National Botanic Gardens of South Africa, Karoo Garden, Worcester.
Orppington, Kents, England.
Published in CACTUS & SUCCULENT JOURNAL (U.S.), Vol. X L V I , 1974
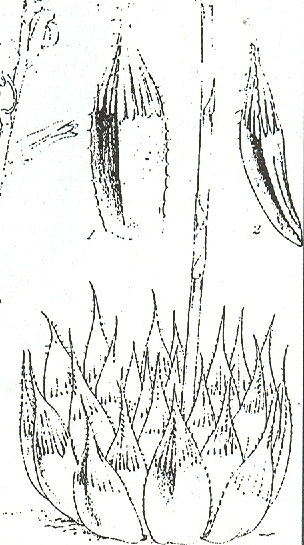
Fig. 1. Water-color by Thomas Duncanson in Kew Library of Haworthia ‘obtusa’ Haw. This is the water-color reproduced by Uitewaal in 1948 in Succulenta on which his name-changes were based.
ABSTRACT: The application of the names H. obtusa Haw., H. pilifera Bale., H. stayneri von Poelln. and H. gordoniana von Poelln. are discussed. It is concluded that Uitewaal (1948) erred in associating H. obtusa and H. pilifera, and that ‘H. obtusa’ is nothing more than a form of H. cymbiformis (Haw.) Duval, perhaps warranting varietal status. The name H. pilifera is thus reinstated for the species centered around the Kingwilliamstown area. Morphological differences are noted in the species H. stayneri and H. gordoniana which suggest, for the present at least, that these names should be retained for species complexes yet to be fully investigated; it may prove that these are merely smaller forms of H. pilifera occurring from SW Grahamstown to Port Elizabeth and on to Humansdorp.
INTRODUCTION: The purpose of this paper is to examine the application of the name H. obtusa Haw. by Uitewaal (1948). Uitewaal bases his argument on a water-color plate in the Kew Library purported to be H. obtusa (fig. 1), but which does not agree with Haworth’s description of the species. He concludes that H. pilifera Bak. must be synonymous and relegates this species to varietal rank under H. obtusa with a new description of the species. Haworth, however, clearly related his species to H. cymbiformis, and in view of problems involved in the application of these names to field populations, it is necessary to question Uitewaal’s conclusions.
INVESTIGATION: Both H. cymbiformis and H. pilifera are widespread and common species in the East Cape. Apart from gross morphological differences, these species differ basically in their habitat preferences, and may be found growing in close association. H. cymbiformis is essentially a riverine species growing on steep rock faces along streams and rivers, and mostly among valley bush vegetation associated with these conditions. It is characteristically a bright green species forming dense mats with compacted roots confined to shallow crevices and cracks of the comparatively soilless habitats. It may become dull green to yellowish-brown or red in its different forms and under different growing conditions. H. pilifera on the other hand, while also found along streams and rivers, occurs in grassland vegetation in sparsely covered rocky situations and usually shallow soils. It is characteristically gray-green in color with pink to purplish hues – never acquiring the brilliant green of H. cymbiformis or becoming dirty brown as Haworth’s description of H. obtusa. It may also form dense mats but with thick fleshy and spreading roots, while the leaf surfaces are level with the soil surface in exposed situations. There should, therefore, be no question of confusing these two species. A form of H. cymbiformis which agrees with the Kew illustration is recorded by G.G. Smith (fig. 2), and this was found growing in a typical population of the species. Other forms have frequently been seen by the writers in cultivation which, by reason of their similarity of form, coloring, habit and floral characters, suggest a similar origin.
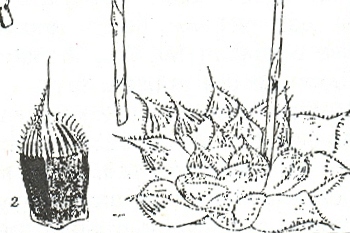
Fig. 2. Haworthia cymbiformis Haw. G. G. Smith’s no. 3389a – a form from Ft. Jackson, Nahoon River
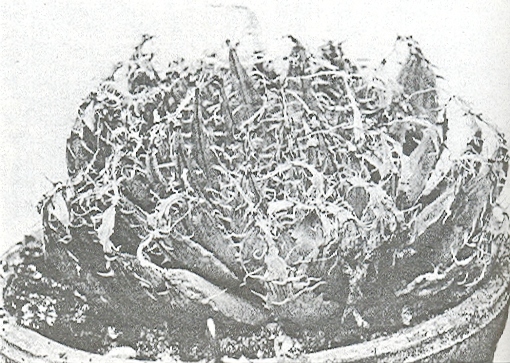
One such plant has been cultivated at the Karoo Garden in shady conditions and then transferred to brighter light for five months (fig. 3) to see how it would compare with the various forms of H. pilifera under similar conditions, and with the Kew illustration. The result confirms that a plant occurs within the writers’ concept of the H. cymbiformis complex which agrees with the Kew illustration but which cannot be associated with H. pilifera. The test plant became brilliantly green under the brighter light conditions, the leaves were shorter but retaining their turgid spreading attitude, neither truncating nor developing necrotic tips. The plant remained above the soil surface and did not recede as H. pilifera does under exposed conditions. Furthermore plants grown in association in cultivation for several years of a plant similar to figure 3 and varieties of H. cymbiformis and H. pilifera, clearly exhibit that the former is in character allied to H. cymbiformis and not H. pilifera.
Fig. 3. Haworthia ‘obtusa. The test plant transferred from shade to sunlight. Origin unfortunately uncertain but alike in every respect.
At the time Uitewaal submerged H. pilifera under H. obtusa, G.G. Smith wrote to J.T. Bates in England that “Mr. Uitewaal and one or two others have recently sunk several of the old well-known names, but I have not accepted all the changes … The H. obtusa, taking the place of H. pilifera, is H. cymbiformis var. obtusa … H. pilifera has never been found on a krantz (rock-face), whereas H. cymbiformis and its varieties are only found on krantzes, and I have plants from a couple of krantzes which are varieties of H. cymbiformis and in a photograph look more like H. pilifera than H. obtusa does … what is more he (Uitewaal) makes up a description from the painting!”.
J.R. Brown in his ‘Notes on Haworthias’ in the American Society’s journal wrote of H. obtusa: “This Haworthia has long been given the name of Haworthia cymbiformis var. obtusa (Haw.) Baker, but the numerous plants under this name which I have seen have only been cultural variations of Haw. cymbiformis Haw.”
The most extraordinary part of Uitewaal’s proposal is that he regards the original description of H. obtusa by Ha worth as “defective and misleading” and, as Smith points out, then proceeds to describe the plant from the painting. The painting itself is by an artist, Thomas Duncanson employed at Kew contemporary with Haworth, and it is more probable that this diverges from the plant it is intended to portray than Haworth’s written description. Anyone who knows Haworthia well (despite Uitewaal using this same claim to support his own contention) must in looking at the Kew watercolor bear Haworth’s description in mind and relate H. obtusa to H. cymbiformis.
Haworth’s description reads as follows: — obtusa H. (small blunt hollow-leaved) foliis ovatis concavis luridis obtusis cum mucronulo, apicam versus pellucentibus, lineis saturatioribus. Habitat C.B.S. G.H. 4 Obs. H. concava Nob.* in Revis.Succ. proxima, at subduplo minor, colore adhuc lurida, et minus virente glaucove quam in ilia, foliis apice crassioribus obtusioribus magis pellucentibusnmagisque striatis. Flores ut in affinibus. (Haworth-Phil. Mag. 1825)
(Translated: — obtusa H . (small blunt hollow-leaved) leaves ovate and concave dingy yellowish-brown obtuse with sharp point, pellucid towards the apex, filled with lines. Habitat C.B.S. G.H.4 Obs. near to H. concava Nob.* in Revis.Succ. but nearly half as small, color until now dingy yellowish-brown, and less glaucous green, leaves thicker and more obtuse, more pellucid and lined. Flowers the same).
* FOOTNOTE: H. concava was subsequently declared synonymous with H. cymbiformis.
Fig. 4. Haworthia vittata Bak. The illustration accompanying Baker’s original description dated March 1871.Fig. 5. Haworthia cooperi Bak. The illustration accompanying Baker’s original description dated September 1870.
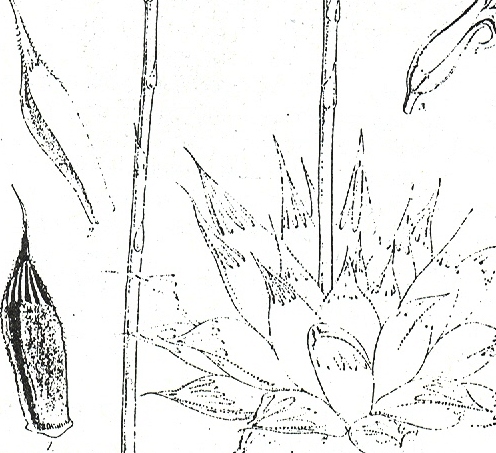
Of particular note are Haworth’s comments on the color, and on the flowers. The flower of H. cymbiformis is a clear white and generally borne on a short, slender peduncle, and Haworth does state that the flower of H. obtusa is the same. In H. pilifera the flower is off-white and borne on a generally stout peduncle. What is difficult to concede, is that Haworth never received specimens of H. pilifera and that it was left to Baker to first describe the species as well as the near relatives H. vittata (fig. 4 ) , H. cooperi (fig. 5) and H. columnaris. These latter species must almost certainly be included in the synonymy of H. pilifera, although the two first named of these may perhaps represent introgression of H. pilifera toward H. blackbeardiana von Poelln. Field observation indicates that there is no good evidence to support varieties (in the botanical sense of sub-species), as variation across the distribution range of H. pilifera is both large and evidently continuous. Observations on the many collections of this species yield some interesting observations. Some variants are very bristly with markedly and acutely pyramidal leaf-tips, and others have rounded and minutely mucronate leaf-tips. Of particular note was H. stayneri from type locality. This species remained small but the leaftips truncated with the end-bristle curving in, and together with the veins of the back upper leaf surfaces, dying to form a whitish necrotic network across the end-area. The differences between sun and shade forms of H. stayneri are shown in figure 6. Variations within a collection of this species from type locality invalidate the variety H. stayneri var. salina (syn. H. pilifera var. salina) von Poelln. H. gordoniana grown under exactly the same conditions became deeply submerged in the ground but the leaves remained erect and narrowly pointed (fig. 7), and also did not assume the deeper purplish to gray-green hues of H. pilifera or H. stayneri.
Fig. 6. Haworthia stayneri von Poelln. Sun and shade grown forms.Fig. 7. Left: Haworthia gordoniana von Poelln. Sun grown plant. Right: Haworthia stayneri von Poelln. Shade grown plant.
CONCLUSIONS: No progress will be made towards a stable and rational nomenclature in Haworthia until names are firmly attached to field populations. It is clear from the above observations that Uitewaal was wrong in regarding H. obtusa to be “unmistakably” associated with H. pilifera and also wrong in amending Haworth’s original description. In view of the confusion now in correctly applying Haworth’s name, and assuming that the illustration at Kew is in fact Haworth’s species, it is proposed to recognize the name H. obtusa as applying to the H. cymbiformis complex, reducing it to synonomy with that species. At the same time it is recognized that the state of naming in the H. cymbiformis complex leaves much to be desired, and it is hoped that further investigation will rationalize this species. The name H. pilifera is retained for the complex found in the Kingwilliamstown, Kei Road and Komgha areas – also requiring further investigation in regard to variation and distribution. This species is more widely distributed than has hitherto been indicated, and is found eastward to Idutywa, north-westward to Bedford, Cradock and westward to Grahams town. It is typified by the illustration accompanying Baker’s description (fig. 8) in Saunders Refugium Botanicum. It is fairly certain that the ‘species’ H. vittata Bak. and H. cooperi Bak. found in the Cathcart area, are northern ecotypes of H. pilifera and will eventually be regarded as synonymous of H. pilifera*.
Fig. 8. Haworthia pilifera Bak. The illustration accompanying Baker’s original description dated September 1870.
It appears to be a practical necessity at this stage to recognize H. stayneri and H. gordoniana as discrete species in consideration of their distribution and the morphological differences recorded here. Both species require further investigation — H. stayneri particularly in relation to the distribution of H. pilifera in the Grahamstown, Alicedale and Bedford areas, and H. gordoniana in relation to the vast complex of the Gamtoos river valley, Humansdorp.
The name H. columnaris Bak. is discarded as being probably a synonym of H. pilifera: described by Baker in 1889 the plant was originally received at Kew from an Exeter nursery, and can be regarded as a variant of H. pilifera.
H. dielsiana von Poelln. and forma acuminata von Poelln. are also discarded as local variants of H. pilifera.
H. obtusa var. pilifera forma truncata Jacobsen is discarded as a misidentification for H. umbraticola von Poelln., cf. original descriptions and photographs of both.
In the context of this paper the following taxa are upheld: — H. pilifera Bak. (formerly H. obtusa var. pilifera (Bak.) Uitew.) H. cymbiformis Haw. H. stayneri von Poelln. (formerly H. obtusa var. stayneri (von Poelln.) Uitew.) H. gordoniana von Poelln. (formerly H. obtusa var. gordoniana (von Poelln.) Uitew.)
Rejected are: — H. columnaris Bak. (subsequently H. obtusa var. columnaris (Bak.) Uitew.) H. dielsiana von Poelln. (subsequently H. obtusa var. dielsiana (von Poelln.) Uitew.) H. stayneri var. salina von Poelln. (subsequentlyH. obtusa var. salina (von Poelln.) Uitew.) H. obtusa Haw.
Questionable are: — H. vittata Bak. H. cooperi Bak.
It is a pity that it has taken so long to refute what was plainly a tenuously-based decision, but it is confidently expected that these new proposals will facilitate investigation and communication regarding the plants concerned and their respective origins.
REFERENCES Collection of Haworthia illustrations in the Library of the Royal Botanic Gardens, Kew, England (to which the authors are grateful for permission to reproduce the painting)
Succulenta i9:101 (1937); 30:49 (1948) – Uitewaal
Cactus & Succulent Society of America Journal 10:57, 88 (1938); 36:75 (1964); 37:115 (1965) – J. R. Brown
Saunders Refugium Botanicum 4:t. 233, t. 234 (1870), t. 263 (1871) – Baker
Desert Plant Life 15:124 (1943) – J. R. Brown National Cactus & Succulent Society Journal 10: 81 (1955)-Jacobsen
*FOOTNOTE: It should be particularly noted here that if such a decision is reached, then the name H. cooperi Bak. will have page priority and will need to be adopted for the whole species complex.
M.B. Bayer, with acknowledgment to P.V. Bruyns and J.D. Venter.
It is unfortunate when a situation arises where individuals compete to provide a classification for a group of plants. The normal process for aspiring taxonomists is to determine what needs to be done that is not being done, and which attracts and interests them and so select a group to study. There are probably not many instances where different people have worked comfortably together to explore and resolve the taxonomy of any plant group. The objective of this article is to show how that it should be possible to look rationally and objectively at a problem and produce a solution which can be taken forward in the same way.
I think it is understood in the taxonomic fraternity that one of the main aims of a classification is that it must presume to account for all of the plants in the group in question. Is this physically possible? Nobody in the least familiar with Haworthia has any doubt that the territory to be covered for a study of the genus is unbelievably big. The nature of habitats and size of populations, coupled with intrinsic variability and the sometimes cryptic nature of many of the plants make it not only possible but probable, that exploration of even a relatively small geographical area may miss something important. This complexity in the field is not unique to Haworthia. A classification therefore can never be considered to be complete and is only an hypothesis based on what is known. For that which is at the time unknown, it should be assumptive and predictive. It is further necessary that the classification meets the needs of a general group of people who will use it. The nature of these needs is in a large part already expressed in the preceding attempts to classify the group and in the literature associated therewith. A classification is important in the way it allows of generalisation, extrapolation to hitherto unknown collections and for communication.
The test of the hypothesis contained in the classification is the subsequent successful identification of plants by means of the key provided and also by the incorporation of new data into the structure of the classification. It can also be tested by the ease with which people communicate about the components of the classification, and, of course, whether it comes to be generally accepted at all. The strongest and most practical test is its acceptance by curators of herbaria and the way it is used to store and retrieve data in an herbarium.
In the case of plants popular in horticulture and with the general public (such as orchids and succulents) there is often controversy over their classification. The reason for this controversy is that, because of their popularity with the general public, untrained and non‑professional people are drawn into the process of classification and identification. Their only justification for this is their own enthusiasm and interest in the subject which they feel generates new and previously unrecorded information which they perceive a real need to express. It should be recognised that they do not necessarily make any more or fewer mistakes than professionals working on obscure groups or at levels of classification (or sophistication) beyond the reach and interest of the layman. There are also many cases where so-called ‘amateurs’ have made contributions unsurpassed by professionals. It should also be noted that many classifications by professionals may very seldom come under any kind of practical and proper scrutiny because those plant groups do not attract the attention of anybody else. Errors, inconsistencies and absurdities remain undetected. The professional also goes to the outer limits of his intellect where he/she is just as error‑prone as any other person operating at their extreme. Both classes of enthusiasts ‑‑ the professional and the layman ‑‑ need to draw on some other wisdom to know (probably) what is right and what is wrong.
One of the particular problems faced by both professional botanists and laymen in a popular group of plants, is the profusion of material that comes to be passed around in the horticultural trade without any information on its origin and frequently under the wrong name. This considerably confuses the picture and this confusion is difficult to dispel without reference to populations in the field.
A further problem, again experienced by anyone who does not have extensive experience in the field (and indeed of pattern recognition generally, and not only in living systems), is the quite extraordinary variability of taxa like Haworthia. The degree of variation is not consistent for species or for populations. In extreme cases, where vegetative propagation has occurred, there may indeed be virtually no variation (eg H. reinwardtii); and at the other, hardly two clones in a population are identical. There are no quantum steps where categories like sub-species, varieties etc. have consistent and invariable connotation. The more fundamental and philosophical concept of even the species becomes questionable.
Haworthia has been one case where conflicting views have produced a fair amount of difference of opinion and acrimony among authors, and subsequent confusion in the minds of the audience that need the classification to serve their interest. At present there are two classifications available for Haworthia, the one by C.L. Scott (Scott 1985) and the other by M.B. Bayer (Bayer 1982). Neither has proved unassailable and both have shortcomings. Particular shortcomings of both treatments are that they did not address typification, they did not comprehensively cite herbarium specimens and they did not provide credible identification keys.
In Haworthia the classifications are largely artificial because there are no definite morphological discontinuities between the different species recognised. This is why I have said elsewhere that a truly objective botanical classification would probably reduce the numbers of species to about half of that recognised even by myself. For such an objective classification a key could perhaps be provided. A key was provided in the older Handbooks (e.g. Bayer 1982) but my new classification will not provide a key. The reasons for this are very obvious in my handbooks and in most of my writing on the subject since 1971:-
(1) there are really not enough tangible characters on which to build a key.
(2) where two keys (Scott 1985, Bayer 1982) have been provided, I have no knowledge that anyone has been able to prove their worth or make anything out of them.
Out of about five published reviews of the two accounts by Scott and Bayer, three were quite ambivalent: they did not attempt to test the classifications and did not espouse either. Since a reliable and useful key cannot be constructed, I have concluded that photographs and distribution information are the simplest, most reliable and most direct route to obtaining an identification. The herbarium specimens in the three main South African herbaria follow the revised scheme (Bayer, in ms.) but this could be happenstance rather than cognitive intention.
Part of my strategy in the Handbook (Bayer 1982) was to retain species and varietal names even if the indications were that their status may have been weak. There were two reasons for this. Firstly, classification is also a communication process and I tried to match my classification to what I felt was the mood of the day. Secondly I tried to avoid, where possible, dramatic change which may have had to be reversed, and where there was uncertainty of some kind. Thirdly I retained names where I felt they had value in terms of the information portrayed if not as substantial taxonomic elements. Whether or not I succeeded is beside the point because classification is an ongoing event, based on a sample that is known, on how well it is known and unfortunately on personal perceptions too.
The description of Haworthia reddii (Scott 1994) provides me with an opportunity to evaluate the respective hypotheses of Bayer (1982) and Scott (1985). This account should also be a guide to aspiring taxonomists in the group who may be tempted to start at their own levels of knowledge and competence, rather than properly build on historical fact.
The population upon which H. reddii is based is referred to in the New Haworthia Handbook (Bayer 1982, p.30) under H. batesiana, as follows ‑ “… a collection from Klipplaat northwest of Cathcart is clearly comparable. However, the plants there are too robust to be regarded as H. batesiana and it appears that there is a tendency towards H. cymbiformis“.
Scott did not seem to make the connection between this reference and the plants collected by Dr Reddi and himself at the same place which had in the meantime become better known as Waterdown Dam. This is unfortunate because he does mention both H. batesiana and H. cymbiformis as possible relatives of his new species, and it would have been significant if this was an independent and credible observation.
In 1982, H. batesiana was not well known and there were very few pointers to the reality of its existence. Since then there has been another collection from the Valley of Desolation to confirm its existence there, as well as two collections by P.V. Bruyns from the Kamdeboo Mountains and another from the Tandjiesberg. These are both in the greater Graaff‑Reinet area. From observations on these collections (and several others pertaining to H. archeri), it seems batesiana must be incorporated in H. marumiana as suggested in 1982. Furthermore, the concept of that species needs also to be broadened to include H. archeri and relevant collections (Bayer in ms.).
In Bayer (Haworthiad, 1996) I wrote with reference to H. reddii ‑ “At the time I commented on the Waterdown plants there was some doubt about the existence, whereabouts and whatever of H. batesiana. Since that time there have been any number of collections which fairly conclusively support its inclusion in H. marumiana. There is, so far as I know, still nothing to show that marumiana comes far enough east to support speculation of linkage with cymbiformis via reddii. The area NW of Cathcart to Tarkastad has not been fine‑combed by any collector and it probably would better be regarded as an under‑collected region. Furthermore, the distance from Cathcart to Tarkastad is considerably less than Tarkastad to Beaufort West and Prince Albert (at the western known limits of marumiana). There are plants in the upper Kei collected by Peter Bruyns which may strengthen the view that reddii is associated with cymbiformis. In which case it may be sensible to consider it with the var. lepida. My inclination is to put it with marumiana“.
It is quite obvious from Bayer (1982) that at the time I did not want to commit myself to a decision on the collection from Waterdown Dam. I did not regard it as substantial enough as a single population to justify formal description and was fairly sure that it would fit into either of two already described species. One of these was batesiana, which I suspected would prove to fit into marumiana. The other was cymbiformis. At the time the odds were heavily against the latter because it was not known at all from the Kei River valley, and only slightly better for the former. The nearest populations included the missing H. lepida and a collection of my own from near that site and both of these came from the middle reaches of the Fish River which is rather far to the south. However there were two collections from much further to the east in the Transkei to hint at a more extensive distribution for H. cymbiformis.
The drainage system of the Kei river and its tributaries is a highly dissected landscape and the terrain is rugged and steep. There are many rocky cliff faces which undoubtedly harbour Haworthias. It will be a very difficult task to thoroughly investigate even a small proportion of possible Haworthia sites. (I did at one time point at the possible significance of river drainage systems regarding species, but it is self‑evident that geographical features of any kind will influence distributions and breeding systems.) Nevertheless, some collections have now been made from which a clearer picture begins to emerge.
The first interesting collections relevant to the ‘reddii’ problem were collected by P.V. Bruyns at Inverbolo and Inversomo on the Kei River east of Cathcart. These were of H. cymbiformis and established for the first time the existence of this species on the Kei River. He also collected what purports to be H. marumiana var. marumiana in several places north of Queenstown, near to Sterkstroom (in an area which, like Waterdown Dam, is also drained by an upper tributary of the Black Kei). This indicates that H. marumiana also occurs much further east than previously thought. These collections have rather attenuate, strongly spined leaves and are highly marked with translucence between the dense reticulation.
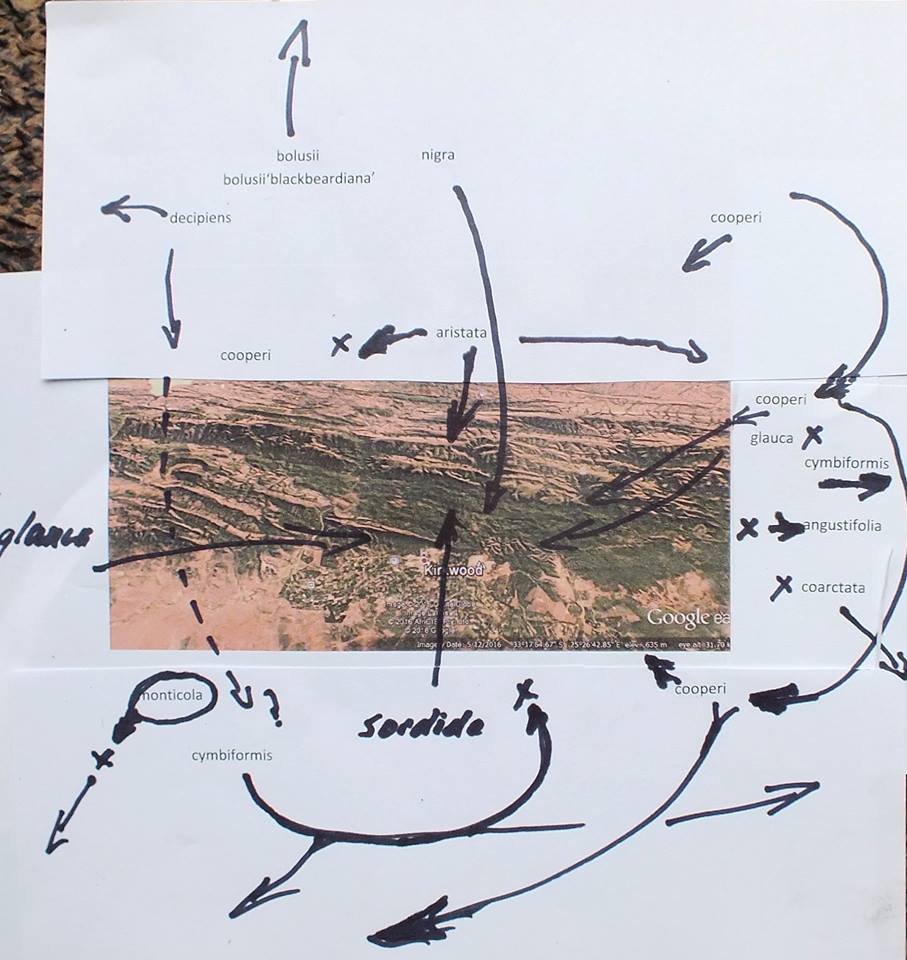
In December 1996, I was fortunate that P.V. Bruyns was able to accompany me on a collecting trip to the Eastern Cape and one of our objectives was the upper Black Kei. It is a tributary of the Black Kei on which the Waterdown Dam was built and the relative location of the populations discussed can be seen on the accompanying map. We were also helped and motivated by a very old specimen in the Pretoria Herbarium collected by Galpin, which I only became aware of earlier in the year and which indicated the occurrence of plants related to H. cymbiformis and ‘reddii’ southeast of Queenstown.
We travelled on a road running northeast from Cathcart in the direction of the Galpin site, but stopped at the bridge over the Black Kei on the farm Turnstream. Peter did the climbing of the huge south‑facing cliff there and came back with several clones of reddii‑like plants. At the same time I found H. bolusii var. blackbeardiana at the eastern base of the same cliff. We then turned back and travelled eastward along the river to the base of a still higher west‑facing cliff on the farm Highclere. Peter again did the very strenuous climbing and again returned with a few clones which he described as difficult to reach on the vertical cliff face.
Peter’s earlier collection along the lower Kei at Inversomo is still further to the south and east. He also collected H. bolusii var. blackbeardiana at this site.
We took the opportunity to revisit the Waterdown Dam on the way home. I was really surprised to find the south‑facing cliff alongside the dam clothed with huge numbers of plants. Although H. marumiana is also a clump‑former, these larger clumps were at lower altitude and much more accessible than H. marumiana usually is. Some of the plants had very distinctive translucent dots and lines while others are unmarked and uniformly opaque with a faint reticulate patterning on the leaves. The floral characters mentioned by Scott are not definitive although the flowers do appear to have strongly coloured veins. We also noted the presence at Waterdown Dam of H. bolusii var. blackbeardiana. The repeated presence of this species may be important in the context of co‑occurrence which forms the basis of my hypothesis relating geographical distribution to the species concept. It is only slightly relevant to this article but it is critical to a broader understanding of the genus[1].
The offsets we collected from Turnstream, Highclere and Waterdown Dam have taken several months under relatively low light to grow out enough to make useful comment. At the moment it is extremely difficult to see any difference between three distinct clones from Turnstream and the collection made on the same trip from Waterdown Dam. I put it like this because the Waterdown plants are quite variable as to the translucent patterning on the leaves. This may be almost absent, or the margins may be translucent, or the face of the leaves may be quite heavily marked with a series of elongated translucent dots or short lines. (It should be noted that in H. cymbiformis as a whole, there is a vast range of translucent patterning, from virtually absent, to only translucent leaf‑margins, to massive reticulate or dotted translucence). The Turnstream collection comprises a very small sample (smaller than I would have liked, and I would have preferred to have seen the plants in situ if I had been fit enough to do so) but the plants are virtually identical in both shape and size to those Waterdown plants which lack the translucent markings. The colour is also the same rather opaque mid‑green. The leaves are sub‑cylindrical, or flatter and slightly recurved with a faint darker reticulation similar to that in H. marumiana var. batesiana, and which is also often evident in H. cymbiformis. The leaf margins in both the Turnstream and Waterdown collections are relatively smooth with evidence of more spination in a few clones of the bigger Waterdown sample. This spination is not comparable with that of the Andriesberg collections.
The Highclere plants looked slightly different at the time of collection. They were bigger, paler in colour and less opaque. The margins were also more heavily spined. I relate these plants to a wider concept of H. cymbiformis var. setulifera V.Poelln. It seems extremely improbable in the context of Haworthia, that these two populations at Turnstream and Highclere could be different species and I cannot harbour any question of this kind.
The Inverbolo and Inversomo plants have been in cultivation for more than eight years and, as they were also grown under brighter light, a straight comparison is perhaps unwise. In comparison with the Turnstream and Waterdown plants they have relatively short obtuse leaves and form tighter smaller rosettes, the coloration is more intense, slightly more glaucous, and the reticulation, opaqueness and/or translucence in either of the two clones (the sample is too small) representing this collection is practically the same. I included this collection among the specimens of H. cymbiformis var. setulifera, which is indicative of the compounding difficulty of making decisions, already difficult, below species level.
These three collections taken together seem to show a definite and tangible connection between the Waterdown Dam plants on the upper reaches of the Black Kei, through the collection at Turnstream and the herbarium collection of Galpin’s, to H. cymbiformis as represented by the Highclere collection and also the Inverbolo and Inversomo collections further to the south.
The connection to H. marumiana is weaker. As one moves northeastwards from Tarkastad, populations of H. marumiana retain their more plentiful and rather slender leaves and do not tend to become more like the Waterdown collections or like H. cymbiformis. The translucence becomes denser and the plants more spinescent. Collections from the western Karoo (Sutherland, Merweville and Carnarvon by Bruyns and Bayer) which enforce the inclusion of H. archeri under marumiana, weaken the argument to include reddii there too. In particular, forms of H. marumiana var. batesiana which do bear resemblance to the Waterdown Dam collections occur only very far to the west around Graaff-Reinet. The clinal trend in this species from Tarkastad northeastward, is thus rather away from a resemblance to the Waterdown Dam plants than towards it. Thus, if one is to seek continuity of variation, the Waterdown Dam populations do not form part of the series exhibited by H. marumiana but fit into the series of variants now known in H. cymbiformis along the upper reaches of the Kei River.
In 1982 I postulated that there were two species viz. H. batesiana and H. cymbiformis, involved in an assessment of the Waterdown Dam collections and was unable to fit it conclusively into either of these. Nevertheless, I was convinced that it could be accommodated here and this was, and has been, the prediction of my classification hypothesis.
More recent collections (mostly by P.V. Bruyns) have filled in much detail in the distributions of both H. cymbiformis and H. marumiana that was, at that time, unknown. These have indicated that H. batesiana and H. archeri can be included in a broader concept of H. marumiana (also as predicted), and they have amplified the known information on H. cymbiformis. This new information shows that, if the species concepts of geographical continuity and co‑occurrence are followed, ‘reddii’ is not a discrete new element standing outside of known and variable species. My prediction that it should not be accommodated in H. batesiana (i.e. H. marumiana) seems to be correct, and our investigations seem to confirm rather that it is an integral part of H. cymbiformis. The important fact then is that recognition as a distinct species is not warranted, and it can adequately be discussed and classified in terms of the structure of the Haworthia Handbooks. The hypothesis has not been disproved and there is a rational basis for development of a still better understanding.
An important implication has been that if reddii and similar individual populations are to be treated as distinct species, then each new discovery of which there could be many, will require a new name and the system will become increasingly disordered and fail. Evidence of exactly this problem is presented by the descriptions of H. batteniae, H. pringlei, H. joeyae, H. venusta and H. mcmurtryi all of which can similarly be accommodated within other variable species. As far as variability is concerned, the plants from Waterdown Dam, and other populations now associated with them, are not exceptional. If we had to continue naming each apparently different element like this, we would end up with a structure that has no coherence, no predictive element and no value in the sense that botanical classification is required to express ‘pattern’ and carry information generally. Such a system may work for the collector in that he may have a name for a particular clone or set of clones, but have no wider or deeper meaning. Many people may be comfortable with and feel justified in using Scott’s treatment. Nevertheless, such an approach simply does not accommodate the incredible variation within the genus.
Because of the conflicting views that seem to be an inevitable part of the process of plant classification, many commentators have said that it is not a science but an art. However, this conflict should not be there. The essence of science is replication i.e. the deduction of conclusions (for example, a classification) from experiments (for example, observations on plants) which should be repeatable. In my work on Haworthia, I have been very conscious of the historic conflict in the genus, the need for credibility, and the responsibility attached to making public statements. Unfortunately, while I may have made mistakes, other authors seem to be less conscientious. Therefore the presentation of differing taxonomic treatments requires the reader to discriminate between them. This demands of the reader that he consider carefully the evidence put forward by authors and then discriminate for himself which author has concluded correctly. Most readers are not prepared to go to this amount of trouble and would rather declare the taxonomy of the group concerned to be ‘controversial’. This is unfair to all authors and also to other readers as it discounts the effort and sacrifice these people put into collecting and communicating information. The opinion that taxonomy is an art with little relevance to the enjoyment of the plants themselves, stems from intellectual laziness and ignorance. It belies the fact that the mere conveyance of a name, which can be forgotten in the very same moment, satisfies some deep psychological need. The audience also has the responsibility to think analytically and critically about what is laid before them. Otherwise they may get a meaningless classification that they have earned, but which is just another yoke around the neck of others who may be striving for the light.
H.cymbiformis var. setulifera. Highclere. NE Cathcart. M.B.Bayer 6873H.cymbiformis var. setulifera. Highclere. NE Cathcart. J.D.Venter 96/70Haworthia marumiana var. batesiana. Tandjiesberg. SE Graaff Reinet.
P.V.Bruyns in J.D.Venter 88/62H.cymbiformis var. reddii. Waterdown Dam. NW Cathcart.
J.D.Venter 96/66H.cymbiformis var. reddii. Turnstream. NE Cathcart. J.D.Venter 96/68/4H.cymbiformis var. reddii. Turnstream. J.D.Venter 96/68/2
(1) My classification hypothesis is built on a definition of species which pre-supposes that they are ‘continuous genetically and morphologically in space and/or time’ (Bayer 1982). Therefore the prediction then is that not only is it probable that H. cymbiformis and H. marumiana will eventually found to be continuous in geographic space, but it is probable that they will also be found to be continuous with H. bolusii. The entire hypothesis should fit within the framework of taxonomic botany, whatever the level of expertise, and satisfy the requirements of scientific discipline. Any two people should come to the same conclusion. If there is conflict there is error.
9. Haworthia cymbiformis (Haw.) Duv., Pl.Succ.Hort.Alenc. :7(1809). Bayer :110(1976). Bayer :35(1982). Scott :91(1985). H. concava Haw. Revis. :58(1821). Aloe cymbiformis Haw., Trans.Linn.Soc. 7:8(1804). Sims, Bot.Mag. 1:21,t.802(1805). Salm-Dyck, Monogr. 11:t.1(1840). Type: Not preserved. Neotype (designated here): icon, t.802, Bot.Mag.. Epitype (ex B&M): Walmer, Port Elizabeth, Smith 2844 (NBG): H. planifolia Haw., Phil.Mag. 44:282(1825). H. cymbiformis var. planifolia (Haw.) Baker JLinn.Soc. 18:209(1880). Aloe planifolia (Haw.) Salm-Dyck, Monogr. 11:t.2(1840). Type: icon. t2, Salm-Dyck, Monogr. 1840: H. cymbiformis var. angustata V.Poelln., Feddes Repert.Spec.Nov. 45:166(1938). Type: Discovery region unknown, T.Foster. Not preserved: H. cymbiformis var. angustata fa subarmata idem. 45:166(1938). Type: Rocklands, Adelaide, W.E.Armstrong in Triebn. 1187. Not preserved. Neotype (designated here): CAPE-3226 (Fort Beaufort): Rocklands, W.E.Armstrong in Smith 2801 (NBG): H. cymbiformis var. compacta Triebn. idem. Type: Cape, west of Peddie, Mrs G. McLaren in Triebn. 1148. Not preserved. Neotype (designated here): CAPE-3327 (Peddie): W Woolridge, Peddie (-AB), Bayer 4648 (NBG). H. planifolia var. exulata V.Poelln., Feddes Repert.Spec.Nov. 43:93(1938). idem. 45:162(1938). Type: Cape, Ubi?, C.H.Woolley in Long 392. Not preserved. Neotype (designated here): Photogr. H.G.Fourcade of Long 392 (NBG): H. planifolia var. planifolia fa agavoides Triebn. et V.Poelln., Feddes Repert.Spec.Nov. 45:162(1938). Type: Cape, Fort Beaufort, W.E.Armstrong in Triebn. 1169. Not preserved. Neotype (designated here): CAPE-3226 (Fort Beaufort): Sulphur Baths (-DC), Bayer 4655 (NBG): et fa alta ibid. Type: Cape, Grahamstown, Mrs Helm in Triebn. 851. Not preserved: et fa olivacea ibid. Type: Cape, Quagga West, Mrs Helm in Triebn. 853. Not preserved: et fa robusta ibid. Type: Cape, Baakens Valley, Mrs I.King 100 in Triebn. 1066. Not preserved. Neotype (designated here): CAPE-3325(Port Elizabeth): Baakens Valley (-DC), Smith 3894 (NBG): et var. incrassata V.Poell. idem. 45:163(1938). Type: Cape, Kowie River, Mrs Archibald 335 in Long 446. Not preserved: et var. sublaevis V.Poelln., Kakteenk. 6:67(1938). idem., Feddes Repert.Spec.Nov. 45:163(1938). Type: Albany district, Mrs Britten in Triebn. 940. Not preserved: et var. longifolia Triebn. et V.Poelln. idem. Type: Cape, Grahamstown, Mrs Helm in Triebn. 864. Not preserved: et var. longifolia fa calochlora ibid. Type: Cape, Port Elizabeth, Mrs Helm in Triebn. 941. Not preserved: H. planifolia var. poellnitziana Res. ibid. 48:133(1940). Type: ex Hort., Hamburg. Not preserved: H. lepida Smith JS.Afr.Bot. 10:21(1944). Type: Cape, Albany district, between Carlisle Bridge and Fort Brown, Smith 5066 (NBG).
cymbiformis: boat-shaped.
Rosette to 130mm φ, partially stemmed, proliferous. Leaves broad ovate to lanceolate, flat to slightly concave, generally <1/3 as thick as wide, usually opaque, green turning yellowish to pink hued on exposure. Inflorescence to 250mm, 10-15 flowers, lax. Flowers white.
1982 – This name has been used interchangeably with H. planifolia for a very widespread and variable species growing in the Eastern Cape. This species has practically the same distribution range as H. cooperi but unlike that species is an opaque, truer green colour. It forms dense mats with a fairly superficial root system and grows on rocky slopes and krantzes along rivers and streams. H. planifolia Haw. was only described in 1825 and was stated to be less proliferous and with distinctly flatter leaves. Baker placed the two species together and a study of Von Poellnitz’s and Smith’s records plainly show that neither could clearly distinguish these two species. Von Poellnitz in fact described H. planifolia var. transiens and H. cymbiformis var. translucens from the same locality, and even stated that he did not know how to clearly separate the two species. Von Poellnitz in Feddes Repert. Spec. Nov. 41:199(1937) records H. cymbiformis from Graaff Reinett (Triebner 861) but in 1938 (ibid. 45:163) describes H. planifolia var. incrassata from the same collection. Berger, in a masterly piece of irrationality, justified separate sections for the two species. Smith transferred H. incurvula V.Poelln. from the section Muticae to the section Obtusatae, on the basis of observed continuity with H. cymbiformis. In Flowering Plants of Southern Africa (pl. 356, 1929) a plant identical to H. incurvula is illustrated under the name H. cymbiformis var. planifolia (Haw.) Baker. It can only be concluded that one name be retained for the entire complex and this is H. cymbiformis.
Uitewaal (1948) put forward the view that H. obtusa was not related to H. cymbiformis at all, but that is was an earlier name for H. pilifera (here a synonym of H. cooperi). He based his observation on a colour plate in the Kew herbarium, and goes as far as to say that Haworth’s original description is faulty. The strongest argument in favour of Uitewaal’s contention is the historical one. None of Haworth’s species can be referred to Baker’s H. cooperi and this is hard to concede knowing how widespread and variable that species is. The name H. obtusa is here considered to be probably synonymous with H. cymbiformis and should probably be rejected as superfluous and as a source of confusion. The distribution range extends from the Bashee River in Transkei, to Prince Alfred’s Pass in the west, and northwards to Fort Beaufort. Variation within individual populations is small but no two populations are quite the same. Thus there is scope for a good many more varieties than previously recognised. However, to stay within manageable limits, only the one really different form viz. fa ramosa with the elongated stem, and varieties not limited to single populations, are upheld. The var. incurvula is apparently restricted to Plutosvale which is a contradiction, but, because Smith discusses continuity with H. cymbiformis (atypical for Smith and not borne out by his field records), because of the possibility of a relationship with H. translucens subsp. tenera, and because of similar forms occurring in the Humansdorp area, it is maintained. The var. transiens is really only more translucent, and at the type locality also larger, than incurvula, otherwise they may have been considered synonymous. The var. umbraticola is a distinctive variety from the Swartwaterpoort west of Alicedale and northeastwards to Fort Beaufort. The leaves are very obtuse and round in cross‑section. The fenestrate blunt tips with shining pellucid areas separated by dark green lines, make it a most attractive variety. H. cymbiformis occurs primarily in the summer rainfall area and is very easy to grow in cultivation. It proliferates to an rapidly and should be exposed to at least some direct sunlight to prevent bloating and excessive softening of the plants. This species is also particularly prone to losing its roots with overwatering so it is also essentially a winter‑growing species which likes at least some resting period during the summer.
1999 – Sims in 1805 commented on the name H. cymbiformis, saying ‘Its name (with too much latitude by the way) is taken from its leaves’, which prompted Haworth to change the name to H. concava. Von Poellnitz repeatedly exclaimed at the poor coloration of these plants in Europe as opposed to the colours which the plants develop under good light, when they are indeed very attractive. The two illustrations in Salm Dyck’s Monograph of Aloe cymbiformis and Aloe planifolia are really very similar in relation to the variation within this one species. One would think thus that a more sensible and conservative approach to species would have been achieved a great deal earlier than it has. It is not obvious to which species H. cymbiformis is most closely allied, but it does seem to be closely associated with H. cooperi. Bayer and Pilbeam may have been in error in their treatment of Uitewaal’s re-appraisal of H. obtusa Haw. as it is not easy to find the obvious field counterpart. The solution suggested below may not be the most appropriate. Col Scott also regarded obtusa as a variety of H. cymbiformis but unfortunately seems to have illustrated the H. cooperi variant. A population on the Kat River near Fort Beaufort sampled by Scott, supports his argument and the decision taken here. The explanation regarding the var. incurvula in the 1982 Handbook is poorly constructed. What was meant, was the recognition of the variety despite its limited distribution, because of the evidence claimed by G.G. Smith, and because of its possible transitional nature towards H. gracilis. H. lepida is regarded as a variant of H. cymbiformis because it could not be re-located despite a detailed description of the one locality where it was recorded. A collection further to the east does not appear to differ dramatically from an already wide range of forms.
Breuer and Metzing nominate a specimen as a neotype when the early illustrations are excellent and are really the basis for the correct and historical application of the name. Haworth cited and accepted the illustrations in Botanical Magazine as well.
a. var. cymbiformis. The typical species is considered to comprise the main body of the species which occurs from Port Elizabeth, eastwards to East London and inland to Adelaide and Committees on the Fish River. Plants in this area tend to have broad, flat smooth leaves without spines.
Distribution: 3226 (Fort Beaufort): Near Alice (-DB), Smith 5635 (NBG); Kat River, W. Alice (-DB), Smith 105 (NBG); 13km S. Fort Beaufort (-DC), Smith 5617, 5617a (NBG); Rocklands (-DC), W.E.Armstrong in Smith 2801 (NBG); Sulphur Baths (-DC), Smith 2795, 3826, 7371 (NBG), Bayer 4655 (NBG); W. Sulphur Baths (-DC), Bayer & Bruyns 6593 (NBG). 3227 (Kingwilliamstown): Debe Nek (-CC), Britten in PRE 39472; Fort Murray Bridge (-CD), Smith 3111, 3317, 3576 (NBG); Bridal Drift (-DC), Smith 2806 (NBG); Umdanzini (-DD), Smith 5336, 5337 (NBG). 3325 (Port Elizabeth): Kranspoort, W. Patterson (-BC), Bayer 4549 (NBG); Slagboom Dam (-BC), Branch 37 (NBG); Below old Fort (-DC), Smith 5040 (NBG); Walmer (-DC), Smith 2790, 2844 (NBG); Baakens Valley (-DC), Smith 3894 (NBG), Paterson 155 (BOL). 3326 (Grahamstown): Fish River valley (-AA), Dyer 4549 (PRE); Carlisle Bridge (-AA), Smith 5567, 5597 (NBG); Cloudlands (-AB), Britten in BOL71308; Howiesonspoort (-AD), Smith 105a, 439, 909, 2843b, 5302, 5305 (NBG); The Fort (-BA), Courtenay-Latimer in Smith 5066 (BOL, PRE), Smith 5066 (NBG, PRE); Between Carlisle Bridge and Fort Brown (-BA), Smith 5066 (NBG); E. Fort Brown (-BA), Bayer 1620 (NBG); Ballinafad (-BB), Smith 3365, 5404a, 5405a (NBG), Bayer 4652 (NBG); Horseshoe (-BB), Smith 2765, 3124, 5311, 5312 (NBG); Committees (-BB), Compton 17834 (NBG), Smith 5071, 5404 (NBG); 3km W. Committees (-BB), Smith 3382 (NBG); S. Committees (-BB), Smith in NBG322/40; Giffords Bush (-BB), Smith 1997 (NBG); 30km E. Peddie (-BB), Smith 3515 (NBG); Grahamstown (-BC), Britten 218 (PRE); Grahamstown (-BC), Britten in PRE 34903; Grahamstown (-BC), Britten in PRE 39480; Grahamstown (-BC), Curator PRE Bot. Garden in PRE 26300; 24km S. Grahamstown (-BC); Stayner in KG254/70; Blaaukranz (-BC), Smith 5544 (NBG); Fernkloof (-BC), Smith 5629 (NBG); Mt. Drive (-BC), Britten (NBG), Dyer 6 (BOL); Kowie (-BC), Dyer 7 (BOL); Kariega (-DA), Branch 43 (NBG); Bussock Farm (-DA), Smith 768 (NBG). 3327 (East London): Peddie (-AA), Smith 3112 (NBG); Paradise, Wooldridge (-AA), Smith 2800, 5602, 5671 (NBG); Gqora (-AA), Smith 5777 (NBG); W Woolridge, Peddie (-AB), Smith 3113, 3115 (NBG), Bayer 4648 (NBG); Kapp-Fish confluence (-AC), Bayer 4654 (NBG); Kaffirdrift (-AC), Smith 655, 5255, 5261, 5262, 5263, 5274 (NBG); E. Fish River (-AC), Smith 5256 (NBG); Wesley to Falloden (-AD), Smith 3157 (NBG); Chalumna (-BB), Smith 571, 2786, 3089, 3391, 3392, 5131, 5313, 5400, 6199 (NBG).
Inadequately located: Cape, Marloth 6284 (PRE), Long in Smith 3892 (NBG), Smith 398, 2804, 3112 (NBG), Stellenbosch 3889, 5560; ex hort, Ross-Frames in NBG76/48; Zaysdorp, NBG101825, Warden (BOL); ex hort, Whitehill (NBG); Albany, Dyer in NBG1806/30, Britten in NBG 734/31, Luyt in NBG309/45.
Haworthia cymbiformis var. cymbiformis JDV92/89 Kleinmond. Can have remarkable colours under stress.Haworthia cymbiformis var. cymbiformis JDV90/54 Beverly Cove. In dense clumps on steep rock faces.
b. var. incurvula (V.Poelln.) Bayer :124(1976). Bayer :36(1982). Scott :94(1985). H. incurvula V.Poelln., Feddes Repert.Spec.Nov. 31:85(1932). H. cymbiformis var. planifolia Flow.Pl.S.Afr. 9:t356(1929). Type: Grahamstown, Plutosvale, Mrs E. Ferguson. Not preserved. Neotype (B&M): Plutosvale, Britten (BOL71307).
incurvula: curved inwards.
As stated above, there is not very much substance to this variety either. It is smaller than the norm and relatively narrow-leaved. It has been collected many times from the same locality. The flower is very similar to that of H. gracilis var. minima, but Smith nevertheless maintained that it is continuous with H. cymbiformis. What he said was this “At the type locality near the top of the slope of a very deep valley, this plant is hardly variable, but as one descends, the plant changes, and at a point in the valley about a mile from the type locality, they are in appearance approaching H. cymbiformis.” The correct way to establish this is with physical evidence, and this is absent.
Distribution: 3326 (Grahamstown): Fish River Ridge (-AB), Britten in PRE 34959; Plutosvale (‑BA), Smith in NBG340/35 (BOL), Smith 5402 (NBG), Britten 11 (BOL), Britten in PRE 34909, in PRE 39477, Long 1029 (PRE), Dyer 3 (BOL), Dyer 2082 (PRE), Fourcade 99 (NBG), Smith 915 (NBG); Road to Plutosvale (-BA), Reynolds 2948 (PRE); 16km from Grahamstown (-BA), Erens in PRE 34910; S. Plutosvale (-BA), Smith 5402, 5403 (NBG); S. Hunts Drift (-BB), Smith 5741, 6508 (NBG).
Inadequately located: Albany, Dyer in NBG1803/30 (NBG), Smith 1123 (NBG), Luckhoff in NBG 404/34, Smith in NBG340/35, Britten in NBG740/31 (BOL); ex hort, NBG705/30, NBG1110/36, Luyt in NBG302/45, in NBG306/45, Whitehill (NBG).
Haworthia cymbiformis var. incurvula JDV93/60 Plutosvale. The relationship of this variety with H. gracilis is uncertain.
c. var. obtusa (Haw.) Baker JLinn.Soc. 18:209(1880). Bayer and Pilbeam, Cact.Succ.J(U.S.)46:166(1974). Scott idem. 48:260(1976). Scott :93(1985). H. obtusa Haw., Phil.Mag. 46:282(1825). Type: Cape ex hort Kew. Not preserved. Lectotype (designated here): Icon Kew library: H. umbraticola V.Poelln., Kakteenkunde 9:134(1937). H. cymbiformis var. umbraticola (V.Poelln.) Bayer :164(1976). Bayer :36(1982). Type: Swartwaterpoort, near Adelaide, W.E. Armstrong. Not preserved. Lectotype (B&M): icon (B): H. hilliana V.Poelln., Desert Pl.Life 9:103(1937). H. umbraticola var. hilliana V.Poelln., Feddes Repert.Spec.Nov. 44:234(1938). Type: Cape, ex hort Kew. Not preserved: H. obtusa var. pilifera fa truncata Jacobs., Handb.Succ.Pl. 2:574(1960). Type: ?. Not preserved.
obtusa: with obtuse leaves.
As discussed under H. cooperi, there is a real problem in understanding the situation concerning the blunt leaved forms of that species and of H. cymbiformis. I have seen very dark green forms with brown venation in cultivation which I would not relate to H. cooperi. The highly translucent forms from Fort Beaufort are greener and seem to be continuous with H. cymbiformis through the Swartwaterpoort. Certainly the plant mentioned above from the Kat River, are very close indeed in appearance to H. cooperi var. obtusata except for the brownish-green coloration. It has been noted that plants of that variety from Inverbolo (Upper Kei River) tend to become greener under lower light intensities. The herbarium record shows that the typical form of H. cymbiformis also occurs along the Kat River. Very robust forms in fact occur 10km S. Fort Beaufort. H. cooperi var. pilifera is also present in very close proximity. At Kagasmond and probably at Olifantsbeen nearby, the plants are very similar to the rather longer leaved forms, still with obtuse tips, which are found in Swartwaterpoort near Alicedale. A Kagasmond collection is also cited under var. dielsiana, which is indicative of the difficulty in using dry herbarium material to make indisputable identifications. G. Marx has made a collection from Swartwaterpoort in which the plants have the coloration of H. cooperi but the boat-shaped leaves of H. cymbiformis.
Distribution: 3226 (Fort Beaufort): Kagasmond (-CD), Bayer & Bruyns 6562 (NBG); Olifantsbeen (-CD), Krynauw in NBG268/43 (NBG); S. Adelaide (‑CD), Krynauw in NBG 67996; Blinkwater (-DA), Smith 6195 (NBG). Scott 600 (PRE), Bayer 4651 (NBG); Kat River, 10km SE. Fort Beaufort (-DC), Scott 1065 (PRE); Rocklands, Adelaide (-DC), Smith 2801 (NBG). 3325(Port Elizabeth (-BB), Bayer 4653 (NBG). 3326(Grahamstown): Thornkloof (-AA), Bayliss in KG382/76; S. Alicedale (-AC), Bayer 4650 (NBG); Alicedale (-AC), Britten in PRE 34905.
Haworthia cymbiformis var. obtusa JDV91/100 north of Fort Beaufort Is very variable in terms of leaf-shape and colour.Haworthia cymbiformis var. obtusa JDV96/63 Kagasmond. The plants have a greyer-blue colour than the Fort Beaufort plants and the leaves are more marginate.
d. var. ramosa (Smith) Bayer comb.nov. H. cymbiformis fa ramosa (Smith) Bayer :149(1976). Bayer :34(1982). H. ramosa Smith, JS.Afr.Bot. 10:22(1940). Type: CAPE-3427 (Peddie): Wooldridge (-AB), Smith 3168 (NBG).
ramosa: branched.
Consistency is a difficult ideal and this variety does not conform well with the principle of substance. It is only known from a long crescent-like rock-face north of Woolridge where plants vary from the normal stemless to increasingly stemmed plants on a gradient from west to east.
Distribution: 3327 (Peddie): Wooldridge (-AB), Smith 3168 (NBG, PRE), Smith 3105 (NBG), Bayer 4648 (NBG); NW. Wooldridge (-AB), Smith 3168, 3169 (NBG).
Haworthia cymbiformis var. ramosa JDV85/53 Wooldridge. The population is quite extensive and not all the plants are as stemmed.
e. var. reddii (Scott) Bayer comb.nov. H. reddii Scott, Cactus Succ.J(US) 66:182(1994). Type: CAPE-3226( Fort Beaufort): Waterdown Dam, Cathcart(-BB), Scott 8968 (PRE).
reddii: for Dr V.B. Reddi.
Plants from this population at Waterdown Dam have been known for a long time and identified (Bayer,1982) as possibly intermediate between H. cymbiformis and H. batesiana. Col Scott similarly mentions both species names in his discussion and the matter appears rather problematic. The population was portrayed as a depauperate one, at least in my perception, with quite considerable variation between the few odd plants at the site. The few clones sampled in 1982 were not as robust as the one described by Scott, who also thought the population to consist of but a few individuals. A re-visit to the site by Bayer and Bruyns in 1996 revealed that the south-facing cliff alongside the dam is clothed with huge numbers of plants. The huge clumps are just like those of H. cymbiformis. Some of the plants have very distinctive translucent dots and lines, others unmarked and uniformly opaque. The floral characters mentioned by Scott are not definitive but the flowers do appear to have strongly colored veins. At the time (Bayer :30, 1982) the species batesiana was still upheld, although not positively associated with field populations. Since then the range of H. marumiana has been shown to extend to at least Queenstown and further north. The Cathcart population does not seem to belong there although the block patterning in the leaves does suggest this.
At the same time the known range of H. cymbiformis has been extended by P.V. Bruyns to the upper reaches of the Black Kei much nearer to Queenstown. This is on a south-facing cliff at the farm Turnstream. Here there are small forms which distantly resemble H. lepida as described and illustrated by Smith, and also suggesting the same possible link with H. marumiana and certainly with reddii. This is particularly so because the Waterdown Dam is on the upper reach of the Black Kei. A little to the southeast is also a population of larger plants on a very high west-facing cliff which are however with spined margins, and thus apparently belonging to the var. setulifera. The block-patterning in the leaves of these two populations is not as marked as the Waterdown plants. There is an old Galpin collection from even nearer to Queenstown. Col. Scott was not correct in his opinions about H. marumiana var. batesiana and it is evident from his book that H. marumiana was unfamiliar to him too. The population at the Waterdown Dam does not seem substantial enough in terms of viability nor range, to justify species rank. What is needed is cognitive exploration of potential new localities to substantiate already expressed opinions. In this case there are new records for var. batesiana as well as for H. marumiana and H. cymbiformis.
Haworthia cymbiformis var. reddii JDV92/113 Waterdown Dam. The translucent markings are quite variable.Haworthia cymbiformis var. reddii JDV96/68 Turnstream. A new collection north-east of Cathcart which confirms relationship with H. cymbiformis. Unfortunately this population is under-sampled and more information is required for a really good decision to be made.
f. var. setulifera (V.Poelln.) Bayer comb.nov. H. planifolia var. setulifera V.Poelln., Kakteenkunde 5:54(1938). idem., Feddes Repert.Spec.Nov. 45:163(1938). Type: East London, Stellenbosch 3332. Not preserved. Neotype (designated here): CAPE-3228(East London): Kwelegha bridge (-CC), Smith 5257 (NBG): H. cymbiformis var. obesa V.Poelln. idem. 45:166(1938). Type: Idutywa, Bashee River (-BA), G.W. Reynolds. Not preserved. Neotype (designated here): CAPE-3228(Butterworth): Xobo River, E. Idutywa (-BA), Smith 7796 (NBG).
setulifera: bearing small bristles.
In the earlier handbooks, the representative specimen for the species was poorly chosen from a Nahoon River specimen. It is apparent from Haworth’s acceptance of the Bot. Mag. illustration that the more western forms such as those at Baakens River or Howiesons Poort, would have been more in keeping with the original circumscription of either the typical species or H. planifolia. East and north of East London, H. cymbiformis begins to develop a thicker and shorter, more deltoid leaf and the teeth become markedly spined and von Poellnitz’ name is re-instated for this variety.
Distribution: 3128 (Umtata): Mquanduli (-DC), Walker in NBG2271/27 (BOL). 3227 (Kingwilliamstown): Highclere (AB), Bayer & Bruyns 6573 (NBG); Inverbolo (-BC), Bruyns (NBG); Bluewater (-DA), Smith 676 (NBG); Near Komgha (-DB), Marloth 6510 (PRE); Tangla River (-DC), Smith 3881, 3882a (NBG); Newlands Location (-DD), Smith 3510 (NBG); Pump Stn. (-DD), Smith 611, 2785, 3096 (NBG); Kings Farm (-DD), Smith 3071 (NBG); McCleantown (-DD), Smith 3126, 3127, 2883, 2884, 2885, 3882 (NBG); Slippery Drift (-DD), Smith 3122, 3125, 3134, 3134a (NBG); below Horseshoe (-DD), Smith 3097 (NBG); Fort Jackson (-DD), Smith 3133, 3388, 3389, 3389a (NBG). 3228 (Butterworth): Willowvale (-AD), Luyt in NBG180/42, in NBG17/46, in NBG57/46, NBG341/38; Xobo River, E. Idutywa (-BA), Smith 7796 (NBG), Smith in NBG341/35 (BOL), Reynolds 2850 (PRE), Reynolds 146 (BOL); Xobo (-BA), Reynolds in PRE 39470, in NBG660/38, Smith 2760, 2796 (NBG); S. Mooiplaas (-CC), Bayer 1706 (NBG); Gonubie (-CC), Smith 6826 (NBG); Kwelegha bridge (-CC), Smith 5257 (NBG); Kwelegha (-CC), Smith 5251, 7183 (NBG); Kei River (-CC), Holmes (BOL).
Haworthia cymbiformis var. setulifera MBB6573 Highclere. This is from the Black Kei north-east of Cathcart, very near to the Turnstream population of the var. reddii. The flowers and seed in this population seem to be very large.Haworthia cymbiformis var. setulifera JDV91/91 north-east of East London. Leaves spinescent and more triangular in cross-section.Haworthia cymbiformis var. setulifera JDV91/91 north-east of East London. ALways in mats on steep rock faces.
g. var. transiens (V.Poelln.) Bayer :162(1976). Bayer :36(1982). H. planifolia var. transiens V.Poelln., Feddes Repert.Spec.Nov.45:163 (1938). Type: Cape, Prince Alfred Pass, Archibald 327. Not preserved. Lectotype (B&M): icon (B): H. cymbiformis var. translucens Triebn. et V.Poelln. idem. 45:166(1938). Scott :94(1985). Type: Cape, Prince Alfred Pass, Lategan in Triebn.1137. Not preserved. Neotype (designated here): CAPE-3323(Willowmore): Prince Alfred Pass, Smith 5709 (NBG): H. cymbiformis var. multifolia Triebn. idem. 45:166(1938). Type: Uitenhage, W.E. Armstrong. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): Hellsgate, UItenhage (-CB), Smith 2794 (NBG): H. cymbiformis var. brevifolia Triebn. et V.Poelln. idem.: 165(1938). Type: Cape, Hellsgate, Uitenhage, Mrs I. King in Treibn. 1068. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth: Hellsgate, Mrs I. King in Smith 2756 (NBG).
transiens: changing into.
In the publication where Von Poellnitz published his two varieties, he actually expresses doubt about identifying three parent species. Yet he states that H. planifolia var transiens is between H. planifolia and H. cymbiformis, but nearer to the former ‘to look at’. Two pages further he describes a var. translucens of H. cymbiformis from the same locality. In this work the circumscription of var. transiens is widened to include the Uitenhage elements at Hellsgate. There are variants in the Gamtoos Valley (eg. Andrieskraal) which are reminiscent of this highly translucent form, and it is not certain just how these variants relate to either H. cymbiformis or H. gracilis. The element H. gracilis var. picturata is applied to those forms of that species which are very similar to H. cymbiformis variants. The ‘clear’ way (a comment made by Dr M. Hayashi) in which var. transiens is related to H. mucronata is symptomatic of the alternative solutions available in classifying Haworthias, which are not always clear. It is generally understood that the Little Karoo species are continuous with the Eastern Cape species and this is commonly expressed in Von Poellnitz identifications, and also a feature of Col. Scott’s species distributions.
Thus H. mucronata can be allied with equal facility to either H. cymbiformis or H. cooperi, when in fact in the field it is more intimately related to H. decipiens. The location of this note is a powerful reminder that distinctions between species are highly blurred and that alternative solutions are possible.
Distribution: 3323 (Willowmore): Prince Alfred Pass (-CC), Smith 5624a, 5709, 5624, 5624a (NBG), Taute (BOL), Fourcade 3490 (BOL); Oskloof (-DA), Bruyns 7077 (BOL); Luiskraal (-DA), Forrester 399 (NBG). 3324 (Steytlerville): Scholtzberg (-CA), Van Jaarsveld 7804 (NBG); Andrieskraal (-DA), Fourcade 176 (NBG). 3325 (Port Elizabeth): Hellsgate, Uitenhage (-CB), Smith 2794 (NBG); Hellsgate, Mrs I. King in Smith 2756 (NBG); Kemachs (-CB), Smith 905 (NBG); Near Port Elizabeth (-DC), Smith 3892 (NBG); Stayner in NBG46/56, Stayner in KG80/70 (NBG), Taute in NBG1283/36, in NBG468/37, Taute .
Inadequately located: ex hort, NBG486/30.
Haworthia cymbiformis var. transiens JDV86/110 Prince Albert Pass. Now known to be more widespread and plants in other populations can be greener.
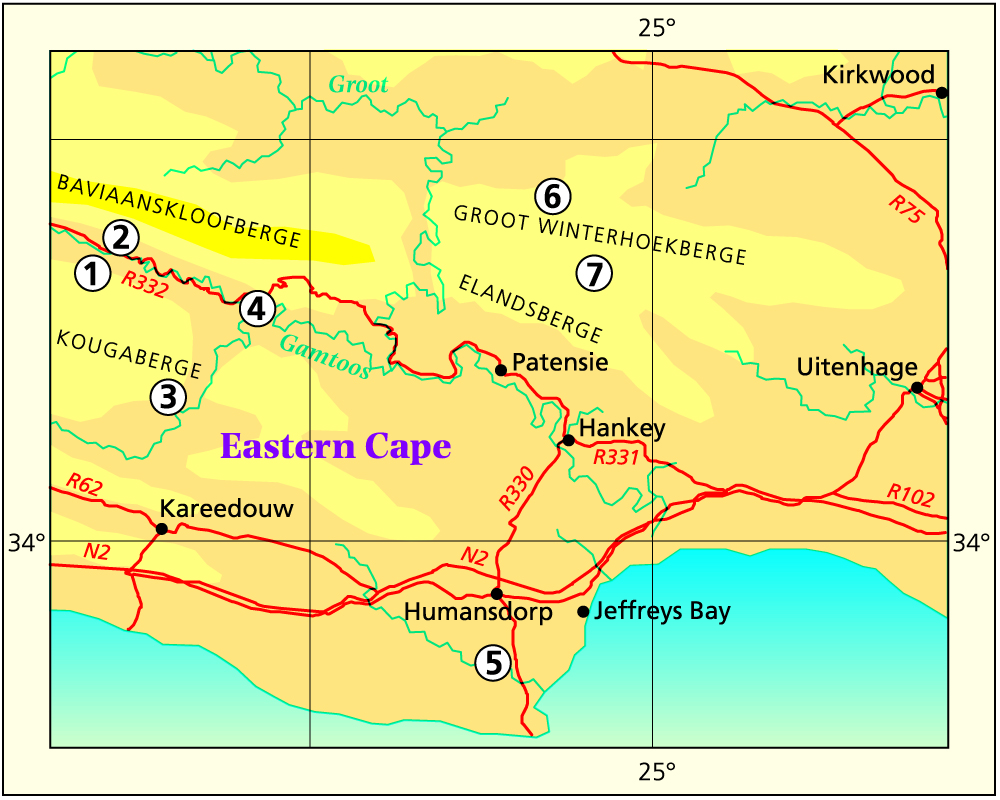
I will be disappointed if anyone had concluded I had any fixed ideas on the classification of these three species and their relationship. It has a problem which has long been on my mind. What happened recently (Nov.1998) is that I was offered the use of a time-share apartment at Jeffrey’s Bay, near the mouth of the Gamtoos River. I used this opportunity to spend six days in the field testing my hypothesis concerning the species Haworthia cymbiformis, Haworthia cooperi and Haworthia gracilis, and this is what I would like to record. Subsequent to that trip (Mar.1999) I planned and executed an excursion through the Baviaanskloof to Grahamstown and Stutterheim in March 1999, and repeated the exploration in Sept. and Oct, 1999.
Baviaanskloof and environs: 1-Engelandkop, 2-Rooikloof, 3-Moordenaarskloof, 4-Rooihoek, 5-Ripon, 6-Ouplaas, 7-Houtkloof
First I must point out again that there is a problem with plant names in that they do not necessarily reflect a set of objects which can be seen and easily recognised. The naming and typification process is partly to blame. Typification is simply the process of ensuring that a name is based on some substantial recognisable item, preferably a specimen rather than an illustration or photograph. (A specimen is preferred because it lends itself to some other analysis e.g. a tissue sample). The type is used to secure and permanently attach a name to for reference purposes. A type specimen and hence a name need not be, and often is not, typical of the taxon which the specimen represents. This does create difficulties particularly when varieties are at issue. In Haworthia there are an inordinate number of populations of uncertain pedigree. This is not because of some obscure evolutionary process, and nor is it just because there is an absence of characters by which to characterise and circumscribe each population. It is what we should expect. Everything is in a state of change and Haworthia is simply following the same unsteady path. I have shown that in Oxalis, in which there are an embarrassment of riches where characters are concerned, that these are of surprising little help in circumscribing the species. I was interested to read this sentence in a book about chameleons, “Without collecting data many chameleons are difficult to identify.” One would expect chameleons to be less difficult to identify than plants. Darwin stated that geographic distribution is the doorway to understanding species, and this is all we have in Haworthia which I can see to make sense.
We simply use ordinary visual observation and sensibility to make judgements about similarities. The less objects we have to examine, the easier it is to assess similarities among them. The problems that then arise are no different from those that develop even with complicated and convoluted discussions and explanations that arise from sophisticated techniques and analysis of characters. In my experience it is very often the direct visual judgement which is used to test the worth of technology rather than the converse. These comments are made because readers seem to think that plant names are as centrally defined as their own unexpressed concept they have of species being groups of things with well-defined limits and readily compartmentalised. Thus they imagine that there is a solution and it is just a question of how it is found. These comments are made because several times in the literature, wild statements have been made about the use of some character or technique or other throwing great light in the darkness. It just does not happen that easily and simply. The more sophisticated the technique, the smaller the sample and the less ‘peer’ review and replication. In statistics it is also pointed out that you do not undertake sophisticated statistical analyses when the figures already show the obvious. Yet how many times do analysts not persevere with test after test until a result does appear significant – itself a matter of probability. Technique may become and end in itself and the purpose for which it is devised lost to view.
There is a nomenclatural code for plant names which regulates about everything except what the binomial stands for. What should be pointed out with some vigour, is that there is a tendency to think that species can be described from single specimens in the paradigm (this is a most useful word because it suggests a pattern, a habit and a model of a time period) of Salm Dyck, Von Poellnitz, G.G. Smith and company. The single plant (and its description) and specimen that is used to typify a name, does not constitute a circumscription of the species. These people described specimens – NOT species. Their work was thus in some measure comparable with stamp collecting. It has little to do with present time description, organisation and understanding of the living variable and changing systems which species are. The different elements in Haworthia named as ‘species’ have been determined by early writers on the basis of vegetative characters such as colour, rosette shape, leaf number and shape, translucence and spination. Nothing much has changed except the scope and extent of the problem. A difficulty with characters is stated in Vavilov’s Law of Homology which states that a set of characters in a taxon will also be present in related taxa. Thus if one finds awned and obtuse rounded leaf tips in the genus, it is possible and indeed probable, that this character set will also occur in lesser ranks. If a set of characters is present in one species, they will possibly and probably occur in related species. This article is to demonstrate the difficulty of classifying plants on this simple basis and in an era when so much more material is there to classify.
It is necessary to discard the name translucens as I have used it in my earlier books. Originally the name really is associated with what is obviously H. herbacea, and I only used it to follow a tradition of misuse. The misfortune of this is that the name gracilis then occupies the nomenclatural hierarchy, but not the same geographic position in my understanding of the species. The consequence is that an outlying variant (Hellspoort, Grahamstown) will become the species name (i.e. gracilis) and a name has to be unearthed for the core of the species in the Gamtoos valley viz. H. gracilis var. isabellae. This in my new book replaces H. translucens var. translucens. Similarly H. cooperi var. cooperi is not central to the plot as is the var. pilifera, and I unearthed the latter name in my 1999 revision for this reason, as also the name gordoniana. I opted in 1985 to use the rank ‘subspecies’ under H. translucens for reasons which should be obvious – a weaker geographical relationship of the known variants of the species associated with a broader distributional phenomenon.
This following discussion is done in terms of Darwin’s statement about the importance of geographic distribution, and the evidence I have in my mind for a wide range of species. The essential problem with the species considered in this article is that we have a set of plant populations from Grahamstown, replicated to a degree by a set of populations in the Humansdorp area. In neither area is the relationship in the set well understood, and this does not imply that they can be so understood either. I set out to deal only with recent limited observations in the Humansdorp area but like a stone thrown into a pond, the ripples soon spread.
The link between the Grahamstown and Humansdorp areas is a curious one. It can be seen in the vegetation types of Acocks, which is a broad view of species associations. Valley Bushveld occurs discontinuously from Natal through to Swellendam. Alexandra Thickets are coastal, east of Port Elizabeth. Knysna forest, west of Humansdorp. Dry fynbos dominates the immediate Humansdorp area. Karoid vegetation dominates north of the Groot Winterhoek Mountains. It is at species level that really interesting facts emerge that impact on our analysis of Haworthia. What happens in the genera Aloe, Gasteria, Astroloba etc.? I leave this as a largely unanswered question because unless the reader can respond, he or she is unlikely to appreciate that it may have implications for understanding Haworthia. Astroloba does not occur in the Humansdorp area. There is instead the curious Haworthiapungens. The Baviaanskloof is home to several endemic Gasterias as well as to Aloe pictifolia. Haworthiaviscosa is ubiquitous. H. nigra with an otherwise fairly similar distribution is not. The distribution of the genus Encephalartos (cycads) is particularly interesting because of its known antiquity and the similarity of present day species to evidence in the fossil record. Aloe must also have an ancient history as evidenced by presence of species of this genus on Madagascar – a continental drift pre-history. It would in fact be instructive for would-be authors to read what R.A. Dyer wrote about the classification of cycads, before trotting out any banal aphorism about ‘active evolution’. There is an interesting break (interval) between the cycad species, east of the Sundays and Bushmans River valleys which separate Grahamstown and Humansdorp. This break can be observed in the distributions of other genera and species and evaluated both for compliance and non-compliance. Haworthia has both conditions e.g. cymbiformis and pilifera happily cross the interval. Angustifolia and coarctata only just do so. Marumiana, and nigra do so in the inland areas. Attenuata does so. Viscosa is almost ubiquitous. Fasciata does not and neither do sordida, longiana nor xyphiophylla. No species jump the Knysna forest interval and the genus as a whole jumps the Transkei interval northward only via limifolia and koelmanniorum (+mcmurtryi). These east-west breaks are interesting because they thus indicate speciation pressures. They may be helpful in deducing possible or probable relationships in a two-directional way i.e. distribution and variation in one set of plant variants provides a tool for the examination of other sets. In the case of H. gracilis, one can argue simply that it complies with the interval, in which case the var. tenera east of Grahamstown is discrete from the Uitenhage, Baviaanskloof look-alikes. Alternatively one can hypothesise non-compliance and that continuity will be found between Grahamstown and Uitenhage. Hellspoort (the site of the var. gracilis) is somewhat supportive of this view, but their is still a wide gap. Perhaps this is only in the known distribution record.
Known collections and observations were the basis of my 1985 classification hypothesis. Many of these were either dry herbarium specimens perhaps supported by photographs of single plants grown in cultivation. This hypothesis for the species in question here was arrived at from specimens from the immediate Grahamstown area and from the lower and eastern Baviaanskloof (Humansdorp) area. For the 1999 revision I have broadened the range of varieties, and relevant to this discussion are:
Haworthia cooperi
var. cooperi
var. leightonii
var. pilifera
var. gordoniana
and included Scott’s two new species as varieties:-
H. joeyii (= var. dielsiana)
H. venusta (var. venusta)
Haworthia gracilis
var. gracilis
var. tenera
var. isabellae
Haworthia cymbiformis
var. cymbiformis
var. incurvula
var. transiens
In this discussion I will largely avoid the issue of both H. bolusii var. blackbeardiana, H. cooperi var. cooperi and H. decipiens although they are closely involved. I will deal primarily with the Humansdorp (Hankey-Patensie); and to lesser degree with the immediate Grahamstown (Plutosvale) area.
I will use the term look-alikes because it is frequently used in the fynbos vegetation to refer to plant species from different genera, which are superficially so similar that close examination and full flowering material is often needed to establish their real identities. In Haworthia this detail is simply not available and thus there are specimens from different species which are geographically and thus probably genetically aeons apart, but which are visually identical.
Grahamstown (Albany) H. cymbiformis – is well represented and discrete. Yellow-green plants with spreading broader, flatter, ovate leaves. Clump forming plants on steep rocky faces. The var. incurvula – is known from many collections but only one locality (Plutosvale). It has look-alikes in the Patensie area particularly. Its relationship as a variety of cymbiformis really follows G.G. Smith who made the combination on the basis of a continuity which he claims to have seen. I recall (!) that its flowers are more reminiscent of tenera and this article may expose this as a possibility. Pale semi-opaque green, small clump-forming, with ovate, obtuse incurved leaves.
H. cooperi – is well represented by the var. pilifera. Blue-green plants tending to purplish, with incurved thicker, shorter ovate leaves, often spined. Solitary plants withdrawn into the soil. (The var. cooperi is not discussed in this appraisal although the concept presents a substantial problem on which I am preparing a manuscript).
H. gracilis – is known only from Hellspoort north-east of Grahamstown, with a second population of larger more cooperi-like plants from nearer Plutosvale (MBB6603-99, Glen Craig – a Gerhard Marx collection). Gracilis does have look-alikes in the Kirkwood area (Paardepoort, and a greater Humansdorp area). Paler grey-green plants tending to clump, with more leaves. The leaves are incurved narrow attentuate, and with or without spines. My opinion, based on my most recent collection from Hellspoort is that the var. tenera is actually the element concerned and is thus a superfluous name for gracilis. This var. tenera – is very well known from several populations north-east and east of Grahamstown and these include a glabrous population. It has look-alikes in the Uitenhage and Hankey areas of Humansdorp. Grey-green, small solitary or clump-forming, with many narrow, spined incurved leaves.
(Note:- the original description of gracilis seems to be of a moderately spined species, and von Poellnitz concept was apparently drawn from several disparate collections. The conception of the species from an illustration in Desert Plant Life, and indeed my own collection from Hellspoort in early 1970s, is of a moderately spineless plant. The recent collection of mine has forms which are relatively spineless as opposed to forms which are indeed similar to the smaller, densely spined, tenera. Thus we again have the problem where a name is neither central to the geographic element nor typical. The name tenera could be dispensed with and replaced with var. gracilis – this is evident from my discussion).
Humansdorp H. cymbiformis – represented by the typical variety in the small Baakens River valley and discrete. It may occur north-east of the Baakens River in the Rocklands area of Port Elizabeth. There is also the population at Hell’s Gate north-west of Uitenhage which I am regarding now as the var. transiens (It needs to be re-examined – hastening to add ‘by someone competent to do so’). This var. transiens – is only recently known to be very well represented in the upper Longkloof and in the Baviaanskloof. Its relationship with cymbiformis may not be as close as varietal rank, and the linking population at Hellsgate east of Uitenhage may be misunderstood. Leaves are incurved, pale-green and less opaque than in cymbiformis. Some of the variants resemble plants of the var. incurvula from Plutosvale.
H. cooperi as the var. pilifera – well represented and largely discrete. However, the typical variety with the necrosing-end area seems to have a western limit just outside Port Elizabeth and also near Uitenhage (the furthest west being at Perseverance, JDV90/40). It is then replaced by a variant which is often smaller, having the same blue-green colour darkening to purplish, but the leaf tips remain slender acuminate even in light conditions. This is what I have regarded as the var. gordoniana which in its typical form is not well represented.
H. gracilis as the var. isabellae – is well represented but apparently variable. The leaves tend to be spreading, pale greyish-green and spined. Some plants resemble gracilis from either Hellspoort (less-spined) or from Plutosvale (densely spined – tenera).
General It is known to me that in the area from Kirkwood, westward to the Little Karoo (north of the Baviaanskloof), there is an interaction between gracilis and decipiens. H. cooperi (not designating any variety) also occurs in that area. There is a problem in the Addo area, where another element (aristata – a separate weak hypothesis) may intrude. It has also been suggested, and this has always been in my own mind, that unicolor (mucronata) from the western Little Karoo may be continuous with either cymbiformis, bolusii or cooperi. It is continuous with arachnoidea, lockwoodii and with decipiens, and I am treating it as a separate issue because there are also other reasons for its exclusion here. The discussion could descend to the point where one says simply that there are not many species in the sub-genus Haworthia.
Results
Grahamstown I first looked at Hellspoort early in 1970 to collect and form an impression of H. gracilis. This was of a largish plant with leaves exceeding 3cm long, fairly spineless and weakly clustering. I also saw tenera in Plutosvale and at Hunts Drift. The former, small, densely spined and strongly clustering. At a later occasion I collected tenera again in Plutosvale as the conventional spiny element, and close by as a glabrous element of the same size. On that occasion I neither looked for nor saw incurvula or cymbiformis in the area. I revisited Hellspoort with J.D. Venter and G.D. Marx in 1996 and Venter made a re-collection again in 1998 (JDV89/42-117). My 1996 collection (MBB6614-118) reminded me forcefully of Plutosvale tenera, the plants were smaller than my previous collection and also spinier. Venter’s 1998 collection (all slightly different points within Hellspoort) was of the conventional gracilis that I visualise from the Von Poellnitz illustration, and from my early collection. These were quite large plants. During my visit of March 1999, I observed and recorded systematically from the bottom of Plutosvale, to the top (Roff’s Rock). On the north side of the kloof from the Cotswold farmhouse to the middle of the kloof which is Plutosvale, there was tenera both large clusters and relatively solitary plants. I could not locate or identify the site where I had collected the glabrous form before. The area has been degraded by the grazing of goats and it is possible the plants are now gone. From there we explored the south side of Plutosvale as the road climbed out of the kloof. Roffs Rock is very accessible and this no doubt accounts for the large number of herbarium records for incurvula. We found if from virtually the top of Plutosvale, to the lower and eastern point across from where we had observed tenera at our western-most search point. It is possible that the collection G.D. Marx in JDV93/73-93 was from further to the south-west than our starting point at Roffs Rock, at the upper point of Plutosvale. Nevertheless we found the same very pointed leaved plants of that collection, tending to be clump-forming. As we moved eastward, so the plants tended have leaves which were less pointed and more incurved. The colour was yellowish-green rather than the greyer or bluer-green of the Baviaanskloof incurvula look-alikes. The leaves seemed to lack the more swollen upper and end margins that may characterise cymbiformis.
My impression from this repeat visit to the Grahamstown area, is that the elements gracilis and tenera may be better considered as the same taxon. This may lay the basis for a better understanding, or be more compatible, with the range of look-alikes in the west around Humansdorp and Uitenhage.
The incurvula problem is now largely resolved and although it may still be necessary to find other links between incurvula and cymbiformis which Smith claimed and which is not supported by herbarium evidence. To the contrary, the collections of the gracilis-gordoniana plants from Glen Craig and Roffs Rock (viz. -99, -89, -93)), as well as the interplay of species in question in the Baviaanskloof, suggest that incurvula may equally be interpreted as a variant of H. gracilis or even H. cooperi. There is a further possibility based on new observations in respect of H. bolusii var. blackbeardiana. It may be possible to restructure the classification to relate the var. bolusii with H. semiviva; and to consider the element blackbeardiana as a variety of cooperi, (as nomenclaturally correct but geographically weak) as the species, with pilifera and gordoniana as principle variants. Nomenclatural considerations in ‘the strict terms of the code’, might confuse the issue, but I see no reason why reasoned and logical argument could not be used to dismiss names which are confusing because of their geographical connotation.
Humansdorp I looked at populations at Jeffrey’s Bay, Zuurbron between Humansdorp and Hankey, two at Draaihoek to the south-west of Patensie, three near Andrieskraal, one further west in the Groot River Poort, and three still further west in the Baviaanskloof, one at Patensie, one at Houtkloof in the Elandriver valley, one at Hankey, one north of Hankey, two south-west of Hankey, and one at the Gamtoos Bridge. Some of these populations were known to me before, and many of them are already represented in the Herbarium. I will also refer to other collections (mostly multiple plants) in my discussion which I have either not seen in the field at all, or in the more distant past. These are collections by credible field botanists and naturalists such as P.V. Bruyns, G.D. Marx, E.J. van Jaarsveld, T. Dold and D. Clarke. This thus reviews some 60 populations, but does exclude some significant ones in the Kleinwinterhoek mountains, where other considerations apply.
MBB6792-20 Jeffrey’s Bay. Fynbos. Very short vegetation on level sandstone outcrop. Early bud stage. Small single plants to 20mm diam. Leaves incurved at tips, no marginal spines and bluish-green, darkening to purplish hue (3) – gordoniana-picturata.
MBB6933-21 E. Humansdorp. Grassy fynbos. Rocky sandstone knoll. Small single plants, bluish-green with coarser spination than MBB6793 (2) – gordoniana.
MBB6553-22 and MBB6793-22 Zuurbron. Valley Bushveld. Scrub vegetation on level alluvial stream bank with loose sandstone rocks. Flowering and seeding. Small single plants to 30mm diam. Leaves thicker, leaves incurved at tips, many small closely spaced marginal spines, bluish- green (2) – gordoniana (possibly close to type locality).
MBB6784-23 SW Patensie. Valley bushveld, west sloping conglomerate. Seeding. Small single plants to 30mm diam. Leaves incurved at tips, forms with and without marginal spines, and blue-green (1). Smaller plants of this order were collected by Tony Dold at Drinkwaterskloof in the Baviaanskloof near Geelhoutboskloof – gordoniana.
MBB6786-24 SW Patensie. Valley bushveld, steeper south facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaves incurved at tips, mostly plants with marginal spines, blue-green (2) – gordoniana-isabellae.
MBB6827-27 Rooihoek. East of Geelhoutboskloof. Steep east facing slope. Plants single, greenish, very small – picturata-transiens.
EvJ14680-28 Vetmaakvlakte, S Rooihoek – isabellae (as for a clone of -41, resembling H. aristata).
MBB6789-29 Groot River Poort (Komdomo). Valley bushveld, very steep west-facing shale cliff. Flowering. Small clustering plants to 30mm diam. Leaves with spreading tips, slightly obtuse, no marginal spines, pale-green (5) – transiens-picturata.
MBB6830-30 E Komdomo. Riverine margins. Large clustering plants on steep soil-covered rock- gracilis.
J.N.Reddi in JDV93/86-31 2km NW Andrieskraal, is a collection which may be the same as -30 – gracilis.
EvJ15927-32.1 N. Groot River Poort. Steep south-facing cliffs. Clustering or single. Fairly similar to -29 – transiens-isabellae, the clone 32.2 to decipiens var. minor?
MBB6790-33 Andrieskraal. Valley bushveld, very steep south-facing conglomerate. Flowering and seeding. Small clustering plants to 30mm diam. Leaf tips moderately incurved, slightly obtuse, no marginal spines, pale yellow green (4+Y) – picturata.
MBB6791-34 Andrieskraal. Valley bushveld, steep east-facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaf tips incurved, slightly obtuse, no marginal spines. Pale yellow green (4+y) – picturata.
MBB6930-35.1 E. Andrieskraal, Nuwelande. Steep south facing conglomerate. Small plants down to 16mm diam. As above (4+y). – picturata. JDV90/55-35.2 from nearby westward, is also small plant with very pronounced windows.
MBB6928-36 Nuwelande. Upper steep conglomerate cliff. Plants nearly spineless and the leaves more slender and terete. Greenish (4). These are similar to JDV94/115-37 from the Paul Sauer Dam wall and also from the site for Gasteria glomerata where it was collected by E.van Jaarsveld (EvJ11076 in JDV90/118-38.1&2). A plant from that collection (-38.3) also resembles the Ripon plants (MBB6932-39) which have broader more deltoid, and more spinose leaves – isabellae.
J.G. Marx298 in JDV94/30-40, also Paul Sauer Dam, include both spined and spineless plants which also appear to be (4) – isabellae.
MBB6799-41.1 also (-9&-10) NW Patensie. Valley bushveld. South facing conglomerate slope. Seeding. More robust single plants to 45mm diam. Leaf tips incurved, with and without marginal spines. Pale green and blue-green (1-3) – gracilis-isabellae. One clone in this quite variable collection, strongly resembled a large specimen of H. aristata (-125,-142,-143).
MBB6801-42 SW Hankey. Early flowering. Valley bushveld. Vertical conglomerate. Small clustering plants to 20mm diam. Leaf tips incurved, with marginal spines. Pale blue-green (2-3) – isabellae.
MBB6802-43 SW Hankey. Valley bushveld. Steep east facing conglomerate. Early flowering. Plants mostly single to 30mm diam. Leaf tips tending to spread, with marginal spines. Pale blue-green (2-3) – isabellae.
PVB7128-44 Holriver, far S Patensie. Steep north-facing cliffs. Similar to above – isabellae.
D.Clark1050-45 Kouenek, Geelhoutboskloof. No detail, similar to above. – isabellae.
MBB6773-46 N Kareedouw. Fynbos. Conglomerate. South facing. Flowering late Dec. Plants single small, leaves incurved, very spiny and spines on leaf surfaces too. Blue-green (2) – isabellae.
MBB6771-47 Moordenaarskloof (N. Kareedouw). Steep south facing slope. Plants in small clusters or single. Relatively spineless. Leaves incurving. Green (4) – picturata.
MBB6805-48 NE Hankey. Valley bushveld. Gradual west facing slope. Seeding. Plants single to 50mm diam., leaf tips incurved, with and without marginal spines. Blue-green and pale-green and (1-4) – gracilis-isabellae.
MBB6804-49 N Hankey. Valley bushveld. Dense vegetation on alluvial stream bank. Seeding. Plants single to 50mm diam., leaf tips incurved, without marginal spines. Pale-green and blue-green (1-4) – gracilis-isabellae.
JDV97/3-50 E Hankey. Valley Bushveld. Plants similar to above – (1-2) – gracilis-isabellae.
MBB6808-51.1 Gamtoos Bridge. Valley bushveld. Steep west facing conglomerate. Seeding. Clustering plants to 50mm diam. Leaf tips spreading, with marginal spines. Blue-green (2) – isabellae. There are very similar plants at Longmore, east of Loerie (MBB6555-51.2).
MBB6798-52 Houtkloof (Upper Elandsriver valley). Fynbos. Rocky valley in sandstones. Neither flower nor seed (flowered in cultivation in Jan.). Plants single to 35mm diam. Leaf tips incurved, very spiny. Blue-green (2) – isabellae.
C. Marais in JDV96/95-53 Forest Glade and JDV92/136-54 Oaklands, both Elandsriver, are similar but more coarsely spined and paler green (4) – isabellae.
MBB1404a in JDV86/13-55 is an old collection of a tenera-like plant from the lower Elandriver valley (Groendal Dam) – cf. tenera.
D.M. Cumming6831 is from nearby and is more typically isabellae-like).
PVB7040 in JDV97/8-57 and PVB7944 in JDV97/7-58 Ouplaas (E. Cockscomb) are very similar to the above from the Elandsriver, but the plants are more robust, with much wider and more obtuse leaves. Particularly interesting because similar plants also occur in the Kleinwinterhoek eg JDV93/41-59 N Campherpoort, JDV87/180-60 S Campherpoort, JDV91/136-61 N Wolvefontein, JDV92/140-62.1 and JDV94/45-62.2 both Wolven (all decipiens var. minor). There is also the collection from the northern Groot River Poort (EvJ15927 – 32.2) linking these, and perhaps also the Cockscomb and Elandsriver plants (ie. isabellae) with transiens.
JDV97/6-63 W Braamrivier. Dry Fynbos. Steep W. facing cliff. Clump-forming, short obtuse incurving leaves. Pale green (5) – transiens. EvJ15342-64 Dieprivier, NE Kareedouw; EvJ17548-65 Horee, Saptou, SE Uniondale; PVB7077 in JDV97/5-66.1 Oshoek, E of Uniondale; MBB6729-66.2 S Uniondale, and Reddi in JDV93/54-67 Kabeljouwsriver, near Jeffrey’s Bay (a doubtful collection), are also this element – transiens.
The latter is similar to -29 Groot River Poort (transiens-picturata), ie. small clustering plants to 30mm diam. Leaves with spreading tips, obtuse, no or very few marginal spines, pale-green. I could not locate this population and it would prove that gracilis, transiens and cooperi co-exist. E. Aslander1247-68 made a collection from the place reported by Reddi, and these plants are unquestionably gordoniana-like.
PVB7093 in JDV97/1-69 Skrikrivier, north of -63 is similar to that collection, but leaves more elongate acuminate and some lightly spined. Flowered late Jan. Very reminiscent of gracilis var. gracilis and particularly a brighter green variant from Paardepoort (MBB6600-70, var. viridis in the new revision) in the Kleinwinterhoek mountains to the north-east. PVB5402 in JDV97/20-71.1 Palmietrivier and MBB6589-71.2 Dorschfontein, both E. Steytlerville, are similar to this latter collection but better related to decipiens var. minor. A problem arises here in trying to limit the scope of the paper because there are similar collections from north of Glenconnor (JDV91/17-72.1), and south of Lake Mentz (JDV91/116-72.2, which grows with gordoniana JDV91/115-73).
MBB6810-74 (also -11&-12) Joubertina. Fynbos. South facing, rocky sandstone slope. Neither flower nor seed. Plants single to 50mm diam. Leaf tips incurved, without marginal spines. Blue-green (1) – gordoniana.
MBB6811 & JDV90/80-75 (also -13) Uniondale Pass. Renosterveld. Rocky defile, steep sandstone. Early bud stage. Plants single to 25mm diam. Leaf tips incurving, with and without marginal spines. Blue-green (1) – gordoniana. PVB7079-76 Saptou, (Upper Longkloof), PVB7062-77 Redcliffe (NE Willowmore), JDV91/80-78 Engelandsekloof (Baviaanskloof), JDV94/95-79 Nuwekloof (W. Baviaanskloof), are all in this class. A collection G. Marx194 in JDV91/81-80 from Apieskloof (Baviaanskloof) is unusually pale green but otherwise also seems to compare with these gordoniana-like plants).
In completing this review of collections I would like to mention five which seem to touch on this issue, but which are even more relevant to the H. cooperi var. cooperi and H. bolusii var. blackbeardiana issue. These are G. Marx in JDV91/14-145 De Plaat (NW Kirkwood), PVB5002 in JDV92/33-147 Kaboega Gorge (Suurberg), JDV96/89-116 Gladhurst (S Adelaide), J.G. Marx in JDV96/4-98 and MBB6603-99 NE Grahamstown (Glen Craig), and JDV93/73-93 upper Plutosvale. These are all plants with elongate acuminate leaves about as wide as thick, and almost or completely without spines. They could be confused with H. cooperi var. leightonii, or with clones of H. bolusii var. blackbeardiana – and of course with H. gracilis var. gracilis.
Discussion In trying to circumscribe each collection I have been aware of my deficient descriptive skills. Can one rationalise such similar things one observes in writing in a way and which can lead others to identifications? I have elsewhere pointed out major weaknesses in the capacity of persons to compare what is written with what is seen, and also to compare plants and illustrations. Apart from the problem of variation in the populations, there is a problem of different habitat conditions (light, soil, temperature and moisture) which will cause phenotypic differences. It should be possible to define these if the plants could all be grown under the same conditions. However, local variation and sample size also becomes a problem. Colour must be critical. There are four main classes in this set of species under discussion – the opaque yellow-green of cymbiformis, the darker blue-green of cooperi which can develop reddish veins and a purplish colour under stress, the mid pale (or light) grey-green of translucens and the subdued pale green of transiens. Under stress the latter becomes very pale. In the descriptions above I have categorised colour on a scale of 1-5 from blue-green through to pale green, and added Y to indicate a more opaque yellow colour. In my book I recognise a variety (i.e. MBB6791-34 picturata from Andrieskraal) in which the transition from opacity to translucence is abrupt. This collecting trip does not strongly support such a geographic entity although their may be populations in that area which do. Certainly in the var. transiens, some plants are also very conspicuously patterned. Flowering time in early summer for all these collections appears to be very much the same. Curiously the Jeffrey’s Bay collection was still at bud stage when most of the other collections were seeding already. When I visited this population six weeks earlier in the previous year, the plants were already in flower.
The off-setting, clustering character is also worth commenting on. As in the case of H. turgida and H. retusa (remembering also an expressed contention of mine that the latter can be regarded as a variant of the former), clustering is usually associated with very steep rocky situations as opposed to plants on level sites being solitary. In this particular study the slopes varied from level through moderate to vertical. Clustering was most pronounced on vertical sites and at Andrieskraal where vertical and sloping sites were nearly adjacent, the plants were clustering on the vertical and solitary on the sloping. Clustering may be accompanied by the capacity of the plant stems to elongate. It is significant that populations around Hankey and Patensie which seemed to be var. gracilis, were invariably on moderate sloping or level ground (NOTE. I have previously always identified these collections as gordoniana), and using the name gracilis now is derived from the product of the papers in the compendium. My later papers suggest that transferring these elements to H. cooperi is worth consideration).
The above collections confirm my observation that Haworthia in the Baviaanskloof are very difficult to classify and circumscribe. It does seem on the basis of collections by Bruyns, Van Jaarsveld, and Venter that transiens is a very substantial element to the west, which may not have a strong connection with cymbiformis. Gordoniana is apparently a much stronger element than I had previously supposed and I am comfortable with the decision to now recognise it as a valid variety of cooperi. The above collections in the Longkloof, and some other recorded collections, seem to show convincingly that it effectively co-occurs with transiens. In the area just west of Patensie, three collections described above seem to indicate that gordoniana intergrades directly with transiens and this transition gives rise to plants which resemble incurvula and which I identify as H. gracilis var. picturata. Similarly south-west of Patensie gordoniana (-24) seems to alter to isabellae. South west of Hankey this latter variety (-42) resembles tenera. The resemblance to tenera is also apparent in the Groendal area of Uitenhage. The Elandsriver collection (-52), and a similar one by Clifton Marais in the same valley (-53), appear to be intermediate in appearance between tenera and isabellae. Isabellae as a separate entity is also represented by the Gamtoos Bridge collection (-51), and an old collection by W.R. Branch from the Krom River estuary at Ripon (-39).
A major difficulty is the Jeffrey’s Bay population (-20). These small solitary plants seem, by virtue of their growth habit and the near co-occurrence with isabellae (perhaps transiens too, if Reddi’s collection -31, was confirmed), to be the local equivalent of cooperi var. pilifera. However, if they had been observed at Grahamstown, they would be taken to be gracilis. In cultivation they have come to bear a stronger resemblance to the -47 which is closer to picturata than to gordoniana. The Zuurbron population (-22) passes for the equivalent of pilifera with some difficulty, and this does transpose to isabellae via Draaihoek, SW of Patensie (-24). The gracilis-like plants around Hankey (-48,-49,-50) and Patensie (-41) too, often present an isabellae facies. Thus we are faced with the possibility of a strange inversion. The upper Longkloof has cooperi as the var. gordoniana, but in the lower Longkloof similar plants may be derived from gracilis. A similar problem is presented by the similarity of gordoniana-like plants to H. bolusii var. blackbeardiana at several different places in the wider Eastern Cape.
The collection -46 is particularly interesting (I am indebted to Ernst van Jaarsveld for its discovery). This is a very small isabellae which has the same colour and (spininess) hairiness of the leaf surfaces as has Scott’s venusta. This may be very significant in providing supporting for an hypothesis that prior to inundation of the continental shelf (12-18000yrs ago) there was less of a vegetation interval east and west of the Sundays and Bushmans Rivers. I have suggested that this is reflected in the relationship and distribution of H. coarctata and H. reinwardtii, and will suggest elsewhere that H. fasciata may also be evidence of such a situation. However, this is very speculative. The surface spination is also evident in some collections from north and south-west of Jansenville (-129,-130,-131,-132 and -133 which I have identified with either decipiens vars. minor or pringlei), although the spination is sparse.
Conclusions These specific observations suggest that my classification hypothesis (the classification as presented in my two handbooks and in my recent revision) is a sound communication medium for the two areas in question, without implying thereby that it cannot be improved. What is problematic is that in the greater Baviaanskloof area, the three elements never seem to occur together. I repeat what I have written in respect of H. cooperi and H. bolusii var. blackbeardiana, and in my handbooks and revision. It is also stated indirectly in the preamble to this paper. “The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain good will.” In this particular article, the key question would have been:-
“Do three elements we can identify as separate living systems representing the species cooperi, cymbiformis and gracilis grow in the same close geographic proximity?”
Evidence available to me and in the herbarium record, is that they do not. There are a few known instances where two of these species are in such proximity – but not all three. What is curious is that it is rare for any two species of the subgenus Haworthia to be in immediate proximity to the extent that they share any specific habitat.
Observations seem to confirm my opinion that there is a connection between the three species in their occurrence east and west of the Sundays and Bushmans Rivers. What does concern me is the relation between cymbiformis and transiens, and the implications these observations have for incurvula, gracilis and cymbiformis around Grahamstown. The set of plants around Grahamstown favours cooperi, cymbiformis and tenera, while the mirror set of the Humansdorp area, favours gordoniana, transiens and isabellae. The relationship of incurvula needs to be explored. In the eventual integration of all these collections, sight should not be lost of the complex situation around Fort Beaufort, where the same set of species and variants seems to be present. Nor should it be forgotten that in the Kleinwinterhoek mountains H. decipiens becomes involved. The situation north of the Groot River is complex and there are substantial records from the Perdepoort and from Willowmore and Steytlerville which indicate a close interaction between decipiens and gracilis as well as gordoniana.
There are a good many other collections from the upper Baviaanskloof, but mostly these are represented only by herbarium records and I would like to see this material in its living state.
A final thrust of this article is to emphasise that Haworthia is not going to be understood by botanists or enthusiasts sitting on some other continent. This is particularly if they depend on the convolutions of the Code of Nomenclature, literature and limited collections for their information and the opinions they feel compelled to express. Less so if they do not and cannot, read and write the English language sufficiently well to communicate properly. Still less so if they have no knowledge and feel of South African geography and vegetation, and are operating in an outdated paradigm where specimens represent entire species. Explanations which only account for a few plants in cultivation are not going to be very satisfactory. The fact must be faced that classification is difficult and more information makes it more difficult. This is not a problem peculiar to Haworthia. It only seems so because the genus has received so much attention. The question now is:
“How is this information recorded and presented in a way that can be accessed, understood and appreciated?”
My conclusion is that we now need to resort to computerisation and digital imagery, recognising that dealing with single plants is not the same as dealing with populations. It is not easy to assess populations objectively as the sample sizes become unmanageable. We should progressively assess and re-assess populations and build up good herbarium documentation. In this way a proper revisionary process is set in motion, which limits Micky Mouse decisions about hypotheses, classification and nomenclature. ♦
1 a. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.1 b. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay.2 a. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 b. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.2 c. MBB6933 H. cooperi var. gordoniana, east of Humansdorp.3 a. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 b. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 c. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.3 d. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron.4 a. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.4 b. MBB6784 H. cooperi var. gordoniana, southwest of Patensie.5 a. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.5 b. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie.6 a. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.6 b. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof.7. MBB6825 H. cymbiformis var. transiens to H. gracilis var. picturata, lower Geelhoutboskloof.8. MBB6827 H. gracilis var. picturata to H. cymbiformis var. transiens, east of Geelhoutboskloof.9. EvJ14680 H. gracilis var. isabellae, Vetmaakvlakte, south of Rooihoek.10 a. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 b. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 c. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 d. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 e. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).10 f. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo).11 a. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 b. MBB6830 H. gracilis var. gracilis, east of Komdomo.11 c. MBB6830 H. gracilis var. gracilis, east of Komdomo.12. JDV93/86 H. gracilis var. gracilis, 2km northwest of Andrieskraal.13 a. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 b. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 c. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 d. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 e. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.13 f. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort.14 a. MBB6790 H. gracilis var. picturata, Andreskraal.14 b. MBB6790 H. gracilis var. picturata, Andreskraal.14 c. MBB6790 H. gracilis var. picturata, Andreskraal.15 a. MBB6791 H. gracilis var. picturata, Andrieskraal.15 b. MBB6791 H. gracilis var. picturata, Andrieskraal.15 c. MBB6791 H. gracilis var. picturata, Andrieskraal.15 d. MBB6791 H. gracilis var. picturata, Andrieskraal.15 e. MBB6791 H. gracilis var. picturata, Andrieskraal.16. 1 a. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 b. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 c. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 1 d. MBB6930 H. gracilis var. picturata, east of Andrieskraal.16. 2 JDV90/55 H. gracilis var. picturata, also east of Andrieskraal.17 a. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 b. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.17 c. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal.18. JDV94/115 H. gracilis var. isabellae, Paul Sauer Dam Wall.19.1. EvJ13809 H. gracilis var. isabellae, upper Paul Sauer Dam.19.2. EvJ11076 in JDV90/118 H. gracilis var. isabellae (smooth), upper Paul Sauer Dam.19.3. EvJ11076 in JDV90/118 H. gracilis var. isabellae (= MBB6932, Ripon), upper Paul Sauer Dam.20. MBB6932 H. gracilis var. isabellae, Ripon, Krom River.21 a. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 a. MBB6799 H. gracilis var. isabellae, northwest of Patensie.21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam.22. 1 c. MBB6799 H. gracilis var. isabellae, northwest of Patensie.22.2. MBB6799 H. gracilis var. gracilis to H. gracilis var. isabellae, northwest of Patensie – a clone resembling H. aristata.23 a. MBB6801 H. gracilis var. isabellae, southwest of Hankey.23 b. MBB6801 H. gracilis var. isabellae, southwest of Hankey.24 a. MBB6802 h. gracilis var. isabellae, southwest of Hankey.24 b. MBB6802 h. gracilis var. isabellae, southwest of Hankey.25 a. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.25 b. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie.26. D.Clark1050 H. gracilis var. isabellae, Kouenek, Geelhoutboskloof.27 a. MBB6773 H. gracilis var. isabellae, north of Kareedouw.27 b. MBB6773 H. gracilis var. isabellae, north of Kareedouw.28 a. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 b. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 c. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.28 d. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw.29 a. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 b. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.29 c. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey.30 a. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 b. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.30 c. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey.31 a. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.31 b. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey.32.1 a. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.1 b. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge.32.2 a. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.32.2 b. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie.33 a. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.33 b. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley.34 a. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.34 b. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver.35 a. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 b. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.35 c. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver.36 a. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.36 b. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage.37. D.M.Cumming6831 H. gracilils var. isabellae, south of Groendal Dam.38 a. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 b. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.38 c. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.39. PVB7043 in JDV97/7 H. gracilis var. isabellae, Ouplaas, east of Cockscomb.40 a. JDV93/41 H. decipiens var. minor, north of Campherpoort.40 b. JDV93/41 H. decipiens var. minor, north of Campherpoort.41. JDV87/180 H. decipiens var. minor, south of Campherpoort.42 a. JDV91/136 H. decipiens var. minor, north of Wolvefontein.42 b. JDV91/136 H. decipiens var. minor, north of Wolvefontein.43.1. JDV92/140 H. decipiens var. minor, Wolven near Wolvefontein.43.2. JDV94/45 H. decipiens var. minor, Wolven near Wolvefontein.44 a. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 b. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.44 c. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier.45 a. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 b. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.45 c. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw.46. EvJ15748 H. cymbiformis var. transiens, Horee southeast of Uniondale.47.1 a. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 b. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 c. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 d. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.1 e. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale.47.2. MBB6729 H. cymbiformis var. transiens, De Vlugt, Prince Alfred’s Pass, south of Uniondale.48. J.N.Reddi in JDV93/54 H. cymbiformis var. transiens, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay.50 b. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].50 c. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c].51 a. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 b. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.51 c. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains.52.1 a. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.1 b. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville.52.2 a. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 b. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.52.2 c. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville.53.1. JDV91/17 H. gracilis var. viridis, north of Glen Connor.53.2. JDV91/116 H. graciis var. viridis, south of Lake Mentz.54 a. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.54 b. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort.55 a. MBB6810 H. cooperi var. gordoniana, Joubertina.55 b. MBB6810 H. cooperi var. gordoniana, Joubertina.56 a. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.56 b. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass.57. PVB7079 H. cooperi var. gordoniana, Saptou upper Longkloof.58. PVB7062 H. cooperi var. gordoniana, Redcliffe northeast of Willowmore.59. JDV91/80 H. cooperi var. gordoniana, Engelandkop, Baviaanskloof.60 a. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.60 b. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof.61 a. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.61 b. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.
One of the greatest difficulties in Haworthia is that of trying to recognise discrete species. This translates into confusion which can be attributed to writers. The initial source of confusion is without doubt the nature of the plants themselves, and this is not a problem confined to Haworthia. The species are often not easily recognisable and discrete entities. I abhor the statement that the genus is in a state of active evolution, but this does at least seem to convey a message that readers understand, even if it is somewhat hackneyed. My observations on Haworthia are based on a definition of species as a system of living organisms which are continuous in time and space. In my New Haworthia Handbook, I suggested that a primary problem lay in separating H. bolusii and H. cooperi, and for the purposes of that work I largely discounted the secondary problems. My first concern was to identify core areas and names as working postulates. This did not mean I was unaware of lesser problems contained within the recognition of those two species. The purpose of this paper is to present my current understanding of the problem.
In my opinion the publication of Scott’s book and whatever merit it had, broke the foundation for understanding. As Bruyns said (Kew Magazine, 1986), it set taxonomy in Haworthia back by 40-50 years. This is because Scott did not attempt to examine what I had done, and was doing, in any objective and cognitive way. Thus there was no progression from a common hypothesis, or from a common concept of ‘species’. In my opinion his work was written in the paradigm of Reynold’s work on Aloe, or of Von Poellnitz and G.G. Smith, and was thus anachronistic. My work was in the intellectual climate of the day, was influenced by researchers of the time and was based on a definition of ‘species’. Scott’s thoughts and actions were both understandable and excusable. His classification reflects the same problems, but differently to mine. Where I recognised H. bolusii he recognised H. bolusii. Where I recognised the var. blackbeardiana he recognised H. cooperi and H. batteniae. Where I recognised cooperi he recognised H. pilifera and H. altilinea. Except for his contention that these latter are separate species, there is really concordance. We recognised different markers along the same continuum.
Essentially I considered that there were two species with two major facies, thus:-
1. H. bolusii – var. bolusii the smaller very densely spined typical variety which seems to diffuse into the species H. semiviva in the west of its distribution range.
– var. blackbeardiana a larger less densely spined element to the south and east which seems to diffuse into H. cooperi. Scott used the name cooperi for this element. There is merit in this, but he had to describe another species, batteniae, because of the inherent tension in the solution (I must point out here that Scott’s problem may have been the inadequate herbarium record that he used for his work, plus the fact that he made few specimens. Thus there is no way of assessing his decisions. Had he had to physically file specimens as I have done, he might have had great difficulty with batteniae and his cooperi, and also in separating his altilinea and pilifera). I regarded bolusii as a more spinose species than cooperi, with thinner and wider leaves. Cooperi then generally less spinose, squatter and with thicker leaves.
2. H. cooperi – var. cooperi itself, with relatively erect slender leaves, and including all the forms with more truncated abrupt leaves.
– var. leightonii – the coastal form with even more attenuate leaves than the typical form, and also very proliferous.
The solutions offered by either myself (1982) in respect of my cooperi and blackbeardiana, or in the case of Scott (1985), altilinea and pilifera, cooperi and batteniae, do have problems. These arise from the nature of the terrain in the area between Graaff Reinet to Queenstown and down to Fort Beaufort. Rugged broken terrain difficult to explore. Like the Baviaanskloof it offers many different classes of habitat, and thus potential ecotypes. The relationship between the varieties is complex and compounded by continuities with other species e.g. gracilis, cymbiformis, and decipiens. It should also always be remembered that any decision is a product of the collecting record and must emphasise that this is the context in which this article is written. Initially I did think that the typical variety of H. cooperi may possibly prove to be well-defined geographically. It has not proved indisputably so, but my classification nevertheless does allow good expression of the continuities which occur. I used the name to cover forms with erect slender leaves as well as forms with relatively blunt truncated leaves. Such plants do occur within the same populations and this is evident in the J.G. Marx collection from Fort Brown. The corresponding Scott names were altilinea and pilifera. To make my system more workable, in my new book (1999) I recognised H. bolusii and its var. blackbeardiana as before, but in H. cooperi, several more varieties, thus:-
– var. dielsiana – a more truncated version of pilifera, the leaves often without end-awns. (Scott’s “joeyii” is synonymous).
– var. truncata – a eastern lighter coloured, proliferous and smaller version of dielsiana.
– var. leightonii – the pinkish, proliferous, slender leaved form growing on granites at Kaisers Beach (Scott maintained this as a separate species).
– var. gordoniana – the erect, slender leaved form in the Hankey and Patensie area, which is very like the typical variety cooperi. In truth var. gordoniana from the type locality is a small very compact plant with short incurved and finely spined leaves.
– var. venusta – a very localised hairy variant from the coast near Alexandria.
I hoped this would explain the variations and resolve the tensions in the solution in an economical way. There is no doubt that Scott’s argument of cooperi sensu Bayer, and blackbeardiana sensu Bayer being the same (and he used only the name cooperi in this context), is correct in the nomenclatural sense. The Thomas River specimen he cites in his book as representative (although Scott does cite the Kew specimen as type, which I have not seen), would have been attributed by Bayer (myself, using the Saunder’s Refugium illustration as type) to my concept of H. cooperi, as also the doubtful gracilis-like elements for the Adelaide and Kingwilliamstown citations. I would have named the Cradock specimen H. bolusii var. blackbeardiana. To bridge the difficulties inherent in his solution, Scott had to later recognise three species viz. batteniae, pringlei and joeyii, with the potential for many more. I have said elsewhere, that a classification which grows with the description of new species, is indicative of a weak system. As in the case of bolusii, the Scott solution is not as economical as mine. His concept of an altilinea and pilifera is essentially the image of my problem in recognising a cooperi/blackbeardiana interface. It would have helped had I initially recognised pilifera as a variety. However, Scott’s interpretation in the sense of the geographical relationships of his various species is problematic throughout his revision, where specimens are not cited and full use has not also been made of the available herbarium record.
I was predicting that an element, namely H. cooperi var. cooperi occurred which could be defined in a geographic context. The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain goodwill. This article thus is written to expand on the problem and indicate where the difficulties lie. These are not difficulties that I was oblivious of, or tried to obfuscate. I have many times said that we can find solutions which work in one area, but they may not work in another. This will again be evident in this paper.
In my revision (Haworthia Revisited, 1999) I made the combination H. decipiens var. pringlei and resurrect the old species name H. aristata. These two elements, and an explanation for the tensions which these two names create and try to address, will be touched on here but will follow in more detail in a later article. As in this immediate case, new material has been seen and collected since the revision was drafted, and a better explanation can now be made.
For this particular issue, the key questions to the hypotheses of Bayer and Scott are difficult to formulate because they are confounded by the different use of names. Therefore I frame the questions in terms of my own classification like this:-
Does an element we can identify as cooperi grow in the same close geographic context as blackbeardiana? (Bearing in mind the assumption that the hypothesis regarding continuity between bolusii and blackbeardiana in fact is valid. Also noting that to maintain the classification hypothesis that there is a difference at the rank of species, the answer must be ‘yes’).
Does the element we can identify as cooperi grow in the same close geographic context as pilifera? (Bearing in mind that to maintain the classification hypothesis of a difference only at the rank of variety, the answer must be ‘no’).
Results The answer to question 1:- has never been a strong point of my classification because I saw cooperi in a broader sense to include pilifera (and altilinea of Scott). The recognition of varieties now strengthens the classification in one respect but weakens it in another. It strengthens the classification in respect of recognising the extent of the variability within my concept of H. cooperi. It weakens it in suggesting both a strong geographical separation of the varieties and a weaker relation with H. bolusii var. blackbeardiana. In Nov.1996, I went with Peter Bruyns to the Eastern Cape and we spent some time in the greater Somerset East area. In Dec.1996 I travelled with Kobus Venter. In Dec.1997, Dec.1998, in Sept. (with Tony Dold, Dez Weekes and Steven Hammer) and again in Oct.1999, I travelled with my wife to these areas. We made many significant collections.
The first of these concerns Scott’s species, H. joeyii and H. pringlei. In the case of joeyii my contention that it is continuous with pilifera barely needs discussion, and I do not think the presence or absence of an end-awn is necessarily diagnostic for such an element – hence synonymy with var. dielsiana. We found three discrete populations within a small radius around Eastpoort (MBB6558-102, 6559 and 6560-103) which support this observation. I do not think there are strong grounds for separating it from pilifera, although I have now done so. At Bedford (there are plants with the similar abrupt leaf-tip as well as plants without. I currently have four batches of seedlings which I want to examine for variation. One from Slagtersnek, south-east of Somerset East (MBB6778-104) is particularly variable. I even have a collection of H. cymbiformis (MBB6847-100,-105) which has the same truncated, awnless leaftips as Scott’s joeyii. Regarding Scott’s pringlei, my conclusion that it is related to decipiens is possibly geographically and otherwise incorrect. (It should be noted that no specimen of pringlei has to date been deposited in the Pretoria herbarium. My understanding of the species is from the description, from two plants sent to J.D. Venter by J.N. Reddi (JDV93/46-106) who collected the plants for Scott, and from two plants given to G. Marx (JDV93/52-107) also by Dr Reddi). It is a problem of ‘look-alikes’ and I will deal with that a little later as a separate issue.
Further south and east of Eastpoort, at Patryshoogte (MBB6561-108 also -4), we found a different population not to my knowledge co-occurrent with a cooperi variety, which must be comparable with Scott’s species i.e. pringlei. It has narrower and more elongate leaves than blackbeardiana generally has, and the keel may be more pronounced. Nevertheless I am sure it has a connection with H. bolusii var. blackbeardiana. In terms of a much wider knowledge of other populations, I decided to link pringlei to decipiens, to emphasise the evident and probable continuity between that species and H. bolusii. This is particularly so in the area north and east of Jansenville.
To the south-west at Somerset East (MBB6776-109, Glen Avon), we found a population of bluer green plants with rather more attenuate leaves than in the vars pilifera and dielsiana. These plants are very like an Adelaide (Koonap Bridge, MBB6563-110) collection, these both satisfy my concept of the var. cooperi. It was not much further to the south (-104), and also not far south of Eastpoort, where we found forms of the vars pilifera and dielsiana in the same population. Thus it seemed that a hypothesis maintaining the species cooperi and H. bolusii var. blackbeardiana, was well supported, but not so the contention that pilifera and dielsiana were good varieties.
The most significant discovery up to Dec.’97 was of two different elements growing together towards the south, near Ripon Station. By together, I mean as populations occupying different niches in the same close proximity – not growing as a common medley of individuals. One population was clearly the typical blunt-leaved pilifera (MBB6557-111) which included dielsiana, but the other (MBB6556-112) was with more slender and erect leaves viz pringlei, and possibly the leaves being broad and spinose enough to even represent blackbeardiana. Thus the added complexity is that it is not possible to separate those two elements. After two years in cultivation as field collected clones and as seedlings, the difference from pilifera (as -111) is maintained. The collected clones grown in Kobus Venter’s collection convincingly demonstrate that the one population is identifiable with pringlei/alias blackbeardiana and the other with cooperi var. pilifera. Thus the answer to the first key question is “Yes, they do grow together in the same geographical context, because there is that obvious connection of blackbeardiana with pringlei. It seems probable that both these elements could, and perhaps should, be interpreted as varieties of H. cooperi. It is a complex problem which cannot be separated from consideration of other elements such as H. gracilis and H. aristata, and these are the issues I am addressing in these papers.
There is not much further evidence that cooperi var. cooperi is a strong, discrete and valid element. There is a collection by J.G. Marx of plants in the Hogsback area (in JDV91/82-113, Woburn) which may be identified as this variety, as can the two collections already mentioned viz. -109 Glen Avon, and -110 Koonap Bridge (originally a Marx collection). However, there are collections generally (one is D.M. Cumming8489-114 of plants south of the Waterdown Dam, Upper Chilton), where the plants can be confused with H cooperi var. gordoniana (in the context of the problem in the Baviaanskloof), whereas geographically they should be blackbeardiana. I also made a collection at Waterdown Dam itself (MBB6569-115), of a plant which is very like a spineless smooth gracilis. These plants tend to be much more blue in colour than the southern look-alikes. There is also JDV96/89-116 Gladhurst, Adelaide; and JDV96/4-98 and MBB6603-99 from Glen Craig, north-east Grahamstown, which pose a similar conundrum. These are gracilis-like and while exploring that problem, the issue became very much more prominent, and is even expressed in the discussion regarding H. cymbiformis var. incurvula. Kobus Venter and I had both re-collected H. gracilis var. gracilis at Hellspoort (JDV89/42-117, MBB6614-118), although we had not concerned ourselves with a search over the whole of the valley. What is now becoming more evident is that there is an archetype which is gracilis-like. Thus Hellspoort needs to be examined as closely as Pluto’s vale.
A later collection of mine (Oct.’99, MBB6927-119, W Ripon) influences the picture dramatically and may further prove a statement of mine true. This is that there instances where there are no real boundaries between species. My collection is further west of -111 (pilifera/dielsiana) and -112 (pringlei/blackbeardiana). Here in -119, the plants include individuals which could be either identified as the vars cooperi or pilifera, with the former collection equally well representing pringlei/blackbeardiana.
The answer to question 2:- has not really been a problem. What we found was what we expected to find and more. South of Adelaide one finds populations of plants which satisfy the tendency towards slender leaves ie. var. cooperi (-109 Glen Avon or -110 Paardefontein, Koonap Bridge); and populations which satisfy the tendency to short obtuse leaves ie. var. pilifera eg. MBB6564-120 Chancery Hall or MBB6591-121 The Tower, S. Fort Beaufort. The same applies just east of Somerset East where in the broader geographical context we can identify var. cooperi (-109, Glen Avon), var. dielsiana (when mostly or wholly without end-awn, MBB6565-122, W. Somerset East, -111 W. Ripon Statio, and vars dielsiana and pilifera as geographically complementary and even in admixture (-104, Slagtersnek). Thus “No, the varieties cooperi and pilifera do not grow together in the same close geographical context as discrete entities.” In fact they are often represented as forms in the same collections as in -119 (far west of Ripon) where both varieties are present as single plants as well as a longer leaved form representing blackbeardiana as decipiens var. pringlei.
The answers given to these two questions thus do still not solve the problem as there remain populations which can not confidently be ascribed to either of the names here used. There are populations which may thus be assigned to cooperi or to blackbeardiana or pringlei. An example is the Marx collection from north-east of Grahamstown (W Fort Brown, in JDV91/85-123) which is ascribable to pilifera although some of the clones are inseparable from blackbeardiana. JDV98/39-124 from Brakkloof, northwest of Grahamstown poses a similar problem with the leaves of the plants tending to lengthen in cultivation. The ultimate difference then between cooperi and blackbeardiana becomes a very subtle one of degree of spination and leaf bulk. The issue is further complicated by the fact that four other elements are involved in the total geographical context, namely H. cymbiformis, H. gracilis, H. decipiens, H. aristata and H. arachnoidea var. xiphiophylla.
My most recent field work, was in following up information and collections concerning H. gracilis. Tony Dold of Rhodes University gave specimens of an Haworthia from the Annsvilla area to Gerhard Marx (MBB6851-125). He thought they may have some bearing on the existence and reality of my interpretation of H. aristata, and this is indeed the case. The plants are small, the darkish blue-green of the cooperi/blackbeardiana elements, and with more, smaller, and quite spinose leaves. Annsvilla is close to three localities for that species cited in my revision viz. Verdun, Stonefountain (re-collected in MBB6852-126 also -4) and Kommadagga. Following Dold’s find, recollection in the vicinity of each of those sites, produced plants which can be interpreted and identified in the light of the Ripon collections. While they equate in some way to H. bolusii var. blackbeardiana, they also do to H. cooperi and to H. decipiens var. pringlei. An (unfortunately doubtful) Kommadagga collection (EvJ sn.-127) is very similar to the long-spined gordoniana-like plants from the eastern Joubertina -74, Uniondale -75, Longkloof -76 and Baviaanskloof (Geelhoutboskloof -78, Nuwekloof -79). The leaves of the plants are rather flatter and broader, and so closer to blackbeardiana. In Sept.’99 Dold and Hammer found a small plant with shorter and squatter leaves than my concept of aristata, south-east of Kommadagga (in MBB6897-128). We also saw plants from the area east of Jansenville which seem to link the greater H. bolusii (i.e. var. blackbeardiana) complex with either H. decipiens or H. arachnoidea var. xiphiophylla. Included in that broad statement is an explanation for the specimens which I have cited as H. decipiens var. pringlei in my revision. These include recent collections from north of Jansenville (MBB6580-129), north-west (MBB6581-130), and south-east of Mt Stewart (MBB6582-131 also -6, 6583-132, also -7). When I first saw a specimen of pringlei my first reaction was recall of plants I had collected at Mt Stewart, and of plants said to have been from Jansenville. Thus I created H. decipiens var. pringlei, expecting to find this east-west continuity between blackbeardiana and decipiens. I did not think that the connection between Middleton (pringlei) and Jansenville would be found to be as evident and as strong as it is. There was already some evidence that there is south to north-east connection between Jansenville and, say, Cradock. This is of a connection between H. decipiens and H. bolusii.
A complicating issue is that MBB6587-133 (further SE Mt Stewart than -132) is a population which is H decipiens var. minor. A small plant with a tendency for incurving leaves but with the widely spaced large spines I associate with the more classical view of H. decipiens. The next collection was MBB6589-71.2 (a little south-east of -133) and it throws us into the H. gracilis context of smaller plants, clustering on cool south-facing rock faces. There are many collections which continue this trend to gracilis, and in fact I suspect to xiphiophylla too, and that will be dealt with as a separate issue. The field-work also revealed populations (J.G. Marx in MBB6845-134, east of Alicedale, and MBB6847-105, also Alicedale) which link cooperi to cymbiformis. This also can be dealt with together with two collections from near Kagasmond (MBB6562-135, south of Adelaide and JDV96/89-116, Gladhurst) as a slightly different issue.
Regarding H. aristata, I resurrected this name for several collections from the Eastern Cape, and somewhat justified by the Dold collection at Annsvilla (-125). However, it seems quite certain that the issue is clouded. Re-collections at the Soutkloof (JDV96/90-136, Addo, Dead Man’s Gulch) were more like pilifera, but there is evidence of deviation. Here D.M. Cumming (DMC3870-137 collected pilifera-like plants, whereas Venter, Marais and Bayer (-136) collected a variant – there was a more typical bluish-green pilifera and a plant tending to the opaque yellow-green of xiphiophylla. I returned there in Oct.1999 because I still had not seen anything like the plants I had seen there on an earlier visit, or like W.R. Branch’s original (WRB459 in JDV87/53-138), and the other cited collections. On this occasion I did find a small population (MBB6920-139) of this kind, and these confirm a relation to the more northern collections mentioned above. At the nearby village of Addo itself, there is the fairly normal pilifera (JDV86/117-140). The Annsvilla collection (Dold and Marx -125) and the recent collection of my own from Stonefountain (-126 also -5) seem also to be in the context of the Ripon collection (-112) of pringlei. These are, however, collections of much smaller plants than from Ripon. Thus it confirms for me that the Haworth names aristata and denticulata could easily have had their origins in plants from this area between Ripon and the Zuurberg, or further south to Soutkloof. My collections MBB6916-141, MBB6917-142 and MBB6901-143, from Kaboega and Hopewell are of the same order.
Discussion and conclusions There is still a very large area unexplored. There are several like collections from the greater area Kirkwood to Uniondale which I have generally ascribed to H. cooperi var. gordoniana. None of these are the short-leaved pilifera type. The teasing probability is that it is in fact blackbeardiana, pringlei, gracilis or aristata (as variants of H. cooperi) which are the main role players. They differentiate (clearly?) in the east to pilifera or cymbiformis and in the west into decipiens. In the south they pass to xiphiophylla, cymbiformis and varieties of gracilis. North-westwards it is to bolusii. My opinion now is that we have an archetype which is in the mould of H. gracilis and this is the root of all the elements I have named here. Curiously Tony Dold has recently sent me specimens from Chalumna (T.Dold3961 in MBB6921-144) which suggest that an aristata-like element is also associated with H. cooperi var. leightonii. Other Chalumna collections (MBB1621 and G.G.Smith 514) bear a very close resemblance to plants (-99, -116) which I have said are gracilis-like.
The above statements all have to be seen in the overall statement about continuity. It barely seems practical (nor legally possible) to sample and analyze plants on the scale that will be necessary, using whatever technique, to get a more definitive answer. The best answer will be continual exploration of the simple kind reported here, which explains occurrences on a smaller and smaller scale. There are herbarium records which need to be corroborated in the field but these do not suggest to me a better solution. Ultimately each recorded population will have to be assigned unequivocally to a taxon (or taxa!) and this is the next necessary step in Haworthia classification. Another revision based on less exacting field observations and a lesser record, will simply exacerbate the regression in time which Scott’s work precipitated, and the consequences which we now suffer. My recent collections are only preserved as living collections and photographically. Specimens need to be made.
I am very conscious of other tensions in my classification and I think it is imperative that we stay with one nomenclatural arrangement and hypothesis to resolve these. The confusion which arises from nomenclatural changes which are nothing but pretentious and cosmetic, is not worth any price. Any difficulties in respect of nomenclature can be resolved by a process of explanation and conservation without formal name change. These changes can be made when it is expeditious to do so and when changes can really offer a better explanation of the genus and hence better communication.
What this discussion should demonstrate is the problem of really observing and discussing variability in Haworthia. There are many possible arrangements of names which can be presented as conclusions in themselves – but done like this they simply cloud and destroy any hope of a broader understanding and good communication. Haworthia is now so much in the public domain, that I would suggest to editors that they move in the direction of encouraging authors to adopt a conserved nomenclature. I regret to be so straightforward and blunt. I see nothing but further confusion if persons feels that they can contribute to an understanding of the situation in the field without:
Consensus on the issue of species definition.
Consensus on the issue of a set of names.
Familiarity with herbarium records and what these represent in terms of fact and fixed reference points.
Familiarity with the written record.
Familiarity with South African geography and the ability to interpret populations in that context.
♦
102 a. MBB6558 H. cooperi var. dielsiana, north of Eastport.102 b. MBB6558 H. cooperi var. dielsiana, north of Eastport.103 a. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 b. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 c. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 d. MBB6560 H. cooperi var. dielsiana, south of Eastport.104 a. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 b. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 c. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 d. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 e. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.105 a. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 b. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 c. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 d. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 e. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 f. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.106 a. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.106 b. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.107 a. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.107 b. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.108 a. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 b. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 c. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.109 a. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 b. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 c. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 d. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 e. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.110 a. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.110 b. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.111 a. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 b. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 c. MBB6557 H. cooperi var. pilifera, west of Ripon station.112 a. MBB656 H. decipiens var. pringlei, west of Ripon station.112 b. MBB656 H. decipiens var. pringlei, west of Ripon station.112 c. MBB6556 H. decipiens var. pringlei, west of Ripon station.112 d. MBB6556 H. decipiens var. pringlei, west of Ripon station.113 a. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.113 b. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.114 a. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.114 b. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.115. MBB659 H. bolusii var. blackbeardiana, Waterdown Dam.116 a. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 b. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 c. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 d. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 e. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.117 a. JDV89/42 H. gracilis var. gracilis, Helspoort.117 b. JDV89/42 H. gracilis var. gracilis, Helspoort.117 c. JDV89/42 H. gracilis var. gracilis, Helspoort.118 a. MBB6614 H. gracilis var. gracilis, Helspoort.118 b. MBB6614 H. gracilis var. gracilis, Helspoort.119 a. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 b. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 c. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.120. MBB6564 H. cooperi var. pilifera, Chancey Hall, south of Adelaide.121 a. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.121 b. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.122 a. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 b. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 c. MBB6565 H. cooperi var. pilifera, west of Somerset East.123 a. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.123 b. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.124 a. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.124 b. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.125 a. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.125 b. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.126 a. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 b. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 c. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 d. MBB6852 H. aristata, Stonefontain, s. Somerset East.127. EvJ H. aristata, Aalwynspoort, east of Commadagga.128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.129 a. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 b. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 c. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 d. MBB6580 H. decipiens var. pringlei, north of Jansenville.130 a. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 b. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 c. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 d. MBB6581 H. decipiens var. pringlei, northwest of Janesville.131 a. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.131 b. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.132 a. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.132 b. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.133 a. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.133 b. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.134 a. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 b. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 c. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.135 a. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 b. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 c. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 d. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.136 a. JDV96/90 H. cf. aristata, soutkloof, Addo.136 b. JDV96/90 H. cf. aristata, soutkloof, Addo.136 c. JDV96/90 H. cf. aristata, soutkloof, Addo.137. D.M.Cumming3870 H. cf. pilifera, Soutkloof, Addo.138 a. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.138 b. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.139 a. MBB6920 H. aristata, Soutkloof, Addo.139 b. MBB6920 H. aristata, Soutkloof, Addo.140. JDV86/117 H. cooperi var. pilifera, Addo.
Introduction Kaboega (also spelt Kabouga) is now an assemblage of farms (De Plaat, Wilgerfontein, Vygeboomfontein, Klipfontein) nestled against the north slopes of the Zuurberg mountains, north of Kirkwood. It is only about 15km away from Kirkwood as the crow flies, but 150km away by road. Oudekraal is about 20km east and it is the source of Haworthia angustifolia var. baylissii and Gasteria baylissiana. There are several records of Haworthia for the Kirkwood area, and von Poellnitz named H. stiemiei (Regarded as insufficiently known and not recognised by Col. C.L. Scott or myself) from there. He also identified plants from Kaboega and Uyepoort, both described as “at Kirkwood”) as H. altilinea var. denticulata (Haw.) v. Poelln. These plants are all in the melange that I attribute to H. cooperi var. gordoniana (the subject of another long essay). The Kaboega farm lies on the Kaboega river which drains an area of about 1m ha and then flows through the long Kaboegapoort into the Sundays River just north-west of Kirkwood. The terrain is very broken with the sandstone Zuurbergs themselves dominating the southern boundary at about 850 to 950m above sea level. The lowest point on the farm is at about 300m and the northern lesser shale or dolerite peaks reach 550 to 650m. The vegetation on the sandstones is Dry Mountain Fynbos. North of this is Karoo Valley Bushveld. Thus Kaboega is at an ecotone of the karoid veld, Eastern Cape grassland and the Noorsveld (Euphorbia thicket) of the Jansenville area.
The name Kaboega means ‘the Big Hole’, referring to the deep gorge which the Kaboega River cuts through the Zuurberg. This river joins the Sundays River where it skirts the eastern end of the Kleinwinterhoek Mountains. Thus Kaboega Gorge is about 20km east of the Sundays River Gorge and 20km west of Oudekraal where the Witrivier also cuts through the Zuurberg flowing southwards.
The only known Haworthia collections from Kaboega prior to this report are two collection by Gerhard Marx (JDV91/14-145, JDV91/15-146) from the easterly part of the farm (DePlaat, north and south aspects), and a similar collection by Peter Bruyns (PVB5002 in JDV92/33-147) from Kaboegapoort itself. Discounting the strange (expected) internal variation in the latter, these three collections are fairly similar. Plants with the brighter yellow-green of H. cymbiformis, but with more terete and slender sub-erect leaves. The plants are quite robust and in cultivation reach about 80mm diam. with leaves up to 90mm long. Look-alikes are found in the gracilis, cooperi and cymbiformis var. transiens complex of the Baviaanspoort, and I have generally referred these all to H. cooperi var. gordoniana. However, that variety is actually quite a distinctive one from the Hankey Pass, north of Humansdorp and perhaps I should never have adopted it for general use in the way I have. Thus in my discussion about H. cooperi and H. bolusii var. blackbeardiana, I speculate that gracilis is an archetypal form which may lie at the root of the Eastern Cape species here being discussed.
The species H. aristata poses similar problem, and so does H. decipiens var. pringlei (Scott) Bayer as well as two collections which I and Bruyns made at Ripon station which is north-east of Kaboega. One of these latter collections is H. cooperi var. pilifera (-111) and the other is H. decipiens var. pringlei (-112). Largely because of that collection, I felt pressured into believing that the latter would best be coupled with H. bolusii var. blackbeardiana rather than with H. decipiens, and I was contemplating a major change of this kind. This would also have involved subsuming that element in an enlarged H. cooperi var. cooperi. There are, however, some other collections from the greater Darlington Dam (Lake Mentz) area to the west, which are relevant to this problem. These include older ones which suggested the link of pringlei with decipiens which I was thus also predicting, and new ones which confirm that this does in fact happen.
Because of the extent and complexity of the problem, this report deals specifically with the Kaboega area. In conjunction with it, a manuscript regarding H. cymbiformis var. incurvula, was written to give another indication of the nature and scope of the problem of classification of Haworthia. However, the chief reason for the visit to Kaboega was somewhat fortuitous. I was intending visiting Pluto’s Vale again, also the farm Thornkloof where Col. R. Bayliss had collected; the place Aalwynpoort to check on an Ernst van Jaarsveld collection and also hoping to cast light on a collection from the Bosberg at Somerset East. Peter Bruyns was hoping I would recollect a Stapelia aff. kougabergensis which he had seen on the Zuurberg, and there are also some other Haworthia records in that general area which need verification. What also materialised was a visit by Steven Hammer to South Africa, and contact with Tony Dold of the Schonland Herbarium at Grahamstown. The best of all was contact by my wife Daphne, with the gracious managers of the Kaboega farming enterprise, Sandy and Ian Ritchie. Through their kindness and hospitality we came to spend four days on the farm and briefly explored what it has to offer. One day there was spent with Steven Hammer, and with Tony Dold and Dez Weekes of Grahamstown. My wife and I returned for a second visit in Sept. ’99, when we also went to Oudekraal via a direct farm road from Kaboega to the east.
Results On our first visit we first explored the western side of the farm known as Wilgefontein. Tony and Dez went up the slopes of the Spekboomberg on the north side of the valley, and the remainder of the party climbed to near the top of the Zuurberg. We climbed straight to the grassveld where the grass was very long and thick. We soon found a solitary-growing greyish-green plant in flower (MBB6904-148), and then higher and on vertical rocks, a less translucent clump-forming plant with a velvety texture – also in flower (MBB6905-149). The plants looked rather different and we were quite excited about it being something apparently out of the ordinary. It is possible in the context of later collections, that these two collections are ecotypes. Looking at similar rocks about 200m to the south-east, we found what at first was obviously H. coarctata var. adelaidensis (MBB6907). This turned out to be a big population of plants which can, as such, be collectively regarded as intermediate between H. glauca and H. coarctata. This is a very significant collection because of the occurrence of glauca in its typical form at Zuurberg Pass, and nearer at Oudekraal, both to the east. Var. coarctata itself is not known nearer than at Patterson 70km eastwards and var. adelaidensis from east of Riebeek East which is still further away. (see also ‘Haworthia Revisited’ p179). What we did hope to see was H. angustifolia known at its western limit from Oudekraal. It must be on Kaboega and we just have not seen it yet.
Across the valley Dez Weekes had collected three specimens from a south facing steep cliff (MBB6903-150). These had long stems with bright green terminal rosettes and I have identified these as H. cymbiformis (with reservations! as I think this could again be a local ecotypic adaptation) without seeing the plants in habitat myself. This species is also not known from nearer than Hell’s Gate 50km to the south, but – it almost certainly has affinities with the plants collected by Marx and Bruyns. Steven, Tony and Dez had to leave after the first day, but Daphne and I continued the exploration the second day with a long climb up the hill east of DePlaat. We soon found Gerhard Marx’ (MBB6909-151) plants at the base of the mountain and continued eastwards and upwards. We came across a very extensive and dense mass of H. glauca (MBB6908). The plants were variously tubercled and lacked the distinctive grey colouration of the species. Any affinity with “coarcata” was less obvious than at Wilgefontein. Another interesting plant there was a dwarf form of Aloe tenuior. We crossed over to the steep north slopes and on the way down came across three plants (MBB6910-152) of what appeared to be similar to the velvety plant of the previous day (-149). It was in the same rocky situation. We looked further for it, but failed to find it again probably only for lack of concerted effort. The terrain was very difficult and we were getting a little hot and tired. We came across the Marx plants again. These were further down the hill and looking rather bleached in the sun.
149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.
On the way home we were travelling across some very stony ground covered with scattered bush and a low-growing Felicia, probably filifolia. Underneath these, in algae and moss, and with Euphorbia stellata and Tylecodon reticulatus were single plants of a cryptic small blue-green species (MBB6917-142). This I relate to collections from Stonefountain and Verdun cited in my revision under H. aristata, again collected as MBB6852-126 and Dold in MBB6851-125 (Modderfontein).
142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
The following day, accompanied by Sandy Ritchie, we ventured into the Kaboegapoort itself. We walked to the boundary with the Addo National Park. On the way we had seen inaccessible clumps of an Haworthia on a very steep cliff and we tried to reach these on the way back. We were lucky to be able to sample four clumps on the first of the rock faces where some plants had established themselves off the face (MBB6911-153). On the other cliff we could not reach anything. The plants were not cymbiformis but relate rather to the Marx plants except that they were clump-forming and bleached. A better comparison is with (-148). They were also in flower.
148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.
Peter Bruyns collection (-147) is of six clones from this poort and each of these plants is different. However, one clone (-147.1) resembles gracilis var. viridis from Perdepoort (-70) and also resembles a collection of Bruyns of cymbiformis var. transiens (-69) from the Langkloof. (This latter collection is involved in the issue of that species or gracilis var. picturata). Kobus Venter also has a collection of similar plants from south of Lake Mentz (-72.2). A second clone (-147.2) was an aberrant plant with very terete, abruptly mucronate leaves which were also unusually translucent. Another clone (-147.3) was of a plant identical to the big gracilis var. isabellae of the Krom River Estuary (Ripon, WRB1, recollected by myself -39), and comparable with a collection (MBB6855-154) from Waterford, east of Jansenville. That collection could be identifiable as H. arachnoidea var. xiphiophylla and perhaps hinting at a link of that element with H. decipiens var. minor. Two clones (147.4, 147,5) are the same as the DePlaat collections (-145, -146), resembling the Gladhurst (-116) and Glen Craig (-98,-99) forms of gracilis. Thus representing my view of an archetype.
147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.
On our last day, Ian Ritchie kindly took us on a drive to territory which had looked quite visitor friendly from the top of the Zuurberg. On closer contact they are anything but so. On the Spitzkop, which is on the northern boundary of the farm, we found aristata (-141) growing on bluish shale in a situation favoured by H. cooperi var. cooperi. It was a little bigger than our previous collection, but smaller than plants we subsequently collected northwards on the road between Riebeek East and Jansenville (Paddafontain, MBB6899-155), which I refer to pringlei). Driving to the east of Spekboomberg we saw plants (MBB6914-156) similar to the Marx plants of DePlaat (-145) in profusion, some of them without the softer translucence of the Marx collection. The leaves were occasionally much flatter and ovate and distinctly reminiscent of cymbiformis. Daphne and I walked down the hill from that point seeing the plants for most of the descent. Ian in the meantime drove back and further to the south-east and then found very similar plants on a steep slope also facing south-east (MBB6915-157). These plants can also occur in large clusters.
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.
Daphne and I drove to Somerset East to reconnoitre the Bosberg and on the way back saw aristata again about 10km north-east of Kaboega (-143). We had looked at the Paddafontein (-155) plants on the way out and they quite obviously can be linked to aristata too. Unlike the other collections of aristata, however, the Paddafontein plants have a large robust inflorescence with many flowers open (usual in decipiens) as opposed to say the Commadagga (-128), aristata, but with fewer and stumpier leaves) dwarfs with only 8-10 flowers per stalk and one open at a time (thus more pilifera-like).
128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.
During our second visit, we again went to the top of the mountain at Wilgefontein, after Ian had shown us H. glauca in the Kaboegapoort itself. This population was not typical of the species and also more like the De Plaat plants. We revisited the site of -148 and -149. Both were in flower and on this occasion we found the grassy ecotype within about 75m of the stone-face plants. The latter had flowered but seed-set was very poor, as opposed to the grass element which had well-formed capsules. From there we went to the Dez Weekes’ slopes via a different route and collected (MBB6925-158) plants ranging from the same greenish cymbiformis-like plants of -156 to specimens which could be nothing else but typical of true cymbiformis. We saw the same plants again at the dam to the north-west of the homestead, and again on the south slopes (-159) behind the previous De Plaat collection (-151). We completed the stay with a visit to Soutkloof where we saw again the true aristata (-139), and also to the office of the Addo National Park on top of the Zuurberg. Here we saw specimens of H. cooperi var. pilifera from that vicinity, and similar to a collection by Ernst Van Jaarsveld from Oudekraal (I had seen these plants when I collected H. angustifolia var. baylissii there many years ago). On our visit to Oudekraal we stopped to the west of my previous visit and probably also west of where Ernst had collected. We found the cooperi-like plants (MBB6922-160) again growing among rocks in dense grassland. The plants had very pronounced reddish-lines in the leaves and this was evident in all the Kaboega collections.
158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.
Curiously a post-graduate botany student busy with a study of succulent endemism brought in a number of plants for identification. This is P. Desmett, and among his collections, two are relevant. One is PD2310-161 from Boplaas. This is north-east of Kirkwood where the Kaboega meets the Sundays River. The plant is a small spinose specimen which could relate to the arachnoidea-like (as von Poellnitz compared it) stiemiei. It could alternatively, and because of its colour and translucent patterning of the leaves, be more probably compared with H. decipiens var. minor represented by several MBB collections from Sapkamma (MBB6618-162, MBB6619-163, MBB6620-164) to the west. The other is PD2309-165 from the southern end of Kaboegapoort. It is the apparently puberulous-like element resembling -148, and also -153. Kobus Venter also collected and aristata-like plant from the Sundays River Poort (-73) which I think compares very favourably indeed with a large number of collections from afar afield as Redcliffe (north-east of Willowmore), the Baviaanskloof, Uniondale, down the Longkloof to Humansdorp and Hankey/Patensie. These are all collections which I have identified as H. cooperi var. gordoniana, and considered in the context of another paper.
161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
Discussion It is apparent to me that there are can be only two elements (species) of the sub-genus Haworthia present on Kaboega. These are from either of the geographical elements cymbiformis, aristata, gracilis and cooperi, and they are directly continuous. In cultivation it is apparent and obvious to me, that aristata from Spitzkop (-141) is mirrored by the gracilis-like -152, which is continuous with the more gracilis-like -151. This latter element leads through several collections to the cymbiformis-like plants in -150, -158 and -159. Similarly a very cooperi-like element in -148 is the apparent ecotype of the very gracilis-like -149. But -148 (and -165) must also be compared with -152 and to -153, which take us back to the gracilis-like archetypes. More significantly these seem to be the elements which best relate to the collections from Oudekraal, and with what occurs still further east at Zuurberg. These collections are considered to be H. cooperi var. pilifera.
There is no doubt that the Spitzkop aristata (-141) must be compared with -155 at Paddafontein and thus connecting aristata to the greater Jansenville area, and to the western elements of H. decipiens var. pringlei. There is the Waterford collection north of Lake Mentz (MBB6855-154) which is problematic as it does not have the opacity nor darker blue-green colouration of pringlei. It is better compared with xiphiophylla (or perhaps this is H. decipiens var. minor) in 72.2 south of Lake Mentz. It also bears a remarkable likeness to the Krom River collection of gracilis var. isabelllae (-39) as indicated above.
Conclusions I conclude that at Kaboega we have a situation where cooperi is excluded by the fact that the archetypal gracilis is represented by an advanced version of gracilis from which aristata and cymbiformis are extended. This pattern of identifications and classification true for one area, are not true for another. Already fully aware of the complex interaction between species like H. bolusii, H. cooperi and H. decipiens, and fast becoming even more aware of the extension of this complexity to H. gracilis, H. cymbiformis, H. arachnoidea var. xiphiophylla and even H. marumiana, I have to express conclusions very guardedly. Any classification of Haworthia will undoubtedly have tensions within it. It has been long apparent to me that sophisticated technology is unlikely to prove of much value in dealing with the nuance of variation between populations. If it is, it has going to have to first take into account the kind of variation one sees at the scale covered by this article. My belief was, and is now confirmed, that this is indeed the scale at which observation is now required. It can still be a lot closer. We did not spend sufficient time at Kaboega to explore the area thoroughly, and neither have we yet made any permanent record of our observations other than this report and accompanying illustrations. The point may now have been passed at which casual generalisation from a memory bank of images is possible. Extensive photographic and herbarium records are going to be essential to create a physical record which can be studied and manipulated. There is a series of eight mountain ranges from near the coast, with the Zuurbergs being the last of these in the north-east. If I calculate how long it would take to explore that area on the scale of our limited survey of Kaboega, I reckon on at least three years of continual search.
At this point I realise that the expectations of “Haworthia Revisited” are not going to be met. There are already snivels and meuls because there is no “data” in my revision. My experience tells me that this is not because the average reader would in fact take any cognisance of such data – but it is part of the illogical and faulty paradigm of modern “science” (“materialism”, the Theosophist would say). My conviction is that I have in my revision presented there a very comprehensive picture of the genus. This can definitely meet the time-worn wishful thought of the platitudinous foreword that “this book will stimulate/encourage/direct/guide further research”. Classification of Haworthia is not simply sorting a few single specimens as they are represented in collections or on herbarium sheets. It is trying to understand a complex system of closely interrelated and similar looking elements, as populations, which do not fit a classic and static image of a genus and so-many discrete species. My contention is that this is not only the case for Haworthia, but for many other genera too. ♦
141 a. MBB6916 H. aristata, Spitzko[, Kaboega.141 b. MBB6916 H. aristata, Spitzko[, Kaboega.141 c. MBB6916 H. aristata, Spitzko[, Kaboega.142 a. MBB6917 H. aristata, Kaboega.142 b. MBB6917 H. aristata, Kaboega.142 c. MBB6917 H. aristata, Kaboega.
143 a. MBB6901 H. aristata, Hopewell.143 b. MBB6901 H. aristata, Hopewell.144 a. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. arista, Chalumna. (H. cooperi var. doldii – ed.)144 b. T.Dold3961 in MBB6921 H. cooperi var. leightonii to H. aristata, Chalumna. (now H. cooperi var. doldii – ed.)145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 b. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 a. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.145 d. J.G.Marx in JDV91/14 H. cooperi var. cooperi, De Plaat, north aspect, Zuurberg.146. J.G.Marx in JDV91/15 H. cooperi var. cooperi, De Plaat, south aspect, Zuurberg.147 a. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 b. PVB5002 in JDV92/33 H. cooperi var. cooperi, Kaboegapoort cf [51 a-c].147 c. PVB5002 in JDV92/33 aberrant.147 d. PVB5002 in JDV92/33 cf [20, 154 a-c].147 e. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].147 f. PVB5002 in JDV92/33 cf.095 [96 a, b, 116a-e].148 a. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 b. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 c. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 d. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.148 e. MBB6904 H. cooperi var. cooperi, in amongst grass, Wilgefontein, west of Kaboega.149 a. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 b. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 c. MBB6905 H. gracilis var. isabellae, on rock face, Wilgefontein, westof Kaboega.149 d. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 e. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 f. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.149 g. MBB6905 H. gracilis var. isabellae, Wilgefontein, west of Kaboega.150. MBB6903 H. cymbiformis var. cymbiformis. south-facing rocks, Wilgefontein.151. MBB6909 H. cooperi var. cooperi, southwest fo De Plaat farmhouse.152 a. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.152 b. MBB6910 H. gracilis var. isabellae to H. cooperi var. cooperi, east of De Plaat farmhouse.153. MBB6911 H. gracilis var isabellae to H. cooperi var. cooperi, rock faces in Kaboegapoort.154 a. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 b. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.154 c. MBB6855 H. arachnoidea var. xiphiophylla to H. decpiens var. minor, Waterford, east of Janesville.155. MBB6899 H. aristata / H. decipiens var. pringlei, Paddafontein, north of Kaboega.156 a. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 b. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.156 c. MBB6914 H. cooperi var. cooperi / H. cymbiformis, east of Spekboomberg, northwest of Kaboega.157 a. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.157 b. MBB6915 H. cooperi var. cooperi / H. cymbiformis, Kok-se-dam, Kaboega.158 a. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 b. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 c. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 d. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.158 e. MBB6925 H. cymbiformis var. cymbiformis, Kok-se-pad.159 a. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.159 b. MBB6935 H. cooperi var. cooperi / H. cymbiformis, south of De Plaat farmhouse.160 a. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 b. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.160 c. MBB6922 H. cooperi var. cooperi. Oudekraal, Zuurberg.161. PVB2310 H. decipiens var. major, Boplas, northeast of Kirkwood.162 a. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.162 b. MBB6618 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma, south of Perdepoort.163 a. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.163 b. MBB6619 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 a. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 b. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 c. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.164 d. MBB6620 H. decipiens var. minor / H. arachnoidea var. xiphiophylla, Sapkamma.165. PD2309 H. gracilis var. isabellae to H. cooperi var. cooperi, southern end of Kaboegapoort.
M. B. Bayer, Cape Town. Ian Ritchie, Somerset East. ♦
This problem of continuity is one I seem to have difficulty in conveying to my readers and listeners. The difference between one species and another is a discontinuity and, if we believe in evolution, it is the resultant of a break-up of continuity in its ancestral parent species. The “model” we have in our minds, is of progressive change from one recognisable entity to another by evolution. Geographic distribution and re-distribution are key elements in this process. But we do not seem accept this in the way we try to classify plants or interpret classifications. Apart from recognising that change could be gradual and therefore manifest continuity, the change may be from a complex variable system which contains different levels of continuity within itself, and not from a simply understood uniform ‘ancestor’.
The result is that in a genus like Haworthia, which is by no means exceptional, the differences between species i.e. the discontinuities between “species”, may be very difficult to either recognise or rationalise. It in fact becomes a statistical operation in which all the characters should be involved i.e. multiple variate analysis. If all the characters could be measured and quantified it is statistically possible to subject all the data so obtained by one of several statistical methods to measure “distance” and “significant difference” between groups of plants which we want to ascertain are species, varieties or even just hybrids. The process of “cladistics” is the use of a system to generate a branching “tree” of relationships base on characters which are also evaluated and loaded for chronological priority (primitive versus advanced). In using such a mathematical package, it is pretended that the classification becomes “objective” and hence replicable to satisfy the scientific requirement. In my estimation, the cladistic process assumes that a two-dimensional “tree” adequately represents the spatial and temporal changes of evolutionary processes, and it does not work.
Somebody might one day try to apply such methods to Haworthia and I say “Good luck to you”. My experience of characterisation and variation in the biological systems I have experience of, and including Haworthia, suggest to me that sensible, practical, experienced “eye-balling” will prove the better bet. Ultimately in Haworthia, I expect that technology and cladistic methods will be testable on the result of my classification. This is not a conceited and arrogant claim. It is a simple reflection on what classification actually is and what it is for. Much of botanical classification has been done by amateurs with no, or minimal, specific training and qualification for that field at all e.g. G.W. Reynolds, L.C. Leach, T.L. Salter, J. Lavranos, C.L. Scott, G.G. Smith, M.B. Bayer etc. Their classifications form the basis of many scientific observations, sometimes by scientists who have no conception of the significance, or insignificance of the names they use or what they may actually mean. The classifications may have little to recommend them except the fact that they appear to conform to the approved nomenclatural style.
Putting that all aside I want to just demonstrate by means of a series of illustrations, what occurs in Haworthia. I have described several situations before but what follows is another account of the continuity which occurs in the southern districts of Somerset East, Eastern Cape. The main area concerned is Kaboega at the north and western Zuurberg near where it fades away north of the Klein Winterhoek Mountains (see map Fig.1). One thing should be engraved on the mind. The following comparisons are made on field impressions and on multiple plant samples. This at least takes note of statistical requirements and considerations and it would be wrong to try and use the individual illustrations to arrive at some other conclusion.
Fig. 1. Map of the western Zuurberg, Kaboega Farms.
Determining a starting point is quite difficult because there simply is none unless a second problem is addressed. This is the one of typification in which names are fixed to a specimen of some kind. After this is done there is the question of how the name is applied to the body of the plants from which it comes. In this article, the question of the application of the names cooperi, pilifera, vittata and blackbeardiana is involved and so I reproduce the illustrations accompanying the origins of these names (Figs.2, 3, 4 & 5). It is my experience that in the field, it is possible to find resemblances to these illustrations in a large number of geographically disparate populations, and hence quite a problem to apply them in a consistent and sensible way. Only the name blackbeardiana can unequivocally be said to have been derived from any particular field population and this is at Bowe’s Farm near Imvani, south of Queenstown, and the single illustration by no means represents all the variants of the many populations which can be said to belong together here. Some of those variants invoke the use of the other names. How all these names are used is something of a personal choice and mine is laid out in the book Haworthia Revisited which I wrote in 1999.
Fig. 2. H. cooperi Baker. Refugium Botanicum, 1871.Fig. 3. H. pilifera Baker. Refugium, Botanicum 1871.Fig. 4. H. vittata Baker. Refugium Botanicum, 1871.Fig. 5. H. Blackbeardiana VPoelln.,1937.
In this chapter I will take a collection (originally by G. Marx) from east of Alicedale and call it H. cooperi var. cooperi ([1]…denoting position on map Fig.6. MBB6845). In doing so I will have to point out again that there are further intrinsic difficulties in the requirements of nomenclature, which is that there is such an entity as a “typical” variety. My belief is that any plant which is not directly assignable to a variety of a species should simply be referred to as that species. Thus the Alicedale collection is H. cooperi var. cooperi and geographically in the distribution area of H. cooperi var. pilifera. The plant illustrated is of a single clone, but the plants in this particular population relate better to populations in the geographical area of the typical var. cooperi. The plants occurs here together with H. angustifolia and H. cymbiformis (in the same indeterminate varietal format as the H. cooperi, but for geographical reasons mainly, this population is referred to as H. cymbiformis var. obtusa. A similar form of H. cooperi depicted recurs on the Zuurberg and I saw it at Oudekraal when I explored there for H. angustifolia var. baylisii ca 1982. I have plants in cultivation from there which were collected by Ernst van Jaarsveld, and I made a new collection MBB6922 slightly west of there in 1999. I saw it again at Vyeboomfontein ([2]Fig.7 MBB6949) much further west and again just north of that at Klipfontein [2] MBB7049. I am unsure if the identification should be var. cooperi or var. pilifera but choose the latter.
Fig. 6. MBB6845 H. cooperi var cooperi. E Alicedale.Fig. 7. MBB6949 H. cooperi var pilifera. Vyeboomfontein.
Slightly further west at DePlaat, the problem starts with plants which are greener (particularly on the south slopes) and with longer more attenuate leaves. There is first the collection from north slopes ([3]Fig.8 JDV91/14) that I attribute to H. cooperi var. gracilis, and then mine from the south slopes ([4]Fig.9 MBB6935). Near the Kaboega farmhouse at Koksedam there is also a greenish element and two plants are illustrated ([5,6]Figs.10a & b MBB6915) to show that it is partly cooperoid and partly cymbiformoid. I propose calling these, with MBB6935, H. cooperi var. viridis because it is complicated (it could invoke the name gracilis of von Poellnitz, but I described a smaller green variant from near west as var. viridis, and this name seems more appropriate). I have, in Haworthia Update Vol.1, described that there is this greenish archetypal form which is quite widely spread and often in an indeterminate zone, between forms of H. cooperi and H. cymbiformis. On Kaboega these plants take three elemental directions:
Fig. 8. JDV-14 H. cooperi var gracilis. DePlaat.Fig. 9. MBB6915 H. cooperi var viridis. De Plaat.Fig. 10a. MBB6915 H. cooperi var viridis. KokseDam.Fig. 10b. MBB6915 H. cooperi var viridis. KokseDam.
1. Directly west to Spekboomberg ([7]Fig.11 MBB6914) and then to Wilgerfontein ([8,9]Fig.12a & b MBB6925) at. At Fig.11 the plants are consistent with Fig.9. At Fig.12a & b the plants resemble H. cymbiformis. Some clones are more similar to that species than others while odd clones are entirely indistinguishable from it. I find it difficult to determine what species it is, never mind add a varietal name, because of its obvious intergradation with H. cooperi variants and its geographical disassociation with H. cymbiformis. My suggestion is that this is where the case calls for the use of the species name, without the addition of the typical variety epithet. But even that is unsatisfactory here because I think the plants, despite their outside similarity with cymbiformis, are actually in the same genetic bag as cooperi var. viridis. It is worth stating that there is a transition from sandstones at DePlaat and eastwards, to Dwyka tillite westwards to Wilgerfontein.
Fig. 11. MBB6914 H. cooperi var viridis. Spekboomberg.Fig. 12a. MBB6925a. H. cooper var viridis=cymbiformis. Wilgerfontein N.Fig. 12b. MBB6925 H. cooperi var viridis=cymbiformis. Wilgerfontein N.
2. Slightly to the south-west, entering the Kaboega gorge is JDV92/33 ([10]Fig.13). This is indeterminate and I would label it “graciloid”, although there is evidence of other variants in the gorge which do not agree with the concept of var. gracilis. That name itself presents problems as described in Update Vol.1. Fig.14 is in sandstones, and on the steep north facing slopes (also sandstone) of Wilgerfontein a little to the west and south of MBB6925, is MBB6904 ([11]Fig.14). I would suggest that this resembles H. cooperi var. gordoniana (subsequent to drafting this manuscript I did a limited exploration south of the Zuurberg, and found a fairly similar populations occurs at [=11] in the Addo National Park. But it is an astonishing ecotypic variant too, as 25 meters away is MBB6905 ([12]Fig.15) which is “isabelloid“. I have coined the name “puberula” for it. Bear in mind that these pictures represent only samples of many plants in small discrete habitats.
Fig. 13. JDV93-33 H. cooperi ‘graciloid’. Kaboega Gorge.Fig. 14. MBB6904 H. cooperi ‘graciloid’. Wilgerfontein S.Fig. 15. MBB6905 H. cooperi ‘puberula’. Wilgerfontein S.
3. In a north-westerly direction from DePlaat is MBB6943([13]Fig.16), which is slightly spinose and indeterminate like the MBB6915 (Fig.10) collection. But it gets very curious. If we ascend the DePlaat hillside towards Klipfontein and [2]MBB7049, we find first MBB6910 ([14]Fig.18) almost indistinguishable from MBB6905 and close to MBB6904. A little further, and less than 250meters from MBB7049 is MBB7017 ([15]Fig.19), a small graciloid=puberula population in which the plants are very small and the leaves have surface spination. If we cross the valley and go now north of Klipfontein to the upper slopes, we find MBB6940 ([16]Fig.19). It is indistinguishable from MBB6904! If instead we stick to the low lying areas and move north of Koksedam, and northwest of DePlaat, there is first MBB6942 ([17]Fig.20) and then MBB6917 ([18]Fig.21). This latter is a relief because I have several collections from widely dispersed points [=18] to suggest these are my version of H. aristata. Not forgetting that the initialisation of that concept at Soutkloof near Addo, also involves H. cooperi. If we look at one collection from Commadagga ([19]Fig.22 MBB6897) we see a plant which is a variant of H. aristata (it could be cooperi var. pilifera!) that has look-alikes at Kirkwood in the arena of H. cooperi var. pilifera. But at Kaboega, the plot thickens further. Going still northwest from DePlaat and now north of MBB6943, is MBB6916 ([20]Fig.23). There is a problem again, and one has to go quite a way (about 20km) to find MBB6899 ([=20]Fig.24). The latter is quite a large plant in the mould of H. aristata. To find a better home for it among several other collections, including some dubious ones to the east and north at Ripon, and including Col Scott’s H. pringlei, I added it there while reducing Scott’s species to H. bolusii var. pringlei. This problem is elucidated elsewhere but I have not decided where best to place MBB6916 I think for practical and geographic reasons they could be placed under H. aristata, but they could equally be put under H. bolusii var. pringlei. I described H. decipiens var. virella to lessen tension on my use of the name pringlei and its relation to H. bolusii var. blackbeardiana and to H. cooperi var. dielsiana (includes Scott’s H. joeyi). While it does so in some areas of the classification, some tension does remain.
Fig. 16. MBB6943 H. cooperi var viridis. Dassieklip.Fig. 17. MBB6910 H. cooperi ‘puberula’. DePlaat.Fig. 18. MBB7017 H. cooperi ‘graciloid=puberula’. Klipfontein.Fig. 19. MBB6940 H. cooperi ‘graciloid’. Klipfontein N.Fig. 20. MBB6942 H. cooperi’graciloid=aristata’. DePlaat Weir.Fig. 20. MBB6942 H. cooperi’graciloid=aristata’. DePlaat Weir.Fig. 21. MBB6917 H. aristata. Kaboega.Fig. 22. MBB6987 H. aristata=cooperi var pilifera. Commadagga.Fig. 23. MBB6916 H. bolusii var pringlei=aristata. Spitzkop, Kaboega.Fig. 24. MBB6899 H. aristata. Paddafontein.
This link to the above variants is enhanced by MBB7012 ([21]Fig.25) from Buffelsnek, overlooking Lake Mentz and west of Wilgerfontein. This collection is also labelled H. cooperi ‘graciloid’ as I have done for MBB6904, but it could just as well be H. aristata or even bolusii var. pringlei (because of the nature of continuity offered by other populations to the northwest [=21]. To show just how complex the situation is, I illustrate JDV92/44 (Fig.26). This was initially a P.V. Bruyns collection from Inverbolo, far distant and northeast of Cathcart. I specifically went there to confirm (MBB6942) that this is H. bolusii var. blackbeardiana. I have said elsewhere that there is a problem of this question of continuity and that H. decipiens can be indistinguishable from H. bolusii var. blackbeardiana. To add insult to injury I illustrate MBB6730 (Fig.27). No! It is not H. aristata, it is H. mucronata var. habdomadis from Seweweekspoort in the Little Karoo, and from an entirely different set of populations. It is in fact extremely disconcerting to have to admit that the green plants of Figs. 9 or 10 can similarly shown to be indistinguishable from H. mucronata var. morrisiae, also from the Little Karoo.
Fig. 25. MBB7012 H. cooperi ;graciloid’. Buffelsnek, W Kaboega.Fig. 26. JDV92-44 H. bolusii var blackbeardiana. Bolokei, NE Stutterheim.Fig. 27. MBB6730 H. mucronata var habdomadis. Seweweekspoort, E Ladismith.
In writing this, I hope that readers will now get to understand that in apparently attacking the work of aspirant Haworthia taxonomists, I am not trying to defend myself or my system. I simply recognise that these persons do not have the competences or the insight to follow and understand the difficult choices that have to be made any better than myself. The main point is, that I know that if we explore still further west of Kaboega (and there are collections which prove this), and then into the Klein Winterhoek mountains to the south-west, still further problems are going to unravel (us) as we roller-coaster between H. decipiens and H. cooperi all the way to Uniondale and beyond. These same problems bedevil most of the species systems in the southern and western Cape. Unless one understands and respects the intricate geographic relationships, the use of the names in the way I recognise them will perhaps not make sense and I truly see no other way in which a nomenclatural system and classification, which will work in the field, can be derived for Haworthia. ♦
This note is not strictly after closure because Cameron MacMaster (Cameron knows the plants, especially the bulbs, of the E Cape intimately and was instrumental in the re-location of H. marumiana many years ago.) sent me a picture (Fig.1) of a Haworthia from Glen Avon Falls east of Somerset East some time ago and this has been a lure to me ever since I saw vdW287(PRE). It should be noted that this specimen is cited, I must note a sentiment of considerable reservation which was not conveyed by the rigidity of print, in Haworthia Revisited (p.67) under H. decipiens var. minor… “3225 (Somerset East): in valley behind Bosberg (-DA), van der Westhuizen 287 (PRE).” I have visited the Bosberg in a weak attempt to locate such a plant after a fruitless attempt to determine who and where the collector was and is. The area is intimidating in its vastness as are so many of the hills and mountains of the Cape and with so much still to explore, this area has not been a priority. In fact I have just recognized that while I wrote Revisited in response to pressure, my subsequent exploration has been to seek validation for my own comfort rather than to try and impress anyone. This recent visit to the Bosberg is only because an odd opportunity arose for me to revisit my friends (Ian and Sandi Ritchie) on Kaboega, coupled with interest from a distant botanist acquaintance in Prof. Richard Cowling. Prof. Cowling is one of those rare botanists from whom I have really learned something to think about rather than just to remember. I had contacted him because in my correspondence with Jan Vlok about the vegetation of the Mossel Bay area, he had copied responses to Prof. Cowling. The outcome was that I was introduced to Dr. Syd Ramdhani who is now contracted under Cowling to study the biogeography of Bulbine as a post-doctoral task. Dr. Ramdhani studied Kniphofia and works in the molecular-biology laboratory of Rhodes University managed by Dr. Nigel Barker. Dr. Ramdhani is now also tasked and occupied with a feasibility study of Haworthia as a target group for extended biogeographical research where H. cooperi has been suggested by me as a possible fruitful area of interest. (These botanists have been warned not to be influenced by Bayer!) So I have been aware that the MacMaster plant could signify a replicate of the Kaboega/Helspoort/Plutos Vale/Baviaanskloof/ complexes which suggest that H. cooperi and H. cymbiformis may be one species. My visit to Glen Avon Falls was then added to the familiarization of Dr. Ramdhani with Haworthia on Kaboega.
With the kind assistance of Bill and Allison Brown of Glen Avon farm and then Bill Playdon of Glen Craig, we gained access to the waterfall which is now a National Heritage site. The road cannot be described as suitable for anything but a rugged 4X4 but we took it on in our less suited vehicle to save time (cost = 1 tyre!). We found just 5 plants and photographed three (figs. 2, 3 & 4). The terrain is really rugged and I have no doubt that the plants will occur in greater abundance nearby and also in the greater area of the Bosberg – and that such populations will follow the same pattern now so familiar to me and so seemingly strange to so many haworthiophile writers of my acquaintance. The plants call to mind an article that I wrote in conjunction with John Pilbeam (Cact. Succ. J (US) 46:166, 1974) where we ironically considered H. obtusa to be a variant of H. cymbiformis rather than of H. cooperi. The latter species is everywhere about the Bosberg while H. cymbiformis is to my knowledge far further to the south and east at Alicedale.
Returning to Kaboega, we took a brief look at some of the populations recorded and discussed in Haworthia Update Vol.1. Dr. Ramdhani left us there and we continued our holiday. We had been intrigued by Ian Ritchie’s discovery of a second population of H. sordida (figs. 5, 6 & 7) and while exploring that took a different route back to the vehicle. On the way we found another population of H. cooperi cf cymbiformis (I will be accused of inconsistency here because I have referred to these variants as graciloid, cf viridis, cymbiformoid if not also by some other name. Figs, 8, 9 & 10). I am not going to try and discuss or describe these plants any further than already done in the literature and because the following considerations come to mind.
A botanist acquaintance is most scornful of my opinions and my solutions for Haworthia and wrote to me… ”I believe taxonomy is an explicit, precise discipline governed by certain rules, if they are obeyed, things will fall into place. If they are ignored, you land up in a mess.” This botanist who might well have been raised on Enid Blyton and the famous five, or Bryce Courtenay and Aloe brevifolia in the Barberton hills from Power of One or the BIG tree in The Four Fires, when it comes to secrecy and drama, field botany and field familiarity in particular. The ICBN is no doubt hot-stuff where nomenclature is concerned but pretty damned useless when it comes to what these are used for. If things are going to fall into place it will be because we all make a more concerted effort towards mutual understanding and common purpose.
Secrecy in respect of localities I do sympathize with. The question is if this serves any purpose. My opinion is that, quite apart from the fact that secrecy has no place in science, it does not have any merit. Where secrecy is practiced there can be no organized skepticism and no shared reality or truth. I see it used to obfuscate and obstruct the healthy organization of skepticism by others and possible alternate solutions. There may be a down-side in that unscrupulous individuals may decimate populations or remove valuable genetic material and diversity from the field. The reality is that this is happening all the time at far greater scale than that of a few collectors. If it were not for collectors our knowledge base would have been infinitely smaller and who knows what the negative impact of the dwindling numbers of taxonomists is going to be. My negative view of conservation is that it is a sentiment with its origins in collectors and that it is now functioning to discourage the very interest that gave birth to it.
On our travel home from Kaboega, I kept looking towards the hills and mountains that accompanied us all the way on a 12 hour drive to complete about 1200kilometers. All of those offer suitable habitat and perhaps 2% or a great deal less of the area has been explored with an eye for Haworthia. This brings me finally to a sort of postscript. Russel Scott writes of his encounter with Poellnitzia rubriflora in the Robertson District. What is odd and welcome too no doubt, is that he does not mention the fact that officially the name is Astrolobarubriflora. Over a 6-hour hike he has to suggest that the plants were relatively rare – where he walked of course. His hike could have covered more than 20 miles and perhaps a 20yard wide band – calculated at a total search area of about 2.2% of one sq. mile! It is quite common south of the Breede River between Eilandia and just east of Bonnievale and as Russel mentions seeing H. herbacea I conclude that he was thus within the 600sq mile area I would say is its range. Thus even a 6-hour hike along what must be conceded was a narrow band, has not provided a lot of insight into quite a large area. Very little indeed as to what all there is to be seen. Yet I do not think it is possible to be looking specifically in species and succulent-rich habitat in the said search area without seeing H. mirabilis. Russel has exposed a reality that for someone to really become familiar with any plant group in the field is a dedicated life-time. Only a select few South Africans seem to have achieved that and then only in respect of particular interests. In my life-time of plant interest and field experience, I think I am personally only beginning to have an idea of what it means to really know what field familiarity can be. ♦
Fig.1. Haworthia sp. Glen Avon Falls, Somerset East 22Jan06 101Fig.2a MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.2b MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.2c MBB7634.1 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.3a MBB7634.2 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.3b MBB7634.2 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.4a MBB7634.3 H. cooperi cf cymbiformis. Glen Avon Falls.Fig.4b MBB7634.3 H.cooperi cf cymbiformis. Glen Avon Falls.Fig.5 MBB7635 H. sordida. Swartkop, Kaboega.Fig.6 MBB7635 H. sordida. Swartkop, Kaboega.Fig.7 MBB7635 H. sordida. Swartkop, Kaboega.Fig.10 MBB7636 H. cooperi cf cymbiformis. N Swartkop, Kaboega.
Kaboega is a set of farms on the northeast of the Zuurberg Mountain range, north of Kirkwood and off the Addo National Park. I wrote about the haworthias that occur there in Haworthia Update Vol.1. There is also an article in Aloe 40:10 (2003) in which there is a discussion of the variation of those haworthias as related to geology and topography. My wife and I frequently visit Kaboega to renew relationships with Ian and Sandy Ritchie who live there. Each time we go we try to explore some different area. We generally end-up with something that is notably new.
Part 1.
There is a real problem in trying to reconcile the populations we see with the names that are available and the way in which I have tried to formalize them myself. The problem is that Kaboega seems to occupy some sort of central and neutral position and it is by no means easy to arrive at any clean rational classification. Three of my species are involved, and I have to say they are “mine” because other authors are in strong disagreement. The three species I see are H. cymbiformis, H. cooperi, and H. aristata. It is firstly necessary to explain that I interpret the name H. aristata in Haworthia Revisited quite differently from what I might have done earlier; and quite differently from other authors who have simply taken the easy route and associated the name with Little Karoo elements for which I use the name H. mucronata. My interpretation of the name will be quite evident from my writings and from the pictures submitted with this article. The use of the name H. cymbiformis with respect to Kaboega is a major problem for someone like myself who is firmly convinced that geographical relationships are foremost in the recognition of species as living systems. On Kaboega, plants that look like H. cymbiformis seem to proceed out of a complex that is surely H. cooperi. If one properly considers all the populations that I ascribe to H. aristata one is seriously confronted with the reality that it is also a geographic variant of H. cooperi.
During June 2008 we were again at Kaboega and found two further populations of ostensible H. aristata. One of these viz. MBB7698 (see figs 1a-d) is only about 1km east of Buffelsnek on the western boundary where I recorded MBB7012. We did also see the same plants on a ridge still further east and north of Wilgerfontein where the very green and proliferous cymbiformis-like plants seem to reach their western limit. MBB7698 is effectively on the top of the Spekboomberg ridge, whereas the cymbiformoids are a little to the west and on the south slopes. It would be really interesting to know what occurs further westwards towards Darlington Dam.
We found H. aristata again practically central to the farm and about 1,5km south east of the Weir (MBB7703, see figs 2a-c, and these plants may have been illustrated before elsewhere incorrectly as MBB7697, a Gasteria). This is still about 1km north of the east-facing riverine cliff that houses the very green cymbiformoid plants (MBB7636, see figs 3a-c) of which other populations have already been reported to the east at De Plaat (3 populations) and also to the west at Koks Dam and Spekboomberg.
I have probably argued for recognition of H. aristata as separate from H. cooperi. Such an entity can probably be recognized as occupying the lowlands to the north of the Zuurberg. Both H. cooperi and H. bolusii var. pringlei occur to the north at Ripon and it appears that the latter may be represented by a population northeast of Stonefountain. There is a problem with proper, as opposed to the formal way either Breuer or myself may have done typification of the name H. gracilis, where we have both apparently just guessed that the name applies to the Helspoort variants of H. cooperi. In fact von Poellnitz’ name H. gracilis rest on the citation of a really mixed bag from not just Hellspoort, but from five quite disparate locations across almost the entire Cape. I use the name ‘gracilis’ only in reference to Hellspoort and it is evident to me that H. aristata is in the H. cooperi continuum. Look at the pictures and see what you think. I have added three pictures (MBB7701, see Figs 4a-c) taken of plants above Klipfontein at the eastern end of Kaboega that I refer to as H. cooperi ‘puberula’ and related to another record MBB6908 at Wilgerfontein.
1a. MBB7698 H. aristata E Buffelsnek, Kaboega1b. MBB7698 H. aristata E Buffelsnek, Kaboega1c.1d.2a. MBB7703 H. aristata N Swartkop, Kaboega2b. MBB7703 H. aristata N Swartkop, Kaboega2c. MBB7703 H. aristata S Swartkop, Kaboega3a. MBB7636 H. cooperi cf. cymbiformis N Swartkop, Kaboega3b. MBB7636 H. cooperi cf. cymbiformis N Swartkop, Kaboega3c. MBB7636 H. cooperi cf. cymbiformis N Swartkop4a. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega4b. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega4c. MBB7101 H. cooperi ‘puberula’ S Klipfontein, Kaboega
Part 2.
Variants of H. cooperi on Kaboega
The various populations of H. cooperi are discussed or referred to in several of my essays published in the Haworthia Updates and elsewhere. I have also been privileged to go back to Kaboega several times and further explore there. Each visit generates something new and different and evokes thoughts of what is still unseen, unreported and unknown.
On this occasion Daphne and I were actually hoping to find and photograph perhaps a chameleon, as workers had reported seeing one abut six months previously in the vicinity of the Klipfontein guest house on Kaboega. This is in the northeastern end of the set of farms that now constitute Kaboega. We thought we should explore the upper slopes where there was more sunlight and basking opportunity for the chameleons in the cold winter sun.
We climbed up the mountain via a very overgrown route that served as a passage for stock to access the higher mountain slopes, but also serve as a donkey pack route for the transport of supplies over the mountain in earlier days. This is of course peripheral to plant exploration. Nevertheless it is so fascinating to dwell on the fact that the enlarged game farm is actually comprised of about seven smaller farms that once were home to farmers and family. All abandoned as circumstances have changed over the years. It is as though man invaded this rich treasure house of natural beauty, sucked it dry and abandoned it again leaving nature to now heal the wounds. This is very unlikely because of the game fences and the absence of predators. Rainfall is marginal and the Kudu population is huge. Nature will have to respond in some dramatic and drastic way if she has anything in mind, and she is perfectly capable of that.
We moved east of our original visit to the mountain top where we had seen and photographed a few scattered plants before. We know that if we find one plant, to ask “Now where are they?” This confinement to such discrete, small habitats is so characteristic that we laugh off the notion that to explain the vicarious distributions over great distance, the plants were wider spread at some distant time in the past. Something else is at work and these populations maintain pattern and coherence beyond material means.
Eventually Daphne, whose eyes are now better than mine, did find the first plant and the body of the population we had only touched before. So these are the pictures. The first three pictures are of MBB6901 H. aristata (See figs 5a-c) on the lower country north of the mountain about 3km away on a neighboring farm Doornkloof, the next one would be a cultivated plant of MBB7701 H. cooperi ‘puberula’ nom. nud. (See fig. 6) in sandstone and Fynbos vegetation in the mountain south of Klipfontein. The rest are H. cooperi (no appellation – MBB6940, See figs 7a-p) just east of my previously reported site north of Klipfontein, also in sandstone and transitional Fynbos. This term is used to simply imply one of these strange anomalies where karoid and Fynbos elements mingle. Many more pictures are really needed to indicate that each plant looks different and we spent a long and enjoyable time calling back and forth to come and observe one more gem.
Some of the clones would correspond to clones in the parent population of H. cooperi ‘viridis’ at Perdepoort about 20km west while others to H. cooperi ‘gracilis’ from Helspoort north of Grahamstown, 50km east. Thus to my mind there is no satisfactory solution to the naming of these plants in our conventional understanding of Latin names.
The last photograph is a view across the Klipfontein valley and over the old greater farmyard to a mountain ridge and over that to the Zuurberg Mountain itself. Between is the Vyeboomfontein farm. The black arrows mark a point on the near ridge where there is a Bushman painting site and a large quartz rock that is home to plants like MBB7101. That mountain ridge extends westward 1-2km to the abandoned DePlaat farmhouse where there are again populations of H. cooperi on northwest and south aspects, at different altitudes and with their own variants. More conventional H. cooperi ‘pilifera’ can be found on the far mountain while Kaboegapoort is a gorge through the mountain with still another set of populations and variants.
It is all a wondrous never ending drama.
5a. MBB6901 H. aristata Doornkloof5c. MBB6901 H. aristata Doornkloof5b. MBB6901 H. aristata Doornkloof6. MBB7701 H. cooperi ‘puberula’ S Klipfontein in cultivation.7a. MBB6940 H. cooperi N Klipfontein7b.7c.7d.7e.7f.7g.7h.7i.7j.7k.7l.7m.7o.7p.7q.
Part 3.
General comments on the plants on Kaboega.
The southern end of Africa is home to one of the six or seven plant kingdoms of the world. This is the so-called Cape Floral Kingdom. Botanists in the past seem to have misinterpreted this flora and not realized that it maintains its integrity for two reasons. The first is that it is on the table Mountain Sandstone formation and the second is that it is in a winter rainfall zone. Its origins are no doubt ancient, but in present day terms it is part of a present day winter rainfall area which includes the Succulent Karoo. Kaboega sits on the eastern verge of this biome and is therefore in a huge tension zone between inland, upland, summer and winter rainfall (and the stress of dryness) vegetation groupings.
Apart from annual rainfall stresses, broken topography, skeletal soils and an enormous range of exposed geological formations and rock series, contribute to an extraordinary range of varied habitats. Thus South Africa and especially the Cape, offer plant species huge opportunities to adapt and change according to ecological differences and stresses. Kaboega is a wonderful example of the way in which plant species have adapted to these different habitats.
Kaboega has plant species from all of the biomes in the Cape, and the vegetation ranges from upland grassy fynbos, renosterveld, succulent karoo, valley bushveld and countless variation of these. The main rocks are quartzitic, dwyka tillite (of glacial origin) and shales. In addition there is a small amount of alluvial terrace.
What is so dramatic is that the different species occupy sometimes very limited habitats and despite this tight habitat choice (requirement) it is very difficult to specify what those requirements are for any group of species.
The genus Haworthia is represented by two of its subgenera. The Subg. Hexangulares is represented by three species:-
a. H. glauca, which is present mostly on the upper quartzitic ridges where Euphorbia polygona and the asclepiad Huernia brevirostris occur. This form of H. glauca includes some evidence of apparent interaction with H. coarctata which comes from further east.
b. H. sordida is an isolated population south of the Zuurberg and then westwards to Steytlerville. It was known on Kaboega from only one small hill where it grew under renoster bush. It was only while researching this visit that a second group was found.
c. H. nigra, which is very widespread in the greater karoo, but rare this far south and so far it is only known from one small population on the river terrace gravels.
The second subgenus Haworthia is only represented by many varieties of but one species viz. H. cooperi and this recognition of one species is only arrived at by discarding the traditional view of plant taxonomy that has no definite and decisive definition for the word species. The Kaboega plants are known from about 30 populations. These vary dramatically, but continuously, according to habitat and this has to be considered in terms of continuing similarities from Kaboega outwards into the Karoo, the Western Cape and the Eastern Cape. This suggests that there are very complex species systems and species cannot be as easily identified and described by visible characters, as a loose definition has in the past allowed. The Kaboega populations are very different from each other, and could be said to be from four or more species if fancy rules. In fact they are probably adapted by habitat and are evidence of the continuity of them all in one species complex.
It is population structure which is so significant and if one considers all the plant species that occur on Kaboega, one comes to realize that they occupy often unique habitats. There is one level of generalists such as Acacia karoo, Olea africana, Aloe ferox, etc which seem to be ubiquitous. But actually most of the species can be seen to have a “home”. There are about seven species of Aloe on Kaboega and seldom do more than two share habitat. As one goes through a general list of names, one finds that one has to visit very specific places to find the plants. One can also find places like this one spot at Klipfontein that can be described as a “hotspot” for Kaboega. These are small species rich areas, which host species which do not occur generally and may in fact be very rare for Kaboega. By also occurring on Kaboega like this, they represent a genetic entity and resource which may be quite unique
Acknowledgement. I am most grateful to Ian and Sandra Ritchie for their generous hospitality and interest for the many times and hours we have spent fossicking in the wilderness of Kaboega. ♦
Kaboega is located in the Zuurberg Mountains north of Port Elizabeth in the Eastern Cape province of South Africa. Read more about this in Haworthia Update Volume 1, Chapter 5:- The Haworthias of Kaboega. There are a mind-boggling array of Haworthia populations here in an area considered to be the meeting point of several vegetation biomes. There is much exposed rock, and the soil is very skeletal, composed of three major groups: sandstone, mudstone, and glacial deposits. These pictures are of a Haworthia cooperi variant that occurs high up on sandstone. I went to this spot because researchers had sent me a picture of a cycad festooned with Haworthia. I did not get to the exact spot but have seen the way it forms hanging bundles in other situations.
Haworthia glauca!! can also be found here. On Kaboega these plants often have a very close resemblance to H. coarctata and it is no co-incidence that the distributions of these two species complement each other. An essential element of species recognition is their juxtaposition and if they occur in very close association or not. Darwin said as much.
I visited four populations of this greenish cooperi. One can find plants like this from east of Grahamstown right through to the Little Karoo. Here they are on Dwyka (glacial) skeletal soil.
These next are in the shales low down in the valley on Kaboega – I name it H. aristata. It is very common in the area but complements H. cooperi while there are populations that are neither. Populations cannot be treated in isolation and there is a distinct possibility/probability that I have been too generous with species. The attempts to find answers via DNA sequencing should make the vendors of that technology thoroughly ashamed.
More of these green things. I would guess that these would class as the simple progenitors of cymbiformis and cooperi. Perhaps even of mucronata?
This is Haworthiopsis sordida that does not occur, as far as I know, north of this. H. nigra also occurs here at it’s most southern at this longitude. Altogether it is quite a complex network of distribution patterns that relate to greater plant geography.
From another population as variants on a theme. have seen about 30 such just on this small mountain area and it just suggests what is still unseen on the length and breadth.
Not a great diagram but a way to appreciate the drammatic choreography of plant distribution and how it impacts on classification. Without it Haworthia names make no sense other than as imagined and fantasized. Cooperi and cymbiformis occur as intertwined species to the east and south. In the south they extend westwards to get lost in H. mucronata. Cymbiformis as an independent species does not enter Kaboega except as an observable variant of H. cooperi. The cooperi gets lost westwards as variants of H. decipiens. Perhaps close northwards as H. aristata. H. glauca does cross the Zuurberg but is here confused with H. coarctata that may occur in recognosable form on the eastern tip. Angustifolia is on the eastern end too but does not enter Kaboega. Neither do H. monticola or H. zantneriana from the west. This is also closely tied to the intrigue of winter vs summer rainfall and still further to the massive geological changes of the very recent.
I was musing over a whole lot of plants in a nursery all labelled cymbiformis and ignoring the formal classification and recognition of varieties. Gordon Rowley was very critical of my work because he felt that the lesser names were just being dumped. I agree that it is a problem and I was thinking of posting pictures that would facilitate the use of old existing names e.g. planifolia. I was very quickly disenchanted as I looked for pictures.
The field situation regarding cooperi and cymbiformis is as complex as the retusa/turgida/mirabilis/emelyae/pygmaea/mutica issue as well as the fact that each such species assemblage runs over in to still other species. I did google cymbiformis and planifolia to see what the internet had to offer and was amazed how poor and confusing that is. I welcome any questions that cooking up an answer for, may help rationalise and enlighten troubled minds.
The picture is from one population at Kaboega where it is evident to me that there is no distinction between cymbiformis and cooperi, and it gets worse.
There is a problem with the name obtusa as a variant of H. cymbiformis, and I think it, with the name translucence, that really covers the transition between cymbiformis and cooperi. Both those names obtusa and translucens can be applied to a large number of populations that belong in a twilight zone of neither “this nor that”. As I was pondering the other day, plant taxonomy is NOT good science. It is largely treated as a field for nomenclatural rules and recommendations and a stage for intellectual display. There is little connection between the minutae of nomenclature and the reality of what occurs in nature. These pictures are of plants in the Bosberg NE Somerset East and are in a geographical cooperi environment far from the range of cymbiformis. They could fit the concept of “obtusa” as so many other cooperoids do.
Let me add – cymbiformis are those green things that grow as clump formers on steep (rock faces, cooperi are those bluish plants that grow solitarily on the flat. They are the same!!! The names are just at the extremities and the more we know the more names we need??
When I posted those two items about cooper and cymbiformis, I had left gracilis somewhere in the closet. It is just a very difficult pill to swallow that all these amazing variants have (?) to be lumped under one species.
I had another look at the 3 DNA phyllograms in the work by Manning et al, and it can be noted that cooperi, cymbiformis and decipiens all see to pan out in the same lineage. So as I said it is far worse than imagined – although there are some improbables in the various lineages of the phyllograms. But this is just as far as sequencing has got and we have yet to see results from “next generation sequencing”. Here are two images of cooperi from different populations west of Uniondale pass that shocked even myself in saying they are the same species.
Many years ago you gave me a challenge to sort out the cymbiformis cooperi and so off I went to the Eastern Cape; many times. I didn’t understand then how plastic, how variable, the plants were and how rigid the names and descriptions. Some I couldn’t type.
Cymbiformis – Rosette to 130mm φ, partially stemmed, proliferous. Leaves broad ovate to lanceolate, flat to slightly concave, generally <1/3 as thick as wide, usually opaque, green turning yellowish to pink hued on exposure. Inflorescence to 250mm, 10-15 flowers, lax. Flowers white.
Cooperi – Rosette to 120mm φ, often proliferous, stemless. Leaves 20-40, fleshy, swollen, oblong-lanceolate, quickly tapering, acuminate or truncating, marginal spines <2mm long if present. Bluish-green in colour, slightly translucent, with veins usually reddening and leaves developing purplish hues in exposed situations. Inflorescence compact, firm peduncle with many closely arranged flowers, to 20cm long. Flowers 20-30, perianth white.
Maybe it’s time to come back for more field work.
Lawrence Loucka
Yes writing those descriptions was a really funny experience but actually they are remarkably as good as you can get. Cooperi is only distinguishable from cymbiformis by colour, inflorescence and habitat. So what do you do with all those other populations that fall in the small cracks between. The “fieldwork” necessary to establish a workable boundary is to weed out the vast majority of plants that will not fit these artificial strictures of botanical classification and nomenclature. And just what is this stuff we weirdly say is “fieldwork”?
Really! I put it best I think in Update Vol 2 or 3 where I wrote about the need for a starting hypothesis for field work and reporting. I can tell you right now what more unplanned field work will produce – more names and more confusion. I have shown time and again that planned field work shows the interconnectedness of all these “species” we so anxiously claim. But we are also very limited by our perceptions and our expectations. We are taught mechanistically and we are not taught about the illusion of time and space or what reality is or might be. We do not live in a finite fixed world that is the same for each of us. We live within our own perceptions and dreams of what we think is real? We work with definitions of words and things and if we do not agree with each other on these, we will not understand each other. The word “species” is the great catch point hen it comes to biology. Science presently defines it for us by default, as a measure of the similarity of DNA in some sort of sampling (ill-defined and inadequate) process. This is going to create serious problems into the future. The polymath who asked plant taxonomists if they knew what they are doing, is going to have his doubts proven too right – they do not.
Bruce Bayer
I am always on about definition, usually of “species”, but checking up some DNA information reveals that even things such as nucleotides, DNA, and what-not are not actually adequately defined. DNA is said in some meanings, to be the nucleotide chains in the cell nucleus. The nucleotides are said to be the four amino-acids that link the paired strands in the chromosomes. But how are the strands constructed?
Then there are different molecular combinations effectively forming short strands in all tissues and these are also referred to as DNA. The Haworthia sequencing used to establish the recent changes within the aloids is based on up to 7 molecular regions, some of nuclear DNA and some on extra-cellular DNA. It bothers me that the sequencing does not actually tell me very much that I was not aware of simple observing the plants. But I see things in each of the phyllograms for the different regions that persuade me that we have a LOT to learn about DNA sequencing. The lineages for each region are not the same and there are some very weird anomalies that people who do not know the species are clearly not aware of. If they were they would be less confident in their conclusions and their faith in the technology (as it stands to date).
Glad that Lawrence posted that DNA paper Manning et al, because those interested can now see the phyllograms and make up their own minds about what they tell us. My personal experience makes me highly suspicious of all this stuff as even in the paper the authors say they opine that the results give a reliable overview – despite limited sampling and despite the variations in the different lineages the phyllograms illustrate. I would actually like to see phyllograms for each of the 5 DNA regions as well as concatenated ones, for just H. floribunda sampled from many populations. also not just single specimens from each population.
Here we have a case where the plastid sequences do not agree with the nuclear. I also do not like using both regional sequences AND concatenated results. Either concatenation is better or it is not. It seems like the statistical ploy viz. if a test does not give you significance, you keep using different statistical tests until you get one that does. The most critical element is that while these results are used to arrive at a generic classification in the aloids, the exercise was initiated to examine the species relationships in Haworthia only. Had that initial intention been observed (using 3 specimens per population) it could have made a really significant contribution instead of leaving us all in vacuum of opinion and belief.
My present view is that classification has devolved into an intellectual exercise that perpetuates confusion and uncertainty, ensuring endless name changes! Posted (above) is a nice retusaXcooperi ‘venusta‘ hybrid. It does not darken when exposed to direct day-long sunlight.
It is interesting to look at the DNA lineages for H. marxii, H. semiviva, H. pulchella var. globifera. H. mutica and H. marumiana var. archeri. They are, in the three data sets, very conflicting. For me highlight the problem of sequencing where sampling is so limited and based on an identification or classification process itself not based on sequencing. Particularly so when the original intention of the project was to examine that very problem.
This might have been called Haworthia dekenah var. argenteo-maculosa! In my opinion all the retusoids east of the Gouritz river are referable to H. pygmaea. But the truth is that one cannot separate retusa, turgida, mutica, mirabilis, pygmea or even emelyae on the basis of any physically observable evidence.
Indeed there is a problem with correct and consistent naming. The fact is that it can be much like pinning the tail on the donkey. It is all very well having a nomenclatural system that can be a juristic nightmare and an intellectual challenge, but does it work?
Haworthia cymbiformis is a case in point. According to the system, if a variety is recognised and described, automatically every other variant becomes the variety cymbiformis. This creates an amazing treadmill and we then have to accommodate all those other variants in a practical working system. How does one deal with this when, apart from the problems of one complex system, H. cymbiformis is inextricably tied into H. cooperi.
My conclusion has to be that the system does not work. While there is the fancy intellectual footwork in the upper echelons of botany, there is also the difficulty of dealing with countless variations that are conveniently set aside because they do not fit. Looking at my own attempts to deal rationally with H. cymbiformis and its variants boggles my mind. How did I set the name ‘planifolia’ aside, or renege on an earlier decision to abandon the name ‘obtusa’, and subsequently change my mind again? No matter how one juggles these names, one is faced with the embarrassment of dealing with many more variants that are neither one nor the other of the names one does decide to recognise and use. Let us say that this picture above is typical of H. cymbiformis and then look at some others …
What I tried to do in Haworthia was establish what groups existed that satisfied some rational species definition in also geographic terms. So I abandoned many of the old names for that reason alone. “Planifolia”is a case in point. I could find no clear geographic evidence and recognised the difficulty of actually separating cymbiformis and planifolia as distinct groups. But it is a subjective decision as so many taxonomic decisions are despite the desperate efforts to hide the doubt and the justify them as objective. as you can see from the two pictures, “planifolia’ has broader and flatter leaves rather than the keeled “boathull” shape of cymbiformis. I did not intend that names in synonymy be abandoned and did suggest that such names be considered available for use by collectors and growers.
Quote … ”The location of this note is a powerful reminder that distinctions between species are highly blurred and that alternative solutions are possible”. This answers my question of the previous post and it comes from Haworthia Revisited after tortuous explanations of the varieties in H. cymbiformis. Here is a photo of a cymbiformis in cultivation that could have come from anywhere, including Plutos Vale where H. cooperi var. tenera (?) and H. cymbiformis var. incurvula obfuscate the difference between the two species.
At this late stage in life, I think an honest submission is called for. My life experience with identification of plants is that botanical classification and naming is largely delusional. Taxonomists name things in relation to their experience and seldom get to truly know all the species and variants. Herbarium cupboards hide scores of unusual specimens filed in doubtful places and many unidentified. I have found it very difficult to deal with variations as this posted picture demonstrates. It is a variant of H. cymbiformis that I initially recognised as var. umbraticola in the belief that it substituted for the name var. obesa. At the time I thought that was a name confused with H. cooperi. I did eventually change my mind and recognise obesa as a variant of cymbifomis. But the fact is that cymbiformis is a riverine cliff hanger versus cooperi that is a grassland element.
There fall is a large spectrum of populations occur in different habitats that are neither one nor the other. This situation is also represented in H. retusa and H. turgida where there is further complexity in plant appearance and behavioural character viz. H. mirabilis, H. pygmaea, H. mutica, H. emelyae and more. These are realities of plant classification that taxonomists seem to dismiss or are unable to recognise. Things simply do not follow the tidy branching nature that fancy evolutionary theory, cladograms, phylograms and revisions illustrate.
This is now from an area in the Prince Alfed’s Pass where H. cymbiformis var. transiens, and H. planifolia var. translucens originated. It is to the west of general distribution of where more characteristic H. cymbiformis and H. cooperi occur. But my experience of these two species is that anything that resembles H. cymbiformis is actually a habitat derived variant of H. cooperi, which includes what was a very complex set of variants that I had treated as yet another species viz. H. gracilis. ♦



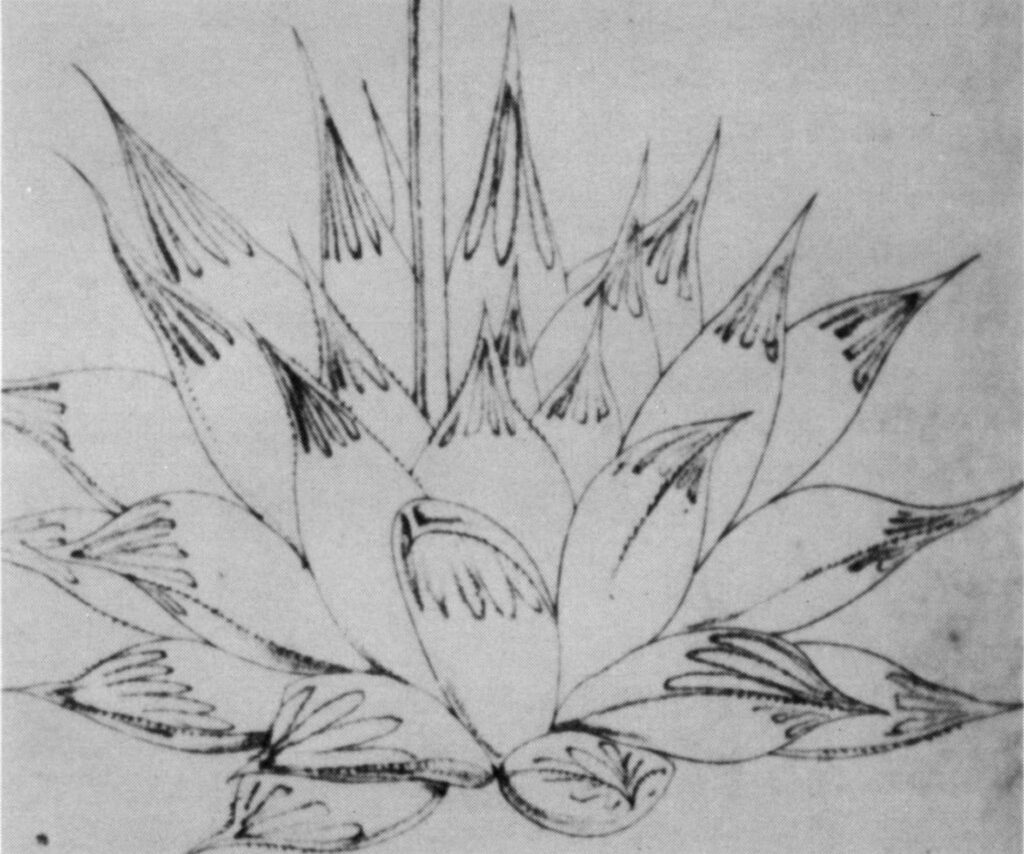
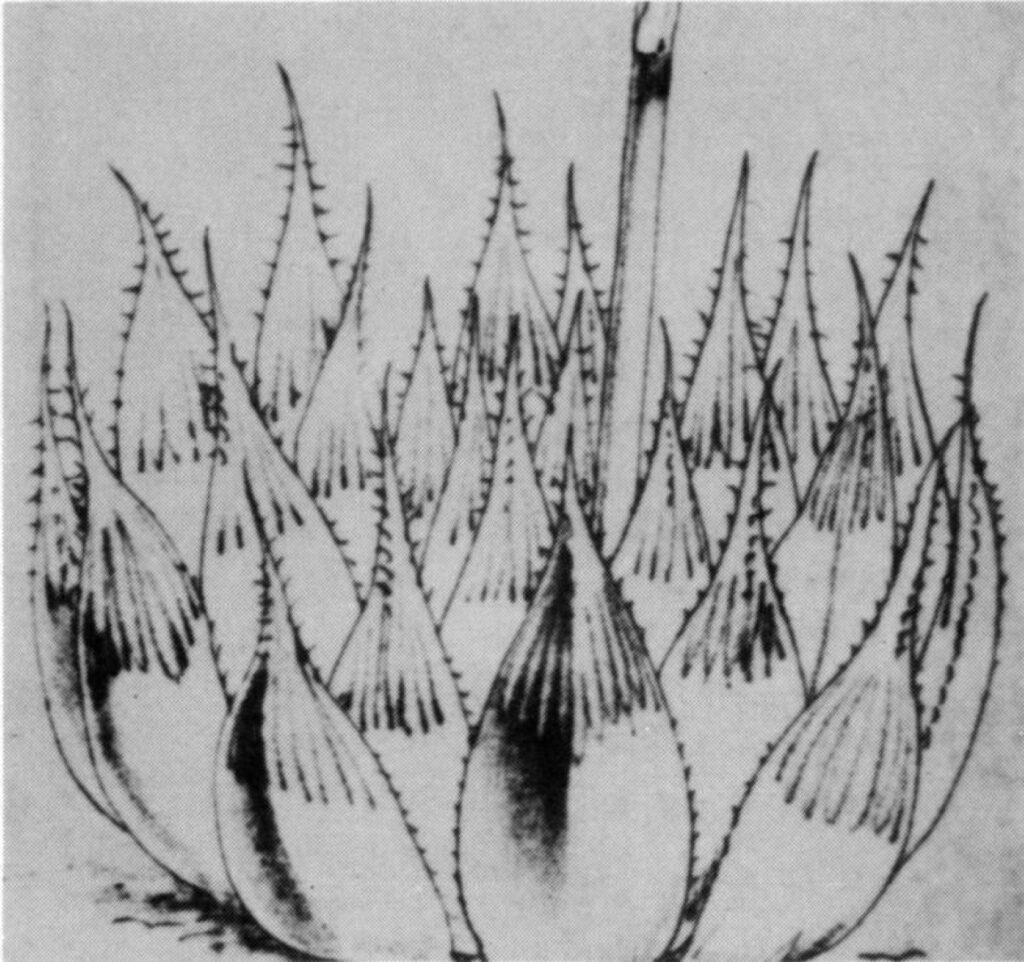


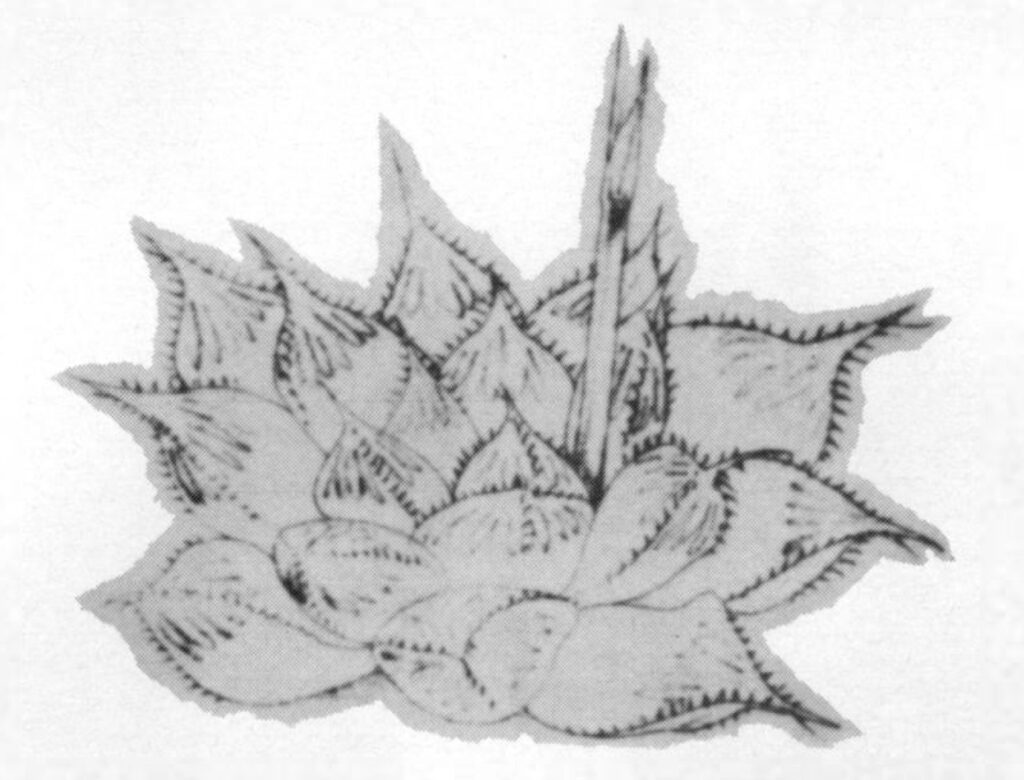
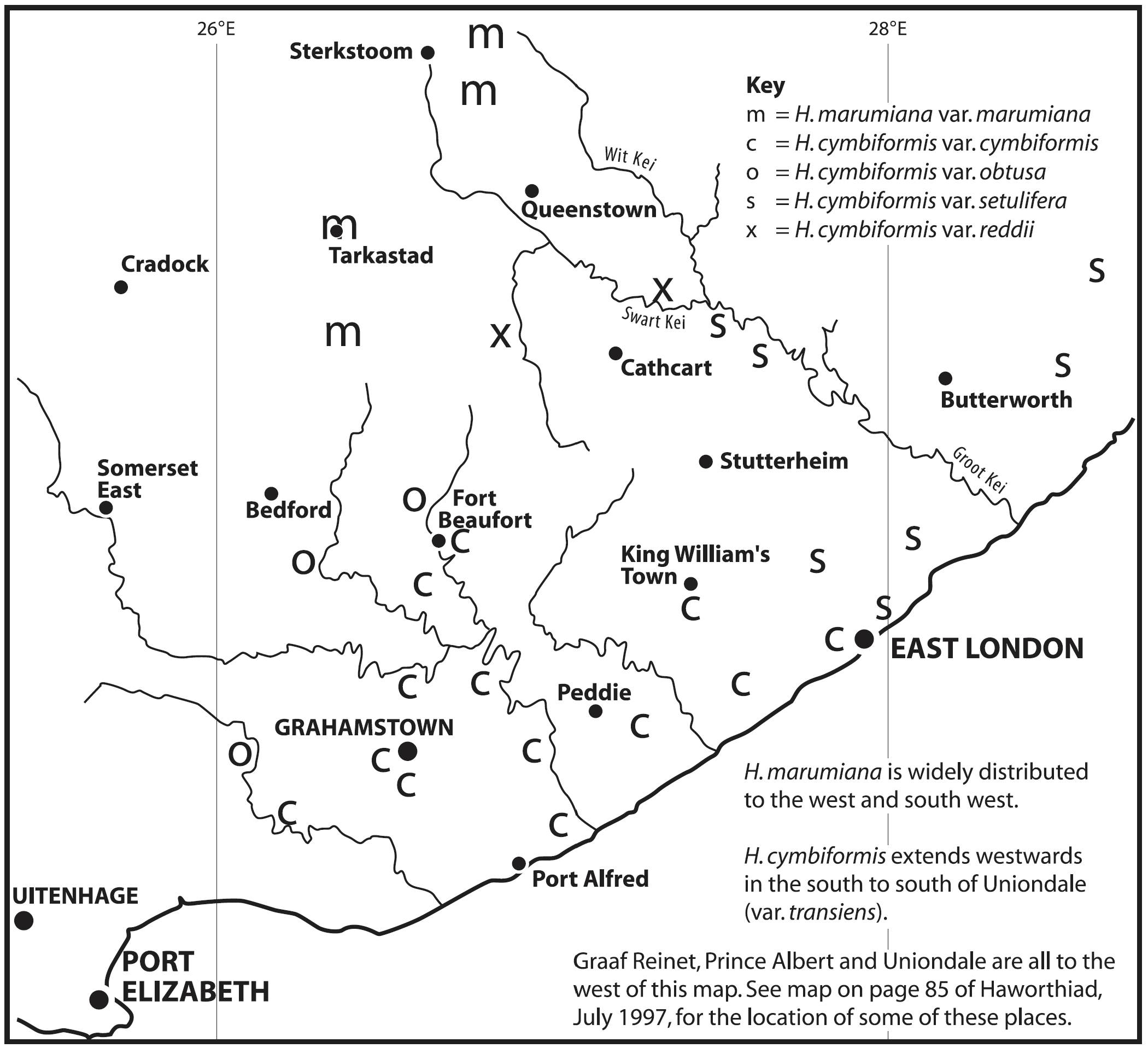






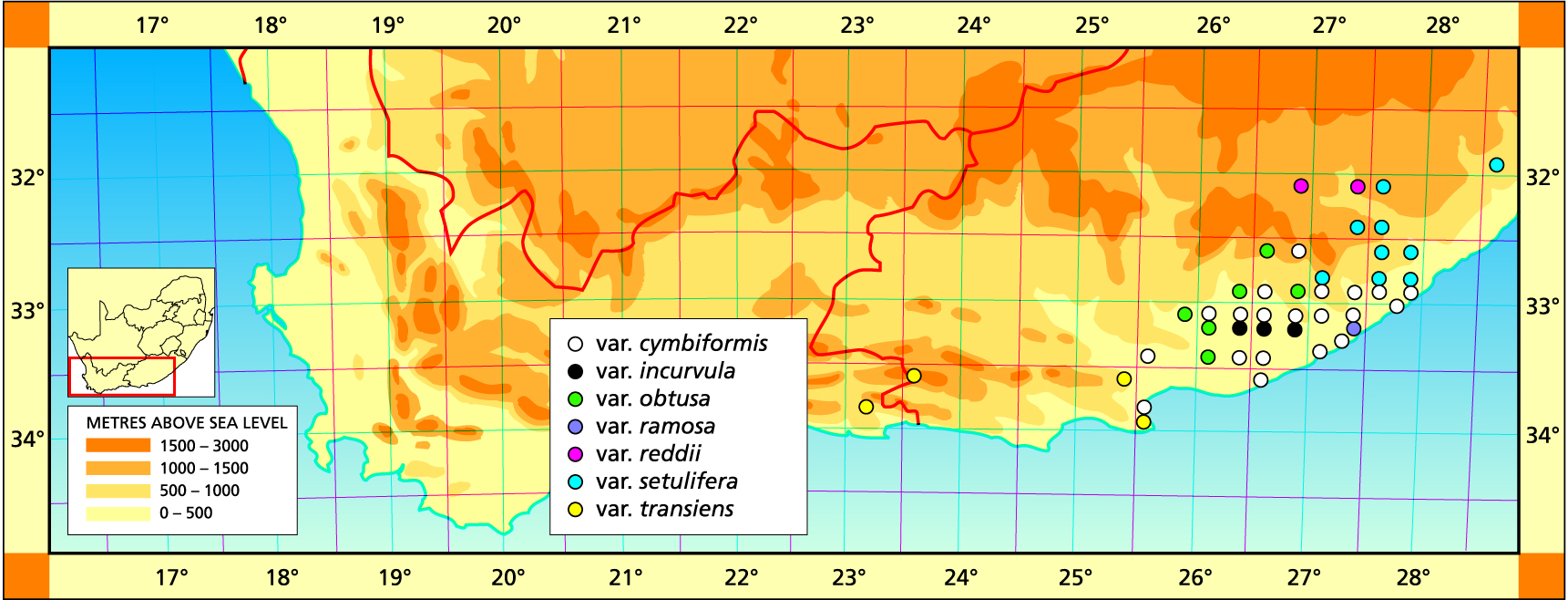












![H. cymbiformis - Curtis’s Botanical Magazine, vol. 21 t. 802 (1805) [S.T. Edwards]8067](https://haworthiaupdates.org/wp-content/uploads/1999/01/H.-cymbiformis-Curtis’s-Botanical-Magazine-vol.-21-t.-802-1805-S.T.-Edwards8067.jpg)
![H. cuspida - Addisonia, vol. 23 t. 741 (1954-1959) [M.E. Eaton] 162903](https://haworthiaupdates.org/wp-content/uploads/1999/01/H.-cuspida-Addisonia-vol.-23-t.-741-1954-1959-M.E.-Eaton-162903.jpg)