Koeisekop populations. The population is smaller and the plants too are quite small. (see Figs 7)
MBB7814 Skeiding, W Heidelberg. This area is also reported on but digital photography makes it so much easier to record the enormous variability that such populations exhibit. Some of the plants are typically ‘atrofusca’ while others have leaves ended in an awn. Leaf surface may be smooth and there is even an indication of smooth leaves. What is notable is the absence of H. floribunda from what is the sort of habitat it can be associated with and can one can speculate its absorption into H. mirabilis with leaf shapes that demonstrate this. H. floribunda does appear again about 15km to the southwest in association with H. retusa and in the absence of H. mirabilis. (see Figs 8)
The Skeiding homestead
MBB7233 Die Plotte, Heidelberg. “H. heidelbergensis” correctly has its origin in this near vicinity and these small summer flowering elements are certainly part of a large assemblage of populations in the central area of what I now come to see as all H. mirabilis. Here discussion must pass to the following. (see Figs 9)
4. Haworthia retusa‘turgida’.
A difficulty that I experience is in the need to dismiss the impression and general idea we have that there are tiered ranks. H. retusa has many populations and past treatment has placed most of the variants in H. turgida. To suggest that these are really two separate systems is not realistic and confounds any explanation for a few anomalies that exist.
MBB7810 Tierkloof, SE Heidelberg. Tierkloof is quite a deep valley that pushes out of the Duiwenhoks to the east. We found this huge population of large clump forming plants and several such populations are known just north of Heidelberg itself. I have applied the epithet ‘longebracteata’ to these and this is no more satisfactory than generating names for each of a very wide range of populations. There are populations along the Duiwenhoks that are far more inclined to the ‘turgida’ element and Essie Esterhuizen wrote a very good account of the host of variable populations that he encountered. (see Figs 10)
H. retusa ‘turgida’ at Tierkloof, Swartrug
MBB7812 Kransriviermond, S Heidelberg. We only saw three plants at this locality and these plant mirror the preceding, if a bit smaller. The problem here is that we also have the element I myself described and named as H. retusa ‘mutica’ and I originally linked this “variety” with H.. mutica following what G.G. Smith had done and for predictive reasons. The fact that ‘turgida’ now occurs with it is a huge hurdle to cross. Think carefully. (see Figs 11)
MBB7804 Kransriviermond.I have written about his population in the chapter “Haworthia enigma”. This explains that the plants are not all leaden-black as perceived. They are spring flowering – as ‘turgida’ is. However, the range of forms and colour virtually replicates the plants in MBB7721 at Morning Star (pictures in the “enigma” chapter) where they are spring flowering. Across the river from both these populations is a weak population of small plants of H. mirabilis (7811 above) summer flowering. We know that this major difference in flowering season is not a total barrier to hybridization. (see Figs 12)
MBB7234 Die Plotte, Heidelberg. This is a population of large clump forming plants on a north slope about 300m south of MBB7233 H. mirabilis ‘heidelbergensis’. It is spring flowering. When I first saw these plants I dismissed (actually I never dismiss anything – I maintain reservations as I do now) these plants as large variants of ‘heidelbergensis’, and actually collected one magnificent specimen that looked to me just like H. mirabilis, say, from west Napier. Looking at the range of variation one is compelled to concede that here again is an infusion of H. mirabilis into a population that is primarily H. retusa in that vast shadowland it casts with ‘turgida’. (see Figs 13)
Conservation I add these comments for several reasons surrounding the poor image that collectors have in the eyes of conservationists, and the very rickety fence I therefore occupy. There is a note reprinted in a recent issue of the newsletter To the Point. This taken from Times magazine and seems to be a rather hyped up condemnation of widespread destruction based on a view of internet websites. These are also monitored by RSA law enforcement that is rightfully disturbed by the high number of locality and collector citations that are indicative of illegal collecting. I personally no longer apply for a “collecting” permit for the prime reason that I am concentrating on photographic record, as a herbarium record no longer serves any better practical short term purpose. The herbaria are overburdened and I prefer living plants to dead ones. After 70 years of field activity I feel treated like a pariah and reduced to subservience under less-informed inexperienced people operating under a rigid and oppressive legislation aimed at acquiring a prosecution rather than wisely trying to direct activity in a positive direction. This activity of collection is, and I have no doubt about this either, a human right imbedded in the very reason for creation and our place in it. Conservation has its roots in the passion that people have for nature and it is nurtured by experience, contact and knowledge of its varied components. It is just ridiculous that one can go to any animal or plant centre here in South Africa and see plants and animals from anywhere else in the world and none indigenous. It is Ok to, say, keep Madagascan Chameleons in captivity where it is someone else’ responsibility to secure them. But keeping local ones is taboo? It is OK to freely sell cacti that do not occur here but are now being dug out in tons in (if that is really true). Recently it has become illegal to so much as possess Encephalartos latifrons and even the National Botanic gardens can no longer propagate and sell this plant. While we talk “plant indigenous” on all public fronts, the fact is that the acquisition, cultivation and sale of indigenous plants is viewed in the most negative light – a viewpoint encouraged by conservators.
This is control by law, a large degree of pure ignorance and by improper application of power derived from legislation. This is not the way to go. It is quite evident from the Times magazine article that legislation has the negative effect of creating a false value and glamour to discovery, Latin names, acquisition and possession. I believe that knowledge and understanding is true power, not legislation and enforcement.
The collecting numbers, localities and information provided here is to promote understanding and knowledge of plants, and thereby to promote their continued existence against all the vagaries of nature that “threaten” them, least of it which may be actually human greed.
Acknowledgement I am glad to acknowledge the company, comments and interest of Gerhard Marx and whose organization sceptisism I also value. Landowners always impress me with their generosity and graciousness in allowing us access. On this occasion they include Mr. and Mrs. Nelie and Wimpie Jacobs of Diepkloof, Mr. Cloete and Belinda Fourie of Melkboom (B&B), Mr. and Mrs. Coetzee and Sarita Uys of Morning Star, Messrs. Christo and Pieter van Deventer of Doornvlei, Mr. and Mrs. Neels and Anne-Lise Uys of Skeiding (B&B) and then Ms. Jennifer Steggie of Heidelberg. In view of the comments on conservation noted above, I note that there are several farms that now offer accommodation and an outdoors experience and the opportunity to view these plants in their extraordinary natural surroundings. ♦
Alsterworthia produced a special edition (No.7) in 2004 to publish new species and combinations subsequent to the publication of Haworthia Revisited. I was given a copy because of my own contributions in respect of primarily new combinations. I had the previous year done some exploration along the Duiwenhoks River south of Heidelberg and found several Haworthia populations notably MBB7227 Witheuwel and MBB7229 Somona. I discussed these in Chapter 6 of my Update Vol 2. dealing with the complexity of the element H. retusa (mutica) var. nigra and the problematic nature of H. mirabilis as it occurs around and south of Heidelberg. So when I saw the picture of H. jakubii I merely glanced at the description to see the words Duiwenhoks River to think this was another of those weird armchair products to befuddle the enthusiast and add another name from an endless production belt. There was nothing about the illustration that suggested anything new to me so it is really fascinating to now only read what the author had to say “When the author first saw them, he thought they were something new because of their features.” This is a very subjective statement and I have no doubt that the author could be misled into thinking that other plants from the same population could also be “something new”. Why “something new” should be allied to a Latin binomial is intrinsic to “namenklutter” and the disrepute into which taxonomy has fallen.
I must dwell on that statement a little because it helps to demonstrate, falling very far short of explaining, why plant classification becomes such a hopeless mess. In 1986 I wrote an article published in Excelsa 12:91 entitled “Confusion reigns again” where I stated that it had become seemingly impossible to write about Haworthia without casting doubts on one’s own, or someone else’s, integrity or sanity. This was because of the apparent lack in the literature of any sense of discrimination and recognition of what might be right or wrong about any written piece. The more painful aspect of course is that it meant that I could write nothing myself that would be believable. I have to state that since 1986, the situation has not improved one iota and I regret that I am not tactless, aggressive or unkind enough to list a set of prominent people, including botanists, who have done nothing more than fuel the fire of enthusiastic writers. So we have “because of their features”. What features? I presume these same unlisted features are what made me sigh and think “Oh dear! – another name for H. mirabilis”.
I did not give the element anymore thought until I happened to see Jakub Jilemicky’s website and e-mailed him to query the principles he was adopting for recognition of “species”. From him I learned that the origin of “jakubii” was in fact the Goukou (ex Kafferkuils) River north of Stilbaai and Jakub kindly also sent me map details. Because there are virtually no records of H. mirabilis further south east of Riversdale than in the Frehse reserve close to town, I suspected that perhaps we had to do with a H. retusa ‘turgida’ variant or possibly a hybrid between that and H. variegata. While not generally prone to concede oddities to hybridization, I have become much more circumspect in this respect after learning a great deal more about hybridization between species despite big discrepancies in flowering time. Jakub’s map was a bit problematic because the designated spot was quite far from the Goukou River and in the middle of cultivated land. Furthermore the farm name Melklhoutkraal is cited and the boundary lay east of the designated spot. However, my wife and I came to the designated place where a road serves two farms south of the road. The first farm deserves special mention in respect of the negative aspects of agriculture where an over zealous new owner has decimated the landscape with no consideration for conservation whatsoever. The invasive Acacia cyclops (Rooikrans) has been bulldozed away and what may have been cultivated before certainly is now, and this also extends into that fragile zone between the productive shales and the non-productive calcrete. The owner initially offered, being busy at the time, to show us a hill that was being “conserved” and we made an appointment to do so. However, when we later arrived back we were informed that here had been a change of mind and we would not be allowed there. The reason will become apparent.
We went on to the second farm where the owners were delightful and allowed us to explore at will. The farm borders the river, but we stayed away from that as we were fairly sure that H. retusa ‘turgida’ would be all there was to see on the steep banks. So we explored the calcrete that is so reminiscent of H. paradoxa and Vermaaklikheid. We found a small and very cryptic population of H. variegata that rather strengthened our suspicion that perhaps ‘jakubii’ was of hybrid origin. Then we went further north to continue some exploration we had started there and we found a strong population of H. mirabilis about 15km SE Riversdale (MBB7818 Windsor – figs 8-10). We connected this population to plants at KomseRante east of Riversdale and also Kruisriver further east and north and suggested the possibility that ‘jakubii’ was then a Duiwenhoks River image of H. mirabilis.
Back home and back onto the internet. Jakub sent a better description of the locality much nearer to the river and on the farm Klipfontein rather than Melkhoutkraal. An error and something I might have worked out had I dug out the publication and recognized that the stated geographical facts were as unreliable as the taxonomic claims. Kobus Venter was thankfully able and anxious to get into the field in connection with other finds, and he kindly offered to drive all the way to Stilbaai to follow newest information. Gerhard Marx was also with us as a most welcome guest, and we then found the evasive ‘jakubii’ growing in a very narrow band right alongside the road and extremely vulnerable to any road works (see MBB7820). Our over zealous farmer had not only erected a new fence and deposited all the Acacia branches on the road reserve, but had also concentrated stock on the river bank on the farm side of the fence. The result was what is to be expected when there is no sympathy for the environment or for the stock. Animals under pressure will eat anything and everything and the concept of selective grazing and suitable stocking rate can fall away. One may as well cultivate the soil for the devastation too many animals in too small an area for too long causes. It is fortunate then that we have the road reserve. We found about 40 plants that appeared from the dry capsules and stalks to have flowered about 6 weeks previously in early February in close synchrony with H. variegata nearby and also with the Windsor population of H. mirabilis. We took Kobus and Gerhard to see the Windsor plants and they confirmed the similarity of individuals there to ‘jakubii’. Among Figures 8 are plants that demonstrate this similarities but also the wide range of variation so evident in most populations of H. mirabilis, thus H. mirabilis ‘jakubii’- a significant population and well worthy of a name outside of formal botany, which brings me back to my near opening remarks. The formal nomenclatural system works very well as no doubt does the horticultural code. However, there are serious limitations that severely affect Haworthia. The first is that few people actually have much experience with plants in the field and the true nature of natural variation. There are too few field-familiar competent taxonomists and the whole field of taxonomy is seriously flawed by any form of peer review outside of the nomenclatural rules and procedures. Another serious flaw is simply the absence of any general and common understanding of what a Latin binomial is actually supposed to be appended to. The same is true of lesser taxonomic ranks. Gordon Rowley many years asked (of me) “I would love to know what is to become of the luckless hybrids and cultivars if they are to be deprived of the rank of genus and species.” I am now not sure if he thought through that question very well but I understand what he means and in recent literature he has highlighted the value of names for the sphere of horticulture in which “collectors’ operate. Short of microchipping, many cultivated plants are going to be deprived of names simply through loss of physical labels. I am not sure, and underlined by that very curious statement about the features that made ‘jakubii’ into something new, that any formal system of cultivar recognition is going to work in Haworthia. The problem is that we are not dealing with flower characteristics and colours of virtually a single species as happens in most horticultural groups. Nor are we dealing with any controlled breeding program. We are dealing with vegetative characters of perhaps 30 species where there is much evidence of gross misidentifications by experienced professionals (botanists) at species level. Also there are any numbers of potential new cultivars by virtue of the simple facts of natural variation. How are we going to ever arrive at descriptive matter and even illustrations that will allow correct identification when growing conditions and season influence the vegetative characters of the plants concerned and we cannot even be sure to what species they belong.
The author of ‘jakubii’ does end by saying that the taxon is intermediate between H. mirabilis ‘paradoxa’ and H. mirabilis ‘magnifica’, listing shape, surface, coloration and solitary growth as reasons. This may seem reasonable on the surface of things. But it is totally unreasonable if one considers that one could take representatives from the same and different populations of these and use the same argument to justify many more names within those two elements as variable components of a much greater cohort of variants. The description itself simply cannot be taken literally. A plant is illustrated with leaves having from 3-9 face lines however one chooses to count them, while the description states 5-7. The colour is given as grass-green (in poor light), but we saw most plants in semi-shaded and poor light microhabitats and none were “grass-green”. The leaf surfaces are also described as “smooth” and this was not evident in the plants we saw. The converse is true and also in respect of either ‘paradoxa’ or ‘magnifica’. In fact the statement underscores the (poetic) license that is taken with descriptions that end up unworkable. In H. mirabilis, the leaves may indeed be smooth (e.g. ‘badia’) but in any population there are plants that have incipiently rough surfaces extending to prominent tubercles and even prominent hair-like spines. So, indeed, if Latin names are said to have a reality they do not in fact possess, this certainly is the case here.
Acknowledgement. Dr. an Mrs. Lindley Kok of Klipfontein, Ms.Sonya Walmsley of Melkhoutkraal (?), Mr. Johannes van Eeden of Windsor. Kobus Venter and Gerhard Marx, for participation and company. I particularly appreciate the interest and assistance of Jakub Jilemicky in resolving an issue his name is associated with. ♦
7818 Windsor 56647818 Windsor 56697818 Windsor 56777818 Windsor 5688Fig. 4a. MBB7820 H. mirabilis ‘jakubii’. Goukou Bridge, Klipfontein. 5746Fig. 4b. MBB7820 H. mirabilis ‘jakubii’. Goukou Bridge 5750Fig. 4c. MBB7820 H. mirabilis ‘jakubii’. Goukou 5752Fig. 4d. MBB7820 H. mirabilis. Goukou 5755Fig. 4e. MBB7820 H. mirabilis. G
My experience with Haworthia dates back to my childhood and on to nearly 70 years of observation. However, my interest was only able to properly manifest when I began work at the Karoo Botanic Garden in 1969 and it has since been through many phases. I wrote a formal taxonomic revision of the genus in 1999 and have spent a good bit of the last nine years adding to and verifying what I wrote. Haworthia has always been regarded as a problem child of botany to be avoided by professional taxonomists for various reasons including an apparent phobia of the many amateur collectors peering over the shoulder while at work. This has puzzled me because it seemed to me that if the need for good classification and identification was so strong there was an obligation on botany to provide the service. So my involvement has been largely by default. I was trained in an agricultural and entomological tradition with a totally different and unsophisticated approach to things like taxonomy, systematics and nomenclature. In the infant science that agriculture then was in South Africa, I can barely claim that my MSc is much more than an indication that I tried to learn something beyond normal schooling. While trained as an agricultural entomologist, my leaning was to plants and I eventually came to the Karoo garden to do what I liked best viz. exploring plants. Unfortunately the route is via identification and names and so I have walked a long road through the minefield that this is. Was this only in respect of Haworthia? No! This is a persistent misconception. Haworthia is only different because it has attracted such close and sustained amateur interest by so many for so long. I experienced failing classification in many other genera. To be fair I think the real reason is the lack of importance attached to the whole function of plant classification. It even seems as if many modern botanists pursue the study of plant relationship under the guise of systematics that is not committed to providing formal names and identifications.
The problem in the genus Haworthia begins with weak botany from the moment Linnaeus decided that there were four small plants that belonged together as one species in the genus Aloe. This alone generated a nomenclatural problem the dust of which is still to settle. A worse and obscured fact is that because of their small flowers these plants were and are still treated as one genus – one set. This is not true. The allied genera of the Alooideae of the Asphodelaceae are not comfortably classified and the three sets of plants that constitute the genus Haworthia are not only florally distinctive, they are also phytogeographically and behaviorally different. Thus while there was or is this huge flaw at this classification level, it is actually bizarre that sense can ever be thought to have reigned at the species level. The proliferation and confusion of names coupled with the associated argumentation about names has even led me to doubt my sanity, or even lose it as my critics will happily claim. My impression now as a non-botanist is that taxonomic botany somehow lost sight of the requirement that a biological classification should mirror and reflect the evolutionary processes or phylogeny that underlies our view of species and their origins. Worse, is the actual absence of a clear and unsullied definition of what a species is and hence what a Latin binomial as a supposedly formal scientific term of reference actually means. Cladistic methodology in plant taxonomy is in my opinion nothing more than a belated and even misguided attempt to arrive at or return to some semblance of scientific truth. Molecular biology is now being desperately seen as a holy grail that will summarily correct reluctantly conceded failures of the past.
There is no doubt that the mess in the classification of Haworthia is the product of non-botanists, but it is absolutely incorrect to exculpate the contributory role of taxonomic botany and professional botanists who are or were taxonomists. My interest has by no means been confined to Haworthia and the need for names has largely been driven by a perpetual curiosity about many different plants in the field environment. I am, by the route my life has taken me, unusually and almost uniquely experienced in the relation between botany, botanists and interest of the layman in plants, as it is reflected in the names we use for them and how we respectively do so. Consequently I think that I am now qualified to express some opinions on what Haworthia can teach us quite apart from what the hostilities that wage across the whole taxonomic front suggest.
I pointed out rather timorously that in Oxalis, morphological characters were not as indicative of species as revisions and identification keys generally seem to imply. The dramatic changes in our classification systems that the results of molecular biology now seem to threaten, is a manifestation of this simple fact rather than any great new understanding of what really goes on in the plant kingdom. My fear is that molecular biology is likely to fudge most of its promise because the basic tenets of classification, the mistakes of the past and the reality of the field are still not properly appreciated. This is what I think Haworthia has to tell us. I made use of Oxalis to test my observations that we can look at Haworthia as composed of elements that reflect the very nature of the habitats and environments where they are found. I did this because Haworthia plants simply do not have any array of characters by which species can be arrived at whereas Oxalis has a plethora of such apparent (taxonomically bewitching, tempting, and misleading) means. While the few facts that there are in respect of differentiating Haworthia into three groups (subgenera as presently constituted) has been lost and confounded in the broader classification of the Alooideae there is not much else to help verbally to define the species. It is not to say that modern technology has not been resorted to, to help clarify the issue. Both cladistic and molecular studies have been done but the results flawed by weak hypothesis and perhaps just weak science too.
How then do I think sense can be made of Haworthia? It is firstly by defining the species as dynamic fractal systems that illustrate the responses of plants to skeletal habitats and geological substrate in the wider context of landscape and topography across the total distribution area. Furthermore, there is an obvious correlation to be made with vegetation and other biological systems both plant and animal. I cannot pretend to know a great deal about any of these things, but also suggest that nothing more is needed other than awareness that this is what biological diversity and species are essentially about anyway.
It has taken me nearly 40 years to arrive at the big picture of Haworthia. It is as a non-taxonomist that I say categorically that the three main elements in the present genus need to be seen and registered as “different”. Then we can focus on the subgenus Haworthia where the greater difficulty lies and which I believe offers the greatest insight into what plant species are and what the difficulties are in recognizing and defining them. Also into what Latin binomials should or might mean.
For this I am going to take just two species systems and ignore some of the peripheral facts that might confuse the issue rather than illustrate the profound nature of species interactions. I take H. retusa (Figures 1 to 4) and H. mirabilis (Figures 5 to 10). In order to do this I have to bypass even my own conservative classification by considering H. turgida and its many variants to be the riverine and cliff dwelling version of H. retusa. H. mirabilis is more complex and I have to include three other species viz H. maraisii, H. magnifica and H. heidelbergensis for which spatial factors and major geological substrate are the prime differentiators. Both my two selected species are highly variable both within and between populations. It is in fact the intra-population variability that I underestimated. Growing plants from field collected seed can result in a hundred or more individual plants that are all different in appearance.
The two species share the same distribution range, but with an extension of H. mirabilis into the Worcester/Robertson Karoo, while H. retusa extends further east from Albertinia to Great Brak. H. mirabilis flowers in late summer and H. retusa flowers in spring. They never grow in direct association! The populations of each are highly localized and when in proximity are usually separated by hundreds of meters, and difficult to ascribe to habitat conditions given the ranges of habitat that respective populations may occupy. I have already noted and warned of the reality that there is evidence of interaction between these two clearly defined systems. In addition, somehow or other, the species H. pygmaea emerges as discrete in the eastern Mossel bay area, while H. mutica sits in an island between Swellendam and Riviersonderend in the west.
What is most striking is the variability within each system and this is where the crunch comes and what my pictures should illustrate. They are but a small sample of many hundreds and I have adopted an informal method of captioning the pictures as the formalities of nomenclature would need to be followed to depart as I have from the existing status. Not only are my pictures supposed to illustrate the problem of variability within species, figures 2 and 3 show variability within a population and figures 3 and 10 show two very similar plants from two quite different species systems. Note that the plant in figure 3 is in flower and that the similarity to figure 10 is rather exaggerated by photograph scale as in H. mirabilis ‘toonensis’ the plants are smaller.
Taken individually from the morphological extremes between and even often within populations, it will stretch the credibility of the most ardent taxonomic lumper to claim that any two of these are representatives of the same species. It is an unpalatable truth that things that look different are the same just as the converse is that things that look the same are different. When this reality is properly conceded by the professional taxonomist, perhaps we will succeed in demystifying and deglamorising the Latin binomial. Botanists who are not taxonomists will arrive at a better understanding of their plant world and consequently we who look to them for understanding will be better served. We need to be more realistic about plant description and identification and also correspondingly more conscious of the real nature of diversity. Conservation is not to be seen as maintaining a list of names for which there is a use. It is to be seen as maintaining the options for change and the conservation of a far greater diversity than any list of names can hold. It is botanists (quoted in Levin, 2000) who wrote “our system of names appears to achieve a reality which it does not in fact actually possess”. I think this implies that even the Latin binomials we so modestly, reluctantly or even portentously use do not convey what they should; but rather the product of our individual limited experience, knowledge and perception. My feeling is that species do have a reality and for Latin names to have any sense, they need to be more circumspectly applied. There is little merit in having a highly regulatory nomenclatural code and little value in what the names actually refer to. ♦
I think we have to find a way to deal with this issue of names. Let us try H. magnifica. It originated in a population from the Frehse Reserve SE Riversdale and it is truly difficult to circumscribe all those individual variants. The name has also been attached to a number of other plants and populations from various places. The name magnifica is not clearly assignable beyond individuals that can be said to resemble the type (an illustration) and there are individual plants in the Frehse population that do not accord with either picture or description. There are individual plants and populations going all the way to near Caledon that confound the name still further. In my opinion the Frehse reserve plants belong to a single system that I consider to be H. mirabilis (and I am not so sure that it is not bigger still). Do we just drop the name ‘magnifica’ and use locality? So is it better to say H. mirabilis magnifica (Frehse Reserve) and H. mirabilis magnifica (3km S Riversdale) or H. mirabilis magnifica (Windsor) for the variants that occur at each of those places? Also H. mirabilis jakubii (Goukou) that I think is a connection between H. mirabilis magnifica and H. mirabilis paradoxa (Vermaaklikheid and on to Infanta). We could use this system and also the Breuer and Hayashi names attached to other mirabilis populations in the Riversdale area. The disadvantage of place names is that they convey nothing to people unfamiliar with local geography and to them there may be no difference. On the other hand that the formal Latin names may be restricted by the accompanying description and illustration, and not convey the variations that occur in the various populations.
I have become increasingly concerned about the poor relations that exist between collectors and the authority of Nature Conservation. The argument that collectors threaten and despoil natural populations is very real and I do not dispute at all that Conservation authorities have a very valid complaint. They have a function to perform. On the other hand there is an interaction between human beings and nature in all its forms that should be fostered to the benefit of both sides.
Nurseries, traders and collectors are as much of the picture as are conservationists, institutions, researchers and landowners. It is unfortunate that there is no non-government party that lobbies for the rights and activities of the former group, but it is not my intention nor within my competence to argue all the aspects of the case.
I strongly believe that people have the right of access to nature in all its forms and the issue is one of individual responsibility and proper consideration of consequences. An appreciation of and sensitivity to nature should be reflected in whatever we do in our lives. My own collecting impulses led me to institutional employment where I could exercise my interest to what I thought were efforts more worthy than my personal interests. From that position I also did try to share and extend privileges to a wider circle. It is in this way that I became involved with Sheilam Nursery. It was not my wish or intention that my collection should have come to be housed there. However, Sheilam has succeeded over a period of nearly 40 years to maintain a fairly true record of my collections obtained as propagated material from the Karoo Garden at Worcester. My offer of permitted collections dating from my revision of Haworthia in 1966 to the Karoo Garden was rejected and for a while resided with Etwin Aslander at Brackenfell. It has since passed to Garth Schwegman at Sheilam who has taken a particular interest in the maintenance and propagation of that collection.
Hand pollination of haworthia has become the norm and Sheilam has made a massive effort to generate plants from this. Of course growing plants from seed is a lengthy process, but is richly rewarded with an astonishing array of variants and beautiful plants.
Need I say how important this activity can be for an understanding of the plants and the way this is expressed in a classification? A good example is of the hybridization found between H. lockwoodii and H. arachnoidea ‘scabrispina’ at the Floriskraal Dam east of Laingsburg (see figs 1-3). The hybrid is very reminiscent of plants that I have seen from between there and Prince Albert; and also southeast of the Rooinek Pass through the Witteberg, and again northeast of Calitzdorp. Roy Mottram underscored the issue of hybridization in Haworthia classification. I have not ignored the issue, but rather viewed it as an imponderable in the continuum between populations that seems to have pattern.
Figures 4 and 5 are seedlings of a collection of mine from north east of Nieuwoudtville of H. nortieri. They could be related to ‘globosiflora’ and it will be interesting to see the flowers. However, it may be foolish to draw any conclusion from whatever flower shape emerges in the broader nature of the variation already observed although not adequately documented, both in H. nortieri and in other species complexes. Plants with these stubby deeply mottled leaves can be the consequence of growing conditions just as much as of genetic variation intrinsic to a ’species’.
Figs 6-12 are seedlings of another collection of mine (MBB7608) from Melkhoutrivier south of the lower Breede River and north of the Potberg Mountain. This collection is reported in Chapter 7 of Update 4, I think very aptly entitled “The brutality of reality in Haworthia”. It is one of 12-15 populations in the greater area that support my suggestion of H. mirabilis as including H. maraisii, H. magnifica and H. heidelbergensis. I suspect that H. mutica is also involved to some degree or other, while it is not difficult to extend comparisons to as far afield as Riversdale and further to H. emelyae in the Little Karoo.
Collectors will no doubt always have a problem with plants that do not match the perceptions they attach to names. Kobus Venter has always expressed anathema for anything but random selection when collecting specimen plants. This is in some degree correct because the specimens are supposed to represent a population. However, the opinion on a population is largely based on field observation. Subsequent cultivation and observation do not greatly influence that, while oddities and unusual forms may help mould opinion. While Nature Conservation may object to the introduction of permitted collections eventually into the collecting arena, I see no ethical grounds for this. I insist that it is an undeniable right that people have right of access to a natural resource with the proviso that there is no pressure on sustainability. ♦
When I was at the Karoo Garden I became a bit befuddled by the way botanists referred to the Cape Floral Kingdom. It seemed to me that they used the term for the vegetation that was on the Table Mountain sandstones and conveniently excluded that which was not. Thus the “Fynbos” vegetation, characterized by its Ericaceae, Proteaceae and Restionacea, was synonymous with this floral kingdom. An official document was published at the time which purported to classify the Southern African vegetation into biomes as major floral assemblages with very broad boundaries. It did not make sense to me because my observations were that the “fynbos”, however different in terms of historical origin, was essentially a flora of the sandstones, and that there was rather a winter rainfall biome which included karoid (Succulent Karoo mainly) flora. The role of geological substrate and skeletal soils seemed to me to be pivotal as there are places where one can virtually take a single step from one vegetation assemblage into another.
My impression is that it is the dramatic range of geological formations and highly eroded and eroding (in geological time) surfaces that is responsible for the extraordinary diversity and variation of the Cape Flora. My experience was also that taxonomists were not really aware that this might be significant in respect of plant classification. Botanising in the north and east of the country is not quite the same as botanising in the Cape and certainly it appears to me that European botanists familiar with the plants of a non-eroding land mass do not, or did not, give enough attention to the relation of substrate to plant variation. Unfortunately I am not a geologist, but I want nevertheless to illustrate a few plant populations which demonstrate the complexity of the problem where dramatic differences in soils over very small distances are reflected in variation in vegetation and in the plant species themselves.
In the Compton herbarium, there is a specimen of a Haworthia collected by Prof. Compton. It is labeled as from “Potberg sandy flats”. Now I digress because there is a problem of semantics. The word “retuse” is currently defined in botany and in the Collin’s Dictionary as “end rounded with a central depression”. In Smith’s “Dictionary of Plant Names”, the name “retusus” is said to mean “with a rounded slightly notched tip”. But this is not how it is understood nor used in Haworthia. Haworth, and other writers of the time, used the term(s) “retuso-deltoid” to describe leaves that are “bent back like the end of the thumb”. Thus “retuso-“ meaning “bent back” and “deltoid” referring to the triangular end-area. It definitely does not refer to the end notching of a flat leaf face. When we speak of the “Retuse” haworthias, we mean plants with bent back leaves and a flat triangular end-area. Prof. Compton’s specimen was of such plants where in the dried herbarium specimens the margins of the leaf ends seem to have been cartilaginous.
No other collections of Haworthia were known to G.G. Smith from the Potberg area other than perhaps H. minima. The Potberg area (see map) is also quite extensive as it is about 35 kilometers long and about 10km wide. My records at the time of writing Haworthia Revisited were virtually limited to reports of Luckhoff, Otzen and Herre of a small black species (maraisii!) and two collections of such a plant by both Frank Stayner and Peter Bruyns. I had twice been down to Infanta to be shown H. variegata by Chris Burgers on one occasion, and when I collected H. turgida south of Malgas on another. However, Chris Burgers had also reported two different plants, and one of these was from Buffelsfontein, south of the Potberg inside the De Hoop Nature Reserve. It was not possible to form any opinion about Burgers’ specimens without seeing live plants and I assumed that they were probably in the “mirabilis” complex. Adam Harrower recently came to light with a collection from Sandhoogte. This is also south of the Potberg, but about 15km east of Buffelsfontein. I was fortunate enough to see this population and was of the opinion that it was close to H. mirabilis. I was not convinced that this was what Prof Compton had recorded, but the phrase “sandy flats” also stuck in my craw. The Sandhoogte and Buffelsfontein populations (figs 1 to 5) are in very rocky streamside slopes and sandy flats are not the place to look for any Haworthia. The leaves in these plants are sub-erect and this seems to be related to habitat where slope and cliff dwelling plants differ from those on the flats. I regret that the pictures are not strictly comparable. We had spent the day trying to relate the De Hoop Oxalis records and scrappy specimens to correct names, and the final search for Burger’s Haworthia was more difficult than we had anticipated. By the time we found the plants, the light was too poor for photographing them. The photographs of the two cultivated plants obscure the close similarity of the plants in the field populations.
While partly motivated by trying to locate Trichodiadema and Acrodon species for Dr. Heidi Hartmann, my wife and I discovered extraordinarily pilose “retuse” plants along the lower Breede River (figs 6-9) This is about 4km over the Potberg from Sandhoogte and I surmised it to be the same, as both those and the Buffelsfontein plants evidence incipient spination. All these collections flower in late summer and I consider them to belong in the greater “mirabilis” group as it is becoming increasingly clear that fewer species is more probable than more.
On our most recent visit to the area, we discovered an almost smooth version of the plants being discussed, at a point about 4km over the mountain from Buffelsfontein (figs 10-12). The habitat is extremely rocky and gravelly, as at the lower Breede River, and I am sure that this is now related to Prof. Compton’s collection. The fact that the leaves are retuse in these latter two populations is related to the levelness of the area. I include all four populations in an informal category I refer to as H. mirabilis “pilosa”, despite the fact that in one population only, the plants are startlingly hairy.
Now I come back to my starting point. The geology of the area is most complex. The Potberg is mostly Table Mountain Sandstone except for the eastern most tip which is Cape Limestone. At the foot of the western end there is Bokkeveld shale which is home to a tiny remnant of renosterveld and all of this vegetation type that is included in the De Hoop Reserve. South of the Potberg it is mostly sandy and the vegetation is “fynbos”. North of the Potberg is a different story all together. While the sandstones meet Bokkeveld shale, the situation is confounded by the extensive and complex tertiary deposits characterized by ferricretes, silcretes, terrace and fluvial gravels and other. This is reflected in the vegetation of the area and close alongside Fynbos genera one can find dense patches of Succulent Karoo species such as Gibbaeum austricola and Acrodon. If one can step from one vegetation type into another like this, it is axiomatic that the complex exposure of this mother material is going to have a dramatic influence on the species that comprise the vegetation.
While I think that the collections I record here are related to what Prof. Compton collected, the habitats can definitely not be described as “sandy flats”. However, I can by no means be said to have explored the area thoroughly. The plants are very cryptic and can be confined to very small and localized habitats so they are very easy to miss. ♦
I wonder. I have written so many words purporting to be my last that my credibility here too must be under stress. Two very recent articles of mine in Alsterworthia deal essentially with that issue, although they also cover the discovery of Haworthia mutica (Buffeljags) (= H. groenewaldii Breuer). They do not cover my subsequent thoughts on actually reading the description of this new “species” by Breuer, Marx and Groenewald. I hope that the present manuscript will explain why I reject this as a Latin binomial although anyone who is in the least familiar with my writing should already know. Spurred on by that discovery, I instigated a search in another area of the Buffeljags valley adjoining the Bontebok Park accompanied by Jannie Groenewald who informed me of what he had found in still another area I had long wanted to explore. So I instigated another search there too and again with Jannie. A discussion of these new finds is submitted to Cactus and SucculentJournal where I trust it will be published. The essence is already in Alsterworthia and this article is written to widen the readership, submit more pictures and maintain continuity with the 6 volumes of HaworthiaUpdate that Harry Mays has been so conscientiously and determinedly publishing. This is all writing that may not otherwise have seen the light of day. I am personally extremely grateful for that as I have had a mania since writing my revision Haworthia Revisited and Update Vol. 1 (both Umdaus), to set the record straight and explore all the unknowns, or at least some of them. So on with Haworthia retusa ‘nigra’.
The second area opened to me by meeting Jannie, was a farm southeast of Tradouw Pass, Heuningklip. In the Tradouw Pass, H. retusa ’turgida’ is found on the steep cliffs at the southern end. The road has been rebuilt since I first saw the plants and the population is greatly reduced from what it was. I simply do not have a picture handy to show what those plants are like, but that is not the need here and it does appear in Haworthia Revisited. Immediately south of the pass is another population of plants that puzzled me from the moment I saw them until now. In Haworthia Revisited there is a picture of one clone under J.D. Kobus Venter’s number 93/35. It is named as H. magnifica var. magnifica. The plants are present in a tension zone between Shale and River gravel, and vegetationally between what was Karoo Valley Bushveld and grassy Renosterveld. In April 1997 I collected seed and grew countless seedlings from there, which were all different. Only one picture, as MBB6666) is included in my long treatise on what I discussed under the title of “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.That picture is also under the name H. magnifica var. magnifica but it was not intended to stay that way. It must be noted that my use of names in the Update volumes was not in a decision-making process so much as a learning and informative one. I read my conclusion to the treatise and note that I made no formal name changes. It is only as further field exploration yielded more and more information that I came to realize that H. magnifica and H. mirabilis, classification-wise, are inseparable. The Tradouw collection still worried me because of the close proximity of H. retusa ‘turgida’. These plants at Tradouw and all the things I referred to as H. mutica ‘nigra’ are in fact better understood as variants of H. retusa in the complexity of its relation to H. mirabilis.
This article thus covers exploration of an area between Swellendam and Heidelberg that I have neglected for too long. It is particularly applied to the problems of classification and naming that have persistently clouded my life and my relations with other writers. But as importantly to me is the unclouding of the way taxonomy and classification is useful and necessary for those interested at all in plants and how it is viewed by them.
Consideration and use of formal names Here I need to explain the naming style I now adopt. I have dropped the use of any rank below that of the species name. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable herbarium specimen that is used to anchor the Latin name. The scientific ranks are aggregates and the terms variety and forma are so too. There is simply too much variation between populations and within populations to enable the certain identification of almost any plant that the binomial system suggests. There are no clear entities and ranks and this article should demonstrate that. There is a separate code of nomenclature that covers cultivars. Note that I do not drop names. I only drop the suggestion of status that additional names have, be they single clones or population references for all the different plants within them. I have used names in Haworthia for 70 years and intensively for the last 40 years and I am very familiar with the difficulties of any system. For the field and all the populations and plants I have seen, I can do no better than offer the list of names I updated in February 2009 in Haworthia Update 5.2:192. This, and the set of Updates, comprises a near total overview of the genus and effectively a revision. The attempted revisions by others fall short for me because they do not cover the range of populations that I have seen nor what history tells me, although they may include populations not seen by me. That is not strange because the area to explore is still vast. Another problem with even trying to be formal, is that which G.G. Smith encountered (Mrs. L. Bolus did too) where several people were generating new names at the same time and trying to keep track formally with concurrent name changes would not have been, is not, possible. The point I should make is that in the last ten years, my field work has confirmed the predictive nature of my classification and these latest finds are in extraordinary agreement with that kind of expectation. What I have found essentially agrees with what I would expect. This is dramatically so for H. mirabilis based on a find by J. Jelimicky at Sandhoogte midway geographically and visually between H. mirabilis ‘bobii’ Hayashi (my nom. nuda ‘pilosa’, ’velcro’, Steven Hammer’s ‘Shaggy dog’) along the Breede river, and a collection of mine further south in the DeHoop reserve, MBB7886 H. mirabilis, discussed (p11) and illustrated (p33) in Haworthia Update 5.2.
Prior to extending exploration in the Buffeljags area, I had also been down to the Haworthia populations in the lower Breede River area. This is where H. mirabilis seems to assume a wide range of forms and H. retusa ‘turgida’ and H. variegata’ are also present. I also, in the company of Kobus Venter and Lawrence Loucka, revisited Kransriviermond and Morning Star south of Heidelberg where H. floribunda is in the mix with H. retusa ‘turgida’ and H. mirabilis (in my broad new sense that includes H. maraisii,H. magnifica and H. heidelbergensis). Thus many of those images were very fresh in my mind when I went to Heuningklip southeast of Tradouw. Looking at all the plants and in editing my pictures I was truly impressed with the reality of continuity. The essence of H. retusa and H. mirabilis is in all those plants. A mobius strip is a strip of paper twisted and joined at the ends to produce a shape that only has one surface. What we have in the Southern Cape, and covered or touched on in this article, are interlinking mobius strips with one main surface as well as loose ends that present identifiable end-points. I tried once to illustrate the continuities between populations with single or multiple lines joining the different populations to express degrees of similarity. I can also visualize a pie-chart in which each population is represented by a circular chart with the contribution of main elements drawn as slices proportional to the strength of contribution. Table 1 is a schematic and very crudely suggested enumeration for such a set of charts. I have put the elements retusa and turgida as separate under an H.retusa heading for no other reason that I want to emphasize turgida as a variant at the two geographic extremes. One is eastwards where H.retusa ‘retusa’ is absent (Brandwacht, Mossel Bay) and one westward where it is present (in mutica, Hasiesdrift, Bredasdorp). H. retusa ‘turgida’ is present both east and west. But there are many populations contributing to the H. retusa pie in all the ranges of the two parts, from 0-50% thus making up the continuity from retusa to turgida.
Consideration and characters.
a. The pie chart Measuring the similarity of any two plants is extraordinarily difficult. Doing this for a population is even more so. Doing so for many populations, including those unseen in order to fulfill the predictive requirement of a proper hypothesis, is next to impossible. Hence all the disputation and name abuse. In table 1, I speculate an arrangement to illustrate the problem. The pie is constructed from the figures in the chart for just that set of genetic characters that are directly relevant to the set of plants in question. The pie is incredibly big and it should be understood that there must surely be pie-parts that spread their influence beyond the restrictions of the slices I conceive. So my table is restricted to the few species that are immediately relevant in respect of geographic association and direct interaction (by possible cross-pollination?). A pie-chart is drawn for each individual population. In the chart I assume that there is a set of genetic information that programs for H. retusa ‘retusa’. I have also suggested a different set for H. retusa ‘turgida’. If one considers only these two elements and all the known populations, my suggestion is that there is a set of charts that moves smoothly from one set to the next with no interruption. There is no discontinuity and the sets average 50:50 for the information. Thus it is one species and H. turgida is noted as H. retusa ‘turgida’.
Table 1. A schematic distribution of similarity
Table 2 is the complete set of pie-charts that should convey the possible relationships of the different populations and from which can deduce and understand to what degree H. retusa and H. mirabilis can be related. Also it will indicate the similarities between, and the difficulties of, trying to explain H. pygmaea in the east and H. mutica in the west. Both arise from the same genetic source. The problem is compounded by the infiltration of H. floribunda in the area midway between Heidelberg and Swellendam with the re-emergence of H. floribunda in the south at Potberg. H, variegata also enters the picture but from the south and also in the Potberg area. H. emelyae is the very probable combined entity entering the little Karoo. H. rossouwii is almost independent except in the very south-west at Bredasdorp and it may possibly link to other elements such as H. herbacea and H. reticulata.
Table 2. Pie Charts
At Heidelberg there is interaction between an element that is a 50/50 retusa/turgida with a variant of H. mirabilis cf ‘heidelbergensis’. The latter name suggests that H. mirabilis is the general name for all the plants of that species, and ‘cf’ is an abbreviation for ‘compare with’ ‘heidelbergensis’ to refer to the small plants in populations previously regarded as independent species.
b. What really constitutes the pie The pie is the totality of the genetic material, the DNA that determines all life forms. It is the genes, the materials of characters and inheritance, and the repository of all genetic information. The pie is very large indeed and it is said that the DNA pie of an elephant only differs by 3% from that of a mouse; that of a man by 1% from a chimpanzee. In Haworthia one species from another ‘species’, I could guess at 0.001% or less regardless of whose concept I follow. This difference does also not reside in one consolidated slice that can be reduced to a single small pie as I have done. In DNA analysis, certain segments of DNA from cellular chloroplasts, mitochondria or the chromosomes themselves are isolated and the sequence of the paired amino-acid is analyzed. It requires a formidable statistical process to arrive at a two-dimensional tree (phyllogram) that illustrates a branching relationship with a measure of branch separation and lengths. This can perhaps also be used to generate pie-charts with an innumerable number of slices. I personally consider the phyllogram generated in the DNA sequencing process grossly inadequate.
c.) The characters This is extraordinarily difficult to specify no matter how obviously the plants seem to differ from one another. Number, size, shape, colour, arrangement, surfaces, spination and attitude of the leaves in the spiral rosette constitute the main things that determine how we see the plants. These are influenced by growing medium and growing conditions so these are complicating factors. Flowers are used in the same way as the leaves but far less so because they are so similar in all the species. Flowers are also not always present either which reduces their usage. Time of flowering is a major factor because it is linked to the reproductive process. It is also difficult to capture any firm picture of the flower because of the aging process in the 3-5 day life of the individual flower.
d.) The relevance here It simply is not necessary to parade the obviousness of this and the way in which will be attempted anyway in the discussion below. It is flowering time that may be critical beyond, or linked, to the facts of spatial geographic relationships of populations. It is traditionally flower structure that has formed the basis of classification of plants because of the reproductive significance and the role of reproduction in distribution of genetic material between plants and populations. Pollination of Haworthia seems to be primarily by a Solitary bee species. These bees make individual nests and are not communal like the Honey bee. They make tunnels in the soil as nests during the summer months. I have observed them as adults in the winter but then they are less numerous and less active. Few Haworthia flower at the height of winter and do so in two main time frames viz. spring and late summer. The implications are that breeding between isolated populations apart from any other consideration of compatibility, is a question of distance and time. A nest bound Honey bee can forage to 13km from its nest. A free flying Solitary bee can be speculated to cover a considerably greater distance. All these things play on the frequency and power of inheritance. Genetic drift is acknowledged as the genetic movement within a population away from the norm for a species because of isolation, while genetic flow is perhaps the holding together of similarity over geographic distance by a travelling pollinator. Thus the statistical probabilities of breeding across distance are impossible to derive.
I cannot quantify any of these things nor suggest any greater knowledge but this suggests to me, given the freedom with which the various species hybridize in cultivation and even in the field, we need to consider geographic relationships. In my extensive experience with other genera, it appears to me that a key consideration is if things grow together and maintain their difference, or not. Haworthia species are invariably in small isolated populations and seldom do different species grow intermixed. They are also influenced by geology of the substrate and particularly so because they favour the lowered competition from more vigorous and larger plants on the rocky sites or shallow soils they favour. That can be stated in the converse, Haworthia are favoured by conditions poor for other plants.
e.) Flowering time and growing together. The existence as discrete species can be judged from the way in which populations share geographic space. I discussed this in some detail in Update 2.1:76. If two sets of plants grow close together and flower at the same time, while still maintaining obvious group differences, then they are most probably different species. If they flower at different times while still maintaining difference then the question of being different species is taken as certain. If the groups are separated and flower at different times while the differences are small and apparently within the experienced range of variation observed between populations, then it may be problematic. One would need to resort to the extended distribution and variability of the groups in question. Generally in Haworthia, species belonging in the same sub-genus do not share habitat and this is indicative of fairly rapid absorption of introduced genetic material (by pollinators). Hand-pollination and the ease with which plants of different species hybridize in cultivation suggest that there is practically no breeding isolation between them. In the following discussion, use is thus made of general distribution of species and proximity of populations in assessing to which species they best belong. It should be borne in mind that variability may be clinal. This means that the plants change along a geographic gradient. The direction of the gradient may also not be of increasing distance i.e. linear. It may also be circular so that ends of a gradient may meet again as two populations that appear wholly different, but linked by an extended set of other populations. A much more complex situation may actually exist where these clinal gradients may be multidirectional and thus involve more than one apparent species. My observation is that populations do differ and differences follow a pattern that becomes predictable i.e. gradual genetic difference over geographic area. This is not just a result of environmental gradient, but also linked to the question of genetic drift (the tendency of isolated populations) to move away from the general norm. It also results from the genetic flow between populations where the change of genetic material between populations at the distribution extremes is only through other populations.
f.) The species definition This is a subject I have covered many times. My view is that species are dynamic chaotic systems oscillating between different character states with time and with different rates of mutational change. Closely related species may not be recognizable on the basis of single character states and the spatial geographical distributions and local juxtapositions of groups of plants may be the only way to understand what the situation is and so make sensible decisions.
g.) Habitat geology I contend that species richness in South Africa is largely due to its varied geology, skeletal soils and comparatively mild climate. This present discussion deals with species that are associated with a geological phenomenon that is the Tertiary deposits of the Southern Cape. I am no geologist and am required to make the pretention I am one to explain why plants are distributed the way they are and what influences difference among them. In this case the vegetation at Buffeljags, Swellendam has been described (by Breuer et al. cited below) as on silcrete, and the general vegetation as by vegetation scientists as Ruens Silcrete Renosterveld. But there are two problems. The first is that vegetation classification is not infallible and I have never found it really useful in dealing with a wide range of species that occupy skeletal habitats. Secondly the substrate geology is definitely not simply silcrete nor is the clay soil either simply bentonite or kaolinite. There are many problems in the classification of clays. Bentonite in common terms seems to be the yellowish clay mined in large quantity from Heidelberg eastwards and there is a transition to ochre bearing clay. Bentonite may be aluminium, potassium, sodium or calcium dominated. As a montmorillomite clay it should have expansive cracking properties. Kaolinite is popularly regarded as the very white clay that is commonly seen under the rocky top layer of the sharp inselbergs that rise above the wheatfields west of Heidelberg. It seems to be less mineral dominated clay and does not have the expansive/contractive properties of bentonite. The distinction between ferricrete and silcrete seems to be that in ferricrete, iron oxides are infused into the petrified water deposited layers. Both the kaolinitic and bentonitic clays seem to be derived from the weathering of Bokkeveld shale that underlies the silcerte/ferricrete layer. It does not require any stretch of the imagination to see now why any one inselberg may be a little different from the next, and sometimes a lot different. What we do not know is how rapidly the landscape has moved from a broad white silcrete/ferricrete underwater plain to the remnant condition of that rocky plain today. Neither do we know what it has meant for the plant species that are linked to, and dependent on, the habitats generated in the erosion processes.
Map of Swellendam to Suurbraak area.
Map Legend – east of Swellendam.
1. MBB7899 H. retusa ‘nigra’. Heuningklip. 2. MBB6666 H. retusa ‘nigra’ ↔ H. mirabilis. S Tradouw Pass. 3. MBB7896 H. retusa ‘nigra’. Heuningklip. 4. MBB7897 H. retusa ‘nigra’. Heuningklip. 5. MBB7898 H. retusa ‘nigra’. Heuningklip. 6. MBB7888 H. mutica. Rotterdam. 7. MBB7889 H. mutica. Rotterdam. 8. MBB7890 H. mutica. Rotterdam. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags. 10. MBB7741 H. mutica. Dankbaar. 11. MBB7892 H. marginata. Rotterdam. 12. MBB7891 H. minima. Rotterdam. 13. MBB6644 H. mirabilis. SW Swellendam. 14. MBB7704 H. mirabilis. Bontebok Park. 15. MBB7805 H. mirabilis. Bontebok Park. 16. MBB7823 H. mirabilis. Klipbult. 17. MBB7887 H. mirabilis. Rotterdam. 18. MBB7876 H. mirabilis. Disselfontein. 19. MBB7901 H. mirabilis. Crodini E. 20. MBB7902 H. mirabilis. Crodini E. 21. MBB7903 H. mirabilis. Crodini E. 22. MBB7904 H. mirabilis. Crodini E. 23. MBB7905 H. mirabilis. Crodini E. 24. MBB7906 H. mirabilis. Crodini E. 25. MBB7907 H. mirabilis. Crodini E. 26. MBB7908 H. mirabilis. Sandkraal W. 27. MBB7909 H. mirabilis. Sandkraal W. 28. MBB7910 H. floribunda. Rietkuil. 29. MBB7912 H. mirabilis. Rietkuil. 30. MBB7913 H. mirabilis. Rietkuil. 31. JDV93/35 H. retusa ‘turgda’. Tradouw Pass.
The populations
Set 1 – THE TRADOUW PASS AREA (H. retusa ‘nigra’) This first section deals with populations that complement those in the long discussion under the title “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50.There I ended with the words … ”The habitat photo (of H. mutica Hasiesdrift) raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.” Added were these words … ”There the saga for the moment must rest.” What I had in mind was exploration that no one seems very anxious to do. This has now been done and I would lay the matter to rest as follows:
1. MBB7899. H. mirabilis, Heuningklip. See figs 1-30 Fig. 1 is a scenic view of the area to show the countryside and the type of inselberg the plants are often found on. Fig. 2 is a closer picture of the erodible clay held in place by grass clumps at the base of which the plants often are. Occasionally they are simply out in the open sun. I term these places pressure bursts because water build-up in the soil causes collapse of the small banks and erosion can be quite fast. Several times I have found plants lying in collapsed soil. Such a soil bank near Heidelberg has retreated by nearly 2 meters in about 15 years albeit accelerated by road maintenance, but it does relate to any attempt to explain how much suitable habitat may have disappeared with time.
Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – bFig. 2. A photograph of the habitat of MBB7899.
Figs 3 to 30 is a selection of plants at the site where one is almost compelled to examine each plant separately and easily generate another 28 practically different images. Fig. 3 is of a plant with the surfaces roughened by small tubercles. SEM (scanning electron microscope) images of similar leaf surface often show incipient tiny spines on the tubercles and they appear as spines or hairs in populations like H. mirabilis ‘pilosa’ at Ballyfar, or H. mirabilis at Windsor Riversdale. This surface roughness and all its degrees is fairly characteristic of H. mirabilis but is does occur in H. retusa too. Plants illustrated in Figs 4 and 16 has one muttering Kruisrivier and Komserante, northeast and east of Riversdale respectively, and which I would have allotted to H. magnifica in the early period of my exploration no doubt have Breuer and Hayashi names. Now I simply say H. mirabilis and add the locality to place the plant to at least give it a geographic reality that is better than using the formal Breuer/Hayashi in hanging commas as I also suggest. Fig. 6 is apparently a seedling and demonstrates the leaf-shape of H. floribunda that led me to suggest that H. floribunda is a perpetuated juvenile form. Of course the evidence suggests to me that H. floribunda coalesces with H. mirabilis in this wider area although it does appear at Swellendam to the west of my map as the large form and at the extreme SE corner of the same map as a convincingly normal form. Fig. 9 demonstrates longer pointed leaves and Fig. 12 short more obtuse leaves. Fig. 17 is of a plant that has relatively smooth leaf surfaces rather like H. retusa and the leaves are conspicuously lined. In Fig. 20 there is a suggestion of muticate (without a point) leaves. Fig. 22 is again quite strongly lined. Fig. 22 shows a leaf with a single line. Fig. 25 demonstrates what is termed rotate leaves where the points of the leaves point around as though to encircle the plant. Fig. 28 made me recall plants of H. mirabilis at Melkhoutrivier in the south west near Malgas and further to Ballyfar. The flowering time is late summer.
Fig. 3. MBB7896 H. retusa ‘nigra’. Heuningklip.Fig. 4. MBB7897 H. retusa ‘nigra’. Heuningklip.Fig. 5. MBB7898 H. retusa ‘nigra’. Heuningklip.Fig. 6. MBB7888 H. mutica. Rotterdam.Fig. 7. MBB7889 H. mutica. Rotterdam.Fig. 8. MBB7890 H. mutica. Rotterdam.Fig. 9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags.Fig. 10. MBB7741 H. mutica. Dankbaar.Fig. 11. MBB7892 H. marginata. Rotterdam.Fig. 12. MBB7891 H. minima. Rotterdam.Fig. 13. 13. MBB6644 H. mirabilis. SW Swellendam.Fig. 14. MBB7704 H. mirabilis. Bontebok Park.Fig. 15. MBB7805 H. mirabilis. Bontebok Park.Fig. 16. MBB7823 H. mirabilis. Klipbult.Fig. 17. MBB7887 H. mirabilis. Rotterdam.Fig. 18. MBB7876 H. mirabilis. Disselfontein.Fig. 19. MBB7901 H. mirabilis. Crodini E.Fig. 20. MBB7902 H. mirabilis. Crodini E.Fig. 21. MBB7903 H. mirabilis. Crodini E.Fig. 22. MBB7904 H. mirabilis. Crodini E.Fig. 23. MBB7905 H. mirabilis. Crodini E.Fig. 24. MBB7906 H. mirabilis. Crodini E.Fig. 25. MBB7907 H. mirabilis. Crodini E.Fig. 26. MBB7908 H. mirabilis. Sandkraal W.Fig. 27. MBB7909 H. mirabilis. Sandkraal W.Fig. 28. MBB7910 H. floribunda. Rietkuil.Fig. 29. MBB7912 H. mirabilis. Rietkuil.Fig. 30. MBB7913 H. mirabilis. Rietkuil.
These images can now be compared with MBB6666 at Tradouw Pass to establish if my assessment is reasonable. My contention is that this MBB7899 population is midway between the other three Heuningklip populations I am about to illustrate and Tradouw Pass. This similarity is repeated widely and eventually contributes to my contention that H. mutica has been generated by drift away from its two contributing parent species viz. H. mirabilis and H. retusa.
2. MBB6666 H. mirabilis. Tradouw Pass See figs 31-71. This population is at the mouth of the Tradouw Pass and relatively low-lying in respect of the previous population. The habitat is quite different being partially coarse river gravels, weathered shale and at one point is an un-weathered shale ridge where the plants are wedged in the cracks. The habitat is of course very different to the steep sandstone cliffs within the pass itself where H. retusa ‘turgida’ occurs. I initially thought these plants fitted with H. magnifica. A decision derived from a very small sample available at the time. Now I looked for the images and photographs in my records under H. mirabilis where they should have been since my inclusion there of ‘magnifica’, and also under H. retusa where I found them in the folder ‘nigra’. The flowering time is late summer. In this respect similar to the previous set MBB7899 H. mirabilis but the plants are more robust. Figs 31-45 were taken in 2009; figs 46-53 in 2011 and figs 54-71 are from seedlings grown by myself. I tried to reduce the pictures and eliminated a few. But I really think they all need to be presented to show again how serious the difficulty of circumscribing them as a single recognizable key-able entity really is. While looking at my pictures I seriously thought that the set taken in 2011 differed from those taken in 2009. The cultivated plants of course are not directly connected to their source and the variation is self-evidently wide. But it is equally so in field plants. There are the smoother surfaces of H. retusa and some very rough surfaces for H. mirabilis. My decision became that they fitted best with the ‘nigra’ populations where my organizing had taken them despite flowering time, but here MBB7899 shows absolute geographic position between H. mirabilis and MBB6666. I consider this key evidence for the basic continuity for H. retusa and H. mirabilis that I have conjectured elsewhere and I would not be able to defend an argument that MBB7889 plants belong better in H. mirabilis and this means MBB6666 belongs there too. I would simply dispute that there is no better solution so stick with history as far as possible and that means H. mirabilis. If Breuer or Hayashi have a name for a variant from this population intended as an aggregate name, I recommend the use in inverted commas. I would actually like the use of the equilibrium diacritic of a double arrow pointing between two names viz. H. mirabilis↔H. retusa ‘nigra’, although this available symbol is not quite the thing.
Fig. 31 6666 H. retusa ‘nigra’
I come to this vexing question of “typical” that is bandied about even by myself when I liken population to population. There is the same range of variables in MBB6666 noted for MBB7899, but the plants do not darken as much and they are larger. So we need now to look at the other three Heuningklip populations asking what significance is attached to their presence in exactly the same habitat as MBB7899, but geographically separated and by perhaps 500m and 2km respectively. The plants are so variable that it is not possible to arrive at a characterization that separates any two plants, or any two populations in any geographic set for that matter.
3. MBB7896 H. retusa ‘nigra’, Heuningklip 1 See figs 72-89. The plants in this and the next two populations were flowering in September as does H. retusa and they are relatively large, green in colour and generally smooth. Of course both vary enormously due to both intrinsic genetic difference and extrinsic local situation. One plant within an older decaying grass tuft was nearly 200mm diameter, and a healthy green colour, another plant in the open white clay was less than 50mm diam. and dark in colouration.
Fig. 72 7896 H. retusa ‘nigra’
4. MBB7897 H. retusa ‘nigra’, Heuningklip 2 See figs 90-105. The first two pictures show the general landscape and the local habitat where the plants occur. The clay is highly erodible and often there are no plants at all in the clay. Some of the plants have very acuminate pointed leaves while in others they may be relatively obtuse and rounded.
Fig. 90 The countryside looking towards the Tradouw Pass.Fig. 91 The ‘white clay pressure burst’
5. MBB7898 H. retusa ‘nigra’, Heuningklip 3 See figs 106 -132. The great variability is again evident and in this population the plants were observed to be generally without much venation or lines as in fig.112. Then one plant, fig. 119, was so striped as to stand out from all the populations seen.
Fig. 106 Silcrete hilltopsFig. 107 7898 H. retusa ‘nigra’.
Set 2 – THE BUFFELJAGS AREA (H. mutica) This second section follows a recent manuscript submitted to Alsterworthia yet to be published entitled “Some last closing thoughts” written in the hope of finding the closure I seek and that will not come. That article is about a population that has been named as a species viz H. groenewaldii, in an article authored by Breuer, Marx and Groenewald (Alsterworthia 11.2:15) that I only saw after the submission of my manuscript. I also only saw the article after the exploration I would have asked for to verify opinions. The description seems to have been prompted by Marx who refers to similarities to H. springbokvlakensis and to H. magnifia var. atrofusca and raises many issues that my manuscript does not cover. I simply find the whole discussion unrealistic and especially so when no further exploration was done to examine anything. One should refer to Breuer’s World of Haworthias Vol 1 to see that the naive promises in the foreword and introduction of exploration and field study, was never achieved. It is true that the plants have an unusual surface texture and it is also true that the habitat is quite unlike any other H. mutica occupies. But it is Marx’s comment that disturbs me viz. “Unfortunately Bruce Bayer keeps on refusing to see it as something new and calls it simply H. mutica ‘Buffeljags”. This is a distortion and misrepresentation, and whose misfortune this is, not specified. My reticence is reflected in the preceding discussion and Gerhard does not seem to follow, or agree with, what I have been writing about and trying to explain in this general context for so many years. In fact in an unpublished (to my knowledge) manuscript he dismisses all my writing as “pretentious” that no doubt it is, because I have never fancied my self as a taxonomist in the tradition of taxonomists. I was as fascinated as Marx by the plants and am even more anxious than he is to see just how they connect to the probable geographic counterparts in the context of the species definition that I apply. I was also very curious to know what the connection to H. mutica and the H. retusa ‘nigra’ issue, only now laid to rest in the preceding discussion, actually might be here. This is why I use the word “instigated” in reference to exploration in the very valley where H. mutica ‘groenewaldii’ occurs and to the obvious habitat for similar plants. It is surely unfair iniquitous to so demand that I see this population as “something new” in an article that rather disregards the history of the subject and what I have been trying to understand of it.
I also recently submitted a manuscript to Cactus and Succulent Journal (US) where I wrote of “H. groenewaldii Breuer”… “I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February/March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time”. I have in many places discussed the problem of “hybridization” and it must surely be well-known from my writing that H. floribunda merges into H. mirabilis. Less well-known will be the fact that evidence for H. variegata doing the same is also available. In respect of floral characters, I wrote a chapter in Haworthia Update Vol.3.2:88 explaining the problems of using floral morphology for species recognition. I would challenge Breuer et al. to produce comparative results from studies of the flowers in any of these problematic species to demonstrate difference. Regarding the curious shiny leaf surface, I refer to Haworthia Update Vol.2.2:141, Chapter 15 where I discuss electron scanning photographs of leaf surfaces. This also demonstrates how problematic leaf surface is. It would be tiresome indeed to parade all the evidence in support of these contentions of mine. In Haworthia Update 5.2:108 I also briefly discussed H. mutica from Buffeljags (‘groenewaldii’) and submitted a large number of photographs explaining the habitat and what the possible connection was to the H. retusa ‘nigra’ conundrum. I may also have written about the issue in Haworthiad. Suffice to say that the authors of H. mutica ‘groenewaldii have certainly not checked or considered “all the facts” as the principal author claims. The very opposite seems to be the case.
Considering my oft repeated observation that geological substrate plays a major role in shaping species appearances, the Breuer, Marx and Groenewald description of the habitat for H. mutica ‘groenewaldii’ is a little misleading (see considerations above). Nowhere else is H. mutica on identical substrate and this would just support my contention of ecotypic differentiation that does not justify a Latin binomial. There is considerable doubt about the actual difference between these river boulder situations and the widely distributed terrace gravels elsewhere in the area, notably the Bontebok Park near where the following populations were found:
6. MBB7888 H. mutica, Rotterdam 1 See figs 133-169.
Fig. 133 7888 H. mutica.
7. MBB7889 H. mutica, Rotterdam 2 See figs 170-188.
Fig. 170 7889 H. mutica.Fig. 185
8. MBB7890 H. mutica, Rotterdam 3 See figs 189-191 The three populations above are separated spatially by several hundred meters and they are all on remnant river gravels on a more gently sloping bank than the Buffeljags east bank. The vegetation is closer to conventional fynbos as in the adjoining Bontebok Park, but grassy. The pictures are self-explanatory and the leaf surfaces of the plants are also generally as for the Buffeljags population. What is significant is that I cannot say I saw a single plant in these populations that resembles the one chosen and pictured as typical for this highly dubious species, H. groenewaldii. It makes more sense to me to identify plants from these populations by collecting number than to generate a supposedly descriptive Latin epithet from an imagined typical representative.
Fig. 189 7890 H. mutica.
9. MBB7801 H. mutica ‘groenewaldii’, Mullershof, Buffeljags See figs 192-215 The following set of photographs has surely seen the light of day somewhere and I repeat them because as far as I am concerned they are H. mutica with the shiny surface claimed for “groenewaldii’. Dankbaar is some 16km west of these Buffeljags populations and the substrate is Table Mountain Sandstone. The flowering time is just normal for H. mutica. Fig. 215 is a photograph of a plant apparently collected by Jannie Groenewald and it is quite uncharacteristic of the population and would suggest that it is a product of hybridization with H. mirabilis in the vicinity (e.g. Klipbult).
Fig. 192 7801 H. mutica ‘groenewaldii’.Fig. 215 7801 H. mutica ‘groenewaldii’.
10. MBB7741 H. mutica, DankbaarSee figs 216-235. My observations in respect of these two sets of plants confirms my widely written convictions about Haworthia taxonomy and the state of classification in the absence of a definition for the concept of “species”, and the intrinsic value of Latin binomials for rational and sensible communication. Of course this Dankbaar population is not identical to other H. mutica populations and I am not driven by fixation on any one character that defines this species. It is a geographic set and the various sets are not simply separable. But I am not finished yet.
Fig. 216 7741 H. mutica.
Set 3 – THE BUFFELJAGS AREA (Robustipedunculares) The western Buffeljags area also produced a population of H. marginata and one of a very curious H. minima, that both complement occurrences in the Bontebok Park a little further west. The conventional small, blue-green H. minima is present in the park as well as an unusually green population.
11. MBB7892 H. marginata, Rotterdam See figs 236-243. This population was quite exceptional and one of the strongest populations known to me. But the plants were in flower in September whereas the flowering time for the species is normally late summer; and the flowers and seed capsules exceptionally large. I do not consider that these factors command another Latin binomial.
12. MBB7891 H. minima, Rotterdam See figs 244-252 and Bontebok Park figs 253-4. These were also extraordinary plants, larger than those seen in the Bontebok Park population where the plants were also green viz. figs 253-4. Fig. 252 confuses the issue because, while the other plants seemed alike to H. marginata and distant hybrids, this one plant appears to be a recent hybrid. This occurrence cannot be overlooked in respect of other introgressive (or seemingly introgressive) populations of Robustipedunculares species. The plants were not in flower and again we have a suggestion or direct evidence of hybridization across a difference of seasonal flowering.
Fig. 244 7891 H. minima.Fig. 254 sn H. minima, Bontebok Park
Set 4 – THE GREATER BUFFELJAGS AREA (H. mirabilis) This is a set of 15 populations of H. mirabilis that occur east, west, south north of the H. mutica ‘groenewaldi’ related populations. I exclude from the discussion and illustrations the several populations of H. floribunda ‘major’ populations to the northwest, while I repeat the statement that H. floribunda contributes to the general gene pool of H. mirabilis and together with H. retusa, they all contribute to the gene pool of H. mutica ‘groenewaldii’. The huge presence of populations more readily assignable to H. mirabilis (sensu lato – in the broadest sense) is used by me to validate my opinion that the “mutica” elements draw most genetic resource from the H. retusa element, and hence why I refer those plants to H. mutica.
The sequence of populations 7901-7908 is located in a horseshoe of the Breede River, southwest of Buffeljags in very promising habitat. Only H. mirabilis was seen. I perhaps need to point out that these populations would have been extraordinarily difficult to name under any previous dispensation of mine where I was still trying to uphold H. magnifica,H. maraisii and H. heidelbergensis as species. These I have since merged in H. mirabilis and I reject the introduction of a group name like “aggregate” as Breuer seems to have done, as a complete fabrication that does nothing to solve the problem other than to highlight it.
The following 6 populations are all associated with tertiary and river gravels. There are differences between most of the populations but of an order that I cannot articulate. It is also not possible to really encapsulate the appearances of the plants in the two photographs for each population that I submit. At the same time it does not seem to me possible to make any communicable distinctions.
13. MBB6644 = 6860 H. mirabilis, SSW Swellendam. See figs 255-256.
14. MBB7704 H. mirabilis, Bontebok ParkSee figs 257-258.
15. MBB7805 H. mirabilis, Bontebok Park See figs 259-260.
16. MBB7823 H. mirabilis, KlipbultSee figs 261-262.
On selecting these two pictures I was reminded of my observation that there was a significant H. floribunda appearance to the plants. I had believed that I had seen H. floribunda in the Bontebok Park nearby many years ago but subsequent exploration has only produced H. mirabilis albeit with some similarities to H. floribunda. My reaction was to suspect that H. mutica ‘groenwaldii’ most probably also then needed to be considered as a product influenced by H. floribunda.
17. MBB7887 H. mirabilis, RotterdamSee figs 263-264.
18. MBB7876 H. mirabilis, DisselfonteinSee figs 265-266. These plants were a little more robust than the others in this set and rather similar to MBB7901.
The following populations are almost entirely in Bokkeveld shale but on a very close interface with tertiary gravels. In some places there was also some wind-blown sand.
19. MBB7901 H. mirabilis, Crodini 344, S Swellendam See figs 267-278.
20. MBB7908 H. mirabilis, Sandkraal, S SwellendamSee figs 279-282.
Set 5 – BACK TO TRADOUW PASS – AS IT HAPPENED In the process of compiling this manuscript I was bothered by the ever present question of what is still out there that might influence the outcome. My wife, Daphne, was planning an outing to see chameleons at Swellendam while simultaneously contacting landowners for me around the Potberg. Always anxious to get two birds with one stone, I asked her to find out who owned the land west of Heuningklip that we had just explored with Jannie Groenewald. The result was that we ended up spending a day with Odette Curtis of Custodians of Rare and Endangered Wildlife, south of Suurbraak and finding three populations that, for me, put the proverbial cherry on the pie.
All three were on the slopes of silcrete inselbergs and almost the last of this habitat westward from Heidelberg to Buffeljags. The first inselberg was different to the second and from the Heuningklip habitats because it was dominated by Aloe arborescens and A. ferox. Aloe brevifolia was also present in an unusually small form. The habitat was very impacted by grazing animals and there is an outside possibility that H. retusa ‘nigra’ may once have been present although I personally doubt it. To our surprise we found…
21 MBB7910 H. floribunda, Rietkuil, Suurbraak See figs 283-288.
There were many plants but severely trampled and disturbed. Their survival was obviously aided by the fact that much of the plant is below soil-level. But my experience of this species is that it struggles to maintain its identity in the presence of either H. mirabilis or H. retusa. They never share immediate habitat. There was no hybrid present and this also adds to the problem of interpreting flowering time and distance apart from other species as part of the process. The plants were not the large green variant of the species present to the west at Appelbos, Swellendam; and the leaves were both with and without marginal spines. Eastwards I know the species to be close only at Blackdown north of Heidelberg and at Goedverwagting south of the N highway about 15km east southeast. It definitely seems to be absorbed into H. mirabilis west and north respectively of those two populations. So while this is a slight contradictory element of surprise, it is the next two that really excited me. These were again inselberg habitat but without the strong silcrete upper layer of the former habitat and without the Aloe element. The second ‘population’ was about 200m from the first and comprised four groups of plants along a longer north-facing ridge.
22. MBB7912 H. mirabilis, Rietkuil, Suurbraak See figs 289-306.
23. MBB7913 H. mirabilis, Rietkuil, SuurbraakSee figs 307-331 I am very hard-pressed to say much more than that these plants met all my expectation for the very variable set of plants and populations that I ascribe to the one single species H. mirabilis. Who could possibly articulate the differences of the many populations of this species from each other and from this one? Variable becomes a done-to-death word. The individual plants, in two populations quite unlike any others I have seen, were generally robust with large specimens up to 100mm. diam. The colour is dark, although green when shaded within or under grass. The leaves were usually sharply pointed but there was always evidence of one or two leaves or odd individuals in which the leaves were muticate i.e. without points. In some plants the leaves were notably lined in others not. The colour also varied from chestnut to deep reddish-brown when the plants were more exposed. The surface texture was generally tubercled but there was evidence of smoothness of the leaves in many plants. Flowering time was obviously late summer gauged by the absence of flower at the time of the visit viz. September. Except for one oddity. This was a single plant with a weak flower spike. I photographed this for the useful information the flower-bud provides. Where Breuer et al. claim to have considered all the facts, this is a character that is conveniently overlooked when claims are made for the significance of floral characters when paraded to substantiate yet another claim to species significant difference. The bud-tip in all the Southern Cape species of subgenus Haworthia with the exception of perhaps H. rossouwii have this very obvious “fish-tail” bud tip. The tips of the upper two petals are flattened and spreading to resemble a fish’s tail. It is most marked in H. herbacea and H. reticulata where the bud tip curves downward from the semi-erect position of the body of the flower and then up again. I am fearful that my inability and reluctance to consider these plants as “new” may be the motive for someone to rush forward and apply a Latin binomial.
My opinion is that these two populations mirror the paradox in the east where H. pygmaea ‘fusca’ is nearer to H. retusa ‘retusa’ andwhere H. retusa ‘turgida’ is nearby. H. mirabilis ‘splendens’, in the doubt about its affinities, in my opinion remains in the context of H. mirabilis. This is a conclusion I reached after exploration of the area between Riversdale and Albertinia where the same elements noted in this article repeat themselves. I do not for one moment concede that better explanation is available by resort to a multitude of Latin binomials and by an artifact like “aggregate”.
Conclusion I have been deeply disappointed that so little progress on the classification front has been made among the fraternity writing about Haworthia. In communication with Lawrence Loucka, he wrote …”You might consider saying that a genus revision is needed and that until amateurs depart no proper botanist will make the investment. Until then chaos will rule, names will proliferate.” This is a useful and insightful comment and generally summarizes what I have often said. Unfortunately the position has been rather exacerbated when proper botanists have made their presence known and I can recount many instances to support this statement. My own view has been that botanists have not been able to confront genera that are so publicly and popularly known, and garlanded with names. Thus amateurs, including myself despite at least an iota of formal training in science, simply have a field day generating names like rabbits out of a hat.
Unless some proper thought is given to this whole question of what the names are supposed to mean apart from catalogue items, taxonomy (classification) will never lift out of the disinterest, disrepute and scorn that collectors justifiably seem to have for it. Latin binomials do have a real value and to demean this by one-dimensional derivation and rationalization is intellectually fraudulent. My writing is replete with examples of the variability and interaction of populations over large area. Taking a single population and one flower, and often a single plant, to generate a species description and a Latin binomial is grossly simplistic.
The one change that this article generates is the confirmation that the population MBB6666 from immediately south of Tradouw Pass is better identified by name as an H. mirabilis variant, and not as H. retusa ‘nigra’. MBB7801 at Buffeljags and now also at Rotterdam, remains as H. mutica and the epithet generated by enthusiasts can be added to make the trinomial H. mutica ‘groenewaldii’. For formality, the name for the form illustrated as typical would be H. mutica ‘Groenewaldii’. A few additional names will be needed for similar clones and a lot more for the cultivars that will be derived from this population set.
My conclusions here make no substantial departure from anything that I have observed or written over 50 years. While the subject has been comprehensively covered in the field and in writing, there is a huge amount of exploration that still needs to be done if the genus is to be still better known. My prediction is that the answer will conform very closely to the species-list I finalized in 2009 (see Alsterworthia 5.1:192). The disclaimer is that botanical classification may eventually reach the point where the real nature of species and their purpose comes to be known. In that case what we now may see and persuade ourselves are discrete species, are not. Instead these highly complex chaotic systems may in fact be quantum units of consciousness and whole elements such as some genera and sub-genera we now recognize will be the species.
Acknowledgement I want particularly to acknowledge Jannie Groenewald whose interest and enthusiasm are boundless. Then Allan Jeptha, an environmental law practitioner with great vision for the Suurbraak community; Odette Curtiss, a consultant for the conservation of Renosterveld vegetation; Lawrence Loucka whose patient guidance and comments I value and who generated the set of pie charts; Kobus Venter and Steven Hammer who have always been supportive and contributory; Max Coetzee of Felix Unite and Samie Lategan of Buffalo Breeding Programme for access to Sandkraal West and Crodini East; Jaap Viljoen for his contribution; my wife Daphne, who trusts me and whom I deeply trust. ♦
The writing of my grand finale was inspired by several things. One of these was another item of a mind-numbing foray into the classification of Haworthia. So I asked that deep thinker and observer, Gerhard Marx, for a devil’s advocate (abbrev. DA) point of view which he has done with the same competence he has as an artist. I have many times in my writing addressed the issue of a species definition and produced one too. Not surprisingly the first thing the DA does is dismiss my definition without producing one of his own. Simply being able to say that an indeterminate number of plants from some population are sufficiently different in respect of a character or two from other haworthia, is motivation enough for the generation of a new name?
The case of H. groenewaldii Breuer, described in an article authored in Alsterworthia 11.2:13-17 by Breuer, Marx and Groenewald is the case in point. It presents the description of this supposed new species from Buffeljags east of Swellendam. The article is written in the first person (Breuer) who quotes extensively from Gerhard’s e-mails, and includes a piece by Jannie Groenewald under the heading “Description of the Vegetation type and distribution”. The overall impression is of an article that conforms to the style of a forgotten era and it is not possible or sensible to attempt a rational dismissal. Who is actually responsible for the article and how does one correct misleading statements without giving offence?
There is probably only one main issue that can be made level-headed sense of. This is the statement in the article…”another (it is not clear what the other was) striking feature was the fine but rough leaf texture…” Then quoted again…”the typical H. mutica grows only 20km to the west at Dankbaar”. But the Dankbaar plants also have the same shiny leaf surface. The word “typical” is generally overused in the article and I would counter claim that the Dankbaar plants are typically H. mutica and reinforce my argument for the similarity to the Buffeljags plants. But this is not the main issue, nor is it the least of many rectifications the article almost demands. In the new-age freedom of communication there are virtually no secrets anymore and this adds to my discomfort because the article only touches on some of the issues clouding Haworthia classification. The DA says I should meet this following argument in support of an H. groenewaldi’…” it is consistently smaller than H. mutica and reminds superficially actually of H. bruynsii or springbokvlakensis with its rounded leaves. Kobus Venter’s feeling was that it is closest to H. magnifica var atrofusca and I agree 100%.” These are subjective statements and I cannot agree about the size as I have seen many H. mutica populations that I do not think supported the opinion. The roundness of the leaves is also subjective. The element H. mirabilis ’atrofusca’ in any case has plants with variably rounded leaves, so which do you mean?. The name ‘mutica’ means leaves without a point and rounded and it is no co-incidence that Col. Scott initially identified H. springbokvlakensis as H. mutica until I pointed out to him that I thought H. mutica was actually the plants (note plural) in the Bredasdorp area. Scott plagiarized my words describing them alongside old wagon tracks that suggested their discovery in earlier times.
DA lists several more indeterminates…”all the unique morphological features like small size, rounded rough-textured and uniquely flecked leaves”. Of course there are not several more as I have already dismissed two and this leaves the third viz. the “uniquely flecked leaves”. Firstly not all the plants have flecked leaves and I have shown that these occur in H. mutica at Klipport too. The main issue is flowering time and I agree this is problematic. But this is actually so in respect of the whole Southern Cape Haworthia complex where it is quite evident that there are two flowering periods, summer and winter and that despite this difference there is hybridization. What happened in the past?
DA writes…”MBB 7801 flowers…with the typical thin tender peduncles and delicate flowers identical to that of H. maraisii and some H. magnifica and H. mirabilis. Normal H. mutica flowers in spring (Sept-Oct) and has more robust peduncles and flowers.” This word “typical” again, does not convey the truth. There are no floral characters by which one can separate the three “species” the sentence suggests. I have also seen small H. mutica (Crodini) with very small weak inflorescences and comment on this as a general variable. See fig. 149, a recent picture of peduncles of H. mutica from Grootvlakte, Riviersonderend. There is a population of H. mirabilis (I presume Breuer and Marx would use the name H. magnifica var. magnifica) from south Riversdale MBB6651. The plants are very small and cryptic and when I collected seed it was from small slender inflorescences with 3-4 seed capsules. In cultivation I raised a large number of plants that were all different and some were huge. The inflorescences were robust as well with 15 -20 capsules. So this reference to floral characters is simply a loose statement and I do not believe that a proper overview of the flowers in H. retusa and H. mirabilis will reveal any dramatic differences. I have taken good note of flowers and in particular the strange fish-tail bud-tip that characterizes the larger set of populations in the southern Cape.
DA states that I… ”made several blatant errors by ignoring the importance of flower features in his studies”. It this one supposed error or several? I have not ignored flowers at all and it is a blatant error to say so. I simply did not and still do not find them useful. It is fraudulent to suggest otherwise. That fish-tail bud is highly significant and I made a great effort to quantify the differences between H. reticulata and H. herbacea. That the attempt failed was only because the sample required to get to statistical difference became too large. Nevertheless I then tackled a good number of H. mirabilis populations that also have the same bud tip. I concluded that there was a difference between the populations south of the Langeberg and those north and then had to discard that as erroneous because the same problem of variability occurred as with the rosette of leaves. Even the general flowering times I gave for the area was quite wrong – flowering time ranged from November to late April. It is odd that the fish-tail bud tip does actually influence the tips of the upper tepals in that the margins tend to be flattened together. But there is a huge difference in that in H. herbacea and H. reticulata the upper tepals are held far out from the mid-axis (hence ‘subregularis’ or ‘cowboy-hat’) while in H. mirabilis the tepals may meet above the middle upper tepal.
More by the DA… ”So, only careful observation and comparison of diagnostic features, starting with flowers, is the solution.” This is a weak sop because it is something that I started to do and found quite unhelpful. The history is that the small aloid genera were virtually recognized by the fact that the flowers were small. The differences between the subgenera were dismissed by Dr L.A. Codd with the statement that “small differences” were being ignored. DA overlooks a single major binding factor between H. retusa and H. mirabilis viz. bud similarity. I have also discussed in detail the whole question of diagnostic features and went to the genus Oxalis to substantiate my observations that small detail is not helpful. It comes back to the issue of what is a species? I maintain that they are not these artificial elements circumscribed by morphological detail. They are systems set in a geographical framework. The species, whatever they are, have surely not arisen from independent sources. My field work is very extensive and I have shown over and over again that the populations are linked so that when one finds a single population like at Buffeljags, one asks how it fits. It is not any misconstrual by G.G. Smith that a sample from a population south of Heidelberg was likened to H. mutica. In his time it was not known where H. mutica even occurred and hence he made a mistake in describing H. otzenii. DA is defending a methodology that failed miserably and is not going to provide any solutions in the future.
Well, well, well one might say. The DA maintains that Bayer has lost his way, to support an argument that he had stopped “doing taxonomy”. DA is not strong on logic but it is intended that way and neither are the DA’s observations necessarily correct. DA writes this … “Bruce’s dilemma is that he spent a lifetime and hundreds of thousands of Rands doing his research and while he did indeed do an invaluable contribution, he is now faced with the fact that he can’t do that full and neat integration of it all because 1) there is more variety in the wild than what his dispensation recognizes and he feels overwhelmed by the idea of having to try and do a real truth-reflecting revision at this ripe age and 2) he made several blatant errors by ignoring the importance of flower features in his studies.” Point two I have already dismissed, but DA cannot be serious when he suggests that there is some kind of dilemma involving money and time spent. This is an issue that I will have to address when I meet the Angel of Death.
I do recognize that there is great ‘variety in the wild” quite contrary to DA’s claim that here is more variety than what my dispensation recognizes. The main gist of my writing has been this extraordinary variation. I am not overwhelmed by it at all and I maintain that my dispensation is the only way to rationally express it. To say it confuses me is not an overstatement because it is clarification that I have been seeking. Now Steven Hammer puts it very well… ”it’s almost as if you are being blamed for nature’s complexities” and I said the same thing in different words long ago. The Latin binomial system fails and I have argued and shown evidence for the need of a new and different model What is more I have proposed such a model too. Anyone who contemplates at any age, the “truth reflecting revision” the DA demands will be faced with a daunting task of citing the synonymy of all the species described or recognized by Breuer and Hayashi (let alone cite all the collecting records). That alone could occupy a lifetime. Not only that, but to also cover the usages and references by the many authors who have written about the plants would be a humungous task. I suspect that DA is implying that I have become senile with a “ripe old-age” comment. My response is that at least my senility was preceded by some degree of clarity in contrast to what I am forced to conclude about contemporary Haworthia taxonomists! I see no reason whatsoever to depart from a logical argument about species definition and the significance of geographical distribution from my species list of February 2009. Effectively this is a revision and it just does not have literature and specimen citations that meet the needs of “a full and neat integration”. Let me state categorically that there is not the slightest hope of such a pie-in-sky solution.
So we ask Lawrence Loucka’s question… ”How best to separate facts and knowledge from personalities and ego?” First present the facts. I want to add to the information that I presented in my article on “Haworthiaretusa ‘nigra’ – A grand finale”. There I discuss and illustrate plants from the wider general area of Buffeljags and especially from the Tradouw area to the northeast. Covered are four populations from the farm Heuningklip, and three from the farm Rietkuil. Here I want to discuss and illustrate eight more populations from the farm Van Reenens Crest that lies between. The gist of my argument is that classification of Haworthia is far beyond this nonsensical and simplistic generation of new names in exactly the same way that a miasma of names was generated in the years before 1948.
It is not just chance that the area I have recently been exploring has been so neglected for so long. It is a high-lying and fairly featureless area and very grassy in comparison to the Karoid Broken Veld, Karoid Valley Bushveld remnants, and the rocky hills, streams and river beds of the areas north east and west. The vegetation is a grassy Renosterveld and the species diversity seems extraordinarily low. There is not much there to attract a succulent plant enthusiast and explorer compared to far more enticing and rewarding countryside elsewhere. My exploration there is a direct result of my having exhausted most other options available to me. An added consideration is that of prediction when long experience has proven that connections between populations exist and need to be found.
So it is not surprising that we found 8 new populations, 6 in the context of H. mirabilis as I circumscribe it, and 2 for H. retusa ‘nigra’ in the same vein of personal circumscription. These populations were all new to me and very exciting. Nevertheless I would not dream of seeing them as new species anymore than I did for H. mutica ‘Buffeljags’. The Van Reenen’s Crest populations fit a pattern and need to be considered in the context of the bracketing populations at Heuningklip and Rietkuil discussed in the preceding chapter. The collections and illustrations are mapped in an accompanying map and are identified as follows:-
The populations.
Set 1 – H. mirabilis. The plants in the following six populations are collectively different from those at either Rietkuil or Heuningklip. Using the erroneous or absent logic that characterizes the description of “H. groenewaldii”, there is no doubt that at least one new Latin binomial should be generated. Far from being confused by the huge variation I have seen in Haworthia and guided by a systems view of species, I see these as being representative of one species. It would be useful if proponents of some other dispensation take these pictures individually and arrange them according to the characters they so diligently use to denote their many different species. The question of flower character I laugh off because I have tried to use it and know it will fail. I will come to flowering time again. Plant size is barely significant because in such clay soils the plants do not grow to the size that they will in cultivation. Generally the plants in these populations fall in the category 35mm to 800mm diam. But it is difficult to gauge age and the direct effects of niche favourability that will favour growth and size.
1. MBB7914 H. mirabilis, Van Reenens Crest See figs 1-19. This was a small population as can be judged from fig.1 where the pressure burst and exposed white clay area barely exceeds 100sq m. The plants were only on the perimeter of the area. This is what makes it quite difficult to assess what the population structure may have been in the era before farming and cultivation. Very seldom indeed have I found plants anywhere in pristine arable or even semi-arable landscapes. Fig. 2 is a view looking across to Heuningklip and the locality of MBB7896 H. retusa ‘nigra’.
Fig. 1 View of locality MBB7914.Fig. 2 View from locality MBB7914 to Heuningklip.Fig. 3 7914 H. mirabilis
2. MBB7915 H. mirabilis, Van Reenens Crest See figs 20-27. Fig. 20 is a view looking south to a very large area that we did not explore because we chose instead a similar valley immediately beyond the one pictured. The MBB7915 locality was rather odd. In fact the whole exercise was in the respect that we kept being reminded how similar the situation was to Kiewietsvlakte where H. mirabilis and H. retusa play out a similar show. In this case there was no actual pressure burst. In this case there was less white clay and there were the characteristic platform-like ridges as if made by grazing animals. There were few plants and while rather scattered, were still confined to what I can only describe as low-biomass bearing area i.e. shallow gravelly soil that does not support dense vegetative cover.
Fig. 20 View from locality MBB7915.Fig. 21 7915 H. mirabilis.
3. MBB7916 H. mirabilis, Van Reenens Crest See figs 28-48. Fig. 28 is a view of the white clay in an area very like that pictured in Fig. 20. Very heavily utilized by grazing animals, it is surprising that there were so many plants. The variability is quite evident from the pictures and some plants stand out as very different.
Fig. 28 The clay soils in which the plants often grow. Locality MBB7916.Fig. 29 7916 H. mirabilis
4. MBB7917 H. mirabilis, Van Reenens Crest See figs 49-70. This was a dramatic population because the plants were not quite where we expected them to be. Fig. 49 shows a more typical habitat in the distance. Here the plants were rather in a very brown gravelly situation and absent a short distance away where there was more white clay. As for all these populations there was a wide range of surface textures and also leaf shapes and endings. Often the leaves had a very distinct mucro (end-awn or point). Equally often the leaves were without any point and rounded and this was not true for all the leaves on any single plant. It is very problematic to make statements about the plants because when we were looking at Rietkuil plants we were reminded of a host of more easterly populations and plants. In this case even variants of H. emelyae came to mind.
Fig. 49 View from locality MBB7917 to that of MBB 7919.Fig. 50 7917 H. mirabilis.
5. MBB7918 H. mirabilis, Van Reenens Crest See figs 71-87. This population was unusual in that the plants were generally small and proliferous. Sometimes there were as many as 20 rosettes in a clump.
Fig. 71 7918 H. mirabilis.
6. MBB7919 H. mirabilis, Van Reenens Crest See figs 88-116. One needs to refer to the map to see how the populations now relate to one another. These were again very variable as I have come to expect in the whole H. retusa/H. mirabilis collective – to coin yet another adjective to do what I do not think the formal system allows. Note carefully fig.100 in which a young bud is clearly visible.
Fig. 88 7919 H. mirabilis.Fig. 100
Set 2 – H. retusa ‘nigra’ We found just these two populations.
7. MBB7920 H. retusa ‘nigra’, Van Reenens Crest See figs 117-137. These plants were in full flower and setting seed as one expects for the summer-flowering H. retusa elements. In fig. 128 a plant is shown with well-developed end-awn and fig. 134 a plant with rounded leaf tips. Fig. 137 is simply a view of the flower that does not even depict the more normal fish-tail bud tip – see next!
Fig. 117 7920 H. retusa ‘nigra’.Fig. 128Fig. 134Fig. 137
8. MBB7921 H. retusa ‘nigra’, Van Reenens Crest See figs 138-148. Fig. 138 is a plant that could have come from an H. retusa ‘retusa’ population from around Riversdale, while Fig. 146 could have come from H. retusa ‘turgida’ from somewhere around Heidelberg. Fig. 147 clearly shows what I mean by the fish-tail buds. Fig. 148 shows that there has been quite extensive exploitation of the white clay (kaolin). Fortunately (?) unlike the bentonite that is mined on large scale, the kaolin pockets are often small and restricted and apparently not commercially viable.
Fig. 138 7921 H. retusa ‘nigra’.
Conclusion The plants in the above populations cannot be categorized without reference to the totality of populations and I will try and summarize my point of view. These populations occupy a position very central to the distribution of both H. retusa and H. mirabilis. I need to quote DA again who wrote … ”His (Bayer’s) emphasis is upon phylogeny and understanding the evolutionary processes at work in Haworthia and he loves getting involved with very complicated arguments and theories which (frustratingly or delightedly) to him nobody quite follows or is really interested to follow.”
DA is most fortunate to be able to dismiss the basis of classification and what all biologists involved in classification try to do. It is quite wounding to say that I enjoy this theorizing. I would be far happier if there was no contradiction and I could wander out in to the veld and be assured that there were smart neat boxes and simple ways of filling them from a manageable list of names and unequivocal identifying characters. My observation is that all this so-called “taxonomy” practiced by too many writers is so flawed because the very basis of biological variation is wholly ignored and forgotten. It does not take rocket science to imagine that science was supposed to set us free from superstition and blind belief and allow us to individually question the nature of creation.
Here we have a real problem that is overridden by a basic problem in formal classification. If I could start now afresh with a clean slate and knowing what I now do, I would still be handicapped by the restrictions of the nomenclatural code. This is that I do not believe that the name H. mirabilis as typified by a plant that comes from an outlying population truly represents the species. Perhaps this is a point not understood by the collector community, and in fact even by botanists in other disciplines. The “type” is only there to serve as a reference point for the NAME and may not be remotely typical of the species that this name is applied to. What is unfortunately true as with the crop of taxonomists who have muddied the classification of Haworthia in more ways than one, is that more often than not the application of names is not to species at all but to an odd assortment of plants that meet no criterion for a basic unit of life forms. In the case of the name and type of H. retusa there has been curiously no problem and the vast variation there is forgotten simply because of familiarity. The interesting thing is that the name H. mirabilis was not even in use before I resurrected it in about 1976, so anything that remotely departs from an unfamiliar norm must be “new”? For H. retusa ‘nigra’, I must have got the name ‘nigra’ from the old literature and reference to the Kransriviermond collection that Smith informally listed as H. mutica. The point is that in my acknowledged ignorance I attached the name to H. mutica because of the absence of an end-awn and a relatively rounded leaf tip to the plants seen. (Looking back to all my pictures I see that in some plants there is definitely an end-awn.) The dark-colour has resulted in a name that is not appropriate for the populations that can be considered to constitute the taxon. Neither is the population at a geographic end-point and as a probable hybrid entity representative. This statement also only scratches at the surface of a real problem of origins and changing faces of the plants with time.
Reflecting about relationship as a taxonomist is required to do, and with due respect to DA and the weird views of my preoccupation with relationships the invitation to him required; it seems to me that the unseen and unknown element, now only evident (H. retusa ‘nigra’) as a widespread and common element in this central area, is important. It may be the precursor to the real way in which H. retusa and H. mirabilis have developed in close synchrony and association in both easterly and westerly directions.
The really strange occurrences are evident from the map herewith (fig. 150). The two species do not grow in direct close association in the area under consideration. There are places where they do e.g. Skeiding about 20km eastwards; KomseRante, Riversdale; Soetmelksrivier, E Riversdale. Why is this? There is no observable difference in habitat or associated vegetation, and still there is pattern in the distribution. It appears to me that they are separated by flowering time (that does not preclude hybridization as field hybrids show) and in character by colour and a tendency to greater surface roughness of various kind and degree in H. mirabilis. The mystery deepens when H. floribunda appears in two places but again as sole occupant of habitat seemingly favourable to either H. retusa or H. mirabilis. Pattern is broken here because the evidence seems to be absorption into H. mirabilis in other places where the two species do closely meet. The conclusion has to be that I have come no nearer to a neat solution. The H. retusa ‘nigra’ populations cited above will attract no takers, but the H.mirabilis populations will. I create no new name because my personal view of Haworthia does not call for one.
Fig. 149 Peduncles of H. mutica, E RiviersonderendMap of Tradouw South
Acknowledgement I particularly want to thank Gerhard Marx for his considered points of view he has crafted as my Devils Advocate. He is an extraordinary grower and observer, and an artist of the highest order. He had driven me to more carefully and thoroughly explore and document what I see and think. I am extremely grateful to him. Lawrence Loucka has been a great help as an advisor, and so has Steven who brings something special to the whole field in the way he “communicates” with plants and about them. Kobus Venter is ever supportive. Access to Van Reenens Crest was through kindness of Trevenan Barry unsoured by the pressures of harvest time. ♦
These field trips, to the Potberg this time, are always made with some objective in mind in respect of new exploration. In this case I wanted to get more pictures of H. mutica as it is a species that I have few digital images of. There were also localities that I remembered from the days when I was sweeping the countryside at a fairly coarse scale and was not much bothered by detail. I confidently expected the number of real species conforming to that in other fields of botany and zoology, to be in the region of 33. I never dreamed that such divergent views would, or even could, arise from less information than even then available to me. So while 450 names were whittled down to the mid-hundreds by me, students of the genus have in recent years pushed that up to the 600 mark. My opinions have been couched in quite conservative terms but it is a problem of the nomenclatural system that an identification in respect of a Latin name evokes a reality that does not exist. I maintain that the problems we experience in Haworthia are no different to that which exists in many animal and plant genera. I think that primarily this is because of the absence of insight into, and understanding of, the actual nature of species and the two dimensional model we use to relate them. Species are very variable systems because they have to be to survive the constantly changing world they occupy. In this article I am just going to present images of plants within populations of four different species viz. H. variegata, H. minima, H. mirabilis and H. mutica.
Set 1 H. variegata H. variegata has an odd distribution in that it is primarily in the lower Goukou River drainage system southeast of Riversdale. Plants of H. monticola from the Uniondale and Willowmore areas, and even those of H. angustifolia from the Eastern Cape, can be very similar. So the distribution south and west of the lower Breede River is quite remarkable. Two populations, one high on the western end of the Potberg Mountain and another northwest of this at Luiperd’s Kop are very similar to those in the Goukou area. Chris Burgers of Cape Nature brought me plants from the lower north slopes of the Potberg that were quite different. Small single plants with outcurving leaf ends barely protruding above the pebbles the plants were growing in. I named this H. variegata ‘hemicrypta’ but not without reservation in the same way as I also named the plants in another population on the Potberg and at Kathoek as H. variegata ‘modesta’.
Chris Burgers did take me to his site many years ago and it is one of those places I cannot recall in the pebbly flats and Fynbos vegetation of that area. So the finds here seem to represent that same element but with subtle differences in respect mainly of variegation and spination of the leaf margins. I have mentioned elsewhere that there is an interaction with an element I refer to as H. floribunda although it is not clear if this indeed that species, or if it is an emergent polytopic element of H. mirabilis in consideration of the variation and distribution of these two apparently different species. We may have a truth for one area that is different from a truth in another. Polytopic means similar looking things arising in different places from a common genetic pool.
1. Map of the Diepkloof area (fig.1)
Map of the Diepkloof area.
2. MBB7923 H. variegata S Diepkloof. (figs 2)
Fig. 2.1 7923 S Diepkloof
3. MBB7924 H. variegata SE Diepkloof (figs 3)
Looking west to the Kleinberg
4. MBB7925 H. variegata SE Diepkloof (figs 4)
Fig. 4.1 7925 SE Diepkloof
5. MBB7927 H. variegata SSE Diepkloof (figs 5)
Fig. 5.1 7927 SSE Diepkloof
6. MBB7928 H. variegata SSE Diepkloof (figs 6)
View looking east to 7927.View looking southwest with Kleinberg off to the right.
7. MBB7515 H. variegata Stoffelsriver (figs 7)
Fig. 7.1 7515 Stoffelsrivier.
Applying the available varietal name, I should refer to these populations as H. variegata var. hemicrypta. However, I think this implies fixity of opinion and a certainty or reality that are neither not strictly true. I have added the Stoffelsriver two pictures just as a reference because while the first 5 populations are associated with recent Tertiary deposits and Renosterveld vegetation, the last is in sandstone gravels and in Fynbos.
Set 2 H. minima This species of course belongs with three others in the Robustipedunculares and in my opinion more closely related to Astroloba than to Haworthia. It is widespread and even occurs in the Little Karoo. None of these populations are significantly different from the general run for the species, but we were awed by the beauty of the plants in their natural settings where exposure results in compactness and beautiful colouration. They can occur in quite different environments and habitats. There are a few aberrant populations and hybrids with other members of the sub-genus.
8. MBB7496a H. minima Diepkloof S (figs 8)
Fig. 8.1 7496a Diepkloof S
9. MBB7926 H. minima SE Diepkloof (fig. 9)
Fig. 9 7926 Diepkloof S
10. MBB7929 H. minima SE Diepkloof S (figs 10)
Fig. 10.1 7929 SE Diepkloof
11. MBB7930 H. minima S Diepkloof (figs 11)
Fig. 11.1 7930 with 7947 SE Diepkloof minima
12. MBB7932 H. minima S Kleinberg (figs 12)
Fig. 12.1 7932 S Kleinberg
Set 3 H. mirabilis H. mirablis is an extremely complex group of many populations and plants and distributed from Albertinia in the east to near Caledon in the west, and from the foothills of the Langeberg to the coast. The relationship with H. retusa hangs on the slender thread of flowering time, with H retusa flowering in late spring and H. mirabilis in late summer. The relationship of these two species to H. pygmaea in the east and H. mutica to the west has been mooted by me elsewhere. A similar scenario can be suggested for the species recognized north of the Langeberg.Included in the following sets is the population at Die Kop that I surmise has its origins in interaction between H. mutica and H. mirabilis.
There are three populations in these sets (nos 13-15) that are suggestive of H. mirabilis ‘atrofusca’, while others have distinct resemblances to H. floribunda (e.g. 16). It is curious that nearly all the populations of H. mirabilis that occur in the same general geographic area as does H. mutica are of smallish plants. These I originally regarded as a separate species viz. H. heidelbergensis. This should be seen as one of those decisions based on inadequate sampling and knowledge of the field. This phenomenon now suggests to me that H. mutica is occupying middle ground between H. retusa in the ‘turgida’ form and H. mirabilis.
13. MBB7496 H. mirabilis Diepkloof S (figs 13)
13.1 7496 Diepkloof SView overlooking habitat 7496.View looking west to north of Kleinberg.
14. MBB7497 H. mirabilis S S Diepkloof (figs 14)
Fig. 14.1 7497 S S DiepkloofView looking north.
15. MBB7933 H. mirabilis S Diepkloof (figs 15)
Fig. 15.1 7933 S 7496 Diepkloof
16. MBB7922 H. mirabilis Diepkloof (figs 16) A locality established by Jakub Jelimicky. The similarities to H. floribunda is only apparent in some of the plants where the leaf tips are twisted, slightly flattened and rounded at the ends.
Fig. 16.1 7922 Diepkloof
17. MBB7931 H. mirabilis S S Kleinberg (figs 17)
Fig. 17.1 7931 SS Kleinberg.
18. MBB7531 H. mirabilis S Kleinberg (figs 18)
S Kleinberg slope. Enthusiasts at play.Figs 18.1 7531 S Kleinberg
19. MBB7935 H. mirabilis Langvlei (figs 19)
7935.1 LangvleiLangvlei, the plants are in a tiny bit of rock just beyond the dumps.
20. MBB7938 H. mirabilis SE Stoffelsriver (figs 20) This locality was recorded when Gerhard Marx and Jakub Jelimicky misunderstood my directions. The plants are incredibly cryptic and very hard to see in a not very well defined habitat. One can only follow my opinion on the classification in the context of 8 other populations in the sandstones and Fynbos, and then relate these to populations in shales, tertiary deposit and transitional vegetation.
Fig. 20.1 7938 SE StoffelsrivierSE StoffelsrivierSE Stoffelsrivier View looking east to Sandhoogte
21. JDV86/2 H. mirabilis Haarwegskloof (figs 21) These are small plants in very close association with H. mutica as discussed above and are in quartz outcrops rather than in the shale ridges where H. mutica is found.
Fig. 21.1 JDV862 Haarwegskloof
Set 4 H. mutica H. mutica is not a species that is well-known and this is probably because it is not proliferous. The name mutica means without a point and “typically” plants of H. mutica should and do have no point (end-awn). The main object of the trip was to obtain more images of this species to convey a better impression of the many variants and how it can be viewed in relation to other species.
22. MBB7075 H.mutica Grootvlakte (figs 22) I am not certain of the origins of H. otzenii G G Smith and the locality that I traced was approximately 8km east of Riviersonderend. That site is now very degraded and I could not find plants on a visit many years ago but where I did see them on my first visit there. This Grootvlakte population occupies a similar habitat in an exposed shale ridge south of the Riviersonderend River.
Fig. 22.1 7075 Grootvlakte7075 View of site at Grootvlakte
23. MBB7935 H. mutica Crodini (figs 23) The name Crodini may have been used for the following collection at nearby Witklipkop, and I say this because I did once get seed from there and Jakub Jelimiky informs he has a plant labeled ‘Crodini’. I only saw two plants here at Crodini itself when the site was severely grazed, and we found only 8 plants on this occasion. The plants appear to be in quartz, but this is a bit of an illusion as there are white quartz bands in the shale and the plants prefer the shale.
Fig. 23.1 7935 CrodiniView across site. Grootvlakte
24. MBB6536 H. mutica Witklipkop (figs 24) This is not a robust population in shale and also marginal to quartz.
Fig. 24.1 6536 Witklipkop, Napky
25. MBB7937 H. mutica Platkop, S Napky (figs 25) The substrate here seems to be silcrete but there is no underlying kaolinite and the area is cultivated, which suggests shale. So that is a bit of an unknown.
Fig. 25.1 7937 N Platkop SE LangvleiN Platkop SE LangvleiN Platkop SE Langvlei
26. ADH2729 H. mutica Ouplaas, DeHoop (figs 26)
Fig. 26.1 ADH2729 Ouplaas, De Hoop
27. MBB7934 H. mutica Wolwefontein (figs 27)
In early 1970 I took Peter Brandham and David Cutler of Kew to look at Haworthia in the field. In fig.27.12 the view is southward to Kathoek and the cottage in the foreground was occupied. It must have been abandoned soon after and the access road obscured. The result was that the place was later difficult to find, but the plants are still there. It is not far from the problematic Die Kop population to come.
Fig. 27.1 7934 WolwefonteinWolwefontein
28. JDV85/17 H. mutica Haarwegskloof (figs 28) This record goes back to 1985. We did not have landowner permission here as we did not know exactly where to go. The area was also unfenced. So we were confronted by the farm manager and never took as many pictures nor explored more widely. It certainly would have been problematic had we not simply being photographing. Anybody else may have ended up at the police station. It is a case of farm land too hilly and rocky to cultivate and bought by apparently wealthy city people with an eye to conservation. This may result in the place being game-fenced, a lodge built for the up-market; and made perhaps even less accessible.
Fig. 28.1 JDV85-17 Haarwegskloof
29. MBB6641 H. mutica Hasiesdrift (figs 29 JDV97/26) This was one of my early encounters in 1969 with Haworthia, where I fortuitously stopped at a river crossing, lifted a branch of a shrublet and saw a magnificent plant just like that illustrated and serving as the type for the name H. mutica (see fig.29.1for an example)). A year later a new road was built and if there had been a population there it was no more. In 1996 when I was writing Haworthia Revisited, I went back there and after a diligent search found a very small group of plants at the base of a small plant of Aloe ferox. The two species seldom grow among each other so this really was odd. I have periodically been back to see what was happening because there were many Aloe seedlings. The very small population has waxed and waned and the mothering Aloe seedling has died. There has been a small spread of plants to occupy an area of about 2 square meters. I know here are more plants about 2km away but have not explored the wider area more thoroughly.
Fig. 29.1 JDV97-26 Hasiesdrift.
30 MBB7500 H.muticaXmirabilis Die Kop (figs 30) So we come to the problem child that has acquired the species Latin binomial H. hammeri. I think this is unfortunate as Steven Hammer deserves a great deal better than this. It should not be necessary to have to also manage personal issues in an inquiry into what the name means or if it is a valid statement in science. This population is one I found when I was exploring widely for an understanding of H. mirabilis. It was not growing in conventional habitat and populations occur at many places in the wider vicinity. So I do not comprehend at all why this single population with absolutely nothing to support its designation as a species in respect of anything but the fact that the plants look different from something else. The plants are also very variable as the set of pictures should show, and the plant suggested by the picture accompanying the description is not typical. The proximity of H. mutica and the nuances of these plants with this species and also with that of other nearby populations of H. mirabilis, suggest to me that this population is of hybrid origin. I also use the term hybrid in the sense of genetic similarity because hybridity itself implies separate existence. My feeling is that H. mirabilis and H. retusa have separated in time from a common gene pool and that the process is not complete and nor will it be. My species definition requires that we see all these populations as one system that is spread in geographic space and is changing with time. The rate of change is a fourth dimension.
I suppose one should comment on the shiny leaf surface (fig. 30.3 especially) that is present in H. mutica, as this character is also described as a diagnostic for H. groenewaldii that is as contentious as H. hammeri.
Fig. 30.1 7500 Die Kop
Conclusion The objectives of the excursion were fully met and I am sure these pictures will contribute to a better view of how these four species vary. Perhaps this will illustrate also the illogic of Latin binomials for every variable, however desirable these names are for traders and collectors.
Acknowledgement Bennie Viljoen of Grootvlakte, Koos Badenhorst of Langvlei, Maree and Elizabeth Prinsloo of Stoffelsriver, Ferdie of Diepkloof Security Services, Adele and Vlooi DuToit for Die Kop, Martin Mijnhardt of Haarwegskloof, Neil Giliomee of Hasiesdrift. In some cases we simply could not track landowners and trust they will forgive us our trespasses remembering that invasion of ones territory is close to an unforgivable sin. ♦
It so happens that Heidi Hartmann first visited the Karoo Garden more than 35 years ago and it has been very difficult for me to pay attention both to her mesembs and all my other plant interests. In the last few years she has been working on Acrodon. This is a small genus of only 5 to 6 species that occurs in the Southern Cape with much the same distribution and habitat requirements as Haworthia. She had had some second thoughts on a species she had described as Acrodon calcicola and intimated that she needed photographs to show what proves to be detaching fruits (capsules). So off we went to get that northeast of Bredasdorp at Rooivlei. But Nick Helme had about a year before sent me an intriguing picture of a greenish soft looking plant from near the De Hoop Reserve entrance road to the east. I had considered that it might be an equivalent of the H. muticaXmirabilis population at Die Kop (MBB7500) that Ingo Breuer usefully described as H. hammeri. I use the name with great trepidation because to say what is correct usage is difficult. It could pass as a cultivar name, a varietal name or a form name. I am quite sure it has its origins in the interaction of two species and that is what a botanical name should reflect that; thus H. muticaXmirabilis or however else the nomenclaturists may require. So these journeys are never without distractions as Rooivlei itself is a remarkable site. I find that I have few images of the populations of Haworthia that occur there. The product is nearly all pictures/images.
Set 1. MBB7941 H. mutica Ouplaas, Wydgelee (figs 1-45)
This population is just 3km west of the Die Kop MBB7500 population. There are two intervening H. mirabilis populations; one small, one large and scattered, and a third a little to the north of east and more to the north. But that species is well covered in an Update article and my attention here is on H. mutica. Why Nick Helme’s plant was so green and soft-looking is a bit odd because the population of H. mutica there at Ouplaas is quite one of the largest and most dramatic of any that I have seen of the species. Daphne and I spent hours examining, reexamining and exclaiming at the sheer beauty and variation of the plants. There were plants squeezed into cracks and flattened so that one could not see any centre, others in the flat clayish areas between the exposed vertical shale ridges, but mostly in the shale. The site is north-facing and so is very hot and dry in summer. The plants had just finished flowering.
There was one particular image (fig. 24) I have used to show five different leaf-end shapes on the same plant.
Fig. 1 7941 OuplaasFig. 44 7941 View west to HaarwegskloofFig. 45 7941 View east to Die Kop
Set 2. MBB7943 H. mutica E Klipbankskloof (figs 46-54)
I saw plants and photographed one (MBB7246) south of Klipbankskloof in a valley mapped as Verbrandskloof. There is another farm, also named Klipbankskloof further west where Jakub Jelimicky recorded H. mutica too. I wanted to see that locality and registered the two Klipbankskloof places as the same. Not so. So I intended up exploring further east where we first found this in a west facing ridge. There was no sign of fruits or flower.
Fig. 46 7943 E KlipbankskloofFig. 54 7943 View west to farmhouse
Set 3. MBB7945 H. mutica SE Klipbankskloof (figs 55-80)
This was on a north facing slope and at first we were a bit taken aback because the plants we first observed had flowered poorly or some time previously. But that changed as from north facing slope we entered a very minor valley with both northeast and northwest aspects (more to follow). Here there were many plants and all just finishing flowering. There were some outstanding, dramatic plants. I wish I was a more accomplished and better equipped photographer. Some of the images would have been stunning.
Fig. 55 7945 SE KlipbankskloofFig. 80 7945 View lookin across depression to west facing slope
Set 4. MBB6638 H. mirabilis Rooivlei (KoeiseKop) (figs 81- 92)
It was fortunate for my story that I took pictures here. As in most of the greater area occupied by H. mutica,H. mirabilis occurs as these smaller, often (how often is often?) dark rough versions of H. mirabilis. Of particular note is the first image (fig. 81). This is of a plant with much smoother textured leaves and had there been and H. mutica in the near vicinity I would surely (mistakenly?) taken this for a hybrid because I know from experience elsewhere that summer and winter flowers can and do hybridize. But wait! Rooivlei is a unique site because there is a population of H. retusa ‘turgida’ nearby that is prominent in my whole explanation of relationships that my critics so dislike. Outside chance is that hybridisation is an explanation.
Fig. 81 6638 Rooivlei
Set. 5 MBB7944a H. mirabilis SE Klipbankskloof (figs 93-101)
I have numbered this as ‘a’ and ‘b’ for a reason. The two ‘populations’ are less than 200m apart. In fact the two H. mutica populations are not all that much more distant and there would be some debate as to whether they constitute populations at all. My argument would be that they occupy discrete separated habitats. This mirabilis population falls into middle ground. It is the upper slope where aspect changes from south to north. It is moister and the southern aspect of the small shale ridges provides mirabilis habitat. It is as if H. mirabilis is found where what I can describe is an escape route is present. These dark plants do not readily tolerate a full days exposure to direct sun and I speculate that this is the same kind of thing that happens on north-facing mountain slopes as around MacGregor. Plants are usually where the aspect allows escape from a full days worth of sunlight. The plants are much like at Rooivlei.
[ed.] images mislabeled as 7943, should be 7944.
Fig. 93 7943a SE Klipbankskloof
Set. 6 MBB7944b H. mirabilis SE Klipbankskloof (figs 102-110)
The different number is because the plants are now sharing both habitat and niche with H. mutica. I do not know any other site where this happens. The two species do occur in the close general area but that is then different habitat and the distance between ranges from about 50m (Haarwegskloof) to 400m (Rondeheuwel). What is now fascinating is that some of the plants resemble fig 1 of Rooivlei and we are faced with the probability that there is exchange of genetic material between H. mutica and H. mirabilis. This is what is suggested at Rooivlei too for H. retusa ‘turgida’ and H. mirabilis (see above).
Fig. 102 7943b SE Klipbankskloof
Conclusion None. This is purely instructive. What I gained from the trip was the realization that H. mutica would be a very good subject by which to explore variability and change with time. Also to look at conservation and impact of land-use on populations. Over to Jakub Jelimicky.
Acknowledgement Again a list as long as an arm. Francois Uys for access to Rooivlei. Nick Helme for information in respect of Ouplaas. Odette Curtis for interest in Renosterveld conservation and contact for Nick. Johan Dutoit of Vleitjie and a friendly combine mechanic for access to Ouplaas. Wynand Dutoit of Klipbankskloof for access to what is his wife, Madrẽ’s farm. ♦
The area concerned is the long and wide contact zone between the Limestone stretching from Bredasdorp to Potberg, and the Bokkeveld shale north of that. The soils and vegetation of the two areas are grossly different. The limestones are agriculturally almost useless, while the shales are prime wheat and pasturage producing soils although relatively low yielding. The vegetation of the shales is Renosterveld and there are very few patches left. Large areas resemble ecological deserts with nothing of the original surface intact. Here and there are shale banks and associated quartz outcrops and also some remnants of tertiary deposits that overlie the shale. Under this deposit layer the shale has decomposed to kaolin and in places there are gravel sheets of fine quartz on white clay. The skeletal nature of these remnants is the saving grace but it is unbelievable to what lengths farmers must have gone to make fields arable. Enormous amounts of stone that have been carted away and dumped to make cultivated lands. Sadly the stone is often dumped on exposed rock and prime Haworthia habitat. The remnants are still under threat and a mindset that has developed in the road construction and maintenance arena is that roads must be clean and scraped fence to fence. Similarly there are farmers who want every square inch under control and in subservience to their production needs. Dense vegetation is abhorred and burnt to control predation of sheep by jackal and lynx. Vegetation adjoining crops is treated with weedkiller to minimize crop contamination. Crops are also grown in conjunction with animal production. When crops are in, the animals are on fallow land and on whatever is left of natural vegetation. It is the harsh reality of conservation.
Haworthias exist on those remnants. I have often pondered that maybe they have not been so severely suffered by habitat destruction but now I am not so sure. There are many remnants where nothing is to be found, and others are species rich for no obvious reason. It is therefore difficult to assess.
This chapter explores the occurrences of two species at 11 localities in the Ouplaas vicinity east of Bredadsorp, and records some of their oddities.
The area in question:
This is shown in the map Fig 1 and I need to re-capitulate what I observed on Die Kop because I do not think I have given proper account for the populations I saw and the variance that I encountered in my first exploration there. This is most unfortunate as the name H. hammeri has since been applied to one or more of the variants and without context. Changes have taken place there too, that now compound the issue. I have in Vol. 3 Pt. 1 given an account of H. mirabilis in the wider area and will not repeat all of it, restricting myself just to those few populations that impact directly in these new observations. What is considerably more relevant is the chapter 13 in Part 2 where the main role players are listed and now again emerging in stark reality..
I have changed the arrangement from that in previous accounts to deal with local groups. Thus three sets, H. mirabilis at Die Kop – 5 populations and re-visits, H. mirabilis at Spitzkop, Haarwegskloof and Langvlei – 5 populations, and H. mutica at Spitzkop,- 3 populations.
Fig 1. Map of area and points covered
SET 1 – Haworthia mirabilis Die Kop
Subset 1 MBB7500 Die Kop E. Figs 2-8
Considering the wide array of populations and variants in the greater area, I concluded that these are most probably the result of interaction between H. mirabilis and H. mutica. The surfaces vary from shiny minutely tubercled to smooth, leaf tips rounded or pointed and flowering time is summer. This particular population is in a deposition area off the Silcrete inselberg.
Fig. 2 7500 Die Kop E
Subset 2 MBB7502 Die Kop N. Figs 9-10
When I first explored here in 2006 I skirted north of the first hill on Die Kop (see fig.1 Map) and observed an array of plants that were more like H. mirabilis and I dismissed them as such. However Jakub Jelimicky and Gerhard Marx both indicated that their observations contradicted mine.
Fig. 9 7502 Die Kop W
Subset 3 MBB7973 Die Kop N. Figs. 11-27
We went further north on this excursion passing through the area touched on in 2006 that was now dramatically different. The previous shrubbery was absent and the area was trampled and grassy with also Bobartia, a large grass-like Irid. It seemed unlikely that Haworthia previously there could have survived, but we had seen them before still further west and they may still be present. This new area we saw was densely populated with plants similar to but smaller than MBB7500.
Fig. 11 7973 Die Kop N
Subset 4 MBB7503 Die Kop mid. Figs. 28-34
My memory of these plants, east of those above, was that they inclined more to H. mirabilis than plants from 7500. Fig. 18 is almost identical to a clone photographed at 7500.
Fig. 28 7503 Die Kop mid
Subset 5 MBB7974 Die Kop mid. Figs. 35-46
Change due to heavy rain and erosion was more apparent here. There was less influence of H. mirabilis and the resemblance to 7500 nearly complete although the plants were generally smaller. This may be because the substrate was more skeletal and eroded than at 7500. What was really curious was the plant pictured figs 45-6 with narrow slender leaves. We did not observe any other species there that could have generated this as a hybrid but we did not have opportunity to comb the entire area small as it is.
Fig. 36 7974 Die Kop mid
Subset 6 MBB7504 N Die Kop. Figs 47-55
This population is about 1km north on an isolated inselberg. The plants are small and more typically mirabiloid with rough surfaces and spined margins and keel. Fig. 55 shows a plant with characteristics of the 7500 plants, but otherwise I would say this population perhaps relates better to Tarentaal MBB6539 (see Updates Vol 3.1 note 17).
Fig. 47 7504 N Die Kop
SET 2 – Haworthia mirabilis Haarwegskloof and Spitzkop.
Subset 7 JDV86-2 Haarwegskloof. Figs 56-63
This subset with subset 11, are included as a reminder of what other populations in the area are like. Originally when so much less was known, this was classified as H. heidelbergensis var. scabra and subsequently so many variants have been found that naming each is not deemed practical by me. It is probably 3-4km west of the following three populations and similar populations occur about the same distance further south at Kathoek and Beyersdal. There are other populations known to me that I have never sampled or photographed and the following task is to explore a large area between Bredasdorp and Spitzkop.
The plants at Haarwegskloof, at the time the pictures were taken in late spring, were both with translucence and without. The plants were small and with leaves erect.
Fig. 56 JDV86-2 Haarwegskloof
Subset 8 MBB7964 Spitzkop West. Figs 64-81
Fig. 64 indicates how difficult it is to find these small plants until you know exactly what you have to look for. One starts the search walking and eventually end upon all fours! Fig. 65 also shows how cryptic the plants can be. The plants are some of the smallest I have ever seen. Fig.72 shows an unusual plant with rounded tips to the leaves and I will show this again in another subset. Why this is so significant is because if the H. floribunda similarity in so many of the populations in the general area. The photographs disappointment me in that my first reaction on seeing the plants, was the association with images of H. parksiana. The pictures do no seem to fully reflect this. Kobus Venter has related his experience of finding H. parksiana-like plants among seedlings of H. mirabilis ‘magnifica’.
The leaves are reflexed as in H. parksiana as they of course are in many populations of H. mirabilis ‘maraisii’. But it is a curious reminder of the floribunda-like leaves in so many populations and the problem of look-alikes.
The plants were on a south slope in an area with white quartz and we had identified this as the most likely place to find them out of the direct north or northwest aspect. But we then did find them there too.
Fig. 64 7964 Spitzkop West
Subset 9 MBB7965 Spitzkop West. Figs 82-86
The plants were again hard to find over quite an extensive area. Fig. 83 again shows the floribundoid leaf tips.
Fig. 82 7965 Spitzkop West
Subset 10 MBB7966 Spitzkop West. Figs 87-103
It is curious how these small localized quartzitic areas can be so different. This was about 500m further north. There was more kaolin present as well as fine gravel. At an intervening similar habitat we found only Acrodon demidiatus, absent from the previous habitats and also from this one where Gibbaeum austricola now appeared. Again all the plants were small with dark green leaves, virtually no translucence and surfaces ranging from mildly spiculed to only minutely tubercled or just rough. Figs 99-101 are of plants with distinctly floribundoid leaves.
These are in white quartz and Aloe ferox is also present. There is moss present which is not the case at Spitzkop but true of Haarwegskloof. Curiously the pinched leaf ends as in fig. 106 have been observed in some plants of H. mutica too.
Although a small area of quartz in a cultivated field, the plants are densely concentrated in small areas. There were some dense clumps in the rocks and this is unusual for H. mutica. It is the variation in these plants that deserves close attention. Some plants have very rounded leaves, they can be quite elongate or short and squat, others have an end-awn, the leaves may be opaque or even translucent; the surfaces may be shiny and minutely tubercled or smooth. Colour may be reddish, golden, or green. Most striking is perhaps the venation. There may be anything from 3-13 straight lines or very anastomizing (veins joining). There is no sign of either true turbercles or surface spination and also none of marginal spines.
Fig. 109 7967 Spitzkop (quartz rock)Gasteria carinata is often severely animal grazed.Looking SE towards Die Kop and PotbergAlso looking SE towards Die Kop and PotbergViewing site from the north
Curiously there is a shale ridge (fig.149) between this population and the first that one would think would be suitable habitat for this species as it is very like the Ouplaas habitat. But they were absent, perhaps because the ridge was lower down the slope. But a little higher where the shale was also exposed in smaller ridges the plants were also absent. Where they did occur was among small quartz pebbles to which the quartz may fragment. Fig. 157 shows a plant that was under the protection of small shrub and shaded.
Plants absentPebbles and plants presentThis is not how one expects to see this species.Fig. 152 7968 Spitzkop (pebbles)Showing the small shale ridgesLooking to the previous habitat of larger quartz rock
Subset 14 MBB7969 Spitzkop East. Figs 160-161
This locality is about 3km east of the previous two at a slightly lower altitude. There is very little quartz but well exposed shale creating apparently suitable habitat as at Ouplaas now a short distance away. The area is well utilized by grazing animals and was very weedy, which may account for the fact that we found only two plants and these were not in the shale where we would have expected them.
Fig. 160 7969 Spitzkop East
Conclusion I must here again relate the story in respect of KG35/70 of which there is only an herbarium specimen. That was found more than 40 years ago at Verfheuwel about 15km eastwards from Spitzkop. I was scrambling up a rough animal track to the top of the Verfheuwel hill when I saw some dried seed stalk less than 10cm high. The plants were tiny, greyish leaved with marginal teeth and rounded tips. I simply did not know what to do with them and identified them tentatively along with a few other populations from that wider area as H. maraisii. I went back to look for them several times with no luck. In 1996 I was with Kobus Venter and we found a strong population of larger plants not far away on the same hill. I recognize now that those first plants I saw were floribunda-like and that this similarity is commonly seen in populations of H. mirabilis in this area.
I speculated in Update 3 about the origins of H. mutica in the context of H. mirabilis and I do contend that these three H. mutica populations show characteristics that support this view. The Die Kop populations suggest to me a more recent amalgamation of H. mutica with H. mirabilis, to virtually reverse the process. The floribunda-like characters and even the parksiana resemblance also support this image of very close common origin and that particularly H. floribunda is simply a polytopic phenomenon in this south western area (polytopic – similar elements arising independently from the same common gene pool) and its genetic resources are embedded in H. mirabilis particularly. This seems to be definitely the case in the greater Swellendam and Heidelberg areas.
What is also so confounding is that now we have H. mutica occurring virtually undifferentiated in several different kinds of habitat. This is quite unlike the Van Reenens Crest area where similar habitat does not even have the same species. The H. mirabilis at Spitzkop west is unlike the other small plants in populations of H. mirabilis in the area and does seem to be differentiated according to habitat.
Acknowledgement: Mr. Johannes and Pat Uys of Die Kop, Mr. Vlooi and Adele duToit of Die Kop, Mr. Gawie de Wet of Spitzkop (West), Mr. Franko de Wet of Spitzkop (East), Mr. Koos Badenhorst of Langvlei and Mr. Martin Mynhardt of Haarwegskloof. ♦