Reprinted from “The Journal of South African Botany”, vol. XXVIII, part III, July, 1962
A NEW VARIETY OF HAWORTHIA LIMIFOLIA FROM NATAL
M.B.Bayer, College of Agriculture, Cedara.
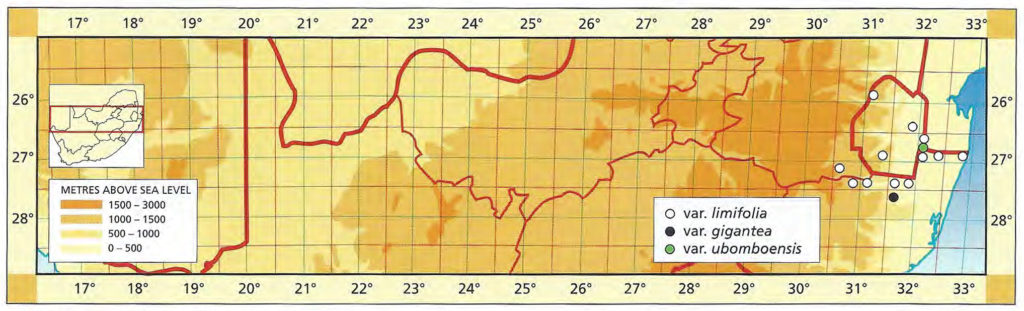
The typical form of Haworthia limifolia is from somewhere west of Delagoa Bay and was described by Marloth (1908). Prof. R. H. Compton has kindly sent me specimens from the Umbuluzi River Valley north of Stegi and also a form from near Gollel on the southern border of Swaziland. Herbarium records show that forms also occur near Barberton; the confluence of the Pongola and Pivaans River 20 miles east of Paulpietersburg; Ndumu, Zululand; Kosi Bay, and the Ubombo Mountains in Zululand. A photograph of a plant collected by Mr. E. R. Harrison 6 miles west of Somkele on the slopes of the Nkonyana Mountains, matches plants collected by me at Ngceba, north of the Black Umfolozi River, between the Umfolozi and Hluhluwe Game Reserves. This form is also reported to occur on the nTsuzi and Ulumbi hills which flank the Umfolozi Reserve on the north and south. Two plants have been collected from this area by Mr. H. Dent. Mr. A. F. Bruyns-Haylett also records it from the Mozaan River Valley between Swaziland and the Pongola River (fig.2.)
49. Haworthia limifolia Marl., Trans.R.Soc.S.Afr. 1:409(1908). Bayer :129(1976). Bayer :69(1982). Scott :31(1985). Smith, Fl.Pl.Afr. 55:24 (1997). Type: W. Delagoa Bay, Marloth 4678 (PRE): H. limifolia var. diploidea Res., Feddes Repert.Spec.Nov. 48:(1940). Type: ex Lisbon. Not preserved: H. limifolia var. tetraploidea ibid. Type: ex hort Lisbon. Not preserved: H. limifolia fa marlothiana Res., Bolm.Soc.Broteriana 14:200(1941): H. limifolia var. marlothiana Res., Mems.Soc.Broteriana: Succ.Afr. 3:93(1943). Type: ex hort Lisbon. Not preserved: H. limifolia var. schuldtiana ibid. Type: ex hort Lisbon. Not preserved: H. limifolia var. stolonifera ibid., Mems.Soc.Broteriana: Succ.Afr. 3:94(1943). Type: ex hort Lisbon. Not preserved: H. limifolia var. stolonifera fa pimentelli ibid. Type: ex hort Lisbon. Not preserved: H. limifolia var. stolonifera fa major ibid. Type: ex hort Lisbon. Not preserved: H. limifolia var. keithii Smith, JS.Afr.Bot. 16:4(1950). Type: Swaziland, Smith 5693 (EL, NBG).
limifolia: file‑like leaves.
Rosette stemless, slowly proliferous, with or without stolons, 5-7cm φ. Leaves 12-30, light to very dark green and even brownish-green, opaque, ovate-lanceolate, spreading, scabrid with white or concolorous tubercles, or with confluent transverse ridges, margins and keel scabrid. Inflorescence slender, to 35cm. Flowers slender, tepals fused and with revolute tips.
1982 – H. limifolia was originally recorded from west of Delagoa Bay but there does not seem to be any confirmed record of this species in Mocambique itself. It is reported to be in the vicinity of Maputo. It is quite well recorded from Swaziland in the eastern mountains. and from Gollel in the south-eastern corner. Other records include Barberton, Ndumu, Umfolozi and Paulpietersburg. It is clearly widespread and the populations differ markedly. It propagates very readily from stolons given optimum growing conditions, as do both H. nigra and H. venosa ssp. tessellata. There is a record of an unusual white‑tubercled form from the Mozaan River in the south‑eastern Transvaal. The var. gigantea is a large variety with small raised tubercles as opposed to the transverse ridges characteristic of the species. The var. ubomboensis is entirely glabrous. However, it has never been recollected at its purported locality east of Stegi and thus remains a mystery. The plants are apparently sought after for their purported medicinal and magical properties and may thus be heavily exploited.
1999 – Despite a very wide distribution range, the species is really poorly known. The herbarium record is fairly comprehensive but the species remains virtually known only from single plant collections from each locality. The var. ubomboensis is apparently from the locality south of Stegi as indicated to me personally by Capt. Keith. This is no doubt also the same general vicinity from which the var. keithii also came. An interesting collection is that of a glabrous light green variant similar to var. ubomboensis from the mountains near Barberton. This seems to be a more geographically discrete element than the original from south of Stegi. Also interesting is the olivaceous and stoloniferous variant reported to have been collected at Hoedspruit. A striated variant in which the tubercles are white and almost confluent in transverse bands was recognised by J. Pilbeam (1983). This does have some substance and appears to have been collected in the Pongola area and specifically along the Mozaan River, a tributary of the Pongola River.
a. var. limifolia.
Distribution: 2531 (Barberton): Saddleback Mt. (-CC), Stayner in KG33/71 (NBG); Near Pongola (-CC), Ross-Frames 13436 (PRE). 2631 (Hlatikulu): Mbuluzi gorge, Blue Jay ranch (-BD), Buitendag 1145 (PRE), Bayer (NBG); Vimy Ridge (-CD), Karstens in PRE39431, in NBG56413. 2632(Bela Vista): Stegi (-CA), Keith in PRE39436, PRE39438; Ubombo Ridge, 13km from Stegi (-CA), Keith in PRE34862, in PRE61130, Smith 5693 (NBG); Ubombo N. of Pongola (-CA), Keith in PRE34863; 16km S. Stegi (-CA), Keith in PRE61128 in PRE61129; Abercorn Drift (‑CC), Keith in NBG69412, Smith 5740 (NBG); Ndumu (-CD), Symons in PRE7532; Kosi Bay (-DD), Cronwright in PRE34875. 2730 (Paulpietersburg): Mozaan River near Moolman (-BA), Smith 7214 (NBG); 30km E. Paulpietersburg, Omdraai (-BD), Roos 4362 (PRE). 2731 (Pongola): Slopes of Ubombo N. Pongola (-AC), De Wet in PRE34874 Near Pongola (-BC), De Wet in PRE34858; 7km W. Gollel (-BD), Dhlamini in NBG646/59. 2832 (Hluhluwe): 8km W. Somkele, Smith 5692a (NBG).
Inadequately located: Zululand, Delagoa Bay, Marloth 4678 (PRE); SuldoSuavo, road to Moamba, Carvealho 755 (NBG).
Haworthia limifolia var. limifolia. Origin unknown. A very dark form, possibly from the Komatipoort area.
Haworthia limifolia var. limifolia JDV87/110 Mbuluzipoort. A stoloniferous and proliferous collection from forest floor habitat.
b. var. gigantea Bayer, JS.Afr.Bot. 28:215(1962). Bayer :118(1976). Bayer :112(1982). Type: NATAL‑2731(Louwsberg): Nongoma (‑DA), Bayer 112 (PRE).
gigantea: very large.
The location of this variety is obscure and although I have occasionally seen large forms with the same small discrete tubercles which characterize it, I have not been able to determine their geographic origin.
Haworthia limifolia var. gigantea west of Pongola. This form with the upright and incurved leaves is well known. A form with coalescing tubercles was informally described as the var. striata by J. Pilbeam.
Haworthia limifolia var. gigantea MBB112 north of Natal. With tubercles rather than ridges, this form can grow very large.
c. var. ubomboensis (Verdoorn) Smith, JS.Afr.Bot. 16:3(1950). Bayer :164(1976). Bayer :69(1982). Scott :33(1985). H. ubomboensis Verdoorn, Fl.Pl.Afr. 21:t818 (1941). Type: SWAZILAND‑2632 (Bela Vista): 16km S. Stegi (-CA), Keith in PRE26392.
ubomboensis: from the Ubombo Mts.
This variety has its origin in several collections from near Stegi (including probably the var. keithii) which seem to indicate that it has no geographic base. However, a very similar bright green glabrous form has also been collected by E. van Jaarsveld near Barberton.
Distribution: 2632 (Bela Vista): 16km S. Stegi (-CA), Keith in PRE26392, 26392b.
Haworthia limifolia var. ubomboensis Roux sn. Barberton. This is a very vigorous handsome grower which is also stoloniferous. A useful introduction to end the monotony of the clone predominant in cultivation.
This chapter was published in Aloe 40:2:41-52, 2003, and I have added an addendum with further explanation and discussion.
My first experience of this species Haworthia limifolia, was with a plant brought from somewhere in Zululand by my late uncle Frank Bayer. He was involved with malaria control in then Zululand and Northern Natal. He must have given the plant to my father while we were staying at New Hanover during the period 1950 to 1954. I have always been attracted to Haworthia, but I did nothing more than admire the plant as it flourished in my mother’s care. I never really concerned myself with the origin of the plant other than recalling my uncle telling that he had stopped for lunch by the roadside, and his Zulu assistant had come back to the car with the plant. I had thought he said “near Nkandhla”, while my father later doubtfully remembered “Nongoma”. It could have been “Ithala”. I recall now that my uncle was stationed at Vryheid during that period and this is a small way inland from either Ithala or Bivane Dam, which feature later in this article.
It was only when I had left University that my interest really developed further. By this time the plant had divided and there were two very big plants each measuring 23cm in diameter. I was working at the Cedara Agricultural College when I decided I should try to learn more about this species and perhaps describe the plant(s) now in my possession. It was obviously quite unlike anything pictured that I was aware of, and certainly the leaf surface did not match that illustrated with the original description of H. limifolia by Marloth in 1908 (see Fig.1). His species was supposed to have come from simply “west of Delagoa Bay”, leaving it to the imagination as to when one switched to “east of Walvis Bay”. Certainly in the way Haworthia were regarded as species, these plants of mine were decidedly different from what I perceived to be H. limifolia. It must be pointed out that no one has ascertained just where Marloth’s species originated, or decided just what should constitute the geographic origin or range of the typical variety. This is a classification problem which is not adequately catered for, and Brummit’s paper (2002) may indicate why.
Fig. 1. H. limifolia var limifolia Marloth as illustrated in Trans. Royal Society S Africa.
A colleague of mine was good enough to accompany me on a trip to look for the species in 1962. We started at the Hluhluwe and Mkuzi Game reserves. We had a very adventurous journey that included being shown a place where a plant had been known to grow. Another experience was to be directed into the veld to occupy us while the police were called to query our presence. What puzzles me now in retrospect is that in my later description of the large H. limifolia var. gigantea (figs.2 & 3), which we never found), is the record in that description of a collection I made at Ngeba just north of the Hluhluwe reserve. I have no recollection of such a plant whatsoever.The big disadvantage my friend and I suffered from on our journey was the inability to communicate with locals. A bigger disadvantage was my sheer ignorance of field collecting and the preparation and effort needed to produce results. During our fruitless journey we went as far north as Stegi (now Isiteki) where I met Capt. D.R. Keith. We also saw and collected plants on the Blue Jay Ranch on the Umbuluzi River to the north. Capt. Keith had collected a plant named for him as the var. keithii (fig.4 – note: a clone, fig.5 in cultivation in J.W. Dodson’s collection as a topotype of the var. keithii seems to differ quite radically from the illustration accompanying the description by G.G. Smith. It is a plant with shorter squatter leaves than the illustrated type). Again I did not record properly the origin of that variety – my notes state “Palata Farm”. This is approximately 12km southeast of Stegi, and the farm Ravelston a little further on. Capt. Keith was also the collector of H. ubomboensis (later H. limifolia var. ubomboensis, figs.6 & 7) from south-east of Stegi. This could have been a single clone in an otherwise tubercled population, as several dried specimens from that general area are in the PRE herbarium.
Fig. 2. H. limifolia var gigantea, MBB112 Nongoma, as illustrated in Journal of SA Botany 1962.Fig. 3. H. limifolia var keithii GGSmith, as illustrated in Journal SA Botany 164, 150.Fig. 4. Haworthia limifolia var gigantea currently in cultivation. Note spreading leaves with discrete tubercles, dark coloration and thin leaves.Fig. 5. J.W.Dodson 2081 H. limifolia var keithii topotype. Note fewer broader and shorter leaves.Fig. 6. H. limifolia var ubomboensis. Clonotype in cultivation. It is not known if this variety has any population basis.Fig. 7. H. limifolia var ubomboensis. The same clone when stressed.
Over the years I simply left H. limifolia to history – for which there is some reason. Nature Conservation authorities made it painfully obvious that they were the only true custodians of nature. Nevertheless I did see several plants of H. limifolia in collections, such as:
a. the Mozaan River plant collected by Bruyns-Haylett (fig.8), illustrated with my description of the var. gigantea.
b. a plant from Barberton collected by Frank Stayner (fig.9).
c. a plant from Gollel mentioned in my varietal description (see fig.10, which is ex hort. Mr. Roux),
Fig. 8. Bruyns-Haylettsn. H. limifolia var gigantea (striata nom. nud.) Mozaan River, Pongola.Fig. 9. F.J.Staynersn. H. limifolia var limifolia. Barberton.Fig. 10. Rouxsn H. limifolia var limifolia. Gollel.
d. a few clones kindly sent to me by Mrs Tony Pooley then at Ndumu which I was sure she said had come from near the entrance gate – subsequently refuted (personal communication).
f. a plant in the garden of Mrs Mairne Hulme (fig.11) near Richmond, Natal. This was a very dark-green plant with distinctive raised wavy undulations across the leaves. These undulations are unquestionably not tubercles in the sense of any departure from the colour or texture of the surrounding leaf surface. Thus we have evidence that Marloth’s assertive “non-tuberculata” is a reality. In my revision (1999), I illustrate a plant from the Umbuluzi gorge which can be seen also to be “non-tuberculate”.
Fig. 11. H. limifolia var limifolia ex Mrs Mairne Hulme, Thornhill, Richmond
I saw plants reported to have been collected at Louwsburg which were rather like the big var. gigantea with the same tubercles on the leaves, rather than the transverse ridges we usually associate with the species. These plants have been distributed as var. gigantea, but they do differ in having thicker leaves which are sub-erect so that the rosette is more upright.
At one stage I also had a plant from the place Goba in the north-east of Swaziland again collected by someone from Johannesburg. There were also other odd plants with no history whatsoever. I made one further trip to the Pongola area in about 1980 to be shown H. limifolia by Denis DeKok some 20km west of the town. This is a rather tuberculate plant, the tubercles somewhat whitish and the leaves form a fairly closed, almost rounded rosette (fig.12).
Fig. 12. MBB sub Denis DeKoksn H. limifolia var gigantea. 20km W Pongola.
A collection with no good history also surfaced ostensibly from near Malelane east of Nelspruit. The same collection has been cited as from Komatipoort and Hectorspruit. I saw it in the Pretoria National Botanic garden in 1996 (cited from Hoedspruit!), and plants obviously from the same place have been sent to me from the USA – having arrived there by courtesy of ‘friends of a collector’. This population is still extant and known to Nature Conservation. The plant has been described as H. limifolia var. arcana (figs.13, 14 & 15) by G.F. Smith and N. Crouch (2001). A population nearer Barberton was said to have been annihilated by medicine collectors.
Fig. 13. H. limifolia var arcana. Hectorspruit, Ex Pretoria NBI.Fig. 14. H. limifolia var arcana. Ex J Hurter, Nelspruit.Fig. 15. H. limifolia var arcana. Ex Bridgevale Nursery.
From the USA (Peter Alonso), I received a plant from a collection by John Lavranos from Bumbeni “on the southern end of the Lebombo mountains” (fig.16).
Fig. 16. J. Lavranos8387 H. imifolia var gigantea. Bumbeni, Hluhluwe.
This is a very handsome plant with striking white markings. It appeared from John Lavranos’ own communication that the population was now extinct, as was said to be true for another collection he once made near Barberton. These are the extracts of letters by John Lavranos concerning these collections of his:-
re Bumbeni…
It is thrilling to know that a plant collected May 31, 1971 is still in cultivation and proof, if this were really needed, of what contributions the hobby can make to conservation, in the true sense of the term! This plant was collected near a place called Bumbeni, on the southern end of the Lebombo Hills, a range 300 miles or so long, which forms the border between Mozambique and S. Africa (Swaziland) and extends south into Natal. It was growing on very rocky east-facing slopes. The climate is very warm with occasional light frosts at night, in winter. Plants were quite common in 1971. However, all have now disappeared it seems. The reason for this was certainly not commercial collecting for the hobby but rather commercial collecting for medicinal purposes, as the plant is prized by African traditional medicine. Your plant, I am afraid, is now a true museum specimen the more so as the form does not seem to sucker. At the time it was identified as var. keithii. The plants varied substantially. The only thing they had in common was their large size and the fact that I never saw one that suckered. One used (upto the mid-60’s) to be able to buy such plants in the African medicine shops. But I have seen none in the last 30 odd years. They were collected out for the medicine trade.
1. The S. end of the Lebombo hills, near Bumbeni is composed of Rhyplit lava, which is acid. The soil is a scanty mixture of quartz sand and humus.
2. Plants in the wild were solitary. We found none with offsets or suckers. My collection number refers to a number of clones. These were all growing in a limited area.
3. I only collected the species once more, in 1956, east of Barberton, but doubt if it is surviving in cultivation.”
re Barberton…
* “The Barberton limifolia was as you describe it. It was collected on 10 June 1956 above Barberton, on the way to Havelock Mine. I remember finding one single large clump of it. I searched again many times in ensuing years but never saw it again. But of course those hills have been heavily grazed and frequently burned since 1956″.
Charles Craib (writes in the literature of plants he collected east of the Mashongololo mountains near the Paris (Bivane) Dam, north-east of Vryheid (figs.17 & 18). Crouch et al. (1999) record “three small populations”, two of which are within the Mkhuze game reserve. They would have been aware through Geoff Nichols of the third at the Bivane Dam.
Fig. 17. C.Craib H. limifolia var gigantea. Mashongololo Mts. environment. There appear to be plants with discrete tubercles as well as with bars.Fig. 18. C. Craibsn. H. limifolia var gigantea. Mashongololo Mts environment. In this plant he tubercles are concolorous.
A dramatic find was by Fanie Venter, ostensibly from near Barberton (Three Sisters) of also a smooth-surfaced, lime-green coloured plant (fig.19) very like the var. ubomboensis. The leaves are seemingly more glaucous, and the plant is bigger – also producing more stoloniferously rather than the close offsetting of var. ubomboensis.
Fig. 19. F.Venter13700 H. limifolia var glaucophylla. Three Sisters, Barberton.
What is now most disconcerting about H. limifolia is the fact that it is regarded as highly endangered. Crouch and Smith (1999) report an unbelievable 22.5 tonnes of H. limifolia traded through Durban markets per annum – an estimated 479 000 plants. Although these authors give places like Kanyayo, Inanda, and Espofu as sources for these plants, no attempt seems to have been made to locate plants in the field. So the “recovery contribution” in the title of their article is limited to “three small populations” which are all within protected areas. This is not in the least surprising considering the logistic problems of searching other areas thoroughly, and probably the greater problem being of moving freely through a politically and socially divided, and troubled area. Actually it is very difficult to make a search at all when working from established roads rather than according to terrain. Also there is no conventional structure of land ownership in a large part of Kwazulu Natal.
Crouch et al. (1999) suggest that the local and inconspicuous nature of the species may have resulted in under-collection and poor representation in herbaria. This is in stark contradiction to the volume of plants that is reported to be collected each year by the locals. Actually the species is fairly well represented in herbaria by the ordinary standard for succulent plants which do not lend themselves to making herbarium specimens – and better than for many other Haworthia species. It is probably only the fact that H. limifolia is away from the traditional succulent-rich areas, that collectors have not paid more attention to it. It is additionally curious that the species is so poorly represented in collections, particularly in regard to label documentation. I never found it an easy species to grow, although I must have propagated many offsets from the leaves of my original gigantea, and still have such a clone grown from the original collection. Some of the older limifolia collections were identified as varieties of H. pumila or H. minima, and similarly a plant of Aloe aristata from the Drakensberg was once thought to be a relative and given the manuscript name of H. natalensis.
Where a species like this is regarded as critically endangered, it certainly seems unwise to disclose known localities. However, whether this is true or not, I actually think it is counterproductive and against a basic ethic of science – no secrecy. Applications for permits to collect plants are granted with reluctance, and many collectors simply ignore the whole process. The consequence is that information from people who are interested in the plants and who can and do locate them in the field is not fed to where it matters. There is confidentiality about locality records and there should perhaps be more discussion about the merit of this secrecy. On the one hand it does nothing to preclude people from looking for the plants, and on the other everything to keep new information concealed. It overlooks and ignores the fact that it is motivated individuals who have contributed so much to the knowledge of our flora. It also minimises the fact that the plant is known virtually only in nature reserves.
Nature Conservation is perceived to keep an extremely strict and close control over permits in a “hands-off” approach. The encouragement for private individuals, or anyone interested in nature for that matter, to pursue any interest is almost non-existent. The converse is probably true, and so severely so that it must encroach on constitutional rights. A consequence may thus be that there appears to have been no tangible or noticeable effort to locate and assess the conservation status of H. limifolia outside of existing nature reserves. My own report of the species in the Hluhluwe/Umfolozi corridor in 1962 has gone unquestioned although this area is now included in the enlarged consolidated reserve. Ironically while permits have been so parsimoniously granted, field collected plants have been flooding the medicine markets. It does not seem to make sense. For forty years it has been almost impossible for anyone to have legally collected plants of H. limifolia, and now they seem to be regarded to occur only in Nature reserves! Is, and was, anything achieved? It may have been far more sensible to have rather embarked on an educational mission to emphasise the value of sustainable activity and the desirability of maintaining a genetic bank of material in cultivation which is adequately recorded and curated. Thus conservation could be seen as for people, and not a mission to dissuade them from nature as happens now. It should be said that as an impersonal public institution, Conservation Authority is perceived to be heavy and high-handed. The absolute converse applies when one is able to deal with individuals in such organisations and speak on a basis of common interest.
At present the known approximate locations for H. limifolia appear to be these.
Fig. 23. J. Bronkhorstsn H. limifolia var giganea. Nr Pagamisa, Magud, SW Pongola.
Armed with some of this information I began to contact interested parties to see if I could set about verifying the continued existence of these populations, less hopefully expand the known record, and build on the report in Aloe (1999). I am rather critical of this last cited paper where I thought the authors had failed to even have made an effort to examine the position in the field, if only in respect of the localities cited above, which are all within the general knowledge of the species. My interest was also driven by the fear that the threat to the continued existence of the species in the wild is real, and that there is a need for ex situ conservation and the establishment of a gene bank of some kind. My past experience has not led me to believe that such ex situ conservation actually is workable – and it will not be into the future unless it is done with very strong attention to identification of the genetic elements in any collection and assurance of continuity. This applies also to institutions such as recognised Botanic gardens where my long experience has shown that their collections are too closely tied to temporary incumbents, and that management has had no real direct interest in specific management of endangered species.
Geoff Nichols kindly reported on his experience with the species in respect of the medicine trade and the plants known to him. Dr Pete Goodman and Rob Scott Shaw of Ezemvelo KZN Wildlife were also most forthcoming with information regarding known populations at Mkhuze, Paris Dam and Ithala. Similarly Mervyn Lotter and Gerhard Strydom of the Mpumalanga Conservation authority offered their co-operation in respect of the Lowveld. A surprise was to learn of Errol Harrison who had collected the specimen cited for “West of Somkhele” near Hluhluwe in 1956 and who was alive and well at Mtubatuba.
Contrary to what I have written about Conservation attitudes above, the Ezemvelo and Mpumalanga authorities issued me with the necessary permits and my excursion was assured. Several other factors precipitated the decision to go, such as the support of my family, offers of accommodation at Barberton, and a 50-year school reunion back in Natal.
My wife Daphne, and I, started our Haworthia epic at the Queen Elizabeth Park headquarters of Ezemvelo KZN Wildlife at Pietermaritzburg. Here Rob Scott Shaw kindly explained the position regarding the only plants they had in cultivation there. This was part of a consignment confiscated from poachers into the Ithala Game reserve north of Louwsberg. Some of these plants had been planted in the open at the Park in Pietermaritzburg where they had flowered well and set seed (figs. 24 & 25). Brigette Church had very successfully germinated and grown that seed. I was allowed to take offsets off the mother plants and, in retrospect, I foolishly did not ask for any of the seedlings. None of the plants were quite like the var. gigantea and I should have considered the variability of the cultivated seedlings more closely.
Fig. 24. MBB7148 H. limifolia var giganea. Queen Elizabeth Park ex Ithala Game reserve.Fig. 25. MBB7148 H. limifolia var gigantea. Queen Elizabeth Park ex Ithala Game Reserve.
At Durban I visited their municipal Bridgevale Nursery in the company of Geoff Nichols. Mark Gillmer was the propagator who had done a marvelous job with plants bought at the local herb market. The plants were rather similar to the Ithala plants and I obtained six clones (figs.26, 27, 28, 29, 30 & 31). Only one of these (fig.31) could I really relate to my original var. gigantea in having the thin leaves, many small concolorous individual tubercles and potential for size. However, the leaves are thicker and more erect than I would have accepted. My conclusion was that, like the Louwsberg (immediately south of Ithala) plants referred to above, the var. gigantea could have originated in that general vicinity but the material I have seen is not sufficiently convincing.
Fig. 26-31. MBB7136 H. limifolia var gigantea. Bidgevale Nursey, Durban Municipality.
From Durban we traveled to Mtubatuba to meet Errol Harrison who had told us that the Somkhele area had now been vastly built up and it was improbable that plants would still be there. We did thus not explore the possibility of plants there or in the now greater Hluhluwe game reserve. This should be done. Errol took us to Phinda where we were disappointed to find that this Resource Reserve did not include the area we needed to see. However, by really good fortune we did find the owners of Pumalanga Game Ranch at home, Mr. and Mrs. Mannie van Rooyen. With the goodwill and kindness that characterised all the people we met on our journey, they took us to a site bordering on the expanded Mkhuze Game Reserve. We found the plants there without too much trouble, but there were only about 20 plants with a total of about 30 rosettes. There was an old ruin nearby which we were told was Bumbeni (a name derived from “bumba” for mud) and the stream which was the source of that mud for building. Thus it appears that this is in fact the same place where John Lavranos had collected more than 30 years before. (See figs.32 & 33 as well as fig.16).
Fig. 32. MBB7137 H. limifolia var gigantea. Pumalanga (Bumbeni), Hluhluwe.Fig. 33. MBB7137 H. limifolia var gigantea. Pumalanga (Bumbeni), Hluhluwe.
From Pumalanga, Daphne and I went on to Mkhuze where we were fortunate to find Mr M.Gumbe, Technical Assistant, who took us immediately on a long and high-speed walk in the failing light (hence no photograph) to find plants on Inxwala Hill. The plants are comparable with those at Bumbeni. Inxwala is one of the localities on Mkhuze reported in the Aloe (1999) article. From Mkhuze we travelled to Pongola where we again met most helpful people, but which did not materialise in the form of many collections despite our conviction that there must be many populations in the area. Why this conviction? From my experience with landowners in the Cape where there are many Haworthia species, people are simply not aware of small cryptic plants. Even with the local Zulus we found on our first trip in 1962, that generally people are just unaware of the plants. The same applies in the Cape where few farmers or their employees are conscious of what actually occurs on their property. Botanists have confused things like Aloe aristata and H. limifolia, and it is hardly surprising that less knowledgeable people make worse mistakes. This is what happened at Pongola and my opinion is that it is just a general level of nescience rather than ignorance, which clouds our perceptions of rarity. I make the distinction between two ignorances viz. when one is expected to know, and that where one is not expected to know.
Fig. 23. J. Bronkhorstsn H. limifolia var giganea. Nr Pagamisa, Magud, SW Pongola.
Johan Bronkhorst reported that there were plants (fig.23) along the road from Pongola westward to Pakamisa and Louwsberg. We stopped at a likely farm where we contacted the owner who in turn referred us to his foreman. This foreman took us to his home where he had a half-drum filled with soil and was growing “umathithibala” – the colloquial name. The plants consisted of H. attenuata, H. attenuata var. clariperla, H. radula and two Gasteria species. I must explain that at Pumalanga there was some confusion too, as apart from two lonely H. limifolia on the rockery, a huge container was filled with H. attenuata var. clariperla (better referred to as a look-alike of which more later). When we explained that we wanted to see the local “umathithibala”, the foreman said that was easy and directed one of the rangers to guide us up the hill to see it. This the ranger did most competently but the plants we were shown was a proliferous and abundant crassula!
We then ventured along the Pakamisa road where the terrain was most promising but simply inaccessible from the point of view of access and ownership – it would have taken more time than we had available to explore the area and determine ownership. Back at Pongola we were lucky to meet Kathy Cameron, a dynamic ex-teacher who was running the information centre. She put us in touch with two people. The first was the local herbalist selling her wares on the town pavement. There we acquired a guide who offered to take us to the township were the plants were. At this village of Ncotshane we were again shown a container, with six plants of H. limifolia consisting of at least three clones carrying seed. We were graciously allowed to take three offsets. But we asked to be shown where the plants had come from. In response a finger was pointed to the nearby hill and we set out, with some apprehension, to climb it. It was also to no avail and we saw nothing in rather unlikely terrain just as we had feared. Ncotshane would class as “north of Pongola”, and I have no doubt that on those many hills, the species does in fact still occur.
Fig. 34. MBB7140 H. limifolia var gigantea. Daaiwater, Pakamisa, Magud.Fig. 35. MBB7140 H. limifolia var gigantea. Draaiwater, Pakamisa, Magud.Fig. 36. MBB7140 H. limifolia var gigantea. Draaiwater, Pakamisa, Magud.
Through Kathy Cameron we also contacted Rynold and Rolene Steenkamp at Magud. Rolene very kindly guided us to where she had known plants on their farm Grootdraai near Pakamisa west of Pongola. There we found 10 plants among rock in a very grassy and somewhat wooded area (figs.34, 35 & 36). Rolene said that the population was far less vigorous than she remembered it and it did seem as if the surrounding vegetation was too competitive for H. limifolia. Time was now very limiting and we left Pongola with much left simply undone, and we travelled to Isiteki. Again we were assisted by really kind and gracious people – Jonathan and Helen Pons who own Mabuda farm. We initially tried on our own to explore the Palata/Ravelston area. This was frustrating in the extreme as the road passes along the escarpment but never approaching near enough to initiate an excursion. Often the vegetation was overgrown with aliens forming an impenetrable thicket while there was the same problem of land ownership and authorization to enter, presented by such communal area. Palata and Ravelston farms had been expropriated by the British Govt. to hand to the Swaziland National Trust on the granting of independence. So our visit there turned out to be no more than a preliminary reconnaissance and we were not able to assess var. ubomboensis. Later Jonathan Pons obtained for me two plants from the approximate area (figs. 37 & 38). Curiously the former does mirror the var. keithii in profile whereas Fig.38 does so the leaf surface.
Fig. 37. J.Ponssn H. limifolia var keithii. SE Isiteki, Swaziland.Fig. 38. J.Ponssn H. limifolia var keithii. SE Isiteki, Swaziland.
From Isiteki we travelled to Barberton where we were met by Mervyn Lotter of the Mpumalanga Parks Board. He took us to see the locality for var. arcana, which unfortunately for us had been fenced and we were faced by a locked gate. However, this was not a serious issue as the plants are well-described and there is adequate material in cultivation to ensure ex situ conservation (in fact it is being grown at Bridgewater nursery and I did acquire an additional clone there for my own record). We went then instead to Kaapmuiden and ended up on the farm Boondoks. We obtained permission to explore a little, and after one unsuccessful excursion Mervyn found a very small population of 84 rosettes in an area of about 2sqm (figs.39 & 40). The area had been burnt and the plants were quite severely singed. The plants do seem to prefer tight rocky habitat with sparse grass cover and so it would take a very severe fire to kill them. It was just very difficult to form an opinion regarding appearance of the plants and I could only surmise a resemblance to var. arcana.
Fig. 39. MBB7143 H. limifolia var limifolia. Boondoks, Kaapmuiden. Plants severely singed, suggest var arcana.Fig. 40. MBB7143 H. limifolia var limifolia. Boondoks, Kaapmuiden.
At Barberton we met Rhona Milstein, who has lived in Barberton all her life and is extremely knowledgeable about all things pertaining to the place. She guided us to a nearby hillside were it was thought the population had been collected out. Fortunately this was not true and we found many plants albeit in a small area again in closely fragmented rock and sparse cover (figs.41). The plants were again badly scorched. We saw the plants again at May Mine on the Mountainlands Game Reserve alongside the Havelock Road. This was in the presence of a joint owner Mr. Nico Oosthuizen, who had very kindly agreed to our entry to the site. The plants were very abundant over an area of about 150m x 10m but again in a very rocky area (figs.42 & 43) and also fire scorched. It was interesting to note the presence in Rhona’s garden of a very stoloniferus form of H. attenuata cf var. clariperla which I have not seen before.
Fig. 41. MBB7141 H. limifolia var limifolia. Rymer’s Ceek, Barberton.Fig. 42. MBB7142 H. limifolia var limifolia. May’s Mine, Barberton.Fig. 43. May’s Mine scene.
Our next excursion was to locate Fanie Venter’s (13700, fig.19) collection at Three Sisters. Again we were overwhelmed by the kindness and helpfulness of John and Bernadette Roux, owners of Three Sisters. They took great pains to show us where to find the plants and stayed in telephone communication while we drove along the tortuous rough road to the near summit of the Sisters (three peaks, with intimidating steep slopes which we would never have negotiated on foot). We found the plants on a nearer locality than indicated to us and there were many in a very small area. The vegetation was almost alpine and very dense. Fire here would surely result in the death of many plants, with survival only in rockier more sparsely vegetated mini-refugia (figs.44 & 45). From here we could look down on the Boondoks site and also identified a likely hillside to the southwest of this. We went there on our descent of the mountain and after a long walk we did find a very small population in an area of 0.5sqm. I refer to this population as Manders Dam. It may have originated from a single clone, which may be true for the Boondoks plants too. The plants were growing in the excavated old soil of a termite mound and badly scorched (figs.46 & 47). Their exposure to fire could not be considered to be more or less severe than had they been anywhere else in that terrain and my opinion is that fire per se is not a hazard to the species. In general if the fuel load is high enough to present a threat to survival of the plants, they are anyway subject to threat by exclusion of light and unfavourable growing conditions.
Fig. 44a. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton. 029Fig. 44b. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton.Fig. 45. MBB7144 H. limifolia var limifolia. Manders Dam, Barberton.Fig. 47. MBB7144 H. limifolia var limifolia. Manders Dam, Barberton.
However, there were odd inflorescences about and it was evident that no seed was set. Fire may affect the pollinators and naturally will also destroy inflorescences and seed if untimely. But this surely is a seasonal factor and there will be years when seed is produced. What one can speculate is that the populations are derived from vegetative propagation as the species does proliferate readily from stolon. If this is so they will probable be non-seed producing as plants are usually self-sterile.
It was sad to leave Barberton with so much to do and such promising unexplored terrain. The drive to Vryheid was also frustrating for that reason. There must be populations of the species in the area Badplaas, Osplaas (Gerhard Strydom of Mpumalanga Parks Board reported rumours of its presence there) on to Lochiel, Amsterdam and Piet Retief. At Vryheid we were taken to the Paris (Bivane Dam) where, pushed for time, we actually saw only one population (figs.48, 49 & 50), surrounded by most promising terrain. We went to Nongoma where we were introduced to a local herbalist and shown “Umathithibala”. This turned out to be a container with Aloe aristata from the Ngome Forest area, and also a plant of H. attenuata. The herbalist explained the use of “umathithibala” to defuse tension and disarm intruders with harmful intention. At Pongola we had been brought three other Aloe viz. bainesii, sessiliflora and komatiensis …) which were said to be grown around the home to tranquillise atmosphere. Thus the usage of the plants is not directly medicinal, but embedded in a paradigm of esoteric values. She did not know H. limifolia. To Jonathan Pons, the Swazi name for the plant is “sidodobala” and they apparently also use it to ward off lightning or else simply explain its use in this way as the Zulus do. The proliferation suggests its use too to encourage fertility in livestock and for that purpose it is grown around kraals.
Fig. 48. MBB7146 H. limifolia var gigantea. Bivane Dam, Vryheid.
Our last stop was at Ithala, which I had not intended to visit because the plants were familiar to us from the Park Headquarters at Pietermaritzburg. There we met the Conservation Manager, Mr. Rob Blok, and were shown plants in the office rockery. What was in the rockery were two plants which I think should not have been there. One was Aloe squarrosa (an exotic) and the other was H. attenuata. This latter plant could perhaps hybridise with H. limifolia. But the plants we were shown were Aloe aristata and said to occur in the Reserve too. As the site for the originally poached plants was on a distant area of the reserve, I asked if I could see and photograph A. aristata. Without the spines, these specimens were remarkably similar in colour and size to H. limifolia as it occurs in the greater Pongola area. We went to the area indicated and must have missed the specific point where the Aloe may well have been and saw instead some plants of H. limifolia which were very like the Paris Dam plants. Our impression was that the species must well be very common at Ithala and surely so between Vryheid and at least Pongola.
Conclusion The prime object of the trip was based on the assumption that the species H. limifolia was critically endangered and hence that there is proper motive to ensure an ex situ gene bank. Thus we collected within the limits of our permit authorisation for this purpose, and these plants will be distributed under accession numbers in due course as they proliferate. The second object was to try to extend knowledge of known localities from the impression given in Aloe (1999), at least to the record cited in Haworthia Revisited (1999) and evident in RSA herbaria. The third object was to derive some kind of estimate of conservation threat. The fourth was to arrive at a new opinion on classification for known and unknown material. Thus:
1. Material of 20 accessions is now held by me of which 12 can be regarded as seed reproducible. It would be very desirable to increase this.
2. The cited record clearly still exists and is largely confirmed. At least five of these new collections can be regarded as new. Hence if one considers the product of a virtually blind-driven effort, there must be many more populations which are still unknown.
3. The conservation threat from fire which has been cited seems to be fallacious, and the same may even be said for the ‘herbal’ trade. The species does not have a particular unique role in herbal medicine or mythology. Where the threat does lie is common to any living thing and far more applicable to any number of animals and other plant species – this is the density of the population living off the land and widespread degradation of the environment due to fire being used to generate grazing for livestock or simply to clear the landscape from vegetation which may harbour vermin, snakes and human intruders. Thus all organisms in communally owned land are at risk. There is a conservation plus in the large number of game ranches and reserves which are being game fenced and patrolled to stop poaching.
4. Classification of plants at least in respect of Haworthia is now bordering on the ridiculous. There are any number of would-be taxonomists all with their private versions of what constitutes species, how old names can be interpreted, what name changes can be implemented and what new names can be invented. As Stevens (2002) stated, botany has been taken over by nomenclature. The peer test of any taxonomic work is largely based on observance of the nomenclatural rules, the viability of the Latin diagnosis, and the protocol of literature and specimen citations. Brummit (2002) also writes that the binomial system does not really work. This has been evident in Haworthia for the last 40 years although here it could be largely the fault of the writers.
My opinion regarding H. limifolia is based on my view of how a species should be defined. This is as a dynamic system of living organisms which are morphologically and genetically continuous in time and space. In Haworthia I have found that the geographic element is essential to the understanding of taxonomic elements. In this respect there appear to be no real discontinuities between all the records cited above despite a wide variation between say var. arcana (in the absence of a population which we can say authoritatively is exclusively and decisively var. limifolia) and var. gigantea, which also has no true provenance. The var. striata could apply to any of the Pongola collections where white striations (as the continuous concolorous bars in var. limifolia) occur as frequently as there are small discrete tubercles as in var. gigantea. What is probable is that the name striata originates in the plants collected at Bumbeni by John Lavranos which were introduced into cultivation. There is no evidence that the single plant from Mozaan which I photographed in Bruyns-Haylett’s possession was ever propagated and distributed. I had simply suggested to John Pilbeam while he was writing his book (1983), that based on what I had seen, a varietal name “striata” was reasonable. I am of the opinion that the Mozaan plants are closer to my variety gigantea than they are to the Bumbeni (and Mkhuze) white-banded variants. Thus the varietal name gigantea could apply to the species as it is known south of the Swaziland border, and the name striata would apply to specific forms and cultivars which have confluent white tubercles or raised white bands across the leaves.
The var. ubomboensis has not been appraised as a population based entity and neither has the old var. keithii. What is evident is that there is another disjunct population of a similarly glabrous element at Three Sisters, Barberton, as FVenter13700 and MBB7145. This latter element is definitely population based and the two collections cited can be recognised as fairly discrete. Unlike the var. ubomboensis which is a smooth yellow-green colour in good cultivation, this Barberton element is a glaucous bluish-green and the name glaucophylla is given to it as follows:
Haworthia limifolia var. glaucophylla (Asphodelaceae: Alooideae) – var. nov.
Type: Mpumalanga, 2531(Barberton): Three Sisters (CB); F.Venter 13700 (Holo. NBG)
Very similar to the var. ubomboensis but glaucous green and geographically discrete.
In habitat small pale yellowish-brown in colour, rosette seldom exceeding 3cm diametre, 8-12 leaves reaching 6cm in length. Surfaces smooth, occasionally with some rounded or barred tubercles. Slowly proliferous. In cultivation very proliferous and off-setting, rosettes to 12cm in diam. with 25-45 leaves, colour glaucous bluish-green, hence the epithet. Differs from the typical in being without raised transverse lines, and from the var. ubomboensis in being bluer in colour and geographically discrete.
There appear to be a number of discrepancies in the description of the var. arcana. In the protologue the leaf surfaces are described as undulate but generally lacking distinct tubercles as in the typical variety. This is not correct. The typical variety was named limifolia, meaning file-like, for the very reason that there were these raised undulate transverse ridges on the leaves – not discrete tubercles. The var. arcana that I have seen generally has such broken ridges which reduce to individual and quite large tubercles. This is quite different to the many smaller discrete tubercles in say var. gigantea where there is no evidence of transverse bars at all. In many of the collections I obtained in the greater Pongola area, there were both continuous bars across the leaves as well as plants in which the leaves were tubercled instead. In these collections it was common for the leaves to be either erect, incurved, sub-erect or spreading. Regarding flowering time – Smith and Crouch state that in contrast to other varieties of the species, the var. arcana flowers in mid- to late winter. This is not true. The collections at both Queen Elizabeth Park and Durban had flowered prior to spring and flowering was completed at the time of our visit in late September. Similarly all the evidence we saw of flowering on our trip was that it had been completed before our visit. We saw late flowers at some sites and collected seed at Draaiwater and at Bivane’s Dam.
This raises the point that we seem to have a convention for how taxa should be described. In the var. arcana as for other Haworthia taxa, there is a long description of the flowers which is quite valueless in the context of the structure and variability of the flowers in the whole subgenus. It lends to the elaboration of the process to give it a validity which it earns from just the fact that the taxon is worth identifying by name for practical communication and comprehension. Furthermore it feeds a perception that this fine detail enhances the credibility and validity of the element when it does nothing more than lengthen the description. Similarly, rather than use an epithet which is derived from a skewed viewpoint of what is known and what is not known (viz. arcana – mysterious, secret) why not a descriptive one such as olivacea, highlighting the extraordinary colours this taxon can get in cultivation? There are still two other things. The mapped distribution of var. arcana is entitled “distribution range” and suggests an area of circa 2600 sq km. when in fact there is one known population occupying perhaps 0.25ha. The suggestion that it is one of more links to H. koelmanniorum is window-dressing with no other truth in that var. aracana is geographically the closest known of the limifolia variants to a not so climatically stressed H. koelmanniorum at Groblersdal and Loskop Dam.)
Fig. 51. Map of Mpumalanga, Swaziland, Kwazulu-Natal to show general distribution of H. limifolia.
Acknowledgement I have seldom made all the acknowledgements in my writing that I might have done. This particular trip of mine was made under special circumstances without which I would not even have thought of going.
Firstly my son Warwick, who suggested I should go and who facilitated the trip by offering a vehicle.
Then to:-
Ms. Adri Henn, a business associate of Warwick’s at Natal Drift Nursery, Barberton. Her offer of accommodation was a vital factor in deciding to go.
Mr. Gordon Morrison and my school contempories who agreed to a 50-year reunion at Ixopo. A most moving occasion.
Ms. Michele Pfaff of the Gauteng Conservation Authority for contacts and discussion on conservation issues.
Mr. & Mrs. L.Alleman of Link Hills, Kloof for hospitality and encouragement.
Mr. Charles Craib for important information and assistance
Mr. Geoff Nichols, a conservation consultant at Durban. Also a naturalist and plant enthusiast par excellence. Also to his acquaintance at Bridgewater Propagation Nursery of the Durban Municipality, Mr. Mark Gillmer.
Ezemvelo KZN Wildlife for the necessary permit and accreditation of my effort, and to Dr. Pete Goodman and Rob Scott-Shaw for valuable co-operation and information. Mr. M.I. Gumbe, technical assistant at Mkhuze Game reserve. Mr. Rob Blok, Conservation Manager, Ithala game reserve. Mr. Alex Wood, Regional manager, Vryheid. Ms. Bridget Church, Horticulturist, Queen Elizabeth Park.
Mr. Errol Harrison of Mtubatuba.
Mr & Mrs. Mannie van Rooyen of Pumalanga Game Farm.
Mr. Kevin Pretorius and Mr. Matheus Wessels of Phinda Resource Reserve.
Ms. Kathy Cameron of Casamia, Pongola.
Mr. & Mrs. Rynold Steenkamp of Magud.
Mr. &Mrs. Jonathan Pons, Mabuda farm, Isiteki.
Mr. Mbabane Matsenjwa, Mabuda farm.
Mpumalanga Conservation Authority for permit and accreditation, and particularly to Mr. Mervyn Lotter and Mr. Gerhard Strydom.
Mr. Johan Hurter of the Lowveld Botanic Garden.
Ms. Rhona Milstein of Barberton.
Mr. Nico Oosthuizen of Mountainlands Game Reserve, Barberton.
Mr. Luke von Johnston of Boondoks Farm, Kaapmuiden.
Mr. & Mrs. John Roux and their son Etienne of Three Sisters, Kaapmuiden.
Mr. & Mrs. Hugh Foster of Rosthwaite farm, Vryheid.
Mr. Barry Killian of Bivane Dam, Vryheid.
Mr. & Mrs. Graham Root of Ntibane Bushveld Hideaway, and herbalist Erica.
Mr. Bob Kent, and Mr. Steven Hammer who were kind enough to comment.
Dr. Paul Vorster was also so good as to offer me his opinions that are not reflected in the article.
Lastly to my wife Daphne whose unfailing trust and friendship defy any evaluation.
To all these and a few whose names I did not record, I must record my appreciation for the courtesy and kindness in listening to my story and offering me so much assistance and advice. There are also many people peripheral to this special journey who played their part and who I omit mentioning. Had I gained no new knowledge or insight into Haworthialimifolia, at least I gained the association of many people whom I would like to call my friends.
Addendum Having written so much about Haworthia in the process of learning about them and knowing them better, I find it incomprehensible that other writers seem to ignore completely what I have had to say. While this may appear to be a very arrogant attitude, it simply mirrors what I consider “science” to be; primarily a review of what is known or thought to be known, and then a statement of intent, action and conclusion. At present there exists a primary field of formal science which resides in students and graduates from schools of learning (professionals), and then the field of informal science where formally unqualified persons (amateurs) try to forward their perceptions and observations. My frustration is that the hearts and boundaries of these two fields in respect of Haworthia have been and now are, so confounded, that “knowledge” and “understanding” are wholly concealed. My frustration is expressed in the following statement sent to the Editor of Alsterworthia…”The actual position is that classification and the naming of plants is not an activity calling for any special skills and it does not class as ‘science’. It can be for science. Brummit, in Taxon, has written that the binomial nomenclatural system does not truly work. If this is true, it means that for all these years taxonomic botanists have been slavishly applying a system which is flawed. Their failure to have properly applied their minds to the task has led to incongruities which are not that difficult to detect. Where leaders in the field have had their doubts and indecisions, nomenclatural niceties have become more important than the goal. Biology being the inexact science that it is has meant that totally amateur classifications have come to be lauded, praised and accepted when they have no intrinsic intellectual credibility at all. Anyone has free entry to botanical classification whether or not they can even sensibly discuss the issue of the definition of the word ‘species’ which is the basic unit of biological classification.
It is in this respect that I quote Lyall Watson, from his book, Dark Nature, where he writes about the similarity science now has to religion, and it is worth recording that the Dalai Lama has said the same thing. In Watson’s opinion scientists are largely defending a position in which they have a vested interest and they say things they wish to hear and what they expect others to want to hear too. Science according to Watson is thus in the same trap as religion … ‘an establishment intent more on protecting its own interests than in looking at the world with curiosity and honesty’.
This is the thrust of many things I have written and my objection to statements made by leading botanists in respect of classification of Haworthia. Offence will be given if I point to these individuals and their statements, but if ‘curiosity and honesty’ are employed, they can easily be found. One manuscript of mine in respect of just one of the papers which I would use to exemplify my point, reads … “it would be more honest just to acknowledge the impracticality of unispecific genera, than parade a host of fallacious arguments to obscure them. The other even more unfortunate aspect is that all this fault‑ridden argument passes over the heads of the unsuspecting community that is interested in the plants and wants to use the names? If this is what professionals do, what can be expected from amateurs? The foundation is secured for the most uninformed and ignorant to parade as taxonomists, and they do. All that actually happens is that credibility becomes a very rare commodity in plant classification and nomenclature.’ Many of us generate opinions and comment which actually have roots in ignorance.”
I therefore wish to encapsulate my observations in this addendum which addresses the above issues in respect of one object viz. Haworthialimifolia.
To do so I have first to recapitulate slightly the issue of definition of the word species. I will not repeat again anything I have written in this regard nor discuss again the paper of Brummit and its implications again. I merely say that I regard the Latin binomial as a reference to a system of living organisms as a group or groups of organisms which are genetically and morphologically continuous in space and time. The binomial itself is derived from a sample and this is formally attached to a single specimen lodged in a recognised herbarium.
What then is the species Haworthialimifolia? We have to go first to the sample and the description. The sample seems to have been a single specimen (perhaps more, the description states… “the plants”) received by R. Marloth in the year 1910 from Medley-Wood, Director of the Botanic Gardens at Durban. He is stated to have obtained (note: not “collected” as Breuer, 2000, interposes) them “originally from the country west of Delagoa Bay”. The description is extremely brief and it is accompanied by an illustration of a single leaf (fig.1). It can be said of the description regarding the flower that it is so vague that one could not even recognise to which of the three major groups in Haworthia to which it belongs. Here there is already room for confusion in the failure of various writers, either amateur or professional, to recognise what these are. This is especially so when there is a distinct probability that the evolutionary roots of those three flower types could be polyphyletic. This means in essence that classification by vegetative or any other “structure” may not necessarily indicate the same grouping.
It is quite clear that Marloth was right in his assumption that he had to deal with a species “quite different from those of any other species” and “sufficient to constitute a new section”. But he was not quite right in respect of the reason or ultimately the rationale. He was right because H. limifolia is a species system in the northeast of Southern Africa (with H. koelmanniorum) and thus widely separated geographically from any of the other species. It is not because the leaves have a specific surface character or marking and this is a point I will have to labour on later. Apart from some similarity to the geographically contiguous H. koelmanniorum, there is H. nigra in the central Karoo which bears some similarity in respect of leaf “markings”. I put “markings” in inverted commas because it is not correct. The “markings” in H. limifolia are specifically transverse undulations on the leaf surfaces giving it the file-like appearance from which the Latin epithet is derived. Marloth stated quite explicitly that the “markings” were not tubercles or tuberculate.
It is really curious that G.G. Smith (1950) – and he was amateur – had no qualms whatsoever in using the word “tubercles” for the leaf surface character. This was when he described a new variety, keithii and also reduced H. ubomboensis Verdoorn to varietal rank in H. limifolia. Var. ubomboensis with a few longitudinally arranged tubercles on the upper and lower leaf surface was in Smith’s opinion, together with the similar longitudinally tubercled var. keithii, recognised to be part of the system “west of Delagoa Bay”, namely H. limifolia. Smith introduces some new perceptions. He describes the var. keithii as lighter in colour, with less rigid and narrower leaves and less prominent tubercles, but similar in its stoloniferous habit, arrangement of back and face tubercles and the longitudinal lines on the face. Referring back to the original description, the only character here which Smith could have correctly taken (he did get the leaf surface wrong) was perhaps in respect of the width of the leaf.
After Smith there was a bizarre contribution by Flavio Resende (1943) in which he described essentially four varieties of H. limifolia, if it is at all necessary (and possible) to try and follow his logic. None of these were of known origin and my assessment of Resende’s total contribution to Haworthia, and specifically to H. limifolia, leads me to counsel complete disregard thereof. Nothing is added to knowledge of the system that can in anyway be currently applicable and useful. All that it does is further confound and confuse the issue in an environment that I am trying to show is already very difficult to function in. It could for example be argued that a variety with the proposed new name viz. gideonii could in fact be Resende’s f. diploidea (= var. marlothiana Res.). The next author to venture information by way of a new epithet was myself in describing the var. gigantea. This was based on a single specimen that for reasons cited in the preceding section, can be reasonably considered to have its origin in the Pongola valley. The last significant contribution was by Pilbeam, possibly as a consequence of my writing, tied to the collection by the very distinguished collector/explorer J. Lavranos, of the var. striata.
Breuer (2000) has subsequently made an excellent compilation of descriptions and illustrations. Other than that it is an excellent compilation which brings all the literature together, it adds neither further information nor new insight. In respect of H. limifolia, it does curiously add a Berlin herbarium photograph to illustrate the “typical” variety. This photograph could just as readily been used to illustrate the var. schuldtiana or one of the other varieties such as even gigantea. There is nothing to suggest that Breuer could have and did consider all the above, nor the three papers which I deal with below. Breuer (2002) has proposed his system for Haworthia in which there is a new varietal structure for H. limifolia. However, I am familiar with his other outstanding compilations and the many less rational imputations and statements which he attaches to names and his perceptions of what those names are as “species”. These often call for the kind of response which Smith and Hilton-Taylor (1996) made to Breuer’s “contribution” to the phytogeography of the genus Haworthia (1996). Breuer clearly cannot know more about H. limifolia than either Smith et al. (1997), nor Crouch et al. (1999). I feel constrained to say that Breuer is not competent to even attempt such taxonomic rearrangements. The title of his 2002 paper includes the words “species concept update”. It is in fact nothing of the kind. Breuer (2002) does not produce any evidence of what his “species” concept is and his list is only his concept of what species there are in Haworthia. Having produced a revision, I am fully aware of many of its limitations and what is needed to verify it and resolve outstanding problems. Among the problems is the major one of the species concept which has to be resolved at polymath level as it is in the realm underlying the whole of taxonomic botany. Breuer has given me no indication of his capacity and willingness to undertake the intellectual activity this impels, and has dismissed most of my attempts as “polemically things” (private communication). He is also not positioned to undertake the arduous and extensive field-work that is required to even verify my revision, let alone add to it or make changes.
The next attempt of note to explain H. limifolia is that by G.F. Smith et al. (1997). Smith et al. paper purports to illustrate and describe the typical variety limifolia on the basis of two specimens both of uncertain geographic origin. The significant thing is that the paper ignores Marloth’s specific statement that the species was not tuberculate. They repeat the curious aberration that G.G. Smith introduced. Despite the key the paper provides, the var. gigantea is misidentified as var. limifolia as can be gleaned from the illustration. I do not doubt that to a degree I am also culpable here because I know a draft of the paper was sent to me for comment. Unfortunately I do not have any recollection of what my comment was and any contribution I may have made is not acknowledged. Certainly it has not corrected this error. I would have thought the authors would have come to realise their error when they became involved in the next paper.
This is the contribution by Crouch and Smith (1999). It must be noted that the title refers to the typical var. limifolia. This paper does acknowledge a contribution from me, but I must say I think this is misplaced and could refer to the earlier paper by Smith (1997). I cannot imagine that this second paper, and certainly not the final draft, by Crouch and Smith would have passed my critical eye. The paper essentially deals with plants either obtained from the Durban herbal market, or from plants shown to the authors by Dr. Peter Goodman, then resident botanist at the Mkhuze Game Reserve. It also contains some quite specious comments about classification. Clearly no effort at all went in to considering and exploring known records or to generating new ones. From what I have discussed in the earlier part of this work, the variety the paper deals with is either gigantea or striata and definitely not the var. limifolia. I have suggested that these two former names are probably synonymous, and I have also suggested that the recognition of a typical variety is somewhat of a problem in botany. The species as a system is typified by ALL its variants. I take Crouch et al. for being remiss in emphasising, as they do, the importance of understanding taxonomic identity and then making such a weak effort at doing this. Here they are dealing with a variable element distributed over an extremely wide area. They have information that the area still extends considerably southward of the known records which they have in addition not properly registered (I will explain this later in this note). No effort was thus made to question distribution. One can hardly take Breuer remiss for thinking he could revise the genus from Nederzier, when local professional approach the “importance of taxonomic identity” in this way.
Now we come to Smith and Crouch (2001). The paper opens with the words “After consideration of available evidence…”. In my opinion very little of the evidence that I have presented here, of that which was then available, has actually been considered. Among my criticisms of the paper is that of distribution. If Smith and Crouch had properly considered this, they would have been struck with the fact that H. limifolia has been reported near Komatipoort as “SildoSuavo to Moamba”, and that the one small locality known for arcana lies between this record and records south-west of Malelane and at Barberton itself. They should also have considered Venter’s collection at Three Sisters which is only about 30km distant. It is beginning to appear to me that as my plants collected from these sites grow in cultivation that these will also class as var. arcana. The area is between Delagoa Bay and the Reef and it is thus highly probable that the var. arcana is a superfluous name for limifolia. Despite their extensive discussion of classification and population variability, the authors simply accept plants of unknown origin to represent Marloth’s species. These plants are the var. gigantea and NOT var. limifolia.
There is very little in Marloth’s description than can be put to good account. The colour is given as “obscura-viridia” which can be taken for dark-green and otherwise it is the transverse non-tuberculate undulate raised lines on the upper leaf surfaces which are the major point of the description. The detail of the flower is quite worthless and it is not possible even to determine the subgenus. Despite this paucity of information, there is little difficulty in saying that all the plants included in the preceding discussion are H. limifolia. What is it then that makes this species recognisable among this welter of words and the confusion they contain?
In their diagnosis for the var. arcana, Smith and Crouch make the following points:- it is “characterised by a combination of vegetative morphological features. It has a distinctive dark-brownish green colour and the leaf-shape is somewhat more reminiscent of H. limifolia var. ubomboensis than of typical H. limifolia. The leaf surfaces are prominently undulate, but generally lack the distinct tubercles characteristic of the typical variety. Furthermore, it differs from the other varieties in the flowering time from the middle to the end of winter.”
Colour I would agree may be distinctive as this is what struck me when I first saw plants at Pretoria. However, colour is a fractious character and largely influenced by growing medium and exposure. My three collections from the close area where arcana occurs (i.e. Boondoks, Manders Dam and Three Sisters), show quite marked differences in colour with the Boondoks plants at present very dark green and Manders Dam plants simply dark green. The glabrous Three Sisters plants are a glaucous bluish-green. At present I have clones of var. arcana which are olive-brown (fig.13 ex PRE), purplish-brown (fig.14 ex J.Hurter) and dark green (fig.15 ex Bridgevale). Smith and Crouch’s remark concerning leaf-shape is misleading as comparison with the Marloth illustration will show. In any case the var. ubomboensis is based on a single clone in which the leaves are quite long and slender. What the Marloth illustration shows is a short-leaf with a reflexed end-area. This is not apparent in all my plants of var. arcana, but there are clones which exhibit this markedly and others which do not. Similarly in plants of var. gigantea from Bridgewater, I have one specific clone in which the leaves are all very reflexed. When it comes to the question of surface character which is the most significant part of the Marloth description outside of origin, then the whole argument falls apart. Marloth’s diagnosis makes the point that the leaf surfaces are prominently undulate whereas Smith et al. say of arcana …”but generally lack the distinct tubercles characteristic of the typical variety”. What Smith et al. seem to have done is to consider that the typical variety limifolia is as they illustrated and described in Bothalia. This is where they have taken the var. gigantea to be the typical, and thus their detailed description does not bear them out as it is incorrect. I have already commented on flowering time.
The conclusion I must reach is that if the var. arcana is properly researched it will be found to contain all the ingredients of the typical variety and prove to be a superfluous name.
I wrote (1962) that “A great deal more information on the distribution and variability of H. limifolia is required before the true relationship (i.e. a good classification) of the various forms can be determined”. Despite this warning and a good many more since, classification is still used as an easy vehicle for publication. I think we have here a series of three publications which are based on a minimum of field-work and knowledge of the subject. It all suggests that my contentions expressed in my letter to the Editor of Alsterworthia, quoted above, are unfortunately true. A new name may be useful as a language tool for collectors and growers even it is quite misleading. It may also be just a source of revenue to traders. On the other hand it maybe worthless as a tool for the meaningful organisation of information and knowledge while it introduces further complexity and confusion into classification. If a name is not rooted in sound reason and science, it can only be such a source of confusion and misunderstanding. In Haworthia there is a constant inflow of poorly researched dubious comment about classification which provokes negation or negative response. The end result is that the literature becomes simply contentious and there is nothing there for the information of the general reader. This is no doubt why nomenclature and name changes are so vilified by the plant growing community. We need a new ethic in classification.
As an end note I must explain the probable source of the statement by Crouch et al. that the distribution of limifolia extends much further south than suggested in any previous reference. While preparing for my exploratory trip to search for the species, I wrote a letter with illustrations for publication in Veld and Flora (The journal of the Botanical Society of South Africa) asking if any readers could help with information. I received seven responses of which only one properly identified the species and this was a photograph of a potted specimen as used in an advertisement by a clothing store. Three re asking if any readers could help with information. I received seven responses of which only one properly identified the species and this was a photograph of a potted specimen as used in an advertisement by a clothing store. Three responses were in respect of Aloe aristata and the others were for H. attenuata. Extraordinarily two of the latter respondents sent me plants or photographs of plants received by them from people living in the Isipofu (Espofu of Crouch et al.) area of Kwazulu-Natal which is in the Sottburgh district south of Durban. The one respondent informed me that his plant had been identified by a botanist as “H. limifolia”. This is clearly the source of the Crouch et al. statement and should be considered in the light of my experiences and encounters with H. attenuata further north related in the main text. ♦
Literature cited
Bayer, M.B. 1962. A new variety of Haworthia limifolia from Natal. Journal of South African Botany 28:215.
Bayer, M.B. 1970. Haworthia as a problem genus. Cactus and Succulent Journal (U.S.) 42:251.
Bayer, M.B. 2000. Thoughts on Haworthia. Spiderwalk Services, Durbanville.
Breuer, I. 1996. A contribution to the phytogeography of the genus Haworthia Duv. Haworthiad 10:35.
Breuer, I. 1998. The World of Haworthia. Vol.1. I. Breuer, Nederzier.
Breuer, I. 2000. The World of Haworthia. Vol.2. I. Breuer, Nederzier.
Breuer, I. 2002. An Haworthia species concept update. Alsterworthia International – Special Issue No.1.
Brummit, R.K. 2002. How to chop up a tree. Taxon 51:41.
Craib, C. 1999. The ecology and status of H. limifolia east of the Masongololo mountain in South Africa’s Kwazulu Natal. Haworthiad 13:49.
Crouch, N.R., Smith, G.F., Nichols, G., Burden, J.A. & Gillmer, M. 1999. A species recovery contribution for H. limifolia var. limifolia, the umathithibala of the Zulu. Aloe 36:8.
Marloth, H.W.R. 1908-10. Some new South African Succulents. Part.2. Liliaceae. Transactions of the Royal Society of South Africa 1:403.
Pilbeam, J.W. 1983. Haworthia and Astroloba, a Collectors Guide, London.
Resende, F. 1943. Succulentas Africanas 3. Memorias de la Sociedade Broteriana 2:94.
Smith, G.F., Crouch, N.R. & Condy, G. 1997. Haworthia limifolia var. limifolia (Asphodelaceae: Alooideae). Flowering Plants of Africa 55:24.
Smith, G.F. & Crouch, N.R. 2001. Haworthia limifolia var. arcana (Asphodelaceae: Alooideae): a new variety from eastern South Africa. Bradleya 19:117.
Smith, G.F. & Hilton-Taylor, C. 1996 A contribution to the phytogeography of the genus Haworthia Duv. – response to Breuer. Haworthiad 10.
Smith, G.G. 1948. Views on the naming of Haworthias. The Journal of South African 14:55.
Smith, G.G. 1950. Some new species and varieties in the genus Haworthia: VII. The Journal of South African Botany. 16:3.
Stevens, P.F. 2002. Why do we name organisms? Some reminders from the past. Taxon 51:11.
Haworthialimifolia is something of an enigma in that both it and Haworthiakoelmanniorum are geographical rather isolated from the rest of the genus. The latter is confined to a small area around Loskop Dam and Groblersdal in what was the old Transvaal province. H. limifolia is much more widespread and occurs from the southern Kruger National Park southwards through Swaziland in to Northern Kwazulu Natal. It was described by Rudolph Marloth from a specimen very loosely said to have come from “west of Delagoa Bay”. No one has ever managed to tie the species down in terms of geographic origin, and the Flowering Plants of Africa (55:24-29, 1997) description of the typical var. limifolia, depicts it as the var. gigantea. My own fieldwork and observations of material of known field provenance show that this var. gigantea and the var. striata are surely synonymous. Neither of them could truthfully said to be from “west of Delagoa Bay”.
The var. arcana is another matter and a very curious one too. It does come from a place very close to the old wagon routes serving the goldfields of the Transvaal viz. near Hectorspruit. However, plants from there do not seem to match Marloth’s original description, and there is some confusion about the question of the “tuberculate” versus the “non-tuberculate” of that original description. Having seen plants which I think are truly non-tuberculate and yet with the file-like ridges across the leaves, I do not think var. arcana fits the bill for the typical variety. When I first saw a specimen of this variety in the Pretoria Botanical Garden while David Hardy was still there, I would also have plumped for a new variety based on its odd colouration. However, that specimen I saw was very olivaceous and I would have named the variety accordingly. The plants which Crouch and Smith collected of var. arcana are more brown coloured.
I was fortunate enough to be able to spend some time in Swaziland and Mpumalanga (Transvaal Lowveld) in 2002 looking for H. limifolia. One of my aims was to see the var. arcana in habitat, and the other to re-locate F. Venter’s collection of a glabrous specimen from very near the same place. The material of Venter’s was illustrated in Haworthia Revisited as var. ubomboensis.
Fig.1 is P.J. Hurter’s 105 of var. arcana from the type locality and Fig.2 is a specimen of Neil Crouch’s ex Bridgewater Nursery, Durban, also from type locality (approximately Hectorspruit – the locality has been variously given as Komatipoort and Malelane. It should also be noted that there are rumours of the occurrence of the species northwards within the Kruger Park, and there is a firm record for Suado Suavo across the border in Mocambique). I photographed the latter Crouch specimen because it is a less tubercled glabrous plant among a few others. It is quite evident from these two pictures that the prominences on the leaves would qualify as tubercles with a little evidence to suggest that they could become concolorous with the rest of the leaf surface and also merge to form the ridges described by Marloth.
Fig. 1. P.J.Hurter102 H. limifolia var arcana. HectorspruitFig. 2. N.Crouch H. limifolia var arcana. Ex Hectorspruit, at Bridgevale Nursery, Durban.
Fig.3 is my MBB7144 from south of Malelane and about 30km west of the Hectorspruit population. Although there were about 100 rosettes they were in a very restricted area and I suspect that they were all of one stoloniferous clone. The three samples I collected are identical. They are the same size as the arcana specimens but the colour is darker and the brownish tint is absent. Fig.4 is my MBB7144 from about 2km to the south-west and again there were only about 15 rosettes in a situation which suggested plainly they were clonal. The plants are larger, the tubercles concolorous and there is a stronger tendency to banding. My MBB7145 (fig.5) is of Fanie Venter’s glabrous element (I have named it elsewhere as var. glaucophylla) from the top of the Three Sister’ mountain. This is about 1,5km from MBB7145. My collection seems to not be from the same precise locality as that of Venter’s and I do suspect, although the plants were plentiful and scattered over a small area in very rough terrain, that the population is derived from vegetative proliferation. My attempts at pollination have so far not been successful and I need to try and pollinate the Venter plants with my own.
Fig. 3. MBB7143 H. limifolia var arcana. Boondoks, Barberton.Fig. 4. MBB7144 H. limifolia var arcana. Manders Dam, Barberton.Fig. 5. MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton.
Lastly I have MBB7142 (fig.6) from very close to Barberton itself and MBB7141 (fig.7) even closer. These I refer to the typical variety. It is highly improbable that these are related clones because of the distance and terrain separating the populations. However, the plants are so consistently alike within and between the populations that it is a possibility that cannot be ruled out. In the previous chapter I illustrated a F. Stayner collection from near Barberton which is very similar to these two collections of mine. Stayner gave the locality as “Barberton Airfield”. This, given the nature of the terrain, is highly unlikely and it appears to me that Stayner was in fact referring to an airstrip near Kaapmuiden, and very close to Three Sisters. The leaves of the plants are more barred and file-like, but there is till a strong tendency for the coalesced tubercles on the upper leaf surface to be broken down the mid-line. This could impact on the integrity of the plants in that area being a geographical entity. I have not been able to successfully pollinate between plants within those Barberton collections and have not tried between them. Although I have plants of var. arcana from three sources, I have also not been able to get them to set seed and it is possible that they are all clonal in origin. This may be true for the original locality too, although it is reported that there are many plants there, and the clone I photographed from Crouch’s sample appears to be different from the other specimens I have.
Fig. 6. MBB7142 H. limifolia var limifolia. May’s Mine, Barberton.Fig. 7. MBB7141 H. limifolia var limifolia. Rymer’s Creek, Barberton.
I have no conclusion to draw other than to say that the problem of genera in the Alooideae is a reflection of the problem in the lower ranks. Alternatively it should be stated in the converse. This may help cladists to better appreciate that building a phylogenetic classification from only the top down probably will not satisfy ordinary people who will use the classification. Where the subgenera are unequivocally (notwithstanding anything Dr. Hayashi has to say in this regard) recognisable on the basis of the floral structure, the species are not. Thus even in the subgenus Hexangulares it is difficult to know if all the species satisfy the requirements of definition. For example H. coarctata and H. reinwardtii, or H. attenuata and H. glabrata, may be synonymous. Even H. koelmanniorum and H. limifolia may be so. A species such as Haworthia pungens (fig.8) also creates problems because of the similarity to Astroloba on the basis of the leaf spirals approaching 5-farious, and the smoothness and rigidity of the leaves. It may well be that vegetative is more compelling than floral structure, and this has to be very carefully considered when speculating about true phylogenetic relationships. ♦
Fig. 8. JDV96-100 H. pungens. Braamriver, Joubertina.
I was contemplating the problem I see in the way we approach classification, contending that there is no resolution to the endless addition of new names and arguments about their validity and usefulness unless we reach some agreement on method and purpose. The confusion about names, descriptions and identifications that had arisen in 1947 came about for four reasons. Small samples and inadequate exploration, too many unqualified experts, lack of insight into what species actually may be, poor methodology.
There is no excuse for this situation to continue. Sampling is at a very high level and because the other three elements have not been removed, confusion is as great as before. Many names may have been lost from sight, but there is no lack of new ones to replace them. Experts are a problem and their qualifications even more so. In my own experience I have communicated with many highly qualified botanists who have contributed absolutely nothing to the resolution of problems as basic as typification of names, or definition of the word ‘species’. So what then contributes to qualification? This is surely an unbridgeable problem when amateurs have dabbled so freely in plant classification with apparent success. But their success has come from the failure of qualified botanists to put something more substantial in place than a nomenclatural code for binomials and nothing to indicate what those should mean, or who is competent to generate or change them.
Neither of those problems can be solved by me. My contribution has been to see as much as I can of what others have seen, and then to myself explore in both the arena of the subject matter and the periphery of biology that touches it. What I have done is to try and relate what I have read and been taught to what I have observed and experienced. In this way “species” have become a reality as systems of populations and individuals that are spread in space and change with time. Whether evolution is a fact or not I cannot say, but change certainly is. Classification since Linnaeus, has always been associated with this change but not necessarily by everyone. But in the time of Linnaeus, the sampling level was very low and anything new could be, or was, interpreted as a new species and diagnostics were put in place to aid identification. Hence even the Latin binomial was a diagnostic. As the sampling level has been raised so has the system come under increasing stress and it is a fallacy to think that only Haworthia is problematic. However, this is the genus under discussion here and we have to admit that the diagnostics and the methodology do not work.
H. limifolia can be used as an example to illustrate the situation.
It is to all intents isolated from the rest of the genus. We do have H. koelmanniorum sitting to the far north-west and then H. tessellata further to the south-west. Otherwise the rest of the genus is in the Cape. The issue of subgenus need not be seen as a consideration here even though the differences there are as great as any elsewhere in the family.
H. limifolia has a very wide distribution from Komatipoort in the north, westwards and southwards to Vryheid, eastwards and southwards to Gollel and then Stanger in KwazuluNatal. The whole of Swaziland and a large part of northern KwazuluNatal may harbor this species. We need to even ask … “Is it a species?”
The fact is that there are very different elements involved in respect of individual appearance and virtually all aspects of that individuality. The possibility is that whole populations may be derived from vegetatively propagating individual clones. Some plants are solitary and propagate with difficulty. Others proliferate as vegetative offsets or by stolons. The leaf shape and surfaces are highly variable. The surfaces may be flat or channeled; absolutely smooth, to having raised wave-like wrinkles, to tuberculate with small individual tubercles, or the tubercles may be fused in groups and ridges. These may be normal leaf-coloured or they may be white. Flowers are extremely variable and so are the seeds and the capsules. Flowering time seems to be opportunistic and in cultivation the plants may flower at any time, usually synchronizing during the summer months.
Do we really have any alternative but to see this as one single very large operating systems, despite all the real problems of isolation by distance and hence the associated problem of gene transfer between individuals and populations? Is it sensible or even practical to insist or suggest that there must be Latin binomials that describe (in the early Linnaean sense) the situation or, more importantly, that there are diagnostic characters by which such single taxa (groups, species, varieties, forms etc) can be identified?
How can the situation we observe in this relatively isolated system and (species?) be extrapolated to the Cape where all three subgenera and the bulk of the species occur?
Recently two authors have proclaimed the value of floral morphology in resolving this ongoing disputation about the species. What their actual view of what species are has not been spelled out but it appears to be a belief that simple structural (morphological) differences are what define them. Behind this must lie the unconscious acceptance of the original definition of species that they were systems of interbreeding or potentially interbreeding organisms. Thus flower structure is assumed to be linked to the breeding system and therefore very important in classification. This view of species still seems to be the power behind classification although it has long been recognized that in plants inter-fertility may extend across generic difference.
In Haworthia the flowers, even across the sub-genera, are fairly similar and especially in regard to size. Within the sub-genera, the similarities between what are considered to be species, is remarkable. There are even exact similarities that can be found in what are truly geographically and vegetative different populations. Flowering time seems to suggest some kind of additional factor but even here it is evident that breeding can occur between populations with flowering times 6 months apart. It has to be considered that flowering time differences like this might even be a characteristic of a species. This has in fact been shown for H. marginata and H. minima that surely can otherwise seen to be different systems.
The pollinating agents are insects and it seems as if flower size may be linked to that fact as it is so consistent across the genera and whatever are thought to be the species. Here I just present a number of pictures of the profiles and faces of flowers from 5 populations H. limifolia of which some are from different clones in those populations. A difficulty in viewing flowers is that they change with age, and place on the peduncle. They only last about three days and it is thus difficult to compare flowers of exactly the same age and condition. Of course these are poor pictures but no essential and detail is lost here by poor focus. But it is quite obvious that there is bigger difference between the two flowers (profiles) of the plants from Ithala than there is between them and other populations. Consider too that it can be very difficult to distinguish any of these flowers from those of other species in the subgenus Hexangulares. This essay is not submitted as conclusive evidence and argument and I will enlarge on this in a following essay. ♦
MBB7145 H. limifolia var glaucophylla. Three Sisters, Barberton.EvJ19330 H. limifolia var gigantea. Zuka, NE Hluhluwe.7137a H. limifolia Bumbeni 0657137a H. limifolia Bumbeni 0667137c H. limifolia Bumbeni 0627137c H. limifolia Bumbeni 0637146a H. limifolia Bivaan Dam 0777146a H. limifolia Bivaan Dam 0787146b H. limifolia Bivaan Dam 0807146b H. limifolia Bivaan Dam 0837147a H. limifola Ithala 0687147a H. limifolia Ithala 0697148.2 H. limifolia Ithala 0737148.2 H. limifolia Ithala 0727142 H. limifolia. Mays Mine 1017142 H. limifola. Mays Mine 0997141 H. limifolia Rymers Creek 059