Rosette from 40-150mm φ, slowly proliferating. Leaves oblong-lanceolate, incurved, not acuminate or truncating, translucent bluish green, with spines on margins and keel >2mm long. Inflorescence robust, simple. Flowers broad and flat across upper basal part.
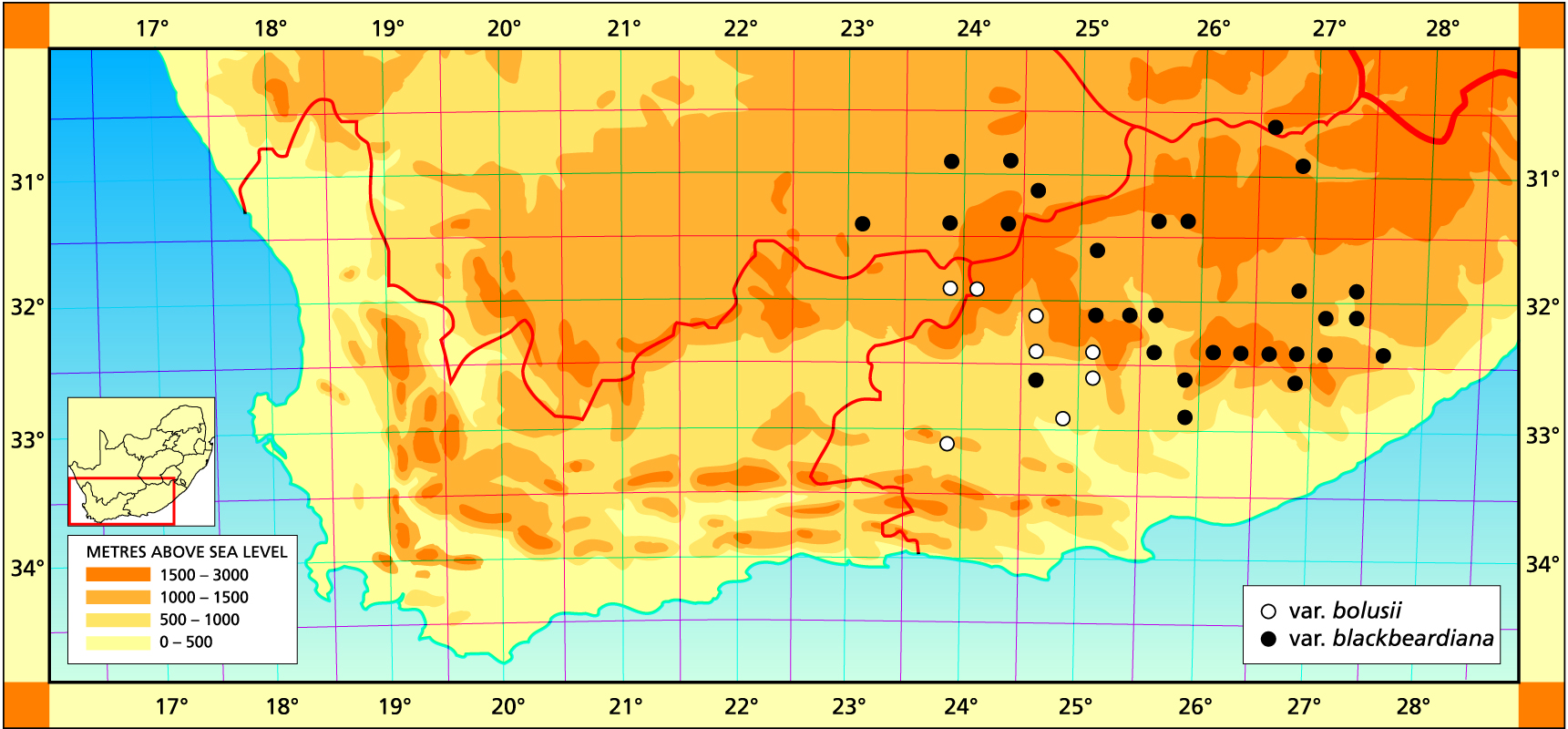
1982 – The previous concept of H. bolusii was based on plants occurring around Graaff-Reinet. Here the plants are 50-60 mm in diameter and the white bristly leaves are tightly closed over the centre of the plant. The essential difference between this species and H. arachnoidea is the colour. H. bolusii has translucent bluish-green or greyish-green leaves. Further investigation in the field has shown that it is not possible to clearly separate H. bolusii and the previously recognised and larger H. blackbeardiana. Thus H. bolusii is now here considered to be a widely distributed species with a fair degree of variability. It extends from Middelburg to Sterkstroom and southwards to Graaff Reinet and Cradock. The distribution extends further southwestwards in the eastern Karoo to Klipplaat and Jansenville, where very small forms occur in some habitats. It is probable that the small, relatively hairless plants south of Klipplaat are extreme forms of H. bolusii. Similarly there are aberrant, small forms with rather fewer but larger leaves than normal in the extreme northwest near Barclay East. At Middelburg the plants are also very large (up to 150 mm diameter) as they can also be at Cradock. There is no justification for upholding H. batteniae in which the supposed distinguishing characters, viz. spiralled florets and raised leaf venation, are wholly fortuitous. The problem really lies in separating H. cooperi and H. bolusii as discussed under the former species. H. bolusii is technically in the summer rainfall region, but responds to winter growing conditions. Good light is necessary if it is to be grown to handsome natural appearances.
1999 – Col Scott’s perceptions of var. blackbeardiana, H. mucronata, H. cooperi and H. batteniae perhaps reflect the problem in this melange of species. Scott’s H. mucronata seems to include the relatively glabrous elements of three different species. His 1983 discussion of that species includes what is quite obviously the spined H. cooperi var. pilifera. My perception is that H. cooperi is a main element and includes several of Scott’s species. It is in fact difficult to exclude H. bolusii if the var. blackbeardiana is placed with cooperi; unless a distinction is made between elements which tend to have at least partially truncated leaf ends. My dilemma is that blackbeardiana could have been placed as a variety of H. cooperi as there is unquestionably a close alliance. However, this is largely a problem of the nomenclatural system. Had H. cooperi not been described, it would have not been presented here as a species but rather as a variety of a more substantial species. I concluded that larger elements with the more lanceolate leaves and longer spines from the higher altitudes of the Eastern Cape formed a more logical and coherent entity. In the northern Transkei (higher altitude) there appear to be elements with truncated leaves which I would assign to H. cooperi var. obtusata).
A similar problem arises in the wider Port Elizabeth area where a similar transition to H. decipiens occurs. I have experienced difficulty with plants collected east of Pearston but have never had adequate material on which to pass an opinion.
a. var. bolusii. This typical variety is a smallish element in which the spination is dominant. The spines are fine and fully cover the plant so that it appears like a white ball. There is no clear integration with H. semiviva, but near Murraysburg and also at Victoria West the plants do show some tendency for leaf tips to die back as in that species. The collection from Jansenville cited below has understandably been confused with H. arachnoidea var. aranea because of the fine spination. It does have the bluish coloration of bolusii. A short distance away, on dolerite soils as opposed to tillite, the plants are larger and more typical of this species too.
Distribution: 3123 (Richmond): E. Murraysberg (-DD), Bayer 2389; Murraysburg (-DD), Banks (BOL). 3124 (Hanover): W. Graaff- Reinet (-CC), Bayer 2380 (NBG). 3224 (Graaff-Reinet): W. Graaff-Reinet (-BA), Smuts (NBG); Graaff-Reinet (-BA), Bolus 158 (BOL, K), Stayner in KG 232/62, (NBG), Ryder in NBG3152/34 (NBG); Bayer 2022, 2072 (NBG); Adendorp (-BC), Stayner in KG284/62 (NBG); Lootskloof (-DD), Bayer 2071 (NBG). 3225 (Somerset East): NE. Ashbourne (-AC), Bayer & Bruyns 6566 (NBG); NE. Pearston (-CA); Bayer 173 (NBG). 3323 (Willowmore): NE. Fullarton (-BB), Bayer 4160 (NBG); Eensaam (-BB), Marx 56 (NBG).
Haworthia bolusii var. bolusii JDV97/71 south of Graaff-Reinet. A small compact plant with fine spination.
Haworthia bolusii var. bolusii JDV87/186 south-west of Graff-Reinet. As one moves further south and eastwards, the plants seem to lose their compactness and dense spination.
b. var. blackbeardiana (V.Poelln.) Bayer Bayer :103(1976). Bayer :31(1982): H. blackbeardiana V.Poelln., Feddes Repert.Spec.Nov. 31:82(1932). Type: Cape, Bowes Farm near Queenstown, Miss G. Blackbeard. Not preserved. Lectotype (B&M): ex cult. V.Poelln. 1932 (B): H. blackbeardiana var. major V.Poelln. 41:196(1937). Type: Cape, Halesowen, Cradock, R. James in Parks 1483/36. Not preserved. Neotype (designated here): CAPE-3225 (Somerset East): beyond Halesowen station (-BA), Smith 2301 (NBG): H. inermis idem. 31:85(1932): H. altilinea var. inermis idem. 41:194(1937). H. altilinea var. limpida fa inermis idem. 49:29(1940). Type: Cape, Halesowen, Cradock (-BA), Stell. Not preserved. Neotype (designated here): CAPE-3225(Cradock): Halesowen (-BA), Smith 2289 (NBG): H. batteniae Scott, Cact.Succ.J(U.S.). 51:268(1979). Type: CAPE-3225(Cradock):, 35km west of Cradock (-AA), C.L. Scott 5272 (PRE): H. cooperi Bak. pp. Scott ibid 53:70(1981) as to Cradock and Thomas River. Scott :104(1985): H. mucronata Haw. pp. Scott ibid. :80(1986) as to Cradock specimen.
blackbeardiana: in honour of Miss G. Blackbeard.
This variety is generally larger than the typical form and the spination is sparser. However it is more densely spined than is H. cooperi, and the spines are longer, exceeding 2mm. The definition is otherwise geographic and the interface with H. cooperi is from south of Cathcart in a southwesterly direction to at least Jansenville. In the general Rippon Station area there are two adjacent populations, one of which is attributable to this species and the other to H. cooperi. The Patryshoogte/Baviaanskranz collection is also this species growing in the same general area of H. cooperi var. dielsiana. The interpretation in this work of H. pringlei Scott as a variety of H. decipiens may be erroneous as it may perhaps be better placed with var. blackbeardiana. This is also suggested by the Patryshoogte specimens.
There are forms of var. blackbeardiana from the Queenstown area which have very few or sparse short spines, and similarly on the Hogsback mountain. It has been reported that var. blackbeardiana has been collected in the southern Orange Free State and that its distribution extends considerably westward from Middelburg. I have no confirmation of this. There are also reports that it intergrades with H. semiviva in the far west of its range, and the sparse herbarium record suggests that this may be true.
Distribution: 3023 (Britstown): Die Puts (DD), Fuller 193 (BOL). 3024 (Philipstown): Hanover to De Aar (-CD), Stayner in KG307/70 (NBG); 3026 (Aliwal North): Elandshoek (-DA), Smith 6828 (NBG); SE. Aliwal North (-DD), Smith 7471 (NBG). 3123 (Richmond): Richmond (-AC), Stayner in KG591/69 (NBG); Richmond (-BD), Wohlman in KG581/69 (NBG). 3124 (Hanover): Cypherwater (-AD), Bruyns 3010 (NBG); Bethesda Road to Goliadskraal (-BA), Smith 3640 (NBG). 3125(Steynsberg): Thebus (-BC), Smith 3635 (NBG); 8km E. Steynberg (-BD), Smith 3634 (NBG); Tafelberg (-CA), Smith 3828 (NBG), Stayner in KG693/71 (NBG). 3126 (Queenstown): Bowes’ farm (-DD), Britten 160 (BOL), Bayer in KG 312/70 (NBG); Finchams Nek (-DD), Smith 7212 (NBG); Nonesses Nek (-DD), Smith 7216, 7217 (NBG); Longhill (-DD), Bursey in KG393/70 (NBG). 3127 (Molteno): Cofimvaba (-CD), Bruyns 4385 (BOL). 3224 (Graaff Reinet): Kendrew (-DA), Rossouw 183 (NBG). 3225 (Somerset East): 35km west of Cradock (-AA), C.L. Scott 5272 (PRE); Doornberg (-AA), Herre STE6632 (BOL); Denmark (-AB), Bruyns 1769 (NBG); Post Chalmers (-AB), Smith 7444 (NBG); 24km SE. Fish River Stn. (-BA), Smith 5791 (NBG); beyond Halesowen station (-BA), Smith 2301 (NBG); Halesowen (-BA), Smith 2289, 2301, 5340 (NBG), Herre STE6668 (BOL); Cradock (-BA), Smith 2263, 2289 (NBG), Britten (BOL); Swagershoek Pass (-BC), James 474 (BOL), Fourcade 40 (NBG), Bruyns 1627 (NBG); Daggaboer (-BC), James (BOL); Baviaanskranz, Patryshoogte (-DB), Bayer & Bruyns 6561 (NBG); NW. Rippon Stn. (-DD), Bayer & Bruyns 6556 (NBG). 3226 (Fort Beaufort): Huntley Glen (-AC), Smith 7493 (NBG); Austrey (-AD), Branch 370 (NBG); Katberg (-BC), Luckhoff in NBG407/34 (NBG); Waterdown Dam (-BD), Bayer & Bruyns 6569 (NBG); S. Estrelle (-BD), Bayer & Bruyns 6570 (NBG); Hogsback (-DB), Smith 375 (NBG). 3227 (Kingwilliamstown): Imvani (-AA), Bayer in KG311/70 (NBG); 24km N. Cathcart (-AA), Smith 3633 (NBG); N. Waku (-AA), Smith 5718 (NBG); 2km N Goshen (-AA), Branch 20 (NBG); Turnstream (-AB), Bayer & Bruyns 6571 (NBG); New Haven (-AB), Branch 18 (NBG); W. Cathcart (-AC), Smith 5744 (NBG); 16km W. Cathcart (-AC), Smith 38, 358 (NBG); SW. Cathcart (-AC), Barker 3427 (NBG), Bayer in KG310/70, KG393/70 (NBG); Inverbolo (-BC), Bruyns (NBG).
Haworthia bolusii var. blackbeardiana JDV91/102 soth of Cradock. This variety is widespread and the spination is coarser and sparser, or occasionally absent.
Haworthia bolusii var. blackbeardiana JDV91/99 south of Cathcart. Here the plants are often sparsely spined and easily confounded with H. cooperi.
Haworthia bolusii var. blackbeardiana JDV97/62 Aliwal North. The variety is not well-known at its north-eastern limits.
One of the greatest difficulties in Haworthia is that of trying to recognise discrete species. This translates into confusion which can be attributed to writers. The initial source of confusion is without doubt the nature of the plants themselves, and this is not a problem confined to Haworthia. The species are often not easily recognisable and discrete entities. I abhor the statement that the genus is in a state of active evolution, but this does at least seem to convey a message that readers understand, even if it is somewhat hackneyed. My observations on Haworthia are based on a definition of species as a system of living organisms which are continuous in time and space. In my New Haworthia Handbook, I suggested that a primary problem lay in separating H. bolusii and H. cooperi, and for the purposes of that work I largely discounted the secondary problems. My first concern was to identify core areas and names as working postulates. This did not mean I was unaware of lesser problems contained within the recognition of those two species. The purpose of this paper is to present my current understanding of the problem.
In my opinion the publication of Scott’s book and whatever merit it had, broke the foundation for understanding. As Bruyns said (Kew Magazine, 1986), it set taxonomy in Haworthia back by 40-50 years. This is because Scott did not attempt to examine what I had done, and was doing, in any objective and cognitive way. Thus there was no progression from a common hypothesis, or from a common concept of ‘species’. In my opinion his work was written in the paradigm of Reynold’s work on Aloe, or of Von Poellnitz and G.G. Smith, and was thus anachronistic. My work was in the intellectual climate of the day, was influenced by researchers of the time and was based on a definition of ‘species’. Scott’s thoughts and actions were both understandable and excusable. His classification reflects the same problems, but differently to mine. Where I recognised H. bolusii he recognised H. bolusii. Where I recognised the var. blackbeardiana he recognised H. cooperi and H. batteniae. Where I recognised cooperi he recognised H. pilifera and H. altilinea. Except for his contention that these latter are separate species, there is really concordance. We recognised different markers along the same continuum.
Essentially I considered that there were two species with two major facies, thus:-
1. H. bolusii – var. bolusii the smaller very densely spined typical variety which seems to diffuse into the species H. semiviva in the west of its distribution range.
– var. blackbeardiana a larger less densely spined element to the south and east which seems to diffuse into H. cooperi. Scott used the name cooperi for this element. There is merit in this, but he had to describe another species, batteniae, because of the inherent tension in the solution (I must point out here that Scott’s problem may have been the inadequate herbarium record that he used for his work, plus the fact that he made few specimens. Thus there is no way of assessing his decisions. Had he had to physically file specimens as I have done, he might have had great difficulty with batteniae and his cooperi, and also in separating his altilinea and pilifera). I regarded bolusii as a more spinose species than cooperi, with thinner and wider leaves. Cooperi then generally less spinose, squatter and with thicker leaves.
2. H. cooperi – var. cooperi itself, with relatively erect slender leaves, and including all the forms with more truncated abrupt leaves.
– var. leightonii – the coastal form with even more attenuate leaves than the typical form, and also very proliferous.
The solutions offered by either myself (1982) in respect of my cooperi and blackbeardiana, or in the case of Scott (1985), altilinea and pilifera, cooperi and batteniae, do have problems. These arise from the nature of the terrain in the area between Graaff Reinet to Queenstown and down to Fort Beaufort. Rugged broken terrain difficult to explore. Like the Baviaanskloof it offers many different classes of habitat, and thus potential ecotypes. The relationship between the varieties is complex and compounded by continuities with other species e.g. gracilis, cymbiformis, and decipiens. It should also always be remembered that any decision is a product of the collecting record and must emphasise that this is the context in which this article is written. Initially I did think that the typical variety of H. cooperi may possibly prove to be well-defined geographically. It has not proved indisputably so, but my classification nevertheless does allow good expression of the continuities which occur. I used the name to cover forms with erect slender leaves as well as forms with relatively blunt truncated leaves. Such plants do occur within the same populations and this is evident in the J.G. Marx collection from Fort Brown. The corresponding Scott names were altilinea and pilifera. To make my system more workable, in my new book (1999) I recognised H. bolusii and its var. blackbeardiana as before, but in H. cooperi, several more varieties, thus:-
– var. dielsiana – a more truncated version of pilifera, the leaves often without end-awns. (Scott’s “joeyii” is synonymous).
– var. truncata – a eastern lighter coloured, proliferous and smaller version of dielsiana.
– var. leightonii – the pinkish, proliferous, slender leaved form growing on granites at Kaisers Beach (Scott maintained this as a separate species).
– var. gordoniana – the erect, slender leaved form in the Hankey and Patensie area, which is very like the typical variety cooperi. In truth var. gordoniana from the type locality is a small very compact plant with short incurved and finely spined leaves.
– var. venusta – a very localised hairy variant from the coast near Alexandria.
I hoped this would explain the variations and resolve the tensions in the solution in an economical way. There is no doubt that Scott’s argument of cooperi sensu Bayer, and blackbeardiana sensu Bayer being the same (and he used only the name cooperi in this context), is correct in the nomenclatural sense. The Thomas River specimen he cites in his book as representative (although Scott does cite the Kew specimen as type, which I have not seen), would have been attributed by Bayer (myself, using the Saunder’s Refugium illustration as type) to my concept of H. cooperi, as also the doubtful gracilis-like elements for the Adelaide and Kingwilliamstown citations. I would have named the Cradock specimen H. bolusii var. blackbeardiana. To bridge the difficulties inherent in his solution, Scott had to later recognise three species viz. batteniae, pringlei and joeyii, with the potential for many more. I have said elsewhere, that a classification which grows with the description of new species, is indicative of a weak system. As in the case of bolusii, the Scott solution is not as economical as mine. His concept of an altilinea and pilifera is essentially the image of my problem in recognising a cooperi/blackbeardiana interface. It would have helped had I initially recognised pilifera as a variety. However, Scott’s interpretation in the sense of the geographical relationships of his various species is problematic throughout his revision, where specimens are not cited and full use has not also been made of the available herbarium record.
I was predicting that an element, namely H. cooperi var. cooperi occurred which could be defined in a geographic context. The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain goodwill. This article thus is written to expand on the problem and indicate where the difficulties lie. These are not difficulties that I was oblivious of, or tried to obfuscate. I have many times said that we can find solutions which work in one area, but they may not work in another. This will again be evident in this paper.
In my revision (Haworthia Revisited, 1999) I made the combination H. decipiens var. pringlei and resurrect the old species name H. aristata. These two elements, and an explanation for the tensions which these two names create and try to address, will be touched on here but will follow in more detail in a later article. As in this immediate case, new material has been seen and collected since the revision was drafted, and a better explanation can now be made.
For this particular issue, the key questions to the hypotheses of Bayer and Scott are difficult to formulate because they are confounded by the different use of names. Therefore I frame the questions in terms of my own classification like this:-
Does an element we can identify as cooperi grow in the same close geographic context as blackbeardiana? (Bearing in mind the assumption that the hypothesis regarding continuity between bolusii and blackbeardiana in fact is valid. Also noting that to maintain the classification hypothesis that there is a difference at the rank of species, the answer must be ‘yes’).
Does the element we can identify as cooperi grow in the same close geographic context as pilifera? (Bearing in mind that to maintain the classification hypothesis of a difference only at the rank of variety, the answer must be ‘no’).
Results The answer to question 1:- has never been a strong point of my classification because I saw cooperi in a broader sense to include pilifera (and altilinea of Scott). The recognition of varieties now strengthens the classification in one respect but weakens it in another. It strengthens the classification in respect of recognising the extent of the variability within my concept of H. cooperi. It weakens it in suggesting both a strong geographical separation of the varieties and a weaker relation with H. bolusii var. blackbeardiana. In Nov.1996, I went with Peter Bruyns to the Eastern Cape and we spent some time in the greater Somerset East area. In Dec.1996 I travelled with Kobus Venter. In Dec.1997, Dec.1998, in Sept. (with Tony Dold, Dez Weekes and Steven Hammer) and again in Oct.1999, I travelled with my wife to these areas. We made many significant collections.
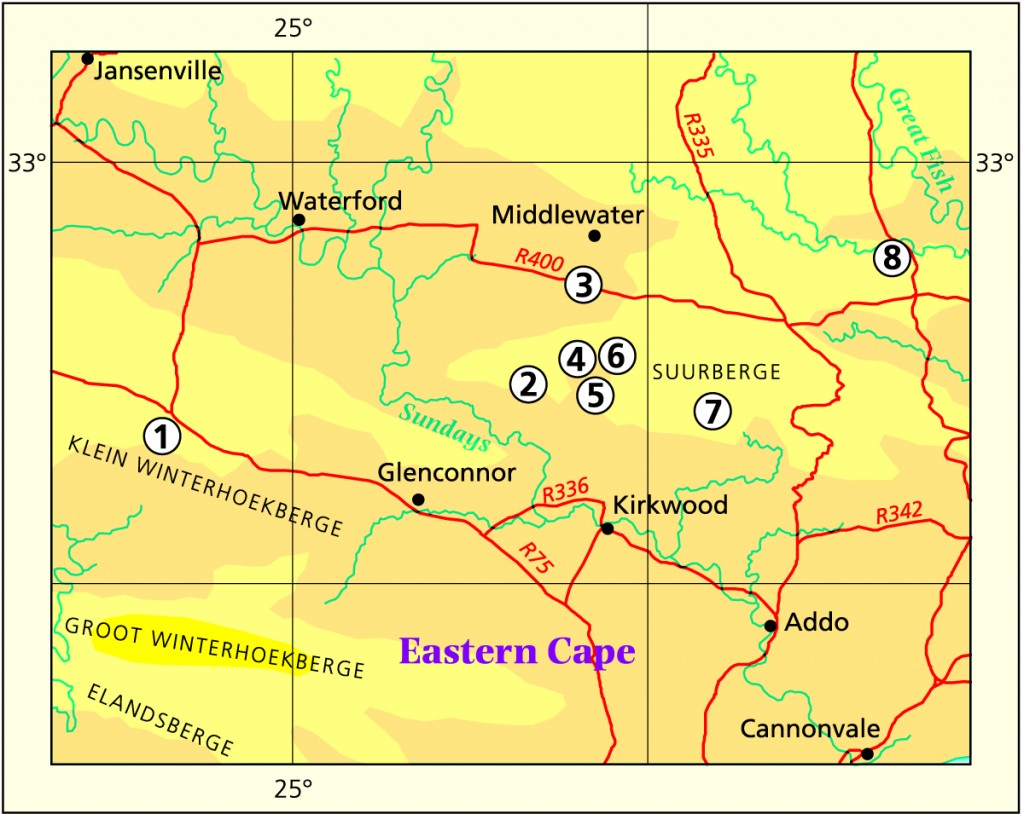
The first of these concerns Scott’s species, H. joeyii and H. pringlei. In the case of joeyii my contention that it is continuous with pilifera barely needs discussion, and I do not think the presence or absence of an end-awn is necessarily diagnostic for such an element – hence synonymy with var. dielsiana. We found three discrete populations within a small radius around Eastpoort (MBB6558-102, 6559 and 6560-103) which support this observation. I do not think there are strong grounds for separating it from pilifera, although I have now done so. At Bedford (there are plants with the similar abrupt leaf-tip as well as plants without. I currently have four batches of seedlings which I want to examine for variation. One from Slagtersnek, south-east of Somerset East (MBB6778-104) is particularly variable. I even have a collection of H. cymbiformis (MBB6847-100,-105) which has the same truncated, awnless leaftips as Scott’s joeyii. Regarding Scott’s pringlei, my conclusion that it is related to decipiens is possibly geographically and otherwise incorrect. (It should be noted that no specimen of pringlei has to date been deposited in the Pretoria herbarium. My understanding of the species is from the description, from two plants sent to J.D. Venter by J.N. Reddi (JDV93/46-106) who collected the plants for Scott, and from two plants given to G. Marx (JDV93/52-107) also by Dr Reddi). It is a problem of ‘look-alikes’ and I will deal with that a little later as a separate issue.
Further south and east of Eastpoort, at Patryshoogte (MBB6561-108 also -4), we found a different population not to my knowledge co-occurrent with a cooperi variety, which must be comparable with Scott’s species i.e. pringlei. It has narrower and more elongate leaves than blackbeardiana generally has, and the keel may be more pronounced. Nevertheless I am sure it has a connection with H. bolusii var. blackbeardiana. In terms of a much wider knowledge of other populations, I decided to link pringlei to decipiens, to emphasise the evident and probable continuity between that species and H. bolusii. This is particularly so in the area north and east of Jansenville.
To the south-west at Somerset East (MBB6776-109, Glen Avon), we found a population of bluer green plants with rather more attenuate leaves than in the vars pilifera and dielsiana. These plants are very like an Adelaide (Koonap Bridge, MBB6563-110) collection, these both satisfy my concept of the var. cooperi. It was not much further to the south (-104), and also not far south of Eastpoort, where we found forms of the vars pilifera and dielsiana in the same population. Thus it seemed that a hypothesis maintaining the species cooperi and H. bolusii var. blackbeardiana, was well supported, but not so the contention that pilifera and dielsiana were good varieties.
The most significant discovery up to Dec.’97 was of two different elements growing together towards the south, near Ripon Station. By together, I mean as populations occupying different niches in the same close proximity – not growing as a common medley of individuals. One population was clearly the typical blunt-leaved pilifera (MBB6557-111) which included dielsiana, but the other (MBB6556-112) was with more slender and erect leaves viz pringlei, and possibly the leaves being broad and spinose enough to even represent blackbeardiana. Thus the added complexity is that it is not possible to separate those two elements. After two years in cultivation as field collected clones and as seedlings, the difference from pilifera (as -111) is maintained. The collected clones grown in Kobus Venter’s collection convincingly demonstrate that the one population is identifiable with pringlei/alias blackbeardiana and the other with cooperi var. pilifera. Thus the answer to the first key question is “Yes, they do grow together in the same geographical context, because there is that obvious connection of blackbeardiana with pringlei. It seems probable that both these elements could, and perhaps should, be interpreted as varieties of H. cooperi. It is a complex problem which cannot be separated from consideration of other elements such as H. gracilis and H. aristata, and these are the issues I am addressing in these papers.
There is not much further evidence that cooperi var. cooperi is a strong, discrete and valid element. There is a collection by J.G. Marx of plants in the Hogsback area (in JDV91/82-113, Woburn) which may be identified as this variety, as can the two collections already mentioned viz. -109 Glen Avon, and -110 Koonap Bridge (originally a Marx collection). However, there are collections generally (one is D.M. Cumming8489-114 of plants south of the Waterdown Dam, Upper Chilton), where the plants can be confused with H cooperi var. gordoniana (in the context of the problem in the Baviaanskloof), whereas geographically they should be blackbeardiana. I also made a collection at Waterdown Dam itself (MBB6569-115), of a plant which is very like a spineless smooth gracilis. These plants tend to be much more blue in colour than the southern look-alikes. There is also JDV96/89-116 Gladhurst, Adelaide; and JDV96/4-98 and MBB6603-99 from Glen Craig, north-east Grahamstown, which pose a similar conundrum. These are gracilis-like and while exploring that problem, the issue became very much more prominent, and is even expressed in the discussion regarding H. cymbiformis var. incurvula. Kobus Venter and I had both re-collected H. gracilis var. gracilis at Hellspoort (JDV89/42-117, MBB6614-118), although we had not concerned ourselves with a search over the whole of the valley. What is now becoming more evident is that there is an archetype which is gracilis-like. Thus Hellspoort needs to be examined as closely as Pluto’s vale.
A later collection of mine (Oct.’99, MBB6927-119, W Ripon) influences the picture dramatically and may further prove a statement of mine true. This is that there instances where there are no real boundaries between species. My collection is further west of -111 (pilifera/dielsiana) and -112 (pringlei/blackbeardiana). Here in -119, the plants include individuals which could be either identified as the vars cooperi or pilifera, with the former collection equally well representing pringlei/blackbeardiana.
The answer to question 2:- has not really been a problem. What we found was what we expected to find and more. South of Adelaide one finds populations of plants which satisfy the tendency towards slender leaves ie. var. cooperi (-109 Glen Avon or -110 Paardefontein, Koonap Bridge); and populations which satisfy the tendency to short obtuse leaves ie. var. pilifera eg. MBB6564-120 Chancery Hall or MBB6591-121 The Tower, S. Fort Beaufort. The same applies just east of Somerset East where in the broader geographical context we can identify var. cooperi (-109, Glen Avon), var. dielsiana (when mostly or wholly without end-awn, MBB6565-122, W. Somerset East, -111 W. Ripon Statio, and vars dielsiana and pilifera as geographically complementary and even in admixture (-104, Slagtersnek). Thus “No, the varieties cooperi and pilifera do not grow together in the same close geographical context as discrete entities.” In fact they are often represented as forms in the same collections as in -119 (far west of Ripon) where both varieties are present as single plants as well as a longer leaved form representing blackbeardiana as decipiens var. pringlei.
The answers given to these two questions thus do still not solve the problem as there remain populations which can not confidently be ascribed to either of the names here used. There are populations which may thus be assigned to cooperi or to blackbeardiana or pringlei. An example is the Marx collection from north-east of Grahamstown (W Fort Brown, in JDV91/85-123) which is ascribable to pilifera although some of the clones are inseparable from blackbeardiana. JDV98/39-124 from Brakkloof, northwest of Grahamstown poses a similar problem with the leaves of the plants tending to lengthen in cultivation. The ultimate difference then between cooperi and blackbeardiana becomes a very subtle one of degree of spination and leaf bulk. The issue is further complicated by the fact that four other elements are involved in the total geographical context, namely H. cymbiformis, H. gracilis, H. decipiens, H. aristata and H. arachnoidea var. xiphiophylla.
My most recent field work, was in following up information and collections concerning H. gracilis. Tony Dold of Rhodes University gave specimens of an Haworthia from the Annsvilla area to Gerhard Marx (MBB6851-125). He thought they may have some bearing on the existence and reality of my interpretation of H. aristata, and this is indeed the case. The plants are small, the darkish blue-green of the cooperi/blackbeardiana elements, and with more, smaller, and quite spinose leaves. Annsvilla is close to three localities for that species cited in my revision viz. Verdun, Stonefountain (re-collected in MBB6852-126 also -4) and Kommadagga. Following Dold’s find, recollection in the vicinity of each of those sites, produced plants which can be interpreted and identified in the light of the Ripon collections. While they equate in some way to H. bolusii var. blackbeardiana, they also do to H. cooperi and to H. decipiens var. pringlei. An (unfortunately doubtful) Kommadagga collection (EvJ sn.-127) is very similar to the long-spined gordoniana-like plants from the eastern Joubertina -74, Uniondale -75, Longkloof -76 and Baviaanskloof (Geelhoutboskloof -78, Nuwekloof -79). The leaves of the plants are rather flatter and broader, and so closer to blackbeardiana. In Sept.’99 Dold and Hammer found a small plant with shorter and squatter leaves than my concept of aristata, south-east of Kommadagga (in MBB6897-128). We also saw plants from the area east of Jansenville which seem to link the greater H. bolusii (i.e. var. blackbeardiana) complex with either H. decipiens or H. arachnoidea var. xiphiophylla. Included in that broad statement is an explanation for the specimens which I have cited as H. decipiens var. pringlei in my revision. These include recent collections from north of Jansenville (MBB6580-129), north-west (MBB6581-130), and south-east of Mt Stewart (MBB6582-131 also -6, 6583-132, also -7). When I first saw a specimen of pringlei my first reaction was recall of plants I had collected at Mt Stewart, and of plants said to have been from Jansenville. Thus I created H. decipiens var. pringlei, expecting to find this east-west continuity between blackbeardiana and decipiens. I did not think that the connection between Middleton (pringlei) and Jansenville would be found to be as evident and as strong as it is. There was already some evidence that there is south to north-east connection between Jansenville and, say, Cradock. This is of a connection between H. decipiens and H. bolusii.
A complicating issue is that MBB6587-133 (further SE Mt Stewart than -132) is a population which is H decipiens var. minor. A small plant with a tendency for incurving leaves but with the widely spaced large spines I associate with the more classical view of H. decipiens. The next collection was MBB6589-71.2 (a little south-east of -133) and it throws us into the H. gracilis context of smaller plants, clustering on cool south-facing rock faces. There are many collections which continue this trend to gracilis, and in fact I suspect to xiphiophylla too, and that will be dealt with as a separate issue. The field-work also revealed populations (J.G. Marx in MBB6845-134, east of Alicedale, and MBB6847-105, also Alicedale) which link cooperi to cymbiformis. This also can be dealt with together with two collections from near Kagasmond (MBB6562-135, south of Adelaide and JDV96/89-116, Gladhurst) as a slightly different issue.
Regarding H. aristata, I resurrected this name for several collections from the Eastern Cape, and somewhat justified by the Dold collection at Annsvilla (-125). However, it seems quite certain that the issue is clouded. Re-collections at the Soutkloof (JDV96/90-136, Addo, Dead Man’s Gulch) were more like pilifera, but there is evidence of deviation. Here D.M. Cumming (DMC3870-137 collected pilifera-like plants, whereas Venter, Marais and Bayer (-136) collected a variant – there was a more typical bluish-green pilifera and a plant tending to the opaque yellow-green of xiphiophylla. I returned there in Oct.1999 because I still had not seen anything like the plants I had seen there on an earlier visit, or like W.R. Branch’s original (WRB459 in JDV87/53-138), and the other cited collections. On this occasion I did find a small population (MBB6920-139) of this kind, and these confirm a relation to the more northern collections mentioned above. At the nearby village of Addo itself, there is the fairly normal pilifera (JDV86/117-140). The Annsvilla collection (Dold and Marx -125) and the recent collection of my own from Stonefountain (-126 also -5) seem also to be in the context of the Ripon collection (-112) of pringlei. These are, however, collections of much smaller plants than from Ripon. Thus it confirms for me that the Haworth names aristata and denticulata could easily have had their origins in plants from this area between Ripon and the Zuurberg, or further south to Soutkloof. My collections MBB6916-141, MBB6917-142 and MBB6901-143, from Kaboega and Hopewell are of the same order.
Discussion and conclusions There is still a very large area unexplored. There are several like collections from the greater area Kirkwood to Uniondale which I have generally ascribed to H. cooperi var. gordoniana. None of these are the short-leaved pilifera type. The teasing probability is that it is in fact blackbeardiana, pringlei, gracilis or aristata (as variants of H. cooperi) which are the main role players. They differentiate (clearly?) in the east to pilifera or cymbiformis and in the west into decipiens. In the south they pass to xiphiophylla, cymbiformis and varieties of gracilis. North-westwards it is to bolusii. My opinion now is that we have an archetype which is in the mould of H. gracilis and this is the root of all the elements I have named here. Curiously Tony Dold has recently sent me specimens from Chalumna (T.Dold3961 in MBB6921-144) which suggest that an aristata-like element is also associated with H. cooperi var. leightonii. Other Chalumna collections (MBB1621 and G.G.Smith 514) bear a very close resemblance to plants (-99, -116) which I have said are gracilis-like.
The above statements all have to be seen in the overall statement about continuity. It barely seems practical (nor legally possible) to sample and analyze plants on the scale that will be necessary, using whatever technique, to get a more definitive answer. The best answer will be continual exploration of the simple kind reported here, which explains occurrences on a smaller and smaller scale. There are herbarium records which need to be corroborated in the field but these do not suggest to me a better solution. Ultimately each recorded population will have to be assigned unequivocally to a taxon (or taxa!) and this is the next necessary step in Haworthia classification. Another revision based on less exacting field observations and a lesser record, will simply exacerbate the regression in time which Scott’s work precipitated, and the consequences which we now suffer. My recent collections are only preserved as living collections and photographically. Specimens need to be made.
I am very conscious of other tensions in my classification and I think it is imperative that we stay with one nomenclatural arrangement and hypothesis to resolve these. The confusion which arises from nomenclatural changes which are nothing but pretentious and cosmetic, is not worth any price. Any difficulties in respect of nomenclature can be resolved by a process of explanation and conservation without formal name change. These changes can be made when it is expeditious to do so and when changes can really offer a better explanation of the genus and hence better communication.
What this discussion should demonstrate is the problem of really observing and discussing variability in Haworthia. There are many possible arrangements of names which can be presented as conclusions in themselves – but done like this they simply cloud and destroy any hope of a broader understanding and good communication. Haworthia is now so much in the public domain, that I would suggest to editors that they move in the direction of encouraging authors to adopt a conserved nomenclature. I regret to be so straightforward and blunt. I see nothing but further confusion if persons feels that they can contribute to an understanding of the situation in the field without:
Consensus on the issue of species definition.
Consensus on the issue of a set of names.
Familiarity with herbarium records and what these represent in terms of fact and fixed reference points.
Familiarity with the written record.
Familiarity with South African geography and the ability to interpret populations in that context.
♦
102 a. MBB6558 H. cooperi var. dielsiana, north of Eastport.102 b. MBB6558 H. cooperi var. dielsiana, north of Eastport.103 a. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 b. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 c. MBB6560 H. cooperi var. dielsiana, south of Eastport.103 d. MBB6560 H. cooperi var. dielsiana, south of Eastport.104 a. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 b. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 c. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 d. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.104 e. MBB6778 H. cooperi var. pilifera, Slagtersnek, southeast of Somerset East.105 a. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 b. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 c. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 d. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 e. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.105 f. MBB6847 H. cymbiformis var. obtusa, southwest of Alicedale.106 a. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.106 b. J.N. Reddi in JDV93/46 H. decipiens var. pringlei, northeast of Middleton.107 a. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.107 b. G. Marx in JDV93/52 H. decipiens var. pringlei, northeast of Middleton.108 a. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 b. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.108 c. MBB6561 H. decipiens var. pringlei, Patryshoogte, E. Somerset East.109 a. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 b. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 c. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 d. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.109 e. MBB6776 H. cooperi var. cooperi, Glen Avon, southwest of Somerset East.110 a. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.110 b. MBB6563 H. cooperi var. cooperi, Koonap Bridge, Adelade.111 a. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 b. MBB6557 H. cooperi var. pilifera, west of Ripon station.111 c. MBB6557 H. cooperi var. pilifera, west of Ripon station.112 a. MBB656 H. decipiens var. pringlei, west of Ripon station.112 b. MBB656 H. decipiens var. pringlei, west of Ripon station.112 c. MBB6556 H. decipiens var. pringlei, west of Ripon station.112 d. MBB6556 H. decipiens var. pringlei, west of Ripon station.113 a. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.113 b. J. G. Marx in JDV91/82 H. cooperi var. cooperi, Woburn, Hogsback.114 a. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.114 b. D.M.Cummings H. bolusii var. blackbeardiana, south of Waterdown Dam, Upper Chilton.115. MBB659 H. bolusii var. blackbeardiana, Waterdown Dam.116 a. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 b. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 c. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 d. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.116 e. JDV96/89 H. gracilis var. gracilis, Gladhurst, Adelaide.117 a. JDV89/42 H. gracilis var. gracilis, Helspoort.117 b. JDV89/42 H. gracilis var. gracilis, Helspoort.117 c. JDV89/42 H. gracilis var. gracilis, Helspoort.118 a. MBB6614 H. gracilis var. gracilis, Helspoort.118 b. MBB6614 H. gracilis var. gracilis, Helspoort.119 a. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 b. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.119 c. MBB6927 H. cooperi var. cooperi, west of [112 a-d], Ripon.120. MBB6564 H. cooperi var. pilifera, Chancey Hall, south of Adelaide.121 a. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.121 b. MBB6591 H. cooperi var. pilifera, The Tower, south of Fort Beaufort.122 a. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 b. MBB6565 H. cooperi var. pilifera, west of Somerset East.122 c. MBB6565 H. cooperi var. pilifera, west of Somerset East.123 a. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.123 b. J.G.Marx in JDV91/85 H. cooperi var. pilifera, west of Fort Brown.124 a. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.124 b. JDV94/39 H. cooperi var. pilifera, Brakkloof, northwest of Grahamstown.125 a. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.125 b. T. Dold in MBB6851 H. aristata, Modderfontein, west of Commadagga.126 a. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 b. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 c. MBB6852 H. aristata, Stonefontain, s. Somerset East.126 d. MBB6852 H. aristata, Stonefontain, s. Somerset East.127. EvJ H. aristata, Aalwynspoort, east of Commadagga.128 a. MBB 6897 H. aristata, southeast of Commadagga.128 b. MBB 6897 H. aristata, southeast of Commadagga.129 a. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 b. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 c. MBB6580 H. decipiens var. pringlei, north of Jansenville.129 d. MBB6580 H. decipiens var. pringlei, north of Jansenville.130 a. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 b. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 c. MBB6581 H. decipiens var. pringlei, northwest of Janesville.130 d. MBB6581 H. decipiens var. pringlei, northwest of Janesville.131 a. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.131 b. MBB6582 H. decipiens var. pringlei, southeast of Mt. Stewart.132 a. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.132 b. MBB6583 H. decipiens var. pringlei, waaipoort, southeast of Mt. Stewart.133 a. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.133 b. MBB6587 H. decipiens var. minor, Die Bordjie, south of Baroe.134 a. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 b. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.134 c. J.G.Marx in MBB6845 H. gracilis var. gracilis?, east of Alicedale.135 a. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 b. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 c. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.135 d. MBB6562 H. cymbiformis var. obtusa Kagasmond, south of Alicedale.136 a. JDV96/90 H. cf. aristata, soutkloof, Addo.136 b. JDV96/90 H. cf. aristata, soutkloof, Addo.136 c. JDV96/90 H. cf. aristata, soutkloof, Addo.137. D.M.Cumming3870 H. cf. pilifera, Soutkloof, Addo.138 a. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.138 b. W.R.Branch459 in JDV87/53 H. aristata, Soutkloof, Addo.139 a. MBB6920 H. aristata, Soutkloof, Addo.139 b. MBB6920 H. aristata, Soutkloof, Addo.140. JDV86/117 H. cooperi var. pilifera, Addo.